
Abstract
The supply of nitrogen (N) and the efficiency with which it is used by phytoplankton serve as two fundamental controls on the productivity of many marine ecosystems. Shifts in nitrogen use efficiency (NUE) can decouple primary production from N-supply but how NUE varies across systems is poorly known. Through a global synthesis of how total N (TN) is apportioned among phytoplankton, particulate, dissolved inorganic, and dissolved organic pools, we demonstrate that NUE underlies broad variations in primary production. Across coastal and open ocean systems, the biomass of autotrophs scales non-linearly with the size of the ecosystem N pool according to a simple equation (chla = 0.004*TN2.38) that captures 68% of the variance in chlorophyll-a (chla) concentration. Such variation in NUE does not arise from organism-level variation in N-use but reflects ecosystem-level shifts in N-distribution among phytoplankton and dissolved organic-N pools. Because these pools differ in their potential for N-retention, shifts in NUE provide a set of common feedback mechanisms that can act to regulate the long-term stock of N in the surface ocean. Cross-system patterns in NUE provide a set of common relationships for predicting how ocean productivity may respond to future perturbations in N-supply.
Similar content being viewed by others

Introduction
The stock of nitrogen (N) in the world’s oceans is apportioned among diverse organic and inorganic pools whose divergent reactivity and residence times govern the global balance of greenhouse gases, productivity of fisheries, and ecological dynamics of microbial and metazoan food webs1,2,3,4. While the supply of N often sets the ultimate upper constraints to marine production5, theoretical predictions also suggest that shifts in the efficiency with which N is acquired and used by primary producers can mediate or decouple the scaling relationship between production and ecosystem N-content (total nitrogen = TN)6. In terrestrial ecosystems, conceptual and empirical models have highlighted the importance of resource use efficiency (RUE) in mediating not only productivity but also stoichiometric feedbacks that regulate nutrient cycling, plant community structure, and trophic interactions7. Indeed, enhanced RUE through complementarity in nutrient use among species is a central mechanistic conceptual underpinning of biodiversity and ecosystem function research8. Nonetheless, in contrast to our understanding of the causal linkages between nutrient supply and primary production in marine ecosystems9, the influence of RUE on marine productivity has remained poorly characterized.
In this study, we define nitrogen use efficiency (NUE) as the proportional efficiency with which ecosystem N (TN) is appropriated by autotrophic biomass (Na) and available for use in net primary production (NPP). Hence, NUE describes the scaling relationship between Na (as proxied by chlorophyll-a) and the total stock of N (TN: sum of all inorganic and organic forms of N) across pelagic systems. If NUE decreases systematically with increasing TN, that would act to dampen cross-system variations in NPP as phytoplankton become less and less efficient at appropriating TN in nutrient-rich systems. Conversely, systematic increases in NUE would accentuate productivity differences between nutrient-poor and nutrient-rich systems. NUE can of course be invariant or vary idiosyncratically across systems, but whether marine production is NUE-neutral or NUE-dependent is not known. Because the supply of N to coastal and open oceans is impacted by direct human alteration of the global N-cycle10 and by climate-sensitive changes in circulation and turbulence-driven N-fluxes11, understanding how NUE mediates the responses of primary production to shifts in N-supply can provide crucial empirical constraints on our forecasts of future ecosystem change. Despite the conceptual utility of NUE, its application to marine ecosystems has been limited. Two prior studies12,13 examined the ratio of chlorophyll-a (chla) to TN and total phosphorus with a focus on identifying limiting nutrient(s) across freshwater and marine systems. Using long-term observations from Chesapeake Bay, Denmark Coast, Tampa Bay, and the Dutch Wadden Sea14, formally applied the NUE concept to examine system-specific responses to nutrient inputs, and identified non-stationary NUE values that narrow options for eutrophication management. Here, we use a global synthesis of the scaling relationship between Na and TN to test if NUE acts as a common determinant of primary production across diverse coastal and open ocean biomes.
Methods
Sample collection
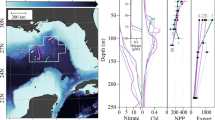
Thirty surface stations on the Oregon inner- to mid-shelves were each sampled 6–10 times from the R/V Elakha as part of a study on the spatial distribution of low oxygen zones by the Partnership for Interdisciplinary Studies of Coastal Ocean (PISCO) program. Surface samples were collected by Niskin bottle casts in the upper 5 m of the water column during April and September 2003 spanning the upwelling season. In addition, thirty-eight open coast stations spanning 1,300 km (between 33° N and 46° N) of the United States west coast and 6 stations on the east and west coasts of the South Island of New Zealand (between 44° S and 42° S) were sampled from the shore from May to August (2002 and 2003) and from Oct to March (2002 to 2003), respectively. All sites were directly exposed to the open ocean and were not located within estuaries or embayments and were previously established to examine the ecological dynamics of rocky intertidal communities15,16. Sample collection is described in17, but briefly, samples were collected by dipping acid-washed 250 ml HDPE bottles into the surf zone. Chla and Particulate nitrogen (PN) samples were filtered in the field onto pre-combusted 25 mm Whatman GF/F glass fiber filters, typically within 30 min, and stored on ice for transport. Samples from New Zealand were held in liquid nitrogen shipping dewar for transport to our laboratory at Oregon State University. The collection and transport of samples in this study were performed in accordance with relevant regulations.
Sample analyses
Chla was determined fluorometrically18 after filters were extracted for 12 h in the dark at – 20 °C in 90% HPLC-grade acetone in a Turner Designs TD700 fluorometer with a detection limit of 0.1 mg and precision of ± 2%. The fluorometer was calibrated against liquid chlorophyll a standard from Turner Designs. Dissolved inorganic nitrogen (DIN), the sum of NO2- + NO3− (N + N), and NH4+ was determined via flow injection analysis colorimetry19 (QuikChem 8500, Lachat Instruments) with detection limits of 0.2 µM, and precision of + /− 5% for both N + N and NH4+. Particulate organic carbon (POC) and particulate nitrogen (PN) was determined via high-temperature combustion and elemental analysis in a CHN analyzer20 (CEC 440HA, Control Equipment Corp), with detection limits of 7 µg and 2 µg for C and N respectively and precision of + /− 0.3% for both analytes. Samples for DIN and POC/PN samples were analyzed at the Analytical Laboratory of the Marine Science Institute, University of California Santa Barbara. Total dissolved organic nitrogen (TDN) was determined via persulfate oxidation and spectrophotometer-based (UV-1201, Shimadzu Corp) colorimetry according to standard methods using EDTA as an oxidation standard21,22. Dissolved organic nitrogen (DON) was calculated as the difference between TDN and DIN. Detection limit and precision after accounting for compounding errors in DIN and TDN analyses were 0.4 µM and ± 8%.
Data compilation and analyses
To increase cross-system coverage, we conducted internet searches for datasets containing observations of chla and total nitrogen (TN), where TN is reported as TN from whole sample digestion or as the sum of total dissolved nitrogen and PN; or DIN, DON, and PN. Not all studies reported or included NH4+. Because we excluded estuarine samples where anthropogenic inputs of NH4+ can be substantial, we anticipate that NH4+ concentrations to be a small portion of the DIN or TN pools consistent with prior studies. For studies that included NH4+ values in our dataset, the mean concentration was 0.46 µM. This sub-micromolar concentration is consistent with values reported from open ocean and shelf systems23,24,25 but remains a potential source of error in estimates of TN. We did not include any data from estuaries or freshwater systems, restricting our data to only comprise open coast, or open ocean observations to focus our analysis on marine ecosystems where a global synthesis has been lacking. Relaxation of nutrient-limitation, light-limitation and/or top-down control can be temporally dynamic and introduce lags between nutrient supply and primary production. Because we are focused on the mean state of an ecosystem rather than the nature of within system variability, for the relationships between chla, PN, POC, DON, and TN, we included only surface data from stations that were occupied more than once in order to arrive at system means. Where available, surface layer followed the definition of the data source; otherwise the surface layer was defined as the upper 25 m. In total, 391 stations from our field sampling and data compilation efforts spanning coastal and open ocean biomes from 80° N to 70° S (Supplementary Table S1, Supplementary Fig. S1) met the criteria of paired chla and TN observations and temporally repeated sampling. We further derived a maximum NUE reference line based on the Redfield ratio for C:N26 and the globally-averaged minimum cellular organic carbon: chla (C:chla) ratio27 (Fig. 1). Depth profiles of TN and DIN were poorly represented within the primary dataset of 391 sites. Consequently, we expanded our criteria to include all readily available data of TN and DIN depth profiles regardless of sampling frequency. A complete list of data sources can be found in Supplementary Table S1. NUE was calculated as the yield of chla relative to TN:

Cross-system pattern of NUE. Reference lines for maximum (100%) and minimum (1%) NUE based on globally averaged C:chla ratio of 48.9 are depicted as A, and C, respectively. Line B reflects maximum NUE if all TN was incorporated as Na, but assumes globally-averaged maximum C:chla ratio of 134.8 from ref39. Back-transformed orthogonal regression equation yields chla = 0.004* TN2.54, n = 391 individual sites. Site means reflect multiple surface layer observations across time.
Orthogonal regressions, accounting for error on both independent and dependent axes were used to compute slope and intercepts on log-transformed data where necessary.
Results and discussion
We synthesized observations of TN and chla that span 170 degrees of latitude representing 37 ecoregions28 or 30 large marine ecosystems29. Our dataset comprised four orders of magnitude in chla concentration, representing nearly the full dynamic range of phytoplankton biomass recorded for the surface ocean. Global variation in chla shows high correlation (R2 = 0.68) with changes in TN (Fig. 1). This scaling relationship between chla and TN is highly non-linear (chla = 0.004*TN2.38) and reflects progressive increases in NUE. That is, NUE values from the most TN-rich upwelling systems are as much as 80-fold higher than those from TN-poor oligotrophic gyres. Relative to a maximum NUE value where Na comprises all ecosystem N, observed NUE increases from < 0.5% to > 95% of its maximum potential value along the global TN gradient (Fig. 1). The magnitude of NUE variation indicates that major variation in marine autotrophic biomass does not simply reflect differences in the size of the ecosystem N pool but is subject to broad control by cross-system variation in the efficiency with which ecosystem N is appropriated by phytoplankton. By examining considerably more marine systems, our findings expand from these previous comparative studies12,13,14 to reveal a global scaling relationship between TN and marine production for ocean ecosystems. Across coastal and open ocean ecosystems, chla-poor (rich) systems are always associated with low (high) NUE values (Supplementary Figs. S2, S3). This pattern holds when coastal and open ocean systems are disaggregated (Supplementary Fig. S13) and suggests that NUE underlies general variations in chla in the sea.
What is the basis of this pattern of increasing NUE with increasing TN and chla? Direct mass balance between TN and chla yield (i.e. maximum NUE line that assumes all TN is converted into phytoplankton biomass, Fig. 1) also indicates that for large areas of the open ocean, there is sufficient TN to support much higher levels of chla than is realized. This suggests that shifts in NUE alone have the potential to drive sizeable changes in the productivity of marine systems. Residuals around the regression line indicate that chla can vary by 1 order of magnitude at any specific TN pool size. Such variation can further reflect factors such as top-down control30, light limitation31, and/or co-limitation by other nutrients32. Across a series of well-monitored temperate systems, chla scales as a declining function of TN with exponents ranging from 0.68 to 0.92, possibly as due to phytoplankton self-shading14.
It is important to consider if the systematic shift in NUE may in fact be a consequence of nutrient or resource co-limitation that covaries with TN. Iron-limited upwelling regions are marked by high nitrate concentrations, with N-limitation most consistently expressed in subtropical gyre systems33. This would suggest high values of NUE in oligotrophic TN-poor (and light-replete) gyre systems and low values in TN-rich upwelling systems where Fe-limitation may constrain production. This prediction is opposite to what was observed in our data (Fig. 1). We do note that chla reported for the Indian Ocean sector of the Southern Ocean, a system where Fe-limitation is prevalent32 is markedly low relative to the global regression line (Fig. 1, Supplementary Fig. S2).
Extremely low values of NUE appear to be a pervasive feature of many open ocean systems. Minimum NUE values indicate that in many nutrient-poor systems phytoplankton may be able to use as little as 1% or less of the nitrogen available (Fig. 1). This pattern of declining NUE with the increasing scarcity of N is surprising given the common conceptualization of oligotrophic ocean gyres as systems where available nutrients are readily depleted tightly recycled and held in the biomass of autotrophs. However, our results indicate that such mechanisms do not lead to disproportionate increases in the amount of N held in the biomass of autotrophs in N-poor systems. Alternative N sinks in marine systems include dissolved inorganic nitrogen (DIN), DON, particulate detritus, bacteria, and other heterotrophs. Distribution of N into these alternative pools can mediate phytoplankton success via direct competition for N or indirectly by increasing herbivore biomass and impacts.
Our assessment indicates that DON progressively dominates the ecosystem N pool as one moves from coastal to open ocean systems and as TN values converge toward global minimum values (Fig. 2, Supplementary Figs. S5, S6). The quantitative dominance of the DON pool has long been recognized in oligotrophic systems34,35. Our results indicate that this dominance is strongly dependent on the size of the ecosystem N pool. Although DON uptake can at times, represent a sizeable fraction of N-uptake needs by phytoplankton35, the dominance of DON in the least productive systems re-enforces the understanding that the bulk DON pool may be of limited bioavailability36.

Global pattern in the relative mass dominance of DON along a gradient of increasing TN pool sizes, n = 194 individual sites.
Extending from the concept of the microbial carbon pump37, it has been recently proposed38 that the turnover of organic-N through a microbial nitrogen pump results in a progressive accumulation of recalcitrant DON that reduces the amount of labile DON available for recycled production. Observationally, in a transect across 75 degrees of longitude in the South Pacific, rates of primary production varied inversely and strongly with the size of the photic zone DON39. This supports the potential for the sequestration of ecosystem N into the DON pool to serve as a fundamental general steady-state constraint on marine production in oligotrophic systems.
The question of bioavailability is important to consider6. If DON is viewed as a biologically unavailable pool, then should it be included in the calculation of TN? DON is of course not a single inert pool but comprises pools that vary widely in lability35,36. Ideally, TN would include only the portion of the DON pool that is bioavailable. While the characterization of DON lability continues to advance40, our quantitative understanding of how DON is apportioned between bioavailable and recalcitrant pools across systems remains limited. In our analysis, we’ve opted to define ecosystem N as the total pool of fixed N that can potentially be available to primary producers. Nonetheless, because DON dominates the ecosystem N pool in oligotrophic systems (Fig. 2), it is important to assess the effects of excluding DON. We find that the exclusion of DON from the TN pool (Supplementary Fig. S7) does not reverse the pattern of increasing NUE with TN (Fig. 1, Supplementary Fig. S2). Chla increases as a non-linear function of PN + DIN (Chla = 0.084* PN + DIN1.65, R2 = 0.71), such that the ratio of chla:PN + DIN in the most oligotrophic system is 4 orders of magnitude lower than that for the most nutrient-rich systems (Supplementary Fig. S7). While the pool of recalcitrant DON likely serves as a general upper constraint to NUE, the variations in DON concentration within and across ecosystems suggest that the degree to which DON serves as a sink or source for autotrophic N is a property of individual systems. The inclusion of DON in the ecosystem N pool provides a means to measure that property.
Our assessment of NUE uses chla concentration as an index of phytoplankton biomass (Na) and net primary production (NPP). What is the justification for this decision? Because chla:Na varies as physiological and phylogenetic responses to changing environmental conditions41, variations in NUE may simply reflect physiological shifts at the organism-scale rather than ecosystem-level changes in N appropriation by autotrophs. Our maximum NUE line (Fig. 1) is calculated from the commonly measured cellular organic carbon: chla (C:chla) ratio and the Redfield C:N ratio, where C:chla reflects the minimum globally-averaged value (48.9) reported in27. A secondary reference line that is based a maximum globally-averaged C:chla value of 134.837 serves to bound the effects of chla:Na variation. The C:chla can vary considerably depending on system and physiological state of phytoplankton42,43. Assuming a lower (higher) C:chla ratio as might be encountered in nutrient-replete and/or light-limited (nutrient deficient and/or light-saturated) cells41 would act to move the maximum NUE line higher (lower). Thus, the two reference lines A, B, (Fig. 1, Supplementary Fig. S8) provide a bound for the effects of a C:chla ratio range of 85.9. The vast majority of chla values fall outside this range indicating limits of organism-scale shifts in chla:Na stoichiometry to account for cross-system variation in NUE.
Direct measures of Na would of course sidestep the need for stoichiometric assumptions but Na is notoriously difficult to ascertain, requiring separation of phytoplankton cells from other particulate materials44, and are currently unavailable for cross-ocean biome comparisons. Besides chla, bulk particulate nitrogen (PN) may provide an alternative index of Na. However, PN comprises detrital and heterotrophic N in addition to autotrophic N. Nonetheless, PN does serve as an upper constraint on potential Na. For example, across systems, PN and chla are strongly coupled (R2 = 0.82) with the chla: PN ratio increasing by approximately 1 order of magnitude across the range of observed values (Supplementary Fig. S9). This shift is consistent with previously documented increases in the ratio of autotroph: heterotroph biomass along marine productivity gradients45,46. Because PN and chla are tightly coupled (Supplementary Fig. S9), like chla, the relative size of the PN pool also increases non-linearly along the TN gradient (Fig. 3).

Particulate nitrogen (PN) and total nitrogen (TN) across systems. Back-transformed orthogonal regression equation PN = 0.027 TN1.69, n = 205 individual sites.
A second assumption in our analysis is the utility of chla in indexing NPP. This reflects the strong empirical relationship between the standing stock of chla and NPP across open ocean and coastal biomes27,47. Nonetheless inferring flux from stock is subject to uncertainties in how phytoplankton turnover rates vary across systems, and NPP is most robustly modeled when environmental factors such as photosynthetically active radiation, mixed layer depth, and temperature accompany chla estimates48. The lowest values of NUE were found in subtropical oligotrophic ocean gyres where the dominance of phytoplankton biomass by picoplankton49 would suggest the potential for phytoplankton turnover rates to compensate for reduced chla. The 4 orders of magnitude of differences in chla described here would require phytoplankton in low chla systems to turnover at rates that are 4 orders of magnitude faster to equal the productivity of high chla systems. Such rates were not documented in in situ phytoplankton growth rate measurements across systems50. Broad scale cross-biome surveys report turnover rates that vary by only 2 to 4 folds, often with the highest rates encountered in productive, chla-rich systems51,52,53. These observations suggest that potential turnover rate differences alone cannot explain the globally divergent patterns in NUE.
Does NUE arise from uptake or production efficiency? Mechanistically, nitrogen use efficiency must ultimately scale from the (1) efficiency with which available N is acquired by autotrophs (uptake efficiency), and (2) rate of NPP per unit of acquired N (production efficiency)54. Production efficiency can be particularly important in terrestrial systems where acquired N is used repeatedly to support the fixation of carbon into N-poor woody biomass55. In aquatic short-lived autotrophs under balanced growth, tissue C: N approximates the ratio of C to N uptake56, and can be used as a measure of production efficiency. Deviations in NUE due to production efficiency would thus manifest as departures from the Redfield ratio in particulate organic-C (POC) and PN. This ratio can exhibit marked within-region variation57. But globally, POC: PN was highly conserved in our dataset, showing strong adherence to the Redfield ratio across the 4 orders of magnitude variation in particulate organic matter concentration (Supplementary Figs. S10, S11) and consistent with previous cross-system compilations58. These patterns suggest that like chla: Na, variation in NPP:Na as proxied by POC:PN is relatively conserved at the cross-biome scale and is unlikely to serve as a determinant of NUE variation in our analysis.
Deviations in phytoplankton production efficiency can also theoretically arise from the extracellular release of dissolved organic carbon (DOC) and/or high C:N compounds. For example, exudation of extracellular DOC comprises on average ~ 20% of NPP, though in oligotrophic systems, extracellular DOC release can comprise up to 50% of NPP59. With respect to C:N, the mean ocean surface C:N ratio of bulk dissolved organic matter does not deviate strongly from the Redfield ratio5, averaging 14.9:1 globally60. These observations suggest extracellular DOC releases or non-Redfield ratio of dissolved organic matter releases alone, may be insufficient to give rise to sufficiently variable production efficiency values given the 4 orders of magnitude variation we observed in chla (Fig. 1).
The size of the ecosystem N pool is surprisingly conserved across a diverse array of marine biomes (Fig. 4a). Coastal sites exhibit greater mean and range than open ocean sites, but the central 90% of all surface TN observations fell between 6.4 and 37.2 µM. Because DIN comprises a major portion of the TN in deep waters (Fig. 4b), mixing and transport can drive increased N-availability in general. This increase in the supply of readily useable N by phytoplankton thus is likely to contribute to the trend towards increased NUE in more TN-rich systems. The absence of TN endmembers > 40 µM in waters of intermediate depth (150–500 m) suggests that high coastal TN values can further reflect processes such as terrestrial inputs10 and/or nutrient accumulation61 on continental shelves.

Depth profiles of TN (a) and DIN:TN (b) to 2000 m across systems. Coastal blue circle and open ocean black circle profiles include additional sites that were sampled only once (Supplementary Fig. S1). A total of 3184 individual profiles are represented. LOWESS regression lines are depicted separately for coastal yeollow line and open ocean red line sites.
The relatively narrow distribution of TN values (Fig. 1) suggests that the size of the marine ecosystem N pool may be subject to strong feedbacks that act to constrain the depletion as well as accumulation of N in surface waters. Our observations reveal a systematic trade-off between the dominance of phytoplankton and DON pools along the TN gradient (compare Figs. 1, 2 and 3). This trade-off offers one general mechanism for regulating ecosystem N pool size in the ocean. Since the sinking of particulate organic matter represents the principal loss of N from surface mixed layers, the increased sequestration of N into phytoplankton biomass, and shifts toward larger, more sedimentation-prone phytoplankton taxa in more productive systems62 would serve to constrain the long-term accumulation of N in high TN waters. The reductions in NUE and dominance of DON—a pool not subject to direct sedimentary losses in oligotrophic systems—can act in an opposing fashion to regulate the loss of N from surface waters and set the lower limit to the size of the TN pool. These feedbacks suggest that NUE may act as a key feature coupling the internal biogeochemical structure of marine ecosystems with the regulation of the size of N pools in the surface ocean.
In terrestrial systems, NUE is often conceptualized as declining along gradients of increasing ecosystem fertility6,63. This pattern arises because mechanisms that act to conserve the loss of limiting resources such as selective resorption of N from senescing leaves, and/or maximize production efficiency, relax with increasing nutrient availability7,54. A reasonable expectation is that analogous mechanisms such as the ability to draw down DIN to exceedingly low concentrations or tight retention and recycling of N in oligotrophic marine systems would also act to maximize NUE. In contrast, marine NUE is lowest in nutrient-poor systems such as sub-tropical gyres and highest in nutrient-rich systems such as upwelling shelves (Fig. 1). This suggests that high uptake affinities for DIN or closed N-cycle associated with oligotrophic systems are insufficient for maximizing NUE at the ecosystem scale. As noted above, NUE is the product of production efficiency in the rate of NPP per unit of Na and N uptake efficiency in the acquisition of N from the environment. Consequently, cross-system differences in NUE can be driven by shifts in the relative importance of production and uptake efficiencies. In terrestrial systems, where long-lived plants exhibit substantial internal recycling of N, production efficiency can vary widely and play a key role in modulating NUE along gradients of N-fertility. For pelagic systems considered here, nutrient uptake efficiency is likely to play the dominant role in determining cross-system variation in NUE.
The mechanism(s) that drive the progressive increase in NUE and uptake efficiency with TN cannot be resolved by our correlative study. Shifts in the strength of top-down control of phytoplankton by consumers along the microbial vs. metazoan food web continuum64, or transient increases in DIN availability in upwelling and seasonally destratified systems65 are alternative processes that can structure and reinforce the ability of autotrophs to effectively appropriate ecosystem N. These represent logical next steps in bringing a mechanistic understanding to controls on NUE across systems.
Future efforts that expand TN and chla observations to systems where limitation or co-limitation by other nutrients will be important to test the stability of the global scaling relationship. In particular, large regions of the ocean are limited by iron but replete in nitrogen. Prominent high-nutrient, low-chlorophyll (HNLC) systems include the Southern Ocean, Northeast Pacific, and Equatorial Pacific upwelling. While these systems are represented in our dataset, the number of observations is limited relative to other coastal and other oceanic sites. As systems that are essentially defined by having low NUE values, a comprehensive representation of HNLC systems would be invaluable. The stability of NUE in time also deserves attention. Carstensen et al.14 found strong evidence of non-stationarity in NUE within individual coastal estuarine systems potentially in response to alterations in nutrient inputs, food web dynamics or climate change.
Our results indicate that in marine ecosystems, realized variation in NUE can impose fundamental constraints on productivity. While the dependence of pelagic production on ecosystem nutrient pool size in freshwater ecosystems is a central tenet of predictive limnology66,67, our results highlight a similarly important and globally pervasive coupling between nutrient pool size and chla across marine ecosystems. Despite the heterogeneous nature of marine biomes, our analyses reveal common responses of marine biogeochemical structure to changes in the size of the ecosystem N pool. The linkage between TN and NUE further indicates incremental changes in TN can result in strongly non-linear responses in marine primary production. Accelerating inputs of anthropogenic N10,68,69 and climatically-driven shifts in transport and mixing11,70 are central perturbations to the N-cycles of marine ecosystems. For many marine systems facing reductions in nitrogen supply from warming waters, the global functional relationship between TN and NUE suggests the likelihood for empirically-constrained nonlinear declines in primary production.
Data availability
The datasets generated during and/or analyzed during the current study are available at: https://github.com/FChanOSU/NUE.
References
Bronk, D.A., Sipler, R.E., Letscher, R.T., McCarthy, M.D. Dissolved organic nitrogen. In Biogeochemistry of Marine Dissolved Organic Matter 343–404. (Academic Press, 2024).
Romera-Castillo, C., Letscher, R. T. & Hansell, D. A. New nutrients exert fundamental control on dissolved organic carbon accumulation in the surface Atlantic Ocean. Proc. Natl. Acad. Sci. 113(38), 10497–10502 (2016).
Checkley, D. M. Jr., Asch, R. G. & Rykaczewski, R. R. Climate, anchovy, and sardine. Ann. Rev. Mar. Sci. 9, 469–493 (2017).
Buchanan, P. J., Aumont, O., Bopp, L., Mahaffey, C. & Tagliabue, A. Impact of intensifying nitrogen limitation on ocean net primary production is fingerprinted by nitrogen isotopes. Nat. Commun. 12(1), 6214 (2021).
Falkowski, P. G. Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2 in the ocean. Nature 387(6630), 272–275 (1997).
Hodapp, D., Hillebrand, H. & Striebel, M. “Unifying” the concept of resource use efficiency in ecology. Front. Ecol. Evol. 6, 233. https://doi.org/10.3389/fevo.2018.00233 (2019).
Vitousek, P. Nutrient cycling and nutrient use efficiency. Am. Nat. 119(4), 553–572 (1982).
Tilman, D., Lehman, C. L. & Thomson, K. T. Plant diversity and ecosystem productivity: theoretical considerations. Proc. Natl. Acad. Sci. 94(5), 1857–1861 (1997).
Howarth, R. W. Nutrient limitation of net primary production in marine ecosystems. Annu. Rev. Ecol. Syst. 19(1), 89–110 (1988).
Schlesinger, W. H. On the fate of anthropogenic nitrogen. Proc. Natl. Acad. Sci. 106(1), 203–208 (2009).
Sydeman, W. J. et al. Climate change and wind intensification in coastal upwelling ecosystems. Science 345(6192), 77–80 (2014).
Guildford, S. J. & Hecky, R. E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: is there a common relationship?. Limnol. Oceanogr. 45(6), 1213–1223 (2000).
Smith, V. H. Responses of estuarine and coastal marine phytoplankton to nitrogen and phosphorus enrichment. Limnol. Oceanogr. 51(1part2), 377–384 (2006).
Carstensen, J., Sánchez-Camacho, M., Duarte, C. M., Krause-Jensen, D. & Marbà, N. Connecting the dots: responses of coastal ecosystems to changing nutrient concentrations. Environ. Sci. Technol. 45(21), 9122–9132 (2011).
Menge, B.A., Lubchenco, J., Bracken, M.E.S., Chan, F., Foley, M.M., Freidenburg, T.L., Gaines, S.D., Hudson, G., Krenz, C., Leslie, H. and Menge, D.N.L. Coastal oceanography sets the pace of rocky intertidal community dynamics. In Proceedings of the National Academy of Sciences of the United States of America, 12229–12234 (2003).
Menge, B. A. et al. Integrating coastal oceanic and benthic ecological approaches for understanding large-scale meta-ecosystem dynamics. Oceanography 32(3), 38–49 (2019).
Menge, B. A., Close, S. L., Hacker, S. D., Nielsen, K. J. & Chan, F. Biogeography of macrophyte productivity: Effects of oceanic and climatic regimes across spatiotemporal scales. Limnol. Oceanogr. 66(3), 711–726 (2021).
Welschmeyer, N. A. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnol. Oceanogr. 39(8), 1985–1992 (1994).
Johnson, K. S. & Petty, R. L. Determination of nitrate and nitrite in seawater by flow injection analysis 1. Limnol. Oceanogr. 28(6), 1260–1266 (1983).
Karl, D. M., Dore, J. E., Hebel, D. V. & Winn, C. Procedures for particulate carbon, nitrogen, phosphorus and total mass analyses used in the US-JGOFS Hawaii Ocean Time-series program. In Marine Particles: Analysis and Characterization (eds Hurd, D. C. & Spencer, D. W.) 71–77 (American Geophysical Union, 1991).
Valderrama, J. C. The simultaneous analysis of total nitrogen and total phosphorus in natural waters. Mar. Chem. 10(2), 109–122 (1981).
Grasshoff, K., Ehrhardt, M., Kremling, K. Methods of Seawater Analysis. (Wiley-VCH, 1999).
Harrison, W. G., Harris, L. R. & Irwin, B. D. The kinetics of nitrogen utilization in the oceanic mixed layer: Nitrate and ammonium interactions at nanomolar concentrations. Limnol. Oceanogr. 41(1), 16–32 (1996).
Rees, A. P., Woodward, E. M. S. & Joint, I. Concentrations and uptake of nitrate and ammonium in the Atlantic Ocean between 60∘ N and 50∘ S. Deep Sea Res. Part II Top. Stud. Oceanogr. 53(14–16), 1649–1665 (2006).
Kokkinakis, S. A. & Wheeler, P. A. Nitrogen uptake and phytoplankton growth in coastal upwelling regions 1. Limnol. Oceanogr. 32(5), 1112–1123 (1987).
Redfield, A. C. The biological control of chemical factors in the environment. Am. Sci. 46(3), 230A-A221 (1958).
Behrenfeld, M. J., Boss, E., Siegel, D. A. & Shea, D. M. Carbon-based ocean productivity and phytoplankton physiology from space. Glob. Biogeochem. Cycles 19(1), GB1006. https://doi.org/10.1029/2004GB002299 (2005).
Spalding, M. D. et al. Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. BioScience 57(7), 573–583 (2007).
Sherman, K. & Duda, A. M. Large marine ecosystems: an emerging paradigm for fishery sustainability. Fisheries 24(12), 15–26 (1999).
Calbet, A. & Landry, M. R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnol. Oceanogr. 49(1), 51–57 (2004).
Ardyna, M. & Arrigo, K. R. Phytoplankton dynamics in a changing Arctic Ocean. Nat. Clim. Change 10(10), 892–903 (2020).
Browning, T. J. & Moore, C. M. Global analysis of ocean phytoplankton nutrient limitation reveals high prevalence of co-limitation. Nat. Commun. 14(1), 5014 (2023).
Karl, D. M. et al. Ecological nitrogen-to-phosphorus stoichiometry at station ALOHA. Deep Sea Res. Part II Top. Stud. Oceanogr. 48(8–9), 1529–1566 (2001).
Knapp, A. N., Casciotti, K. L. & Prokopenko, M. G. Dissolved organic nitrogen production and consumption in eastern tropical South Pacific surface waters. Glob. Biogeochem. Cycles 32(5), 769–783 (2018).
Berman, T. & Bronk, D. A. Dissolved organic nitrogen: a dynamic participant in aquatic ecosystems. Aquat. Microb. Ecol. 31(3), 279–305 (2003).
Aluwihare, L. I., Repeta, D. J., Pantoja, S. & Johnson, C. G. Two chemically distinct pools of organic nitrogen accumulate in the ocean. Science 308(5724), 1007–1010 (2005).
Jiao, N. et al. Microbial production of recalcitrant dissolved organic matter: long-term carbon storage in the global ocean. Nat. Rev. Microbiol. 8(8), 593–599 (2010).
Yamaguchi, Y. T. & McCarthy, M. D. Sources and transformation of dissolved and particulate organic nitrogen in the North Pacific Subtropical Gyre indicated by compound-specific δ15N analysis of amino acids. Geochim. Cosmochim. Acta 220, 329–347 (2018).
Raimbault, P., Garcia, N. & Cerutti, F. Distribution of inorganic and organic nutrients in the South Pacific Ocean—evidence for long-term accumulation of organic matter in nitrogen-depleted waters. Biogeosciences 5(2), 281–298 (2008).
Broek, T. A. et al. Dominant heterocyclic composition of dissolved organic nitrogen in the ocean: A new paradigm for cycling and persistence. Proc. Natl. Acad. Sci. 120(49), e2305763120 (2023).
Geider, R. J., MacIntyre, H. L. & Kana, T. M. Dynamic model of phytoplankton growth and acclimation: responses of the balanced growth rate and the chlorophyll a: carbon ratio to light, nutrient-limitation and temperature. Mar. Ecol. Progr. Ser. 148, 187–200 (1997).
Riemann, B., Simonsen, P. & Stensgaard, L. The carbon and chlorophyll content of phytoplankton from various nutrient regimes. J. Plankton Res. 11(5), 1037–1045 (1989).
Graff, J. R., Westberry, T. K., Milligan, A. J., Brown, M. B. & G. Dall’Olmo, V. van Dongen-Vogels, K. M. Reifel, M. J. Behrenfeld.,. Analytical phytoplankton carbon measurements spanning diverse ecosystems. Deep Sea Res. Part I Oceanogr. Res. Pap. 102, 16–25 (2015).
Hessen, D. O., Andersen, T., Brettum, P. & Faafeng, B. A. Phytoplankton contribution to sestonic mass and elemental ratios in lakes: implications for zooplankton nutrition. Limnol. Oceanogr. 48(3), 1289–1296 (2003).
Gasol, J. M., Del Giorgio, P. A. & Duarte, C. M. Biomass distribution in marine planktonic communities. Limnol. Oceanogr. 42(6), 1353–1363 (1997).
Kang, H. C. et al. Food web structure for high carbon retention in marine plankton communities. Sci. Adv. 9(50), eadk0842 (2023).
Marañón, E. et al. Resource supply overrides temperature as a controlling factor of marine phytoplankton growth. PLoS One 9(6), e99312 (2014).
Westberry, T. K., Silsbe, G. M. & Behrenfeld, M. J. Gross and net primary production in the global ocean: An ocean color remote sensing perspective. Earth-Sci. Rev. 237, 104322 (2023).
Agawin, N. S., Duarte, C. M. & Agustí, S. Nutrient and temperature control of the contribution of picoplankton to phytoplankton biomass and production. Limnol. Oceanogr. 45(3), 591–600 (2000).
Laws, E. A. Evaluation of in situ phytoplankton growth rates: a synthesis of data from varied approaches. Annu. Rev. Mar. Sci. 5, 247–268 (2013).
Marañón, E., Holligan, P. M., Varela, M., Mouriño, B. & Bale, A. J. Basin-scale variability of phytoplankton biomass, production and growth in the Atlantic Ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 47(5), 825–857 (2000).
Huete-Ortega, M., Calvo-Díaz, A., Graña, R., Mouriño-Carballido, B. & Marañón, E. Effect of environmental forcing on the biomass, production and growth rate of size-fractionated phytoplankton in the central Atlantic Ocean. J. Mar. Syst. 88(2), 203–213 (2011).
Henderikx Freitas, F., White, A. E. & Quay, P. D. Diel measurements of oxygen-and carbon-based ocean metabolism across a trophic gradient in the North Pacific. Glob. Biogeochem. Cycles. 34(11), e2019GB006518 (2020).
Bridgham, S. D., Pastor, J., McClaugherty, C. A. & Richardson, C. J. Nutrient-use efficiency: a litterfall index, a model, and a test along a nutrient-availability gradient in North Carolina peatlands. Am. Nat. 145(1), 1–21 (1995).
Berendse, F. & Aerts, R. Nitrogen-use-efficiency: a biologically meaningful definition?. Funct. Ecol. 1, 293–296 (1987).
Ågren, G. I. The C: N: P stoichiometry of autotrophs–theory and observations. Ecol. Lett. 7(3), 185–191 (2004).
Sterner, R. W. et al. Scale-dependent carbon: nitrogen: phosphorus seston stoichiometry in marine and freshwaters. Limnol. Oceanogr. 53(3), 1169–1180 (2008).
Galbraith, E. D. & Martiny, A. C. A simple nutrient-dependence mechanism for predicting the stoichiometry of marine ecosystems. Proc. Natl. Acad. Sci. 112(27), 8199–8204 (2015).
Roshan, S. & DeVries, T. Efficient dissolved organic carbon production and export in the oligotrophic ocean. Nat. Commun. 8(1), 2036 (2017).
Liang, Z., Letscher, R. T. & Knapp, A. N. Global patterns of surface ocean dissolved organic matter stoichiometry. Glob. Biogeochem. Cycles 37(12), e2023GB007788 (2023).
Flynn, R. F. et al. On-shelf nutrient trapping enhances the fertility of the southern Benguela upwelling system. J. Geophys. Res. Oceans. 125(6), e2019JC015948 (2020).
Buesseler, K. O. The decoupling of production and particulate export in the surface ocean. Glob. Biogeochem. Cycles 12(2), 297–310 (1998).
Shaver, G. R. & Melillo, J. M. Nutrient budgets of marsh plants: efficiency concepts and relation to availability. Ecology 65, 1491–1510 (1984).
Vargas, C. A. et al. The relative importance of microbial and classical food webs in a highly productive coastal upwelling area. Limnol. Oceanogr. 52(4), 1495–1510 (2007).
Fawcett, S. E. & Ward, B. B. Phytoplankton succession and nitrogen utilization during the development of an upwelling bloom. Mar. Ecol. Progr. Ser. 428, 13–31 (2011).
Peters, R. H. The role of prediction in limnology. Limnol. Oceanogr. 31(5), 1143–1159 (1986).
Graeber, D. et al. Consistent stoichiometric long-term relationships between nutrients and chlorophyll-a across shallow lakes. Nat. Commun. 15(1), 809 (2024).
Jickells, T. D. et al. A reevaluation of the magnitude and impacts of anthropogenic atmospheric nitrogen inputs on the ocean. Glob. Biogeochem. Cycles 31(2), 289–305 (2017).
Yamamoto, A. et al. Competing and accelerating effects of anthropogenic nutrient inputs on climate-driven changes in ocean carbon and oxygen cycles. Sci. Adv. 8(26), eabl9207 (2022).
Moore, J. K. et al. Sustained climate warming drives declining marine biological productivity. Science 359(6380), 1139–1143 (2018).
Acknowledgements
We thank all the originators of data used in this synthesis (see Supplementary Table S1 for a complete listing). We thank P. Wheeler, and R. W. Howarth for their comments on previous versions of this manuscript. We also thank four anonymous reviewers whose comments greatly improved the manuscript. Funding was provided by NSF and the David and Lucile Packard Foundation. This is contribution 543 of the Partnership for the Interdisciplinary Studies of Coastal Oceans (PISCO) program.
Author information
Authors and Affiliations
Contributions
F.C. conceptualized the paper, performed the analyses, and was the primary writer of the manuscript, J.L. and K.N. worked in conceptualizing the paper. F.C., K.N., and B.M collected the field data. J.L., K.N, and B.M. reviewed and edited the paper. B.M., and J.L., acquired the core funding that enabled this work.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chan, F., Nielsen, K.J., Lubchenco, J. et al. Nitrogen use efficiency underlies cross-ecosystem variation in marine primary production. Sci Rep 14, 32146 (2024). https://doi.org/10.1038/s41598-024-84019-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-84019-6
