
Abstract
Clam shrimps are a group of freshwater crustaceans who prospered during the Late Triassic. They were abundant in lacustrine sedimentary records of continental basins distributed throughout Pangea during this time. However, they show significant taxonomic differences between the clamp shrimp faunas from the rift basins of central Pangea and the southern Gondwanan basins. In this contribution, we show new fossil clam shrimp assemblages from the lacustrine sedimentary successions of the Eastern Cordillera of Colombia (the Bocas and Montebel formations), providing information on the Late Triassic species that inhabited the northwestern Gondwana basins. This study demonstrates that the basins of northwestern Gondwana shared Norian clamp shrimp species with rift basins of central Pangea and differed in their faunas with the basins of the southern portion of Gondwana. In addition, the Late Triassic clam shrimps paleobiogeographic distribution reflects the dispersal of this fauna throughout fluvial-lacustrine environments established in the rift valleys along the central Pangea. Therefore, the rift valleys produced during the early fragmentation of central Pangea could have acted as corridors for dispersion. Simultaneously, rift valleys also provided paleobiogeographic barriers that isolated the central Pangea clam shrimp faunas from southern Gondwana.
Similar content being viewed by others
Introduction
The first stage of the breakup of the Pangea supercontinent was a continuous process that developed from the Ladinian to the Triassic-Jurassic boundary. As a result of rifting processes, a system of fissures crossed the central part of the supercontinent from the Caribbean to the Tethys1,2,3,4,5,6,7. This fragmentation resulted in the separation of Gondwana and Laurasia and the simultaneous fragmentation of Laurasia into North America and Eurasia2,4,5.
The best-known examples of this early fragmentation are the sedimentary records of the Germanic Basin in central Europe and the rift basins of the central Atlantic margins in the United States, Canada, and Morocco3,8,9,10,11,12 (rift basins of central Pangea). These basins were propitious areas for the establishment and accumulation of lacustrine and fluvial successions8,13,14,15,16.
The sedimentary record of the rift basins of central Pangea hosts diverse and abundant groups of clam shrimps, also known (with different systematic and phylogenetic meanings) as “conchostracans” or spinicaudatans17,18,19,20. These non-marine invertebrates show global distribution along Pangea during the Late Triassic, allowing them to be used as biostratigraphic markers21,22,23,24,25,26,27. However, for this period, the central Pangea basins share a clam shrimp fauna, which shows substantial taxonomic differences compared to the faunas of the southern Gondwana basins23,28 (Supplementary Table 1). The factors responsible for the isolation of clam shrimp faunas within a single continental mass remain under debate.
This study focuses on two fossil assemblages from the Upper Triassic successions in the Eastern Cordillera of Colombia (the Bocas and Montebel formations), presenting a novel association of clam shrimps with high-resolution biostratigraphy and providing information on the Late Triassic species that inhabited the northwestern Gondwana basins. The clamp shrimp faunas of the northwestern Gondwana units differ from those of the southern Gondwana basins. However, the northwestern Gondwana basins share clam shrimp faunal components with the rift basins of central Pangea. Combining the biostratigraphic and paleobiogeographic data provides additional insights into the spatial distribution of the clam shrimp fauna. This paleobiogeographic data reflects the early breakup of central Pangea, whith the dispersal of clam shrimps through fluvial-lacustrine environments formed in the rift valleys during the Norian.
Geological setting
The Colombian basins in northern South America were located at low paleolatitudes (0º-5º N) at the western margin of Pangea during the Late Triassic29. At this paleolatitude, the western margin of Pangea was affected by intracontinental rifting, resulting in a series of extensional basins forming during the Late Triassic30,31,32,33,34,35. In the Sierra Nevada de Santa Marta (SNSM) regions and the Eastern Cordillera, the filling of the basins is represented by the Los Indios, Corual, Tiburón, Bocas, Palermo, and Montebel formations.
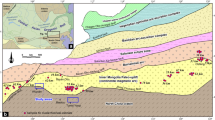
The Upper Triassic Bocas and Montebel formations are located in the Eastern Cordillera of Colombia (Fig. 1A–C). These units host fossil assemblages characterized by freshwater invertebrates (e.g. clam shrimps, darwinuloid ostracods, unionid bivalves), vertebrate teeth, fish scales, and plant remains36,37,38,39,40,41,42,43. The lithological and fossiliferous characteristics of the Bocas and Montebel formations indicate a lacustrine environment38,42,43,44.

(A) The Eastern Cordillera of Colombia in northern South America. (B,C) Detail of the studied areas from the Bocas and Montebel formations. (D) Stratigraphic columns of the Bocas and Montebel formations showing the stratigraphic position of the studied clam shrimps. Maps drawn with QGIS 3.36 (https://www.qgis.org/en/site/index.html). Stratigraphic columns drawn with Inkscape 1.2.2 (https://inkscape.org/).
Results
Clam shrimp assemblages
The identified clam shrimp assemblages exhibit significant species diversity and are abundant within the Bocas and Montebel formations (Fig. 1D; Supplementary systematic paleontology). The clam shrimp faunas in both units are characterized by an assemblage of species with the following components and percentage abundance: Howellisaura colombianus Bock, 1953 (51.5%), Shipingia hebaozhaiensis Shen, 1976 (28.5%), Euestheria winterpockensis Bock, 1953 (17.6%), Gregoriusella sp. (1.5%), and Shipingia olseni Kozur and Weems, 2005 (0.9%; Fig. 2).

Fossil clam shrimps from the Upper Triassic of the Eastern Cordillera of Colombia. (A) Shipingia olseni Kozur and Weems, 2005. (B–D) Shipingia hebaozhaiensis Shen, 1976. (E,F) Howellisaura colombianus Bock, 1953. (G) Euestheria winterpockensis Bock, 1953. (H) Gregoriusella sp. (I) Detailed growth bands (white box in (A)) with pitted ornamentation of Shipingia olseni. (J) Detailed growth bands (white box in (B)) with smooth ornamentation of Shipingia hebaozhaiensis. (K) Detailed growth bands (white box in E) with radial lirae ornamentation of Howellisaura colombianus. The yellow arrows indicate the radial lirae. (L) Detail of the growth bands with radial lirae ornamentation of Howellisaura colombianus.
Biostratigraphy
The assemblage described from the Bocas and Montebel formations in Colombia allows for correlation with the Shipingia hebaozhaiensis zone defined in the Germanic Basin and the Newark Supergroup22,23,25. This correlation assigns a middle-late Alaunian age (middle Norian) to the Bocas and Montebel formations22,23,25. However, in the middle to upper part of the Bocas Formation, the occurrence of Shipingia olseni, a marker taxa of the Sevatian21,22,23,25, in association with Shipingia hebaozhaiensis, indicates the transition between the Alaunian and Sevatian for this specific interval. The overlap of S. hebaozhaiensis and S. olseni has also been reported in the Newark Supergroup, specifically in the Groveton Member of the Bull Run Formation in the Culpeper Basin21,22.
Discussion and conclusions
Paleobiogeography
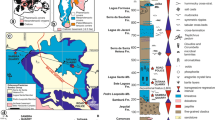
The clam shrimp taxa described from the Bocas and Montebel formations in the Eastern Cordillera of Colombia (Shipingia hebaozhaiensis, Euestheria winterpockensis, Gregoriusella sp., Shipingia olseni, and H. colombianus) reveal common elements between the northwestern Gondwana basins and the rift basins of central Pangea. The common taxa (Shipingia hebaozhaiensis, Euestheria winterpockensis, Gregoriusella sp., and Shipingia olseni) have previously been reported in the Newark Supergroup for the Culpeper, Gettysburg, Fundy, and Newark basins (U.S.A. and Canada), as well as in the Germanic Basin (Germany). In addition, H. colombianus was reported in the Tinacoa Formation, along the Serranía del Perijá45 (Venezuela, South America). Therefore, this paleobiogeographical distribution would encompass the rift basins of central Pangea extending to the northwestern Gondwana basins during the middle-late Norian (middle Alaunian-early Sevatian; Fig. 3; Supplementary Table 2).

Correlation of stratigraphic units and common clam shrimp species of the Upper Triassic in the basins of northern South America and the rift basins of central Pangea. Species reports and unit ages are based on previous studies12,21,22,23,24,25,34,45. Figure drawn with Inkscape 1.2.2 (https://inkscape.org/).
Nevertheless, older reports of clam shrimps preserved in the Los Indios Formation34 (the Sierra Nevada de Santa Marta, northern Colombia) provide evidence that the distribution of clamp shrimp assemblages in northwestern Gondwana and rift basins of central Pangea could have begun earlier, probably during the Lacian (early Norian). This early distribution is supported by Euestheria ovata Lea, 1856, Euestheria cf. E. hausmanni Schmidt, 1938, Euestheria buravasi Kobayashi, 1975 and Shipingia weemsi Kozur et al., 2012 present in the Los Indios Formation as well as in units of the Newark Supergroup and the Germanic Basin (Fig. 3; Supplementary Table 2).
Distribution and dispersion of clam shrimps across rift valleys
The paleobiogeographic distribution of clam shrimps throughout central Pangea demonstrates that this fauna (i.e., Euestheria ovata, Euestheria cf. E. hausmanni, Euestheria buravasi, Shipingia weemsi, Shipingia hebaozhaiensis, Euestheria winterpockensis, Gregoriusella sp., Shipingia olseni, H. colombianus) extended across central Europe, the eastern margin of North America (Newark Supergroup), and northern South America during the Norian. This paleobiogeographic distribution coincides with the rift zones associated with the Late Triassic early fragmentation of central Pangea (Fig. 4).

Paleobiogeographic distribution of common clam shrimp species from the Upper Triassic of the basins of northern South America and the rift basins of central Pangea. (A) Late Tuvalian-early Lacian. (B) Lacian. (C) Alaunian-early Sevatian. The red lines represent the rifting zones of central Pangea during the Late Triassic2,5,6. The scheme was developed based on paleogeographic reconstructions for the Late Triassic57,58. Figure drawn with Inkscape 1.2.2 (https://inkscape.org/).
During the latter, the fractured areas and related subsidences led to the formation of large rift valleys and a network of lakes and aligned river systems46. An analogous tectonic scenario can currently be observed in the Great Rift Valley of East Africa, resulting from the fracturing of the African Plate. The tectonic depressions are occupied by elongated and narrow lakes and river systems, reflecting the rift morphology that extends regionally (> 3000 km) from Mozambique to Ethiopia47,48.
The freshwater environments, supplied by a network of fluvial and lacustrine systems in the rift valleys that developed in a large continentalized Pangea may have provided uniquely favorable habitats for the establishment of the clamp shrimp faunas during the Late Triassic. Additionally, the clamp shrimp reproductive adaptations (i.e. accelerated biological cycle, dioecious reproduction, and the presence of abundant eggs capable of withstanding desiccation28,49,50,51,52,53,54) could have favored their high abundance in these habitats and the survival resistance of their eggs during transport across rift valleys.
This conditioned dispersion of clam shrimp fauna throughout the central Pangea basins associated with the early fragmentation (rift basins of the central Atlantic margins; Germanic Basin; northwestern Gondwana) could explain the taxonomic difference with the southern Gondwanan clamp shrimp faunas (Supplementary Table 1). Therefore, the rift valleys of central Pangea could have acted as a paleobiogeographic barrier favoring the isolation of these independently evolving faunas (provincialism).
Methods
A total of 330 remarkably well-preserved specimens were collected from the Bocas and Montebel formations for systematic identification. These individuals were obtained from dark lacustrine claystones and siltstones (Fig. 1D). The specimens were observed with a binocular stereo microscope Leica S9D and photographed with a Leica Flexcam C1 camera. The S.E.M. (scanning electron microscope) images were obtained with a JEOL 5800LV microscope at the Secretaría General de Ciencia y Técnica, Universidad Nacional del Nordeste (UNNE). The previously proposed methodology was followed for taxonomic identification and biostratigraphic schemes18,23,25,55,56. All specimens are housed in the Instituto y Museo de Ciencias Naturales collection, Universidad Nacional de San Juan (registered numbers PISJ 111–117).
Data availability
No datasets were generated or analysed during the current study.
References
Rogers, J. J. & Santosh, M. Rifting of Pangea and formation of present ocean basins. In Continents and Supercontinents (eds Rogers, J. J. & Santosh, M.) (Oxford University Press, 2004).
Blakey, R. C. Gondwana paleogeography from assembly to breakup—A 500 m.y. odyssey. In Resolving the Late Paleozoic Ice Age in Time and Space (eds Fielding, C. R. et al.) 1–28 (Geological Society of America Special Paper 441, 2008).
Olsen, P. E. Fossil great lakes of the Newark Supergroup–30 years later. In In Field Trip Guidebook, New York State Geological Association: 83nd Annual Meeting, College of Staten Island, 101–162 (2010).
Frizon de Lamotte, D., Fourdan, B., Leleu, S., Leparmentier, F. & de Clarens, P. Style of rifting and the stages of Pangea breakup. Tectonics 34, 1009–1029 (2015).
Müller, R. D. et al. Ocean basin evolution and global-scale plate reorganization events since pangea breakup. Annu. Rev. Earth Planet. Sci. 44, 107–138 (2016).
Schettino, A. & Turco, E. Breakup of Pangaea and plate kinematics of the central Atlantic and Atlas regions. Geophys. J. Int. 178, 1078–1097 (2009).
Olsen, P. E. Stratigraphic record of the early Mesozoic breakup of Pangea in the Laurasia-Gondwana rift system. Annu. Rev. Earth Planet. Sci. 25, 337–401 (1997).
Kozur, H. W. & Bachmann, G. H. The Middle Carnian wet intermezzo of the Stuttgart formation (Schilfsandstein), Germanic Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 290, 107–119 (2010).
Manspeizer, W. & Cousminer, H. L. Late Triassic-Early Jurassic synrift basins of the U.S. Atlantic margin. In The Atlantic Continental Margin (eds Sheridan, R. E. & Grow, J. A.) 197–216 (Geological Society of America, 1988). https://doi.org/10.1130/DNAG-GNA-I2.197.
Schlische, R. W. Anatomy and evolution of the Triassic-Jurassic continental rift system, eastern North America. Tectonics 12, 1026–1042 (1993).
Withjack, M. O., Schlische, R. W., Malinconico, M. L. & Olsen, P. E. Rift-basin development: Lessons from the Triassic-Jurassic Newark Basin of eastern North America. Geol. Soc. Lond. Spec. Publ. 369, 301–321 (2013).
Kent, D. V., Olsen, P. E. & Muttoni, G. Astrochronostratigraphic polarity time scale (APTS) for the Late Triassic and Early Jurassic from continental sediments and correlation with standard marine stages. Earth Sci. Rev. 166, 153–180 (2017).
Olsen, P. E. A 40-million-year lake record of early Mesozoic orbital climatic forcing. Science 1979(234), 842–848 (1986).
Olsen, P. E. Tectonic, Climatic, and Biotic Modulation of Lacustrine Ecosystems-Examples from Newark Supergroup of Eastern North America. In Lacustrine Basin Exploration: Case Studies and Modem Analogs (ed. Katz, B.) 209–224 (American Association Petroleum Geologists, 1990).
Olsen, P. E. & Kent, D. V. High-resolution early Mesozoic Pangean climatic transect in lacustrine environments. Zentralblatt für Geologie und Paläontologie, 1475–1495 (2000).
Tourani, A. et al. Characterization of the Carnian Pluvial Episode in the Argana Basin (Western High Atlas, Morocco): An approach based on sedimentology, clay mineralogy and paleosols. Palaeogeogr. Palaeoclimatol. Palaeoecol. 627, 111720 (2023).
Martin, J. W. & Davis, G. E. An Updated Classification of the Recent Crustacea Vol. 39 (Natural History Museum of Los Angeles County, 2001).
Scholze, F. & Schneider, J. W. Improved methodology of ‘conchostracan’ (Crustacea: Branchiopoda) classification for biostratigraphy. Newsl. Stratigr. 48, 287–298 (2015).
Olesen, J. Phylogeny of Branchiopoda (Crustacea)—Character evolution and contribution of uniquely preserved fossils. Arthropod. Syst. Phylogeny 67, 3–39 (2009).
Olesen, J. & Richter, S. Onychocaudata (Branchiopoda: Diplostraca), a new high-level taxon in branchiopod systematics. J. Crustacean Biol. 33, 62–65 (2013).
Kozur, H. W. & Weems, R. E. Conchostracan evidence for a late Rhaetian to early Hettangian age for the CAMP volcanic event in the Newark Supergroup, and a Sevatian (late Norian) age for the immediately underlying beds. Hallesches Jahrb. Geowiss. 27, 21–51 (2005).
Kozur, H. W. & Weems, R. E. Upper Triassic conchostracan biostratigraphy of the continental basins of eastern North America: Its importance for correlating Newark Supergroup events with the Germanic Basin and the international geologic time scale. Glob. Triassic N. M. Mus. Nat. Hist. Sci. Bull. 41, 137–188 (2007).
Kozur, H. W. & Weems, R. E. The biostratigraphic importance of conchostracans in the continental Triassic of the northern hemisphere. Geol. Soc. Lond. Spec. Publ. 334, 315–417 (2010).
Kozur, H. W., Franz, M. & Bachmann, G. Shipingia weemsi n.sp., a biostratigraphically important conchostracan species from the uppermost Carnian and lowermost Norian of Central Europe. In Triassic Geology and Paleontology (eds Tanner, L. H. et al.) 1–6 (New Mexico Museum of Natural History and Science Bulletin, 2012).
Weems, R. & Lucas, S. A revision of the Norian Conchostracan Zonation in North America and its implications for Late Triassic North American tectonic history. In Fossil Record 4 (eds Sullivan, R. & Lucas, S.) 303–317 (New Mexico Museum of Natural History and Science Bulletin 67, 2015).
Geyer, G. & Kelber, K.-P. Spinicaudata (“Conchostraca”, Crustacea) from the Middle Keuper (Upper Triassic) of the southern Germanic Basin, with a review of Carnian-Norian taxa and suggested biozones. PalZ 92, 1–34 (2018).
Kozur, H. W. & Mock, R. The importance of conchostracans for the correlation of continental and marine beds. Nonmar. Triassic N. M. Mus. Nat. Hist. Sci. Bull. 3, 261–266 (1993).
Tasch, P. Fossil Conchostraca of the Southern Hemisphere and Continental Drift: Paleontology, Biostratigraphy, and Dispersal (Geological Society of America, 1987).
Jiménez, G., Geissman, J. W. & Bayona, G. Unraveling tectonic inversion and wrench deformation in the Eastern Cordillera (Northern Andes) with paleomagnetic and AMS data. Tectonophysics 834, 229356 (2022).
Cochrane, R. et al. Permo-Triassic anatexis, continental rifting and the disassembly of western Pangaea. Lithos 190–191, 383–402 (2014).
Spikings, R. et al. The geological history of northwestern South America: From Pangaea to the early collision of the Caribbean Large Igneous province (290–75Ma). Gondwana Res. 27, 95–139 (2015).
van der Lelij, R., Spikings, R., Ulianov, A., Chiaradia, M. & Mora, A. Palaeozoic to Early Jurassic history of the northwestern corner of Gondwana, and implications for the evolution of the Iapetus, Rheic and Pacific oceans. Gondwana Res. 31, 271–294 (2016).
Toro-Toro, L. M., Cardona-Ríos, J. J., Moreno-Sánchez, M. & de Gómez-Cruz, A. J. Petrografía y geoquímica de las rocas piroclásticas y efusivas de la Formación Bocas (Triásico Superior-Jurásico Inferior) y efusivas de la Formación Nogontova (Macizo de Santander, Colombia). Bol. Geol. 43, 53–75 (2021).
Gómez, C., Kammer, A., Bernet, M., Piraquive, A. & von Quadt, A. Late Triassic rift tectonics at the northernmost Andean margin (Sierra Nevada de Santa Marta). J. S. Am. Earth Sci. 105, 102953 (2021).
Bustamante, C. et al. Middle Triassic to Jurassic convergence at the north-western margin of Gondwana: Insights from the Central Cordillera of Colombia. Int. Geol. Rev. 66, 109–129 (2024).
Langenheim, J. H. Late paleozoic and early mesozoic plant fossils from the Cordillera oriental of Colombia and correlation of the Giron Formation. Bol. Geol. 8, 95–132 (1960).
Bock, W. American Triassic estherids. J. Paleontol. 27, 62–76 (1953).
Geyer, O. Das präkretazische mesozoikum von kolumbien. Geol. Jahrb. 5, 1–156 (1973).
Rabe, E. Zur Stratigraphie Des Ostandinen Raumes von Kolumbien (Giessener geologische Schriften, 1977).
Gallego, O. et al. Comparisons of two Triassic “conchostracan” associations from Morocco and Colombia. In Reunión de comunicaciones de la Asociación Paleontológica Argentina, 118 (2019).
Alarcón, C. M., Ezcurra, M. D., Colombi, C. & Drovandi, J. M. Registro corpóreo de un tetrápodo Triásico para Colombia (Formación Montebel, Triásico Superior). In XXXVI Jornadas Argentinas de Paleontología de Vertebrados, 21–22 (2023).
Alarcón, C. M., Clavijo-Torres, J., Mantilla-Figueroa, L. C. & Rodríguez, J. G. Nueva propuesta de edades para el registro sedimentario de las formaciones Bocas y Jordán y su relación con el desarrollo de la actividad magmática del Grupo Plutónico de Santander (Cordillera Oriental, Colombia). Rev. Acad. Colomb. Cienc. Exactas Fis. Nat. 44, 1137–1151 (2020).
Remy, W., Remy, R., Pfefferkorn, H., Volkheimer, W. & Rabe, E. Neueinstufung der Bocas- Folge (Bucaramanga, Kolumbien) in den unteren Jura anhand einer Phlebopteris-branneri- und Classopollis-Flora. Argum. Paleobot. 4, 55–75 (1975).
Alarcón, C. M., Colombi, C. & Drovandi, J. M. Depósitos de grano fino asociados a flujos de fango en ambientes lacustres. Ejemplo Formación Montebel (Triásico Superior), Andes del Norte, Colombia. In IX Congreso Latinoamericano de Sedimentología. XVIII Reunión Argentina de Sedimentología, 236–236 (2023).
Odreman, O. & Benedetto, G. Paleontología y edad de la Formación Tinacoa, Sierra de Perijá, Venezuela. Mem. Congreso Geol. Venezuela 1, 15–32 (1977).
Olsen, P. E. Comparative Paleolimnology of the Newark Supergroup: A Study of Ecosystem Evolution (Yale University, 1984).
WoldeGabriel, G., Olago, D., Dindi, E. & Owor, M. Genesis of the East African rift system. In Soda Lakes of East Africa (eds WoldeGabriel, G. et al.) 25–59 (Springer International Publishing, 2016). https://doi.org/10.1007/978-3-319-28622-8_2.
Scoon, R. N. The East African rift system. In Geology of National Parks of Central/Southern Kenya and Northern Tanzania (ed. Scoon, R. N.) 19–28 (Springer International Publishing, 2018). https://doi.org/10.1007/978-3-319-73785-0_3.
Zhang, W., Shen, Y. & Niu, S. Discovery of Jurassic Conchostracans with well-preserved soft parts and notes on its biological significance. In Palaeontologia Cathayana Vol. 5 (ed. Lu, Y.) 311–351 (Springer, 1990).
Thiéry, A. Large Branchiopods (Crustacea: Anostraca, Notostraca, Spinicaudata, Laevicaudata) from temporary inland waters of the Arabian Peninsula. In Fauna of Saudi Arabia Vol. 15 (eds Krupp, F. & Mahnert, V.) 37–98 (Pro Entomologia, Natural History Museum, 1996).
Fryer, G. Diapause, a potent force in the evolution of freshwater crustaceans. Hydrobiologia 320, 1–14 (1996).
Tasch, P. Branchiopoda. In Treatise on Invertebrate Paleontology (ed. Moore, R. C.) 128–191 (Geological Society of America and University of Kansas, 1969).
Webb, J. A. A reappraisal of the palaeoecology of conchostracans (Crustacea: Branchiopoda). Neues Jahrb. Geol. Palaontol. Abh. 158, 259–275 (1979).
Weeks, S. C., Chapman, E. G., Rogers, D. C., Senyo, D. M. & Hoeh, W. R. Evolutionary transitions among dioecy, androdioecy and hermaphroditism in limnadiid clam shrimp (Branchiopoda: Spinicaudata). J. Evol. Biol. 22, 1781–1799 (2009).
Hethke, M., Hartmann, K., Alberti, M., Kutzner, T. & Schwentner, M. Testing the success of palaeontological methods in the delimitation of clam shrimp (Crustacea, Branchiopoda) on extant species. Palaeontology https://doi.org/10.1111/pala.12634 (2023).
Sun, X. & Cheng, J. Sexually dimorphic ornamentation in modern spinicaudatans and the taxonomic implications for fossil clam shrimps. Acta Palaeontol. Pol. 67, 475–492 (2022).
Scotese, C. R. The PALEOMAP Project PaleoAtlas for ArcGIS, version 2, Volume 3, Triassic and Jurassic Plate Tectonic, Paleogeographic, and Paleoclimatic Reconstructions, Map 33–48. PALEOMAP Project, Evanston, IL (2014).
Deep Time Maps. https://deeptimemaps.com/.
Acknowledgements
We thank the economic support of the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). This research is also supported by grants (PI18Q005 to O.F.G. and PI23Q001 to M.D.M.) of the Secretaría General de Ciencia y Técnica (UNNE). Additionally, we thank the Universidad Nacional de San Juan (UNSJ) and Instituto y Museo de Ciencias Naturales (IMCN) for providing the facilities to host the materials.
Author information
Authors and Affiliations
Contributions
C.M.A. designed the research project and conducted the fieldwork. C.M.A., O.F.G., and M.D.M. described all the specimens. C.E.C. and J.M.D. revised the manuscript. All the authors wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Alarcón, C.M., Colombi, C.E., Gallego, O.F. et al. Dispersal of Late Triassic clam shrimps across Pangea linking northwestern Gondwana and central Pangea rift basins. Sci Rep 14, 15025 (2024). https://doi.org/10.1038/s41598-024-66015-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-66015-y
