
Abstract
Fibroblast-like synoviocytes (FLSs) are critical for promoting joint and surrounding soft tissue damage in Rheumatoid arthritis (RA). β-Sitosterol has the potential to attenuate RA; however, the underlying mechanism remains largely unknown. This study aimed to investigate the effect of β-Sitosterol on the biological functions of FLSs. FLSs were isolated from the synovial tissues of patients with RA, and cellular behaviors were evaluated using cell counting kit-8, 5-ethynyl-2’-deoxyuridine, scratch test, and enzyme-linked immunosorbent assay. The binding between β-Sitosterol and LDHA was evaluated using molecular docking and surface plasmon resonance. The lactylation of GPI was identified using immunoprecipitation (IP), western blotting, and protein stability assay. The results showed that β-Sitosterol suppressed FLS proliferation, migration, and the levels of IL-1β, IL-6, and TNF-α in a dose-dependent manner. Next, we found that β-Sitosterol bound to LDHA and decreased its protein levels. Moreover, overexpression of LDHA elevated the lactylation levels of GPI and increased GPI protein levels. Knockdown of GPI abrogated the effects on cellular behaviors induced by LDHA. In conclusion, β-Sitosterol inhibits the proliferation, migration, and inflammatory response of FLSs by suppressing LDHA-mediated lactylation of GPI, thereby attenuating RA. These findings provide insights into the molecular mechanisms of β-Sitosterol and suggest β-Sitosterol may be a therapeutic agent for RA.
Similar content being viewed by others
Introduction
Rheumatoid arthritis (RA) is a chronic systemic autoimmune inflammatory disease characterized by synovial hyperplasia and progressive joint destruction, leading to significant morbidity and disability1. Early diagnosis and treatment can enhance clinical outcomes for individuals with RA to a notable degree2. Despite advancements in therapeutic options3, a significant proportion of patients continue to suffer from persistent pain, systemic complications, functional disability, and even death4,5. Therefore, it is necessary to develop new therapeutic agents to treat RA more effectively.
The therapeutic potential of traditional Chinese medicine in managing RA has garnered wide concern6. Si Miao San, a traditional Chinese prescription known for its anti-inflammatory and antioxidant properties, has been used in the treatment of rheumatic diseases, with particular efficacy in alleviating RA symptoms7,8. β-Sitosterol is an active ingredient of Phellodendri Chinensis Cortex (Huangbai), which is a key herb in the Si Miao San formula9. Based on its potential health benefits, β-Sitosterol has demonstrated promising therapeutic potential for various diseases, such as cancer, diabetes, and liver diseases10. Recent studies have revealed that β-Sitosterol possesses a regulatory effect on macrophage polarization, inflammation, and synovial angiogenesis, highlighting its potential as a therapeutic agent in RA11,12. However, the precise mechanisms through which β-Sitosterol exerts its effects are not yet fully elucidated.
The synovium of RA undergoes metabolic reprogramming to a glycolytic phenotype in response to hypoxia and inflammatory stimulation, enabling rapid energy acquisition and further promoting pathogenic growth. Lactylation, a novel lysine modification involving the addition of lactate to lysine residues, has been identified as a significant player in the glycolysis process13. It is known that lactylation is involved in various diseases, especially cancer, but also Alzheimer’s disease, fatty liver disease, etc14. A previous study has reported that artemisinin modulates the proliferation and migration of fibroblast-like synoviocytes (FLSs) by regulating PKM2 lactylation15, suggesting the critical role of lactylation in the pathogenesis of RA. Despite this, how lactylation affects the progression of RA remains not well documented.
Lactate dehydrogenase A (LDHA) is a key glycolytic enzyme converting pyruvate to lactate, which serves as a bridge between glycolysis and lactylation16. It has been reported that LDHA expression was increased in RA-FLSs and drives glycolysis and viability of FLSs17. In addition, inhibiting LDHA controls bone resorption of osteoclasts in RA by reducing glycolysis18. Glycosylphosphatidylinositol (GPI) is an important regulator in glycolysis that can facilitate the conversion between the isomers of glucose 6-phosphate19. It has been revealed that soluble GPI concentration and anti-GPI IgG are elevated in the serum and synovial fluid in patients with RA20. Moreover, GPI can induce RA to establish the animal model21. However, how LDHA and GPI regulate functions of FLSs and whether LDHA can modulate lactylation of GPI in RA pathogenesis remain unexplored.
In this study, we investigated the role of β-Sitosterol in RA. FLSs play a critical role in promoting joint damage and synovial inflammation in RA22. Thus, we used in vitro experiments to identify its regulation on FLS proliferation, migration, and inflammatory responses, and conducted animal experiments to evaluate the role of β-Sitosterol in vivo. We further explored the molecular mechanisms of β-sitosterol, focusing on the involvement of lactylation modification. This study contributes to clarifying the mechanism of β-Sitosterol action and provides theoretical support for its treatment of RA.
Materials and methods
Cell culture
Human FLSs were isolated from the synovial tissues (STs) of patients with RA who underwent synovectomy in our hospital. FLSs were also isolated from patients with osteoarthritis (OA) as the negative control. RA and OA were diagnosed according to the criteria of the 1987 American College of Rheumatology. Written informed consent was provided by each participant. This study was approved by the Ethics Committee of The Second Affiliated Hospital of Guangzhou Medical University.
STs were cut into small pieces (about 1 mm3) and transferred to a T25 culture flask. The Dulbecco’s modified Eagle’s medium/F12 (DMEM/F12) supplemented with 10% fetal bovine serum (FBS) was added into the culture flask to culture the STs at 37 °C with 5% CO2. The culture medium was replaced every 2–4 days. After the FLSs that migrated from the tissue explant grew to more than 90% confluence, the tissues were removed and the cells were passaged. The FLSs at passages 3–5 were used in the following experiments.
Cell counting kit-8 (CCK-8) assay
FLSs were seeded into 96-well plates at the density of 2.5 × 103 cells/well. The cells were treated with 5, 25, and 50 µM β-Sitosterol (MedChemExpress, Monmouth Junction, NJ, USA) for 24 h. The cells treated with dimethyl sulfoxide (DMSO; MedChemExpress) for 24 h were considered the control. Subsequently, cell viability was measured using CCK-8 (Dojindo, Kumamoto, Japan). In brief, FLSs were incubated with 10 µL CCK-8 solution for 2 h at 37 °C. The absorbance was measured at 450 nm using a microplate reader.
5-Ethynyl − 2’- Deoxyuridine (EdU) assay
Cell proliferation was evaluated using the EdU kit (Ribobio, Guangzhou, China). FLSs after β-Sitosterol or DMSO stimulation were incubated with complete culture medium containing 50 µM EdU solution for 2 h. Next, the cells were fixed with 4% paraformaldehyde for 30 min and permeated with 0.5% TritonX-100 for 10 min, followed by incubation with 1× Apollo staining solution protecting from the light at room temperature for 30 min. DNA was stained using 4’,6-diamidino-2-phenylindole (DAPI; MedChemExpress) for 15 min. After washing with PBS, the stained cells were observed under a fluorescence microscope.
Scratch test
Cell migratory ability was evaluated using a scratch test. FLSs were seeded in six-well plates and cultured until 100% confluence. A sterile pipette tip was used to scratch the cell layer. Cell debris was removed by PBS washing. The cells were cultured at 37 °C with 5% CO2 for 24 h. The images of cells were photographed at 0 and 24 h under a light microscope.
Enzyme-linked immunosorbent assay (ELISA)
The levels of IL-1β, IL-6, and TNF-α were measured using their corresponding commercial kits (AnAnti, Wuhan, China) in line with the manufacturer’s protocols.
Quantitative real-time polymerase chain reaction (qPCR)
Total RNA was isolated from FLSs using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The Prime Script RT Reagent kit (Takara, Tokyo, Japan) was used for reverse transcription. Afterwards, qPCR was conducted using the SYBR Premix Ex Taq kit (Takara) following the manufacturer’s instructions. The expression of mRNAs was calculated using the 2−ΔΔCt method by normalizing to β-actin.
Western blotting
FLSs and synovial tissues were lysed using the radio-immunoprecipitation assay buffer containing 1% phenylmethanesulfonyl fluoride on ice. The concentration of proteins in the lysate was detected using a BCA kit (Beyotime, Shanghai, China). After boiling, 40 µg protein was separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. The membranes were blocked with 5% skim milk for 1 h. Next, the membranes were incubated with primary antibodies at 4 °C overnight on a shaker, and labeled with secondary antibody at room temperature for 1 h. The membranes were exposed to Immobilon ECL Ultra Western HRP substrate (Sigma-Aldrich, St. Louis, MO, USA) to show the protein bands. The primary antibodies included anti-LDHA (19987-1-AP; Proteintech, Wuhan, China), anti-β-actin (20536-1-AP; Proteintech), anti-GPI (15171-1-AP; Proteintech), anti-lactyl lysine (kla; PTM-1401RM; PTMBio, Hangzhou, China). The secondary antibody was HRP-conjugated goat anti-rabbit IgG (SA00001-2; Proteintech).
Molecular Docking
The protein structure of LDHA was obtained from the PDB database. The water molecules, co-crystallized ligands, and ions were removed using the Pymol software. Additionally, the 3D structure of β-Sitosterol was acquired from the PubChem database, and this file was converted into the Mol2 format. The docking was performed using the AutoDock Tools and visualized using the Pymol software.
Surface plasmon resonance analysis
The binding ability and affinity between β-Sitosterol and LDHA were analyzed using surface plasmon resonance according to the previously described23.
Cell transfection
FLSs were seeded into six-well plates and cultured for 24 h before transfection. LDHA overexpression plasmids, empty vector, short hairpin RNA targeting GPI (shGPI), shLDHA, short hairpin RNA negative control (shNC), GPI wild-type (GPI-WT) plasmids, and GPI mutant plasmids at K34, K454, and K466 sites (K34R, K454R, and K466R) were transfected into FLSs using Lipofectamine 2000 (Invitrogen) for 48 h.
Bioinformatic analysis
The proteins related to LDHA were predicted using the STRING database.
GPI lactylation (GPI-kla) level determination
GPI protein was immunoprecipitated using the Pierce classic magnetic IP/co-IP kit (Thermo Fisher Scientific, Waltham, MA, USA). FLSs and synovial tissues were lysed using the IP lysis buffer, and the lysate was collected after centrifugation. Kla antibody (5 µg) was incubated with magnetic beads at room temperature for 1 h. The lysate was incubated with the antibody/beads complex at room temperature for 2 h. The beads were washed with IP buffer and the antigen was eluted and collected. The levels of GPI-kla were measured using western blotting using the anti-GPI primary antibody.
Protein stability detection
FLSs were treated with 100 µM cycloheximide (CHX; MedChemExpress) for 0, 8, 16, and 24 h. Proteins were isolated from the cells and the levels of LDHA and GPI were measured using western blotting.
Animals and in vivo study
Sprague Dawley (SD) rats (male, 200–230 g), purchased from Guangdong Medical Laboratory Animal Center (Guangdong, China), were housed in the specific pathogen-free environment with 12/12 h light/dark cycles at 22–25 °C.
The rats were randomly divided into three groups, control, collagen-induced-arthritis (CIA), and CIA + β-Sitosterol groups, with 6 rats in each group.
A CIA rat model was generated as previously described15. Briefly, bovine type II collagen was dissolved in 0.01 M acetic acid and emulsified with an equal volume of complete Freund’s adjuvant (CFA) to prepare the CI emulsion. Additionally, bovine type II collagen was emulsified with an equal volume of CFA to prepare the CII emulsion. The rats received a subcutaneous injection of 0.1 mL CI emulsion at the tailhead. After one week, the rats received a subcutaneous injection of 0.1 mL CII emulsion at the tailhead. After 7 days, the therapeutic potential of β-Sitosterol was evaluated. β-Sitosterol (1 mg/kg/day) was administered by gavage to rats, once a day for 4 weeks.
The severity of arthritis was analyzed using the arthritis index as previously described15. The score ranged from 0 to 4 points: 0 point, normal; 1 point, mildly swollen of ankle and/or redness; 2 points, moderate redness; 3 points, severe redness and swelling of ankles and feet; 4 points, excess edema and joint rigidity. The total score was the cumulative score of the two hind limbs of rats, with a maximum of 8 points.
The toe volume meter (KEW basis, Nanjing, China) was used to detect paw volume. Foot swell was calculated by subtracting the baseline value from the value measured on the last day.
The area and intensity of the hind paw were analyzed using the gait analysis24. Briefly, the rats were subjected to adaptive training for 2 days. After fasting and water restriction for 4 h, the rats were placed in the CatWalk system and freely walked back and forth. The rats successfully crossed the glass track 5 times without stopping halfway and were given food rewards. Formal experiments were the same as adaptive training. Paw area and intensity were recorded.
The arthritis index, foot swell, and gait analysis were all evaluated by two independent observers who were unaware of the grouping and administration details.
This study was approved by the Ethics Committee of The Second Affiliated Hospital of Guangzhou Medical University. All animal experiments should comply with the ARRIVE guidelines. All methods were carried out in accordance with relevant guidelines and regulations.
Histopathological analysis of ankle joints
After euthanizing all rats with isoflurane, the ankles were collected and fixed with 4% paraformaldehyde for 24 h, followed by decalcification with 0.5 M ethylene diamine tetraacetic acid for three weeks. Then, the ankles were used to prepare paraffin Sect. (5 μm). After dewaxing and rehydrating, the sections were stained by hematoxylin for 10 min and eosin for 2 min. The results were observed under a light microplate.
Statistical analysis
Data were analyzed using the GraphPad Prism 8 software and were represented as means ± standard deviation (SD). Differences were analyzed using the Student’s t-test (two groups) and ANOVA (multiple groups). P < 0.05 was considered statistically significant.
Results
FLSs from patients with RA show high proliferative and migratory ability
To identify whether FLS proliferation and migration were involved in RA, primary FLSs were isolated from patients with RA and OA. Cell proliferation was evaluated using CCK-8 and EdU assay. The results revealed that cell viability and the percentage of EdU positive cells were higher in the RA-FLSs group than that in the OA-FLS group (Fig. 1A and C). Next, cell migration was analyzed using scratch test, and we found that the migration rate was also higher in the RA-FLSs group than that in the OA-FLS group (Fig. 1D and E). The results suggest that RA is associated with high proliferation and migration of FLSs.

FLSs from patients with RA show high proliferative and migratory ability. Primary FLSs were isolated from patients with RA, and those from patients with OA were used as the negative control. (A) Cell viability was evaluated using CCK-8. (B, C) Cell proliferation was evaluated using EdU assay, and representative images were shown. EdU positive cells were quantified. (D, E) Cell migration was assessed using scratch test, and representative images were displayed. Migration rate was quantified. n = 6.
β-Sitosterol inhibits the proliferation, migration, and inflammatory response of FLSs
To investigate the function of β-Sitosterol on FLSs, FLSs from patients with RA were treated with various concentrations of β-Sitosterol. Cell proliferation and migration were evaluated. We found that β-Sitosterol at 5, 25 and 50 µM significantly inhibited cell viability and reduced EdU positive cells, compared with the 0 µM group (Fig. 2A and C). Next, β-Sitosterol suppressed FLS migration in a dose-dependent manner (Fig. 2D and E). Additionally, the levels of inflammation-related cytokines were detected using ELISA. As shown in Fig. 2F and H, the levels of IL-1β, IL-6, and TNF-α were reduced by β-Sitosterol treatment in a dose-dependent way. In short, β-Sitosterol inhibits the proliferation, migration, and inflammatory response of FLSs.

β-Sitosterol inhibits the proliferation and migration of FLSs. (A-H) FLSs were exposed to 0 (DMSO), 5, 25, and 50 µM for 24 h. (A) Cell viability was evaluated using CCK-8. (B, C) EdU assay was performed to analyze cell proliferation, and representative images were shown. EdU positive cells were quantified. (D, E) Cell migration was assessed using scratch test, and representative images were displayed. Migration rate was quantified. The levels of (F) IL-1β, (G) IL-6, and (H) TNF-α were measured using ELISA. NS: no significance.
β-Sitosterol binds to LDHA and downregulates its expression
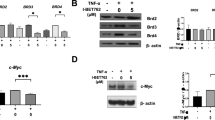
LDHA is associated with histone lactylation. Thus, we identified the effect of β-Sitosterol on LDHA. The results of qPCR and western blotting showed that β-Sitosterol did not affect the mRNA expression of LDHA, but decreased its protein levels (Fig. 3A and C). Next, we found that β-Sitosterol reduced the protein levels of LDHA (Fig. 3D and E). Additionally, molecular docking results showed that β-Sitosterol had moderate interaction with LDHA protein (binding energy=-5.801 kcal/mol; Fig. 3F). Surface plasmon resonance (SPR) analysis results also indicated that β-Sitosterol bound with LDHA (Fig. 3G). The results demonstrated that β-Sitosterol binds to LDHA and decreases LDHA expression in FLSs.

β-Sitosterol binds to LDHA and downregulates its expression. (A-E) FLSs were exposed to DMSO or β-Sitosterol. (A) The mRNA expression of LDHA was detected using qPCR. (B) The protein levels of LDHA were measured using western blotting. (C) Protein levels of LDHA were quantified. (D) CHX was used to treat FLSs for 0, 6, 12, and 24 h, and protein levels of LDHA were measured using western blotting. (E) Protein stability of LDHA was quantified. (F) The binding between β-Sitosterol and LDHA protein was visualized using molecular docking. (G) The binding between β-Sitosterol and LDHA was confirmed using surface plasmon resonance analysis. n = 6.
Overexpression of LDHA reversed the impacts on cellular behaviors caused by β-Sitosterol
To investigate the effect of LDHA on cellular behaviors, we overexpressed LDHA in FLSs. After LDHA overexpression plasmid transfection, the expression of LDHA was elevated (Fig. 4A). Next, cellular biological functions were analyzed. We found that β-Sitosterol inhibited cell proliferation and migration, whereas LDHA overexpression counteracted the inhibition induced by β-Sitosterol (Fig. 4B and F). In addition, the levels of IL-1β, IL-6, and TNF-α were reduced by β-Sitosterol treatment, which was abrogated by LDHA overexpression (Fig. 4G and I). Taken together, β-Sitosterol inhibits FLS proliferation, migration, and inflammatory response via decreasing LDHA expression.

Overexpression of LDHA reversed the impacts on cellular behaviors caused by β-Sitosterol. (A) FLSs were transfected with LDHA overexpression plasmids and empty vectors, and the levels of LDHA were detected using qPCR to confirm successful transfection. (B-I) FLSs were transfected with LDHA overexpression plasmids and treated with β-Sitosterol. (B) Cell viability was evaluated using CCK-8. (C, D) Cell proliferation was assessed using EdU assay, and representative images were shown. EdU positive cells were quantified. (E, F) Cell migration was assessed using scratch test, and representative images were displayed. Migration rate was quantified. ELISA was performed to measure the levels of (G) IL-1β, (H) IL-6, and (I) TNF-α. n = 6.
LDHA promotes the lactylation of GPI at K454 site
To identify which protein could be affected by LDHA, related proteins were predicted using the STRING and shown in Fig. 5A. Among these proteins, GPI is an inducing factor of RA21 and thus was selected for subsequent study. We analyzed the effect of LDHA on lactylation modification of GPI. The results indicated that overexpression of LDHA increased GPI-kla levels and also elevated GPI protein levels (Fig. 5B and C). Additionally, lactylation sites in GPI were analyzed. We found that as compared to the WT group, mutations at K454 site decreased GPI-kla and GPI protein levels, while mutations at K34 and K466 sites did not affect their levels (Fig. 5D and F). Moreover, overexpression of LDHA enhanced the stability of GPI protein (Fig. 5G and H). In summary, LDHA stabilizes GPI protein by promoting its lactylation.

LDHA promotes the lactylation of GPI at K454 site. (A) The protein-protein interaction network of LDHA was predicted using the STRING database. (B) FLSs were transfected with LDHA overexpression plasmids, and the levels of GPI-kla and GPI were measured using western blotting. (C) The levels of GPI-kla and GPI were quantified. (D) The lactylation sites in GPI were evaluated using western blotting, and protein levels of (E) GPI and (F) GPI-Kla were quantified. (G) FLSs were overexpressed with LDHA and treated with CHX, and LDHA protein levels were measured using western blotting. (H) Protein stability of GPI was quantified. n = 6.
Knockdown of GPI reversed the effects on cellular behaviors induced by LDHA overexpression
Rescue experiments were carried out to analyze the functions of GPI. After shGPI transfection, the expression of GPI was decreased, compared with shNC transfection (Fig. 6A). We found that overexpression of LDHA promoted the proliferation, migration, and inflammatory response of FLSs treated with β-Sitosterol, whereas GPI knockdown reversed the effects induced by LDHA overexpression (Fig. 6B and I). The results demonstrated that LDHA mediates the proliferation, migration, and inflammatory response of FLSs by regulating GPI expression.

Knockdown of GPI reversed the effects on cellular behaviors induced by LDHA overexpression. (A) FLSs were transfected with shGPI and shNC, and the levels of GPI were detected using qPCR to confirm successful transfection. (B-I) FLSs were transfected with LDHA overexpression plasmids and shGPI, followed by β-Sitosterol treatment. (B) Cell viability was evaluated using CCK-8. (C, D) Cell proliferation was assessed using EdU assay, and representative images were shown. EdU positive cells were quantified. (E, F) Cell migration was assessed using scratch test, and representative images were displayed. Migration rate was quantified. ELISA was performed to measure the levels of (G) IL-1β, (H) IL-6, and (I) TNF-α. n = 6.
LDHA inhibits FLS proliferation and migration by promoting GPI lactylation
We analyzed the effect of GPI lactylation on cellular behaviors. We transfected shLDHA to FLSs, and LDHA expression was found to be downregulated (Fig. S1A). Knockdown of LDHA inhibited cell proliferation and migration, and decreased the levels of IL-1β, IL-6, and TNF-α, whereas GPI-WT reversed these effects induced by LDHA knockdown. Furthermore, the mutant plasmids of GPI at K454 site reduced these cellular processes, compared with GPI-WT plasmids (Fig. S1B-1I). The results demonstrate that LDHA mediates the proliferation, migration, and inflammatory response of FLSs by facilitating GPI lactylation.
β-Sitosterol attenuates RA in CIA rats
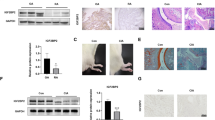
The effect of β-Sitosterol on RA in vivo was analyzed. CIA rat model was established, and we found CIA promoted synovial injury, while β-Sitosterol alleviated synovial damage in CIA rats (Fig. 7A). TNF-α levels were measured in the synovial tissues using western blotting. The results showed that CIA induced the upregulation of TNF-α levels, which was abrogated by β-Sitosterol (Fig. 7B and C). Additionally, the arthritis index was elevated in CIA rats, while β-Sitosterol decreased the arthritis index of CIA rats (Fig. 7D). Foot swell of rats induced by CIA was reversed after β-Sitosterol treatment (Fig. 7E). Moreover, the reduction of paw mean area and paw mean intensity in CIA rats was increased after β-Sitosterol treatment (Fig. 7F and G). Besides, the levels of LDHA, GPI, and the lactylation levels of GPI in the synovial tissues were measured. As shown in Fig. 7H and K, LDHA, GPI, and GPI-kla levels were increased in the synovial tissues of CIA rats, compared with the control group rats. However, β-Sitosterol decreased their levels in CIA rats. Together, β-Sitosterol attenuates RA in CIA rats though inhibiting LDHA-mediated lactylation of GPI.

β-Sitosterol attenuates rheumatoid arthritis in CIA rats. (A) Histological evaluation of the ankle joint in rats was conducted by haematoxylin and eosin staining, and representive images are shown. Scale bar = 50 μm. (B, C) The protein levels of TNF-α were measured using western blotting. (D) The severity of arthritis was analyzed using the arthritis index. (E) Foot swell was detected using the toe volume meter. (F) Paw mean area and (G) intensity were analyzed through gait analysis. (H-K) The levels of LDHA, GPI, and GPI-kla in the synovial tissues of mice were measured by western blotting. n = 6.
Discussion
Our study provides novel insights into the therapeutic potential of β-Sitosterol in RA by demonstrating its inhibitory effects on FLS proliferation, migration, and inflammation. Moreover, we found that β-Sitosterol directly bound to LDHA and subsequently reduced GPI lactylation. Our work offers a novel comprehensive understanding of the molecular mechanisms underlying the effects of β-Sitosterol on FLS.
β-Sitosterol has anti-inflammatory properties, so it has received attention in the treatment of RA. Several studies have explored how β-Sitosterol regulates RA progression. For example, Liu et al.11 have discovered that β-Sitosterol promotes macrophage M2 polarization and inhibits M1 polarization, and thus reduces inflammation in vivo. Qian et al.12 have found that β-Sitosterol treatment suppresses synovial angiogenesis and cartilage injury in collagen-induced arthritis mice by inhibiting the proliferation and migration of endothelial cells. Additionally, β-Sitosterol improves aberrant oxidative stress to serve an antiarthritic effect25. In the joints of patients with RA, FLSs exhibit excessive proliferation and migration capability26. Nevertheless, whether β-Sitosterol affects these features of FLSs is unknown. In this study, we found that β-Sitosterol reduced the proliferation, migration, and pro-inflammatory factor levels in a dose-dependent manner. Moreover, we found that β-Sitosterol attenuated RA in CIA rats. The findings suggest that β-Sitosterol has the potential for the treatment of RA. However, the concentration of β-Sitosterol in the synovial tissues of rats remains unknown due to the limited experimental conditions, which will be further studied in our future work.
Additionally, we have learned that a variety of drugs are used to treat RA, such as methotrexate, salazosulfapyridine, and anti-TNF therapy. However, conventional therapeutic agents such as methotrexate and salazosulfapyridine can induce certain side effects, including gastrointestinal toxicity, malignant diseases, disabilities, and even life-threatening conditions27,28. β-Sitosterol, as a naturally occurring plant sterol, demonstrates favorable safety profiles29. Furthermore, RA is a disease involving multiple pathophysiological mechanisms, highlighting the importance of multi-target therapeutic approaches. β-Sitosterol, with its ability to influence RA progression through various targets and mechanisms11,12,25, holds a broader prospect compared to single-target therapies like anti-TNF treatments.
The underlying mechanism of β-Sitosterol is investigated. The STs in RA have significant changes in glucose metabolism30. Notably, FLSs in patients with RA tend to obtain energy through glycolysis, and inhibition of glycolysis will prevent cell proliferation and migration31. Lactic acid produced by glycolysis drives the occurrence of lactylation modification. However, little is known about the role of lactylation in RA. Only one study indicates that PKM2 lactylation inhibits FLS proliferation and migration15. LDHA is a key enzyme in regulating glycolysis that promotes the production of lactate. In our work, the results indicated that β-Sitosterol bound to LDHA and decreased its expression levels. LDHA is highly expressed in RA FLSs and contributes to the glycolysis and apoptosis of FLSs17. In the present study, we focused on its impacts on FLSs and found that overexpression of LDHA abrogated the inhibition of cellular processes induced by β-Sitosterol. These findings suggest that β-Sitosterol may attenuate RA by decreasing LDHA expression, and LDHA-regulated lactation may be involved in RA progression. A limitation has been resolved in this part. We only identified the interaction between β-Sitosterol and LDHA, but their binding sites remain unclear. According to the molecular docking results, we hypothesized that the binding of β-Sitosterol to LDHA at LEU-164, THR-247, and ARG-168 may influence the ubiquitination or lysosomal degradation pathway of the LDHA protein, subsequently altering protein expression, which will be further investigated in our future research.
The lactylation of GPI promoted by LDHA was found in our work. GPI is a glycolysis-related enzyme that has an arthritogenic role32. It is a potential diagnostic biomarker for RA33. Moreover, GPI is associated with angiogenesis, inflammation, and bone destruction21,34. Zong et al.35 have revealed that GPI promotes FLS proliferation and inhibits apoptosis in RA. Our study indicated that knockdown of GPI counteracted the promotion of proliferation, migration, and inflammatory response induced by LDHA in β-Sitosterol-treated FLSs. Moreover, GPI-WT reversed the inhibition of these cellular behaviors mediated by LDHA knockdown, while GPI-K454R further reversed these behaviors. These findings are the first to demonstrate that β-Sitosterol regulates FLS function in RA by regulating LDHA-mediated lactylation of GPI. However, we found that LDHA was associated with several proteins. Whether they are indeed regulated by LDHA and play a role in the biological behaviors of FLSs still requires further research. This limitation indicates that the role of β-Sitosterol is not merely confined to GPI lactylation. Additionally, we have no evidence to prove that LDHA can directly act as a lactylation-related enzyme. Therefore, we speculated that LDHA induced GPI lactylation by regulating lactic acid production, which will be further studied.
In conclusion, β-Sitosterol inhibits the proliferation, migration, and inflammatory response of FLSs by targeting LDHA, which promotes the lactylation of GPI protein, and thus attenuates RA in CIA rats. Our study highlights the potential of β-Sitosterol as a therapeutic agent in RA and advances our understanding of the molecular mechanisms involved in RA.
Data availability
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Di Matteo, A., Bathon, J. M. & Emery, P. Rheumatoid arthritis. Lancet 402 (10416), 2019–2033 (2023).
Cush, J. J. Rheumatoid arthritis: early diagnosis and treatment. Rheum. Dis. Clin. North. Am. 48 (2), 537–547 (2022).
Radu, A. F. & Bungau, S. G. Management of rheumatoid arthritis: an overview. Cells 10 (11), 2857 (2021).
Diaz-Gonzalez, F. & Hernandez-Hernandez, M. V. Rheumatoid arthritis. Med. Clin. (Barc). 161 (12), 533–542 (2023).
Wu, D. et al. Systemic complications of rheumatoid arthritis: focus on pathogenesis and treatment. Front. Immunol. 13, 1051082 (2022).
Wang, Y. et al. Traditional herbal medicine: therapeutic potential in rheumatoid arthritis. J. Ethnopharmacol. 279, 114368 (2021).
Lower-Nedza, A. D., Kuess, C., Zhao, H., Bian, B. & Brantner, A. H. In vitro anti-inflammatory and antioxidant potential of Si-Miao-San, its modifications and pure compounds. Nat. Prod. Commun. 8 (8), 1137–1141 (2013).
Shen, P. et al. Si Miao San attenuates inflammation and oxidative stress in rats with CIA via the modulation of the Nrf2/ARE/PTEN pathway. Evid. Based Complement. Alternat Med. 2021, 2843623 (2021).
Ma, Y. et al. Efficacy and mechanism of nourishing Yin and purging fire therapy for central precocious puberty based on meta-analysis and network Pharmacology. Med. (Baltim). 102 (48), e36395 (2023).
Khan, Z. et al. Multifunctional roles and Pharmacological potential of beta-sitosterol: emerging evidence toward clinical applications. Chem. Biol. Interact. 365, 110117 (2022).
Liu, R. et al. beta-Sitosterol modulates macrophage polarization and attenuates rheumatoid inflammation in mice. Pharm. Biol. 57 (1), 161–168 (2019).
Qian, K. et al. beta-Sitosterol inhibits rheumatoid synovial angiogenesis through suppressing VEGF signaling pathway. Front. Pharmacol. 12, 816477 (2021).
Gan, P. R., Wu, H., Zhu, Y. L., Shu, Y. & Wei, Y. Glycolysis, a driving force of rheumatoid arthritis. Int. Immunopharmacol. 132, 111913 (2024).
Li, X. et al. Lactate metabolism in human health and disease. Signal. Transduct. Target. Ther. 7 (1), 305 (2022).
Wang, Y. H. et al. Small-molecule targeting PKM2 provides a molecular basis of lactylation-dependent fibroblast-like synoviocytes proliferation Inhibition against rheumatoid arthritis. Eur. J. Pharmacol. 972, 176551 (2024).
Sun, W., Jia, M., Feng, Y. & Cheng, X. Lactate is a Bridge linking Glycolysis and autophagy through lactylation. Autophagy 19 (12), 3240–3241 (2023).
Zhang, M. et al. Influences of the LncRNA TUG1-miRNA-34a-5p network on fibroblast-like synoviocytes (FLSs) dysfunction in rheumatoid arthritis through targeting the lactate dehydrogenase A (LDHA). J. Clin. Lab. Anal. 35 (9), e23969 (2021).
Nishioku, T. et al. Lactate dehydrogenase A Inhibition prevents RANKL-induced osteoclastogenesis by reducing enhanced Glycolysis. J. Pharmacol. Sci. 153 (4), 197–207 (2023).
Wu, S. T. et al. Esculetin inhibits Cancer cell Glycolysis by binding tumor PGK2, GPD2, and GPI. Front. Pharmacol. 11, 379 (2020).
Schaller, M., Burton, D. R. & Ditzel, H. J. Autoantibodies to GPI in rheumatoid arthritis: linkage between an animal model and human disease. Nat. Immunol. 2 (8), 746–753 (2001).
Ebbinghaus, M. et al. Contribution of inflammation and bone destruction to pain in arthritis: A study in murine Glucose-6-Phosphate Isomerase-Induced arthritis. Arthritis Rheumatol. 71 (12), 2016–2026 (2019).
Tsaltskan, V. & Firestein, G. S. Targeting fibroblast-like synoviocytes in rheumatoid arthritis. Curr. Opin. Pharmacol. 67, 102304 (2022).
Wang, X. et al. Paeoniflorin directly binds to TNFR1 to regulate podocyte necroptosis in diabetic kidney disease. Front. Pharmacol. 13, 966645 (2022).
Zhang, Y., Lin, H. & Yi, W. B. Evaluation of the effects of ketamine on spinal anesthesia with Levobupivacaine or ropivacaine. Exp. Ther. Med. 12 (4), 2290–2296 (2016).
Zhang, F. et al. beta-Sitosterol-loaded solid lipid nanoparticles ameliorate complete freund’s adjuvant-induced arthritis in rats: involvement of NF-small ka, CyrillicB and HO-1/Nrf-2 pathway. Drug Deliv. 27 (1), 1329–1341 (2020).
Promila, L., Joshi, A., Khan, S., Aggarwal, A. & Lahiri, A. Role of mitochondrial dysfunction in the pathogenesis of rheumatoid arthritis: looking closely at fibroblast- like synoviocytes. Mitochondrion 73, 62–71 (2023).
Wang, W., Zhou, H. & Liu, L. Side effects of methotrexate therapy for rheumatoid arthritis: A systematic review. Eur. J. Med. Chem. 158, 502–516 (2018).
Nozaki, Y., Inoue, A., Kinoshita, K., Funauchi, M. & Matsumura, I. Efficacy of Iguratimod vs. salazosulfapyridine as the first-line CsDMARD for rheumatoid arthritis. Mod. Rheumatol. 30 (2), 249–258 (2020).
Wang, H., Wang, Z., Zhang, Z., Liu, J. & Hong, L. beta-Sitosterol as a promising anticancer agent for chemoprevention and chemotherapy: mechanisms of action and future prospects. Adv. Nutr. 14 (5), 1085–1110 (2023).
Hu, Z. et al. Metabolic changes in fibroblast-like synoviocytes in rheumatoid Arthritis: state of the Art review. Front. Immunol. 15, 1250884 (2024).
Garcia-Carbonell, R. et al. Critical role of glucose metabolism in rheumatoid arthritis Fibroblast-like synoviocytes. Arthritis Rheumatol. 68 (7), 1614–1626 (2016).
Matsumoto, I. et al. Revisit of autoimmunity to glucose-6-phosphate isomerase in experimental and rheumatoid arthritis. Mod. Rheumatol. 30 (2), 232–238 (2020).
Soleimani, N., Hosseinzadeh, M. & Habibagahi, Z. Value of serum glucose-6-phosphate isomerase in patients with rheumatoid arthritis and correlation with disease activity: A case-control study. J. Educ. Health Promot. 8, 125 (2019).
Lu, Y. et al. Glucose-6-Phosphate isomerase (G6PI) mediates Hypoxia-Induced angiogenesis in rheumatoid arthritis. Sci. Rep. 7, 40274 (2017).
Zong, M. et al. Glucose-6-phosphate isomerase promotes the proliferation and inhibits the apoptosis in fibroblast-like synoviocytes in rheumatoid arthritis. Arthritis Res. Ther. 17 (1), 100 (2015).
Funding
The work was supported by Guangzhou Third-level Famous Traditional Chinese Medicine Studio Construction Project under grant number 202377; Guangdong Provincial Medical Scientific Research Fund under grant number A2023420.
Author information
Authors and Affiliations
Contributions
All authors participated in the design, interpretation of the studies and analysis of the data and review of the manuscript. Y T and X Z drafted the work and revised it critically for important intellectual content; X H, Y Y, Y D, J H and Y Z was responsible for the acquisition, analysis and interpretation of data for the work; C X and H P made substantial contributions to the conception or design of the work. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study was approved by the Ethics Committee of The Second Affiliated Hospital of Guangzhou Medical University. Informed consent was obtained from all individual participants included in the study. This study was performed in line with the principles of the Declaration of Helsinki. All animal experiments should comply with the ARRIVE guidelines. All methods were carried out in accordance with relevant guidelines and regulations.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Tan, Y., Zhong, X., He, X. et al. Beta sitosterol inhibits the proliferation and migration of synoviocytes in rheumatoid arthritis via lactylation of GPI. Sci Rep 15, 25045 (2025). https://doi.org/10.1038/s41598-025-10928-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-10928-9
Keywords
This article is cited by
-
Biochemical investigations, in silico study and targeted delivery aspects of plant phytosterols: β-sitosterol
Inflammopharmacology (2025)
