
Abstract
Atopic dermatitis (AD) is a common chronic inflammatory skin disease characterized by chronic inflammation, barrier impairment, and immunoglobulin E-mediated sensitization. Sodium dodecyl benzene sulfonate (SDBS) is a widely used anionic surfactant which are believed to exacerbate AD according to the hygiene hypothesis. However, recent studies have shown that SDBS does not significantly upregulate the expression of key proinflammatory cytokines, which is inconsistent with previous hypotheses. Furthermore, it is not known whether SDBS affect the inflammatory response to AD. Herein, we used mice with MC903-induced AD-like dermatitis and tumor necrosis factor (TNF)-α/interferon (IFN)-γ (T/I)-treated HaCaT cells to investigate the effects of SDBS on AD. We found that topical use of 0.1% and 1% SDBS did not exacerbate dermatitis in mice. Instead, clinical and histological remission of MC903-induced AD-like dermatitis were observed following the administration of both 0.1% and 1% SDBS, where 1% SDBS treatment showed a greater degree of relief compared to 0.1% SDBS. SDBS also reduced the expression of major cytokines involved in the pathogenesis of AD both in vivo and in vitro. Furthermore, our results showed that SDBS alleviated AD via the nuclear factor kappa B/mitogen-activated protein kinase pathways in a signal transducer and activator of transcription 3-dependent manner. In conclusion, our findings suggest for the first time that the surfactant SDBS has anti-inflammatory properties, which raises new possibilities for detergent use among patients with AD.
Similar content being viewed by others


Introduction
Atopic dermatitis (AD) is a common inflammatory skin disease that affects up to 25% of children and 6–10% of adults1,2. The hyperactivation of Th2-type inflammation and skin barrier dysregulation are two cardinal players in the pathogenesis of AD.
Skin cleansing is an essential component of AD management, as proper hygiene can help remove irritants and allergens from the skin. Surfactants, are the main components of cleansers that aid in reducing the surface tension of the skin to remove dirt, sebum, and sweat in an emulsified form3. However, the hygiene hypothesis suggests a positive correlation between household economic status, hygiene conditions, and the incidence of AD4,5,6, indicating that excessive exposure to detergents may lead to skin irritation and aggravation of AD symptoms7,8.
Sodium dodecyl benzene sulfonate (SDBS) is a widely used anionic surfactant, primarily employed as a detergent and emulsifier in household, industrial, and cosmetic formulations due to its surfactant properties9. In cosmetic applications, SDBS is used as a cleansing agent, with its dermal absorption dependent on pH. Toxicological assessments indicate that dodecyl benzene sulfonate salts exhibit low acute toxicity in oral and dermal animal studies, with no systemic toxic effects observed in repeat-dose dermal exposure. However, at high concentrations (15%), SDBS has been reported to cause severe skin irritation in animal models, similar to other detergents. The Cosmetic Ingredient Review (CIR) Expert Panel has recommended that formulations containing SDBS should be optimized to minimize irritancy potential9.
The cytotoxic effects of SDBS on human keratinocytes have been investigated, revealing that at concentrations of 20 µg/ml and above, SDBS induces direct cytotoxicity and morphological changes in HaCaT cells. However, unlike those of other well-studied anionic surfactants, such as sodium lauryl sulfate (SLS), SDBS does not significantly upregulate the expression of key proinflammatory cytokines such as IL-1α, IL-6, IL-8, and TNF-α, suggesting a different mechanism of action in skin inflammation10. Reinartz et al.11 suggested that variations in chain length among anionic surfactants may lead to differences in their biological effects, highlighting the need for further investigation into SDBS.
Our previous studies have provided preliminary evidence that SDBS may modulate inflammatory responses in human keratinocytes10. In contrast, SLS have been shown to upregulate thymic stromal lymphopoietin (TSLP) expression12—a key cytokine in AD pathogenesis. Our unpublished data indicate that SDBS does not induce proinflammatory cytokine expression and may, in fact, inhibit TSLP production. These findings suggest that SDBS might play a distinct role in AD-related inflammation, prompting us to further investigate its effects in an AD-like model.
In this study, we investigated the effects and underlying mechanisms of SDBS in the MC903-induced AD-like mouse model and in TNF-α/IFN-γ (T/I)-treated keratinocytes. We examined AD clinical features, histology, immune responses, and associated signaling pathways.
Materials and methods
Animal experiments
Six-week-old male BALB/c mice were purchased from Beijing Vital River Laboratory Animal Technology Company (China). Mice were maintained under specific pathogen-free conditions. All animal experiments were performed following the ARRIVE guidelines to ensure proper welfare and ethical treatment. The study protocol was reviewed and approved by the ethics committee of Peking University Health and Science Center and performed following their guidelines.
To establish an AD-like murine model, male BALB/c mice were topically treated with 20 µl (2 nmol) of MC903 per ear on the dorsal ear surface daily for 20 d13,14. Mice treated with ethanol served as the control group. From d 15 to 20, the mice were topically treated with 20 µl of vehicle (ethanol), 0.1% SDBS (Sigma-Aldrich, USA; dissolved in ethanol), 1% SDBS, desonide cream (Huapont Pharm, China), or tacrolimus ointment (Astellas Pharma, Japan) as a positive control (Fig. 1B). From d 15 onward, a signal transducer and activator of transcription 3 (STAT3) agonist colivelin was injected intraperitoneally into the mice 2 h before topical intervention at a dose of 7 nmol/animal (Fig. 4B). On d 15, 17, 19, and 21, the mice were anesthetized with 2% isoflurane, and photographs were taken using a digital camera (Canon EOS 1200D, Japan). The severity of dermatitis was evaluated and scored for erythema/hemorrhage, edema, and scaling/dryness (0, none; 1, mild; 2, moderate; or 3, severe). Ear thickness was also measured and recorded using a Vernier caliper (Satatool, USA).

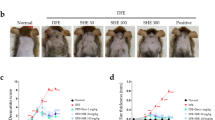
Topical sodium dodecyl benzene sulfonate (SDBS) treatment attenuated MC903-induced Atopic dermatitis (AD)-like dermatitis. (A) The molecular structural formula of SDBS. (B) Schematic diagram of the timeline for the induction of the mouse MC903-induced AD-like model. (C) Representative images of clinical features of MC903-induced AD-like mice treated with ethanol or SDBS or desonide or tacrolimus at d 21. (D) Thicknesses of mouse ear lesions (mm) (left panel) and dermatitis severity scores of mouse lesions (right panel) at d 21. (E) Serum levels of total immunoglobulin E (IgE) in the MC903-induced AD-like mice treated with ethanol or SDBS or desonide or tacrolimus. (F) Representative images of histological sections of the skin stained with hematoxylin and eosin (H&E) (400×) at d 21 (left panel) and thickness of the epidermis (µm) (right panel). (G) Toluidine blue staining images of ethanol or SDBS or desonide or tacrolimus-treated skin tissue from MC903-induced AD-like mice to show the numbers of infiltrating mast cells. Right panel shows mast cell counts which was performed in 5 randomly selected bright high-power field (HPF) (400×). The data are presented as the mean ± standard error of the mean (n = 8). Statistically significant differences indicated as ####, p < 0.0001, ##, p < 0.01 compared to the Control/MC903 group; ****, p < 0.0001, ***, p < 0.001, *, p < 0.05 compared to the MC903 + Treatment/MC903 group.
Histological analysis
At the end of the experiment, each mouse’s ears were removed and fixed in 4% methyl aldehyde for 24 h. The skin specimens were then dehydrated, embedded in paraffin, and stained with hematoxylin and eosin (H&E) or toluidine blue for mast cell detection. The number of mast cells was counted in 5 high-power fields (HPF) chosen at random in each slide by 3 different pathologists. Images were captured using a light microscope (Leica DM4000B, Germany).
Immunohistochemical staining
The slides were treated with xylene and ethanol, rinsed with phosphate-buffered saline containing 0.025% TritonX-100, and subjected to a heat-induced antigen retrieval step. The slides were incubated with normal goat serum for 20 min at room temperature. Then, an anti-TSLP antibody (1:500, rabbit polyclonal, Abcam, UK) was applied and the slides were incubated at 4 °C overnight, followed by incubation with horseradish peroxidase-conjugated anti-rabbit IgG for 30 min at room temperature and incubation with the final developmental chromogen, diaminobenzidine. The number of TSLP+ cells were counted in five high-power fields (HPF) chosen at random in each slide by 3 different pathologists.
Cell culture
Human keratinocyte HaCaT cells (Sigma-Aldrich, USA) were cultured in high glucose DMEM (Sigma-Aldrich, USA) with 10% FBS (Sigma-Aldrich, USA) in a 5% CO2 cell incubator at 37 °C.
Cell viability
HaCaT cells were seeded at a density of 5 × 104 cells/well in a 96-well culture plate and treated with SDBS or SLS of gradient concentration. After 24 h of SDBS or SLS treatment, 300 µL of MTT (3-(4,5-dimethylthiazol-2-yl)−2,5-diphenyl-2 H-tetrazolium) measuring the metabolic activity of cells was added per well. The cells were incubated at 37 °C for 2–4 h and then dissolved in crystal violet with DMSO. The absorbance intensity at a wavelength of 550 nm was measured by the microplate reader (Perkin-Elmer, USA).
Enzyme-linked immunosorbent assay
Mouse blood was collected on the final day of the experiments. Serum was obtained by centrifugation at 3000 rpm for 10 min. Mouse immunoglobulin E (IgE), TSLP, and interleukin (IL)−6 in serum were measured using enzyme-linked immunosorbent assay (ELISA) kits (Abcam, UK) according to the manufacturer’s instructions.
To assess proinflammatory cytokine expression, HaCaT cells were pretreated with different concentrations of SDBS (0.5, 1, 2.5, 5, 10, 15, 20, 25, or 30 µg/ml) for 24 h and then stimulated with T/I for an additional 24 h. To assess the effects of the STAT3 agonist colivelin, HaCaT cells were pretreated with colivelin (2 µg/ml) for 1 h prior to incubation with 10 µg/ml SDBS for 24 h, and then stimulated with T/I for an additional 24 h. The supernatant of the HaCaT cells was collected, and the cytokines (IL-6, TSLP) were measured using ELISA kits (Abgent, USA) according to the manufacturer’s instructions.
Quantitative real-time polymerase chain reaction
Total RNA was extracted using TRIzol (Invitrogen, USA) according to the manufacturer’s instructions. cDNA was synthesized from 500 ng of each RNA sample in a reaction volume of 20 µl with a cDNA synthesis kit (Invitrogen, USA). Transcripts were detected using SYBR Green (Applied Biosystems, Sweden). Duplicate reactions were set up for each sample. The sequences of the primers used were as follows: Tslp (forward: 5’-TGAAGCCTATGACACCACTGA-3’, reverse: 5’-TCCCCTACGCTTTCTTGTCCT-3’); Il4 (forward: 5’-GGTCTCAACCCCCAGCTAGT-3’, reverse: 5’-GCC-GATGATCTCTCTCAAGTGAT-3’); Il6 (forward: 5’-TAGTCCTTCCTACCCCAATTTCC-3’, reverse: 5’-TTGGTCCTTAGCCACTCCTTC-3’); Il13 (forward: 5’-CCTGGCTCTTGCTTGCCTT-3’, reverse: 5’-GGTCTTGTGTGATGTTGCTCA-3’); and β-actin (Actb; forward: 5’-CATGTACGTTGCTATCCAGGC-3’, reverse: 5’-CTCCT-TAATGTCACGCACGAT-3’). TSLP (forward: 5’- ATGTTCGCCATGAAAACTAAGGC-3’, reverse: 5’- GCGACGCCACAATCCTTGTA − 3’); IL6 (forward: 5’- ACTCACCTCTTCAGAACGAATTG − 3’, reverse: 5’- CCATCTTTGGAAGGTTCAGGTTG − 3’); IL33 (forward: 5’- GTGACGGTGTTGATGGTAAGAT − 3’, reverse: 5’- AGCTCCACAGAGTGTTCCTTG − 3’); CCL17 (forward: 5’- TTCTCTGCAGCACATCCACGCA − 3’, reverse: 5’-CTGGAGCAGTCCTCAGATGTCT − 3’); and GAPDH (forward: 5’- GGAGCGAGATCCCTCCAAAAT − 3’, reverse: 5’-GGCTGTTGTCATACTTCTCATGG − 3’).
Western blots
Protein was extracted from HaCaT cells and mouse ears using radioimmunoprecipitation assay buffer (Beyotime, China), and the total protein concentration was determined with a bicinchoninic acid protein assay kit (Beyotime, China). Protein samples were loaded and separated on 10% sodium dodecyl sulfate–polyacrylamide gels and transferred to polyvinylidene fluoride membranes (PALL, USA). The membranes were blocked in 5% bovine serum albumin for 1 h and incubated at 4 °C overnight with the following specific primary antibodies: human phospho-STAT3, STAT3, phospho-p38, p38, phospho-inhibitor kappa B (IκB)-α, IκB-α, phospho-cytoplasmic NF-κB p65, cytoplasmic NF-κB p65, phospho-nuclear NF-κB p65, nuclear NF-κB p65, phospho-extracelluar regulating kinase (ERK)1/2 (Cell Signaling Technology, USA), ERK1/2 (Abcam, USA), β-actin, and laminin B (Connoisseur, China). β-actin was used as the cytoplasmic control, while Lamin B was used as the nuclear control15,16. Mouse antibodies against phospho-STAT3, STAT3, phospho-p38, p38, phospho-IκB-α, IκB-α, phospho-total NF-κB p65, total NF-κB p65, phospho-ERK1/2, ERK1/2, β-actin (Cell Signaling Technology, USA), and GAPDH (Proteintech, USA) were used for murine samples. After being washed in Tris-buffered saline with 0.1% Tween 20 three times, the membranes were then incubated with horseradish peroxidase-conjugated anti-rabbit or anti-mouse antibodies (Cell Signaling Technology, USA) for 1 h. Enhanced chemiluminescence imaging was performed and relative protein levels were analyzed using Fiji/ImageJ version 2.16.0/1.54 g software.
Statistical analyses
Statistical analysis was performed with GraphPad Prism version 10.3.1 (GraphPad Software, USA). Two-sample independent t-tests were used for comparisons between the two groups. One-way analysis of variance was used for comparisons among groups. A P-value < 0.05 was considered indicative of statistical significance in all analyses.
Results
Topical SDBS treatment attenuated MC903-induced AD-like dermatitis
To determine the effects of SDBS on the pathogenesis of AD, MC903 was applied to mouse ears to induce AD-like skin lesions and then SDBS was topically applied (Fig. 1B). The application of MC903 successfully induced AD-like dermatitis. The topical use of 0.1% and 1% SDBS significantly inhibited dermatitis, as indicated by dermatitis scores (p < 0.001) and ear thicknesses (p < 0.001) (Fig. 1C-D). Histopathology revealed that MC903-induced AD-like ear lesions displayed pronounced hyperkeratosis, parakeratosis, acanthosis, and lymphocyte infiltration, and 0.1% or 1% SDBS treatment significantly alleviated those pathological alterations, which was consistent with the reduction in epidermal thickness (p < 0.001) (Fig. 1F). Furthermore, treatment with 0.1% and 1% SDBS reduced mast cell infiltration (p < 0.001) (Fig. 1G) and decreased the serum IgE level (p < 0.0001) (Fig. 1E) in mice with MC903-induced AD-like dermatitis. The effects of both 0.1% and 1% SDBS on MC903-induced dermatitis remission were similar to those of desonide cream and tacrolimus ointment in terms of the dermatitis score, ear thickness, epidermal thickness, mast cell infiltration, and serum IgE level (Fig. 1C-G).
SDBS inhibited inflammatory cytokine production in both AD-like lesions and T/I-induced HaCaT cells
We next explored the effects of SDBS on AD cytokines at both the protein and mRNA levels. We first found abundant cytoplasmic staining for TSLP in the epidermis of the ears of mice with MC903-induced AD-like dermatitis by immunohistochemical staining (Fig. 2A). Moreover, ELISA showed that the serum TSLP and IL-6 levels were higher than in control mice (Fig. 2B). Nevertheless, through quantitative real-time polymerase chain reaction, we found that Tslp, Il4, Il6, and Il13 mRNA levels were significantly elevated in mice with MC903-induced dermatitis (Fig. 2C). However, treatment with 0.1% or 1% SDBS resulted in lower cytoplasmic TSLP staining (p < 0.001), serum TSLP (p < 0.0001) and IL-6 (p < 0.0001) levels, and mRNA levels of Tslp (p < 0.01), Il4 (p < 0.0001), Il6 (p < 0.001), and Il13 (p < 0.0001) compared with MC903-induced AD-like mice group to a similar extent as treatment with desonide or tacrolimus (Fig. 2A-C). Similarly, in vitro ELISA experiments revealed that different concentrations of SDBS (2, 5, or 10 µg/ml) can suppress the increase in TSLP (p < 0.01) and IL-6 (p < 0.01) protein levels (Fig. 2E) and mRNA levels of TSLP, IL6, IL33, and CCL17 (p < 0.05) (Fig. S1A) induced by cotreatment with T/I. Treatment with SLS at concentrations of 2, 5, and 10 µg/ml did not result in suppression of TSLP, IL6, IL33, and CCL17 mRNA levels in T/I-induced HaCaT cells. Instead, a statistically significant increase in the mRNA levels of TSLP, IL6, and IL33 was observed (p < 0.05) (Fig. S1A). Notably, SDBS at a concentration of < 20 µg/ml did not affect the viability of HaCaT cells in culture (Fig. 2D). Under treatment with the same concentrations of SLS, we observed that concentrations above (and including) 5 µg/ml significantly reduce the viability of HaCaT cells (Fig. S1A).

SDBS decreased the production of proinflammatory cytokines in MC903-induced AD-like lesions and tumor necrosis factor (TNF)-α/interferon (IFN)-γ (T/I)-treated HaCaT cells. (A) Representative immunohistochemical analysis with specific antibody showed the expressions of TSLP, indicated in brown color (400×). Right panel shows quantification of the cells staining positive of TSLP antibody from 5 randomly selected bright HPF per Sect. (400×) (n = 8). (B) Serum levels of thymic stromal lymphopoietin (TSLP) (left panel) and interleukin (IL)−6 (right panel) in MC903-induced AD-like mice treated with ethanol or SDBS or desonide or tacrolimus (n = 8). (C) Relative expression of Tslp, Il4, Il6, and Il13 mRNA normalized to beta-actin in MC903-induced AD-like ears treated with ethanol or SDBS or desonide or tacrolimus (n = 8). (D) Cell viability of HaCaT cells after pretreatment with different concentrations of SDBS (0, 5, 10, 15, 20, 25 and 30 µg/ml) (n = 3). (E) TSLP and IL-6 protein levels were analyzed in T/I-induced HaCaT cells treated with different concentrations of SDBS (0, 2, 5, 10, 20 µg/ml) by enzyme-linked immunosorbent assay (ELISA) (n = 6). The data are presented as the mean ± standard error of the mean. Statistically significant differences indicated as ####, p < 0.0001, ###, p < 0.001, #, p < 0.05 compared to the Control/MC903 group or Control/TI group; ****, p < 0.0001, ***, p < 0.001, **, p < 0.01*, p < 0.05 compared to the MC903 + Treatment/MC903 group or SDBS/model group.
SDBS suppressed the activation of the STAT3, NF-κB, and MAPK signaling pathways in vivo and in vitro
We next investigated the effects of SDBS on molecular signaling pathways in both the in vivo and in vitro models. Western blot analysis of the ears of mice with MC903-induced AD-like dermatitis revealed that both 0.1% and 1% SDBS suppressed the phosphorylation of STAT3, total NF-κB P65, IκB-α, and P38 MAPK, which had been significantly higher than control levels in MC903-induced AD-like lesions (Fig. 3A-B). Similarly, T/I-induced phosphorylation of the STAT3, P38 MAPK, and ERK1/2 proteins in HaCaT cells was markedly inhibited following SDBS treatment. Furthermore, although the cytoplasmic NF-κB P65 protein levels did not differ, the nuclear NF-κB P65 levels were markedly lower in all the SDBS groups than in the T/I-treated group. Moreover, the level of phospho-IκB-α was markedly lower in the 20 µg/ml SDBS and 10 µg/ml SDBS groups than in the T/I-treated group (Fig. 3C-D).

SDBS suppressed the activation of signal transducer and activator of transcription 3 (STAT3), nuclear factor kappa B (NF-κB), and mitogen-activated protein kinase (MAPK) signaling pathways in vivo and in vitro. (A) Protein levels of p-STAT3, STAT3, p-total NF-κB p65, total NF-κB p65, p-inhibitor kappa B (IκB)-α, IκB-α, p-p38 and p38 in MC903-induced AD-like ears treated with ethanol or SDBS or desonide or tacrolimus were analyzed using Western immunoblotting (n = 5). (B) The protein levels of p-STAT3, STAT3, p-total NF-κB p65, total NF-κB p65, p-IκB-α, IκB-α, p-p38 and p38 in MC903-induced AD-like ears treated with ethanol or SDBS or desonide or tacrolimus were quantified by band density and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (n = 5). (C) Protein levels of p-STAT3, STAT3, p-p38, p38, p-extracellular signal-regulated kinase (ERK) 1/2, ERK1/2, p-cytoplasmic NF-κB p65, cytoplasmic NF-κB p65, p-nuclear NF-κB p65, nuclear NF-κB p65, p-IκB-α, IκB-α in T/I-induced HaCaT cells treated with different concentrations of SDBS (0, 2, 5, 10, 20 µg/ml) were analyzed using Western immunoblotting (n = 3). (D) The protein levels of p-STAT3, STAT3, p-p38, p38, p-ERK 1/2, ERK1/2, p-cytoplasmic NF-κB p65, cytoplasmic NF-κB p65, p-IκB-α and IκB-α in T/I-induced HaCaT cells treated with different concentrations of SDBS (0, 2, 5, 10, 20 µg/ml) were quantified by band density and normalized to beta-actin. The protein levels of p-nuclear NF-κB p65 and nuclear NF-κB p65 were quantified by band density and normalized to Lamin B (n = 3). The data are presented as the mean ± standard error of the mean. Statistically significant differences indicated as ##, p < 0.01, #, p < 0.05 compared to the Control/MC903 group or Control/T/I group; **, p < 0.01, *, p < 0.05, compared to the MC903 + Treatment/MC903 group or SDBS/model group.
SDBS suppressed MC903-induced AD-like dermatitis and underlying NF-κB and MAPK pathways in a STAT3-dependent manner
To further investigate whether the effects of SDBS on T/I-treated HaCaT cells were STAT3-dependent, we administered the STAT3-selective agonist colivelin to HaCaT cells. As expected, both TSLP and IL-6 levels in the T/I-treated group were higher than in the control group, and were decreased after SDBS treatment. The addition of colivelin to T/I + SDBS-treated HaCaT cells resulted in significantly higher protein levels of TSLP and IL-6 compared to cells treated with T/I + SDBS alone (p < 0.0001) (Fig. 4A), indicating that colivelin attenuates the suppressive effect of SDBS on TSLP and IL-6 protein levels in T/I-treated HaCaT cells (Fig. 4A). Then, we administered colivelin to mice with MC903-induced AD-like dermatitis, as described in Fig. 4B. As expected, both the clinical and histopathological manifestations of MC903-induced dermatitis were alleviated by SDBS, as were the dermatitis score and epidermal thickness. The dermatitis score (p < 0.05) and epidermal thickness (p < 0.01) were significantly higher in the colivelin + T/I + SDBS group than in the T/I + SDBS group (Fig. 4C-E), suggesting that colivelin diminishes the ameliorative effects of SDBS on both the clinical and pathological manifestations of MC903-induced AD-like dermatitis. Finally, we investigated the impact of the STAT3-selective agonist colivelin on signaling pathways. The phosphorylation of STAT3, IκB-α, and ERK1/2 was markedly greater in MC903-induced AD-like lesions than in control skin and was downregulated by SDBS. The ratios of p-STAT3/STAT (p < 0.05), p-IκB-α/IκB-α (p < 0.0001), and p-ERK1/2/ERK1/2 (p < 0.0001) were significantly higher in the colivelin + T/I + SDBS group (both 0.1% and 1%) compared with the T/I + SDBS group (Fig. 4F-G). This suggests that colivelin interferes with the suppressive effect of SDBS on the phosphorylation of STAT3, IκB-α, and ERK1/2 in the context of MC903-induced AD-like lesions.

SDBS suppressed MC903-induced AD-like dermatitis and underlying NF-κB and MAPK pathways in a STAT3-dependent manner. (A) TSLP and IL-6 protein levels were analyzed in T/I-induced HaCaT cells treated with 10 µg/ml SDBS and/or colivelin (n = 6). (B) Schematic diagram of the timeline for the induction of the mouse MC903-induced AD-like model with SDBS treatment and intraperitoneal injection of colivelin. (C) Representative images of clinical features and H&E staining of histological Sect. (200×) of MC903-induced AD-like mice treated with ethanol or SDBS or desonide and/or colivelin at d 21 (n = 8). (D) Dermatitis severity scores of mouse lesions (n = 8). (E) Epidermal thicknesses of mouse lesions (µm) (n = 8). (F) Protein levels of p-STAT3, STAT3, p-IκΒ-α, IκΒ-α, p-ERK1/2 and ERK1/2 in MC903-induced AD-like ears treated with ethanol or SDBS or desonide and/or colivelin were analyzed using Western immunoblotting (n = 5). (G) The protein levels of p-STAT3, STAT3, p-IκΒ-α, IκΒ-α, p-ERK1/2 and ERK1/2 in MC903-induced AD-like ears treated with ethanol or SDBS or desonide and/or colivelin were quantified by band density and normalized to beta-actin (n = 5). The data are presented as the mean ± standard error of the mean. Statistically significant differences indicated as *P < 0.05, **P < 0.01, *** P < 0.001, and **** P < 0.0001, ‡P < 0.05, ‡‡, P < 0.01 compared to the MC903 + SDBS group.
Discussion
Sodium dodecyl benzene sulfonate (SDBS) is a widely used anionic surfactant. Contrary to what might be expected based on the hygiene hypothesis, previous studies have indicated that SDBS exposure does not lead to a significant upregulation of key proinflammatory cytokines, including IL-1α, IL-6, IL-8, and TNF-α. In this study, we performed in vivo and in vitro experiments to investigate the effects of SDBS on the pathogenesis of AD. We found that 0.1% and 1% SDBS alleviated clinical symptoms, including dermatitis severity, lesion thickness, and decreased mast cell infiltration and serum IgE levels in mice with MC903-induced AD-like dermatitis. In addition, 0.1% or 1% SDBS reduced serum TSLP and IL-6 levels and inhibited cutaneous Tslp, Il6, Il4, and Il13 mRNA expression. In vitro experiments showed that different concentrations of SDBS (2, 5, or 10 µg/ml) inhibited TSLP and IL-6 protein levels and mRNA levels of TSLP, IL6, IL33, and CCL17 in T/I-stimulated HaCaT cells without affecting cell viability. Furthermore, we found that SDBS suppressed the activation of the STAT3, NF-κB, and MAPK signaling pathways both in vitro and in vivo. The inhibitory effects of SDBS on MC903-induced AD-like dermatitis—from the perspective of clinical symptoms, histology, and phosphorylation of STAT3, IκB-α, and ERK1/2—were relatively abolished by in vivo administration of a STAT3 agonist. Collectively, our findings revealed that 0.1% or 1% SDBS exerted a protective effect against MC903-induced AD-like dermatitis, which was associated with downstream activation of proinflammatory NF-κB and MAPK in a STAT3-dependent manner (Fig. 5).

Schematic diagram for the effects of SDBS on AD-like inflammation. SDBS decreased mast cell infiltration and serum IgE levels, reduced serum TSLP and IL-6 levels and inhibited cutaneous Tslp, Il6, Il4, and Il13 mRNA expression. The suppressing effects of SDBS were highly associated with underlying NF-κB/MAPK pathway in a STAT3-dependent manner.
AD, one of the most common inflammatory skin diseases, is characterized by a chronic, relapsing form of skin inflammation, epidermal barrier function disturbance, and IgE-mediated sensitization17. The topical MC903-induced AD-like mouse model has been proven as a convenient preclinical AD model for exploring therapeutic avenues, identifying factors involved in the development of AD, and characterizing various aspects of AD pathogenesis14. Therefore, we used mice with MC903-induced AD-like dermatitis as a clinical model to investigate the effects of SDBS on AD in vivo. In this study, the MC903-treated group exhibited pronounced erythema, edema, crusting, and telangiectasia, which were alleviated by SDBS, as were ear thickening and dermatitis score. It is worth noting that 1% SDBS showed a greater degree of relief for dermatitis score and ear thickness compared to 0.1% SDBS. The typical histological manifestations of AD are epidermal hyperplasia and spongiosis, with perivascular infiltrates of lymphocytes. In this study, while the MC903-treated group manifested typical AD histological features by H&E staining, treatment with SDBS resulted in remarkable improvement. Histopathology also confirmed a significant reduction in epidermal thickness in the SDBS groups, where 1% SDBS treatment exerted a greater reduction than 0.1% SDBS treatment.
The proximate mechanism for AD is predominantly type 2-skewed immune dysregulation and IgE-mediated hypersensitivity18,19. Elevated total serum IgE is a major hallmark of AD20. In this study, we found that SDBS decreased total serum IgE levels in mice with MC903-induced AD-like dermatitis. Mast cells, which are activated by IgE, play a major role in AD and are the first-line responders to allergen stimulation21. This study revealed that SDBS reduced mast cell infiltration of MC903-induced AD-like lesions. TSLP is a major regulator and a key initiator of Th2 immune responses in AD22,23,24,25. Our study revealed that SDBS suppressed lesion TSLP protein and mRNA expression and serum TSLP levels in MC903-induced AD-like mice. Many studies have shown that the combination of TNF-α and IFN-γ can stimulate keratinocytes and induce the expression of inflammatory mediators, thereby promoting inflammation26,27. Therefore, we used T/I-treated HaCaT cells to investigate the effects of SDBS on proinflammatory factors derived from keratinocytes in AD. SDBS also decreased TSLP protein levels and mRNA levels of TSLP in T/I-treated HaCaT cells. IL-4 and IL-13, which induce IgE class switching and promote inflammation1,28, are key factors in the pathogenesis of AD29. This study showed that SDBS reduced cutaneous Il4 and Il13 mRNA expression in MC903-induced AD-like lesions. IL-6, a multifunctional cytokine that has been shown to prevent T cell apoptosis30,31, promotes Th2 differentiation and restrains the function of Treg cells, contributing greatly to the progression and protraction of AD32,33. Our study revealed that SDBS downregulated both lesion Il6 mRNA expression and serum IL-6 levels in MC903-induced AD-like mice. In vitro experiments showed that SDBS decreased IL-6 protein levels and mRNA levels of IL6 in T/I-treated HaCaT cells. IL-33, an epithelial-derived alarmin cytokine overexpressed in atopic dermatitis, drives type 2 inflammation by activating ILC2s, mast cells, and eosinophils34,35. CCL17, a chemokine that recruits CCR4⁺ or CCR8⁺ cells and plays a key role in Th2-driven inflammation, correlating with disease severity, serves as a potential biomarker for disease activity and early risk prediction36,37. This study showed that SDBS decreased mRNA levels of IL33 and CCL17 in T/I treated HaCaT cells. The well-studied anionic surfactant SLS was tested at the same concentration. Unlike SDBS, SLS does not suppress the expression of cytokines (TSLP, IL6, IL33, and CCL17) induced by T/I in HaCaT cells; instead, it further stimulates their expression (TSLP, IL6, and IL33).
STAT3 is a transcription factor belonging to the STAT family that mediates signal transduction from the membrane to the nucleus38. The STAT3 signaling pathway and associated key cytokines play a central role in the pathogenesis of AD39. It has been reported that targeting STAT3 activation is a new modality for treating AD40. Our study revealed that SDBS prominently suppressed the activation of STAT3 in the ears of mice with MC903-induced AD-like dermatitis, in accordance with the in vitro results, in which STAT3 phosphorylation was markedly reduced by SDBS in T/I-treated HaCaT cells. The NF-κB pathway, which mediates the activation of multiple transcription factors involved in inflammation, is interconnected with the MAPK signaling pathway; the phosphorylation of MAPK induces proinflammatory cytokine expression and enhances the inflammatory responses of various immune cells. Multiple studies have confirmed that AD pathogenesis is closely associated with the activation of and synergistic effects between the NF-κB and MAPK signaling pathways41,42. Inhibition of the P38 MAPK, ERK, and NF-κB signaling pathways has been shown to alleviate AD43,44. This finding is consistent with those of our study, in which we found that SDBS suppressed the phosphorylation of total NF-κB P65, IκB-α, and P38 MAPK in the ears of mice with MC903-induced AD-like dermatitis. Similarly, our in vitro study showed that SDBS inhibited the phosphorylation of P38 MAPK, and ERK1/2 in T/I-treated HaCaT cells. Activation of NF-κB involves the phosphorylation and ubiquitination of IκB-α followed by nuclear translocation of NF-κB P6545. In our in vitro study, SDBS blocked the phosphorylation of cytosolic IκB-α and the nuclear translocation of NF-κB P65 in T/I-treated HaCaT cells. In conclusion, our results suggested that SDBS suppressed the activation of the STAT3, MAPK, and NF-κB signaling pathways in MC903-induced AD-like mice and T/I-treated HaCaT cells.
To further explore the mechanisms underlying the effects of SDBS in AD, we first administered the selective STAT3 agonist colivelin to T/I-treated HaCaT cells. Our results showed that colivelin largely abolished the suppressive effects of SDBS on TSLP and IL-6. Then, we administered colivelin to mice with MC903-induced AD-like dermatitis and found that colivelin mitigated the suppressive effects of SDBS on clinical and histopathological manifestations, dermatitis scores, and epidermal thickness, indicating a STAT3-dependent mechanism for SDBS’s suppression of MC903-induced dermatitis. Several studies have indicated that reciprocal crosstalk among the STAT3, NF-κB, and MAPK signaling pathways forms a key axis in the pathogenesis of inflammatory diseases46,47. It has been reported that STAT3 activation can robustly lead to increased activity of the NF-κB and MAPK signaling pathways48,49. Our data revealed that the pharmacological activation of STAT3 by colivelin significantly increased the activation of both IκB and ERK1/2 and largely abolished the suppressive effects of SDBS on these two signaling pathways in MC903-induced AD-like dermatitis, indicating that SDBS suppresses NF-κB and MAPK activation in a STAT3-dependent manner.
Our findings regarding SDBS’s effects on AD may be explained by several interconnected mechanisms. Firstly, SDBS’s reported ability to disrupt bacterial cell membranes50,51, and thus reduce the Staphylococcus aureus load often associated with AD exacerbation, could directly attenuate lesion severity. This antimicrobial action may modulate inflammation and immune responses, potentially influencing the STAT and NF-κB pathways, which are key regulators of inflammatory processes in AD. Secondly, SDBS has been shown to alter the production of short-chain fatty acids (SCFAs)52. Given that decreased SCFA levels are linked to increased eczema risk and that SCFAs play a role in improving skin barrier function and reducing inflammation53, SDBS’s impact on SCFA production could represent another microbiome-mediated mechanism contributing to AD pathogenesis. Finally, previous research has shown that low-dose SDBS can inhibit CYP1A1/CYP1A2 and GSTs, enzymes involved in redox balance54. Considering the established role of oxidative stress in AD, this SDBS-induced modulation of oxidative stress homeostasis could further contribute to the suppression of inflammatory signaling pathways, including STAT and NF-κB, ultimately influencing the course of the disease.
There are several limitations to the present study. First, the scope of the study is limited, and since barrier function is a crucial aspect of AD and surfactants have been proven to be potent skin irritants, the effects of SDBS on the barrier function of AD patients warrant future clinical research. Second, although the suppressive effects of SDBS on AD are STAT3-dependent, the precise mechanisms underlying the effects of SDBS on STAT3 activation require further exploration.
SDBS is a widely used anionic surfactant; however, little is known about the effects of SDBS on the skin, and the complex biological responses underlying the effects of SDBS on AD are poorly understood. Our study showed, for the first time, that 0.1% and 1% SDBS could alleviate both clinical and histological manifestations of MC903-induced AD-like dermatitis. SDBS inhibited the expression of key cytokines involved in AD pathogenesis both in vivo and in vitro, and protected against MC903-induced AD-like dermatitis via the NF-κB/MAPK pathway in a STAT3-dependent manner. These results provide new insights into the complex relationship between surfactants and AD, and offer important clues for potential strategies to develop novel surfactant formulations for AD patients.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Langan, S. M., Irvine, A. D. & Weidinger, S. Atopic dermatitis. Lancet396, 345–360. https://doi.org/10.1016/S0140-6736(20)31286-1 (2020).
Mao, D. et al. Prevalence and risk factors of atopic dermatitis in Chinese adults: A nationwide population-based cross-sectional study. Chin. Med. J. (Engl.)136, 604–606. https://doi.org/10.1097/CM9.0000000000002560 (2023).
Corazza, M. et al. Surfactants, skin cleansing protagonists. J. Eur. Acad. Dermatol. Venereol.24, 1–6. https://doi.org/10.1111/j.1468-3083.2009.03349.x (2010).
Lunjani, N. et al. Environment-dependent alterations of immune mediators in urban and rural South African children with atopic dermatitis. Allergy77, 569–581. https://doi.org/10.1111/all.14974 (2022).
Lee, J. S., Kim, J. M., Seok, J. & Kim, B. J. Correlation between socio-economic status and atopic dermatitis in Korean adults: The Korea National Health and Nutrition Examination Survey (2007–2014). J. Eur. Acad. Dermatol. Venereol.31, 1509–1515. https://doi.org/10.1111/jdv.14343 (2017).
Ofenloch, R. F. et al. Socioeconomic status and the prevalence of skin and atopic diseases in five European countries. Acta Derm. Venereol.99, 309–314. https://doi.org/10.2340/00015555-3082 (2019).
Lambrecht, B. N. & Hammad, H. The immunology of the allergy epidemic and the hygiene hypothesis. Nat. Immunol.18, 1076–1083. https://doi.org/10.1038/ni.3829 (2017).
Celebi Sozener, Z. et al. Epithelial barrier hypothesis: Effect of the external exposome on the microbiome and epithelial barriers in allergic disease. Allergy77, 1418–1449. https://doi.org/10.1111/all.15240 (2022).
Becker, L. C. et al. Amended safety assessment of dodecylbenzenesulfonate, decylbenzenesulfonate, and tridecylbenzenesulfonate salts as used in cosmetics. Int. J. Toxicol. 29, 288S–305. https://doi.org/10.1177/1091581810386124 (2010).
Mu, Z., Liu, X., Zhao, Y. & Zhang, J. Cytotoxic effects of sodium dodecyl benzene sulfonate on human keratinocytes are not associated with proinflammatory cytokines expression. Chin. Med. J. (Engl.)127, 3777–3781 (2014).
Reinartz, J. et al. Activation of the plasminogen activator system in a keratinocyte cell line (HaCaT) by alkyl sulfates. Biomed. Biochim. Acta. 50, 743–748 (1991).
Kumari, V., Babina, M., Hazzan, T. & Worm, M. Thymic stromal lymphopoietin induction by skin irritation is independent of tumour necrosis factor-α, but supported by interleukin-1. Br. J. Dermatol. 172, 951–960. https://doi.org/10.1111/bjd.13465 (2015).
Li, M. et al. Induction of thymic stromal lymphopoietin expression in keratinocytes is necessary for generating an atopic dermatitis upon application of the active vitamin D3 analogue MC903 on mouse skin. J. Invest. Dermatol.129, 498–502. https://doi.org/10.1038/jid.2008.232 (2009).
Li, M. et al. Topical vitamin D3 and low-calcemic analogs induce thymic stromal lymphopoietin in mouse keratinocytes and trigger an atopic dermatitis. Proc. Natl. Acad. Sci. U. S. A.103, 11736–11741. https://doi.org/10.1073/pnas.0604575103 (2006).
Liu, Y. et al. PD-L1 compartmentalization suppresses tumorigenesis and overcomes immunocheckpoint therapy resistance in mice via histone macroH2A1. J. Clin. Invest. 134:e181314. https://doi.org/10.1172/JCI181314
Gao, W. et al. Network pharmacology and experimental verification to investigate the mechanism of Isoliquiritigenin for the treatment of Alzheimer’s disease. Sci. Rep.15, 4379. https://doi.org/10.1038/s41598-025-88542-y (2025).
Weidinger, S. et al. Atopic dermatitis. Nat. Rev. Dis. Primershttps://doi.org/10.1038/s41572-018-0001-z (2018).
Feld, M. et al. The pruritus- and TH2-associated cytokine IL-31 promotes growth of sensory nerves. J. Allergy Clin. Immunol. 138, 500–508e24. https://doi.org/10.1016/j.jaci.2016.02.020 (2016).
Wilson, S. R. et al. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell 155, 285–295. https://doi.org/10.1016/j.cell.2013.08.057 (2013).
Kasperkiewicz, M., Schmidt, E., Ludwig, R. J. & Zillikens, D. Targeting IgE antibodies by immunoadsorption in atopic dermatitis. Front. Immunol.9, 254. https://doi.org/10.3389/fimmu.2018.00254 (2018).
Schuler, C. F. et al. Novel insights into atopic dermatitis. J. Allergy Clin. Immunol. 151, 1145–1154. https://doi.org/10.1016/j.jaci.2022.10.023 (2023).
Liu, Y.-J. Thymic stromal lymphopoietin and OX40 ligand pathway in the initiation of dendritic cell-mediated allergic inflammation. J. Allergy Clin. Immunol.120(2), 238–244. https://doi.org/10.1016/j.jaci.2007.06.004 (2007).
Soumelis, V. et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol.3, 673–680. https://doi.org/10.1038/ni805 (2002).
Leyva-Castillo, J. M. et al. Skin thymic stromal lymphopoietin initiates Th2 responses through an orchestrated immune cascade. Nat. Commun.4, 2847. https://doi.org/10.1038/ncomms3847 (2013).
Ito, T. et al. TSLP-activated dendritic cells induce an inflammatory T helper type 2 cell response through OX40 ligand. J. Exp. Med. 202, 1213–1223. https://doi.org/10.1084/jem.20051135 (2005).
Han, E.-J. et al. (-)-Loliolide isolated from Sargassum horneri suppressed oxidative stress and inflammation by activating Nrf2/HO-1 signaling in IFN-γ/TNF-α-stimulated HaCaT keratinocytes. Antioxidants10, 856. https://doi.org/10.3390/antiox10060856 (2021).
Cho, S.-H. et al. Eckol from Ecklonia cava ameliorates TNF-α/IFN-γ-induced inflammatory responses via regulating MAPKs and NF-κB signaling pathway in HaCaT cells. Int. Immunopharmacol.82, 106146. https://doi.org/10.1016/j.intimp.2019.106146 (2020).
Bieber, T. Atopic dermatitis. N. Engl. J. Med.358, 1483–1494. https://doi.org/10.1056/NEJMra074081 (2008).
Dubin, C., Del Duca, E. & Guttman-Yassky, E. The IL-4, IL-13 and IL-31 pathways in atopic dermatitis. Expert Rev. Clin. Immunol.17, 835–852. https://doi.org/10.1080/1744666X.2021.1940962 (2021).
Neurath, M. F. & Finotto, S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine Growth Factor. Rev. 22, 83–89. https://doi.org/10.1016/j.cytogfr.2011.02.003 (2011).
Piconese, S. et al. Mast cells counteract regulatory T-cell suppression through interleukin-6 and OX40/OX40L axis toward Th17-cell differentiation. Blood 114, 2639–2648. https://doi.org/10.1182/blood-2009-05-220004 (2009).
Doganci, A. et al. The IL-6R alpha chain controls lung CD4 + CD25 + Treg development and function during allergic airway inflammation in vivo. J. Clin. Invest. 115, 313–325. https://doi.org/10.1172/JCI22433 (2005).
Yoshida, H., Hashizume, M., Suzuki, M. & Mihara, M. Anti-IL-6 receptor antibody suppressed T cell activation by inhibiting IL-2 production and inducing regulatory T cells. Eur. J. Pharmacol. 634, 178–183. https://doi.org/10.1016/j.ejphar.2010.02.026 (2010).
Savinko, T. et al. IL-33 and ST2 in atopic dermatitis: Expression profiles and modulation by triggering factors. J. Invest. Dermatol.132, 1392–1400. https://doi.org/10.1038/jid.2011.446 (2012).
Imai, Y. Interleukin-33 in atopic dermatitis. J. Dermatol. Sci. 96, 2–7. https://doi.org/10.1016/j.jdermsci.2019.08.006 (2019).
Kakinuma, T. et al. Thymus and activation-regulated chemokine in atopic dermatitis: Serum thymus and activation-regulated chemokine level is closely related with disease activity. J. Allergy Clin. Immunol.107, 535–541. https://doi.org/10.1067/mai.2001.113237 (2001).
Halling, A-S. et al. Skin TARC/CCL17 increase precedes the development of childhood atopic dermatitis. J. Allergy Clin. Immunol. 151, 1550–1557e6. https://doi.org/10.1016/j.jaci.2022.11.023 (2023).
Huang, I.-H., Chung, W.-H., Wu, P.-C. & Chen, C.-B. Jak-stat signaling pathway in the pathogenesis of atopic dermatitis: An updated review. Front. Immunol.13, 1068260. https://doi.org/10.3389/fimmu.2022.1068260 (2022).
Guttman-Yassky, E. et al. The role of Janus kinase signaling in the pathology of atopic dermatitis. J. Allergy Clin. Immunol.152, 1394–1404. https://doi.org/10.1016/j.jaci.2023.07.010 (2023).
Kim, W.-H. et al. Beneficial effects of melittin on ovalbumin-induced atopic dermatitis in mouse. Sci. Rep.7, 17679. https://doi.org/10.1038/s41598-017-17873-2 (2017).
Du, H.-Y. et al. The expression and regulation of interleukin-33 in human epidermal keratinocytes: A new mediator of atopic dermatitis and its possible signaling pathway. J. Interferon Cytokine Res.36, 552–562. https://doi.org/10.1089/jir.2015.0159 (2016).
Fan, H.-J. et al. Anti-inflammatory and immune response regulation of Si-Ni-San in 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin dysfunction. J. Ethnopharmacol.222, 1–10. https://doi.org/10.1016/j.jep.2018.04.032 (2018).
Park, H.-J. et al. Corticotropin-releasing hormone (CRH) downregulates interleukin-18 expression in human HaCaT keratinocytes by activation of p38 mitogen-activated protein kinase (MAPK) pathway. J. Invest. Dermatol.124, 751–755. https://doi.org/10.1111/j.0022-202X.2005.23656.x (2005).
Lim, J.-M. et al. Effect of peiminine on DNCB-induced atopic dermatitis by inhibiting inflammatory cytokine expression in vivo and in vitro. Int. Immunopharmacol.56, 135–142. https://doi.org/10.1016/j.intimp.2018.01.025 (2018).
Jayasinghe, A. M. K. et al. Fucoidan isolated from Sargassum confusum suppresses inflammatory responses and oxidative stress in TNF-α/IFN-γ- stimulated HaCaT keratinocytes by activating Nrf2/HO-1 signaling pathway. Mar. Drugs20, 117. https://doi.org/10.3390/md20020117 (2022).
Bode, J. G., Ehlting, C. & Häussinger, D. The macrophage response towards LPS and its control through the p38(MAPK)-STAT3 axis. Cell. Signal.24, 1185–1194. https://doi.org/10.1016/j.cellsig.2012.01.018 (2012).
Jiang, Y. et al. Bergapten ameliorates combined allergic rhinitis and asthma syndrome after PM2.5 exposure by balancing Treg/Th17 expression and suppressing STAT3 and MAPK activation in a mouse model. Biomed. Pharmacother.164, 114959. https://doi.org/10.1016/j.biopha.2023.114959 (2023).
Lee, C. & Cheung, S. T. STAT3: An emerging therapeutic target for hepatocellular carcinoma. Cancers (Basel)11, 1646. https://doi.org/10.3390/cancers11111646 (2019).
Sims, J. T. et al. Imatinib reverses doxorubicin resistance by affecting activation of STAT3-dependent NF-κB and HSP27/p38/AKT pathways and by inhibiting ABCB1. PLoS One8, e55509. https://doi.org/10.1371/journal.pone.0055509 (2013).
Wang, S-H. et al. Inhibition of bacterial adherence to biomaterials by coating antimicrobial peptides with anionic surfactant. Colloids Surf. B Biointerfaces. 196, 111364. https://doi.org/10.1016/j.colsurfb.2020.111364 (2020).
Han, W. et al. Ecotoxicity and micellization behavior of anionic surfactant sodium dodecylbenzene sulfonate (SDBS) and its mixtures with nonionic surfactant fatty alcohol-polyoxyethylene ether (AEO). Aquat. Toxicol.216, 105313. https://doi.org/10.1016/j.aquatox.2019.105313 (2019).
Zhang, P., Chen, Y. & Zhou, Q. Effect of surfactant on hydrolysis products accumulation and short-chain fatty acids (SCFA) production during mesophilic and thermophilic fermentation of waste activated sludge: Kinetic studies. Bioresour. Technol.101, 6902–6909. https://doi.org/10.1016/j.biortech.2010.03.124 (2010).
Xiao, X. et al. The role of short-chain fatty acids in inflammatory skin diseases. Front. Microbiol.13, 1083432. https://doi.org/10.3389/fmicb.2022.1083432 (2023).
Sousa, B., Domingues, I. & Nunes, B. Biological responses in Danio rerio by the disinfectant SDBS in SARS-CoV-2 pandemic. Environ. Toxicol. Pharmacol.107, 104431. https://doi.org/10.1016/j.etap.2024.104431 (2024).
Funding
Incubating project of The General Hospital of the People’s Liberation Army (22QNFC117).
Author information
Authors and Affiliations
Contributions
H.Z., Z.M. and J.Z. designed the experiments. S.Z. and D.Z. performed data curation. Y.W. and Y.Wg. carried out formal analysis. H.Z., Z.L. and X.Z. conducted the investigation. H.Z. and Z.M. developed the methodology. B.L., X.L. and C.L. managed the project. S.Z., X.Z. and W.X. provided resources. Y.Z. managed software. C.Z., C.L. and J.Z. supervised the work. Y.W., D.Z. and W.X. validated the results. Z.L. and Y.Z. handled the visualization. H.Z. and Z.L. wrote the original draft. C.Z., C.L. and J.Z. reviewed and edited the manuscript. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, H., Li, Z., Mu, Z. et al. The anionic surfactant Sodium Dodecyl Benzene Sulfonate alleviates inflammatory responses in atopic dermatitis via NFκB/MAPK pathways in a STAT3-dependent manner. Sci Rep 15, 28019 (2025). https://doi.org/10.1038/s41598-025-12776-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-12776-z
