
Abstract
Increased dietary salt intake is a well-established risk factor for hypertension and related cardiovascular diseases, involving complex vascular remodeling processes. However, the specific role of hypoxia-inducible factor-1α (HIF-1α) in vascular pathophysiology under high-salt conditions remains poorly understood. This study investigates the role of HIF-1α in high-salt-induced vascular remodeling using human aortic vascular smooth muscle cells (HA-VSMCs) cultured in vitro. HA-VSMCs were divided into three groups: high-salt with HIF-1α knockdown (shHIF-1α + HS), negative control (shcontrol), and high-salt (HS). Cell viability, migration, gene expression, and protein levels were evaluated. High-salt conditions significantly increased mRNA expression of α-smooth muscle actin (α-SMA), smooth muscle protein 22 (SM22), angiotensin II type 1 receptor (AT1R), collagen I, and collagen III (p < 0.0001). HIF-1α knockdown partially attenuated these increases, particularly for α-SMA, SM22, and AT1R (p < 0.01). At the protein level, high-salt exposure markedly elevated expression of collagen III, HIF-1α, osteopontin (OPN), and angiotensin II (Ang II) (p < 0.0001). HIF-1α knockdown significantly reduced the high-salt-induced increases in collagen III and HIF-1α protein levels (p < 0.001) but had a limited effect on OPN and Ang II upregulation. Interestingly, SM22 protein expression was significantly decreased under high-salt conditions (p < 0.0001), an effect partially reversed by HIF-1α knockdown (p < 0.0001). These findings demonstrate that high-salt conditions induce complex changes in gene and protein expression in HA-VSMCs, with HIF-1α playing a crucial role in mediating many of these alterations. The study highlights the differential effects of HIF-1α on various markers of vascular remodeling and suggests that HIF-1α may be a potential therapeutic target for mitigating salt-induced vascular pathology. Further research is warranted to elucidate the mechanisms underlying the HIF-1α-dependent and -independent effects observed in this study.
Similar content being viewed by others

Introduction
Cardiovascular disease represents a paramount global public health concern, its significance deriving not only from its high prevalence but also from its status as a primary contributor to global mortality. Vascular remodeling (VR) emerges as a crucial etiological factor in the development of cardiovascular diseases, characterized by aberrant proliferation, migration, and phenotypic transformation of vascular smooth muscle cells (VSMCs)1,2,3. These cellular alterations are intrinsically associated with vessel wall thickening, sclerosis, and luminal narrowing. The pathogenesis of VR is a complex and multifactorial process. It involves various contributing factors, including genetic predisposition, environmental influences, and lifestyle factors4. However, the precise underlying mechanisms driving VR remain elusive. Among the myriad contributing factors, high salt (HS) intake strongly correlates with the progression of vascular remodeling, yet it remains one of the most frequently overlooked environmental determinants5,6.
Under physiological conditions, VSMCs exhibit a non-proliferative contractile phenotype7, essential for modulating vascular tone and maintaining tissue perfusion. Upon exposure to external stimuli, VSMCs can undergo phenotypic switching to a synthetic state characterized by proliferative capacity and the synthesis or secretion of osteogenic substances (e.g., alkaline phosphatase (ALP) and osteopontin (OPN) and collagens (e.g., type I and type III)8. This phenotypic alteration promotes cellular proliferation, migration, and hypertrophy and ultimately culminates in VR9. Hansen-Smith et al.5 documented structural alterations in the microvasculature of normotensive and hypertensive rats after merely three days of HS diet exposure. This observation underscores that HS stimulation is not only a critical factor in VR but also highlights the imperative nature of scientific salt regulation in the prevention and management of VR10.
Furthermore, HS-induced VR is intricately linked to angiotensin II (Ang II), a pivotal bioactive peptide within the renin–angiotensin–aldosterone system (RAAS)11. Ang II primarily mediates VR-related cellular processes, including reactive oxygen species (ROS) production, migration, and proliferation, through the angiotensin II type 1 receptor (AT1R)12. Extant literature indicates that HS intake can activate local RAAS, elevate AT1R mRNA and protein expression, and induce arterial wall thickening and collagen deposition in salt-sensitive rats13,14. Notably, angiotensin-converting enzyme inhibitors or angiotensin receptor antagonists have demonstrated efficacy in preventing HS-induced arterial collagen deposition and structural remodeling10,15. However, the potential of HS stimulation to promote Ang II expression in human aortic vascular smooth muscle cells (HA-VSMCs) cultured in vitro and its mechanistic association with VR warrants further investigation.
Hypoxia-inducible factor-1 (HIF-1) plays a significant role in the pathogenesis of VR16,17. HIF-1 is a heterodimeric transcription factor composed of HIF-1α and HIF-1β subunits, with HIF-1α primarily responsible for regulating its functional activities, including energy metabolism, angiogenesis, and cellular proliferation and migration9,18,19,20. These functions are integral to the processes of VR, establishing HIF-1α as a critical hypoxia-related transcription factor. Beyond hypoxic conditions, HS stimulation can also induce HIF-1α expression and its target genes in the renal medulla, contributing to salt-sensitive hypertension21. HIF prolyl hydroxylase-2 (PHD2), highly expressed in the renal medulla, has been identified as a major enzyme promoting HIF-1α degradation. In rats with normal blood pressure, the HS diet suppresses the expression and activity of PHD2 in the renal medulla, leading to increased expression of HIF-1α-related genes and influencing the sensitivity of blood pressure to salt22.
Extensive research has shown that HS stimulation increases the expression of HIF-1α. However, the specific mechanisms behind this upregulation and its role in VR are not fully understood. HIF-1α is a key point of convergence for multiple signaling pathways, but the regulatory mechanisms of HIF-1 signaling are not yet fully defined. We hypothesize that HIF-1α plays a crucial role in HS-induced, Ang II-mediated vascular remodeling in HA-VSMCs and that the knockdown of HIF-1α will inhibit this process.
To investigate this hypothesis, we used an in vitro model to explore the relationship between HS stimulation, HIF-1α, and Ang II and to understand how HIF-1α induces VR in HA-VSMCs. By employing HIF-1α knockdown techniques and simulating high salt conditions, we aimed to gain insights into the regulatory role of HIF-1α in VR and its potential as a therapeutic target.
Materials and methods
Materials
HA-VSMCs were procured from Wuhan Procell Biological Technology Co., Ltd.. The lentiviral packaging was executed by Shanghai Genechem Co., Ltd. Cells were cultivated in Dulbecco’s Modified Eagle Medium (DMEM, Gibco) containing 132 mmol/L Na+ (Gibco, USA), supplemented with New Zealand fetal bovine serum (FBS; Gibco, USA), and 100 U/mL penicillin and 100 μg/mL streptomycin (Biosharp Life Sciences, China). Sodium chloride (purity > 99.9%) was sourced from Beijing Solarbio Science & Technology Co., Ltd. The Cell Counting Kit-8 (CCK-8) was obtained from Proteintech Group, Inc. Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) reagents were acquired from Novozymes A/S. Antibodies used in this study included primary antibodies against HIF-1α (Cell Signaling Technology)23, SM22 (Cell Signaling Technology), Collagen III (Proteintech), Ang II (Proteintech), OPN (Proteintech)24, and β-actin (Proteintech). Secondary antibodies were horseradish peroxidase-conjugated goat anti-rabbit IgG (Cell Signaling Technology) and horse anti-mouse IgG (Cell Signaling Technology).
Cell culture and transfection
HA-VSMCs were initially subjected to varying sodium concentrations (137, 142, 147, 152, 157, 162, and 167 mmol/L Na+) by supplementing DMEM complete medium with sodium chloride. Cells were incubated at 37 °C in a humidified atmosphere containing 5% CO2 to determine the optimal HS stimulation concentration for subsequent experiments. The study design comprised three experimental groups: the high-salt knockdown group (transfected with lentivirus expressing shHIF-1α), the control group (transfected with negative control shRNA sequences), and the high-salt group (152 mmol/L Na+). All HA-VSMCs were cultured in DMEM complete medium supplemented with 15% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin.
HA-VSMCs in the logarithmic growth phase were harvested using trypsin digestion, centrifuged, and seeded into six-well culture plates. After 20–24 h, when cell confluence reached approximately 60%, the medium was replaced with 2.5 mL of serum-free and antibiotic-free medium. Following transfection, the high-salt group and the high-salt knockdown group were exposed to high-salt conditions (152 mmol/L Na+) for 72 h, after which cellular proteins and RNA were extracted for analyses. The shHIF-1α target sequence was 5’-GCCGAGGAAGAACTATGAACA-3', while the negative control shRNA (shcontrol) target sequence was 5’-TTCTCCGAACGTGTCACGT-3'. Lentiviral vectors expressing green fluorescent protein (GFP) were constructed. Cells were seeded in 6-well plates (1 × 105 cells/well) and transduced with lentiviral particles at an MOI of 100, supplemented with 5 μg/mL polybrene. Seventy-two hours post-transduction, GFP expression was visualized using a fluorescence microscope (Model DP73, Olympus) equipped with a GFP filter set (excitation: 488 nm, emission: 509 nm). Images were captured using identical exposure settings (exposure time: 160 ms seconds) for both groups to ensure comparable fluorescence intensity. While GFP fluorescence indicated successful transduction, HIF-1α knockdown efficiency was quantified separately using RT-qPCR, with HIF-1α expression levels normalized to GAPDH as an internal control.
Cell proliferation assay
Cell proliferation was assessed using the Cell Counting Kit-8 (CCK-8) assay. HA-VSMCs were enzymatically dissociated using trypsin, enumerated, and subsequently seeded into 96-well plates at a density of 2.5 × 103 cells per well. Following overnight adherence, the culture medium was replaced with a standard medium containing 1% New Zealand fetal bovine serum to facilitate a 12-h serum-free synchronization period. Post-synchronization, the cells were exposed to media containing varying sodium concentrations (137, 142, 147, 152, 157, 162, and 167 mmol/L Na+) and incubated for 24, 48, 72, and 96 h.
As per the CCK-8 kit protocol, 100 μL of the premixed solution (CCK-8 solution: DMEM complete medium = 10:100) was added to each well. The HA-VSMCs were then incubated at 37 °C in a humidified atmosphere with 5% CO2 for 1.5 h to allow for chromogenic reaction development. Subsequently, the absorbance of each well was quantified at 450 nm using a microplate spectrophotometer.
Cell migration assay
HA-VSMCs were seeded in six-well plates at a density of 1.5 × 105 cells/L and allowed to reach confluency (> 90%) overnight. A sterile 200 μL pipette tip was used to create a linear scratch wound across the monolayer. The wells were then gently rinsed several times with phosphate-buffered saline (PBS) to remove cellular debris. To mitigate the confounding effects of cell proliferation on the assay results, the cultures were maintained in a medium supplemented with 1% New Zealand fetal bovine serum.
Wound closure was monitored at 0 and 6 h post-wounding. Digital images were captured at these time points using phase-contrast microscopy. Cell migration was quantified by measuring the reduction in scratch width over time. The migration distance was calculated as the difference between the initial and final wound widths, providing a measure of the cells’ migratory capacity under the experimental conditions.
Colorimetric method
Cells that underwent the intervention were harvested and collected in a centrifuge tube. The cells were resuspended in 180 µl of PBS. Cell lysis was performed using an ultrasonic cell crusher with the following parameters: 200 watts power, 30 cycles of 3-s ultrasonic pulses each followed by a 10-s interval. The centrifuge tube was then subjected to the ultrasonic cell crusher for complete cell disruption. Following sonication, the lysate was centrifuged at 12,000 rpm for 10 min at 4 ℃ in a refrigerated high-speed centrifuge. The resulting supernatant was collected and used for SOD and MDA assays according to the manufacturer’s instructions provided in the reagent manual.
ELISA
After the intervention, the cells were collected, and the cell suspension was diluted with PBS (without calcium) to a concentration of approximately 106/ml. The cells were frozen and thawed several times to disrupt them and release the intracellular assay material. They were then centrifuged for approximately 20 min (2500–3000 rpm), the supernatant was carefully collected, and the OPN level was assayed according to the ELISA instructions.
RT-qPCR analysis
Total RNA was isolated using an RNA-easy Isolation Reagent. cDNA synthesis was performed using HiScript III All-in-one RT SuperMix Perfect for qPCR in a one-step reverse transcription process. RT-qPCR was conducted using ChamQ Universal SYBR qPCR Master Mix. The thermal cycling conditions were as follows: initial denaturation at 95 °C for 30 s, followed by 40 cycles of 95 °C for 10 s and 60 °C for 30 s, with a final dissociation stage of 95 °C for 15 s, 60 °C for 1 min, and 95 °C for 15 s.
The primer sequences utilized were:
- HIF-1α:
-
: forward 5’-TACCCACCGCTGAAACGC-3'.
reverse 5’-TGCTTCCATCGGAAGGACTAG-3'.
- AT1R:
-
: forward 5’- TAAGATTGCTTCAGCCAGCGTCAG-3'.
reverse 5’-TTGGGTGAACAATAGCCAGGTATCG-3'.
- α-SMA:
-
: forward 5ʹ-GATGGGCATCTATCAGATAC-3ʹ.
reverse 5ʹ-AAGCATTTCTGATGGTGATG-3ʹ.
- SM22:
-
forward 5’- GTGGAGTGGATCATAGTGCAGTGTG-3'.
reverse 5ʹ-AATCACGCCATTCTTCAGCCAGAC-3'.
- Collagen I:
-
: forward 5’-GATTCCCTGGACCTAAAGGTGC-3’.
reverse 5’-AGCCTCTCCATCTTTGCCAGCA-3’.
- Collagen III:
-
forward 5’-CTCAGGGTGTCAAGGGTGAAAGTG-3’.
reverse 5’-TGTACCAGCCAGACCAGGAAGAC-3’.
- β-actin:
-
forward 5ʹ-CCTGGCACCCAGCACAAT-3'.
reverse 5ʹ-GGGCCGGACTCGTCATAC-3'.
β-actin was used as an internal control, and the relative expression levels were analyzed using the 2^(-ΔΔCt) relative quantification method25.
Western blot analysis
HA-VSMCs in the high-salt and high-salt knockdown groups were cultured in a medium containing 152 mmol/L Na+ for 72 h. Cells were then washed twice with PBS and lysed for 20 min using RIPA premixed lysate (lysate: PMSF = 100:1). The cell lysates were collected and centrifuged at 12,000 rpm for 5 min. The supernatants were denatured at 98 °C.
Protein concentrations were determined using the BCA method. A total of 20 μg protein per sample was separated by 12% SDS-PAGE and transferred to PVDF membranes. Membranes were blocked with 5% skim milk for 1 h, then incubated with primary antibodies overnight at 4 °C. After washing three times with TBST (10 min each), membranes were incubated with HRP-labeled secondary antibodies for 1 h at room temperature. Following three additional TBST washes, protein bands were visualized using ECL chemiluminescence detection.
Statistical analysis
Data are presented as mean ± standard deviation from at least three independent experiments. All statistical analyses were performed using GraphPad Prism 10 (Chicago, IL, USA). Differences among multiple groups were compared using a one-way analysis of variance (ANOVA) followed by an appropriate post-hoc test (specified in each figure legend). For comparisons between the two groups, a Student’s t-test was applied. Statistical significance was set at P < 0.05. Specific P-values or significance levels are indicated in each figure as asterisks or other appropriate symbols, with explanations provided in the corresponding figure legends.
Results
RNA interference-mediated knockdown of HIF-1α expression in HA-VSMCs
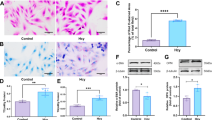
Lentiviral-mediated RNA interference effectively suppressed HIF-1α expression in human aortic vascular smooth muscle cells (HA-VSMCs). Fluorescence microscopy (Fig. 1A, B) confirmed successful transduction, while RT-qPCR (Fig. 1C) revealed a significant reduction in HIF-1α mRNA levels. The knockdown efficiency reached approximately 90% compared to the control group (p < 0.0001), demonstrating the high efficacy of the engineered lentiviral construct in silencing HIF-1α expression. This robust knockdown provides a solid foundation for further investigations into the role of HIF-1α in HA-VSMC function under various experimental conditions.

Validation of HIF-1α knockdown efficacy in HA-VSMCs using RNA interference. (A, B) Representative fluorescence microscopy images (100× magnification) illustrating the transduction efficiency of lentiviral vectors in HA-VSMCs. Cells were transduced with either control shRNA (shcontrol) (A) or HIF-1α-targeted shRNA (shHIF-1α) (B) constructs. Green fluorescence indicates successful transduction. Scale bars: 100 μm. (C) Quantitative assessment of HIF-1α mRNA expression levels in HA-VSMCs following lentiviral transduction. Expression was analyzed using real-time quantitative PCR (RT-qPCR) and normalized to the shcontrol group. Data are presented as mean ± standard deviation. Statistical significance was determined using Student’s t-test. ****P < 0.0001 compared to the shcontrol group.
Effect of high-salt conditions on HA-VSMCs proliferation
To elucidate the potential proliferative effects of HS on HA-VSMCs, cell proliferation was assessed using the CCK-8 assay. As illustrated in Fig. 2A, exposure to a sodium concentration gradient ranging from 132 to 152 mmol/L resulted in a dose-dependent increase in HA-VSMC proliferative activity. Conversely, sodium concentrations between 157 and 167 mmol/L elicited a progressive decline in HA-VSMC proliferation relative to the control group, with evidence of cellular apoptosis at higher concentrations.

Effects of elevated sodium concentrations on HA-VSMCs viability and the role of HIF-1α. (A) Time- and dose-dependent effects of varying sodium concentrations on HA-VSMC viability. Cells were exposed to a range of sodium concentrations (132–167 mmol/L) for 24, 48, 72, and 96 h. Cell viability was assessed and expressed as a percentage. (B) Comparative analysis of HA-VSMCs viability under high-sodium conditions in shcontrol, HS, and shHIF-1α + HS groups at 72 h post-treatment. Data are presented as mean ± standard deviation. Statistical significance was determined using one-way ANOVA followed by Tukey’s post-hoc test. *P < 0.05, **P < 0.01, ***P < 0.001 compared to the shcontrol group. shcontrol, negative control group; HS, high salt group (152 mmol/L Na+); shHIF-1α + HS, high salt knockdown group treated with high salt; HA-VSMCs, human aortic vascular smooth muscle cell; HIF-1α, hypoxia-inducible factor 1-alpha.
We investigated the effects of various sodium concentrations (137, 142, 147, 152, 157, 162, and 167 mmol/L) on HA-VSMC viability over 48, 72, and 96 h. The results showed that exposure to 152 mmol/L Na+ led to a statistically significant enhancement in cell viability compared to the control group (p < 0.001). Based on these findings, we selected 152 mmol/L Na+ as the optimal high-sodium stimulus concentration for the subsequent experiments.
Under these optimized conditions (152 mmol/L Na+), both the HS group and the shHIF-1α + HS groups exhibited enhanced proliferative capacity relative to the shcontrol group. However, the proliferative potential of the shHIF-1α + HS group was attenuated compared to the HS group (Fig. 2B). These findings suggest a potential role for HIF-1α in mediating the proliferative response of HA-VSMCs to elevated sodium concentrations.
Effect of high-salt conditions on HA-VSMCs migration
Our investigation into the migratory behavior of HA-VSMCs under high-salt conditions revealed significant effects. As illustrated in Fig. 3, HS treatment markedly enhanced HA-VSMCs migration compared to the control group (p < 0.001). Interestingly, while knockdown of HIF-1α (ShHIF-1α + HS) partially attenuated this effect, it still resulted in significantly increased migration relative to the control (p < 0.01). The wound healing assay demonstrated that after 6 h, HS-treated cells exhibited approximately 35% cell mobility, compared to 25% in the ShHIF-1α + HS group and 17% in the control. These results suggest that high-salt conditions promote HA-VSMCs migration through both HIF-1α-dependent and independent mechanisms, highlighting the complex interplay between salt exposure and vascular cell behavior.

High-salt stimulation promotes migration of HA-VSMCs, partially mediated by HIF-1α. (A) Representative images of the scratch wound healing assay for HA-VSMCs under different conditions. Images were taken at 0 h and 6 h after scratch formation. Yellow lines indicate the edges of the scratch wound. (B) Quantification of cell motility expressed as cell mobility (%). Compared to the shcontrol group, both the high salt (HS) and HIF-1α knockdown (shHIF-1α + HS) groups showed significantly increased cell mobility. The HS group demonstrated the highest mobility, while the shHIF-1α + HS group showed an intermediate level of mobility between shcontrol and HS. ****P < 0.0001; *** P < 0.001. Scale bar: 200 μm.
Effect of high salt stimulation on oxidative stress and calcification in HA-VSMCs
Changes in the levels of superoxide dismutase (SOD) and malondialdehyde (MDA) in cells are good indicators of oxidative stress. To determine whether high salt stimulation can promote or accelerate the oxidative stress process in HA-VSMCs, we examined the changes in SOD and MDA levels in cells using a colorimetric method. The results (Fig. 4A,B) indicate a significant decrease in SOD content and a significant increase in MDA in the HS group but not in the shHIF-1α + HS group compared to the shcontrol group. Our results suggest that HIF-1α knockdown partially alleviates HS-induced oxidative stress, as evidenced by a modest reduction in oxidative stress markers. However, the effect was minor, indicating that other factors likely contribute to the HS-induced oxidative stress in HA-VSMCs. Additionally, OPN expression exhibited a similar change pattern under this stimulus (Fig. 4C). Thus, high salt stimulation may promote oxidative stress and calcification in HA-VSMCs. These effects appear to be mitigated by the downregulation of HIF-1α under high salt conditions, potentially delaying or reducing the oxidative stress and calcification induced by high salt stimulation.

High-salt stimulation induces changes in oxidative stress markers and OPN expression in HA-VSMCs. (A–C) Quantitative analysis of (A) SOD activity (U/mgprot), (B) MDA levels (nmol/mgprot), and (C) OPN expression (ng/ml) in negative control (shcontrol), high salt (HS), and high-salt with HIF-1α knockdown (shHIF-1α + HS) groups. Data are presented as mean ± standard deviation. ns: not significant, *p < 0.05, **p < 0.01, ***p < 0.001. SOD, superoxide dismutase; MDA, malondialdehyde; OPN, osteopontin.
Effects of high salt stimulation on cellular phenotypic transformation and related mRNA and proteins expression in HA-VSMCs
Figure 5 demonstrates the effects of high-salt and HIF-1α knockdown (shHIF-1α + HS) on mRNA expression levels of various genes in HA-VSMCs. The results show that, compared to the control group, high-salt stimulation significantly decreased the expression of contractile markers α-SMA and SM22, while increasing the expression of synthetic markers collagen I and III, as well as HIF-1α and AT1R. Notably, these high-salt-induced changes were partially reversed in the HIF-1α knockdown condition, suggesting that HIF-1α may play a crucial role in the phenotypic transformation of vascular smooth muscle cells.

mRNA expression levels of key genes in response to high salt and HIF-1α knockdown. (A–F) Quantitative analysis of mRNA expression levels for various genes in negative control (shcontrol), high salt (HS), and high-salt with HIF-1α knockdown (shHIF-1α + HS) groups. (A) α-SMA, (B) SM22, (C) Collagen I, (D) Collagen III, (E) HIF-1α, and (F) AT1R. Data are presented as mean ± standard deviation. ns: not significant, **p < 0.01, ***p < 0.001, ****p < 0.0001. α-SMA, alpha-Smooth Muscle Actin; SM22, Smooth Muscle Protein 22-alpha; HIF-1α: Hypoxia-Inducible Factor 1-alpha; AT1R: Angiotensin II Type 1 Receptor.
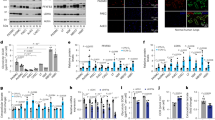
Figure 6 further confirms these findings at the protein level through Western blot analysis. The results indicate that high-salt stimulation increased the expression of collagen III, HIF-1α, OPN, and Ang II, while decreasing the expression of SM22. Knockdown of HIF-1α was able to partially inhibit these high-salt-induced changes, particularly in the expression of collagen III, HIF-1α, OPN, and Ang II. Collectively, these results suggest that HIF-1α plays a key regulatory role in the high-salt-induced phenotypic transformation of vascular smooth muscle cells, and that inhibiting HIF-1α may be a potential strategy for maintaining the normal phenotype of vascular smooth muscle cells under high-salt conditions.

Expression levels of key proteins in different experimental groups. (A) Representative western blot images showing the protein expression of Collagen III, HIF-1α, OPN, Ang II, SM22, and β-actin in the negative control (shcontrol), high salt (HS), and high-salt with HIF-1α knockdown (shHIF-1α + HS) groups. (B–F) Quantitative analysis of protein expression levels normalized to β-actin. (B) Collagen III, (C) HIF-1α, (D) OPN, (E) Ang II, and (F) SM22. Data are presented as mean ± standard deviation. n = 3 per group. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. HIF-1α: Hypoxia-Inducible Factor 1-alpha; OPN, Osteopontin; Ang II, Angiotensin II; SM22, Smooth Muscle Protein 22-alpha.
Discussion
This study provides novel insights into the critical role of HIF-1α in high-salt induced phenotypic transformation of HA-VSMCs. By successfully suppressing HIF-1α expression through lentiviral-mediated RNA interference with approximately 90% knockdown efficiency, we established a solid foundation for investigating its function under high-salt conditions.
Our findings demonstrate that high-salt conditions, optimally at 152 mmol/L Na+, significantly enhance HA-VSMCs proliferation and migration. These results align with previous studies, which also observed increased VSMCs proliferation and migration under high-salt conditions26,27. However, our study goes further by revealing the regulatory role of HIF-1α in this process. The partial attenuation of high-salt-induced proliferation and migration in HIF-1α knockdown cells suggests that HIF-1α is a key mediator in the cellular response to elevated sodium levels, providing a new perspective on salt-induced vascular remodeling.
The research also sheds light on the impact of high-salt stimulation on oxidative stress and calcification in HA-VSMCs. We observed decreased SOD content and increased MDA and OPN levels under high-salt conditions, indicating enhanced oxidative stress and calcification processes. These findings are consistent with previous studies, which also reported salt-induced oxidative stress in vascular smooth muscle cells26,28. Our study extends this knowledge by demonstrating that HIF-1α knockdown partially mitigates these HS-induced changes, suggesting a potential regulatory role for HIF-1α in modulating oxidative stress and calcification under high-salt conditions.
A key finding of our study is the high-salt-induced alteration in gene expression profiles and phenotypic transformation of HA-VSMCs. High-salt conditions significantly decreased the expression of contractile markers (α-SMA and SM22) while increasing synthetic markers (collagen I and III), HIF-1α, and AT1R at both mRNA and protein levels. These results corroborate the findings of some previous studies, which also observed a shift from contractile to synthetic phenotype in VSMCs under high-salt conditions29,30. Our study advances this understanding by demonstrating that HIF-1α knockdown partially reverses these high-salt-induced changes, highlighting its crucial role in regulating VSMCs phenotypic transformation.
Based on our results and previous literature, we propose several potential mechanisms through which HIF-1α may mediate high-salt-induced VSMCs phenotypic transformation. Firstly, HIF-1α might influence cell proliferation and migration by regulating angiotensin II (Ang II) expression. Our observation of increased Ang II expression under high-salt conditions, which was partially reversed by HIF-1α knockdown, aligns with the finding on HIF-1α's role in regulating the renin-angiotensin system31,32. Secondly, HIF-1α may modulate cellular redox state by regulating oxidative stress-related genes32, as evidenced by the changes in SOD and MDA levels. Additionally, HIF-1α could influence vascular remodeling by regulating the expression of extracellular matrix proteins such as collagen.
The strengths of this study lie in its systematic approach to investigating HIF-1α's role in high-salt-induced VSMCs phenotypic transformation, the use of multiple experimental methods to comprehensively assess the effects of high-salt on VSMCs, and the novel mechanistic insights provided by the HIF-1α knockdown experiments.
While our study provides valuable insights, it also has limitations. The experiments were conducted in vitro, and in vivo validation would strengthen the findings. The transfection of HA-VSMCs prior to high-salt stimulation, necessitated by technical constraints, may not fully replicate chronic high-salt exposure conditions. Additionally, we observed varying HIF-1α knockdown efficiencies between normal salt (90%) and high-salt (20%) conditions, likely due to salt-induced HIF-1α stabilization. This discrepancy could lead to an underestimation of HIF-1α's effects under high-salt conditions—however, the significant effects observed despite partial knockdown underscore HIF-1α's potential importance in salt-induced vascular remodeling. In addition, although we observed HIF-1α's regulatory effects, the specific molecular mechanisms remain to be fully elucidated. Furthermore, the study did not explore other transcription factors or signaling molecules that might be involved in HS-induced VSMCs phenotypic transformation.
Future research directions could include validating HIF-1α's role in salt-induced vascular remodeling in animal models, further exploring the specific molecular mechanisms of HIF-1α-mediated VSMCs phenotypic regulation through techniques such as Chromatin Immunoprecipitation-Sequencing, investigating potential interactions between HIF-1α and other known regulators of VSMCs phenotype (e.g., Krüppel-like factor 433, myocardin34), and exploring therapeutic strategies targeting HIF-1α for preventing or treating salt-sensitive cardiovascular disorders.
In conclusion, this study provides new insights into the mechanisms of high-salt-induced vascular smooth muscle cell phenotypic transformation, highlighting the critical regulatory role of HIF-1α in this process. These findings not only deepen our understanding of the pathogenesis of salt-related cardiovascular diseases but also suggest potential molecular targets for developing new preventive and therapeutic strategies. By elucidating the complex interplay between high-salt conditions, HIF-1α, and VSMCs phenotype, this research contributes significantly to the field of vascular biology and opens new avenues for addressing salt-sensitive cardiovascular disorders.
Data availability
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
References
Tang, H. Y. et al. Vascular smooth muscle cells phenotypic switching in cardiovascular diseases. Cells https://doi.org/10.3390/cells11244060 (2022).
Wang, G. et al. MicroRNA regulation of phenotypic transformations in vascular smooth muscle: Relevance to vascular remodeling. Cell. Mol. Life Sci. 80, 144. https://doi.org/10.1007/s00018-023-04793-w (2023).
Wei, C. et al. Resolvin D1 attenuates Ang II-induced hypertension in mice by inhibiting the proliferation, migration and phenotypic transformation of vascular smooth muscle cells by blocking the RhoA/mitogen-activated protein kinase pathway. J. Hypertens. 42, 420–431. https://doi.org/10.1097/hjh.0000000000003610 (2024).
Guo, J. et al. Aging and aging-related diseases: From molecular mechanisms to interventions and treatments. Signal Transduct. Target Ther. 7, 391. https://doi.org/10.1038/s41392-022-01251-0 (2022).
Hansen-Smith, F. M., Morris, L. W., Greene, A. S. & Lombard, J. H. Rapid microvessel rarefaction with elevated salt intake and reduced renal mass hypertension in rats. Circ. Res. 79, 324–330. https://doi.org/10.1161/01.res.79.2.324 (1996).
Grillo, A., Salvi, L., Coruzzi, P., Salvi, P. & Parati, G. Sodium intake and hypertension. Nutrients https://doi.org/10.3390/nu11091970 (2019).
Wang, H. et al. The organ specificity in pathological damage of chronic intermittent hypoxia: An experimental study on rat with high-fat diet. Sleep Breath. 17, 957–965. https://doi.org/10.1007/s11325-012-0784-1 (2013).
Cao, G. et al. How vascular smooth muscle cell phenotype switching contributes to vascular disease. Cell Commun. Signal. 20, 180. https://doi.org/10.1186/s12964-022-00993-2 (2022).
Chen, J. et al. HIF-2α upregulation mediated by hypoxia promotes NAFLD-HCC progression by activating lipid synthesis via the PI3K-AKT-mTOR pathway. Aging 11, 10839–10860. https://doi.org/10.18632/aging.102488 (2019).
Liang, B. & Leenen, F. H. Prevention of salt induced hypertension and fibrosis by angiotensin converting enzyme inhibitors in Dahl S rats. Br. J. Pharmacol. 152, 903–914. https://doi.org/10.1038/sj.bjp.0707472 (2007).
Zeng, Z. H. et al. Control of vascular changes by renin-angiotensin-aldosterone system in salt-sensitive hypertension. Eur. J. Pharmacol. 503, 129–133. https://doi.org/10.1016/j.ejphar.2004.09.022 (2004).
Ziaja, M., Urbanek, K. A., Kowalska, K. & Piastowska-Ciesielska, A. W. Angiotensin II and angiotensin receptors 1 and 2-multifunctional system in cells biology, what do we know?. Cells 10, 66. https://doi.org/10.3390/cells10020381 (2021).
Hengel, F. E., Benitah, J.-P. & Wenzel, U. O. Mosaic theory revised: Inflammation and salt play central roles in arterial hypertension. Cell. Mol. Immunol. 19, 561–576. https://doi.org/10.1038/s41423-022-00851-8 (2022).
Russo, E. et al. Non-haemodynamic mechanisms underlying hypertension-associated damage in target kidney components. Int. J. Mol. Sci. 24, 9422 (2023).
Kagiyama, S. et al. Aldosterone-and-salt-induced cardiac fibrosis is independent from angiotensin II type 1a receptor signaling in mice. Hypertens. Res. 30, 979–989. https://doi.org/10.1291/hypres.30.979 (2007).
Rey, S. & Semenza, G. L. Hypoxia-inducible factor-1-dependent mechanisms of vascularization and vascular remodelling. Cardiovasc. Res. 86, 236–242. https://doi.org/10.1093/cvr/cvq045 (2010).
Lim, C. S., Kiriakidis, S., Sandison, A., Paleolog, E. M. & Davies, A. H. Hypoxia-inducible factor pathway and diseases of the vascular wall. J. Vasc. Surg. 58, 219–230. https://doi.org/10.1016/j.jvs.2013.02.240 (2013).
Joshi, S., Singh, A. R., Zulcic, M. & Durden, D. L. A macrophage-dominant PI3K isoform controls hypoxia-induced HIF1α and HIF2α stability and tumor growth, angiogenesis, and metastasis. Mol. Cancer Res. 12, 1520–1531. https://doi.org/10.1158/1541-7786.Mcr-13-0682 (2014).
Harris, A. L. Hypoxia—A key regulatory factor in tumour growth. Nat. Rev. Cancer 2, 38–47. https://doi.org/10.1038/nrc704 (2002).
Ke, Q. & Costa, M. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharmacol. 70, 1469–1480. https://doi.org/10.1124/mol.106.027029 (2006).
Wang, Z. et al. Hypoxia-inducible factor prolyl-hydroxylase 2 senses high-salt intake to increase hypoxia inducible factor 1alpha levels in the renal medulla. Hypertension 55, 1129–1136. https://doi.org/10.1161/hypertensionaha.109.145896 (2010).
Li, N. Hypoxia inducible factor-1α-mediated gene activation in the regulation of renal medullary function and salt sensitivity of blood pressure. Am. J. Cardiovasc. Dis. 2, 208–215 (2012).
Kim, E.-J. et al. Transcriptional activation of HIF-1 by RORα and its role in hypoxia signaling. Arterioscler. Thromb. Vasc. Biol. 28, 1796–1802. https://doi.org/10.1161/ATVBAHA.108.171546 (2008).
Wang, T., Li, Y., Chen, J., Xie, L. & Xiao, T. TGF-β1/Smad3 signaling promotes collagen synthesis in pulmonary artery smooth muscle by down-regulating miR-29b. Int. J. Clin. Exp. Pathol. 11, 5592–5601 (2018).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. https://doi.org/10.1006/meth.2001.1262 (2001).
Bkaily, G. et al. High Na+ salt diet and remodeling of vascular smooth muscle and endothelial cells. Biomedicines 9, 883 (2021).
Liu, G. et al. High sodium augments angiotensin II-induced vascular smooth muscle cell proliferation through the ERK 1/2-dependent pathway. Hypertens. Res. 37, 13–18. https://doi.org/10.1038/hr.2013.108 (2014).
Ying, K. E. et al. Dietary salt initiates redox signaling between endothelium and vascular smooth muscle through NADPH oxidase 4. Redox Biol. 52, 102296. https://doi.org/10.1016/j.redox.2022.102296 (2022).
Liu, N. et al. The dynamic change of phenotypic markers of smooth muscle cells in an animal model of cerebral small vessel disease. Microvasc. Res. 133, 104061. https://doi.org/10.1016/j.mvr.2020.104061 (2021).
Kusche-Vihrog, K., Schmitz, B. & Brand, E. Salt controls endothelial and vascular phenotype. Pflügers Arch. 467, 499–512. https://doi.org/10.1007/s00424-014-1657-1 (2015).
Nangaku, M. & Fujita, T. Activation of the renin-angiotensin system and chronic hypoxia of the kidney. Hypertens. Res. 31, 175–184. https://doi.org/10.1291/hypres.31.175 (2008).
Kurlak, L. O., Mistry, H. D., Cindrova-Davies, T., Burton, G. J. & Broughton Pipkin, F. Human placental renin-angiotensin system in normotensive and pre-eclamptic pregnancies at high altitude and after acute hypoxia-reoxygenation insult. J. Physiol. 594, 1327–1340. https://doi.org/10.1113/jp271045 (2016).
Yap, C., Mieremet, A., de Vries, C. J. M., Micha, D. & de Waard, V. Six shades of vascular smooth muscle cells illuminated by KLF4 (Krüppel-like factor 4). Arterioscler. Thromb. Vasc. Biol. 41, 2693–2707. https://doi.org/10.1161/atvbaha.121.316600 (2021).
Long, X., Bell, R. D., Gerthoffer, W. T., Zlokovic, B. V. & Miano, J. M. Myocardin is sufficient for a smooth muscle-like contractile phenotype. Arterioscler. Thromb. Vasc. Biol. 28, 1505–1510. https://doi.org/10.1161/atvbaha.108.166066 (2008).
Acknowledgements
We sincerely thank the staff from the Department of Pathology at the First Affiliated Hospital of Dali University for their invaluable contributions to this study.
Funding
Our work was supported by the National Natural Science Foundation of China (Nos. 82260076 and 81860084), the Xingdian Famous Physician Special Project of Yunnan Province, the Youth Top-notch Talents Program of the Ten Thousand Talents Plan of Yunnan Province, Yunnan Key Specialties, and the Key Discipline of the First Affiliated Hospital of Dali University.
Author information
Authors and Affiliations
Contributions
LLH conceptualized and designed the study. DWB, HSQ, YLS, and WX carried out experimental work. All authors reviewed and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Deng, W., Huang, S., Yu, L. et al. HIF-1α knockdown attenuates phenotypic transformation and oxidative stress induced by high salt in human aortic vascular smooth muscle cells. Sci Rep 14, 28100 (2024). https://doi.org/10.1038/s41598-024-79892-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-79892-0
