
Abstract
Uveitis is a leading cause of blindness in the world and autoimmune uveitis is an ocular tissue specific autoimmune disease. Utilizing experimental autoimmune uveitis (EAU), we can interrogate different immune responses in the mouse that are relevant to the human disease. cytotoxic T-lymphocyte antigen 4 (CTLA-4) is an immune checkpoint molecule that has different roles depending on the target tissue. In this work we investigate the role of CTLA-4 on CD4 T cells in ocular tissue during EAU. We find that CTLA-4 is needed for both the severity of disease but also timely resolution of disease. Regulatory T cells (Tregs) that emerge in the spleen during resolution of EAU require CTLA-4 to suppress disease, but ocular Tregs that emerge in the eye do not require CTLA-4 to suppress disease. This report provides an additional understanding of CTLA-4 on Tregs that is specific for ocular tissue. The implications of this work are that circulating Tregs in uveitis patients may require CTLA-4 to suppress ocular inflammation but once in the eye the function of CTLA-4 is dispensable.
Similar content being viewed by others
Introduction
Uveitis is an ocular inflammatory condition that causes blindness worldwide1,2,3. Experimental autoimmune uveitis (EAU) serves as a widely used mouse model of autoimmune uveitis4. Tregs are key players in maintaining immune tolerance and preventing chronic inflammation, providing a buffer against autoimmune diseases like uveitis6. An association of Tregs with uveitis remission has been observed in the clinic, and ocular antigen specific Tregs that emerge in the spleen at remission of uveitis (post-EAU Tregs) provide resistance to EAU5,6,7,8,9. The emergence of post-EAU Tregs is a dynamic process over the course of EAU7, and how these Tregs function is of interest.
Tregs that recognize ocular antigen emerge in the secondary lymphoid tissue and the eye EAU8. The Caspi lab has been demonstrated that Tregs are needed for the normal resolution of EAU10. However, other Treg subsets that provide resistance to relapsing EAU but do not contribute to the resolution of EAU have been identified11. Therefore, multiple Treg subsets exist that have different functional roles during uveitis and in the prevention of relapsing uveitis. We have identified distinct TIGIT+ Tregs and PD-1+ Tregs that suppresses EAU, and each have different homing requirements9, but it is not known which effector molecules are utilized to suppress. We have previously identified that cytotoxic T-lymphocyte antigen 4 (CTLA-4) is expressed on each of these Treg subsets8, so we examined if CTLA-4 is required for post-EAU Tregs to suppress disease.
CTLA-4 is expressed on Treg cells and limits the immune response, CTLA-4 knockout mice die at four weeks of age of lymphoproliferative disease12,13. The Sakaguchi lab first developed a floxed CTLA-4 mouse on a BALB/c background and found that Treg specific deletion of CTLA-4 resulted in T cell mediated autoimmune disease and reduced CD80 and CD86 expression on dendritic cells14. The Sharpe lab developed a conditional CTLA-4 mouse on a C57BL/6 background and discovered a role for CTLA-4 in Treg development and effector T cell development15. Sharpe, et al., also showed that mice were resistant to experimental autoimmune encephalomyelitis (EAE) and less capable of limiting tumor growth due to an immunocompensatory mechanism that resulted in an upregulation of IL-10, LAG3, and PD-1 15. Klocke, et al., then examined collagen induced arthritis (CIA) and EAE with the conditional CTLA-4 on a C57BL/10.Q background and found that a CTLA-4 deficiency caused CIA to be more severe, but protected against EAE or delayed EAE onset when a much larger peptide (residues 1–125) is used16. These studies suggest CTLA-4 may have a tissue specific role and suggests CTLA-4 may have a role in ocular antigen specific Tregs.
In this report we asked if a CTLA-4 deficiency has an impact on EAU, if post-EAU Tregs from the spleen require CTLA-4 to suppress disease, and if ocular post-EAU Tregs can suppress disease when administered systemically and if this suppression requires CTLA-4. Ctla-4fl/fl were provided by the Sharpe lab and bred to Cd4Cre−ERT2 mice to determine the role of CTLA-4 specifically on CD4 T cells during autoimmune uveitis and on post-EAU Treg cells. We found that a CTLA-4 deficiency did not affect the onset of disease, but delayed resolution, post-EAU Tregs required CTLA-4 to suppress, and we show for the first time that post-EAU Tregs taken from the resolved eye are capable of suppressing disease and is not dependent on CTLA-4.
Materials and methods
Mice
The University of the University of Massachusetts Medical School IACUC (UMMS IACUC) approved all mouse procedures described in this study and all mouse study methods were carried out in accordance with the relevant guidelines approved by the UMMS IACUC and by the ARRIVE. The C57BL/6 J and CD4Cre−ERT2 mice used in this study were purchased from Jackson Laboratories. The Ctla-4fl/flwere generously provided by Dr. Arlene Sharpe and described in Paterson, et al.15,.
Experimental autoimmune uveoretinitis (EAU)
Induction of EAU was done by immunizing mice as previously described17. Briefly, complete Freund’s adjuvant (CFA) with 5 mg/mL desiccated M. tuberculosis (Difco Laboratories, Detroit, MI) and 2 mg/ml interphotoreceptor retinoid binding protein (residues 1–20) (IRBPp) (Genscript, Piscataway, NJ) was emulsified together. Two 100 µL subcutaneous injections of the emulsion was given in the lower back, then 0.5 µg pertussis toxin was delivered by intraperitoneal injection. Fundus examination every 3–4 days with a slit lamp microscope was done to determine the severity of retinal inflammation over the course of EAU. Prior to retinal examination, dilation of the iris was accomplished with 1% tropicamide, and the cornea was flattened with a glass coverslip. A 5-point scale as previously described18 was used to score the clinical signs of observable infiltration and vasculitis in the retina. The greater score of both eyes was used to represent the clinical score of that mouse for that day, the average score for the group of mice was then calculated for that day. Another masked member of the lab verified the scores of the mice.
Administration of Tamoxifen
Tamoxifen (Sigma) was dissolved in corn oil a concentration of 20 mg/mL and administered to mice by oral gavage every other day for a total of three doses.
In vitro stimulation
The mouse spleen was collected in 5% FBS in RPMI-1640 (RPMI) supplemented with 10 µg/ml gentamycin (Sigma), 10 mM HEPES, 1 mM sodium pyruvate (BioWhittaker), non-essential amino acids 0.2% (BioWhittaker). The spleen was then made into a single cell suspension that was depleted of red blood cells using RBC lysis buffer (Sigma). Serum free media (SFM) that consisted of RPMI with 0.2% ITS + 1 solution (Sigma) and 0.1% BSA (Sigma) was used to resuspend the RBC depleted splenocytes and IRBPp was added at 50 µg/mL for 48 h at 37oC and 5% CO2 to reactivate antigen specific T cells. Following the reactivation cells were collected for adoptive transfer into recipient mice or stained for flow cytometry analysis.
In vivo imaging
Imaging of mouse retinas was done using a Micron IV microscope imaging system (Phoenix Technologies, Milpitas, CA) containing filters for green and red fluorescence. Mice were anesthetized and the eye was dilated with 1% tropicamide, placed into a holder, and the optic nerve was used as a reference point to obtain images of the retina. The GFP fluorescence was quantified using Imaris Software (Oxford Instruments, Abingdon, United Kingdom).
Flow cytometry
Staining of mouse cells for flow cytometry analysis was done after cells were collected and washed with PBS with 1% BSA (staining buffer), and then Fc blocked with mouse IgG. Conjugated antibodies used were anti-CD4 (clone RM4-5, Biolegend, San Diego, CA), anti-CD25 (clone PC61, Biolegend), anti-PD-1 (clone 29 F.1 A12, Biolegend), anti-TIGIT (clone 1G9, Biolegend), anti-CTLA-4 (UC10-4B9, Biolegend), and anti-FoxP3 (clone FJK-16 s, Invitrogen). Mouse spleen cells were washed with staining buffer, blocked with mouse IgG in staining buffer, then stained with antibodies. Intracellular staining for FoxP3 was done using the FoxP3 transcription factor staining kit (Invitrogen), and intracellular staining for CTLA-4 was done using intracellular staining permeabilization buffer (Biolegend).
Antibody stained cells were analyzed in the University of Massachusetts Chan Medical School Flow Cytometry Core Facility. Full spectrum flow cytometry data was unmixed with SpectroFlo software (Cytek) and data was analyzed using FlowJo Software (Tree Star, Inc., Ashland, OR) or FCS Express (DeNovo Software, Pasadena, CA).
Cell sorting
Cell sorting was done by magnetic sorting using the EasySep Mouse CD4 + Tcell Isolation kit (STEMCELL Technologies, Kent, WA) or CD4 + CD25 + Treg Iso Kit II (STEMCELL Technologies). Cells for adoptive transfer were washed with sterile PBS, resuspended to 1 × 105 in 50 µL, and then injected by tail vein into recipient mice.
Statistics
Two-way ANOVA was also used to assess significant changes in the tempo of disease between the groups of treated EAU mice. Area under the curve (AUC) was calculated for each mouse to obtain an individual AUC, and all individual AUC values were pooled for each cohort. Statistical significance between experimental groups for the maximum EAU scores and AUC was determined using nonparametric Mann-Whitney U test between groups of mice. Statistical significance was determined when P ≤ 0.05 (two-sided). All statistical tests were conducted with Graphpad Prism software.
Study approval
Approval of animal studies was obtained by the University of Massachusetts Medical School IACUC approval was obtained before all relevant experiments involving mice were done.
Results
CTLA-4 is expressed on multiple post-EAU Treg subsets
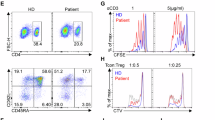
We previously identified multiple post-EAU Treg subsets based on PD-1 and TIGIT expression8,9, but did not determine how these PD-1+ or TIGIT+ Tregs mediate suppression. We therefore asked if CTLA-4 is expressed on either of these Treg subsets and capable of driving functional suppression by Tregs. EAU mice were monitored until disease resolved and the spleen cells were isolated to determine the expression of CTLA-4 on each post-EAU Treg subset. Flow cytometry analysis revealed similar expression of CTLA-4 on each post-EAU Treg subset (Fig. 1A and B). These observations suggest that CTLA-4 may be utilized by multiple post-EAU Treg subsets to suppress EAU.

CTLA-4 is expressed on multiple post-EAU Treg subsets. C57BL/6 J mice were immunized for EAU and monitored by slit lamp examination until resolution. At the resolution of disease, spleen cells were collected and reactivated in vitro with IRBPp, then stained for flow cytometry analysis. Representative density plots showing CTLA-4 and TIGIT expression (A) or CTLA-4 and PD-1 expression (B) of cells gated on FoxP3±CD4+ is shown. Bar graphs show the mean ± SEM of the percentage of cells that are CTLA-4+TIGIT+ or CTLA-4+PD-1+ of the parent FoxP3+ or FoxP3− group. Each experiment consisted of 4–5 mice, each experiment was repeated four separate times, and the mice from all experiments are pooled. Significance was assessed by nonparametric Mann-Whitney U test. Statistical significance (P ≤ 0.001) is designated by ****.
CTLA-4 is needed for resolution of EAU
Based on our observation that CTLA-4 is expressed on post-EAU Tregs, that Tregs are needed for resolution of EAU11, and that CTLA-4 is needed for EAE15, we asked if CTLA-4 has a role in EAU. Conditional Ctla4fl/fl mice were provided by the Sharpe Lab as previously described15. We crossed the Ctla4fl/fl mice with Cd4Cre/ERT2+ mice to study the role of CTLA-4 on CD4+ T cells. The resulting Cd4Cre/ERT2+Ctla4fl/fl mice were given tamoxifen one week before immunization for EAU and the retina was monitored for ocular inflammation to determine the impact of CTLA-4 deficiency on the course of ocular inflammatory disease. Tamoxifen administration resulted in greater than two-fold reduction in CTLA-4 expression was observed in CD4 + cells, and greater than five-fold reduction in FoxP3 + CD4 + cells (Supp Fig. 1). CTLA-4 sufficient mice had a typical disease course compared to wild-type (WT) mice (Fig. 2A). In contrast, a delayed onset and a delayed resolution was observed in CTLA-4 deficient mice (Fig. 2B, Supp Fig. 3), as well as a significant reduction in severity as demonstrated by the max EAU scores (Fig. 2C). These observations show CTLA-4 on CD4 T cells is needed for the onset and severity of uveitis. More importantly, CTLA-4 expressing CD4 T cells are needed for resolution of uveitis.

A CTLA-4 deficiency drives a less severe and long lasting disease. C57BL/6 J (WT), Cd4Cre/ERT+; Ctla4fl/fl, and Cd4Cre/ERT−; Ctla4fl/fl mice were given tamoxifen one week before EAU immunization and monitored by slit lamp examination until resolution. The mean ± SEM EAU scores of mice at the indicated time point of EAU induction is shown for WT (n = 15) with Cd4Cre/ERT−; Ctla4fl/fl (n = 8, Cre-, A) or Cd4Cre/ERT+; Ctla4fl/fl (n = 10, Cre+, B). The images under the graph correspond to the CTLA-4 sufficient or deficient mouse. Area under the curve (AUC) was also calculated for each mouse and the bar graph shows the mean ± SEM for each group. The maximum score for each mouse over the course of disease was identified to determine the severity (Max Score) with the bar graph showing the mean ± SEM for each group. Each experiment consisted of 4–8 mice, each experiment was repeated two separate times, and the mice from all experiments are pooled. Significance of EAU scores for each day was determined by two-way ANOVA with Bonferroni post-test. Significance of AUC and Max Scores was determined by nonparametric Mann-Whitney U test. Statistical significance (P ≤ 0.05) is designated by *, (P ≤ 0.01) is designated by **, and no significance is designated by “ns”.

CTLA-4 is needed for post-EAU Tregs to suppress disease. Spleen cells of post-EAU Cd4Cre/ERT+; Ctla4fl/fl, and Cd4Cre/ERT−; Ctla4fl/fl mice given tamoxifen one week before spleen cells were collected were reactived in vitro with IRBPp, sorted for CD4 + CD25 + cells and transferred to recipient mice by tail vein injection at 1 × 105. The recipient mice were immunized for EAU and the mean ± SEM EAU scores of recipient mice at the indicated time point of EAU induction is shown for EAU mice that did not receive a cell transfer (no AT, n = 18) with mice that received cells from Cd4Cre/ERT−; Ctla4fl/fl mice (AT Cre-, n = 14, A) or mice that received cells from Cd4Cre/ERT+; Ctla4fl/fl mice (AT Cre+, n = 26, B). On day 35 brightfield (BF) imaging and optical coherence tomography (OCT) was done, the line in the BF indicates the OCT laser location. The images under the graph correspond to the mouse that received CTLA-4 sufficient or deficient T cells. Area under the curve (AUC) was also calculated for each mouse and the bar graph shows the mean ± SEM for each group. The maximum score for each mouse over the course of disease was identified to determine the severity (Max Score) with the bar graph showing the mean ± SEM for each group (C). Each experiment consisted of 4–9 mice, and each experiment was repeated 3–4 separate times, and data shown are pooled from all experiments. Significance was assessed by two-way ANOVA with Bonferroni post-test for EAU scores over the course of disease or by nonparametric Mann-Whitney U test for maximum severity scores. Significance (P ≤ 0.05) is designated by *, or (P ≤ 0 0.01) is designated by **, and no significance is designated by “ns”.
Systemic post-EAU Tregs require CTLA-4 to suppress EAU
Based on our observations that post-EAU Tregs that suppress EAU express CTLA-4 (Fig. 1)8, we asked if CTLA-4 is needed for post-EAU Tregs to suppress EAU. One week before post-EAU Tregs were collected from the spleen, Cd4Cre/ERT2+Ctla4fl/fl donor mice were given tamoxifen to delete Ctla4 in CD4 + T cells before adoptive transfer into recipient EAU mice. The unsorted reactivated cells had less than 3% CD4+CD25+ cells that were greater than 50% FoxP3+ (Supp Fig. 2). Post-sort analysis of CD4+CD25+ cells revealed > 98% purity of CD4+ cells, and > 80% CD25+ cells (Supp Fig. 2). Mice that received post-EAU CTLA-4 sufficient Tregs had a delayed onset and accelerated resolution of disease compared to mice that did not receive any cells (Fig. 3A). In contrast, mice that received post-EAU CTLA-4 deficient T cells had a similar onset of disease and delayed resolution compared to mice that did not receive a cell transfer (Fig. 3B, Supp Fig. 3). The maximum EAU score for each mouse showed CTLA-4 sufficient Tregs were able to reduce the severity of disease compared to mice that received CTLA-4 deficient T cells or mice that received no cells (Fig. 3C). These observations show that post-EAU Tregs from the spleen require CTLA-4 to suppress disease and also delays the tempo of resolution when CTLA-4 is absent.
Post-EAU ocular Tregs do not require CTLA-4 to suppress EAU
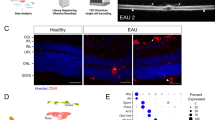
We have previously observed that Tregs migrate into the eye at the onset of disease and persist through resolution9. Therefore, we asked if CTLA-4 is expressed on Tregs in the eye that accumulate in the eye at resolution of disease. We observed CTLA-4 expression on FoxP3+ Tregs in the eye (Fig. 4A and B) and modest CTLA-4 expression was observed on TIGIT+ Tregs (Fig. 4A) and PD-1+ Tregs (Fig. 4B). These observations that post-EAU ocular Tregs express CTLA-4 prompted us to ask if ocular Tregs have the capacity to suppress EAU.

Ocular post-EAU Tregs do not require CTLA-4 to suppress EAU. At the resolution of EAU eyes from WT mice were collected, the retina was dissected out, and cells were immediately stained for flow cytometry analysis. Representative density plots showing CTLA-4 and TIGIT expression (A) or CTLA-4 and PD-1 expression (B) of cells gated on FoxP3±CD4+ is shown. Bar graphs show the mean ± SEM of the percentage of cells that are CTLA-4+TIGIT+ or CTLA-4+PD-1+ of the parent FoxP3+ or FoxP3− group. Post-EAU Cd4Cre/ERT+; Ctla4fl/fl mice and post-EAU Cd4Cre/ERT−; Ctla4fl/fl mice were given tamoxifen one week before eyes were collected, sorted for CD4 + cells and transferred to recipient mice by tail vein injection. The recipient mice were immunized for EAU and the mean ± SEM EAU scores of recipient mice at the indicated time point of EAU induction is shown for EAU mice that did not receive a cell transfer (no AT, n = 12) with mice that received cells from Cd4Cre/ERT−; Ctla4fl/fl mice (AT Cre-, n = 19, C) or mice that received cells from Cd4Cre/ERT+; Ctla4fl/fl mice (AT Cre+, n = 8, D). On day 24–32 brightfield (BF) imaging was done. The images to the right of the graph correspond to the mouse that received CTLA-4 sufficient or deficient T cells. Area under the curve (AUC) was also calculated for each mouse and the bar graph shows the mean ± SEM for each group. The maximum score for each mouse over the course of disease was identified to determine the severity (Max Score) with the bar graph showing the mean ± SEM for each group (E). Each experiment consisted of 4–9 mice, and each experiment was repeated 2–3 separate times, and data shown are pooled from all experiments. Significance was assessed by two-way ANOVA with Bonferroni post-test for EAU scores over the course of disease or by nonparametric Mann-Whitney U test for maximum severity scores. Significance (P ≤ 0.05) is designated by *, or (P ≤ 0 0.01) is designated by **, or (P ≤ 0.001) is designated by ****, and no significance is designated by “ns”.
Cd4Cre/ERT2+Ctla4fl/fl and Cd4Cre/ERT2− Ctla4fl/fl mice immunized for EAU were monitored until resolution. Tamoxifen was administered one week before eyes were collected and CD4+ T cells were collected from the retina and transferred systemically to recipient mice immunized for EAU. Mice that received CTLA-4 sufficient post-EAU ocular CD4 + T cells showed significantly suppressed disease compared to mice that did not receive any cells (Fig. 4C). We also observed a significant suppression of disease in mice that received CTLA-4 deficient post-EAU ocular T cells (Fig. 4D, Supp Fig. 3). The severity of disease was also significantly reduced in both recipient cohorts compared to control mice that did not receive an adoptive transfer (Fig. 4E). These observations show systemically delivered post-EAU ocular CD4 + T cells have the capacity to suppress EAU independently of CTLA-4 expression.
Discussion
Utilizing a CD4-specific deletion of CTLA-4, we determined the impact of a CTLA-4 deficiency on CD4+ T cells on EAU. We observed a delay in resolution of EAU in mice with a CD4+ T cell deficiency for CTLA-4. We further observed that a deletion of CTLA-4 on CD4+CD25+ post-EAU Treg cells prevented suppression of disease when transferred into recipient EAU mice. Since Tregs emerge in the eye over the course of EAU and persist through resolution8,9, we asked if these post-EAU ocular Tregs could suppress disease when transferred systemically to recipient EAU mice. While suppression of disease in mice that received post-EAU ocular Tregs was observed, the suppression of disease severity was not dependent on expression of CTLA-4. Since the expression of CTLA-4 was lower on T cells from the eye compared to the spleen at resolution, this may reflect the reduced dependency on CTLA-4 by ocular T cells to suppress disease severity.
It has been previously observed by Klocke, et al., and Paterson, et al., that CTLA-4 deficient mice were resistant to EAE15,16. In contrast, we observed a slight delay in onset of disease with a slight delay in resolution. In our study, we focused on a CD4-specific deletion of CTLA-4, whereas Paterson, et al. utilized a global tamoxifen induced Cre, so CTLA-4 was removed from both CD4+ and CD8+ T cells, but also found resistance to EAE when CTLA-4 was deleted in FoxP3-expressing cells15. Paterson, et al. also found an increase in the regulatory molecules, IL-10, PD-1, and LAG3 on CD4+ T cells, suggesting a compensatory mechanism15. Since we observed a dependency on CTLA-4 post-EAU Tregs from the spleen to suppress EAU, the lack of EAU resistance with a CTLA-4 deficiency could be due to dysfunctional Tregs. Paterson, et al. also found differences in antigen presenting cells that were tissue specific15. The Klocke, et al. demonstration that CTLA-4 deficient mice exhibit systemic autoimmune disease and more severe CIA16 further supports a tissue specific role for CTLA-4. Therefore, it is possible our EAU findings suggest a different role for CTLA-4 in the eye compared to the brain.
It has been previously observed that CTLA-4-Ig, a fusion of the extracellular domain of CTLA-4 with Ig, was effective in suppressing induction of EAU through blockade of T cell activation, by Iwahashi, et al.19. One interpretation of our observation that resolution of EAU is extended in CTLA-4 deficient CD4+ T cells is that newly activated uveitic T cells are not suppressed by ocular antigen specific Tregs. Egwaugu, et al., first published that whole IRBP drives a chronic EAU model20, suggesting that activation of different T cell clones against multiple epitopes of the same retinal protein could drive a much longer lasting uveitis model. Epitope spreading has been described in mouse, rats, and horse uveitis20,21,22,23 and is thought to contribute to the chronic nature of uveitis in the clinic24, therefore it is possible that CTLA-4 may be employed by Tregs to limit uveitis by preventing T cell activation to new ocular epitopes. Another possible explanation is that the antigen resides in at the injection site, so extension of the resolution phase may be due to additional antigen being released and the CTLA-4 deficiency prevents Tregs from suppressing this new release of antigen.
There have been indirect reports that CTLA-4 has a role on uveitis in the clinic. For example, it has been reported that oncology patients being treated with a CTLA-4 inhibitor may develop a novel uveitis as anterior, posterior, intermediate, panuveitis, and optic neuritis25,26,27,28. The emergence and persistence of Tregs in the eye during EAU suggests they may remain to prevent relapse. It has been shown that Tregs are needed for resolution of EAU. Since resolution of EAU can occur without emergence of post-EAU Tregs that prevent relapse, this suggests that there are multiple Treg subsets with distinct functions. Our observation that post-EAU ocular T cells do not require CTLA-4 to control the disease severity but post-EAU spleen T cells do require CTLA-4 demonstrates that within the post-EAU Treg subset there are additional subsets that suppress through different mechanisms. It should be noted that the transfer of ocular T cells contained conventional T cells that also expressed CTLA-4, so the observed suppression could also be due to CTLA-4 expressing conventional T cells. Given how heterogeneous uveitis is from a clinical perspective, these multiple mechanisms and subsets may explain why some uveitis patients fail multiple therapies, while others go into remission with one type of therapy. Fortunately, the multiple mechanisms and multiple subsets of Tregs suggests there are multiple paths to remission for the uveitis patient.
Research standards
All animal studies adhere to the editorial policies involving animals, with all work done in adherence to the NIH Guide for the Care and Use of Laboratory Animals.
Data availability
“The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.”
References
Darrell, R. W., Wagener, H. P. & Kurland, L. T. Epidemiology of uveitis. Incidence and prevalence in a small urban community. Arch. Ophthalmol. 68, 502–514 (1962).
Hwang, D. K., Chou, Y. J., Pu, C. Y. & Chou, P. Epidemiology of uveitis among the Chinese population in Taiwan: a population-based study. Ophthalmology 119 (11), 2371–2376 (2012).
Suhler, E. B., Lloyd, M. J., Choi, D., Rosenbaum, J. T. & Austin, D. F. Incidence and prevalence of uveitis in veterans affairs medical centers of the Pacific Northwest. Am. J. Ophthalmol. 146 (6), 890–896e898 (2008).
Caspi, R. R. Animal models of autoimmune and immune-mediated uveitis. Drug Discovery Today: Disease Models. 3 (1), 3–9 (2006).
Gilbert, R. M. et al. Clinical remission of Sight-Threatening Non-Infectious uveitis is characterized by an upregulation of peripheral T-Regulatory cell polarized towards T-bet and TIGIT. Front. Immunol. 9, 907 (2018).
Lee, D. J. The relationship between TIGIT + regulatory T cells and autoimmune disease. Int. Immunopharmacol. 83, 1–11 (2020).
Lee, D. J., Preble, J., Lee, S., Foster, C. S. & Taylor, A. W. MC5r and A2Ar deficiencies during experimental autoimmune uveitis identifies distinct T cell polarization programs and a biphasic regulatory response. Sci. Rep. 6, 37790 (2016).
Muhammad, F. et al. TIGIT(+) A2Ar-Dependent anti-uveitic Treg cells are a novel subset of Tregs associated with resolution of autoimmune uveitis. J. Autoimmun. 111, 102441 (2020).
Peters, K. et al. A2Ar-dependent PD-1 + and TIGIT + Treg cells have distinct homing requirements to suppress autoimmune uveitis in mice. Mucosal. Immunol. 16, 422–431 (2023).
Zhou, R. et al. The living eye disarms uncommitted autoreactive T cells by converting them to Foxp3(+) regulatory cells following local antigen recognition. J. Immunol. 188 (4), 1742–1750 (2012).
Silver, P. B. et al. Retina-specific T regulatory cells bring about resolution and maintain remission of autoimmune uveitis. J. Immunol. 194 (7), 3011–3019 (2015).
Tivol, E. A. et al. Loss of CTLA-4 leads to massive lymphoproliferation and fatal multiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4. Immunity 3 (5), 541–547 (1995).
Waterhouse, P. et al. Lymphoproliferative disorders with early lethality in mice deficient in Ctla-4. Science 270 (5238), 985–988 (1995).
Wing, K. et al. CTLA-4 control over Foxp3 + regulatory T cell function. Science 322 (5899), 271–275 (2008).
Paterson, A. M. et al. Deletion of CTLA-4 on regulatory T cells during adulthood leads to resistance to autoimmunity. J. Exp. Med. 212 (10), 1603–1621 (2015).
Klocke, K., Sakaguchi, S., Holmdahl, R. & Wing, K. Induction of autoimmune disease by deletion of CTLA-4 in mice in adulthood. Proc. Natl. Acad. Sci. U S A. 113 (17), E2383–2392 (2016).
Lee, D. J. & Taylor, A. W. Recovery from experimental autoimmune uveitis promotes induction of antiuveitic inducible Tregs. J. Leukoc. Biol. 97 (6), 1101–1109 (2015).
Namba, K., Kitaichi, N., Nishida, T. & Taylor, A. W. Induction of regulatory T cells by the Immunomodulating cytokines alpha-melanocyte-stimulating hormone and transforming growth factor-beta2. J. Leukoc. Biol. 72 (5), 946–952 (2002).
Iwahashi, C. et al. CTLA4-Ig suppresses development of experimental autoimmune uveitis in the induction and effector phases: comparison with Blockade of interleukin-6. Exp. Eye Res. 140, 53–64 (2015).
Oh, H. M. et al. Autoreactive memory CD4 + T lymphocytes that mediate chronic uveitis reside in the bone marrow through STAT3-dependent mechanisms. J. Immunol. 187 (6), 3338–3346 (2011).
Boldison, J. et al. A novel pathogenic RBP-3 peptide reveals epitope spreading in persistent experimental autoimmune uveoretinitis. Immunology 146 (2), 301–311 (2015).
Deeg, C. A. et al. Uveitis in horses induced by interphotoreceptor retinoid-binding protein is similar to the spontaneous disease. Eur. J. Immunol. 32 (9), 2598–2606 (2002).
Diedrichs-Mohring, M., Hoffmann, C. & Wildner, G. Antigen-dependent monophasic or recurrent autoimmune uveitis in rats. Int. Immunol. 20 (3), 365–374 (2008).
Caspi, R. R. Mechanisms underlying autoimmune uveitis. Drug Discovery Today: Disease Mech. 3 (2), 199–206 (2006).
Sun Mm Md, P. et al. Uveitis in patients treated with CTLA-4 and PD-1 checkpoint Blockade Inhibition. Ocul Immunol. Inflamm. 28 (2), 217–227 (2020).
Eikenberry, J. et al. Ocular side effects of target therapy and immunotherapy in patients with cutaneous malignant melanoma. Eur. J. Ophthalmol. 31 (3), 1391–1398 (2021).
Kuo, H. T. et al. Association between immune checkpoint inhibitor medication and uveitis: a population-based cohort study utilizing TriNetX database. Front. Immunol. 14, 1302293 (2023).
Zhang, H., Houadj, L., Wu, K. Y. & Tran, S. D. Diagnosing and managing uveitis associated with immune checkpoint inhibitors: A review. Diagnostics (Basel) 14(3), 1–48 (2024).
Acknowledgements
We would like to thank Arlene Sharpe for graciously providing the Ctla4fl/fl mice used in this study. We would like to thank the UMass Chan Flow Core Facility for their expertise, instrumentation was purchased with the NIH S10OD028576 grant. This work was supported by National Institutes of Health/National Eye Institute grants EY024951 (DJL), and EY029240 (DJL).
Author information
Authors and Affiliations
Contributions
All experimental data curation-formal analysis and investigation of this work was done by DJL, MJA, PH, and JD. The conceptualization, project administration/oversight of this work and review, editing, and revision of this manuscript was done by DJL. The writing of this manuscript was a collaborative effort between DJL and MJA.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflict of interest
“The authors declare that they have no competing interests.”
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ahn, M., Dostal, J., Hegde, P. et al. CTLA-4 expression on tregs is needed for suppression of autoimmune uveitis. Sci Rep 15, 17745 (2025). https://doi.org/10.1038/s41598-025-02816-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-02816-z
