
Abstract
Senecio vulgaris L. (Asteraceae), a European-origin invasive plant, has established widespread populations in China since the nineteenth century, posing both ecological and health-related threats due to its herbicide resistance, toxic pyrrolizidine alkaloids, and broad habitat adaptability. Despite being classified as a Level 4 invasive species in China, its potential risk remains underestimated. In this study, we applied an optimized MaxEnt model using occurrence data from both native (Europe) and invasive (China) ranges to predict the species’ potential distribution. Our results revealed high environmental suitability in central, eastern, southwestern, and northeastern China, with human activity, temperature, and precipitation as dominant predictors. Niche analysis indicated that S. vulgaris in China occupies environmental conditions largely overlapping with its native niche, demonstrating niche conservatism rather than expansion. These findings suggest that even moderately ranked invasive species can maintain ecological stability while gradually expanding, especially under increasing anthropogenic pressure. Our results underscore the need for targeted monitoring and preemptive management in agriculturally sensitive regions. This study provides a practical framework for assessing and managing adaptable invasive plants beyond those classified as high risk.
Similar content being viewed by others

Introduction
Senecio vulgaris L. (commonly known as common groundsel) is a member of the family Asteraceae, originally native to parts of Europe and Asia. Due to its robust reproductive capacity and environmental adaptability, it has become a globally distributed invasive species. S. vulgaris is commonly found in open hillsides, grasslands, and along roadsides1. The species disperses its seeds via wind, enabling rapid colonization of open spaces, farmlands, and other disturbed habitats, demonstrating high environmental tolerance and ecological plasticity2. In addition to threatening native plant communities, S. vulgaris exacerbates weed competition in agricultural fields, thereby impacting crop production3. With the acceleration of globalization, this species has spread to numerous countries and regions, emerging as a prominent example of ecological invasion4,5. Although S. vulgaris is currently classified as a Grade IV invasive species in China, it possesses several ecological traits that justify scientific attention40. This species exhibits a rapid life cycle, prolific seed production, and strong environmental plasticity, which enable it to establish and spread under a wide range of climatic conditions5. Globally, it has invaded numerous temperate and subtropical regions, suggesting a high potential for expansion, particularly under ongoing climate change scenarios40. Recent distribution data indicate that its suitable habitat is increasing in eastern and northeastern China, highlighting a trend of gradual expansion6. Moreover, in contrast to well-documented aggressive invaders such as Spartina alterniflora7 or Lolium temulentum8, the ecological niche dynamics and climatic responses of moderate-risk species like S. vulgaris remain underexplored. Studying such species offers valuable insights into the early stages of invasion processes and helps develop tiered management strategies.
Species Distribution Models (SDMs) and Ecological Niche Models (ENMs) are widely used predictive tools in ecology. These models integrate species distribution data with environmental variables to estimate potential suitable habitats within a given geographic region9,10,11. In recent years, SDMs and ENMs have been extensively applied in predicting and managing the distribution of invasive species, aiding ecologists in assessing invasion risks and habitat occupancy12,13. Among these models, the MaxEnt model has gained significant popularity for its excellent performance with small sample datasets14. By analyzing the contributions of environmental variables to species distributions, MaxEnt effectively predicts potential distribution areas and has proven particularly valuable in managing invasive species15. The integration of SDMs and ENMs also facilitates the study of ecological niche dynamics in invasive species, including niche stability during invasion processes and niche expansion in response to novel climatic conditions. These approaches offer powerful tools for unraveling the mechanisms underlying species invasions9,10,11,12,13,14,15,16. For instance, Xian et al.17 employed the Biomod2 model in combination with the ecospat package to predict the global distribution shifts of three invasive Ambrosia species and compared their native and invaded range niches. Their findings revealed that the ecological niches of these species had undergone changes in the invaded regions, indicating further invasion risks and a poleward shift of potential distribution centers under climate change. Similarly, Wan et al.18 used the MaxEnt model alongside the ecospat package to simulate the climatic niches of eight representative invasive weeds from the Americas in both their native and invaded ranges. Modeling results under climate change scenarios showed significant niche differences between the Americas (native range) and China (invaded range), with broader niche widths observed in the invaded range, suggesting a high likelihood of continued expansion into high-altitude and high-latitude areas in China.
This study aims to analyze the distribution patterns and niche dynamics of S. vulgaris in China using Species Distribution Models (SDMs) and Ecological Niche Models (ENMs). The specific objectives are as follows: (1) to predict the potential distribution patterns of S. vulgaris in China using the MaxEnt model, thereby assessing its dispersal capacity; (2) to investigate whether S. vulgaris exhibits niche conservatism or expansion during its invasion of China by employing ecological niche models; and (3) to provide theoretical support for the management of alien invasive plants in China by systematically analyzing the potential distribution and niche changes of S. vulgaris.
Method and materials
The process of occurrence of S. vulgaris
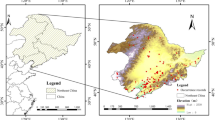
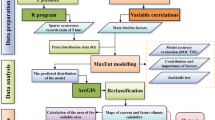
To ensure accuracy in species distribution modeling, comprehensive and reliable occurrence data were compiled from both the native and invasive ranges of S. vulgaris. For the native range (primarily Europe, latitude: − 10° to 40° N; longitude: 35° to 70° W), we retrieved 10,000 initial records using the rgbif package in R from the Global Biodiversity Information Facility (GBIF)19. Additional records were supplemented from databases such as iPlant (species distribution images), the Global Invasive Species Database (GISD, https://www.iucngisd.org/), and the National Species Observation Platform (http://sso.especies.cn/). After removing duplicates and correcting coordinates using Google Earth (standardized to the WGS84 system), a total of 2380 valid occurrence points were retained for the native range. For the invasive range in China, we collected 366 records from multiple sources, including the Chinese Virtual Herbarium (CVH), literature sources, and online biodiversity platforms. After spatial filtering to reduce autocorrelation, 247 valid occurrence points were retained. To minimize spatial bias in both datasets, we applied a thinning process using the spThin package in R and the ENMtools toolkit, ensuring a minimum distance between retained points. we first applied spThin20 to reduce sampling bias by enforcing a minimum distance of 10 km between occurrence records. The function thin() was run with 100 replicates per dataset to ensure stability, and the replicate with the highest retained points was selected. Next, we used ENMtools21 to cross-validate and confirm spatial independence between points. ENMtools performs additional checks by computing pairwise distances and generating spatially independent datasets through random subsampling. This step helps reduce potential pseudoreplication and ensures that models are not overly influenced by spatially clustered data. These curated datasets formed the basis for subsequent species distribution and niche modeling. (Supplementary Table S5 and Fig. 1).

The distribution map of S. vulgaris within its native (blue, 2380) and invasive area (red, 247).
Acquisition and processing of environmental variables
In this study, environmental variables were classified into three categories: bio-climate, topography, and human activity factors. Bio-climatic variables, particularly those associated with temperature and precipitation, are widely recognized as the primary determinants of plant distribution and spread. A total of 19 bio-climatic variables were selected from the Bioclim dataset, sourced from the WorldClim database (version 2.1), with a spatial resolution of 10 arc-minutes (approximately 18.5 km). Topographic variables included elevation data (Digital Elevation Model, DEM), which were further derived into slope and aspect using GIS tools. Human activity factors were represented by the Human Influence Index (HII), obtained from the Center for International Earth Science Information Network (CIESIN), Columbia University. The HII integrates factors such as population density, land use and cover, transportation networks, and light pollution22. All environmental variables were resampled to a consistent 10 arc-minute resolution in GIS, and the coordinate system was set to WGS84 for compatibility with species occurrence data, ensuring consistency in subsequent analyses. A detailed summary of these variables is presented in Supplementary Table S1.
To perform the predictive analysis, all environmental variables, along with valid species occurrence points, were imported into the MaxEnt model. The model was run 10 times, and variables with a contribution of 0% were excluded. The remaining variables were subjected to Pearson correlation analysis to identify highly correlated factors (|r|> 0.8). Only the variables with higher contributions and lower correlation coefficients were retained23. Ultimately, the following 10 environmental variables were selected for further analysis: annual mean temperature, mean diurnal temperature range, temperature seasonality, annual temperature range, precipitation in the coldest quarter, precipitation in the driest quarter, elevation, slope, aspect, and the Human Influence Index (Supplementary Table S2 and Fig. S1).
Parameter optimization and construction of MaxEnt model based on ENMeval package
The MaxEnt model is widely used for predicting species’ potential distributions due to its accuracy and robustness. However, the performance of the MaxEnt model largely depends on the optimization of model parameters, particularly the choice of regularization multiplier (RM) and feature combinations (FC). In this study, the ENMeval package in R was used to establish six feature combinations (FCs), representing different combinations of mathematical functions in MaxEnt: H (Hinge), L (Linear), LQ (Linear + Quadratic), LQH (Linear + Quadratic + Hinge), LQHP (Linear + Quadratic + Hinge + Product), and LQHPT (Linear + Quadratic + Hinge + Product + Threshold). The regularization multiplier (RM) ranged from 0 to 4 with a gradient of 124. Based on existing literature, the parameter set with the minimum AICc value (Delta_AICc = 0) was selected to build the final model.
The optimal set of parameters derived from ENMeval was then applied to the MaxEnt model. The model was trained with 75% of the data as the training set and tested with 25% of the data. A maximum of 10,000 background points was used, and the model was run 10 times. The final prediction results were based on the average of the 10 runs, with the output provided as an ASCII file. The importance of each environmental variable was assessed based on its contribution to the MaxEnt model and the average score from the Jackknife test of regularized training gain. Additionally, response curves for the most influential environmental variables were plotted to examine their effects on species’ suitability probabilities25.
Quantification of ecological niche dynamics of S. vulgaris in its native and invasive ranges using the R package hypervolume
The hypervolume (The hypervolume refers to an n-dimensional space where each dimension represents an environmental variable, and the volume encapsulates the range of conditions suitable for a species’ survival and reproduction) package in R26 was used to quantify the dynamic ecological niches of S. vulgaris in its native range (Europe) and invasive range (China). First, Principal Component Analysis (PCA) was performed to reduce the dimensionality of the environmental variables, projecting the high-dimensional data into a two-dimensional space. The hypervolume_gaussian function was then applied to construct the ecological niche hypervolume for both the native and invasive ranges. Furthermore, many studies suggest that a niche expansion index greater than 10% indicates a shift in the ecological niche26,27. In this study, the Biodiversity Assessment Tools (BAT) package28 was used to calculate the overall similarity index (β_total) between the native and invasive range niches using the kernel.beta function. The β_total index was further decomposed into two potential processes: spatial displacement between hypervolumes (β_replacement), which reflects the degree of niche shift, and richness expansion or contraction (β_richness), which reflects niche expansion or contraction. The β_total index ranges from 0 to 1, with values closer to 0 indicating higher niche similarity, and values closer to 1 indicating lower similarity. The relationship is given by the equation: The total niche dissimilarity index (β_total) is the sum of niche displacement (β_replacement) and niche expansion/contraction (β_richness): β_total = β_replacement + β_richness26,27,28.
Model accuracy and habitat suitability classification based on MaxEnt
The accuracy of the MaxEnt model is judged by the AUC value, which ranges from 0 to 1. If the AUC value is less than 0.5, it is considered a prediction failure. If 0.5 < AUC ≤ 0.6, it is poor; if 0.6 < AUC ≤ 0.7, it is poor; if 0.7 < AUC ≤ 0.8, it is average; if 0.8 < AUC ≤ 0.9, it is good; and if 0.9 < AUC ≤ 1, it is excellent29. The prediction results of the MaxEnt model are raster data between 0 and 1, where the value represents the high or low probability of adaptation. The establishment of the adaptation zone level is based on the natural breakpoint method in GIS tools, which means that the non adaptation zone is 0–0.2, the low adaptation zone is 0.2–0.4, the medium adaptation zone is 0.4–0.6, and the high adaptation zone is 0.6–130.
Research results
Accuracy evaluation of the MaxEnt model
The MaxEnt model, constructed using the optimized parameters (FC = LQ, RM = 1) based on ENMeval package results, demonstrated good accuracy and stability in predicting the potential distribution of S. vulgaris. The AUC value for the model’s prediction was 0.884 in Fig. 2a, indicating high reliability. Furthermore, the difference in AUC values between the training and testing datasets was only 0.001, much smaller than the gap observed under default conditions (Supplementary Table S3)(Fig. 2b), suggesting that the optimized model provides robust and reliable predictions.

(a) The AUC value of MaxEnt model by ggplot2 and (b) Optimized MaxEnt model by ENMeval package.
Influence of environmental variables on the potential distribution of S. vulgaris
According to the Jackknife test of regularized training gain, the environmental variables with the greatest impact on the distribution and spread of S. vulgaris were human activity, annual mean precipitation, temperature diurnal range, and annual mean temperature. The contributions of these variables in the MaxEnt model (Supplementary Table S4) indicated that human activity and annual mean temperature are the dominant factors limiting the potential distribution of S. vulgaris, with a cumulative contribution of 78.5%. Other factors, such as annual temperature range, elevation, temperature seasonality, mean diurnal temperature range, slope, annual precipitation, aspect, and precipitation variability, contributed 7.1%, 4.9%, 4.7%, 2%, 1.4%, 0.7%, 0.4%, and 0.3%, respectively. This analysis suggests that S. vulgaris is primarily influenced by human activity, temperature, and precipitation, while topographic factors have a relatively weaker effect.
The single-factor response curve in Fig. 3 analysis revealed that as human activity intensity increases, the probability of occurrence of S. vulgaris also increases. When the human activity intensity exceeds 23.2, the species’ probability of occurrence is greater than 0.5. At an intensity of 50, the occurrence probability approaches 1, indicating that high human activity levels significantly promote the spread of S. vulgaris. Similarly, the optimal annual mean temperature range for the species was found to be between 1.28 °C and 18.72 °C, with the highest suitability occurring around 10 °C. The optimal annual precipitation range for S. vulgaris was 419.9 mm to 1450.5 mm, with peak suitability occurring at 935.2 mm (Fig. 4).

The simple factor response curve of environmental variable.

The regularized training gain. Note: WV is without variable, WOV is with only variable, and ALL is using all variables.
Ecological niche changes between the native and invasive ranges of S. vulgaris
This study employed multidimensional ecological niche modeling to quantify the dynamic changes in the ecological niche of S. vulgaris across different geographic regions in Fig. 5. The results showed significant differences in the ecological niches of S. vulgaris between its native range (Europe) and invasive range (China), especially in terms of the niche indicators β_total and β_richness, which reflected the species’ adaptability to new environments. Principal Component 1 (PC1) was primarily determined by the temperature diurnal range, accounting for 49% of the variance, while Principal Component 2 (PC2) was determined by temperature seasonality, also accounting for 49%. These results suggest that S. vulgaris is gradually shifting towards areas with a larger temperature diurnal range. In comparison to the native range (HV = 22.89), the ecological niche in China (HV = 94.95) shows an expansion trend. The β_total value of 0.791 indicates relatively low niche overlap between the native and invasive ranges, with an Euclidean distance of 3.77 between the centroids of the two niches. The β_richness value of 0.739 explains 73.9% of the niche differences, suggesting significant niche expansion. In contrast, the β_replacement value of 5.2% indicates that there has been little change in the position of the species’ niche across regions, implying that the species has not significantly migrated to a different environmental space. Instead, S. vulgaris has retained a similar niche position across both regions. Our findings suggest that, although S. vulgaris has adapted to some new environmental conditions during its invasion from Europe to China, it has not migrated to completely different environmental spaces. Rather, it continues to adapt to similar environmental types, maintaining a similar ecological niche in both its native and invasive ranges.

Quantitative analysis of the niche of S. vulgaris in its native range (Europe) and invaded range (China) based on the hypervolume package. Note: The picture above depicted that niche shift of S. vulgaris between native area (Europe) and invaded area (China), and the blue dash line indicate that the environment range space of native area and similarly the red dash indicate that the environment range space of invaded area. The blue and red dots symbolize the species density of S. vulgaris in each range. The black arrow is the direction of niche shift of S. vulgaris from native area to invaded area.
Potential distribution of S. vulgaris in China at present
The potential distribution of S. vulgaris in China during the current period is shown in Fig. 6. The non-suitable areas, low-suitability areas, moderate-suitability areas, and high-suitability areas are represented in white, green, orange, and red, respectively. The overall suitable area is primarily concentrated in central, eastern, southwestern, and northeastern China, with scattered patches in the northwest. The total suitable area covers 2.744 × 104km2, accounting for approximately 28.6% of China’s total land area. The low-suitability area covers 156.64 × 104km2, accounting for approximately 16.3% of China’s total land area, and is primarily distributed around the periphery of the moderate-suitability zone. This includes regions such as Xinjiang, Gansu, Ningxia, southeastern Tibet, eastern Inner Mongolia, northern Shaanxi, northeastern Shandong, southern Liaoning, and parts of Heilongjiang. The moderate-suitability area spans 63.42 × 104km2, or about 6.6% of China’s total area, and is sporadically distributed around the high-suitability zone. Key locations in this category include southern Shaanxi, Henan, Anhui, Jiangsu, southern Shandong, northern Hubei, Beijing, Tianjin, southern Hebei, and central Shandong. The high-suitability area encompasses 54.34 × 104km2, or 5.6% of China’s total land area, and is concentrated primarily in the southwestern and central regions of China. Major regions include Sichuan, Chongqing, Guizhou, northwestern Yunnan, southern Gansu, northern Sichuan, southern Shaanxi, southern Ningxia, eastern Qinghai, and northern Taiwan.

The potential distribution area of S. vulgaris in China at present using Arcmap10.8 software (URL: https:// www. arcgis. com/ index. html).
Discussion
Niche quantification between native and invasive ranges using hypervolume
Previous studies commonly employed the ecospat package developed by Petitpierre et al.31, based on sorting analysis under the COUR framework(COUR) framework, which is widely used in niche conservatism studies33. However, literature suggests that this method might miss key information compared to multidimensional spatial analysis, potentially affecting result accuracy32,33. Therefore, in this study, the hypervolume method proposed by Blonder et al.26 was used to quantify the ecological niches of S. vulgaris in its native (Europe) and invasive (China) ranges. The results indicated that although the niche of S. vulgaris in the invasive range (China) expanded more than in the native range, this expansion was primarily due to the larger ecological volume in the invasive range, rather than a significant niche shift. Despite these changes, the species occupied regions with similar climatic conditions to its native range during its spread.
Potential distribution and influencing factors of S. vulgaris in China
The MaxEnt model predicted that the potential distribution of S. vulgaris in China is primarily concentrated in areas such as Gansu, Ningxia, eastern Inner Mongolia, Heilongjiang, Liaoning, Beijing, Tianjin, Shaanxi, Hebei, Henan, Shandong, Anhui, Jiangsu, Hubei, Sichuan, Chongqing, Guizhou, Yunnan, eastern Qinghai, Xinjiang, southern Tibet, and northern Taiwan. This result is consistent with the findings of Cheng et al.34 and is nearly identical to the collected distribution points, supporting the reliability of the predictions. The primary factors driving the species’ further spread and distribution in China include human activity, temperature, and precipitation, which further corroborates the crucial role of climatic conditions such as temperature and precipitation in shaping plant distribution35. Additionally, with the acceleration of global economic processes, human activity—both intentional and unintentional—has contributed to the introduction of invasive species into vacant ecological niches. Lacking natural enemies, intra- and inter-species competition, and other ecological checks, invasive species can exploit local resources, leading to exponential population growth and posing a significant threat to biodiversity and ecosystem stability36,37. Finally, after the optimized MaxEnt model, the model prediction ability has been largely improved. Zhao et al.38 also used optimized MaxEnt model RM = 0.5; FC = LQ, to predict the potential distribution of China fir. The model had the lowest complexity under this parameter. The average AUC value of 10 times of repeated training set was 0.9101 ± 0.0012 and the average AUC value of the test set was 0.9036 ± 0.0149, which showed that this model was accurate for Chinese fir prediction. In addition, Zhao et al.39 used optimized MaxEnt model with ENMeval to predicted the potential distribution area of Ziziphus jujuba in China. RM = 3.5, FC = LQH, and delta. AICc = 0, according to the AIC, the model is the optimal model.
Recommendations for the control of S. vulgaris
Given the strong influence of climatic factors on the distribution of S. vulgaris, it is essential to develop proactive management tools that can forecast its potential spread under changing environmental conditions. Climate adaptability warning system is a modeling-based tool that can clearly forecasts high-risk invasion zones by combining species distribution data with climate change projections40. While this study focuses on the role of human activity, bioclimatic, and topographical factors in shaping the niche dynamics of S. vulgaris between its native and invasive ranges, and its potential spread within China, it does not address how climate change may further influence its distribution patterns. Additionally, other environmental variables such as solar radiation intensity, normalized difference vegetation index (NDVI), and land use classification have not been incorporated into the analysis41. These will be the focus of future research. Based on the current findings, the following recommendations are made for the early warning and control of S. vulgaris in China.
-
1.
Target High-Risk Areas for Monitoring and Control: Focus on high-risk regions in central, eastern, southwestern, and northeastern China, particularly areas with high human activity. Strengthen monitoring of such areas and adopt a combination of physical, chemical, and biological control methods to suppress the spread of pests and diseases.
-
2.
Establish a Climate Adaptability Warning System: Establish a climate based early warning system for the spread of S. vulgaris, guide ecological restoration in high-risk areas, and improve the resilience of ecosystems through measures such as habitat restoration and strengthening the protection of local species.
-
3.
The potential role of citizen science in detection and monitoring: it could provide cost-effective, scalable surveillance.
-
4.
Improve Legal Framework and Global Cooperation: Strengthen legal regulations by classifying S. vulgaris as a key invasive species to be controlled. Draw on international best practices and enhance control strategies through policy support and global collaboration.
Data availability
All data generated or analysed during this study are included in this published article [and its supplementary information files].
References
Xue-Tian, L., Jing, Z. & Dan-Dan, C. Diversity of endophytic nitrogen-fixing and phosphorus-solubilizing bacteria in the invasive plant Trichoderma europaea. Microbiol. Bull. 50(2), 454–470. https://doi.org/10.13344/j.microbiol.china.220453 (2023).
Jianmeng, F. et al. Risk assessment and spatial distribution pattern of invasive alien plants in China. J. Southwest Univ. Nat. Sci. Ed. 33(2), 57–63. https://doi.org/10.13718/j.cnki.xdzk.2011.02.010 (2011).
Bo, L. I., Bingsheng, X. U. & Jiakuan, C. H. E. N. General characterization of plant invasions from exotic weeds in Shanghai. Biodiversity 9(4), 446. https://doi.org/10.3321/j.issn:1005-0094.2001.04.018 (2001).
Xiaoling, Y. et al. Classification and geographic distribution of invasive alien plants in China. Biodiversity 22(5), 667–676. https://doi.org/10.3724/SP.J.1003.2014.14069 (2014).
Cheng, D., Nguyen, V. T. & Ndihokubwayo, N. Comparison of the performance of native and invasive plants of Senecio vulgairs L. PeerJ Preprints 4, e2012v1. https://doi.org/10.7287/PEERJ.PREPRINTS.2012 (2016).
Kadereit, J. W. The origin of Senecio vulgaris (Asteraceae). Plant Syst. Evol. 145, 135–153. https://doi.org/10.1007/BF00984036 (1984).
Wan, S. et al. The positive and negative effects of exotic Spartina alterniflora in China. Ecol. Eng. 35(4), 444–452. https://doi.org/10.1016/j.ecoleng.2008.05.020 (2009).
Yang, M. et al. Potential global geographical distribution of Lolium temulentum L. under climate change. Front. Plant Sci. 13, 1024635. https://doi.org/10.3389/fpls.2022.1024635 (2022).
Zhongsai, T. et al. A preliminary survey on the invasion status of European Senecio vulgaris L in China. Saf. Environ. Eng. 25(2), 7–14. https://doi.org/10.13578/j.cnki.issn.1671-1556.2018.02.002 (2018).
Melo-Merino, S. M., Reyes-Bonilla, H. & Lira-Noriega, A. Ecological niche models and species distribution models in marine environments: A literature review and spatial analysis of evidence. Ecol. Model. 415, 108837. https://doi.org/10.1016/j.ecolmodel.2019.108837 (2020).
Iannella, M. et al. A step towards SDMs: A “couple-and-weigh” framework based on accessible data for biodiversity conservation and landscape planning. Divers. Distrib. 27(12), 2412–2427. https://doi.org/10.1111/ddi.13410 (2021).
Mendes, P. et al. Dealing with overprediction in species distribution models: How adding distance constraints can improve model accuracy. Ecol. Model. 431, 109180. https://doi.org/10.1016/j.ecolmodel.2020.109180 (2020).
Guisan, A. et al. Predicting species distributions for conservation decisions. Ecol. Lett. 16(12), 1424–1435. https://doi.org/10.1111/ele.12189 (2013).
Thapa, S. et al. Understanding the dynamics in distribution of invasive alien plant species under predicted climate change in Western Himalaya. PLoS ONE 13(4), e0195752. https://doi.org/10.1371/journal.pone.0195752 (2018).
Elith, J. et al. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 17(1), 43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x (2011).
Phillips, S. J. & Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 31(2), 161–175. https://doi.org/10.1111/j.0906-7590.2008.5203.x (2008).
Xian, X. et al. Climate change has increased the global threats posed by three ragweeds (Ambrosia L.) in the anthropocene. Sci. Total Environ. 859, 160252. https://doi.org/10.1016/j.scitotenv.2022.160252 (2023).
Wan, J. Z. et al. Climatic niche divergence and habitat suitability of eight alien invasive weeds in China under climate change. Ecol. Evol. 7(5), 1541–1552. https://doi.org/10.1002/ece3.2684 (2017).
Chamberlain, S. A. & Boettiger, C. R Python, and Ruby clients for GBIF species occurrence data. PeerJ Preprints https://doi.org/10.15468/dl.t3vyoi (2017).
Aiello-Lammens, M. E. et al. SpThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 38(5), 541–545. https://doi.org/10.1111/ecog.01132 (2015).
Warren, D. L., Glor, R. E. & Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 33(3), 607–611. https://doi.org/10.1111/j.1600-0587.2009.06142.x (2010).
Li, S. et al. Mapping human influence intensity in the Tibetan Plateau for conservation of ecological service functions. Ecosyst. Serv. 30, 276–286. https://doi.org/10.1016/j.ecoser.2017.10.003 (2018).
Xu, D. et al. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling. Global Ecol. Conserv. 19, e00691. https://doi.org/10.1016/j.gecco.2019.e00691 (2019).
Muscarella, R. et al. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 5(11), 1198–1205. https://doi.org/10.1016/j.ecoser.2017.10.003 (2014).
Syfert, M. M., Smith, M. J. & Coomes, D. A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 8(2), e55158. https://doi.org/10.1371/journal.pone.0055158 (2013).
Blonder, B. et al. The n-dimensional hypervolume. Glob. Ecol. Biogeogr. 23(5), 595–609. https://doi.org/10.1111/geb.12146 (2014).
Blonder, B. et al. Package ‘hypervolume’. Glob. Ecol. Biogeogr. 23, 595–609. https://doi.org/10.1111/geb.12146 (2015).
Cardoso, P., Rigal, F. & Carvalho, J. C. BAT–biodiversity assessment tools, an R package for the measurement and estimation of alpha and beta taxon, phylogenetic and functional diversity. Methods Ecol. Evol. 6(2), 232–236. https://doi.org/10.1111/2041-210X.12310 (2015).
Peterson, A. T., Papeş, M. & Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 213(1), 63–72. https://doi.org/10.1016/j.ecolmodel.2007.11.008 (2008).
West, A. M. et al. Field validation of an invasive species Maxent model. Eco. Inform. 36, 126–134. https://doi.org/10.1016/j.ecoinf.2016.11.001 (2016).
Di Cola, V. et al. Ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 40(6), 774–787. https://doi.org/10.1111/ecog.02671 (2017).
Mammola, S. & Cardoso, P. Functional diversity metrics using kernel density n-dimensional hypervolumes. Methods Ecol. Evol. 11(8), 986–995. https://doi.org/10.1111/2041-210x.13424.DOI:10.1111/2041-210x.13424 (2020).
Petitpierre, B. et al. Climatic niche shifts are rare among terrestrial plant invaders. Science 335(6074), 1344–1348. https://doi.org/10.1126/science.1215933 (2012).
Cheng, D. & Xu, L. Predicting the potential distributions of Senecio vulgaris L. in China. PeerJ PrePrints 3, e1612v1. https://doi.org/10.7287/peerj.preprints.1612v1 (2015).
Morecroft, M. D. & Paterson, J. S. Effects of temperature and precipitation changes on plant communities. Plant Growth Clim. Change https://doi.org/10.1002/9780470988695 (2006).
Chapman, D. S. et al. Invasion of freshwater ecosystems is promoted by network connectivity to hotspots of human activity. Glob. Ecol. Biogeogr. 29(4), 645–655. https://doi.org/10.1111/geb.13051 (2020).
Leprieur, F. et al. Fish invasions in the world’s river systems: When natural processes are blurred by human activities. PLoS Biol. 6(2), e28. https://doi.org/10.1371/journal.pbio.0060322 (2008).
Zhao, Y. et al. Predicting potential suitable habitats of Chinese fir under current and future climatic scenarios based on Maxent model. Ecol. Inform. 64, 101393. https://doi.org/10.1016/j.ecoinf.2021.101393 (2021).
Zhao, G. et al. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Ind. 132, 108256. https://doi.org/10.1016/j.ecolind.2021.108256 (2021).
Gozlan, R. E. et al. Pan-continental invasion of Pseudorasbora parva: Towards a better understanding of freshwater fish invasions. Fish Fish. 11(4), 315–340. https://doi.org/10.1111/j.1467-2979.2010.00361.x (2010).
Bowen, A. K. M. & Stevens, M. H. H. Temperature, topography, soil characteristics, and NDVI drive habitat preferences of a shade-tolerant invasive grass. Ecol. Evol. 10(19), 10785–10797. https://doi.org/10.1002/ece3.6735 (2020).
Author information
Authors and Affiliations
Contributions
Conceptualization: Z.E.X., W.X.S. Methodology: Z.E.X., W.X.S. Investigation: Z.E.X., W.X.S. Project administration: L.B., H.Y.W.,S.J. Supervision: L.B., H.Y.W.,S.J. Writing—original draf: Z.E.X., W.X.S. Writing—review & editing: Z.E.X., W.X.S., L.B., H.Y.W.,S.J.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, E., Wang, X., Lei, B. et al. The potential distribution area and dynamic niche variation analysis of an alien invasive plant Senecio vulgaris L. Sci Rep 15, 26742 (2025). https://doi.org/10.1038/s41598-025-08144-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-08144-6
