
Abstract
Changing climates threaten crop growth and fodder yields in dryland farming. This study assessed two radish genotypes (LINE 2, ENDURANCE) under three water regimes (W1 = well-watered, W2 = moderate stress, W3 = severe stress) and two leaf harvesting options over two seasons (2021/22 and 2022/23). Key findings revealed that water regime significantly (P < 0.05) affected WUE, with W2 yielding (4.71 kg ha-1 mm-1) higher values. The combination of W3 and LINE 2 biomass were reduced by ≈ 60.09% in 2021/22 and ≈ 71.06% in 2022/23, whereas ENDURANCE declined by ≈ 63.9% and ≈ 53.33%. Tuber yield was highest under W1 and W2, with ENDURANCE yielding 59 t ha-1 (W1) and 48 t ha-1 (W2). Generally, W2 reduced micronutrient concentration (iron, zinc, β-carotene and vitamin C). For example, W2 improved vitamin E and key findings showed that human dietary for women and children can be met. The W3 exceeded vitamin E, iron, and zinc recommendations for all ages. The genotypes showed similar biomass and CP yield under W2 and W1, indicating moderate water stress can sustain yields. These findings highlight the importance of strategic water management for food and fodder security, while meeting nutritional needs in water-scarce regions.
Similar content being viewed by others

Introduction
The Global Aridity Index suggests that dryland production of food will gradually expand, despite the challenge of changing climates1. South Africa, already ranked as one of the thirty driest countries globally2, is already experiencing water stress and faces the looming possibility of worsening aridity towards 20503. Predictions indicate that by 2030, South Africa is projected to experience a significant increase in both human population and livestock numbers. With the population anticipated to grow considerably, reaching an estimated 68.8 million people with rapidly increasing in-migration4, the demand for food, water and resources will intensify. Concurrently, the livestock sector is expected to expand to meet the growing protein requirements of the populace, with a projected increase in cattle and beef consumption5.
The global challenge of food insecurity is pressing, with the imperative to guarantee universal access to a substantial, safe and nutritious food supply more crucial than ever. Regarding the severity of adverse impacts concerning Sustainable Development Goal (SDG) 2, aiming for Zero Hunger for all by 2030, global food insecurity is significant. Currently, 720 to 811 million individuals suffer from undernourishment, with around 2.3 billion people affected by malnutrition6. According to the Food and Agricultural Organisation6, drought caused a 34% drop in crop and livestock production in the least developed and lower-middle-income countries, resulting in a $37 billion loss. Furthermore, in impoverished rural areas of South Africa, drought has resulted in higher livestock mortality rates due to the unavailability of forage, adversely impacting food supply and increasing food prices7. In addition to drought, warmer temperatures are expected to promote the growth of woody plants over grass in certain grasslands of South Africa, potentially worsening the challenge of providing adequate feed for livestock8. This prediction highlights the urgent need for the country to support a larger population amid weakening water resources sustainably. However, the solution extends beyond mere quantity; it requires intensive effort to produce highly nutritious crops efficiently.
Since the United Nations General Assembly adopted the SDGs in September 2015, several strategies have been implemented to achieve SDG 1 (No Poverty) and SDG 2 (Zero Hunger). Amidst South Africa’s water stress and the impending rise in population, the narrative shifts towards exploring viable strategies to navigate this challenge. Water use efficiency (WUE) and Water Productivity (WP) emerge as crucial concepts. While WUE focuses on maximising the yield per unit of water used, WP considers the overall output per unit of water consumed, including crop and livestock production2. In crop production, selecting the right genotype is one of the most critical agronomic practices for achieving high productivity9. The choice of an optimal crop variety depends primarily on its ability to efficiently utilise nutrients10,11, its resilience to environmental stresses12, and its overall yield potential13. Hence, the selection of crops becomes pivotal, emphasising the importance of choosing varieties that not only meet human nutritional needs but also cater to the requirements of livestock, ensuring a holistic approach to water management and food security.
Other approaches include climate-smart forages or crops, dietary modifications, distributing vitamin-rich supplements and fortifying staple foods. However, during the COVID-19 period in South Africa, while many supplemented with vitamin C, rural resource-poor households with limited resources could not afford these supplements. Since 34% of rural South African families rely on agriculture, implementing food-based initiatives could alleviate nutritional food insecurity in the country14. With a focus on utilising sweet potato leaves as a source of green vegetables during the summer season to alleviate food scarcity, Nyathi15 examined the feasibility of integrating sweet potatoes into both animal feed and human diets. Senekal16 recommended consuming dark green vegetables, including Brassicaceae, for food nutritional security in South Africa.
In South Africa’s water scarcity and the need for efficient water management strategies, the relevance of fodder radish emerges as a potential solution. To ensure the efficient use of scarce water resources, it is critical to implement strategies that conserve water while maintaining crop productivity17. One effective approach is deficit irrigation, where water is applied at levels below the crop’s full requirement, a practice that can improve water-use efficiency in irrigation programs18,19. However, this practice often reduces soil moisture, limiting nutrient availability and uptake20. Moreover, insufficient water supply can expose plants to drought stress, leading to nutrient imbalances that ultimately decrease both yield and crop quality21.
Fodder radish, known for its deep taproots and ability to thrive in diverse soil conditions, holds promise as a water-efficient crop. Therefore, there is a pressing need to evaluate the WUE of fodder radish genotypes and assess their suitability in meeting the nutritional requirements of humans and livestock. As far as it can be ascertained, there is no study has evaluated the dual usage (leafy vegetable and storage root) of fodder radish, and these genotypes were bred for biomass enhancement22. Findings of this study will serve as a benchmark. The study’s objectives were (i) to assess the effect of different water regimes on the growth, biomass accumulation, and nutrient content of fodder radish, and (ii) to examine the impact of leaf harvesting on tuber yield. We hypothesised that: (i) leaf harvesting will reduce the tuber yield of fodder radish, and (ii) no leaf harvesting combined with a well-watered irrigation regime will improve the tuber yield of fodder radish. Additionally, a well-watered regime is expected to enhance growth, biomass accumulation, and nutrient content in fodder radish.
This study revealed several constraints that may affect the generalisability and practical applicability of its findings. First, it was conducted at a single site (Roodeplaat, Pretoria, South Africa), thereby limiting the generalisability of the findings to regions with differing climatic, soil, and environmental conditions. The study did not examine the impact of various cooking methods, such as boiling, frying, steaming, or grilling, on the nutrient composition of fodder radish, thereby leaving a gap in the understanding of its nutritional value in practical dietary contexts. The study focused solely on two genotypes, “LINE 2” and “ENDURANCE,” thereby restricting insights into the performance of other potentially relevant genotypes. While covering two seasons, it did not account for the variability of long-term climatic conditions, potentially limiting the applicability of its findings in extreme or exceptional weather situations. While nutrient concentrations were evaluated, the study did not examine the bioavailability of these nutrients to humans or cattle, which may vary based on factors such as soil characteristics, genetic factors, and water stress conditions.
Materials and methods
Site description and soil quality attributes
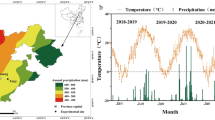
The experiment was conducted in Roodeplaat, Pretoria, Gauteng Province (25° 60′ S; 28° 35′ E; 1168 m a.s.l.), South Africa. While normal field conditions were used, rainfall data were excluded since the rain shelter is designed to close when rainfall starts23 (Fig. 1a,b). Nyathi15 presented long-term climate data from 1990 to 2015, showing that a place receives 650 mm of summer rainfall annually, most occurring between October and March, the main summer rainfall season. A maximum average temperature of 30 °C is normally experienced during January. Soil classification, done according to the USDA24, indicated that the soil is a sandy clay loam with a field capacity of 291 mm m− 1 and a permanent wilting point of 20 mm m− 1. The chemical properties of the topsoil (0.3 m) were used as a guide to supply fertiliser requirements for radish (Table 1).

Rain shelter site: (a) Rain shelter closed during rainfall, (b) Rain shelters are open during non-rainy periods22.
Plant material
Germplasm was obtained from the Agricultural Research Council (ARC) of South Africa (API Cedara seed bank) to develop the two genotypes used for the study: ENDURANCE and LINE 2. The genotypes were bred to provide high-quality forage for grazing livestock in late winter. They were derived from a cross between a very late flowering fodder radish line “PG 1” from Pyne Gould Wrightson Seeds, New Zealand, and ARC- AP Cedara fodder radish genotypes Geisha and Sterling. ENDURANCE and LINE 2 were selected for late flowering and high-yielding traits. These fodder radish genotypes have similar characteristics to annual cool season crops, hairless soft leaves, a shoot system (broad-leafy plants), and a root system (large tube but differed in size). However, they differ in flowering behaviour and flower colour22.
Trial layout and design
The experiment was conducted under a rain shelter at ARC-VIMP Roodeplaat, Pretoria, South Africa, across two winter seasons (2021/2022 and 2022/2023) using a 3 × 2 × 2 factorial design. The factors were three irrigation water regimes [well-watered (W1), moderate water stress (W2), and severe water stress (W3)], two fodder radish varieties [variety 1 (ENDURANCE) and variety 2 (Line 2)], and leaf harvesting [no leaf harvesting (H1) and leaf harvesting (H2)]. The experiment was a randomised complete block design, replicated three times. Table 2 presents the meteorological conditions [maximum and minimum temperatures (°C), total solar radiation (MJ m− 2), cumulative reference evapotranspiration (mm), U2 Average Wind Speed (ms− 1), CU Total Cold Units (unitless), HU Total Heat Units (unitless) and vapour pressure deficit (kPa)]. Total rainfall (mm) was excluded as the rain shelter excluded rain (Fig. 1). Before planting, aluminium access tubes were installed in the middle of each plot to a depth of 1 m. A neutron water meter (CPN, 503 DR Hydroprobe, USA) calibrated for the site with wet and dry profile measurements was utilised to measure soil water content. Compensating non-leaking (CNL) Urinam dripper lines maintain an irrigation discharge rate of 2.3 L per hour. Irrigation scheduling was based on irrigation regimes (W1, W2 and W3).
Irrigation management and scheduling
For plant establishment, fodder radish crops were irrigated with 14 mm water over three weeks in the 2021/22 season. In the 2022/23 season, they received 13.8 mm of water over the same period before the treatments were imposed. Irrigation was applied three times every week. Plants were irrigated in the mornings to ensure water availability during peak demand. The total amount of irrigation water applied was recorded for all irrigation levels, including water applied before the introduction of treatments. A neutron probe reading monitored the soil water status during the growing period. Plants were irrigated to refill capacity when they experienced a 30% loss of available water (W1), and the same procedure was followed for 50% (W2) and 80% (W3) loss. The field capacity was 291 mm m− 1 and a permanent wilting point of 20 mm m− 1. During the trial, treatments received different water levels; for the 2021/2022 season, W1 = 305 mm, W2 = 221 mm and W3 = 180 mm. and for the 2022/2023 season, W1 = 366 mm, W2 = 245 mm and W3 = 188 mm.
Soil water content
Figure 2 shows that the interaction between soil water content and season was not significantly different (P = 0.54). However, the water regime and season were significantly different (P < 0.0001). During the 2021/22 season, soil water content was 42 mm for well-watered, 36 mm for moderate stress, and 28 mm for severe stress. In the 2022/23 season, values increased to 47 mm (well-watered), 40 mm (moderate stress), and remained at 28 mm (severe stress). Overall, soil water content was higher in the 2022/23 season, regardless of the water regime.

Soil water content on different water regimes during S1 = 2021/22 and S = 2022/23 season, Error bars represent standard errors of the means. The letters above the error bars indicate significant differences (P < 0.0001).
Agronomic practice and data collection
Before initiating land preparation, soil samples were collected from each plot using a 30 cm auger for subsequent chemical analysis (Table 1). The land underwent mechanical ploughing before planting seeds. Seeds were sown in individual plot size of 4.6 m2 with inter-row and intra row spacing of 0.3 m × 0.3 m making it a total of 111.111 plants ha− 1. Manual weed removal occurred before planting and continued throughout the experiment. Hand sowing was performed at a depth of 1 cm, and twelve plants were tagged to measure plant height and leaf number.
Throughout the growing seasons, the Leaf Area Index (LAI) was measured at two-week intervals utilising the LAI-2000 canopy analyser (Licor, Lincoln, NE, USA). Chlorophyll content index (CCI) readings were obtained using a SPAD-502 Plus chlorophyll meter (Konica Minolta, Osaka, Japan) on the adaxial surface of the first fully expanded, fully exposed leaf between 13:00 and 15:00, from crop establishment to the end of flowering. Stomatal conductance measurements were recorded 4, 8, and 12 weeks after planting with the SC-1 leaf porometer (Decagon Devices, Pullman, WA, USA).
Fodder radish leaves were harvested four times during the growing seasons, with data from the middle rows (1.8 m2) utilised to avoid border effects. Fresh mass was determined by weighing freshly harvested aboveground biomass (AGB, including leaves, stems, and tuber) and aboveground edible biomass (AGEB, leaves only without stems). Subsequently, samples underwent oven drying at 75 °C for 3–4 days, and the dry biomass was measured. Various plant parameters, including Relative Crop Growth (RGR), Relative Height Rate (RHR), Harvest Index (HI), Fractional Interception of Photosynthetically Active Radiation (PAR), Radiation Use Efficiency (RUE), and Water use efficiency (WUE) were all calculated (Table 3). Canopy extension coefficient (k) values for PAR were obtained from Archontoulis25. Plant height measurements were taken using a tape measure, and the number of leaves counted at harvest occurred after four months. Tubers’ length and diameter were measured using a ruler and digital vernier calliper. Each plant was separated into leaves and tubers, placed in labelled sampling bags, and weighed to determine fresh mass. Dry mass was determined after drying the leaf plant material at 75 °C for 3–4 days and tubers at 75 °C for 4–5 days.
Determination of β-carotene, vitamin C, vitamin E, iron and zinc mass concentrations
The aboveground edible biomass (AGEB) of fodder radish was separated into leaf blades and petioles. Leaf blades, collected at 12 weeks post-planting, were thoroughly washed with distilled water to remove debris. Subsequently, 500 g of the samples were placed in transparent, airtight plastic polythene bags and promptly transported to ARC-VOP laboratories to analyse β-carotene, vitamin C, vitamin E, iron, and zinc mass concentrations14. Iron and zinc mass concentrations were determined following the method recommended by the Association of Official Analytical Chemists26. The leaf samples were oven-dried at 80 °C for 24 h, and their water contents were calculated. The samples were stored in airtight containers after grinding in a Wiley mill with a number 20 stainless steel sieve. The protocol by the Association of Official Analytical Chemists26 was followed to determine iron and zinc nutrient concentration using an inductively coupled plasma atomic emission spectrometer. For β-carotene, vitamin C, and vitamin E analysis, duplicate samples were processed at the ARC-VOP biotechnology laboratory. Fresh mass was determined, and the samples were frozen and stored at −80 °C before freeze-drying. The extraction of β-carotene, vitamin C, and vitamin E was performed using tetrahydrofuran methanol (1:1 vol/vol), following the method outlined by Moyo27. Extracts were analysed using an HPLC-DAD system (Shimadzu, Kyoto, Japan) at a wavelength of 450 nm. A 5-point standard curve covering the concentration range of the samples was constructed for the quantitative analysis of β-carotene, vitamin C, and vitamin E.
The possible contribution to human nutrition
The concentration of β-carotene was transformed into Vitamin A (µg RAEs - retinol activity equivalents), following the guidelines of Trumbo28, where 1 µg RAE equals 1 µg retinol, corresponding to 12 µg of β-carotene. The daily recommended nutrient intakes (DRNI) for iron, zinc, β-carotene, and vitamins C and E across different age groups were obtained from Uusiku29. The percentage contribution to DRNI was determined by dividing the nutrient concentrations (iron, zinc, vitamins C, E and β-carotene, mg 100 g⁻¹) by the nutrient requirements in mg day⁻¹, then multiplied by 100. To assess the potential contribution of 1 hectare for a family of six individuals, nutritional yield data (iron, zinc, vitamin C, E and β-carotene NYs, g ha⁻¹) were multiplied by 10 (mg day⁻¹ ha⁻¹) and divided by DRNI (mg day⁻¹ per person).
The vitamin A, C, and E, iron, and zinc requirements for a family of six individuals
“Vitamin A requirements for a household of six [(one adult male = 600 µg RAE day− 1; one adult female = 500 µg RAE day− 1; two 1–3-year infants (2 × 400 µg = 800 µg RAE day− 1); two 4 -18-year children (2 × 600 µg = 1200 µg RAE day− 1); total = 3100 µg RAE day-1)]. For vitamin C requirements for a household of six [(one adult male = 90 mg day− 1; one adult female = 75 mg day− 1; two 1–3-year infants (2 × 15 mg = 30 mg day− 1); 4–18-year children (2 × 65 mg = 130 mg day − 1); total = 334 mg day− 1). Vitamin E [(one adult male = 15 mg day− 1; one adult female = 15 mg day− 1; two 1–3-year infants (2 × 6 mg = 12 mg day− 1); 4–18-year children (2 × 15 mg = 60 mg day− 1); total = 102 mg day− 1). Iron requirements for a household of six [(one adult male = 13.7 mg day− 1; one adult female = 29.7 mg day− 1; two 1–3-year infants (2 × 5.8 mg = 11.6 mg day− 1); 4–18-year children (2 × 32.7 mg = 65 mg day− 1); total = 120 mg day− 1)]. Zinc requirements for a household of six [(one adult male = 14 mg day− 1; one adult female = 9.8 mg day− 1; two 1–3-year infants (2 × 8.3 mg = 16.6 mg day− 1); 4–18-year children (2 × 14.4 mg = 28.8 mg day− 1); total = 69.2 mg day− 1)]”. The approach used for these calculations followed the methodology outlined by Nyathi14.
The possible contribution to livestock nutrition
The percentage contribution to livestock requirement was determined by dividing the nutrient concentrations (CP mg 100 g⁻¹) by the nutrient requirements in mg day⁻¹, then multiplied by 100. To assess the potential contribution of 1 hectare for six livestock, nutritional yield data (CP NYs, g ha⁻¹) were multiplied by 10 (mg day⁻¹ ha⁻¹) and divided by the recommended CP level (mg day⁻¹ per class of animal). Crude protein requirements for cattle, goat and sheep [(2 x cattle (7000 mg 100 g⁻¹ = 1400 mg 100 g⁻¹; 2 x goat (7000 mg 100 g⁻¹ DM; 2 x = 1400 mg 100 g⁻¹; sheep (7000 mg 100 g⁻¹ DM = 1400 mg 100 g⁻¹; total = 4200 mg 100 g⁻¹. We considered that these values may vary based on factors such as growth stage, lactation, pregnancy, and the specific nutritional needs of individual animals. Livestock requirements are based on the recommended levels for individual classes of ruminants suggested by NRC30 and summarised by McDowell31, while the CP was adapted from Ncisana22.
Statistical analysis
The IBM SPSS statistical software version 25 (https://www.ibm.com/spss) was used to conduct statistical analysis. Soil water content and CP nutritional yield were subjected to the two-way ANOVA, water regimes and season as fixed factors. Variables that were not normally distributed were successfully log10 transformed for analysis. The fresh tuber biomass, dry tuber biomass, tuber diameter, and tuber length were subject to analysis of variance four-way ANOVA, with water regimes, genotype, harvest, and season as fixed factors. Then, a three-way ANOVA was used to test the effects of water regimes, genotype, and season on LI, FI, RUE, AGB, AGEB, HI, RGR RHR, LAI, WUE, β-carotene, vitamin C, vitamin E, Zn, Fe, nutritional yield, stomatal conductance and chlorophyll content. The LSD post hoc test was used to compare means. Statistical significance was determined (P < 0.05).
Results
Stomatal conductance, total biomass, above ground biomass, above edible biomass, tuber yield, harvest index and water use efficiency
Stomatal conductance
Stomatal conductance showed no significant differences (P = 0.489) among water regimes, genotypes, or seasons; however, there was a significant interaction (P < 0.0001) between water regime and genotype was found. Additionally, water regime and season significantly (P < 0.0001) affected stomatal conductance. In the 2021/22 and 2022/23 seasons, well-watered, moderate stress, and severe stress treatments differed significantly (P < 0.05), with severe stress showing higher mean values of stomatal conductance than well-watered and moderate stress in both seasons (Fig. 3a,b).

Effect of water regimes and genotypes on stomatal conductance during the season (a) 2021/22 and (b) 2022/23, W1; well-watered; W2; moderate water stress; W3; severe water stress, V1; ENDURANCE and V2; LINE 2; Error bars represent standard errors of the means. The letters above the error bars indicate significant differences (P < 0.05).
Total biomass
There was no interaction effect among water regime, genotype, and season on the total biomass (P = 1.000). However, the water regime significantly (P = 0.010) influenced total biomass, with moderate stress resulting in higher biomass, though not significantly different from the well-watered treatment for both genotypes in both seasons (Table 4). In the 2021/22 season, LINE 2, under moderate stress, produced 1036.68 kg ha− 1 and ENDURANCE 943.90 kg ha− 1. In the 2022/23 season, LINE 2 had 1227.73 kg ha− 1, and Endurance 820.03 kg ha− 1 under the same conditions (Table 4). Severe water stress significantly reduced total biomass in both genotypes. Fore example, LINE 2 declined by ≈ 60.09% in 2021/22 and ≈ 71.06% in 2022/23, whereas ENDURANCE reduced by ≈ 63.9% and ≈ 53.33%, respectively.
Above ground biomass (AGB)
There was no significant interaction (P = 0.100) effect among water regime, genotype and season. Only the water regime significantly (P < 0.001) affected AGB. In 2021/22, W1 and W2 were similar but higher than W3 for both genotypes (Table 4). In 2022/23, LINE 2 had higher biomass under well-watered conditions, while ENDURANCE performed best under moderate stress. LINE 2 produced 2352.67 kg ha− 1 (W1), 2490.25 kg ha− 1 (W2), and 1204.83 kg ha− 1 (W3) in 2021/22, and 2824.77 kg ha− 1, 2025.27 kg ha− 1, and 955.57 kg ha− 1 in 2022/23. ENDURANCE yielded 2428.95 kg ha− 1, 2524.73 kg ha− 1, and 789.45 kg ha− 1 in 2021/22, and 2487.43 kg ha− 1, 2755.30 kg ha− 1, and 1059.27 kg ha− 1 in 2022/23. Severe water stress reduced AGB in LINE 2 by ≈ 48.91% (2021/22) and ≈ 71.06% (2022/23), while ENDURANCE showed reductions of 63.09% (2021/22) and ≈ 53.32% (2022/23).
Above ground edible biomass (AGEB)
There was no interaction effect observed among water regime, genotype, and season on the AGEB (P = 0.334). Only the water regime significantly (P < 0.001) affected AGEB. LINE 2 showed no difference between well-watered and moderate stress in both seasons. In the first season, yields were 668.94 kg ha− 1 (well-watered), 603.79 kg ha− 1 (moderate stress), and 467.91 kg ha− 1 (severe stress). In the second season, the trend continued with 972.28 kg ha− 1, 997.31 kg ha− 1, and 495.63 kg ha− 1, respectively (Table 4). For ENDURANCE, moderate stress produced the highest biomass in both seasons, with 1053.62 kg ha− 1 in 2021/22 and 1161.38 kg ha− 1 in 2022/23, while severe stress yielded the lowest, at 251.59 kg ha− 1 and 426.55 kg ha− 1, respectively (Table 4). LINE 2 resulted in AGEB reductions of ≈ 30.05% in 2021/22 and ≈ 49.02% in 2022/23 under severe water stress. ENDURANCE showed larger declines, with ≈ 74.23% in 2021/22 and ≈ 53.35% in 2022/23.
Tuber yield
Effect of water paired with no leaf harvest
There was no significant effect (P = 1.000) was observed on tuber yield in terms of fresh and dry weight concerning water regime, genotype and season. However, the water regime significantly (P < 0.001) affected tuber yield, both as fresh and dry tuber weights. In the 2021/22 season, LINE 2 yielded 55 t ha− 1 under well-watered conditions, 48 t ha− 1 under moderate stress, and 22 t ha− 1 under severe stress. In 2022/23, yields were 56 t ha− 1 (well-watered), 48 t ha− 1 (moderate water stress), and 21 t ha− 1 (severe water stress). For ENDURANCE, the 2021/22 season yielded 59 t ha− 1 (well-watered), 48 t ha− 1 (moderate water stress), and 22 t ha− 1 (severe water stress). In 2022/23, ENDURANCE produced 59 t ha− 1 (well-watered), 20 t ha− 1 (moderate water stress), and 8 t ha− 1 (severe water stress). Well-watered and moderate stress treatments showed no significant differences but yielded more than severe stress (Fig. 4a,b). Severe stress sharply reduced tuber yield (fresh weight), with ENDURANCE reduced by ≈ 61.88% in 2021/22 and ≈ 59.83% in 2022/23, and LINE 2 decreasing ≈ 61.49% and ≈ 61.5%, respectively. For dry weight, ENDURANCE declined ≈ 42.8% in 2021/22 and ≈ 51.98% in 2022/23, while LINE 2 reduced by ≈ 54.78% and ≈ 50.59% in those seasons (Fig. 4c,d).

Effect of leaf harvesting and different water regimes on tuber yield data as fresh and dry in season 2021/22 and season 2022/23, W1 – H1; well-watered with leaf harvest, W1 – H2; well-watered with no leaf harvest, W2- H1; moderate water stress with no leaf harvest; W2- H2; moderate water stress with no leaf harvest, W3-H1; severe water stress with leaf harvest; W3-H2; severe water stress with no leaf harvest. Genotypes V1 = ENDURANCE, V2 = LINE 2; Error bars represent standard errors of the means. The letters above the error bars indicate significant differences (P < 0.05).
Effect of leaf harvesting on tuber yield
There was no significant (P = 1.000) interaction effect among harvest, genotype and season on fresh or dry tuber weight. However, for both seasons, leaf harvesting significantly (P < 0.001) affected fresh tuber yield. Treatments without leaf harvesting produced higher mean tuber yields compared to those with leaf harvesting (Fig. 4a,b). Leaf harvest reduced tuber yield by ≈ 53.19% in ENDURANCE and ≈ 57.55% in LINE 2. For dry tuber yield, it caused a reduction of ≈ 56.25% in ENDURANCE and ≈ 41.4% in LINE 2 (Fig. 4c,d).
Harvest index (HI)
There was no significant (P = 0.150) interaction that was observed among water regime, genotype and season for the harvest index (HI). Water regime significantly influenced HI, with LINE 2 showing higher values under severe stress than moderate and well-watered conditions in both seasons. For ENDURANCE, well-watered treatment produced higher yields in the first season (P < 0.05) and slightly higher yields in the second season, though not significantly different from moderate stress. LINE 2 under well-watered conditions reduced HI by ≈ 22.12% in 2021/22 and ≈ 32.48% in 2022/23, while ENDURANCE resulted in reductions of ≈ 43.66% and ≈ 51.07% under severe stress (Table 4).
Water use efficiency (WUE)
The WUE did not show a statistically significant (P = 0.993) interaction among water regime, genotype, and season. Only the water regime significantly (P = 0.030) impacted WUE. The highest WUE was attained at the moderate water stress, whereas the lowest WUE was attained at the severe stress (Table 4). There was seasonal variation; for example; WUE was higher in 2021/22 than in 2022/23. Severe stress reduced WUE for line 2 by ≈ 51.59% in 2021/22 and ≈ 63.78% in 2022/23. For endurance, it was reduced by ≈ 58.04% in 2021/22, improving to ≈ 15.16% in 2022/23.
Crop growth
Tuber diameter
No statistical effect was observed on the tuber diameter concerning water regime, genotype, harvest and season (P = 1.000). However, the water regime significantly influenced storage root yield diameter (P < 0.001). Well-watered and moderate water stress treatments did not display significant differences. Still, they significantly showed larger diameters than severe water stress conditions (Fig. 5). Severe water stress resulted in tuber diameter reduction, with ENDURANCE experiencing decreases of ≈ 34.61% in the 2021/22 season and ≈ 34.46% in the 2022/23 season. Similarly, LINE 2 had reductions of ≈ 30.95% in the 2021/22 season and ≈ 30.79% in the 2022/23 season. The harvesting alone had a statistically significant effect on storage root diameter for both seasons (P < 0.001), with no harvesting resulting in improved values compared to leaf harvest. Leaf harvesting led to a reduction of ≈ 32.08% in ENDURANCE and ≈ 36.88% in LINE 2.

Effect of leaf harvesting and different water regimes on tuber diameter in season 2021/22 and season 2022/23, W1-H1; well-watered with leaf harvest, W1-H2; well-watered with no leaf harvest, W2- H1; moderate water stress with no leaf harvest; W2- H2; moderate water stress with no leaf harvest, W3-H1; severe water stress with leaf harvest; W3-H2; severe water stress with no leaf harvest. Genotypes V1 = ENDURANCE, V2 = LINE 2; Error bars represent standard errors of the means. The letters above the error bars indicate significant differences (P < 0.05).
Tuber length
No interaction effect was observed among water regime, genotype, harvest and season on storage root length (P = 1.000). None of the fixed factors affected storage root length including water regime (P = 0.197) (Fig. 6).

Effect of leaf harvesting and different water regimes on tuber length in season 2021/22 and season 2022/23, W1-H1; well-watered with leaf harvest, W1-H2; well-watered with no leaf harvest, W2- H1; moderate water stress with no leaf harvest; W2- H2; moderate water stress with no leaf harvest, W3-H1; severe water stress with leaf harvest; W3-H2; severe water stress with no leaf harvest. Genotypes V1 = ENDURANCE, V2 = LINE 2; Error bars represent standard errors of the means. The letters above the error bars indicate significant differences (P < 0.05).
Relative growth rate (RGR)
The relative growth rate (RGR) showed no significant interaction among water regime, genotype, and season (P = 1.000). Only the water regime significantly affected RGR (P = 0.020), with well-watered and moderate stress conditions yielding higher values than severe stress across genotypes and seasons (Table 5).
Relative height rate (RHR)
The relative height rate (RHR) showed no significant interaction among water regime, genotype, and season (P = 0.250). Only the water regime had a significant effect on RHR (P < 0.0001), with well-watered and moderate stress conditions yielding higher values than severe stress across genotypes and seasons (Table 5).
Leaf area index (LAI)
Leaf area index (LAI) showed no interaction effect among water regime, genotype, and season (P = 0.665). Water regime significantly (P = 0.030) affected LAI, and season had a statistical effect (P < 0.035) on LAI, with well-watered having higher values but not significantly different from moderate stress for both seasons (Table 5).
Crop height
The was no significant (P = 0.890) interaction effect among water regime, genotype and season. The water regime significantly (P = 0.01) influenced the crop height. While well-watered and moderately stressed conditions were not significantly different for both seasons (P < 0.05), ENDURANCE consistently showed taller growth compared to LINE 2, especially in the second season of 2022/23 (Table 5).
Leaf number
No significant interaction was observed among water regime, genotype and season for the number of leaves (P = 1.000). However, the water regime significantly impacted the number of leaves (P < 0.002), with LINE 2 producing one more leaf than ENDURANCE (Table 5).
Chlorophyll content
Figure 7. Effect of water regimes and genotypes on chlorophyll content during the season (a) 2021/22 and (b) 2022/23, W1; well-watered; W2; moderate water stress; W3; severe water stress, V1; ENDURANCE and V2; LINE 2. Error bars represent standard errors of the means. The letters above the error bars indicate significant differences (P < 0.05).

Effect of water regimes and genotypes on chlorophyll content during the season (a) 2021/22 and (b) 2022/23, W1; well-watered; W2; moderate water stress; W3; severe water stress, V1; ENDURANCE and V2; LINE 2. Error bars represent standard errors of the means. The letters above the error bars indicate significant differences (P < 0.05).
Light intercepted (LI), radiation use efficiency (RUE) and fractional interception (FI)
The interaction effect between water regime, genotype, and season did not significantly impact (P < 0.05). However, the water regime and the season individually had a significant effect (P < 0.05) on light interception (LI) (Table 5). While well-watered and moderately stressed treatments did not significantly differ (P < 0.05) from each other, they differed significantly from severely stressed treatments. Fractional interception (FI) showed a similar pattern regarding interaction effects, with the water regime significantly (P < 0.05) influencing FI. Radiation use efficiency (RUE) was only influenced by season and water (P < 0.05), without the second season possessing relatively higher values (Table 5).
Micronutrients, β-carotene, vitamins C & E, nutritional yield
In the case of zinc, there was no interaction was observed among water regime, genotype, and season (P = 0.77). Only water regimes had statistical influence (P < 0.001) (Table 6). ENDURANCE showed 135.36 to 132.4 mg 100 g− 1 DW levels under well-watered conditions, 108.70 to 104.4 mg 100 g− 1 DW under moderate stress, and 46.3 to 44.3 mg 100 g− 1 DW under severe water stress across both seasons. LINE 2 displayed levels of 136.93 to 133.93 mg 100 g− 1 DW under well-watered conditions, 107.23 to 104.40 mg 100 g− 1 DW under moderate stress, and 52.03 to 49.87 mg 100 g− 1 DW across the seasons 2021/22 and 2022/23, respectively.
Iron
Iron showed no interaction among water regime, genotype, and season (P = 1.000). The interaction between genotype and water was evident (P < 0.001) and water had statistically influenced the iron content (P < 0.001). ENDURANCE showed levels of 88.97 to 83.96 mg 100 g− 1 DW under well-watered conditions, 82.20 to 77.20 mg 100 g− 1 DW under moderate stress, and 92.03 to 87.03 mg 100 g− 1 DW under severe water stress for seasons 2021/22 and 2022/23. LINE 2 showed 100.9 to 95.9 mg 100 g− 1 DW levels under well-watered conditions, 100.7 to 95.7 mg 100 g− 1 DW under moderate stress, and 85.2 to 80.2 mg 100 g− 1 DW under severe water stress across both seasons (Table 6).
Regarding β-carotene, there was no significant interaction effect among water regimes, genotype, and season (P = 1.000) (Table 6). However, an interaction between the water regime and genotype affected β-carotene (P < 0.05), and the water regime influenced the β-carotene (P < 0.001) (Table 6). Across both seasons, the ENDURANCE showed higher concentrations under well-watered conditions than water stress treatments. LINE 2 showed concentrations of 40.43 mg 100 g− 1 DW and 37.2 mg 100 g− 1 DW, while ENDURANCE displayed 46.6 mg 100 g− 1 DW and 31.4 mg 100 g− 1 DW for the seasons 2021/2022 and 2022/2023, respectively. Under moderate stress, LINE 2 demonstrated concentrations of 29.6 mg 100 g− 1 DW and 30.6 mg 100 g− 1 DW, while ENDURANCE showed 29.8 mg 100 g− 1 DW and 30.8 mg 100 g− 1 DW for the seasons 2021/2022 and 2022/2023, respectively (Table 6). Severe water stress resulted in concentrations of 30.23 mg 100 g− 1 DW and 33.93 mg 100 g− 1 DW for LINE 2, and 29.00 mg 100 g− 1 DW and 29.60 mg 100 g− 1 DW for ENDURANCE for the seasons 2021/2022 and 2022/2023, respectively.
There was no significant (P = 1.000) difference in vitamin C concentration between the water regime, genotype, and season interaction. Instead, only the water regime showed an effect (P < 0.0001) (Table 6). Well-watered conditions significantly differed from moderate and severe stress treatments (Table 6). Under well-watered conditions, the LINE 2 genotype exhibited 31.1 mg 100 g− 1 DW concentrations for 2021/2022 and 27.3 mg 100 g− 1 DW for 2022/2023. In contrast, ENDURANCE displayed 31.16 mg 100 g− 1 DW in 2021/2022 and 26.5 mg 100 g− 1 DW in 2022/2023. Moderate stress conditions resulted in LINE 2 producing 24.6 mg 100 g− 1 DW for 2021/2022 and 26.5 mg 100 g− 1 DW for 2022/2023. Conversely, ENDURANCE yielded 26.5 mg 100 g− 1 DW for 2021/2022 and 29.33 mg 100 g− 1 DW for 2022/2023. In the severe stress treatments, LINE 2 showed 23.8 mg 100 g− 1 DW concentrations for 2021/2022 and 22.6 mg 100 g − 1 DW for 2022/2023. Meanwhile, ENDURANCE resulted in 22.3 mg 100 g− 1 DW for 2021/2022 and 26.5 mg 100 g− 1 DW for 2022/2023.
The vitamin E showed no significant (P = 1.000) difference among water regimes, genotype, and season. Only water regime significantly (P < 0.0001) influenced the vitamin E concentration, with moderate stress treatment demonstrating significantly higher concentrations compared to both the well-watered and severe stress treatments (Table 6).
LINE 2 displayed concentrations of 547.57 mg 100 g− 1 DW for the season 2021/2022 and 476.50 mg 100 g− 1 DW for the season 2022/2023 under well-watered conditions, while ENDURANCE showed 548.03 mg 100 g− 1 DW for the season 2021/2022 and 345.00 mg 100 g− 1 DW for the season 2022/2023. Under moderate stress, LINE 2 showed concentrations of 644.80 mg 100 g− 1 DW for the season 2021/2022 and 686.80 mg 100 g− 1 DW for the season 2022/2023, while ENDURANCE displayed 714.20 mg 100 g− 1 DW for the season 2021/2022 and 610.80 mg 100 g− 1 DW for the season 2022/2023.
Severe stress conditions resulted in LINE 2 producing 316.70 mg 100 g− 1 DW for 2021/2022 and 414.90 mg 100 g− 1 DW for 2022/2023. Conversely, ENDURANCE yielded 316.90 mg 100 g− 1 DW for 2021/2022 and 552.6 mg 100 g− 1 DW for 2022/2023.
Significant effects were not observed among water regime, genotype, and season for zinc (P = 1.000) and iron nutritional yield (P = 0.770). Interestingly, genotype alone did not yield statistically significant effects (P = 0.294). Only the water regime affected the nutritional yield of zinc (P < 0.001), demonstrating that the well-watered treatment yielded higher values than both moderate and severe water stress treatments. Severe water stress led to an ≈ 81% reduction in zinc nutritional yield for ENDURANCE across both seasons, while LINE 2 experienced an ≈ 82% reduction in both seasons. Similarly, for iron nutritional yield, significant effects were found between water regime, genotype, and season and between water regime and genotype (P < 0.014).
The water regime significantly influenced iron nutritional yield (P < 0.001), with both well-watered and moderate water stress treatments showing similar and superior values compared to severe water stress treatments, which reduced the concentration by ≈ 60% for ENDURANCE and ≈ 47% for LINE 2 in both seasons (Table 7). The results revealed no significant interactions between water regime, genotype, and season for β-carotene, vitamin C, and vitamin E nutritional yield (P = 1.000). Only the water regime showed a statistically significant difference in all the variables (P = < 0.001) (Table 7). Regarding crude protein (CP) nutritional yield, a statistically significant difference was observed among the water regime and genotype (P = 0.038), whereas genotype alone did not yield a significant effect (P = 0.918). Water regime significantly influenced CP nutritional yield (P < 0.001), with no significant difference observed between well-watered and moderate stress treatments (Fig. 8). In ENDURANCE, severe water stress reduces CP nutritional yield by ≈ 70% and ≈ 54% in LINE 2.

Crude protein nutritional yield (CPNY, g ha− 1) on different water regimes during the 2021/22 season, error bars represent standard errors of the means. The letters above the error bars indicate significant differences (P < 0.0001). Well-watered = W1; moderate water stress = W2 and severe water stress = W3; Genotypes V1 = ENDURANCE, V2 = LINE 2.
Discussion
Drought leads to several critical dysfunctions in the photosynthesis process32, including damage to plant pigments, disruption of stomatal performance and reduced CO2 flow to the photosynthetic machinery32. Additionally, reducing the crop’s water requirement impairs nutrient uptake and utilisation, while disturbing osmotic potential33. The generation of reactive oxygen species (ROS) under deficit irrigation further exacerbates plant damage, injuring pigments, photosynthetic systems, and overall metabolism34. Consequently, deficit irrigation ultimately results in reduced crop growth and yield35,36.
On the other hand, the soil water content showed significant variation among the three water regimes: well-watered, moderate water stress, and severe water stress. As anticipated, the well-watered treatment displayed the highest soil water content, followed by moderate water stress, with severe water stress showing the lowest soil water content. These results align with previous findings14,37, confirming the expected relationship between water regime and soil water content.
The reduction in stomatal conductance of selected fodder genotypes under drought stress has been extensively documented in the literature38,39. However, contrary to this prevailing observation, certain studies have reported an improvement in stomatal conductance under moderate and severe water stress conditions40,41, which aligns with the findings of the current study where drought stress led to higher stomatal conductance. This discrepancy may be attributed to the response of crops to stress, wherein their stomata, small pores on the surface of leaves, tend to close to prevent excessive water loss through transpiration42.
The importance of water use efficiency (WUE) in plant production cannot be overstated, as it plays a critical role in ensuring better productivity, especially in challenging environmental conditions43. High WUE allows plants to maintain essential metabolic processes crucial for their growth and development44. Interestingly, Mandal45 observed lower WUE in well-watered plots of certain forage brassicas; this finding was partially supported by our study, where plots under moderate stress exhibited higher WUE compared to both well-watered and severely stressed plots. Similarly, Akram38 documented higher WUE in water-stressed radish plots. However, Henschel39 reported no significant difference in WUE between well-watered and drought-stressed radish plots. Despite limited studies focusing specifically on WUE in fodder radish or forage brassicas, research across various crops indicates an increase in WUE under water-stressed conditions46,23,47.
The reduction in WUE observed under well-watered conditions could be attributed to amplified soil water evaporation rather than transpiration48. Additionally, the higher values of WUE under moderate water stress may be due to more efficient stomatal regulation in water-stressed plots, as the closure of stomata reduces water loss through transpiration. Conversely, well-watered plants might keep their stomata open longer, leading to higher transpiration rates and lower water use efficiency. Water use efficiency measures the biomass or yield produced per unit of water the crop uses. Our expectations were that WUE would be higher under stressed conditions, as the crop is expected to produce more with less water. This higher WUE under stress implies that the crop can make more efficient use of available water resources, potentially leading to increased productivity per unit of water.
When crops experience stress, their stomata, which are small pores on the surface of leaves, tend to close to prevent excessive water loss through transpiration. While this mechanism helps conserve water, it also affects crop productivity. When stomata close, photosynthesis is inhibited, reducing carbon dioxide uptake and decreasing yield potential. The extent of yield reduction under stressed conditions varies depending on factors such as the severity and duration of stress, crop species, and growth stage. In general, prolonged or severe stress can lead to significant yield losses. Our study findings concurred with severe water stress having low biomass and higher stomatal conductance values. However, moderate water stress in both seasons for ENDURANCE resulted in higher biomass. In contrast, for the LINE 2 genotype in both seasons, well-watered and moderate stress conditions resulted in similar biomass. Nyathi14 observed a reduction in both total aboveground and edible aboveground biomass of leafy vegetables under severe water-stressed conditions.
Unsurprisingly, well-watered treatments resulted improved biomass because the leaf area index was improved by well-watered conditions, given that the higher the LAI, the higher the biomass. The higher light use efficiency (LI) might have facilitated increased CO2 assimilation and subsequently improved biomass; this might explain the favourable outcomes in moderate stress and well-watered treatments due to their higher LI, Fractional interception (FI), and superior Radiation Use Efficiency (RUE) in mean values49,60 (Table 4). Throughout both seasons, the average daily maximum temperature remained around 25 °C (Table 2), which remained optimal for radish growth. Stagnari40 reported similar findings in radish genotypes, where well-watered plots exhibited significantly higher mean values for aboveground biomass. The high total biomass under moderate stress in both seasons reveals that these genotypes ENDURANCE and LINE 2 conserve water because for 2021/2022 season W1 = 305 mm, W2 = 221 mm and 2022/2023 season W1 = 366 mm, W2 = 245 mm. Therefore, using these genotypes, farmers can save between 84 and 121 mm to achieve maximum yield. This implies that the goal of WUE in crops is to maximise the yield or productivity of the crop while minimising the amount of water used. In the South African context, which is deemed as a water-stressed country ranked 30th in the world, WUE is a crucial factor for ensuring sustainable water management practices and improving overall crop performance.
The study findings supported our first hypothesis that leaf harvesting significantly reduces fodder radish storage root/tuber yield in the 2021/22 and 2022/23 seasons. This reduction persisted across different irrigation water regimes, highlighting the consistent negative effect of leaf harvesting on root/tuber yield. The observed phenomenon aligns with global trends, as similar root/tuber yield reductions due to leaf harvesting have been documented in different regions. Studies from India50, and Congo51 revealed decreased yields with multiple leaf harvests, while research in South Africa, Nyathi15 reported losses in root or tuber yield with repeated harvesting. However, there were contrasting findings by52, indicating that leaf harvest did not influence root/tuber yield. Hauser53 even noted positive effects on root or tuber yield in specific conditions, claiming that shorter dry seasons could be attributed to their results.
The second hypothesis was that no leaf harvesting paired with a well-watered irrigation water regime would improve fodder radish storage root yield, and the study results did not consistently support it. The well-watered and moderate stress treatments were not significantly different. Both well-watered and moderate stress with no leaf harvest improved root/tuber yield during the 2021/22 and 2022/23 seasons. The unexpected result might be attributed to that radish crops store water in their tubers, allowing them to withstand periods of water stress, so moderate stress conditions positively affected the tuber yield, implying that water applied in the moderate stress plots combined with the water reservoir in the tubers was enough to produced yield same as in the well-watered conditions. Although not all were conducted on fodder radish, some studies align with these findings15,54. Stagnari40 noted the reduction in radish’s root storage/tuber yield under water stress treatments. Access soil resources for regrowth, possibly explaining the lack of significant differences among water regimes, particularly in the fodder radish from the severely water-stressed treatment paired with leaf harvest. This indicates that the ENDURANCE and LINE 2 fodder radish genotypes can achieve higher tuber yield without excessive water requirements. The total water supplied for well-watered conditions was 305 mm and 366 mm for the 2021/22 and 2022/23 seasons, respectively, while moderate stress received 221 mm and 245 mm across both seasons.
Both well-watered and moderate stress, coupled with no leaf harvest, yielded larger tuber diameters, as expected, given the strong correlation between stem diameter and plant biomass7. The well-watered regime improves radish tuber diameter55. Cunha56 found no genotype-specific effects on radish tuber diameter, indicating that the total water applied may have a more pronounced impact, which aligns with our study’s results. In our study results, the lack of genotype impact might be due to limited genomic diversity between ENDURANCE and LINE 2. Tuber length remained unaffected by leaf harvest, water regime, and genotype. The root/tuber, essential for nutrient and water uptake from the soil, may prioritise growth by enlarging to access soil resources for regrowth, possibly explaining the lack of significant differences among water regimes, particularly in the fodder radish from the severely water-stressed treatment paired with leaf harvest. Similar findings of no water regime effects on radish tuber length were reported by57,58, consistent with the results of this study. This indicates that the ENDURANCE and LINE 2 fodder radish genotypes can achieve high tuber yield without excessive water requirements. The total water supplied for well-watered conditions was 305 mm and 366 mm for the 2021/22 and 2022/23 seasons, respectively, while moderate stress received 221 mm and 245 mm across both seasons.
Crop growth indicators such as total aboveground biomass, leaf area index (LAI), harvest index (HI), relative growth rate (RGR), relative height rate (RHR), plant height, and the number of leaves all showcased positive responses to well-watered conditions as hypothesised. Stagnari40 reported similar findings in radish genotypes, where well-watered plots exhibited significantly higher mean values for aboveground biomass, RGR, and LAI. As noted by Silva42, water stress treatments impacted the growth indicators of Salvia hispanica L., including RGR, suggesting a reduction due to water stress. Galmes59 also noted reduced RGR in different genotypes under water-stressed conditions, potentially linked to leaf senescence and decreased leaf area index60.
The reduction in dry weight growth in ENDURANCE and LINE 2 could be largely attributed to diminished light interception, radiation use efficiency, and fractional interception under severe stress treatments. In a radish study, Marcelis and Van Hooijdonk60 observed decreased light interception under water stress treatments, attributing it to low growth rates in the leaf area. Understanding light interception, radiation use efficiency, and fractional interception is pivotal in comprehending the photochemical efficiency of plant photosynthesis. While our findings on light interception align with Nyathi14, differences were noted in radiation use efficiency, where they found a reduction with increased water stress levels, contrary to our study where only season influenced RUE, although on Amaranthus.
The decline in chlorophyll content in Raphanus sativus L. due to drought stress is a well-documented38,39. However, contrary to this, certain studies have reported improved chlorophyll content under moderate and severe water stress conditions40,41. This opposes our study result, which found no effect on water regimes. This discrepancy might be attributed to the inherent genetic traits that ENDURANCE and LINE 2 possess to maintain consistent chlorophyll content levels in different water availability conditions. Similarly, some studies, consistent with our findings, observed no significant difference in chlorophyll content under various water regimes.
Brassica microgreens are a rich source of microelements Fe and Zn61. The lack of micronutrients and vitamin A in human diets leads to “hidden hunger”, a condition whose effects may not manifest immediately. Still, it can result in severe consequences such as stunted growth, delayed cognitive development, and reduced immunity62. Inadequacy of essential nutrients can directly influence the body’s immune response. These nutrients act as antioxidants, safeguarding cells, fortifying immune cells’ growth and function, and initiating antibodies’ production. Our study strongly supports the hypothesis that well-watered conditions lead to elevated zinc concentrations in both genotypes. Our findings align with those of Maseko63, who observed increased zinc concentration under well-watered conditions. However, our results contradict those of Schlering64, who reported a rise in zinc levels under water-stressed treatments in radish leaves. Furthermore, a similar pattern was noted by Nyathi14, where severe water stress resulted in higher concentrations of iron and zinc in Amaranth and Spider flower.
Although the study hypothesis was partially supported for the iron, well-watered and moderately water-stressed treatments exhibited comparable values in LINE 2. Conversely, ENDURANCE under severe water-stressed treatment demonstrated higher values than well-watered and moderately water-stressed treatments. This contradicts the findings of Nyathi14, who reported an approximately 46% decrease in iron concentration for Swiss chard under severe water stress. The disparity between these genotypes could be attributed to the resilience of ENDURANCE to water scarcity, as it maintained or even increased iron concentration as a stress response. At the same time, LINE 2 experienced a decline in iron levels.
Our study results supported our hypothesis that well-watered will improve the β-carotene and vitamin C concentration. The water-stressed treatments reduced the concentration of β-carotene in both genotypes and seasons. Maluleke65 assessed cucumber under different water regimes and found 1.6 to 1.5 mg 100 g− 1 DW β-carotene under moderate and severe water stress treatments; this aligns with our findings. While Park66 found no effect of different water regimes β-carotene of green leafy vegetables. In an extensive review conducted by Gamba67, radish leaves’ mineral and vitamin composition was compared to various vegetables such as cabbage, cauliflower, broccoli, arugula, and turnip. The β-carotene concentration in radish leaves was particularly remarkable, which reached the highest level at 3.96, surpassing other vegetables by up to threefold. This high β-carotene content establishes radish leaves as a significant source of vitamin A within the Brassicaceae family, highlighting their nutritional importance compared to the studied vegetables.
For vitamin C, the current study results showed a reduction in the concentration of vitamin C under water-stress treatments, aligning with the findings of Ahmed68, who observed a decrease in vitamin C in fruits under water-stress treatments. Conversely, Park65 have demonstrated increased vitamin C under water stress conditions. Stagnari69 have acknowledged the limited data reporting on vitamin C under varying water regimes.
Regarding vitamin E, our hypothesis was not supported because moderate water stress treatment had higher values of vitamin E than well-watered treatment. This might be attributed to that moderate water stress conditions impacted photosynthetic processes in plants. Vitamin E protects the photosynthetic apparatus from oxidative damage, and an increase in its levels may be a response to maintaining or enhancing photosynthetic efficiency under stress conditions. An increase in vitamin E in soybean under water stress treatments was found70. This finding aligns with our study results, indicating a higher concentration of vitamin E under moderate stress. However, Oh71, in their study on lettuce, found no effect of water regime on Vitamin C and E. Our findings suggest that under well-watered conditions, both ENDURANCE and LINE 2 varieties can exceed the daily recommended nutrient intake (DRNI) for vitamin A across all age groups. Meanwhile, under moderate stress conditions, they meet the DRNI for the most vulnerable groups, including women and infants aged 1 to 3 years. Under severe water stress conditions, both ENDURANCE and LINE 2 varieties surpass the daily recommended nutrient intake (DRNI) for vitamins E and iron and zinc across all age groups and genders. Vitamin C only meet the daily recommended nutrient intake (DRNI) for 1–3 years old.
Nutritional yield is the amount of essential nutrients a crop provides per area, considering both how much it produces and how concentrated the nutrients are within it14. The study on different types of fodder radish explored how water levels affect the nutritional yield of these crops. Under moderate water stress conditions (W2), the levels of essential nutrients, including vitamin E, CP yield and other important micronutrients, were typically found to be ideal. In contrast, severe water stress (W3) often resulted in lower nutrient levels per unit, but still met the necessary vitamin requirements for extreme dietary needs. These findings align with the observations of Nyathi14, who reported a decrease in zinc and β-carotene nutritional yield under conditions of severe water stress. The results highlight how effectively managing water is crucial for maximising the amount of essential nutrients provided by crops in regions with limited water resources72. Nutritional yield is essential for ensuring food and fodder security, particularly in regions such as South Africa, where dealing with water scarcity necessitates growing crops like fodder radish, valued for efficient water use and nutritional advantages for both humans and livestock. Therefore, nutritional yield is a crucial measure for evaluating how strategic water management and selecting plant varieties contribute to improving the sustainability of diets and agriculture.
Conclusion and future research
This study examined how efficiently water was used, along with the growth of biomass and nutrient levels, in two types of fodder radish, “LINE 2” and “ENDURANCE,” under different water conditions and leaf harvesting practices. Conducted over two growing seasons (2021/22 and 2022/23) in South Africa, the research assessed the impact of well-watered, moderate water stress, and severe water stress environments. The results showed that when experiencing moderate water stress, the efficiency of water use and biomass yield were around 90% compared to full irrigation, while also increasing vitamin E concentrations and CP yield. Under severe stress, total biomass decreased by 30%, tuber yield by 25%, and nutrient content by 15%. LINE 2 showed 5% better resilience than ENDURANCE in this scenario. Additionally, leaf harvesting substantially lowered tuber yield. Our findings proposes that using moderate water stress as an irrigation method can optimise resource use efficiency in environments with limited water supply. For instance, moderate water stress encourages the growth of deeper roots, which enhances water absorption and utilization, ultimately boosting plant health and productivity.
The study concludes that moderate water stress is a sustainable alternative to full irrigation for cultivating fodder radish. It optimises both water use efficiency and nutrient concentration, leading to improved plant health and nutritional value, all while maintaining a competitive yield. Severe water stress had negative effects, leading to a significant decrease in both biomass and nutritional value. Both genotypes showed resilience under moderate stress, but LINE 2 outperformed ENDURANCE in specific aspects. This research contributes to developing climate-smart agricultural practices tailored for arid and semi-arid regions. For instance, implementing precision irrigation techniques can optimise water use in arid environments, ensuring sustainable crop production and food security while preserving soil quality.
Future research should broaden its evaluation to include additional fodder radish genotypes and evaluate their performance over the long term in various climatic conditions. Investigating the effects of different cooking methods on nutrient bioavailability and exploring soil amendments to mitigate nutrient loss under stress conditions are crucial for promoting human health and sustainable agricultural practices. Additionally, integrating multi-season trials across various agro ecological zones will enhance the generalisability of findings. Studying scalable water management technologies and their economic viability for farmers with limited resources will strengthen the position of fodder radish in sustainable agriculture.
Data availability
The datasets utilized and/or analyzed in this study are available from the corresponding author upon reasonable request.
References
Wang, L. et al. Dryland productivity under a changing climate. Nat. Clim. Chang. 12, 981–994. https://doi.org/10.1038/s41558-022-01499-y (2022).
Mabhaudhi, T. et al. Options for improving agricultural water productivity under increasing water scarcity in South Africa. In 3rd World Irrigation Forum (WIF3) on Development for Water, Food and Nutrition Security in a Competitive Environment, Bali, Indonesia 1–7. (2019).
Du Plessis, A. South Africa’s Water predicament: Freshwater’s unceasing decline. 101. (Springer Nature, 2023).
Go, A., Moyer, J., Rafa, M. & Schunemann, J. Population futures: revisiting South Africa’s National Development Plan 2030. Inst. Secur. Stud. Papers (7), 1–16. (2013).
Meissner, H. H., Scholtz, M. M. & Palmer, A. R. Sustainability of the South African livestock sector towards 2050 part 1: worth and impact of the sector. South. Afr. J. Anim. Sci. 43 (3), 282–297 (2013).
FAO The impact of disasters and crises on agriculture and food security. Rome https://doi.org/10.4060/cb3673en (2021).
Ncisana, L. et al. A comparative study of teaching approaches in agro-ecology: an investigation of 10th-grade agricultural sciences learners in selected schools. Sustainability 15 (5). https://doi.org/10.3390/su15054048 (2023).
Ncisana, L., Mkhize, N. R. & Scogings, P. F. Warming promotes growth of seedlings of a woody encroacher in grassland dominated by C4 species. Afr. J. Range Forage Sci. 39 (3), 272–280. https://doi.org/10.2989/10220119.2021.1913762 (2022).
Noureldin, N. A., Saudy, H. S., Ashmawy, F. & Saed, H. M. Grain yield response index of bread wheat cultivars as influenced by nitrogen levels. Ann. Agric. Sci. 58 (2), 147–152. https://doi.org/10.1016/j.aoas.2013.07.012 (2013).
Saudy, H. S., El–Momen, A., El–Khouly, N. S. & W. R., & Diversified nitrogen rates influence nitrogen agronomic efficiency and seed yield response index of sesame (Sesamum indicum L.) cultivars. Commun. Soil Sci. Plant Anal. 49 (19), 2387–2395. https://doi.org/10.1080/00103624.2018.1510949 (2018).
Saudy, H. S., El–Samad, G. A. A., El–Temsah, M. E. & El–Gabry, Y. A. E. G. Effect of iron, zinc, and manganese nano-form mixture on the micronutrient recovery efficiency and seed yield response index of sesame genotypes. J. Soil. Sci. Plant. Nutr. 1–11. (2022).
Shahin, M. G. et al. Physiological and agronomic responses and nutrient uptake of soybean genotypes cultivated under various sowing dates. J. Soil Sci. Plant Nutr. 23 (4), 5145–5158. https://doi.org/10.1007/s42729-023-01389-y (2023).
Saudy, H. S., Hamed, M. F., El–Metwally, I. M., Ramadan, K. A. & Aisa, K. H. Assessing the effect of biochar or compost application as a spot placement on broomrape control in two cultivars of faba bean. J. Soil Sci. Plant Nutr. 21, 1856–1866. https://doi.org/10.1007/s42729-021-00485-1 (2021).
Nyathi, M. K., Van Halsema, G. E., Beletse, Y. G., Annandale, J. G. & Struik, P. C. Nutritional water productivity of selected leafy vegetables. Agric. Water Manag. 209, 111–122. https://doi.org/10.1016/j.agwat.2018.07.025 (2018).
Nyathi, M. K., Du Plooy, C. P., Van Halsema, G. E., Stomph, T. J., Annandale, J. G., & Struik, P. C. The dual-purpose use of orange-fleshed sweet potato (Ipomoea batatas var. Bophelo) for improved nutritional food security. Agric.Water Manag. 217 23–37. (2019).
Senekal, M., Nel, J., Malczyk, S., Drummond, L. & Steyn, N. P. Provincial Dietary Intake Study (PDIS): micronutrient intakes of children in a representative/random sample of 1-to < 10-year-old children in two economically active and urbanized provinces in South Africa. Int. J. Environ. Res. Public Health 17 (16), 5924. https://doi.org/10.3390/ijerph17165924 (2020).
Saudy, H. S., El-Metwally, I. M. & Abd El-Samad, G. A. Physio-biochemical and nutrient constituents of peanut plants under bentazone herbicide for broad-leaved weed control and water regimes in dry land areas. J. Arid Land 12 (4), 630–639. https://doi.org/10.1007/s40333-020-0020-y (2020).
Saudy, H. S. Mohamed El–Metwally, I. Effect of irrigation, nitrogen sources, and metribuzin on performance of maize and its weeds. Commun. Soil Sci. Plant Anal. 54 (1), 22–35. https://doi.org/10.1080/00103624.2022.2109659 (2023).
Salem, E. M., Kenawey, K. M., Saudy, H. S. & Mubarak, M. Soil mulching and deficit irrigation effect on sustainability of nutrients availability and uptake, and productivity of maize grown in calcareous soils. Commun. Soil Sci. Plant Anal. 52 (15), 1745–1761. https://doi.org/10.1080/00103624.2021.1892733 (2021).
Saudy, H. S. & El–Metwally, I. M. Nutrient utilization indices of NPK and drought management in groundnut under sandy soil conditions. Commun. Soil Sci. Plant Anal. 50 (15), 1821–1828. https://doi.org/10.1080/00103624.2019.1635147 (2019).
El-Metwally, I. M. & Saudy, H. S. Interactional impacts of drought and weed stresses on nutritional status of seeds and water use efficiency of peanut plants grown in arid conditions. Gesunde Pflanz. 73 (4), 407–416. https://doi.org/10.1007/s10343-021-00557-3 (2021).
Ncisana, L. et al. Water regimes in selected fodder radish (Raphanus sativus) genotypes: effects on nutritional value and in vitro ruminal dry matter degradability. Heliyon 10 (8). https://doi.org/10.1016/j.heliyon.2024.e29203 (2024).
Mabhaudhi, T., Modi, A. T. & Beletse, Y. G. Response of taro (Colocasia esculenta L. Schott) landraces to varying water regimes under a rain shelter, Agric. Water Manag. 121 102–112. https://doi.org/10.1016/j.agwat.2013.01.009 (2013).
Soil Classification Working Group. Soil classification: a taxonomic system for South Africa. Pretoria: Department of Agricultural Development. (1991).
Archontoulis, S. V. et al. Temporal dynamics of light and nitrogen vertical distributions in canopies of sunflower, kenaf and cynara. Field Crops Res. 122 (3), 186–198. https://doi.org/10.1016/j.fcr.2011.03.008 (2011).
AOAC (Association of official analytical chemists). Official methods of analysis of the association of official. Analytical Chemists, 15th edition. AOAC, Washington DC, 992–995. https://law.resource.org/pub/us/cfr/ibr/002/aoac.methods.1.1990 (1990).
Moyo, M. et al. Determination of mineral constituents, phytochemicals and antioxidant qualities of Cleome gynandra, compared to Brassica oleracea and Beta vulgaris. Front. Chem. 5, 128. https://doi.org/10.3389/fchem.2017.00128 (2018).
Trumbo, P. R., Yates, A. A., Schlicker-Renfro, S. & Suitor, C. Dietary Reference Intakes: revised nutritional equivalents for folate, vitamin E and provitamin A carotenoids. J. Food Compos. Anal. 16, 379–382 https://doi.org/10.1016/S0889-1575(03)00025-5 (2003).
Uusiku, N. P., Oelofse, A., Duodu, K. G., Bester, M. J. & Faber, M. Nutritional value of leafy vegetables of Sub-saharan Africa and their potential contribution to human health: a review. J. Food Compos. Anal. 23, 499–509. https://doi.org/10.1016/j.jfca.2010.05.002 (2010).
NRC. Nutrient Requirements for beef Cattle 7th edn (National Academy, 1996).
McDowell, L. R. Minerals for grazing ruminants in tropical regions (3rd edn). Extension Bulletin. (Gainesville: University of Florida, 1997).
Shaaban, A., Mahfouz, H., Megawer, E. A. & Saudy, H. S. Physiological changes and nutritional value of forage clitoria grown in arid agro-ecosystem as influenced by plant density and water deficit. J. Soil Sci. Plant Nutr. 23 (3), 3735–3750. https://doi.org/10.1007/s42729-023-01294-4 (2023).
Ali, M. A. A. et al. Melatonin as a key factor for regulating and relieving abiotic stresses in harmony with phytohormones in horticultural plants — a review. J. Soil Sci. Plant. Nutr. 24, 54–73. https://doi.org/10.1007/s42729-023-01586-9 (2024).
Doklega, S. M. et al. Rhizospheric addition of hydrogel polymer and zeolite plus glutathione mitigate the hazard effects of water deficiency on common bean plants through enhancing the defensive antioxidants. J. Crop Health 76, 235–249. https://doi.org/10.1007/s10343-023-00947-9 (2024).
Abdo, R. A., Hazem, M. M., El-Assar, A. E. M., Saudy, H. S. & El-Sayed, S. M. Efficacy of nano-silicon extracted from rice husk to modulate the physio-biochemical constituents of wheat for ameliorating drought tolerance without causing cytotoxicity. Beni Suef Univ. J. Basic Appl. Sci. 13 (1), 75. https://doi.org/10.1186/s43088-024-00529-2 (2024).
Saudy, H. S., El-Bially, M. E., Hashem, F. A., Shahin, M. G. & El-Gabry, Y. A. The changes in yield response factor, water use efficiency, and physiology of sunflower owing to ascorbic and citric acids application under mild deficit irrigation. Gesunde Pflanz. 75 (4), 899–909. https://doi.org/10.1007/s10343-022-00736-w (2023).
Shelembe, S. C. Water use and nutritional water productivity of taro (colocasia esculenta L. Schott) Landraces (MSc dissertation, 2020).
Akram, N. A., Noreen, S., Noreen, T. & Ashraf, M. Exogenous application of trehalose alters growth, physiology and nutrient composition in radish (Raphanus sativus L.) plants under water-deficit conditions. Brazilian J. Bot. 38, 431–439. https://doi.org/10.1007/s40415-015-0149-7 (2015).
Henschel, J. M. et al. Drought stress mitigation by foliar application of L-carnitine and its effect on radish morphophysiology. Physiol. Mol. Biol. Plants 29 (4), 579–590. https://doi.org/10.1007/s12298-023-01308-6 (2023).
Stagnari, F., Galieni, A., D’Egidio, S., Pagnani, G. & Pisante, M. Responses of radish (Raphanus sativus) to drought stress. Ann. Appl. Biol. 172 (2), 170–186. https://doi.org/10.1111/aab.12409 (2018).
Salazar-Garcia, G., Balaguera-Lopez, H. E. & Hernandez, J. P. Effect of plant growth-promoting bacteria Azospirillum brasilense on the physiology of radish (Raphanus sativus L.) under waterlogging stress. Agronomy 12 (3), 726. https://doi.org/10.3390/agronomy12030726 (2022).
Silva, M. R. F. et al. Trading a little water for substantial carbon gains during the first years of a Eucalyptus globulus plantation. Agric. For. Meteorol. 318, 108910. https://doi.org/10.1016/j.agrformet.2022.108910 (2022).
Flexas, J. et al. Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. Plant Cell Environ. 39 (5), 965–982. https://doi.org/10.1111/pce.12622 (2016).
Jacobs, J., Ward, G. & McKenzie, F. Effect of irrigation strategies on dry matter yields and water use efficiency of a range of forage species. (2004).
Mandal, K. G., Hati, K. M., Misra, A. K. & Bandyopadhyay, K. K. A. Assessment of irrigation and nutrient effects on growth, yield and water use efficiency of Indian mustard (Brassica juncea) in central India. Agric. Water Manag. 85 (3), 279–286. https://doi.org/10.1016/j.agwat.2006.05.004 (2006).
Zhang, X., Qin, W., Chen, S., Shao, L. & Sun, H. Responses of yield and WUE of winter wheat to water stress during the past three decades—A case study in the North China Plain. Agric. Water Manag. 179, 47–54. https://doi.org/10.1016/j.agwat.2016.05.004 (2017).
Kunrath, T. R., Lemaire, G., Sadras, V. O. & Gastal, F. Water use efficiency in perennial forage species: interactions between nitrogen nutrition and water deficit. Field Crops Res. 222, 1–1. (2018).
Sunith, S., Ravindran, C. S., George, J. & Sreekumar, J. Effect of pruning on cassava stem, foliage and tuber yield. J. Root Crops 41 (2), 36–41 (2015).
Ullah, H., Santiago-Arenas, R., Ferdous, Z., Attia, A. & Datta, A. Improving water use efficiency, nitrogen use efficiency, and radiation use efficiency in field crops under drought stress: a review. Adv. Agron. 156, 109–157 (2019).
Dahniya, M. T., Oputa, C. O. & Hahn, S. K. Effects of harvesting frequency on leaf and root yields of cassava. Exp. Agric. 17 (1), 91–95 (1981).
Munyahali, W. et al. Responses of cassava growth and yield to leaf harvesting frequency and NPK fertilizer in South Kivu, Democratic Republic of Congo. Field Crops Res. 214, 194–201. https://doi.org/10.1016/j.fcr.2017.09.018 (2017).
Hauser, S., Bakelana, Z., Bungu, D. M., Mwangu, M. K. & Ndonda, A. Storage root yield response to leaf harvest of improved and local cassava varieties in DR Congo. Arch. Agron. Soil Sci. 67 (12), 1634–1648. https://doi.org/10.1080/03650340.2020.1800647 (2021).
Ochieng’Orek, C. Morphological, physiological and molecular characterization of drought tolerance in cassava (Manihot esculenta Crantz) (Doctoral dissertation, ETH Zurich). (2014).
Abdel, C. G. Yield analysis of four radish (Raphanus sativus L. var. sativus) genotypes storage roots grown in controlled cabinets under varying temperatures and irrigation levels. (2015).
Cunha, F. F. D., Souza, I. P. D., Campos, W. D. O., Andrade, V. C. D. & Magalhães, T. A. Agronomic performance of radish genotypes under different irrigation depths. Engenharia Agrícola 39, 182–190. https://doi.org/10.1590/1809-4430 (2019).
Lima, E. M. C., Maller, A., Hara, A. T., Rezende, F. C. & Carvalho, J. A. Efeito De diferentes níveis de água no solo na produção do rabanete cultivado em dois tipos de ambientes protegidos. Rev. Engenharia Agric. Reveng. 23 (4), 346–354 (2015).
Galmes, J., Cifre, J., Medrano, H. & Flexas, J. Modulation of relative growth rate and its components by water stress in Mediterranean species with different growth forms. Oecologia 145, 21–31. https://doi.org/10.1007/s00442-005-0106-4 (2005).
Shabani, A., Sepaskhah, A. R. & Kamgar-Haghighi, A. A. Growth and physiologic response of rapeseed (Brassica napus L.) to deficit irrigation, water salinity and planting method. Int. J. Plant. Prod. 7 (3), 569–596 (2013).
Mahesh, K., Balaraju, P., Ramakrishna, B. & Rao, S. S. R. Effect of brassinosteroids on germination and seedling growth of radish (Raphanus sativus L.) under PEG-6000 induced water stress. Am. J. Plant Sci. https://doi.org/10.4236/ajps.2013.412285 (2013).
Marcelis, L. F. M. & Van Hooijdonk, J. Effect of salinity on growth, water use and nutrient use in radish (Raphanus sativus L.). Plant Soil 215, 57–64 (1999).
Partap, M. et al. A tiny plant with superfood potential. J. Funct. Foods 107, 105697. https://doi.org/10.1016/j.jff.2023.105697 (2023).
Mabhaudhi, T., Chibarabada, T. & Modi, A. Water-food-nutrition-health nexus: linking water to improving food, nutrition and health in Sub-saharan Africa. Int. J. Environ. Res. Public Health 13 (1), 107 (2016).
Maseko, I. et al. Du Plooy. Moisture stress on physiology and yield of some indigenous leafy vegetables under field conditions. South. Afr. J. Bot. 126 (201), 85–91. https://doi.org/10.1016/j.sajb.2019.07.018 (2019).
Schlering, C. et al. Chemical composition of field grown radish (Raphanus sativus L. var. Sativus) as influenced by season and moderately reduced water supply. J. Appl. Bot. Food Qual. 92 https://doi.org/10.5073/JABFQ.2019.092.046 (2019).
Maluleke, M. K., Moja, S. J., Nyathi, M. & Modise, D. M. Nutrient concentration of African horned cucumber (Cucumis metuliferus L.) fruit under different soil types, environments, and varying irrigation water levels. Horticulturae 7 (4), 76. https://doi.org/10.3390/horticulturae7040076 (2021).
Park, T. et al. Combined effects of drought and soil fertility on the synthesis of vitamins in green leafy vegetables. Agriculture 13 (5), 984. https://doi.org/10.3390/agriculture13050984 (2023).
Gamba, M. et al. Bioactive compounds and nutritional composition of Swiss chard (Beta vulgaris L. var. cicla and flavescens): a systematic review. Crit. Rev. Food Sci. Nutr. 61 (20), 3465–3480. https://doi.org/10.1080/10408398.2020.1799326 (2021).
Ahmed, A. F., Yu, H., Yang, X. & Jiang, W. Deficit irrigation affects growth, yield, vitamin C content, and irrigation water use efficiency of hot pepper grown in soilless culture. HortScience 49 (6), 722–728 (2014).
Stagnari, F., Galieni, A. & Pisante, M. Drought stress effects on crop quality. Water Stress Crop Plants Sustain. Approach 2, 375–392 (2016).
Britz, S. J. & Kremer, D. F. Warm temperatures or drought during seed maturation increase free α-tocopherol in seeds of soybean (Glycine max [L.] Merr.). J. Agric. Food Chem. 50 (21), 6058–6063 (2002).
Oh, M. M., Carey, E. E. & Rajashekar, C. B. Regulated water deficits improve phytochemical concentration in lettuce. J. Am. Soc. Hortic. Sci. 135 (3), 223–229 (2010).
Bumgarner, N. R., Scheerens, J. C. & Kleinhenz, M. D. Nutritional yield: a proposed index for fresh food improvement illustrated with leafy vegetable data. Plant Foods Hum. Nutr. 67, 215–222. https://doi.org/10.1007/s11130-012-0306-0 (2012).
Acknowledgements
This study received financial support from the Range and Forage Unit of the Agricultural Research Council and the National Research Foundation of South Africa. Additionally, we express our gratitude to the University of Limpopo Research Office for funding the nutritional analysis conducted in this research. Special acknowledgement is extended to Aron Maema and Sivuyisiwe Mvundlela for their valuable assistance in data collection. We would also like to recognise Mr Dave Goodenough and Dr Alan Stewart for their significant contributions to the breeding of ENDURANCE and LINE 2 genotypes.
Author information
Authors and Affiliations
Contributions
Lusanda Ncisana, Melvin Kudu Nyathi, Ntuthuko Raphael Mkhized, Tafadzwa Mabhaudhi, Tlou Julius Tjelelef, Lwando Mbambalala and Albert Thembinkosi Modi planned the study and designed the experiments. Lusanda Ncisanaa and Melvin Kudu Nyathiperformed the experiments. Lusanda Ncisana conducted data analysis and wrote the paper. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ncisana, L., Nyathi, M.K., Mkhize, N.R. et al. Water use efficiency (WUE) and nutrient concentration of selected fodder radish (Raphanus sativus L.) genotypes for sustainable diets. Sci Rep 14, 31315 (2024). https://doi.org/10.1038/s41598-024-82727-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-82727-7
