
Abstract
Faba bean is a nutritionally and medicinally rich popular legume crop. However, vicine-convicine remain as potential threats for “favism” in human beings. In this study, 189 diverse faba bean accessions have been evaluated for yield component traits and vicine content in seeds followed by a correlation study. Combined genetic variability analysis shows that traits like days to pod initiation (DPI), pod length (PL), test weight (TW) and grain yield have minimally been influenced by the environment. PCA revealed that TW, PL and PW were the primary indicators for deciding yield performance. LC–MS/MS confirms that vicine concentration varied in between 3.489 and 10.025 g/kg and a significant positive correlation (0.40***) was observed between vicine conc. and grain yield of faba bean. Thus, present study demonstrated that the faba bean genotypes containing lower vicine were mostly poor yielding, which might be regulated by vicine in faba bean. Therefore, complete elimination of vicine or development of near-zero vicine faba bean could drastically reduce the yield potential of the crop, hence one has to be very cautious and follow efficient selection strategies while optimizing lower concentration of vicine for development of low vicine varieties. This study shows that faba bean genotypes containing 4.0–5.5 g/kg vicine were fairly productive and also have considerably lower vicine.
Similar content being viewed by others


Introduction
Legume crops have been recognized as the second most valuable plant source for health and nutrition and mainly used to meet the protein requirements. Over a dozen legume crops are grown in different parts of the world, and faba bean is the fourth most widely grown cool-season grain legume globally after pea, chickpea and lentil, with an annual production of 5.4 million tonnes from an area of 2.57 million ha (FAOSTAT 2021: www.faostat.fao.org). It is considered as an underutilized legume with great potential to improve the nutritional status of both humans and animals and people who can’t afford regular animal-based protein for their dietary requirements. Faba bean is a rich source of proteins (22–38.2%), essential amino acids and dietary fibres (12–25%), and also possesses excellent medicinal properties1,2. Additionally, it has good amount of carbohydrates (57.3%), vital vitamins (B vitamins), minerals, and energy (320 kcal/100 g)3,4. It is also a rich source of folic acid (vitamin B9, 148 mg/100 g), which is essential for the synthesis and repair of nucleic acids, metabolism of amino acids, and prevention of anemia by promoting the production of red blood cells5.
Phenolic compounds particularly flavonoids occur in faba bean and are known to have antioxidant, anti-inflammatory, antiviral, anticancer and anti-atherosclerotic effects6. Faba bean contains L-DOPA (L-3,4-dihydroxyphenylalanine), which is the precursor of dopamine (The Happiness Hormone) and has the ability to cross blood brain barrier and hence used for treatment of Parkinson’s disease (PD), the second most common neurodegenerative disease in elderly people7,8. It is also a suitable food for diabetic patients, heart and cardiovascular diseases because of its biochemical composition3.
Faba bean is considered as a cool-season legume, which grows well at temperature around 22 °C. Major faba bean breeding challenges are mainly due to its mixed breeding system, unknown wild progenitor, and large genome size of nearly 13 Gb, which is the largest among diploid field crops9,10. Based on size of the seeds, the faba bean has four subspecies: V. faba major, (primarily grown in China and South Mediterranean regions); V. faba equina, (grown primarily in North Africa and Middle Eastern regions); V. faba minor, (grown primarily in Ethiopia and North Europe); and V. faba unijuga, (primitive form primarily found in Afghanistan and India)11. The large seeded cultivars are known as “Broad Bean” in Australia, United Kingdom and New Zealand, whereas, small seeded cultivars are known as “Field Bean”, which are mostly used as fodder for cattle. The global resources for faba bean genomics exhibit significant variation, and there is an enormous collection of faba bean accessions within 37 international repositories, where more than 43,500 faba bean accessions are preserved1,9,12, (FAOSTAT 2016: http://faostat.fao.org/site/567/default.aspx#ancor/; ICARDA: https://www.icarda.org/; CAAS: https://www.caas.cn/en/; GENESYS: https://www.genesys-pgr.org/c/fababean; NBPGR: http://pgrportal.nbpgr.ernet.in/; accessed on 15.07.2024).
However, the potential usage of faba bean as a rich source of protein is hampered due to certain anti-nutritional factors (ANFs) such as vicine-convicine (VC), tannin, phytic acid etc. VC are the thermostable, glucosidic aminopurine derivatives that build up in the cotyledons of faba bean mainly during the seed development and maturity stage13. When people with genetic deficiency of glucose-6-phosphate dehydrogenase (G6PD) activity (~ 4% of the global population) eat faba bean containing higher VC and their hydrolysed products may suffer a condition called favism or hemolytic anemia due to hemolysis of red blood cells1,9,14.
After screening a large number of faba bean accessions, Duc et al.15 reported that, VC locus, which contains two distinct alleles viz., VC + (wild type) and vc − (mutant allele), controls the VC characteristic. The mutant single recessive allele, vc − was responsible for low vicine characteristic in a Greek mutant and the allele reduces VC concentration by almost 95% (10–20-fold reduction). In wild-type faba bean, seeds contain 6–14 g/kg of VC, however, low VC or VC-free genotypes (mutant) contain only 5–10% of this amount16. Therefore, homozygosity at the mutant allele (vc −/vc −) has the potential to prevent the genetic disorder of favism in humans and eliminate various dietary disadvantages in animals, whereas, heterozygous mutant allele leads to intermediate VC concentration17,18,19.
The VC locus has been fine mapped to a region of 0.13 cM on the long arm of chromosome 114,17,20,21. According to Björnsdotter et al.14, the gene-to-metabolite correlation study shows that, VC are generated in maternal tissues and eventually transferred to the embryo, and their contents in seed are maternally determined. The gene expression profiling revealed that the gene VC1 encodes an isoform of 3,4-dihydroxy-2-butanone-4-phosphate synthase/GTP cyclohydrolase II, that is a bifunctional riboflavin biosynthesis enzyme and is responsible for catalyzing the crucial step in the biosynthesis of VC. VC1 expression in the seed coat was found to be 7.4 times higher than the embryo. The activity of this enzyme is relatively low in immature seeds, highest in ripe seeds, and decreases in older/dried seeds14. Their results strongly suggest that VC are the by-products of riboflavin biosynthesis from GTP. They proposed the reason of the low VC phenotype was a frame-shift insertion in the vc1 gene located at vc-allele and consequent changes in the amino acids composition of RIBA1 protein, whose GTP cyclohydrolase II function was lost due to the 2 bp (AT) insertional mutation14,21.
There are multiple reports of low or near zero VC genotypes and the mutant allele responsible for low vicine had been identified, but the consequent effect of low vicine on the reproductive yield and crop performance of faba bean has not been explored yet. Although faba bean production has been increased over the past 20 years, possibly as a result of selection efforts and the development of some high-yielding cultivars through various breeding programs, there is still a pressing need to boost the production and productivity of safe and high-quality faba bean varieties with low ANFs in order to maintain global food security and fulfil the requirement of Safe and Nutritious Food (SNF) for an ever-increasing population of 10 billion by 2057. Moreover, as per UN’s anticipation, India will be the most populous country with an estimated population of 1.67 billion by 2050 (Report from UN Population Division 2022: https://population.un.org/wpp/).
Therefore, evaluation of faba bean genotypes for growth and yield component traits and study of correlation between variation in vicine concentration and reproductive yield performance of faba bean are highly desirable to investigate whether any significant correlation is present or not and as to why some faba bean genotypes grow well in a particular agroclimatic region but poor in their reproductive yield performance. After a detailed phenotypic evaluation using more than 20 morphological/yield component traits and an extensive vicine phenotyping of 189 faba bean accessions, here we report a significant positive correlation between vicine concentration and grain yield performance of faba bean. Moreover, we hypothesized how low vicine concentration could impact the reproductive growth and yield performance in faba bean.
Results
Identification of genotypes with desirable morphological and yield component traits
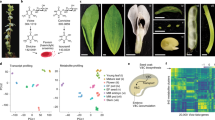
A total of 189 different faba bean accessions, including 40 indigenous and 128 exotic collections from NBPGR, 19 landraces from Northeast (NE) India and 2 national varieties (PS and PU) were grown and evaluated during two consecutive cropping seasons (Supplementary Fig. S1) and data were recorded on both qualitative and quantitative variables. Based on seed shape, there were mainly two types of seeds: round (121 accessions) and flattened seeds (68 accessions) (Fig. 1A and B). Majority of the landraces from NE India and the indigenous collections from NBPGR had flattened seeds, whereas, most of the exotic collections comprised of round shaped seeds. There were no accessions with angular seeds. There were 174 accessions (92%), which had hilum pigmentation and 15 accessions (8%) with colorless hilum. Majority of the Indian landraces do not show hilum pigmentation. Faba bean usually shows indeterminate and semi-determinate type of growth and very rarely shows determinate growth. In the entire collection of germplasm, 164 accessions (87%) were indeterminate and 25 accessions (13%) were semi-determinate. However, there were no accessions with determinate growth habit. The faba bean accessions show various degree of pod curvature: strong (1 acc.), medium (34 acc.), weak (52 acc.) and very weak or absence of curvature (102 acc.). Based on anthocyanin coloration on wings and standard, there were mainly two types of flowers: white flower and colored flower (Fig. 1C and D). In the entire faba bean collection, majority of the accessions had colored flowers and only few accessions had white flowers.

Field evaluation of morphological and yield component traits. (A) accession Fb-13 (indigenous collection from NBPGR, New Delhi, acc. no. ET-218751) shows flattened seeds. (B) accession Fb-33 (indigenous collection from NBPGR, New Delhi, acc. no. ET-218736) shows round shaped seeds. (C) accession Fb-39 shows white flowers with early flowering. (D) accession Fb-116 had coloured flowers showing black pigmentation in standard. (E) variation in LL, where Fb-151 had short (< 6.5 cm), Fb-168 (> 6.5 cm) had medium and Fb-18 had long (> 8 cm) leaflet. (F) variation in PL (varies between 6–13 cm) and PDC curvature, Fb-6 and Fb-49 (absence of curvature), Fb-92 (weak curvature), Fb-64 (medium curvature) and Fb-3 and Pusa Udit (strong curvature).
Data on various morphological and yield component traits were recorded throughout the seasons (Fig. 1E and F) and the descriptive statistics and combined genetic variability components obtained from pooled analysis of combined data are presented in Table 1 and the variability observed on some of the important yield component traits are briefly described here. DPI varied between 52 and 133 days with a mean value of 97.92. It was found that the majority of NE Indian landraces had emerged pods at a very early stage as compared to other indigenous and exotic accessions. However, BLUP estimation shows that DPI varies in the range of 87.06 to 104.5 (Fig. 2A). The NPPN varied from 1 to 3 and there was significant variation in the NPPP among the faba bean accessions. Though several accessions had emerged quite a good number of flowers, but due to excessive flower drop, low pod setting has been observed. The highest NPPP recorded was > 130 with an average pod number of 24.33, whereas, the BLUP values for NPPP varied in between 22 and 30 (Fig. 2B). The longest PL was recorded 12.57 cm and smallest was 4.33 cm, whereas, mean PL was 6.41 and BLUP values vary in between 5 and 9 (Fig. 2C). PW varied in the range of 0.97 to 3.22 cm with a mean value of 1.52, and BLUP values for PW varied in between 1.16 and 1.82. The highest NOPP recorded was 4 and lowest was 2. TW is an important yield component trait in faba bean and it varied considerably among the accessions. The maximum TW of 1192 g and minimum 257 g was recorded, whereas, the mean TW was 486.99 g and BLUP values varied in between 299 and 1078 (Fig. 2D). Throughout the crop seasons, the maximum, minimum and mean yield recorded were 23.28, 1.2 and 3.92 t/ha respectively, whereas, pooled analysis shows BLUP values for yield varies in between 1.51 and 7.16. The distribution of grain yield performance of 189 accessions was presented in the Fig. 2E.

Analysis of agronomical and yield component traits. (A) considerable variation observed on DPI (from 87.07 to 105.07 days) with low PCV and GCV. (B) variation in NPPP- majority of genotypes showed 23–26 NPPP. (C) PL varies in between 5.817 and 8.967 cm, whereas, majority show 6.5–7.5 cm. (D) variation in test weight of faba bean accessions, minimum (299.44 g), maximum (1078.06 g) and average (478.21 g). (E) distribution of grain yield performance (t/ha) of 189 accessions, both the lower and higher extreme groups are highlighted in red and green respectively and two national varieties namely PS and PU are indicated using arrows. (F) PCA of 12 morphological and yield component variables using 189 accessions, PC1 and PC2 together contribute nearly 40% of the total variation, whereas, PC1, PC2, PC3 and PC4 together describe > 60% of the total variation.
Principal component analysis (PCA)
PCA was used to reduce the dimension of the variables and to prioritize the morphological and yield component traits based on their relative significance. From the analysis results, four principal components (namely PC1 PC2, PC3 and PC4) with eigenvalues > 1 were extracted using varimax rotation matrix, and these four PCs collectively explained more than 60% of the total variation (Supplementary Table S1). Highly influential traits were identified based on factor loadings in different PCs, where TW, PL, PW, DPI, LL and LW demonstrated substantial loadings in PC1 and also exhibited significant correlations. Factor loadings in PC1 also demonstrated that TW, PL and PW are the major or primary indicators of yield performance in faba bean and PC1 was more relevant to yield contributing traits (Fig. 2F).
Vicine concentration varied between 3.489 and 10.025 g/kg
A standard curve with R2 value of 0.9998 was generated and the concentration of vicine from faba bean accessions were determined using the equation y = 4E − 05x + 0.0152 (Supplementary Fig. S2, here “x” is the concentration and y means absorbance). Homogenous fine powdered samples from seeds were used for vicine estimation. The accession Fb-33 had the highest vicine concentration of 9.108 g/kg, whereas the accession Fb-40 had the lowest vicine concentration of 2.584 g/kg (Fig. 3A). The average vicine concentration of 6.024 g/kg was observed and a substantial number of accessions exhibited vicine concentrations in between 5.037 and 6.909 g/kg across the entire faba bean accessions. Two national varieties, namely Pusa Sumeet (PS) and Pusa Udit (PU) exhibited vicine content 7.402 and 5.129 g/kg respectively. The Fig. 3A and Supplementary Table S2 provide information on the spectrophotometry-based vicine concentration of each faba bean accessions.

Estimation of vicine from seed samples. (A) Vicine concentration (g/kg) of 189 faba bean accessions based on spectrophotometric estimation, accessions with the highest (Fb-33) and lowest (Fb-40) vicine concentrations are highlighted in red and green respectively. (B) Chromatogram of vicine obtained from the sample Fb-33 in liquid chromatography (retention time- 3.117 min, wavelength-273 nm). (C) Chromatogram of vicine obtained from the sample Fb-40 in liquid chromatography (retention time- 3.117 min, wavelength-273 nm). (D) Mass-spectrum of vicine, which shows a base peak of 305.5 as obtained from LC–MS/MS estimation of vicine from faba bean.
Based on the above observation, 30 selected accessions (including both the extremes) were further analysed using LC–MS/MS method (Table 2). A linear calibration curve with a R2 value of 0.9998 was obtained (Fig. S3). The mass spectrum of vicine exhibited a solitary ion and base peak at m/z 305 and few product ions. The solitary ion was fragmented by collision energy, producing the resulting MS/MS spectra. As vicine could be ionized in both positive and negative ionization modes, the mass spectra (m/z 290–320) was obtained by positive ionization. The best separation was achieved with gradient elution at a temperature of 40 °C and a flow rate of 0.4000 ml/min. In LC–MS/MS, the accessions Fb-33 and Fb-40 (extreme genotypes in terms of vicine content from spectrophotometric results) were identified with vicine concentrations 8.045 and 3.925 g/kg respectively. However, among the 30 selected accessions, in LC–MS/MS, Fb-29 and Fb-41 were identified with the lowest and highest vicine concentration of 3.489 and 10.025 g/kg respectively. The chromatograms of vicine obtained from samples Fb-33 and Fb-40 are given in the Fig. 3B and C and the mass-spectrum of vicine from faba bean was given in the Fig. 3D.
Concentration of vicine is positively correlated with the reproductive yield performance of faba bean
All the accessions were evaluated in field for two consecutive cropping seasons (Supplementary Fig. S1) and simultaneously grown in pots containing fertile soil as medium in a controlled condition and their growth and reproductive performance were observed. The Fig. 4A shows the growth and reproductive performance of some selected faba bean accessions after 70 days on soil-based pot culture in a controlled condition, where growth of plants with lower concentration of vicine was comparatively slower than the plants with higher vicine. In case of plants with higher vicine, the number of emerged flowers at the age of 70 days were more as compared to the plants with lower vicine (Fig. 4A; Table 3). Plants with higher vicine show comparatively a greater number of pods and more grain yield per plant. Additionally, some of the important seed morphological traits such as hilum colour, seed shape etc. were evaluated critically to ascertain any correlation between seed morphology and vicine content. Figure 4B represents the two types of hilum present in faba bean: accessions Fb-4, Fb-30, Fb-33, Fb-40, Fb-41, Fb-162, and PS had black pigmentation in hilum, whereas, accessions Fb-173, Fb-177, Fb-181 show no hilum pigmentation. Data on seed shape show that the round seeds contain comparatively higher vicine content than the flattened seeds. Figure 4C represents seed size, test weight and vicine concentration of eight important faba bean accessions.

Correlation between yield performance of faba bean and vicine content. (A) growth and reproductive performance of some contrasting (low/high vicine) faba bean accessions at the age of 70 days in pots in controlled condition and their respective vicine concentrations. (B) variation in hilum pigmentation among some of the important faba bean accessions, where Fb-4, Fb-30, Fb-33, Fb-40, Fb-41, Fb-162, and PS had black pigmentation in hilum, on the other hand, Fb-173, Fb-177, Fb-181 had colourless hilum. (C) Seed size (visual observation made on 5 seeds per accession), test weight (g) and vicine concentration (g/kg) of eight important selected faba bean accessions, a strong significant negative correlation (− 0.60***) between TW and vicine concentration can be seen. (D) Pearson’s correlation coefficients between vicine concentration and major yield component parameters and morphological traits recorded from field evaluation of faba bean accessions.
The correlation analysis performed between vicine concentration and morphological and yield component traits recorded from field evaluation of 189 faba bean accessions showed that vicine is positively correlated with NPPP (0.41***), NGPP (0.53***) and overall grain yield (0.40***) in faba bean (Fig. 4D). Moreover, vicine is also positively correlated with hilum pigmentation (0.32***) and seed shape (0.40***). However, vicine is negatively correlated with DPI (− 0.36***), where accessions with higher vicine enter pod initiation stage at an early growth phase. Additionally, vicine shows significant negative correlations with PL (− 0.37***), PW (− 0.24***) and TW (− 0.60***) of dry seeds. Based on the correlation analysis and overall performance of the faba bean accessions in the agroclimatic condition of Meghalaya, 42 superior accessions were selected for further evaluation and advanced breeding studies.
Discussion
All the faba bean genotypes have been evaluated in the field and their morphological and yield component traits were analyzed. The variability observed in growth habit was found similar with the previous reports by López-Bellido et al.22, Azarpour et al.23, Al Barri24 and Karaköy25 where predominantly indeterminate and occasionally semi-determinate patterns of growth were noted. Another important trait of faba beans is hilum color, which has mostly been found to be linked with the anti-nutritional properties20,21,26 of faba bean. Therefore, the observed variation in hilum color shows the diverseness of the faba bean accessions in terms of genetic and anti-nutritional properties. Variability also observed in seed shape and size of the faba bean genotypes, where similar findings had also been reported previously5,27,28.
ANOVA with treatments and blocks effects on 189 accessions shows significant variations for the traits PH, NNPP, DPI, NPPP, LL, LW, PL, PW, TW, yield etc. at p < = 0.01. In faba bean, DPI varies under irrigated conditions and are largely dependent on soil moisture and weather variables, rain-fed circumstances etc.29,30. In this study, DPI varied moderately in between 87.06 and 104.5, where a low GCV (5.74%) and a low PCV (5.86%) were observed, hence suggesting a lower environmental influence on the trait. In faba bean, NPPP is determined by the number of flowers per plant, proportion of flowers, number of stems per plant and number of nodes per plant13,22,31. Studies show that rather than increasing seed size or number of seeds per pod, by increasing the number of pods per plant at maturity, there is potential to increase seed yield in faba bean and most other legumes. Pooled analysis shows that NPPP in between 22 and 29 with a medium GCV (15.58%) and high PCV (24.02%), suggesting moderate environmental influence on the trait. Combined genetic variability studies on PL and PW show that pod length (GCV-12.38% and PCV 14.71%) has minimally been influenced by the environment, whereas, pod width (GCV-9.30% and PCV-37.40%) has been influenced greatly by the environmental factors. These results are found similar with the previous studies32,33,34,35.
Pooled analysis and study of combined genetic variability components show that TW (BLUP values = 299–1078; GCV-37.33% and PCV-37.40%) and overall grain yield (BLUP values = 1.51–7.16; GCV-41.18% and PCV-42.28%) have minimally been influenced by the environment and the observed variations on the traits was mostly due to the genotypic differences. A high hBS (95.72%) and a high GAM/GG (77.32%) on TW indicate a highly heritable nature of test weight and significant effect of selection on the trait. Moreover, a low hBS (25.04%) and high GAM/GG (43.58%) were observed on yield, suggesting a highly significant effect of selection on yield. For simplification of multivariate datasets into few dominant components and to understand the major sources of variation among the faba bean accessions, PCA was performed and it shows that, PC1 was strongly correlated with mainly three yield related variables, namely TW, PL and PW, suggesting that an efficient selection for these variables could substantially improve the yield potential of the crop and these results were found similar with the previous study by Sultan et al.28.
Studies on vicine demonstrated that the concentration of vicine in faba bean varies in between 3 and 10 g/kg1,14,16,17,20,36,37,38,39,40,41, and in mutant low VC or VC-free genotypes, the concentration is at least 10–20 times lesser14,15,16,17,39,42. Several authors have reported high-throughput genomics approaches for selection of low VC genotypes14,15,16,17,20. However, there is lack of information regarding the growth and yield performance of the previously reported low VC genotypes and no information available about the effect of low VC on the yield performance of this crop. Moreover, growth and reproductive yield performance of faba beans containing low or near-zero VC have not been evaluated adequately by the previous studies and there is lack of evidence that the faba bean yield performance had not been compromised in the low VC genotypes14,15,42.
In this study, vicine estimation using spectrophotometry and LC–MS/MS shows a substantial variation in vicine concentration in the entire faba bean collections, where vicine concentration varied in between 3.489 and 10.025 g/kg. In order to investigate the effects of low/high vicine on the growth and yield performance of faba bean, a correlation analysis was performed using field phenotyping data, where vicine shows significant positive correlations with NPPP, NGPP and overall grain yield and significant negative correlations with DPI, PL, PW and TW. There were nearly 35 accessions with good height and vegetative growth, but beared only few pods and majority of them contained comparatively lower vicine (2.5–5.0 g/kg). The study of growth and reproductive abilities of faba bean genotypes at the age of 70 days in controlled environment (pot culture) shows significantly higher growth in higher vicine genotypes, which emerge comparatively more flowers than the plants with lower vicine.
These findings suggest a possible reduction in yield as a consequence of selection for low vicine trait and the lower yield could be due to slower growth, delay in flowering and pod setting, lower number of pods and grains per plant and loss of seed viability, which might be someway regulated by VC in faba bean. Because, studies show that, most of the antinutritional factors (ANFs) and plant secondary metabolites have role in plant defense, reproduction and stress related functions and provide means to survive in adverse growing conditions43,41,42,46. They are highly biologically active and rapidly synthesized at certain stages of critical growth and stored in organs, which are highly important for survival and reproduction47,45,46,47,51. Vicine (purine glycosides) is an ANF and considered as plant secondary metabolite. Though consumption of faba bean containing higher VC induce hemolytic anemia in G6PD-deficient individuals52, but in plant systems, they might have crucial regulatory roles in various biochemical pathways and stress endurance, reproductive systems and chemical defense mechanisms against biotic stress factors. Therefore, after considering all these facts and our present study, here we proposed an efficient selection strategy, which involves extensive phenotyping of a large number of genotypes especially for yield and yield component traits and their genotyping at VC locus for potential alleles/haplotypes and simultaneous vicine analysis using high-throughput techniques (Fig. 5A). A hypothetical model describing how VC could be involved in maintaining improved reproductive and grain yield performance is given in the Fig. 5B.

A proposed model of efficient selection strategy for high-yielding and low-vicine faba bean and hypothetical roles of vicine in improving reproductive and grain yield performance. (A) precise and efficient selection for low vicine-convicine (LVC) involving extensive phenotyping of large number of genotypes for yield component traits, their genotyping at VC locus for potential haplotypes/alleles and high-throughput vicine estimation for identification of high-yielding faba bean variants/mutants containing low vicine, whereas, inefficient selection without adequate phenotyping could lead to reduced reproductive/grain yield in faba bean. (B) vicine-convicine which are potent ANFs in faba bean might be involved in defense mechanism against biotic stress factors, stress regulatory pathways in adverse growing conditions, maintaining reproductive performance and seed viability, nucleic acid metabolism and regulation of unknown metabolic pathways, which ultimately lead to improved reproductive and grain yield performance.
Hence, complete elimination of these compounds or development of near-zero VC varieties could have the potential for drastic reduction in the yield of faba bean. Therefore, one has to be very cautious and highly precise while optimizing the lower concentration of vicine for the development of low vicine varieties, without compromising the yield potential of the crop. Further, we need to investigate in details how exactly vicine plays roles in maintaining higher reproductive growth.
Conclusions
As vicine-convicine (VC) pose threat to G6PD-deficient individuals, there is a great need to develop low VC varieties, but the effect of low vicine on the reproductive yield and crop performance of faba bean has not been explored yet. Our study demonstrated that the faba bean genotypes containing lower vicine were mostly poor yielding, which might be someway regulated by vicine in faba bean. Therefore, complete elimination of vicine or development of near-zero vicine faba bean could drastically reduce the yield potential of the crop, if the selection for low vicine is not sufficiently fortified with an adequate and efficient phenotyping for both vicine/convicine and yield component traits. Therefore, one has to be very cautious and follow efficient selection strategies while optimizing lower concentration of VC for development of low VC varieties. The present study also demonstrated that, vicine concentration varied substantially (3.489–10.025 g/kg), and faba bean genotypes containing vicine in the range of 4.0–5.5 g/kg were fairly productive and also have comparatively reduced amount of vicine.
Methods
Faba bean accessions and their cultivation
A total of 189 diverse (global collections) faba bean accessions (Supplementary Table S3) were cultivated for two consecutive cropping seasons during 2021 to 2023 at the experimental farm of College of Post-Graduate Studies in Agricultural Sciences (CPGS-AS), Central Agricultural University (CAU, Imphal), Umiam, Meghalaya, India. The farm land was located at an altitude of more than 900 m above the MSL, longitude of E91.910919 and latitude of N25.68197. The region where the farm land was located has a humid, cool and subtropical climate, where nearly 4 months starting from June to September receive the maximum rainfall. The sandy loam soil of the experimental farm had an acidic pH of nearly 6.5. The crop was sown on an 800 square meter plot of land during the second fortnight of October of each season and 10 seeds from each accession were planted following the Augmented Block Design (ABD). All the accessions were also planted in pots containing fertile soil as medium in a controlled environment.
Data collection and evaluation of morphological and yield component traits
All the 189 accessions were grown and assessed in accordance with the faba bean descriptors established by the International Board for Plant Genetic Resources (IBPGR), 1985, as well as the Plants Protection Varieties and Farmer’s Rights Act, 2001 (PPV and FRA 2001: https://plantauthority.gov.in/) guidelines for conducting tests for DUS (Distinctiveness, Uniformity, and Stability) in faba beans. A total of five healthy plants from each accession were selected and observations were noted, with the exception of the border plants, which have not been evaluated. Data on following morphological and yield component traits were recorded: days to 50% flowering (DF), flower color, growth habit, plant height (PH), days to pod initiation (DPI), number of stems per plant (NSPP), number of pods per nod (NPPN), pod length (PL), pod width (PW), number of pods per plant (NPPP), pod degree of curvature (PDC), pod angle, number of ovules per pod (NOPP), number of grains per plant (NGPP), leaflet length (LL) and leaflet width (LW), seed shape (SS), hilum colour (HC), test weight (TW) and yield. In faba bean, the yield is contributed by the following parameters: TW, NPPP, PL and PW, NPPN, NSPP and NOPP53. Out of these, NPPP and TW have the largest range of variation among the faba bean accessions. Freshly harvested dry seeds were used for estimation of vicine from the accessions listed in the Supplementary Table S3.
Statistical analysis
Initially, data from a single season were analysed by using “augmentedRCBD” package in R54 and adjusted mean (AM) values were obtained for all the parameters. Analysis of variance (ANOVA), standard errors (SE), coefficient of variance (CV), genetic variability components55,53,54,58 etc. for each season were obtained from the same analysis in R. Principal component analysis (PCA) was conducted on 12 morphological and yield component variables using factoextra, FactoMineR and ggplot2 packages in R. Adjusted mean data from two different seasons were pooled by using lme4 package in R and BLUP values (Best Linear Unbiased Prediction values represent the actual variability in genotype, because the algorithms used by lme4 package are so designed that it could minimize the environmental effects) were obtained for each of the parameter. The combined genetic variability components and descriptive statistics were also obtained from the results of lme4 in R. The pooled data in the form of BLUP values were used for correlation analysis among various parameters including vicine content. Higher yielding and better performing genotypes in the state of Meghalaya in the Eastern Himalayan region were identified and further evaluated for vicine content from freshly harvested dry seeds of faba bean.
Estimation of vicine content of faba bean accessions
Previous literatures reported that generally vicine and convicine are present in ~ 2:1 ratio in faba bean seeds20,38,59,60, therefore, this study mainly focused on vicine alone, hence a thorough vicine estimation was done for each of the 189 faba bean accessions in 3 different biological replications and the average content was calculated in g/kg. Initially, all the 189 faba bean accessions were screened for vicine content using a modified spectrophotometric method as established by Debnath et al.21, and Sixdenier et al.37. The concentration of vicine in some selected genotypes was further determined and confirmed by Liquid Chromatography-Tandem Mass Spectrometry (LC–MS/MS) as established by Debnath et al.1 and Elkins et al.61.
Spectrophotometric estimation of vicine
Standard vicine was procured from Toronto Research Chemicals, Toronto, ON, Canada. Using 10% LC–MS grade methanol in water, a stock solution of 1000 ppm was prepared, which was used to prepare a working stock of 100 ppm in 80% methanol and then serially diluted to different concentrations of 80 ppm, 70 ppm, 60 ppm, 50 ppm, 40 ppm, 30 ppm, 20 ppm and 10 ppm vicine standard solutions for generation of a standard curve at 274 nm wavelength. Homogenous fine powdered form (< 0.5 mm) of faba bean seed samples were prepared by grinding about 4–5 dry seeds from each accession. Exactly 200 mg of sample powder from every accession was soaked in 1980 µl of distilled water in microcentrifuge tubes (2 ml) for nearly 3.5 h. in water bath at 90 °C. About 20 µl of normal HCL (32%) was mixed with the resulting solution and followed by centrifugation at 13,000 rpm for 15 min. The resulting supernatant was diluted 30 times with ddH2O (1:30 v/v) and recorded OD at 274 nm wavelength using a UV Visible Spectrophotometer (Motras Scientific, Plot 175, Sector 7, IMT Manesar, Gurugram, 122,050, Haryana, India). Data was analyzed and compared with the molecular analysis and sequencing results for allelic variation under study.
LC–MS/MS estimation of vicine from selected accessions
The 100 ppm vicine standard was serially diluted to 50 ppm, 25 ppm, 12.5 ppm, 6.25 ppm, 3.125 ppm and 1.56 ppm vicine in 80% methanol (≥ 99.9% pure, LC–MS grade) and obtained the calibration curve at 273 nm wavelength. Exactly 10.0 mg (0.01 gm) of the previously prepared powdered sample from each sample was used for sample preparation and analysis. Samples were extracted using 0.5 ml of 80% methanol in Milli-Q water, vortexed for 1 min, sonicated for 10 min (Aczet 2.5 Ltr Ultrasonic Cleaner CUB-2.5), and centrifuged at 13,000 rpm for 12 min. The pellets were extracted again by using same protocol and the resulting combined supernatant (1 ml) was filtered using PTFE Chromatography Syringe Filters (25 mm Dia, 0.45 μm Pore) and finally transferred to pre-labelled LC–MS vials (2.0 ml) for analysis (at 273 nm).
A Shimadzu HPLC, I-series LC-2050C and a Triple Quadrupole Shimadzu 8045 mass spectrometer linked via an Electrospray Ionization (ESI) Interface (Model No. LCMS-8045, Nishinokyo Kuwabara-cho, Nakagyo-ku, Kyoto 604–8511, Japan) were used to perform LC–MS/MS analysis. A Shimadzu Shim Pack GIST C18 column (2.1 × 150 mm), which has been maintained at 40 °C was utilized and acetonitrile with 0.1% formic acid (≥ 98.5% pure) were used as solvents in the column. Nitrogen served as the ion source gas and argon as the collision gas. Methanol, acetonitrile and water were the constituents of the mobile phase, which had a pressure of 7252 psi and a flow rate of 0.4000 ml/min. The automated instrument control, method acquisition, and data processing were all handled by LabSolution software. The vicine concentration was determined by analysing the LC–MS/MS data and applying the formula provided by Debnath et al.1, Majumder et al.62, and Lee and Jang63.
Correlation between vicine concentration and yield performance of faba bean
In order to study the effect of low vicine on the reproductive yield performance of faba bean, a correlation analysis was performed between vicine concentration of faba bean accessions and their morphological/yield parameters recorded from the field experiments. The morphological and yield component traits used for the correlation analysis were DF, PH, DPI, NSPP, PL, PW, NPPP, NOPP, NGPP, LL, LW, SS, HC, TW and yield. The correlation analysis was performed using “metan” package in R.
Selection of agronomically superior genotypes containing lower concentration of vicine
Selection was done mainly on the basis of yield performance over the years and lower content of vicine. Based on PH, NSPP, NPPP, NPPN, PL, PW, NOPP and overall yield, plants which performed well in the agroclimatic condition of the state in the Eastern Himalayan region were identified. Stable and better performing plants which also had lower concentration of vicine were finally selected for further evaluation and selection for other important quality and nutritional traits.
Data availability
All the supporting data of this study are available with the manuscript and supplementary information. Additional data will be made available on request.
References
Debnath, S., Majumder, S., Tyagi, W., Rai, M. & Meetei, N. T. An optimized simple, rapid and reliable method for large-scale estimation of vicine from Faba Bean (Vicia faba L.). Food Anal. Methods 17, 436–449. https://doi.org/10.1007/s12161-024-02571-3 (2024).
Akgun, D. & Canci, H. Selection of faba bean (Vicia faba L.) genotypes for high yield, essential amino acids and low anti-nutritional factors. Agriculture 13, 932. https://doi.org/10.3390/agriculture13050932 (2023).
Baginsky, C. et al. Phenolic compound composition in immature seeds of fava bean (Vicia faba L.) varieties cultivated in Chile. J. Food Compos. Anal. 31, 1–6 (2013).
Karaköy, T. et al. in Agriculture for Life, Life for Agriculture” Conference Proceedings. 72–78.
Singh, J. Folate content in legumes. Biomed. J. Sci. Tech. Res. 3, 3475–3480 (2018).
Nijveldt, R. J. et al. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 74, 418–425 (2001).
Topal, N. & Bozoğlu, H. Determination of L-Dopa L-3, 4-dihydroxyphenylalanine content of some Faba bean Vicia faba L. Genotypes. J. Agric. Sci. 22, 145–151 (2016).
Oviedo-Silva, C. A., Elso-Freudenberg, M. & Aranda-Bustos, M. L-DOPA trends in different tissues at early stages of Vicia faba growth: effect of tyrosine treatment. Appl. Sci. 8, 2431. https://doi.org/10.3390/app8122431 (2018).
Khazaei, H. et al. Recent advances in faba bean genetic and genomic tools for crop improvement. Legume Sci. 3, e75 (2021).
Jayakodi, M. et al. The giant diploid faba genome unlocks variation in a global protein crop. Nature 615, 652–659. https://doi.org/10.1038/s41586-023-05791-5 (2023).
Cubero, J. I. On the evolution of Vicia faba L. Theor. Appl. Genet. 45, 47–51 (1974).
Kaur, S., Cogan, N. O., Forster, J. W. & Paull, J. G. Assessment of genetic diversity in faba bean based on single nucleotide polymorphism. Diversity 6, 88–101. https://doi.org/10.3390/d6010088 (2014).
Khamassi, K., Makay, I. J., Ali, M. B., Rezgui, S. & O’Sullivan, D. Agro-morphological variation and nutritional qualitative trait screening among field bean (Vicia faba L. var minor). Legume Genom. Genet. 5, 7–24 (2014).
Björnsdotter, E. et al. VC1 catalyses a key step in the biosynthesis of vicine in faba bean. Nat. plants 7, 923–931 (2021).
Duc, G., Sixdenier, G., Lila, M. & Furstoss, V. Search of Genetic Variability for Vicine and Convicine Content in Vicia faba L. A First Report of a Gene Which Codes for Nearly Zero-Vicine and Zero-Convicine Contents 305–313 (Wageningen Academic Publishers, 1989).
Duc, G., Marget, P., Esnault, R., Le Guen, J. & Bastianelli, D. Genetic variability for feeding value of faba bean seeds (Vicia faba): Comparative chemical composition of isogenics involving zero-tannin and zero-vicine genes. J. Agric. Sci. 133, 185–196 (1999).
Tacke, R., Ecke, W., Höfer, M., Sass, O. & Link, W. Zooming into the genomic vicinity of the major locus for vicine and convicine in faba bean (Vicia faba L.). bioRxiv https://doi.org/10.1101/2021.02.19.431996 (2021).
Gallo, V. et al. No red blood cell damage and no hemolysis in G6PD-deficient subjects after ingestion of low vicine/convicine Vicia faba seeds. Blood J. Am. Soc. Hematol. 131, 1621–1625. https://doi.org/10.1182/blood-2017-09-806364 (2018).
Crépon, K. et al. Nutritional value of faba bean (Vicia faba L.) seeds for feed and food. Field Crops Res. 115, 329–339 (2010).
Khazaei, H. et al. Flanking SNP markers for vicine–convicine concentration in faba bean (Vicia faba L.). Mol. Breed.ng 35, 1–6. https://doi.org/10.1007/s11032-015-0214-8 (2015).
Debnath, S., Meetei, N. T. & Rai, M. Advancement in genomics and molecular marker technologies for breeding of faba bean with low vicine-convicine content: A review. Legume Res. Int. J. 47(5), 687–694. https://doi.org/10.18805/LR-5041 (2024).
López-Bellido, F., López-Bellido, L. & López-Bellido, R. Competition, growth and yield of faba bean (Vicia faba L.). Eur. J. Agron. 23, 359–378 (2005).
Azarpour, E. et al. Path coefficient analysis of seed yield and its components in faba bean (Vicia faba L.) under nitrogen and zinc fertilizer management. Int. J. Agric. Crop Sci. 21(4), 1559–1561 (2012).
Al Barri, T. H. M. Phenotypic characterization of faba bean (Vicia faba L.) landraces grown in Palestine. J. Agric. Sci. 5(2), 110–117. https://doi.org/10.5539/jas.v5n2p110 (2012).
Karaköy, T., Baloch, F. S., Toklu, F. & Özkan, H. Variation for selected morphological and quality-related traits among 178 faba bean landraces collected from Turkey. Plant Genet. Resour. 12, 5–13 (2014).
Gutierrez, N. et al. CAPs markers to assist selection for low vicine and convicine contents in faba bean (Vicia faba L.). Theor. Appl. Genet. 114, 59–66 (2006).
Chaieb, N., Bouslama, M. & Mars, M. Growth and yield parameters variability among faba bean (Vicia faba L.) genotypes. J. Nat. Prod. Plant Resour 1, 81–90 (2011).
Sultan, S. M., Raina, S. K., Ahmad, S. & Bhat, S. S. Characterization and evaluation of Faba Bean (Vicia faba L.) germplasm collected from Kashmir. India. Indian J. Plant Genet. Resour. 36, 346–355. https://doi.org/10.61949/0976-1926.2023.v36i03.02 (2023).
Nanda, H., Yasin, M., Singh, C. & Rao, S. Effect of water stress on dry matter production, harvest index, seed yield and its components in faba bean (Vicia faba L.). FABIS Newslett. 21, 26–30 (1988).
Musallam, I., Al-Karaki, G., Ereifej, K. & Tawaha, A.-R.M. Chemical composition of faba bean genotypes under rainfed and irrigation conditions. Int. J. Agric. Biol. 6, 359–362 (2004).
Turk, M. A. & Tawaha, A.-R.M. Impact of seeding rate, seeding date, rate and method of phosphorus application in faba bean (Vicia faba L. minor) in the absence of moisture stress. Biotechnol. Argon. Soc. Environ. 6(3), 171–178 (2002).
Nadal, S., Suso, M. & Moreno, M. Management of Vicia faba genetic resources: Changes associated to the selfing process in the major, equina and minor groups. Genet. Resour. and Crop Evol. 50, 183–192 (2003).
Terzopoulos, P., Kaltsikes, P. & Bebeli, P. Collection, evaluation and classification of Greek populations of faba bean (Vicia faba L). Genet. Resour. Crop Evol. 50, 373–381 (2003).
Terzopoulos, P., Kaltsikes, P. & Bebeli, P. Characterization of Greek populations of faba bean (Vicia faba L) and their evaluation using a new parameter. Genetic Resour. Crop Evol. 51, 655–662 (2004).
Duzdemir, O. & Ece, A. Determining relationships among plant characteristics related to plant seed yield of broad bean (Vicia faba L.) sown in winter and summer seasons in transitional climate areas of Turkey. Bulg. J. Agric. Sci. 17, 73–82 (2011).
Olsen, H. S. & Andersen, J. H. The estimation of vicine and convicine in faba beans (Vicia faba L.) and isolated faba bean proteins. J. Sci. Food Agric. 29, 323–331 (1978).
Sixdenier, G., Cassecuelle, F., Guillaumin, L. & Duc, G. Rapid spectrophotometric method for reduction of vicine and convicine in faba bean seed. FABIS Newslett. 38(39), 42–44 (1996).
Pulkkinen, M. et al. Determination of vicine and convicine from faba bean with an optimized high-performance liquid chromatographic method. Food Res. Int. 76, 168–177 (2015).
Khazaei, H. et al. Development and validation of a robust, breeder-friendly molecular marker for the vc-locus in faba bean. Mol. Breed. 37, 1–6 (2017).
Debnath, S. et al. Discovery of potential haplotypes associated with varying levels of vicine content due to the InDel1.4 and a coding-SNP in the VC1 gene in faba bean (Vicia faba L.). Plant Gene 41, 100481. https://doi.org/10.1016/j.plgene.2024.100481 (2025).
Khamassi, K. et al. A baseline study of vicine-convicine levels in faba bean (Vicia faba L.) germplasm. Plant Genet. Resour.: Characterisation Util. 11(3), 250–257. https://doi.org/10.1017/S1479262113000105 (2013).
Ugwuanyi, S. et al. VC2 regulates baseline vicine content in faba bean. bioRxiv. https://doi.org/10.1101/2024.08.06.606773 (2024).
Liu, Z. et al. The roles of cruciferae glucosinolates in disease and pest resistance. Plants 10, 1097. https://doi.org/10.3390/plants10061097 (2021).
Al-Khayri, J. M. et al. Plant secondary metabolites: The weapons for biotic stress management. Metabolites 13, 716. https://doi.org/10.3390/metabo13060716 (2023).
Kumar, K., Debnath, P., Singh, S. & Kumar, N. An overview of plant phenolics and their involvement in abiotic stress tolerance. Stresses 3, 570–585. https://doi.org/10.3390/stresses3030040 (2023).
Muthusamy, M. & Lee, S. I. Abiotic stress-induced secondary metabolite production in Brassica: Opportunities and challenges. Front. Plant Sci. 14, 1323085. https://doi.org/10.3389/fpls.2023.1323085 (2024).
Gemede, H. F. & Ratta, N. Antinutritional factors in plant foods: Potential health benefits and adverse effects. Int. J. Nutr. Food Sci. 3, 284–289. https://doi.org/10.11648/j.ijnfs.20140304.18 (2014).
Bora, P. Anti-nutritional factors in foods and their effects. J. Acad. Ind. Res. 3, 285–290 (2014).
Parca, F., Koca, Y. O. & Unay, A. Nutritional and antinutritional factors of some pulses seed and their effects on human health. Int. J. Second. Metab. 5, 331–342. https://doi.org/10.21448/ijsm.488651 (2018).
Mattila, P. H. et al. Contents of phytochemicals and antinutritional factors in commercial protein-rich plant products. Food Qual. Saf. 2, 213–219. https://doi.org/10.1093/fqsafe/fyy021 (2018).
Eom, S. H. et al. Plant response to cold stress: Cold stress changes antioxidant metabolism in heading type kimchi cabbage (Brassica rapa L. ssp. Pekinensis). Antioxidants 11, 700. https://doi.org/10.3390/antiox11040700 (2022).
Arese, P. et al. Effect of divicine and isouramil on red cell metabolism in normal and G6PD-deficient (Mediterranean variant) subjects. Possible role in the genesis of favism. Progr. Clin. Biol. Res. 55, 725–746 (1981).
De Costa, W., Dennett, M., Ratnaweera, U. & Nyalemegbe, K. Effects of different water regimes on field-grown determinate and indeterminate faba bean (Vicia faba L.). II. Yield, yield components and harvest index. Field Crops Res. 52, 169–178. https://doi.org/10.1016/S0378-4290(96)01060-X (1997).
R Core Team, R. A Language and Environment for Statistical Computing 2021 (R Foundation for Statistical Computing, 2021).
Sivasubramaniam, S. & Madhava Menon, P. Heterosis and inbreeding depression in rice. (1973).
Robinson, H. & Comstock, R. E. Genotypic and Phenotypic Correlations in Corn and Their Implications in Selection (North Carolina State University. Dept. of Statistics, 1950).
Johnson, H. W., Robinson, H. & Comstock, R. Estimates of genetic and environmental variability in soybeans. Agronomy J. 47(7), 314–318 (1955).
Allard, R. Principles of plant breeding. Soil Sci. 91, 414 (1961).
Goyoaga, C. et al. Content and distribution of vicine, convicine and L-DOPA during germination and seedling growth of two Vicia faba L. varieties. Eur. Food Res. Technol. 227, 1537–1542 (2008).
Purves, R. W., Khazaei, H. & Vandenberg, A. Quantification of vicine and convicine in faba bean seeds using hydrophilic interaction liquid chromatography. Food Chem. 240, 1137–1145 (2018).
Elkins, A. C., Rochfort, S. J., Maharjan, P. & Panozzo, J. A simple high-throughput method for the analysis of Vicine and Convicine in Faba bean. Molecules 27, 6288. https://doi.org/10.3390/molecules27196288 (2022).
Majumder, S. et al. A liquid chromatographic method for determination of acetamiprid and buprofezin residues and their dissipation kinetics in paddy matrices and soil. Environ. Sci. Pollut. Res. 29, 1401–1412 (2022).
Lee, Y.-D. & Jang, S.-W. Determination of buprofezin residues in rice and fruits using HPLC with LC/MS confirmation. Korean J. Environ. Agric. 29, 247–256 (2010).
Acknowledgements
The study was supported by the funds from CPGS-AS, CAU (Imphal), Umiam, Meghalaya, India and facilities and instrumentation available at the Institute. It was also supported by the Senior Research Fellowship (SRF) grant awarded by the ICAR, New Delhi, India. Authors are highly thankful to ICAR-NBPGR, New Delhi for sharing diverse germplasm collections and ICAR-IARI, New Delhi for sharing two very important national varieties (Pusa Sumeet and Pusa Udit). We duly acknowledge Dr. Kuldeep Singh, ICRISAT, Patancheru, Hyderabad, India and former Director of ICAR-NBPGR for his generous support in accessing the germplasm collections of NBPGR and his valuable suggestions. We are also highly thankful to our technical staffs and skilled labors for their tireless support throughout the cropping seasons.
Author information
Authors and Affiliations
Contributions
SD, MR, WT and NTM conceptualized the study and designed experiments. NTM, MR and WT supervised the study. SD performed both field and biochemical experiments, curated and analysed phenotypic and biochemical data and wrote the original draft manuscript. SM contributed in the design of biochemical experiments and analysis of results. All the authors have read and edited the draft manuscript and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Debnath, S., Rai, M., Tyagi, W. et al. Lower vicine content reduces the reproductive yield performance in faba bean (Vicia faba L.). Sci Rep 15, 311 (2025). https://doi.org/10.1038/s41598-024-83488-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-83488-z
