
Abstract
Global epigenetic resetting in the gonadal primordial germ cells (PGCs) enables transition from early PGCs to gametogenesis and eventual restoring of totipotency after fertilization. This reprogramming process involves global DNA demethylation, changes in nuclear morphology and remodeling of repressive histone modifications. Here, using combined cytological and Hi-C-based methods, we reveal that, following the epigenetic reprogramming and concomitant with their commitment to gametogenesis, premeiotic gonadal germ cells display a distinct chromosome and genome architecture. This involves separation of individual chromosomes, anchoring of centromeres at the nuclear periphery, reduction in interchromosome interactions and disentangling of chromosome ends. Furthermore, genome-wide contact mapping documents remodeling of the three-dimensional (3D) genome architecture across all observable levels, including disruption of topologically associating domains (TADs), loss of detectable loops and reduced active–active compartment interactions. We further show that the diminished TADs correlate with the reduced levels of CCCTC-binding factor, thus providing an in vivo physiological model to understand genome folding principles. Lastly, we show that PGC-like cells, derived from embryonic stem cells, do not exhibit the same chromatin organization as embryonic germ cells. Collectively, our findings uncover the existence of a distinct chromatin architecture in premeiotic male and female gonadal germ cells and show that, alongside global DNA demethylation, the germline epigenetic reprogramming involves erasure of memory at the genome architectural level through profound reorganization of the 3D genome.
Similar content being viewed by others

Main
The germline is a unique cell lineage as it provides an enduring link to next generations. This property is exemplified by an early separation from soma (during early gastrulation in mammals) and manifested not only by the change in transcriptional regulation and in the underlying gene regulatory network in the nascent primordial germ cells (PGCs) but also by a profound change in global epigenetic landscape and regulation1. Following their specification (embryonic day (E6.25) in the mouse) and subsequent migration, the PGCs arrive to the embryonic developing gonad where they complete their epigenetic resetting (Fig. 1a). This involves global changes in DNA methylation and histone post-translational modifications2,3,4, as well as notable nuclear morphological changes4. Completion of the epigenetic resetting represents an important transition in the germline development; proliferating gonadal PGCs have reached the lowest level of their genomic DNA methylation and, developmentally, they are now primed to progress toward gametogenesis (meiotic prophase in the case of female PGCs and cell-cycle arrest for male PGCs). We previously reported the mechanistic link between epigenetic resetting and the transcriptional activation of the meiotic program; our work suggested that the successful execution of the gonadal reprogramming enables activation of the meiotic transcriptional program and, hence, progression toward gametogenesis5,6. Although global loss of DNA methylation and changes in repressive modifications in gonadal PGCs have been well documented5,6, we have still only very limited understanding regarding the unique nuclear morphology changes observed in the premeiotic gonadal PGCs, whether the observed cytological changes are linked to changes in three-dimensional (3D) chromatin architecture, regarding the molecular mechanisms involved or the biological relevance.

a, Top: schematic overview of nuclear architecture and heterochromatin changes during mouse PGC development. Bottom: timeline of the generation of PGCLCs and similarities to the development in vivo. b, Representative transmission electron microscopy images for somatic cells and germ cells at E13.5 (n = 2 embryos). c, Nuclear distribution of CENPA using IF staining in embryo (E9.5) or gonadal (E11.5–E13.5) sections (n = 3 biological replicates). OCT4, germ cell marker. Yellow arrowheads indicate PGCs. The square marks the cell selected for the zoomed-in view. DNA was stained with DAPI (blue). Scale bars, 10 µm (main images), 1 µm (zoomed-in view). d, Measurement of the shortest distance of each CENPA focus to the nuclear periphery. The mean value of each nucleus was calculated. n represents the number of cells analyzed. P values were calculated by two-tailed Mann–Whitney U-test (n = 3 embryos derived from independent litters and analyzed with 3 independent experiments). Box plots were plotted using Tukey’s method. The median and the first and third quartiles are indicated. The whiskers represent the furthest value within 1.5× the IQR. e, Percentage of cells with periphery (average distance of centromere to the nuclear periphery < 1 µm) and nonperiphery centromere association pattern. f, Minor satellite DNA-FISH on gonadal sections. DNA was stained with DAPI (blue). Data are representative of two independent experiments. Scale bar, 10 µm. g, Representative 3D reconstruction image. The total number of CENPA foci was calculated for each nucleus. The mean value is shown. Each dot represents the number derived from an individual cell (mean ± s.d.). The P values were calculated using a two-tailed Mann–Whitney U-test. h, Western blot analysis of CENPA. A total of 10,000 GFP-positive (PGC) and GFP-negative (Soma) cells were sorted using FACS from gonads of the Oct4–GFP mice (GOF 18ΔPE–EGFP) (n = 2 embryos derived from independent litters and analyzed with 2 independent experiments). *P < 0.05, ***P < 0.001.
Recent developments of in vitro gametogenesis protocols allow for the differentiation of PGC-like cells (PGCLCs) and mature gametes from pluripotent stem cells7. While these protocols represent true technical breakthroughs, mature gametes are generated only with low efficiency and display frequent abnormalities in meiosis progression. Given the link between successful epigenetic resetting and progression toward meiosis, it is essential to determine whether these PGCLCs derived in vitro, used as a surrogate for early germline development, undergo epigenome resetting equivalent to that observed in developing embryonic germ cells in vivo.
Results
Chromosome anchoring at the nuclear periphery in PGCs
We previously reported that gonadal PGCs exhibit chromocenter disappearance and chromatin decondensation4 (Fig. 1a). This observation is further supported by transmission electron microscopy. In differentiated gonadal somatic cells, electron-dense heterochromatin is clearly visible around the nucleolus and at the nuclear periphery, while PGCs exhibit an overall less electron-dense, dispersed chromatin state (Fig. 1b). Mouse cells are characterized by clustering of constitutive pericentromeric heterochromatin (PCH) in chromocenters. Given the disappearance of these structures in PGCs, we first examined the nuclear localization of centromeres in gonadal PGCs and in the neighboring somatic cells. Of note, mouse cells contain acrocentric chromosomes and, hence, the centromere is located near one end of the chromosome. As a proxy for centromere localization, we used immunofluorescence (IF) staining of CENPA, a centromere-specific H3 histone variant. We found that CENPA foci resided within conventional chromocenters (DAPI-dense regions) in somatic cells, whereas, in PGCs, these foci were positioned close to the nuclear periphery with few attached to nucleoli near the periphery (Fig. 1c). The striking repositioning of centromeres is apparent when assessing the average distance between the CENPA signal and the nuclear lamina (Fig. 1d). Centromeres are in proximity to the nuclear envelope (NE) in both male and female gonadal PGCs, while the distribution showed a greater variation in somatic cells (Fig. 1d,e). To provide further support for our findings, we confirmed the PGC-specific localization of centromeres at the nuclear periphery using minor satellite DNA fluorescence in situ hybridization (FISH) (Fig. 1f).
Following their entry into the gonadal anlagen, the gonadal PGCs undergo global epigenetic reprogramming8 and the onset of sex differentiation (Fig. 1a). In females, this precedes entry into the meiotic prophase, whereas, in males, PGCs enter cell-cycle arrest after E14.5. Notably, the observed centromere repositioning is observable both in the male and female PGCs, before E13.5 (that is, before the entry into the meiotic prophase in the female). Further progression of meiosis in the females is associated with the bouquet formation demarcated by the coclustering of centromeres at the nuclear periphery of female fetal germ cells (Extended Data Fig. 1a). To the contrary, the peripheral localization of centromeres persists in male germ cells following mitotic arrest; it is still apparent at E18.5 and in undifferentiated spermatogonia in the postnatal testis (P10), but dissolves once the spermatogonia undergo further differentiation (Extended Data Fig. 1b).
The peripheral distribution of centromeres in PGCs is apparent in 3D reconstruction of confocal images (Fig. 1g and Supplementary Video 1). Using this approach, we identified, on average, 31.8 CENPA foci in PGCs compared to 8.6 foci in somatic cells (Fig. 1g and Extended Data Fig. 1c). As mouse cells contain 40 chromosomes, our data indicate that each centromere is separated in PGCs, which is in stark contrast to mouse somatic cells where centromeres tend to aggregate within chromocenters. Surprisingly, despite the declustering of centromeres, the signal in the individual CENPA foci seemed more pronounced in PGCs (Fig. 1c and Extended Data Fig. 1a–c), suggesting higher CENPA expression in the germ cells. This was confirmed using western blot that showed considerably higher CENPA levels in E13.5 PGCs compared to the differentiated gonadal somatic cells (Fig. 1h).
To further characterize the spatial organization of chromosomes in PGCs, we examined telomere localization. IF combined with 3D telomere DNA-FISH revealed a significantly higher number of telomere foci in PGCs compared to somatic cells (♂: Soma, 39; PGC, 66; ♀: Soma, 46; PGC, 60) (Fig. 2a,b). We also observed the mean intensity of individual telomere FISH signals to be markedly lower in germ cells, consistent with a more dispersed telomeric configuration (Fig. 2c and Supplementary Video 2). To assess the telomere’s spatial distribution, we measured the shortest distance from each telomere focus to the nuclear periphery. In each cell, the radius, derived from the nuclear volume, was normalized and divided into five bins. Bin 1 represents the closest proximity to the nuclear periphery, while bin 5 corresponds to the center of the nucleus (Fig. 2d and Extended Data Fig. 1d). Approximately half of the telomere FISH signal resided in bin 1, whereas the other half was located in bins 2–5 in both male and female germ cells (Fig. 2d) (bin 1: ♂, 54%; ♀, 53%). Collectively, these findings support a model in which one end of the chromosome (comprising the proximal telomere and centromere) is tethered at the nuclear periphery, while the distal telomere is positioned inside the nucleoplasm.

a, 3D telomere DNA-FISH combined with IF on gonadal sections (n = 3 biological replicates). MVH, germ cell marker. White and yellow squares indicate somatic cells and PGCs, respectively. DNA was stained with DAPI (blue). Scale bars, 10 µm (main images), 1 µm (zoomed-in view). b, The total number of telomere foci was calculated for each nucleus (mean ± s.d.). The number of nuclei analyzed is shown. Scale bar, 1 um. c, Fluorescence intensity was measured for each DNA-FISH telomere signal. Each dot represents the mean value of each cell (mean ± s.d.). d, Distribution of telomere foci in the nuclei. The shortest distance from telomere foci to the nuclear periphery was measured on the basis of 3D reconstructed images. The distance was normalized to the radius, which was calculated from the nuclear volume. The distance of each focus was classified into five bins, where bin 1 is the closest bin to the nuclear periphery. Each dot represents each cell (mean ± s.d.; n = 3 embryos analyzed with 3 independent experiments). In b and c, P values were calculated using a two-tailed Mann–Whitney U-test. *P < 0.05, **P < 0.01, ***P < 0.001.
Considering the possible mechanism underlying the observed perinuclear localization of centromeres, we first focused on the histone modifications associated with constitutive heterochromatin. These modifications are known to have a crucial role in facilitating chromatin interaction with the nuclear lamina, leading to the formation of lamina-associated domains (LADs)9. Consistent with previous reports2,10, we found significant reductions in global H3K9me2 and H4K20me3 levels in PGCs (Extended Data Fig. 2a,b). Interestingly, H3K9me3, a modification typically associated with PCH, exhibited enrichment primarily around the nuclear periphery in PGCs starting from E11.5, with further pronounced peripheral localization at E13.5. (Fig. 3a,b and Extended Data Fig. 2c). On average, ~80% of H3K9me3-enriched regions were located in the vicinity of the NE in individual PGCs starting from E11.5 (Fig. 3b and Methods), with the majority of PGCs displaying this perinuclear H3K9me3 pattern (E11.5: 61.5%; E13.5: ♂, 70%; ♀, 77.6%) (Fig. 3c). This observed PCH localization was also independently validated using DNA-FISH targeting major satellite repeats (MSRs) (Fig. 3d).

a, Representative IF image of H3K9me3 and lamin B1 (n = 3 biological replicates). DNA was stained with DAPI (blue). b, Quantification of total H3K9me3 area and H3K9me3 area within the perinuclear area (Methods). Each dot represents the percentage within each cell (mean ± s.d.). P values were calculated using a two-tailed Mann–Whitney U-test (n = 3 embryos analyzed with 3 independent litters). c, The proportion of cells with perinuclear H3K9me3 (>70% of total H3K9me3 area). d, Representative image of major satellite DNA-FISH combined with IF staining. e, Representative IF image of ATRX. f, Representative image of single molecular RNA-FISH of major satellite combined with IF. Yellow arrowheads indicate signals from PGCs. White arrowheads indicate signals from somatic cells. DNA was stained with DAPI (blue). For the scatter plot, colored dots represent the average percentage of each embryo and gray dots represent the percentage of each tissue section (n = 3 biological replicates; mean ± s.d.). P values were calculated using a two-tailed Mann–Whitney U-test. g, Western blot analysis of lamin B1 and LBR. In total, 10,000 GFP-positive (PGC) and GFP-negative (Soma) cells were sorted using FACS from the gonads of Oct4–GFP mice (GOF 18ΔPE–EGFP) (n = 2 animals, representative of 2 independent experiments). h, Comparative gene expression using published scRNA-seq data27. i, Representative IF image of LBR. Germ cell markers, OCT4 (E9.5) and MVH (E13.5). j, Gonads were cultured with chaetocin for 12 and 24 h. Measurement of the shortest distance of each CENPA focus to its nuclear periphery. The mean value of each nucleus was calculated. n represents the number of cells analyzed. Each cell is represented by a gray dot. Colored dots represent the mean value of each individual biological replicate (mean ± s.d.). P values were calculated using a two-tailed Mann–Whitney U-test. In d, e and i, data are representative of three independent experiments (three embryos from independent litters). Scale bars, 10 µm (a,d–f,i,j). **P < 0.01, ***P < 0.001.
Distinct perinuclear blocks of H3K9me3 were observed in early germ cells of both sexes until E13.5. As female germ cells entered meiotic prophase, the H3K9me3 signal became more diffuse with the conventional chromocenter H3K9me3 organization reestablished only in the postnatal (P10) ovary (Extended Data Fig. 2c,d). In the male germline, a diffuse pattern was visible in mitotically arrested male germ cells at E18.5, with clear perinuclear enrichment of H3K9me3 persisting through to the undifferentiated spermatogonia stage in P10 testes (Extended Data Fig. 2c,d).
PCH tends to be amongst the late DNA-replicating regions that display a distinctive proliferating cell nuclear antigen PCNA distribution pattern11. Consistent with this, we observed PCNA enrichment around the edges of chromocenters in somatic cells during late S phase, whereas, in PGCs, PCNA displayed a perinuclear distribution (Extended Data Fig. 2e).
Regarding other factors typically associated with constitutive heterochromatin, we did find the levels of heterochromatin-associated proteins HP1α greatly reduced in PGCs, suggesting a potential contribution to the overall more decondensed chromatin structure (Extended Data Fig. 3a,b). To the contrary, ATRX, a chromatin remodeler that is critical for maintaining PCH stability, was associated with perinuclear PCH in both interphase and during mitosis (Fig. 3e and Extended Data Fig. 3c), indicating that some components of constitutive heterochromatin remain detectable throughout the cell cycle, although their spatial organization is markedly altered in PGCs.
Recent studies documented that transcripts originating from MSRs are required for the establishment or stabilization of constitutive heterochromatin12,13,14,15. Hence, we carried out single-molecule RNA-FISH combined with IF to ask whether the perinuclear association of PCH is linked to its altered transcriptional activity. Although all PGCs show discrete perinuclear PCH, only 5–10% of PGCs expressed detectable MSR transcripts (Fig. 3f), with the level of PCH transcription in PGCs comparable to that observed in the neighboring somatic cells (Fig. 3f). In addition, while transcriptionally active genomic loci are often associated with the nuclear pore complex (NPC)16, electron-dense peripheral heterochromatin was not found in direct vicinity of the NPC in PGCs (Extended Data Fig. 3d). Collectively, our findings suggest that perinuclear localization of PCH in PGCs is not associated with altered transcriptional activity of MSRs.
Considering other possible mechanisms affecting heterochromatin formation or localization, association with nuclear lamina has been shown to stabilize heterochromatin9,17. In line with this, we observed a marked alteration of nuclear lamina composition characterized by a reduction in nuclear lamins, including lamins A/C and B1 specifically in PGCs at both mRNA and protein levels (Fig. 3a,g,h and Extended Data Fig. 3e). Interestingly, in contrast to the low levels of lamins A/C and B1, we found the expression of lamin B receptor (LBR) to be greatly elevated in gonadal germ cells at E13.5 and in the postnatal testis (Fig. 3g–i and Extended Data Fig. 3f). Lastly, to test whether H3K9 methylation is required for perinuclear heterochromatin localization in PGCs, similarly to what was reported in Caenorhabditis elegans17, we cultured the dissected mouse E13.5 gonads in the presence of the H3K9 methyltransferase inhibitor chaetocin. After 24 h of treatment, centromeres were visibly detached from the nuclear periphery in PGC nuclei, whereas centromere tethering remained intact in DMSO-treated controls (Fig. 3j and Extended Data Fig. 3g,h). Collectively, our findings show that PGCs are characterized by an altered composition of nuclear lamina with H3K9me3 required for anchoring of (peri)centromeres at the nuclear periphery.
Chromosome interactions and genome organization in PGCs
The disappearance of chromocenters and broader alterations in heterochromatin architecture prompted us to investigate global genome organization during this critical stage of germline development. We performed low-input Hi-C on isolated male and female E13.5 PGCs (ΔPE–Oct4, GFP-positive) and corresponding somatic cells (GFP-negative) (Fig. 4a), generating almost half a billion uniquely mapped reads for each biological replicate (Supplementary Table 1). Following an assessment of replicate reproducibility (Extended Data Fig. 4a), we merged the two biological replicates to enhance the resolution of the Hi-C contact matrices.

a, Experimental scheme for the isolation of somatic cells and PGCs for low-input Hi-C experiments. Two biological replicates were collected from different litters. b, Hi-C matrices of merged replicates showing chromatin interactions of chromosomes 1–6 in somatic cells and PGCs. Right: heat map illustrating the log2-transformed ratio of contacts between PGC and Soma, where red indicates an enrichment of contacts and blue indicates a depletion of contacts. Hi-C matrices at 1-Mb resolution were normalized at a whole-genome level. The matrix comparison was based on observed/expected matrices. Chr, chromosome. c, Interchromosomal contact frequency of regions that are proximal to the centromere (4–5 Mb) and distal regions (20–21 Mb). For each sample, each dot is the contact frequency between a region from a chromosome and the same region in a different chromosome. All possible interactions are shown (female, n = 190; male, n = 210). d, Bar chart showing the fraction of cis (intrachromosomal) contacts (cis/(cis + trans)) derived from all autosomes. e, Hi-C contact probability as a function of distance (P(s) curves, where P is probability and s is genomic distance) in somatic cells and PGCs. Expected contacts were calculated for each chromosome using Hi-C matrices at 10-kb resolution; plots show the mean of all autosomes. f, Box plots showing the ratio of cis interactions that span more than 1 Mb in somatic cells and PGCs. Each point represents an autosome (n = 19). The P values were calculated using a two-sided Wilcoxon signed-rank test. In the box plot, central lines represent median values; upper and lower limits show the 75th and 25th percentiles, respectively; box heights represent the IQRs; and whiskers are 1.5× the IQR.
Like somatic cells, PGCs displayed a pattern of chromosome territories without Rabl-like features (a specific interphase chromosome arrangement in which centromeres and telomeres are polarized to opposite sides of the nucleus) (Fig. 4b and Extended Data Fig. 4c). Consistent with our cytological observations, our data showed a clear reduction in interchromosomal interactions in both male and female PGCs (Fig. 4c), resulting in a relatively higher proportion of cis contact compared to somatic cells (Fig. 4d and Extended Data Fig. 4b,c). Notably, intrachromosomal interactions on the X chromosome were greatly reduced in female PGCs in line with reactivation of the inactive X chromosome in these cells (Extended Data Fig. 4c). Within intrachromosomal interactions, short-range interactions (Fig. 4e) were relatively more abundant in PGCs, whereas long-range chromatin interactions (>1 Mb) were significantly reduced (Fig. 4e,f). Our findings are, thus, consistent with the possibility that PGCs exhibit a distinct and dispersed chromosome distribution, with more ‘stretched’ chromosomes within their chromosome territories.
On the basis of its organization, the genome can be classified into the A compartment, associated with active euchromatin, and the B compartment, associated with a repressive chromatin state. Our data revealed a marked reduction in compartmentalization in both male and female PGCs compared to somatic cells (Fig. 5a and Extended Data Fig. 4d). Furthermore, although B–B interactions were largely maintained, A–A interactions were surprisingly substantially diminished in both male and female PGCs (Fig. 5b,c and Extended Data Fig. 4e), a trend also evident in the contact matrices (Fig. 5d). Despite the overall weakening in compartment strength, we observed only limited switching between A and B compartments between somatic cells and PGCs (Fig. 5e), suggesting that the reduction in compartment strength does not reflect widespread compartment identity changes.

a, Strength of compartmentalization of somatic cells and PGCs. b, Saddle plots showing compartment interactions. A–A compartment (active–active) interactions were significantly reduced in PGCs compared with somatic cells. c, Box plots showing compartment interaction strength between the same or different types of compartments. Each point represents an autosome (n = 19). P values were calculated using a paired Wilcoxon test (two-sided). Box plots were plotted using Tukey’s method. The median and the first and third quartiles are indicated. The whiskers represent the furthest value within 1.5× the IQR. d, Hi-C contact matrices of female somatic cells and PGCs for the whole chromosome 2. The plaid pattern reflects genome compartmentalization, where compartments of the same type interact more frequently with each other than with other contacts. The heat map illustrates the difference in Hi-C matrices. Red indicates an enrichment of contacts in PGC and blue indicates a depletion. This comparison was based on observed/expected (O/E) matrices. Bottom: eigenvector of the correlation matrix obtained at 500-kb resolution was used to derive compartment type. Positive values were assigned to compartment A, whereas negative values were assigned to compartment B. e, River plots showing compartment changes between somatic cells and PGCs in Mb.
Further analysis of the Hi-C contact matrices revealed a striking reduction in topologically associating domains (TADs) in PGCs (Fig. 6a–c and Extended Data Fig. 5a,b). Using TADs previously defined in mouse embryonic stem cells (ESCs)18, we observed a substantial decrease in TAD strength in PGCs compared to somatic cells (♀: Soma, 0.689; PGC, 0.21; ♂: Soma, 0.699; PGC, 0.392) (Fig. 6b and Extended Data Fig. 5b). This was further confirmed by de novo detection of TAD boundary scores that detected a significantly lower TAD boundary strength in PGCs (Fig. 6c). Notably, despite the striking loss of TAD boundaries, comparison with neighboring gonadal somatic cells revealed no clear correlation between changes in TAD boundary strength and transcriptional activity (Extended Data Fig. 5c,d). We next asked whether the observed TAD loss is linked to specific local chromatin environments. Using our H3K9me3 or H3K27me3 ultralow-input native chromatin immunoprecipitation (ChIP) data6, we performed k-means clustering and identified four distinct boundary clusters. Despite differences in compartment identity and heterochromatin marks, all clusters exhibited a general reduction in TAD strength, as measured by changes in insulation (Extended Data Fig. 5e).

a, Normalized Hi-C interaction frequencies (40–50 Mb, chromosome 2) in somatic cells (top) and PGCs (bottom), male (right) and female (left). b, TAD aggregate plots showing diminished TADs in both male and female PGCs. TAD strength values (median) are indicated for each group. c, Boundary strength was calculated at boundaries identified in our Hi-C libraries. Boundary strength is significantly reduced in PGCs compared with somatic cells. P values were calculated using a two-sided Student’s paired t-test (female, n = 5,512 boundaries; male, n = 6,251 boundaries). Outliers are not depicted. d, Loop aggregate plots for somatic cells (left) and PGCs (right). Loop strength is shown by the genome-wide averaged contact signals (Methods). The contact map was normalized using matrix balancing and distance (O/E). Positive enrichment is shown in red and negative signals are displayed in blue. The ratio of contact enrichment for the center pixels is annotated within each plot. e, Normalized Hi-C interaction frequencies (50.7–52.7 Mb, chromosome 6) in somatic cells and PGCs, male and female. Bottom: insulation plot. Identified TADs and subTADs are labeled with different colors. Loops are indicated by circles. f, Left: Gaudí plot generated by METALoci (Methods) showing genomic regions at 10-kb resolution, overlaid with corresponding TADs (as shown in e). Different colors represent distinct TADs. Right: log10-transformed sum of ATAC-seq signal within each 10-kb bin is mapped onto the same graph layout, illustrating chromatin accessibility. g, Representative IF image of CTCF (n = 3 biological replicates). Germ cell markers, AP2γ and OCT4. Yellow arrowheads indicate PGCs. DNA was stained with DAPI (blue). Scale bar, 10 µm. h, Image quantification analysis showing normalized CTCF intensity (Methods) from three biological replicates (mean ± s.d.). P values were calculated using a two-tailed Mann–Whitney U-test. i, CTCF footprints of corrected ATAC-seq signal at CTCF-binding sites in TAD boundaries (mild, severe and total loss categories). CTCF footprint scores were calculated from published ATAC-seq data for E13.5 PGCs27. j, CTCF motif score for TAD boundaries with mild loss, severe loss and total loss. The motif with the highest score was considered for each TAD boundary. P values were calculated using a two-sided Student’s t-test. Box plots were plotted using Tukey’s method. The median and the first and third quartiles are indicated. Whiskers represent the furthest value within 1.5× the IQR.
In line with the diminished TAD architecture, we detected only 17 chromatin loops in male and female PGCs, in contrast to 300 in male and 1,392 in female somatic cells, respectively (Fig. 6d). Loop strength was also notably diminished in PGCs, indicating reduced loop formation in germ cells (Fig. 6d,e).
To further explore alterations in 3D genome organization, we applied the METALoci method19 for genome modeling, followed by the integration of assay for transposase-accessible chromatin with sequencing (ATAC-seq) data onto the resulting structures (Fig. 6e,f). This analysis confirmed profound changes in genome architecture in PGCs. We also noted that the accessible chromatin sites in somatic cells tended to cluster at TAD boundaries, while this spatial organization was less apparent in PGCs (Fig. 6f).
Epigenetic perturbations do not recapitulate PGC-specific 3D genome reorganization
Given the unique epigenetic landscape of PGCs at this stage of germline development (Fig. 1a), we considered whether the observed changes in 3D chromatin architecture could be driven by global changes in the epigenome. To test this, we examined whether global epigenetic perturbations, such as chromatin decondensation and genome-wide global DNA demethylation, could induce similar 3D genome architecture remodeling in cultured cells.
DNA methyltransferase (DNMT1, DNMT3A and DNMT3B) triple-knockout (Dnmt-TKO) ES cells displayed canonical chromocenters with a conventional CENPA distribution and H3K9me3 pattern, similar to their parental J1 ES cell line (Extended Data Fig. 6a). However, this stability may reflect long-term adaptation to gene knockout. To induce acute loss of DNA methylation, we cultured ES cells in a 2i medium supplemented with vitamin C. This culture condition has been shown to induce genome-wide DNA demethylation20. Notably, this treatment did not result in chromocenter loss and relocalization to the nuclear periphery (Extended Data Fig. 6b). Furthermore, to artificially induce chromosome decondensation, we treated cells with TSA and SGC0946, potent and selective inhibitors of histone deacetylases (HDACs) and the DOT1L histone lysine methyltransferase, respectively21,22,23. After 48 h of treatment, chromocenter appearance and CENPA distribution was barely altered in both cell lines (Extended Data Fig. 6a,b). These results demonstrate that neither reduction of DNA methylation or chromosome decompaction lead to chromocenter disappearance and centromere redistribution. Lastly, analysis of published Hi-C data from Dnmt-TKO and J1 (parental cell line control) ES cells24 confirmed that loss of DNA methylation does not globally alter TAD strength and compartment interactions (Extended Data Fig. 6c–e).
Together, these findings indicate that the profound reorganization of 3D genome architecture observed in PGCs is not simply a consequence of global DNA demethylation or chromatin decompaction and suggests the involvement of additional, PGC-specific regulatory mechanisms.
Low CCCTC-binding factor occupancy underlies genome organization changes in PGCs
CCCTC-binding factor (CTCF) and RAD21 are critical regulators of higher-order genome architecture25,26. Surprisingly, we found that CTCF protein levels were significantly reduced in postmigratory gonadal PGCs compared to the neighboring somatic cells (Fig. 6g,h). This reduction was further supported by western blot and RNA-seq analysis, which revealed a gradual downregulation of Ctcf expression in gonadal PGCs27 (Extended Data Fig. 6f–h). Notably, reduced CTCF levels persisted to postnatal testis (Extended Data Fig. 6i). In contrast, RAD21 remained highly expressed in PGCs (Extended Data Fig. 6g,j).
These findings prompted us to investigate whether the reduction in CTCF levels contributes to the attenuation of TAD boundaries in PGCs. Comparative analysis between somatic and germ cells revealed very few PGC-specific boundaries. Where present, these typically arose from expansions of existing TAD boundaries or more intricate rearrangements rather than through the emergence of genuinely new or stronger boundaries (Extended Data Fig. 7a,b). In contrast, we observed a clear loss of strength in somatic-specific insulation boundaries (Extended Data Fig. 7b).
On the basis of the degree of attenuation observed in PGCs relative to somatic cells, these boundaries could be categorized into three groups: ‘mild loss’, ‘severe loss’ and ‘total loss’ (Extended Data Fig. 7c–f and Supplementary Table 2). In agreement with the reduced CTCF level, analysis of published PGC ATAC-seq data27 revealed reduced CTCF footprinting scores across all three boundary groups, with the most pronounced reduction observed at boundaries with total loss (Fig. 6i). Interestingly, the degree of boundary loss negatively correlates with the sequence conservation of the CTCF-binding motif (Fig. 6j) suggesting that the strength of CTCF binding might determine the retention of TAD boundaries when CTCF availability is limiting. We also note that, using a Gene Ontology (GO) term analysis, we did not observe specific association of germline-related or meiosis-related genes with any of our TAD boundary groups. (Extended Data Fig.7g).
3D genome reorganization in PGCs occurs before meiotic entry and independently of cell-cycle effects
Genome organization can be influenced by mitotic chromosome condensation28 and synaptonemal complex (SC) formation during meiotic prophase29,30. Gonadal PGCs at E13.5 are still proliferative, exhibiting a cell-cycle pattern comparable to ES cells31,32,33. This is supported by our data; at E13.5, 21% of male and 25% of female PGCs were in G2 phase (positive for cyclin B1), while only 2.7% and 2.2%, respectively, were in M phase (Extended Data Fig. 8a,b), indicating that mitotic cells do not dominate the population. Furthermore, consistent with previous reports, fewer than 2.7% of female PGCs exhibited weak expression of the meiotic marker SCP3 (ref. 34) at E13.5 (Extended Data Fig. 8c,d), confirming that our analysis captures premeiotic germ cells.
Overall, our data show that the dramatic 3D genome remodeling observed in male and female PGCs, characterized by the global reduction in compartment strength, weakening of TAD structure and loss of detectable loops, occurs in proliferative gonadal germ cells before their respective entry into the meiotic prophase.
Comparison of in vivo embryonic PGCs and in vitro PGCLC differentiation
PGCLCs have been successfully derived from mouse pluripotent ES cells as an in vitro model for early germline development35. While these cells faithfully recapitulate early premigratory and migratory stages of PGC development (up to E9.5 in the mouse36), PGCLCs do not have the intrinsic capacity to enter meiosis and can progress toward gametogenesis only when reaggregated with somatic cells of the gonad7 (Fig. 1a). Previous studies have indicated that PGCLCs exhibit nuclear morphology similar to the embryonic PGCs, including reduced lamin B1 levels and loss of chromocenters37.
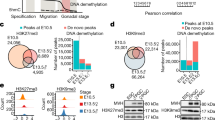
To assess whether PGCLCs can recapitulate the 3D genome reorganization observed in gonadal PGCs, we differentiated PGCLCs (PGCLCd4) from ES cells and extended their culture to generate PGCLCd4c6, a stage reported to undergo epigenetic reprogramming similar to that seen in vivo (Extended Data Fig. 9a,b and Methods). Contrary to the uniform nuclear pattern observed in in vivo PGCs, PGCLCs exhibited more diverse nuclear organization patterns across the cell population. Centromere distribution also showed significant cell-to-cell variations (Extended Data Fig. 9c); despite reduced chromosome compaction previously reported in PGCLCs, conventional H3K9me3-enriched chromocenters were readily detected both in PGCLCd4 and PGCLCd4c5 (Extended Data Fig. 9d,e). Moreover, we did not observe a decrease in CTCF protein levels and RAD21 expression remained comparable to non-PGCLCs (Extended Data Fig. 9f). Comparative analysis of our Hi-C with the published dataset from PGCLCs and germline stem cell (GSC)-like cells (GSCLCs)38 revealed that in vivo PGCs form a distinct cluster, clearly separate from both somatic cells and GSCs derived in vitro (Extended Data Fig. 10a). Notably, PGCLCs did not exhibit the same degree of TAD weakening as observed in premeiotic gonadal PGCs in vivo (Extended Data Fig. 10b–d). In contrast, GSCs derived from neonatal spermatogonia showed diminished TADs comparable to PGCs.
Together, these results suggest that, in vivo, the genome of premeiotic gonadal germ cells undergoes a unique transition phase that coincides with the acquisition of competency by PGCs to progress toward gametogenesis and meiosis. This transition cannot be robustly recapitulated in the current PGCLC in vitro differentiation system, in agreement with the reduced capacity of these in vitro derived cells to undergo meiosis.
Conservation between mouse and human PGC development
To determine whether the striking chromatin architecture reorganization observed in premeiotic PGCs is specific to mice or represents an evolutionarily conserved feature of embryonic premeiotic germ cells, we examined human fetal gonads. Given that chromocenters are a characteristic feature of mouse nuclei but not of human cells, we asked whether other aspects of nuclear organization, such as centromere distribution and chromatin-associated factors, are conserved between the two species.
Unlike the mouse, the human genome contains metacentric chromosomes. Interestingly, examination of human embryonic gonads (9–14 weeks after conception) revealed several features reminiscent of mouse PGCs, such as elevated CENPA levels, separated individual CENPA foci (Fig. 7a–c), low lamin B1 levels, very high LBR levels (Fig. 7d,g), low CTCF levels and high RAD21 levels (Fig. 7e,f). These features were specific to germ cells, suggesting evolutionary conservation of nuclear reorganization during this developmental window. Strikingly, we also identified a distinct subpopulation of germ cells with perinuclear distribution of centromeres, with over 80% of CENPA foci positioned at the NE. This population increased from ~4% at postconception week 9 (PCW9) to ~15% at PCW14 in both testis and ovaries (Fig. 7g,h). It is important to note that, unlike mouse fetal gonadal germ cells, human germ cells represent a developmentally heterogenous population at these stages, ranging from mitotically proliferating germ cells to cells in the various stages of the meiotic prophase (in the female embryos)39. It is, thus, possible that the observed perinuclear localization of centromeres is characteristic of a specific subpopulation of germ cells. Together, these observations suggest that key features of nuclear reorganization are conserved between mouse and human germ cells, highlighting a potentially fundamental transition in genome architecture that accompanies the acquisition of meiotic competence.

a, Schematic of human early germline development. Asynchronized germ cell populations exist after PCW8. hPGCs, human primordial germ cells. b,c, Representative IF image of maximum projection from z-stack images (b) and corresponding quantification (c). The total number of CENPA foci was calculated for each nucleus. The mean value is shown. Each dot represents the number derived from an individual cell. DNA was stained with DAPI (n = 54 and n = 17, where n represents the number of cells analyzed; n = 2 embryos analyzed with 2 independent experiments; mean ± s.d.). d–f, Representative IF image of LBR (d), CTCF (e) and RAD21 (f) from human embryonic gonadal sections. Germ cells are indicated by yellow arrowheads. g, Representative IF images for CENPA using human gonadal sections. Germ cell markers, MVH (PCW14) and AP2γ (PCW9). h, Germ cells with perinuclear CENPA foci (>80% of total foci tethered to the NE) were calculated at the assigned gestational stage (PCW9 and PCW10, n = 1; PCW14, n = 2). In d–g, data are representative of two independent experiments (two independent embryos). Scale bars, 10 µm (b,d–g), 5 µm (h).
Discussion
We and others have previously shown that gonadal PGCs undergo comprehensive epigenetic resetting. This includes genome-wide DNA demethylation, remodeling of repressive histone modifications and changes in nuclear size and morphology4,40. Here, through a combination of cytological and Hi-C analyses, we demonstrate that post-reprogramming premeiotic PGCs acquire a unique and distinctive genome organization (Fig. 8). Specifically, we show that both male and female premeiotic gonadal PGCs exhibit globally remodeled 3D genome organization, characterized by weakened compartmental insulation, diminished TAD structure and a pronounced loss of detectable chromatin loops. Importantly, this reorganization is likely linked to the observed reduction in CTCF levels.

Following the germline specification and their migration, PGCs undergo epigenetic resetting that involves remodeling of chromatin architecture. This includes chromatin decondensation, disruption of chromocenters and tethering of the H3K9me3-enriched PCH around the nuclear periphery. At the molecular level, we observe weakening of interchromosomal interactions, attenuation of TAD strength and a loss of loop formation specifically in PGCs compared with the neighboring somatic cells. The schematic is based on the mouse model. Mouse chromosomes are acrocentric (that is, one of the telomeres is located near the centromere).
Recent studies using degron-mediated acute depletion of CTCF in ES cells showed that only a small subset of enhancer–promoter interactions are disrupted within 3 h after CTCF depletion, while global gene expression remains largely unaffected41. Our findings extend this observation by providing a physiological example in which a germline-specific transcription program is executed in the context of low CTCF levels and minimal high-order genome structure. We propose that epigenetic reprogramming during early germline development resets genome architecture to a more primitive state, in which transcription is maintained independently of canonical 3D genome features such as chromatin loops and TADs. In this context, transcription may be supported by local chromatin environment or transcription factor enrichment alone.
Alternatively, TAD-independent transcriptional regulation may represent a germline-specific strategy. During spermatogenesis, for example, transcription remains active in pachytene spermatocyte despite the loss of TADs as homologous chromosomes pair and condense into loop arrays42. In this case, enhancer–promoter contacts can be identified even when domain formation has mostly disappeared43. Interestingly, some organisms, such as Arabidopsis thaliana and C. elegans, execute transcription and meiosis in the absence of TADs, suggesting that a domain-level genome organization could be predominantly relevant for the formation of complex somatic tissues, especially in metazoans44,45.
Although the attenuation of TADs and loss of chromatin loops in PGCs are likely linked to reduced CTCF levels, the extent of 3D genome architectural disruption we observe exceeds that reported in CTCF degron systems26, suggesting that additional mechanisms may be involved. One such factor is WAPL, a cohesin-release factor that promotes the dissolution of cohesin-mediated loops. Supporting this possibility, Wapl expression is elevated in PGCs compared to somatic cells (Extended Data Fig. 6g). In addition, the stoichiometry of structural maintenance of chromosomes (SMC) proteins could impact loop extrusion dynamics. Our analysis revealed variable gene expression of cohesin subunits between PGCs and somatic cells (Extended Data Fig. 6g and Supplementary Table 3). For example, RAD21 expression is generally higher in PGCs, although it shows variability in females at E13.5 (Extended Data Fig. 6g,j). Further work will, thus, be required to dissect the dynamics of loop extrusion in this physiological context.
Our results indicate that the observed dissolution of conventional chromocenters in premeiotic gonadal PGCs occurs through a process where one end of the chromosome (specifically the pericentromere) attaches to the nuclear periphery (Fig. 1c). We also show that individual chromosome ends are more dispersed in germ cells (Figs. 1g and 2b). Supporting this observation, the frequency of interchromosomal interactions is greatly reduced in germ cells (Fig. 4b,c,e). We hypothesize that the spatial chromosome separation is the first step to allow nonhomologous chromosomes to be disentangled from chromocenters where pericentromeric repeats are normally clustering and envisage that this could increase the probability of chromosomes finding the homolog. Intriguingly, during the early stages of oogenesis in Drosophila, the unpairing and separation of homologous chromosomes occur in mitotic female GSCs before they enter the meiotic prophase46. We observed similar changes in nuclear architecture in human germ cells, despite the fact that human cells have metacentric chromosomes and, in general, lack conventional chromocenters. Our results document the existence of dispersed centromeres in human germ cells and a subset of germ cells exhibiting the perinuclear anchoring of centromeres (Fig. 7b,h). Importantly and similarly to mouse cells, human fetal germ cells show reduced CTCF and increased RAD21 expression (Fig. 7e,f), further pointing toward the evolutionary conservation of our findings.
Importantly, despite diverging along sex-specific developmental trajectories after E12.5, both male and female embryonic germ cells undergo remarkably similar changes in nuclear morphology, chromosome organization and 3D genome architecture during the described gonadal stages. These shared features suggest that the reprogramming of genome architecture occurring in gonadal germ cells of both sexes may serve to establish a chromosomal configuration primed for meiotic entry. Supporting this notion, substantial evidence indicates that male germ cells at this stage possess the intrinsic capacity to initiate meiosis. For example, exogenous application of retinoic acid to fetal testes or genetic ablation of the retinoic acid-degrading enzyme CYP26B1 in male embryos could induce premature meiotic entry in male gonadal germ cells47,48,49,50.
A hallmark of meiotic prophase is the reorganization of telomeres to the nuclear periphery and formation of the bouquet—a configuration that facilitates chromosome movement and homologous chromosome pairing51. How and when telomeres are attached is currently unclear. Our findings reveal that the initial chromosome anchoring occurs through centromeres in mitotic (premeiotic) germ cells and propose a possible role for H3K9me3 in this process (Fig. 3j). We also note that proteins that are responsible for telomere perinuclear binding, such as membrane-anchored junction protein and telomere repeat-binding bouquet formation protein 1, are lowly expressed in gonadal premeiotic PGCs52 (Supplementary Table 3). Collectively, this highlights the necessity to revisit the current model. In the male germline, once established, perinuclear centromere anchoring appears to be a stable feature, persisting from late embryonic stages (E18.5) into the postnatal testis, where it is maintained in undifferentiated spermatogonia (P10) (Extended Data Figs. 1a,b and 3f). This continuous nuclear organization may support the eventual transition to meiosis and underscores the importance of early chromatin reprogramming in germ cell development.
Achieving full recapitulation of meiosis progression remains a notable hurdle in the context of in vitro gametogenesis. Our study suggests that, despite chromatin decondensation, canonical H3K9me3-enriched chromocenters remain observable and the distribution of centromeres exhibits significant variability among the PGCLC population (Extended Data Fig. 9c), unlike what is observed in gonadal PGCs in vivo. In addition, a direct comparison of genome-wide chromatin interactions between in vivo PGCs and in vitro PGCLCs documented that PGCLCs do not undergo the same degree of genome architectural remodeling as PGCs38. The addition of bone morphogenetic protein 2 and vitamin A to the extended PGCLC culture or reconstitution of the developmental niche using female gonadal tissue can induce the expression of meiosis-related structural proteins and formation of the SC In PGCLCs53,54. However, the resulting in vitro gametogenesis shows low efficiency of homologous chromosome pairing and frequent meiotic defects, pointing toward the existence of an unknown prerequisite that exists in germ cells preceding the meiotic prophase in vivo. Our findings are in direct support of this hypothesis.
In summary, our study revealed striking genome architecture remodeling that occurs at the transition to gametogenesis in mitotic PGCs. In addition to offering an unparalleled model for physiological rewiring of 3D genome organization, our study provides a previously missing understanding regarding the structural rearrangements occurring before meiosis, which will be critical for our attempts to recapitulate meiosis and complete gametogenesis in vitro.
Methods
All experiments performed in this study complied with all relevant ethical regulations. All animal experiments were approved by the UK Home Office.
Mice
All animal experiments were approved by the UK Home Office under project license PPL pp1838178 and followed all relevant guidelines and regulations. Mouse rooms have a 12-h light–dark cycle and the room temperature was maintained at 20–24 °C. The relative humidity was kept at 45–65%.
For PGC isolation, ΔPE–Oct4–GFP mice (GOF 18∆PE–EGFP)55 were crossed with C57BL/6 mice. The time of mating is determined by the appearance of vaginal plug at noon, which is defined as E0.5. No randomization or blinding was performed. All replicates of samples were processed and analyzed in parallel using the same settings.
Cryosection IF staining
The dissected embryonic trunk or genital ridges from embryos (E11.5–E18.5) were fixed with 2% paraformaldehyde (PFA) for 30 min at 4 °C, washed three times for 10 min in PBS and incubated overnight in 30% sucrose in PBS at 4 °C. The samples were then mounted in OCT mounting medium (VWR) and then snap-frozen in liquid nitrogen. The block was stored at −80 °C. The mounted samples were sectioned for 10 μm using Leica cryostat (Leica, CM1950). The cryosections were postfixed with 2% PFA in PBS for 3 min, washed three times for 5 min in PBS and permeabilized with blocking buffer (1% BSA and 0.1% Triton in PBS). For optimal histone modification staining, the slides underwent mild antigen retrieval by incubation with boiled sodium citrate buffer (10 mM sodium citrate, pH 6.0) at 90 °C for 10 min and then washed with PBS. The primary antibodies were added in blocking buffer and incubated with slides overnight at 4 °C. The slides were subsequently washed three times in blocking buffer and incubated with Alexa dye-conjugated secondary antibodies (Molecular Probes) in blocking buffer for 1 h at room temperature in the dark. Then, the slides were washed twice for 5 min in blocking buffer and once for 5 min in PBS. Finally, the slides were treated with DAPI (0.1 µg ml−1) for 20 min, mounted in Vectashield (Vector Laboratories) and imaged using a Leica SP5 confocal microscope (Supplementary Table 4).
DNA-FISH combined with IF
Cryosection and IF staining were performed as described above. After washing with PBS, the slide was postfixed with 4% formaldehyde in PBS for 10 min at room temperature and then washed three times for 5 min in PBS. The slides were dehydrated by incubating with 70%, 80%, 90% and 100% ethanol for 5 min each. DNA-FISH probes were diluted in 70% formamide, 10 mM Tris pH 7.2 and 0.5% blocking reagent (Roche, 11096176001). The mixture was applied to each slide, followed by codenaturation with the sections for 3 min at 80 °C on the heating block. After 24 h of hybridization at 37 °C in the dark, slides were washed twice for 15 min in 70% formamide, 20 mM Tris pH 7.4, followed by three washes of 5 min in 100 mM Tris pH 7.4, 150 mM NaCl and 0.08% Tween-20. Finally, the slides were treated with DAPI (0.1 µg ml−1) for 20 min, mounted in Vectashield (Vector Laboratories) and imaged using a Leica SP5 confocal microscope. Major satellite DNA-FISH probes were prepared using a FISH tag DNA green kit (F32947) with pγsat plasmid (Addgene, 39238) following the manufacturer’s instruction. Telomere and centromere PNA DNA-FISH probes were purchased from Panagene (F1002 and F3006).
RNA-FISH combined with IF
RNA-FISH probes were designed using the Stellaris website (https://www.biosearchtech.com/support/tools/design-software). The RNA-FISH experiment was conducted following the manufacturer’s instructions (Stellaris RNA-FISH) with minor modifications56. The mounted samples were sectioned into 10 μm using a Leica cryostat (Leica, CM1950). After immunostaining, the cryosections were postfixed with 2% PFA in PBS for 2 min and washed twice for 5 min in PBS. The slides were permeabilized with blocking buffer with the RNase inhibitor (1% BSA, 0.1% Triton and 40 U per μl of Protector RNase inhibitor (Roche, RNAINH-RO) in PBS). For the negative control, the slides were incubated with 1 μg μl−1 RNase A (Invitrogen, 12091021) in PBS for 1 h at 37 °C. The primary antibodies were then added in blocking buffer and incubated with the slide for 30 min at room temperature. The slides were subsequently washed three times in blocking buffer and incubated with Alexa dye-conjugated secondary antibodies (Molecular Probes) in blocking buffer for 30 min at room temperature in the dark. Then, the slides were washed twice for 5 min in blocking buffer and once for 5 min in PBS. The slides were postfixed with 2% PFA in PBS for 10 min and washed twice for 5 min in PBS. The slides were immersed in 70% (v/v) ethanol for at least 1 h at room temperature and then washed with RNA-FISH wash buffer A (Biosearch Technologies, SMF-WA1-60) for 5 min. The slides were then incubated with the RNA-FISH probes diluted in 125 nM hybridization buffer (Biosearch Technologies, SMF-HB1-10) in a humified chamber in the dark for 16 h at 37 °C. After probe hybridization, the slides were immersed in RNA-FISH wash buffer A for 30 min at 37 °C. The slides were treated with DAPI (0.1 µg ml−1) for 20 min and then incubated with RNA-FISH wash buffer B (Biosearch Technologies, SMF-WB1-20). Finally, the slide was mounted in Vectashield (Vector Laboratories, H-1000) and imaged using a Leica SP5 confocal microscope. Ten probes were synthesized: 5′-TTTCCACCTTTTTCAGTTTT-3′, 5′-TCCTACAGTGGACATTTCTA-3′, 5′-AGTTTTCTTGCCATATTCCA-3′, 5′-GTTTCTCATTTTCCATGATT-3′, 5′-TTTTTCAAGTCGTCCAGTGG-3′, 5′-ACGTTTTTTAGTGATTTCGT-3′, 5′-TTCAGTGTGCATTTCTCATT-3′, 5′-TTTTCTCGCCATATTCCAGG-3′, 5′-TCTCATTTTCCGTGATTTTC-3′ and 5′-TTCACGTCCTAAAGTGTGTA-3′.
Western blot for PGCs
PGCs were isolated as previously described6. The dissected genital ridges were digested for 5 min at 37 °C using 0.05% trypsin–EDTA (Gibco, 25300054) or TrypLE Express (Gibco, 12563011). Enzymatic digestion was followed by neutralization with prewarmed DMEM (Gibco, 21969-035), containing 10% FBS (Gibco, 16000-044) and pipetting up and down 20–30 times, followed by centrifugation at 400g for 3 min. Cells were subsequently resuspended in DMEM/F12 supplemented with hyaluronidase (300 μg ml−1; Sigma, H3506) by pipetting up and down ten times to generate single-cell suspensions. Following centrifugation at 400g for 3 min, the cell pellet was resuspended in ice-cold PBS supplemented with polyvinyl alcohol (10 μg ml−1; Sigma-Aldrich, 341584) and EGTA (0.4 mg ml−1; Sigma-Aldrich). GFP-positive cells were isolated by fluorescence-activated cell sorting (FACS) using an Aria IIu (BD Bioscience) or Aria III (BD Bioscience) flow cytometer. The gating strategy of FACS is provided.
After dissociation, 10,000 Oct4–GFP-positive PGCs and 10,000 OCT4–GFP-negative cells were sorted and centrifuged at 400g for 3 min. Cells were lysed with 10 μl of RIPA buffer (Sigma, RO278) with protease inhibitor cocktail (Roche cOmplete tablets mini, 11836153001). Total lysate was loaded into 12% acrylamide–Bis gel and separated in running buffer (25 mM Tris base, 190 mM glycine and 0.1% sodium dodecyl sulfate (SDS)). The protein was transferred to a nitrocellulose membrane (Amersham) after electrophoresis in transferring buffer (25 mM Tris base, 190 mM glycine, 0.1% SDS and 20% methanol). The membrane was blocked by 5% BSA in PBST (0.1% Tween in PBS) for 30 min at room temperature. Primary antibodies were added in PBST with 5% BSA and incubated with the membrane overnight at 4 °C (GFP, 1,000; CENPA, 1:2,000; H3, 1:5,000). The membrane was washed three times with PBST for 10 min. Horseradish peroxidase (HRP)-conjugated secondary antibodies (1:10,000) were added before incubating for 1 h at room temperature and then washing three times with PBST. Luminata Crescendo western HRP substrate (WBLUR0100, Milipore) was used for detection on an Amersham Imager 680. Quantification of the western blot bands was performed using ImageJ Fiji (1.53c).
Electron microscopy
For transmission electron microscopy morphological analysis, gonads were fixed with 2.5% glutaraldehyde (Sigma, G7776) in H2O for 2 h at room temperature, before rinsing in PBS and H2O. Then gonads were incubated in 2% (w/v) OsO4 (Sigma, 419494) in H2O for 2 h at room temperature and rinsed in distilled water. The specimens were dehydrated in acetone, infiltrated with epoxy resin overnight (Sigma, 31185) and polymerized in gelatin capsules at 60 °C for 48 h. Thin sections (60–70 nm thick) were cut on a Leica UCT ultramicrotome with a diamond knife and collected on 300-mesh nickel grids. Saturated aqueous uranyl acetate and lead citrate was used to stain the grids. An FEI Titan Krios electron microscope operated at 300 kV with high tension was used for the acquisition.
PGCLC differentiation
Blimp1–RFP ES cells, derived from mouse line B6.Cg-Tg(Blimp1–RFP)4Rbrc (RBRC01833)57, were used to generate PGCLCs. ES cells were maintained in 2i medium. Epiblast-like stem cell (EpiLC) and PGCLC induction was performed as previously described58,59. In brief, 0.5 × 105 cells were seeded in one well of a fibronectin-coated 12-well plate with EpiLC differentiation medium (N2B27 medium supplemented with 12 ng ml−1 basic fibroblast growth factor (Thermo Fisher, 13256-029), 20 ng ml−1 activin A (Peprotech, 120-14E-10) and 1% KnockOut SR (Gibco, 10828028). Cells were cultured for 2 days and the medium was changed after 1 day of culture. After 2 days of culture, EpiLCs were dissociated and the cell suspension was diluted with PGCLC differentiation medium (GK15 medium containing 500 ng ml−1 BMP4 (R&D system, 314-BP-500), 100 ng ml−1 mouse stem cell factor (SCF; R&D Systems, 455-MC-010), 1,000 U per ml leukemia inhibitory factor (LIF) and 50 ng ml−1 epidermal growth factor (R&D Systems, 2028-EG-010) to a final concentration of 2 × 104 cells per ml. Then, 100 µl of the cell suspension was pipetted into a 96-well Lipidure-Coat plate (A-U96, NOF) and cultured for 4 days.
To isolate PGCLCd4 from the aggregates, aggregates were transferred and dissociated with TrypLE solution for 7–8 min at 37 °C. The suspension is neutralized with MEF medium with DNase I (Roche, 04716728001). After centrifugation 220g for 5 min, cell pallets were resuspended with FACS buffer. Blimp1–RFP-positive cells were isolated by FACS using an Aria IIu (BD Bioscience) or Aria III (BD Bioscience) flow cytometer (100-μm nozzle).
Sorted cells were centrifuged and plated on m220 feeder-coated eight-well plates (Ibidi). PGCLCs were further cultured for 5–6 days with PGCLC propagation medium (DMEM/F12 with 10% KnockOut SR, 2.5% FBS, 0.1 mM nonessential amino acids, 2 mM glutamine, 0.1 mM β-mercaptoethanol, 100 ng ml−1 SCF, 10 µM forskolin (R&D system, 1099/10), 10 µM rolipram (StemCell Technologies, 73384), penicillin–streptomycin (Gibco, 15140122). All cells were grown at 37 °C under 5% CO2.
Image analysis
CENPA and H3K9me3
The measurements of CENPA foci and H3K9me3 area were conducted using a custom pipeline developed in CellProfiler (version 2.2.0; https://cellprofiler.org/). Briefly, child objects were identified by CENPA foci and the parent objects were identified by DAPI. Germ cells were identified by germ cell markers, such as MVH. The minimum distance between child (CENPA foci) and parent (edited nuclei) objects was calculated after identification. H3K9me3 area was measured for each nucleus and the nuclear periphery area was defined by thresholding of the lamin B1 staining and expanding from the edges of the nuclei objects, followed by subtraction of a shrunken nuclei area. The number of CENPA foci was identified after maximum projection of z-stack images using ImageJ. Three-dimensional confocal z-stack visualizations were produced using Imaris (version 6.1; BitPlane).
Telomere
The quantification of telomere DNA-FISH was conducted using Imaris (versions 6.1 and 10.1) and the ‘Imaris Cell’ package. Telomere DNA-FISH foci were identified as ‘vesicles’, using an estimated focus diameter before performing background intensity subtraction and automated focus detection. Germ cells were identified as MVH-positive cells. The total number of telomere foci and the mean value of vesicle intensity (vesicles spot intensity sum) for each nucleus was measured and calculated.
The closest distance between individual telomere foci and the nuclear membrane was measured (vesicles spot distance to cell membrane). This distance measured was normalized to the radius, which was derived from the nuclear volume of each nucleus. The ratio of the focus number in different bins was calculated after normalization.
RAD21 and CTCF intensity quantification
Nuclei were segmented and labeled image masks were created using Cellpose 2.0 (https://www.cellpose.org/)60,61 with the following parameters: cell diameter, 70; flow threshold, default (0.4); cyto2 model.
After segmentation and export of labeled masks, quantification was performed using a custom pipeline developed in CellProfiler (version 4.0)62. PGCs were identified by the germ cell marker (MVH). The mean intensity within each nucleus was measured for PGCs and somatic cells. The mean and s.d. were calculated for each image. To compare the difference between PGCs and somatic cells, z scores were calculated for the individual nucleus in each image. Mean values of z scores for biological replicates were calculated and are shown in the figures.
Ki67
The focal plane images from every fifth cryosection throughout the entire gonad were acquired using a Zeiss Axio Scan Z1 slide scanner. PGCs and somatic cells were distinguished by MVH expression. Mitotic cells were defined by Ki67 positivity and a chromosome-coating pattern. Three biological replicates from different litters were used.
Inhibitor treatment
Gonads were dissected from E12.5 mouse embryos and male and female gonads were separated on the basis of the gross appearance. The sex was later confirmed by genotyping PCR. Gonads were treated with chaetocin (Sigma, C9492-1MG; 100 nM) and control (DMSO) in FBS-containing ES cell medium in Transwells (Corning, 3413) for 12 h and 24 h, respectively. After treatment, gonads were fixed with 2% FBS medium consisting of DMEM/F12 (Gibco, 2133-020) supplemented with 15% FBS (Gibco, 16000-044), 0.1 mM nonessential amino acids (Gibco, 11140-050), 2 mM l-glutamine (Lonza, BE17605E) and 0.1 mM 2-mercaptoethanol (Gibco, 31350-010). J1 and Dnmt-TKO ES cells63 were cultured in serum-free 2i medium (DMEM/F12, Neurobasal medium (Gibco, 2113-049) containing B27 (Gibco, 17504-044) and N2 supplement (Gibco, 175022-048)) with inhibitors: 1 μM PD0325091 (Cambridge Bioscience,1643-5) and 3 μM CHIR99021 (Cambridge Bioscience, SM13-10). Then, 1,200 U per ml LIF (ESG1107, Sigma) was added without adding any antibiotics. ES cells were grown on 0.1% gelatin-coated plates and passaged every other day. The cell line was verified by genotyping. All cell lines tested negative for Mycoplasma contamination. Chemical inhibitors were dissolved in DMSO. Cells were cultured with inhibitors with final concentration: tryptic soy agar (Sigma-Aldrich, T1952, 1 nm) and SGC0946 (Sigma-Aldrich, SML1107, 2 μM). Vitamin C (Sigma-Aldrich, A8960) was added into the medium with final concentration 100 µg ml−1.
Collection of human embryonic samples
Following written informed consent and anonymization, human embryonic gonads were collected by the University College London Great Ormond Street Institute of Child Health and were obtained from the Human Developmental Biology Resource (HDBR, project 200744; https://www.hdbr.org/). Samples were staged on the basis of HDBR guidelines (https://www.hdbr.org/factsheets). Briefly, to describe the stage of the developing embryo, HDBR uses PCWs. This measurement starts from the moment of fertilization. Notably, there exists an approximate 2-week difference between the number of gestation weeks and the PCWs for an embryo or fetus. All fetal samples in the HDBR collection are staged as PCWs. The samples were destroyed after the experiments.
In situ Hi-C
Hi-C was performed as previously described64. Cells were fixed in 2% PFA for 10 min, lysed in lysis buffer (30 min on ice) and digested with DpnII (New England Biolabs) overnight at 37 °C while rotating (950 rpm). Restriction overhangs were filled-in with Klenow (New England Biolabs) using biotin-14-dATP (Jena Bioscience) and ligation was performed in ligation buffer for 4 h at 16 °C (T4 DNA ligase; Life Technologies). After overnight decrosslinking at 65 °C, the ligated DNA was tagmented to produce fragments of 300–700 bp. Ligation products were isolated using MyOne C1 streptavidin beads (Life Technologies), followed by washing with wash and binding buffer and nuclease-free water. Isolated Hi-C ligation products on the beads were then used directly for PCR amplification and the final Hi-C library was purified with AMPure XP beads (Beckman Coulter).
Hi-C analysis
Hi-C matrix generation
Hi-C reads were trimmed using Trim Galore (version 0.6.6; https://github.com/FelixKrueger/TrimGalore). To obtain data as homogeneous as possible and given that male reads were 100 bp and female reads were 50 bp long, male reads were initially hard-trimmed 50 bp from their 3′ end to get 50-bp reads for both male and female samples. Then, quality trimming and adaptor removal were performed on all reads using trim_galore with default parameters.
Hi-C matrices were obtained using FAN-C (version 0.9.23)65. We used the fanc auto command, establishing the restriction enzyme with -r DpnII and adding a ligation error filter with --le-inward-cutoff and --le-outward-cutoff set to 10 kb. Reads were aligned to the mm39 genome. Normalization of Hi-C matrices was conducted using the Knight–Ruiz balancing method per chromosome. Reproducibility of replicates was assessed by principal component analysis (PCA) using 500-kb-resolution bins derived from all autosomes. After confirming the reproducibility, replicates were merged for further analysis. Similar to Rao et al.66, a maximum resolution of 10 kb was chosen for all merged samples by identifying the smallest bin size at which over 80% of bins contained more than 1,000 contacts.
For the figures displaying multiple chromosomes or sex chromosomes (Fig. 4b and Extended Data Fig. 4c), new matrices were produced for each individual replicate of the female sample excluding chromosome Y from the reference genome before alignment. Replicates were merged. Then, matrices were normalized at the whole-genome level using fanc hic with the following arguments: --normalize, --whole-matrix and --subset (main chromosomes: chr1–chr19 and chrX, plus chrY in males). Publicly available Hi-C datasets24,38 were processed using the same approach. Correlation of insulation scores between Hi-C data from PGCs and Hi-C data from in vitro gametogenesis38 was calculated using the Pearson correlation coefficient comparing insulation scores at 300-kb Hi-C matrices.
TAD boundaries
TAD boundaries were calculated on the basis of the insulation score using FAN-C (version 0.9.23). Insulation was calculated using different bin sizes (for example, 100 kb, 300 kb and 500 kb) and TAD boundaries were initially calculated for each sample at different bin sizes. Boundaries overlapping the ENCODE blacklist67 were removed. After careful examination of the results, 300-kb bins and a minimum score of 0.3 in at least one of the samples were selected. To create a merged list of TAD boundaries across all four cell types (PGC/soma and male/female), boundaries at the same bin or less than two bins away were considered the same. The strength of each boundary was compared between PGCs and somatic cells and boundaries were classified into soma-specific (n = 747 in female, n = 938 in male), shared (stronger in soma) (n = 3951 in female, n = 3914 in male), shared (stronger in PGCs) (n = 406 in female, n = 846 in male) and PGC-specific (n = 408 in female, n = 553 in male). Visual inspection of the four groups of boundaries revealed that PGC-specific boundaries were mainly false positives or lifted boundaries and we focused downstream analyses on the soma-specific or shared (stronger in soma) categories, which encompassed the majority of boundaries in both female and male.
Shared boundaries with a score stronger in soma were further classified into two groups reflecting a mild or severe disruption of the boundary in PGC, relative to soma. For this classification, the boundary score in PGCs and soma were compared; if (score in PGC)/(score in soma) < 0.5, then boundaries were classified as severe loss, whereas, if (score in PGC)/(score in soma) > 0.5, they were classified as mild loss. Previously identified TADs in mouse ES cells were used to compare TAD strength in somatic cells and PGCs18 (Fig. 6b). TADs in sex chromosomes were excluded. TAD strength was obtained with the FAN-C fanc aggregate command with the --tads and --tad-strength arguments.
k-means clustering
An unsupervised classification of boundaries was conducted by k-means clustering of the change in insulation calculated as the log2(insulation in PGC/insulation in soma), using insulation scores calculated at 30-kb bins and a window of 400 kb around each TAD boundary. Then, k-means clustering was performed using deepTools (version 3.5.5). A total of four clusters were selected after calculating up to ten clusters, as the four clusters were conserved across sexes and not redundant.
Identification of compartments
Enrichment profiles (saddle plots) and compartment strength were calculated using FAN-C considering autosomes exclusively and on Hi-C matrices binned at 500-kb bins (Fig. 5a,b). The strength of compartment interactions between same or different types of compartments was also calculated for each individual autosome after subsetting each autosome from the whole-genome Hi-C matrix (Fig. 5c).
Identification of loops
Loops were called on merged Hi-C matrices at 10-kb resolution using Mustache68 with the following arguments: --sparsityThreshold 0.9, --resolution 10 kb, --pThreshold 0.01, --sigmaZero 1.6. Loops with one or both anchors overlapping the ENCODE blacklist67 were removed. Because false loops were called at highly repetitive regions, we used genome mappability tracks for the mm39 genome from https://bismap.hoffmanlab.org/ (k36.merged.GRCm39_umap.bed)69 to filter out loops with one or both anchors overlapping regions with <90% mappability.
METALoci
To visualize Hi-C interaction patterns in regions of interest (Fig. 6f), we used a publicly available METALoci method (https://github.com/3DGenomes/METALoci/)19 to generate two-dimensional (2D) graph-based representations. The model was run on FAN-C hic files at 10-kb resolution, with a significance cutoff for interactions set to 1 and a smoothing parameter of 1. We then overlaid the corresponding ATAC-seq signal and TAD boundary annotations onto the METALoci 2D Gaudi plots to examine spatial organization and interactions.
Single-cell ATAC-seq and motif analysis
Single-cell ATAC-seq data processing
Single-cell ATAC-seq reads for mouse gonad were retrieved from a previous study27, processed and aligned to the mm39 genome using cellranger-atac-1.2.0. Using the cell type identities assigned to each cell in the publication of origin, the resulting BAM files were subset using subset-bam to get separate files with E13.5 somatic or PGC reads. In female, for the somatic BAM file, reads assigned the following identities were selected: pregranulosa cell, stromal progenitor cell, mesothelial cell, endothelial cell, erythroblast and macrophage. In male, for the somatic BAM file, reads assigned the following identities were selected: Sertoli cell, presupporting cell, stromal progenitor cell, mesothelial cell, fetal Leydig cell, endothelial cell, erythroblast and macrophage. For the PGC BAM file, reads with PGC identity (and meiotic germ cells in females) were selected.
Transcription factor footprinting
CTCF footprinting analysis was performed using TOBIAS (version 0.15.1)70 Briefly, ATACorrect was used on somatic and PGC BAM files, as described above, to correct the cutsite signal with regard to Tn5 transposase biases. BINDetect was used to predict transcription-factor-binding sites using the JASPAR 2022 CORE vertebrate collection of nonredundant position frequency matrices71. Aggregate ATAC-seq signals or footprints were calculated using PlotAggregate at CTCF (CTCF_MA0139.1) motifs after classifying the motifs by their overlap with stable, intermediate or lost TAD boundaries. Aggregate signals were smoothed for plotting using the smoothing function from the modelbased R package (https://easystats.github.io/modelbased/).
CTCF motif score
A CTCF motif score was assigned to TAD boundaries as follows. First, CTCF (CTCF_MA0139.1)-binding sites were retrieved from the TOBIAS BINDetect step of the footprinting analysis. Then, the CTCF-binding site score (TFBS_score column of BINDetect output) was retrieved for all CTCF-binding sites overlapping with each TAD boundary. Lastly, the maximum binding site score was assigned to each TAD boundary.
RNA-seq
Soma versus PGC gene expression
To compare gene expression levels between somatic cells and PGCs, scRNA-seq data were retrieved from a previous study27. Reads were processed and aligned to the mm39 genome using cellranger-7.2.0. Reads from somatic cells and PGC from the E13.5 embryo were then subset from the resulting alignments as for single-cell ATAC-seq. Coverage tracks were obtained with SAMtools bamCoverage, normalizing using BPM while ignoring sex chromosomes.
Gene expression and boundary score
To calculate the strength of boundaries in relation to genes with different levels of expression (Extended Data Fig. 5d), genes were first split into five equally sized groups of genes on the basis of their levels of expression in E13.5 PGCs. Transcripts per million values were acquired from the previously published RNA-seq data6. Bins in female samples contained genes with expression levels between the following ranges: bin 1, 0–0.0199; bin 2, 0.0199–0.298; bin 3, 0.298–2.98; bin 4, 2.98–28.1; bin 5, 28.1–1.26 × 104. In male, genes were split within the following expression ranges: bin 1, 0–0.0245; bin 2, 0. 0245–0.26; bin 3, 0.26–2.46; bin 4, 2.46–26.4; bin 5, 26.4–1.6 × 104. Then, the nearest TAD boundary was assigned to each gene and its strength was retrieved and grouped on the basis of the gene’s expression bin. Lastly, TAD strengths were compared using Student’s t-test for all pairs of bins.
GO enrichment analysis
Genes that overlapped with TAD boundaries with a minimum overlap of 1 bp were identified and GO analysis was performed with clusterProfiler (version 4.12.0)72,73. For ‘all’ GO terms, we applied pAdjustMethod = BH and no P-value or q-value thresholds (pvalueCutoff and qvalueCutoff = 1.1). Then, the top ten most enriched GO terms were retrieved for each TAD boundary group. The adjusted P values of all resulting GO terms in all boundary groups are shown for comparison (Extended Data Fig. 6h).
ChIP-seq
H3K27me3 ChIP-seq data from a previous publication were retrieved (GSE141182)6. Quality trimming was applied using trimGalore with default parameters, plus the --paired setting. Alignment to mm39 was performed with bowtie2 (version 2.4.4) using the following parameters: --end-to-end --very-sensitive --no-mixed --no-discordant -k 10. Nonprimary alignments were filtered out using SAMtools view -f 0×02 -F 0×900 -F 0×4 (SAMtools, version 1.14). PCR duplicates were removed using sambamba (version 0.8.2). Coverage tracks were obtained with SAMtools bamCoverage, normalizing using RPGC and ignoring sex chromosomes. The ENCODE blacklist67 was also excluded at this step. Heat maps combining insulation, scATAC-seq, scRNA-seq and ChIP-seq signal at TAD boundaries were generated using the EnrichedHeatmap R package (version 1.34.0)74.
Statistical analysis
All statistical analyses were performed using R or GraphPad software and are described in the figure legends. Biological replicates for all experiments were based on embryos from independent litters. No statistical methods were used to predetermine sample size. P values were calculated using a two-tailed Mann–Whitney U-test (Figs. 1d,g, 2b,c, 3b,f,j and 6h and Extended Data Figs. 3h, 6j, 8a,b and 9c), paired Wilcoxon signed-rank test (Figs. 4f and 5c), two-sided Student’s t-test (Fig. 6c, paired; Fig. 6j, not paired), one-way ANOVA with Tukey correction (Extended Data Figs. 5d, 6d and 10d). Enrichment of GO terms in genes located at TAD boundaries (Extended Data Fig. 7g) was calculated using clusterprofiler, with Benjamini–Hochberg correction for multiple testing. Insulation scores were compared across multiple samples (Extended Data Fig. 10a) by calculating Pearson correlation for all pairs of samples.
Box plots (Figs. 1d, 4c,f, 5c and 6c,j and Extended Data Figs. 1d, 3h, 5d, 6d and 10d) were plotted using the Tukey method. The upper and lower hinges represent the first and the third quartiles. The central line represents the median. The upper end of the whisker represents the lowest value from either the third quartile plus 1.5× the interquartile range (IQR) or the maximum value from the dataset. The lower end of the whisker represents the largest value from either the first quartile minus 1.5× the IQR or the minimum value from the dataset. Box plots were generated using the R package ggplot2 (with argument outlier.shape = NA; Extended Data Figs. 6d and 10d).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The Hi-C data produced in this study were submitted to the Gene Expression Omnibus and are available under accession number GSE272501. In addition, we analyzed data from the following publicly available datasets: J1 and Dnmt-TKO ES cells, GSE138102; PGCLCs, GSCs and GSLCLCs, GSE184129; RNA-seq of E10.4 and E14.5 germ cells, GSE76973; RNA-seq and ChIP-seq of E13.5 PGCs, GSE141182; and scRNA-seq and ATAC-seq of PGCs, CRA003910. Source data are provided with this paper.
Code availability
All analyses were conducted using published packages. No custom code was created. All computational analysis was described in Methods and code is also available from GitHub (https://github.com/vaquerizaslab/Huang_Rigau_et_al-NSMB-PGC_3D_genome).
References
Shirane, K. & Lorincz, M. Epigenetic mechanisms governing female and male germline development in mammals. Sex Dev. 16, 365–387 (2022).
Seki, Y. et al. Cellular dynamics associated with the genome-wide epigenetic reprogramming in migrating primordial germ cells in mice. Development 134, 2627–2638 (2007).
Daujat, S. et al. H3K64 trimethylation marks heterochromatin and is dynamically remodeled during developmental reprogramming. Nat. Struct. Mol. Biol. 16, 777–781 (2009).
Hajkova, P. et al. Chromatin dynamics during epigenetic reprogramming in the mouse germ line. Nature 452, 877–881 (2008).
Hill, P. W. S. et al. Epigenetic reprogramming enables the transition from primordial germ cell to gonocyte. Nature 555, 392–396 (2018).
Huang, T. C. et al. Sex-specific chromatin remodelling safeguards transcription in germ cells. Nature 600, 737–742 (2021).
Saitou, M. & Hayashi, K. Mammalian in vitro gametogenesis. Science 374, eaaz6830 (2021).
Hajkova, P. Epigenetic reprogramming in the germline: towards the ground state of the epigenome. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366, 2266–2273 (2011).
van Steensel, B. & Belmont, A. S. Lamina-associated domains: links with chromosome architecture, heterochromatin, and gene repression. Cell 169, 780–791 (2017).
Magaraki, A. et al. Silencing markers are retained on pericentric heterochromatin during murine primordial germ cell development. Epigenetics Chromatin 10, 11 (2017).
Schonenberger, F., Deutzmann, A., Ferrando-May, E. & Merhof, D. Discrimination of cell cycle phases in PCNA-immunolabeled cells. BMC Bioinformatics 16, 180 (2015).
Burton, A. et al. Heterochromatin establishment during early mammalian development is regulated by pericentromeric RNA and characterized by non-repressive H3K9me3. Nat. Cell Biol. 22, 767–778 (2020).
Johnson, W. L. et al. RNA-dependent stabilization of SUV39H1 at constitutive heterochromatin. eLife 6, e25299 (2017).
Shirai, A. et al. Impact of nucleic acid and methylated H3K9 binding activities of Suv39h1 on its heterochromatin assembly. eLife 6, e25317 (2017).
Velazquez Camacho, O. et al. Major satellite repeat RNA stabilize heterochromatin retention of Suv39h enzymes by RNA–nucleosome association and RNA:DNA hybrid formation. eLife 6, e25293 (2017).
Capelson, M. et al. Chromatin-bound nuclear pore components regulate gene expression in higher eukaryotes. Cell 140, 372–383 (2010).
Gonzalez-Sandoval, A. et al. Perinuclear anchoring of H3K9-methylated chromatin stabilizes induced cell fate in C. elegans embryos. Cell 163, 1333–1347 (2015).
Bonev, B. et al. Multiscale 3D genome rewiring during mouse neural development. Cell 171, 557–572 (2017).
Mota-Gómez, I. et al. Sex-determining 3D regulatory hubs revealed by genome spatial auto-correlation analysis. Preprint at bioRxiv https://doi.org/10.1101/2022.11.18.516861 (2022).
Blaschke, K. et al. Vitamin C induces Tet-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature 500, 222–226 (2013).
Yang, M. et al. Chemical-induced chromatin remodeling reprograms mouse ESCs to totipotent-like stem cells. Cell Stem Cell 29, 400–418 (2022).
Malla, A. B. et al. DOT1L bridges transcription and heterochromatin formation at mammalian pericentromeres. EMBO Rep. 24, e56492 (2023).
Toth, K. F. et al. Trichostatin A-induced histone acetylation causes decondensation of interphase chromatin. J. Cell Sci. 117, 4277–4287 (2004).
Jiang, Q. et al. G9a plays distinct roles in maintaining DNA methylation, retrotransposon silencing, and chromatin looping. Cell Rep. 33, 108315 (2020).
Rao, S. S. P. et al. Cohesin loss eliminates all loop domains. Cell 171, 305–320 (2017).
Nora, E. P. et al. Targeted degradation of CTCF decouples local insulation of chromosome domains from genomic compartmentalization. Cell 169, 930–944 (2017).
Jiang, S. et al. Single-cell chromatin accessibility and transcriptome atlas of mouse embryos. Cell Rep. 42, 112210 (2023).
Nagano, T. et al. Cell-cycle dynamics of chromosomal organization at single-cell resolution. Nature 547, 61–67 (2017).
Alavattam, K. G. et al. Attenuated chromatin compartmentalization in meiosis and its maturation in sperm development. Nat. Struct. Mol. Biol. 26, 175–184 (2019).
Patel, L. et al. Dynamic reorganization of the genome shapes the recombination landscape in meiotic prophase. Nat. Struct. Mol. Biol. 26, 164–174 (2019).
Western, P. S., Miles, D. C., van den Bergen, J. A., Burton, M. & Sinclair, A. H. Dynamic regulation of mitotic arrest in fetal male germ cells. Stem Cells 26, 339–347 (2008).
Kagiwada, S., Kurimoto, K., Hirota, T., Yamaji, M. & Saitou, M. Replication-coupled passive DNA demethylation for the erasure of genome imprints in mice. EMBO J. 32, 340–353 (2013).
Ter Huurne, M., Chappell, J., Dalton, S. & Stunnenberg, H. G. Distinct cell-cycle control in two different states of mouse pluripotency. Cell Stem Cell 21, 449–455 (2017).
Soygur, B. et al. Intercellular bridges coordinate the transition from pluripotency to meiosis in mouse fetal oocytes. Sci. Adv. 7, eabc6747 (2021).
Hayashi, K. et al. Offspring from oocytes derived from in vitro primordial germ cell-like cells in mice. Science 338, 971–975 (2012).
Hayashi, K., Ohta, H., Kurimoto, K., Aramaki, S. & Saitou, M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell 146, 519–532 (2011).
Kurimoto, K. et al. Quantitative dynamics of chromatin remodeling during germ cell specification from mouse embryonic stem cells. Cell Stem Cell 16, 517–532 (2015).
Nagano, M. et al. Nucleome programming is required for the foundation of totipotency in mammalian germline development. EMBO J. 41, e110600 (2022).
Li, L. et al. Single-cell RNA-seq analysis maps development of human germline cells and gonadal niche interactions. Cell Stem Cell 20, 891–892 (2017).
Seisenberger, S. et al. The dynamics of genome-wide DNA methylation reprogramming in mouse primordial germ cells. Mol. Cell 48, 849–862 (2012).
Hsieh, T. S. et al. Enhancer–promoter interactions and transcription are largely maintained upon acute loss of CTCF, cohesin, WAPL or YY1. Nat. Genet. 54, 1919–1932 (2022).
Ur, S. N. & Corbett, K. D. Architecture and dynamics of meiotic chromosomes. Annu. Rev. Genet. 55, 497–526 (2021).
Kitamura, Y. et al. CTCF-mediated 3D chromatin sets up the gene expression program in the male germline. Nat. Struct. Mol. Biol. 32, 1227–1240 (2025).
Dekker, J. & Heard, E. Structural and functional diversity of topologically associating domains. FEBS Lett. 589, 2877–2884 (2015).
Acemel, R. D. & Lupianez, D. G. Evolution of 3D chromatin organization at different scales. Curr. Opin. Genet. Dev. 78, 102019 (2023).
Rubin, T. et al. Premeiotic pairing of homologous chromosomes during Drosophila male meiosis. Proc. Natl Acad. Sci. USA 119, e2207660119 (2022).
Bowles, J. et al. Retinoid signaling determines germ cell fate in mice. Science 312, 596–600 (2006).
Koubova, J. et al. Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc. Natl Acad. Sci. USA 103, 2474–2479 (2006).
MacLean, G., Li, H., Metzger, D., Chambon, P. & Petkovich, M. Apoptotic extinction of germ cells in testes of Cyp26b1 knockout mice. Endocrinology 148, 4560–4567 (2007).
Trautmann, E. et al. Retinoic acid prevents germ cell mitotic arrest in mouse fetal testes. Cell Cycle 7, 656–664 (2008).
Shibuya, H. & Watanabe, Y. The meiosis-specific modification of mammalian telomeres. Cell Cycle 13, 2024–2028 (2014).
Shibuya et al. MAJIN links telomeric DNA to the nuclear membrane by exchanging telomere cap. Cell 163, 1252–1266 (2015).
Hamada, N. et al. Germ cell-intrinsic effects of sex chromosomes on early oocyte differentiation in mice. PLoS Genet. 16, e1008676 (2020).
Miyauchi, H. et al. Bone morphogenetic protein and retinoic acid synergistically specify female germ-cell fate in mice. EMBO J. 36, 3100–3119 (2017).
Hajkova, P. et al. Epigenetic reprogramming in mouse primordial germ cells. Mech. Dev. 117, 15–23 (2002).
Raj, A., van den Bogaard, P., Rifkin, S. A., van Oudenaarden, A. & Tyagi, S. Imaging individual mRNA molecules using multiple singly labeled probes. Nat. Methods 5, 877–879 (2008).
Sugimoto, M. & Abe, K. X chromosome reactivation initiates in nascent primordial germ cells in mice. PLoS Genet. 3, e116 (2007).
Hayashi, K. & Saitou, M. Generation of eggs from mouse embryonic stem cells and induced pluripotent stem cells. Nat. Protoc. 8, 1513–1524 (2013).
Ohta, H. et al. In vitro expansion of mouse primordial germ cell-like cells recapitulates an epigenetic blank slate. EMBO J. 36, 1888–1907 (2017).
Stringer, C., Wang, T., Michaelos, M. & Pachitariu, M. Cellpose: a generalist algorithm for cellular segmentation. Nat. Methods 18, 100–106 (2021).
Pachitariu, M. & Stringer, C. Cellpose 2.0: how to train your own model. Nat. Methods 19, 1634–1641 (2022).
Stirling, D. R. et al. CellProfiler 4: improvements in speed, utility and usability. BMC Bioinformatics 22, 433 (2021).
Tsumura, A. et al. Maintenance of self-renewal ability of mouse embryonic stem cells in the absence of DNA methyltransferases Dnmt1, Dnmt3a and Dnmt3b. Genes Cells 11, 805–814 (2006).
Ho, J. S. Y. et al. TOP1 inhibition therapy protects against SARS-CoV-2-induced lethal inflammation. Cell 184, 2618–2632 (2021).
Kruse, K., Hug, C. B. & Vaquerizas, J. M. FAN-C: a feature-rich framework for the analysis and visualisation of chromosome conformation capture data. Genome Biol. 21, 303 (2020).
Sanborn, A. L. et al. Chromatin extrusion explains key features of loop and domain formation in wild-type and engineered genomes. Proc. Natl Acad. Sci. USA 112, E6456–E6465 (2015).
Amemiya, H. M., Kundaje, A. & Boyle, A. P. The ENCODE blacklist: identification of problematic regions of the genome. Sci. Rep. 9, 9354 (2019).
Roayaei Ardakany, A., Gezer, H. T., Lonardi, S. & Ay, F. Mustache: multi-scale detection of chromatin loops from Hi-C and Micro-C maps using scale-space representation. Genome Biol. 21, 256 (2020).
Karimzadeh, M., Ernst, C., Kundaje, A. & Hoffman, M. M. Umap and Bismap: quantifying genome and methylome mappability. Nucleic Acids Res. 46, e120 (2018).
Bentsen, M. et al. ATAC-seq footprinting unravels kinetics of transcription factor binding during zygotic genome activation. Nat. Commun. 11, 4267 (2020).
Rauluseviciute, I. et al. JASPAR 2024: 20th anniversary of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 52, D174–D182 (2024).
Wu, T. et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation (Camb.) 2, 100141 (2021).
Yu, G., Wang, L. G., Han, Y. & He, Q. Y. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS 16, 284–287 (2012).
Gu, Z., Eils, R., Schlesner, M. & Ishaque, N. EnrichedHeatmap: an R/Bioconductor package for comprehensive visualization of genomic signal associations. BMC Genomics 19, 234 (2018).
Acknowledgements
We thank J. Elliot, T. Adejumo and B. Patel for help with FACS, L. Game and I. Andrew for help with next-generation sequencing and members of the P.H. and J.M.V. laboratory for discussions and revisions of the paper. Work in the P.H. laboratory is supported by Medical Research Council funding (MC_US_A652_5PY70) and a European Research Council grant (ERC-CoG-648879, dynamic modifications) to P.H. Work in the J.M.V. laboratory is supported by the Medical Research Council (MC_UP_1605/10 to J.M.V.), the Academy of Medical Sciences and the Department of Business, Energy and Industrial Strategy (APR3\1017 to J.M.V.). The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
T.C.H. and P.H. conceptualized the study. T.C.H. performed the experiments and analyzed the data. T.C.H. and V.M. carried out the Hi-C experiments. M.R. analyzed the Hi-C, transcriptomic and epigenomic data. S.S. carried out the electron microscopy. J.L. carried out the METALoci modeling. I.B.B. and C.D. processed the human gonad samples. C.W. facilitated the image analysis. M.S. and P.A. facilitated the data analysis. T.C.H., M.R., J.M.V. and P.H. wrote the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Structural & Molecular Biology thanks Déborah Bourc’his and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available. Primary Handling Editors: Carolina Perdigoto and Dimitris Typas, in collaboration with the Nature Structural & Molecular Biology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Nuclear distribution of chromosome ends.
Nuclear distribution of CENPA using IF staining of E18.5 embryonic gonadal sections (a) and postnatal P10 testis sections (b). Biological replicates N = 3. MVH: germ cell marker. DNA stained by DAPI (blue). Scale bar: 10 µm. c, Representative IF image of max projection from Z-stack confocal images. Yellow arrowheads indicate PGCs. Scale bar: 10 µm. Representative of 3 independent experiments (3 embryos from independent litters). d, Distribution of telomere foci in the nuclei. The shortest distance was measured between telomere foci to the nuclear periphery based on 3D reconstructed images. The distance was normalized to the radius, which was calculated from the nuclear volume. The distance of each focus was classified into 5 bins where Bin 1 is the closest bin to the nuclear periphery. Each dot represents an individual cell. N = 3 embryos analysed with 3 independent experiments. Boxplot central lines represent median values, upper and lower limits show 75th and 25th percentiles, box heights represent interquartile ranges (IQRs) and whiskers are 1.5× the IQR.
Extended Data Fig. 2 Heterochromatin-associated histone marks in germ cells.
Representative IF image of a, H3K9me2. b, H4K20me3. c, H3K9me3 at different embryonic stages and in postnatal testis and ovary (d). e, Representative images showing PCNA distribution during early and late S phase. Biological replicates N = 3. MVH or OCT4: germ cell marker. Yellow arrowheads indicate PGCs. DNA stained by DAPI (blue). Scale bar: 10 µm. In male germ cells at E18.5, the H3K9me3 pattern transitions from a nuclear punctate arrangement to a more dispersed distribution. In female germ cells at E18.5, the H3K9me3 pattern is dispersed with enrichment at the nuclear periphery, potentially resulting from bouquet formation.
Extended Data Fig. 3 Heterochromatin-associated proteins in germ cells.
a, Representative IF image showing downregulation of Hp1 α. Scale bar: 10 µm. b, Comparative gene expression of of Hp1 α(Cbx5), HP1β (Cbx1), HP1Ƴ (Cbx3) using published scRNA-seq data26. c, ATRX is retained in the mitotic chromosomes in PGCs. Scale bar: 10 µm. d, Representative cropped transmission electronic microscope image highlighting nuclear pores and perinuclear heterochromatin. Scale bar: 100 nm. e, Representative IF image of LAMIN A/C. Scale bar: 10 µm. f, Representative IF image of LBR. Yellow arrowheads indicate germ cells. White arrowheads indicate surrounding somatic cells. Scale bar: 10 µm. g, Representative IF image of H3K9me3 after Chaetocin treatment. Scale bar: 5 µm. h, Gonads were cultured with Chaetocin for 24 hours. Scale bar: 5 µm. Measurement of H3K9me3 intensity. Mean value of each nucleus was calculated. n represents the cell number analyzed. P values were calculated by 2 tailed Mann-Whitney U test. Box plots were plotted using Tukey’s method. The median and the first and third quartiles are indicated. The whiskers represent the furthest value within 1.5 × interquartile range (IQR). (a, c, d, e, f, g) Representative of 3 independent experiments (3 embryos from independent litters).
Extended Data Fig. 4 Inter-chromosomal interaction changes in PGCs.
a, PCA plot showing correlations between biological replicates for somatic cells and PGCs. Rep: biological replicates. b, Bar chart showing the fraction of Cis(intra) contacts (Cis/(Cis+Trans)) derived from all autosomes. c, Hi-C matrices showing chromatin interactions of all chromosomes in somatic cells and PGCs. The heatmap on the right illustrates the log2-transformed ratio of contacts between PGC and Soma, where red color indicates an enrichment of contacts PGC and blue a depletion. X chromosome exhibits relatively depleted intra-chromosomal interactions, potentially resulting from X chromosome reactivation in the female germ cells. Hi-C matrices at 1 Mb resolution were normalised at whole-genome level. The matrix comparison was based on observed/expected matrices. Chr: chromosome. d, Total compartment strength of somatic cells and PGCs for biological replicates. e, Saddle plots showing compartment interactions for biological replicates.
Extended Data Fig. 5 Insulation and TADs are diminished in both male and female germ cells.
a, Heatmaps showing normalized Hi-C interaction frequencies (102-104 Mb, chromosome 5) in somatic cells and PGCs, male and female, respectively. The heatmap on the right illustrates the log2-transformed ratio contacts between PGC and Soma, where red color indicates an enrichment of contacts in PGC and blue a depletion. This comparison was based on observed/ expected (O/E) matrices. b, TAD aggregated plots showing diminished TAD structure in individual biological replicates from male and female, somatic cells and PGCs. Numbers on the top-right indicate the median TAD strength. c, Heat maps illustrating insulation profiles and corresponding transcriptional activity in somatic cells and PGCs. Each row represents an identified boundary, ranked according to the extent of insulation strength loss between somatic and germ cells. Transcription levels were obtained from published scRNA-seq data26 d, Strength of boundaries overlapping with genes with different levels of expression in PGCs. Each bin contains one fifth of genes sorted by the expression level, from lowest to highest (Methods). Bin 1 (female n = 1306, male n = 1446), Bin 2 (female n = 1848, male n = 1908), Bin 3 (female n = 1997, male n = 1988), Bin 4 (female n = 2652, male n = 2649). Bin 5 (female n = 2681, male n = 2707). Box plots were plotted using Tukey’s method. The median and the first and third quartiles are indicated. The whiskers represent the furthest value within 1.5 × interquartile range (IQR). P values were calculated one-way ANOVA with Tukey correction (all P values can be found in the Source Data). e, Heatmaps showing four clusters by k-mean clustering based on the differential insulation between somatic cells and PGCs. RPGC: Reads Per Genomic Content. Bar plots showing the proportions of different types of compartments in each cluster. Insulation is greatly reduced across both A and B compartments in PGCs.H3K27me3 and H3K9me3 ChIP-seq data are from previously published study6.
Extended Data Fig. 6 Epigenetic perturbation does not result in drastic genome organization changes in embryonic stem cells.
a, IF staining of CENPA and H3K9me3 in J1 and Dnmt TKO ES cells. HDAC inhibitor: Trichostatin A (TSA). DOT1L inhibitor: SGC0946. Scale bar: 10 µm. b, CENPA distribution in J1 ES cells after a prolonged culture with HDAC inhibitor or vitamin C. DNA stained by DAPI (blue). Scale bar: 10 µm. (a, b) Representative of 3 independent experiments. c, Aggregate TAD plot showing normalized observed/expected Hi-C counts in J1, Dnmt TKO ES cells and PGCs. d, Box plot showing TAD strength in J1 ESCs, Dnmt TKO ESCs, PGCs and somatic cells. Boxplot central lines represent median values, upper and lower limits show 75th and 25th percentiles, box heights represent interquartile ranges (IQRs) and whiskers are 1.5× the IQR. For each sample, the TAD strength was calculated based previously defined TADs18. TAD number n = 2822. n.s.: not significant. ***: P < 0.001. P values were calculated one-way ANOVA with Tukey correction (all P values can be found in the Source Data). e, Saddle plots showing compartment interactions of control cell line J1 and Dnmt TKO ES cells. Hi-C data obtained from a published study24. f, Dynamic changes of gene expression levels in germ cells across developmental stages from bulk RNA-seq data5. TPM: transcript per million. g, Comparative gene expression profile using published scRNA-seq data27. h, Western blot analysis of CTCF. 10000 GFP positive (PGC) and negative (Soma) cells were fluorescence-activated cell (FAC) sorted from gonads of the Oct4-GFP mice (GOF 18ΔPE-EGFP). N = 2 animals, representative of 2 independent experiments. i, Representative IF image of CTCF from P10 testis. Yellow arrowheads indicate germ cells. White arrowheads indicate surrounding somatic cells. Scale bar: 10 µm. Representative of 3 independent experiments. j, RAD21 is highly expressed in germ cells. DNA stained by DAPI (blue). Scale bar: 10 µm. Quantitative analysis of RAD21 abundance in PGCs and somatic cells. Grey dot: each cell. Color dot: mean values from each biological replicate. 3 embryos analysed in 3 independent experiments (mean ± s.d.). P values were calculated by 2 tailed Mann-Whitney U test.
Extended Data Fig. 7 TAD boundaries.
a, Scatter plots showing the identified TAD boundaries (boundary strength > 0.3. Methods.) and their boundary score/strength in somatic cells and PGCs. b, Examples of PGC-specific and soma specific boundaries (grey boxes). Raw and final TAD boundary calls in two selected regions. At the top, heatmaps showing normalized Hi-C interaction frequencies in soma and PGC in somatic and PGC, respectively. Below, tracks representing 1) insulation scores for both soma and PGC, 2) final TAD classified boundaries, 3) raw boundary calls for Soma and PGC before merging and filtering boundaries with low-scores or in low-mappability regions (Methods). PGC-specific boundaries are mostly false positive calls and thus were not considered in further analysis. c, Number of boundaries and fraction per boundary type in females and males, respectively. d, Boundaries were classified based on the degree of strength loss in comparison between somatic cells to PGCs. Shared (stronger in soma) boundaries were classified into two groups: “Mild loss” and “Severe loss”. Soma-specific boundaries were defined as “Total loss” (Methods). e, Heat map of “mild loss”, “severe loss” and “total loss” boundaries. f, Aggregate plot of boundaries. g, Top enriched GO terms from the neighboring genes of mild loss, severe loss and total loss boundaries. P-values obtained performing hypergeometric tests and Benjamini-Hochberg FDR correction.
Extended Data Fig. 8 Germ cells are mitotic at E13.5.
a, Mitotic cells were identified based on the chromosome-coating pattern of Ki67. The percentage of mitotic cells (male: n = 3618, female: n = 3603) was calculated from N = 3 embryos derived from independent litters and analysed with 3 independent experiments (mean ± s.d.). DNA stained by DAPI (blue). Scale bar: 10 µm. b, Cytoplasmic CYCLIN B1 positive germ cells were calculated from gonadal sections from N = 3 embryos analysed with 3 independent experiments (mean ± s.d.). Scale bar: 50 µm, 10 µm. c, Representative image of SCP3 in gonadal sections at E13.5 and d, E16.5. Yellow arrowheads indicate germ cells. SCP3 positive and negative germ cells were counted from N = 3 embryos analysed with 3 independent experiments (mean ± s.d.). Scale bar, 100 µm, 10 µm. (a-b) P values were calculated by 2 tailed Mann-Whitney U test.
Extended Data Fig. 9 Chromatin configuration of embryonic germ cells cannot be recapitulated in PGCLC differentiation.
a, Experimental scheme of PGCLC induction and expansion. Equivalent stages of mouse germline development are shown above the experimental timeline. Blimp1-RFP embryonic stem cells (ESCs) were induced into PGCLCs and further cultured to expand (Methods). b, Gating strategy to determine the efficiency of PGCLC induction. BLIMP1-RFP high, SSEA1 high for PGCLCd4, BLIMP1-RFP high for PGCLCd4c6. Experiments were repeated independently with similar results. c, Nuclear CENPA distribution pattern is heterogeneous among PGCLCd4 and PGCLCd4c5 cell population. Representative images are shown. DNA stained by DAPI (blue). Scale bar: 10 µm. Dot plot showing the measurement of the shortest distance between each CENPA focus to the nuclear periphery. Mean value of each nucleus was calculated. Each data point represents one cell analyzed (mean ± s.d.). N = 3 biological independent experiments. The P values were calculated by two-tailed Mann–Whitney U-test. Percentage of cells with periphery (average distance of centromere to the nuclear periphery < 1 µm) and non-periphery pattern. d, Representative IF image of H3K9me3 and e, H3K9me2. f, IF staining of CTCF and RAD21 in ESCs, EpiLCs and PGCLCs. AP2Ƴ positive cells were identified as PGCLCs. Yellow arrowheads indicate PGCLCs. White arrowheads indicate non PGCLCs. DNA stained by DAPI (blue). Scale bar: 10 µm. (d-f) Representative of 3 independent experiments.
Extended Data Fig. 10 In vitro gametogenesis does not recapitulate PGCs’ 3D genome re-organisation.
a, Unsupervised hierarchical clustering showing the correlation of insulation score between our samples and published data sets38. Pearson correlation was calculated based on the bin size 300 K. b, Heat maps showing normalized Hi-C interaction frequencies in ESC, EpiLC (epiblast like cell), PGCLC (primordial germ cell like cell), GSCLC (germline stem cell like cell) and GSC (germline stem cell), respectively. c, TAD aggregated plots. Numbers on the top-right are the median of TAD strength. d, Quantification of TAD strength. For each sample, the TAD strength was calculated based previously defined TADs17. TAD number n = 2822. Boxplot central lines represent median values, upper and lower limits show 75th and 25th percentiles, box heights represent interquartile ranges (IQRs) and whiskers are 1.5× the IQR. P values were calculated one-way ANOVA with Tukey correction (all P values can be found in the Source Data).
Supplementary information
Supplementary Table 1 (download XLSX )
Hi-C quality check.
Supplementary Table 2 (download XLSX )
TAD boundaries.
Supplementary Table 3 (download XLSX )
Expression of SMC genes and lamina-associated genes.
Supplementary Video 1 (download MP4 )
3D reconstruction of confocal images. Gray balls, CENPA; green, lamin B1.
Supplementary Video 2 (download MP4 )
3D telomere DNA-FISH combined with IF. Red, telomere; green, DAPI.
Supplementary Table 4 (download XLSX )
Light microscopy reporting table.
Source data
Source Data Fig. 1 (download PDF )
Unprocessed western blots.
Source Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download TIF )
Unprocessed western blots.
Source Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 6 (download PDF )
Unprocessed western blots.
Source Data Extended Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 8 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 9 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 10 (download XLSX )
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Huang, TC., Rigau, M., Malysheva, V. et al. Global reorganization of genome architecture at the transition to gametogenesis. Nat Struct Mol Biol 33, 433–447 (2026). https://doi.org/10.1038/s41594-026-01747-1
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41594-026-01747-1
