
Abstract
Circular RNAs (circRNAs) are highly stable and abundant in tumor-derived exosomes, where they often function as microRNA (miRNA) sponges to regulate gene expression and biological processes. Previous research showed that Transducer of ERBB2.1 (TOB1) overexpression induces autophagy and suppresses gastric cancer progression through exosome secretion. This study investigates the relationship between TOB1, exosomal circRNAs, and their target miRNAs in gastric cancer. RNA sequencing and bioinformatics analyses identified differentially expressed circRNAs and miRNAs in exosomes from gastric cancer cells. qRT-PCR revealed that hsa_circ_0008719 was downregulated in gastric cancer tissues compared to normal tissues. Further analysis showed that TOB1 upregulates hsa_circ_0008719 and downregulates miR-3615 in gastric cancer cells and their exosomes. Functional assays, including lentiviral infection, Cell Counting Kit-8 (CCK-8), colony formation, and Western blot, demonstrated that exosomal hsa_circ_0008719 inhibits proliferation and promotes autophagy in gastric cancer cells. Differential expression analysis identified 87 upregulated circRNAs and 1 upregulated miRNA, as well as 2 downregulated circRNAs and 8 downregulated miRNAs in exosomes from TOB1-overexpressing AGS cells. Twelve novel circRNAs were discovered. GO and KEGG analyses linked the host gene of hsa_circ_0008719, AKT2, to gastric cancer and autophagy pathways. hsa_circ_0008719 was found to localize in the cytoplasm, where it binds to miR-3615. These findings highlight the TOB1-hsa_circ_0008719-miR-3615 axis in gastric cancer, showing that exosomal hsa_circ_0008719 suppresses cancer progression by enhancing autophagy and reducing proliferation. This study provides insights into the role of TOB1-mediated exosomal circRNAs in gastric cancer regulation.
Similar content being viewed by others

Introduction
Gastric cancer still ranks the fifth most common malignant cancer worldwide, which continues to be the fourth leading cause of cancer-related mortality1. Genetic, epigenetic and external environmental factors can induce the development and progression of gastric cancer2. Transducer of ERBB2.1 (TOB1) is a tumor suppressor gene, similar to many others, the inactivation of TOB1 has been reported to contribute to the development of cancer in the lung, liver, and lymph nodes3. We found elevated levels of phosphorylated TOB1 (p-TOB1) in gastric cancer cells, with p-TOB1 representing one of its inactive forms4. In addition, increased p-TOB1 also suggests that lymph node-negative breast cancer has a poor prognosis5. In earlier studies, we revealed TOB1 as a tumor suppressor in gastric cancer and demonstrated that TOB1 nuclear retention is critical for its antiproliferative activity6,7,8. Down-regulation of TOB1 expression is associated with shorter survival in gastric cancer patients, thus TOB1 may serve as a potential novel marker for predicting the prognosis9. Recently, we found that TOB1 can induce autophagy10 by secreting exosomes11 in gastric cancer cells.
Exosomes are nanosized vesicles of cellular origin, ranging from 30 to 150 nm in diameter, that transport nucleic acids, proteins, lipids, and other bioactive substances to recipient cells, thereby regulating physiological and pathological processes within the body12. Among the transported cargoes, the producing cell and its homeostatic condition influence the variations in non-coding RNAs (ncRNAs)13. RNA sequencing analysis revealed that circular RNAs (circRNAs) are enriched in exosomes compared to their originating cells, and that the sorting of circRNAs into exosomes may be regulated by the levels of associated miRNAs14. CircRNAs are covalently closed loop structures formed by back-splicing of mRNA without 5’ caps and 3’ tails, which have been classified as a distinct subset of non-coding RNAs15. Existing evidence indicates that circRNAs can act as microRNA sponges, thereby impairing the activity of miRNAs16. Autophagy is the primary mechanism by which autophagic vesicles are generated to engulf cellular targets and deliver various cellular cargoes to lysosomes for degradation and recycling.
Autophagy plays a crucial protective role in various diseases; however, it appears to have an opposing effect in the early prevention and metastasis of cancer17,18. The interactions between exosome and autophagy pathways are complex, as autophagy and exocytosis can simultaneously mitigate cellular stress19. Furthermore, exosomes can exhibit cytoprotective effects by inducing intracellular autophagy, while autophagy also regulates both exosome biogenesis and degradation20. Thus, exosomal circRNAs and miRNAs may reflect the autophagy of gastric cancer cells in an environment characterized by TOB1 overexpression. In the present study, we conducted an analysis of specific exosomal circRNAs and their target miRNAs, as well as their correlation with TOB1 in gastric cancer. We selected hsa_circ_0008719 for our investigation, which is produced by the AKT2 gene. Our findings revealed that hsa_circ_0008719 is positively correlated with TOB1 and downregulated in gastric cancer tissues. Furthermore, we verified that hsa_circ_0008719 can bind to miR-3615 and negatively regulate its expression. Functional studies demonstrated that exosomal hsa_circ_0008719 can reduce proliferation of gastric cancer cells while enhancing autophagy. Collectively, this study provides a novel mechanism underlying gastric cancer progression and elucidates a potential target for the prevention and treatment of gastric cancer.
Materials and methods
Cell lines and clinical tissues
The gastric cancer cell line AGS was procured from the American Type Culture Collection (ATCC, Manassas, VA, USA) cultured in F-12 K medium (Gibco BRL, USA) supplemented with 10% fetal bovine serum (absin, abs972, China). The gastric cancer cell line HGC-27 was obtained from the Cell Resources Center of Shanghai Life Sciences, Chinese Academy of Sciences (Shanghai, China), and cultured in RPMI-1640 medium (Gibco BRL, USA) supplemented with 10% fetal bovine serum. In our previous study, we successfully established gastric cancer AGS cells overexpressing TOB1 (AGS-TOB1) along with a control group (AGS-Vector)10, and HGC-27 cells with TOB1 knockout (HGC-27-TOB1-6E12) was generated using CRISPR/Cas9 genome editing technology by Cyagen (Suzhou, China). HEK-2937T cell line was purchased from Shanghai Institute of Cell Biology, Chinese Academy of Sciences, and cultured in DMEM medium (Gibco BRL, USA) supplemented with 10% fetal bovine serum. All of these cell lines were maintained in a 37 °C incubator with 5% CO2. Tumor and paratumor tissues were collected from gastric cancer patients (n = 52) admitted to the Second Affiliated Hospital of Harbin Medical University and the Affiliated Cancer Hospital between 2018 and 2023. Samples were rapidly collected and transported in liquid nitrogen within 30 min post-ex vivo, then stored at − 80 °C.
Ethics approval and consent to participate
This experiment was conducted in accordance with the Declaration of Helsinki. The Ethics Committee of the Second Affiliated Hospital of Harbin Medical University approved the study, and informed consent to participate was obtained from all participants (Approval Number: YJSKY2022-425).
Exosome isolation
Conditioned medium (15 ml) was collected from AGS-TOB1, AGS-Vector, and HGC-27 cells which were respectively cultured in F-12 K medium and RPMI-1640 medium supplemented with 10% exosome-depleted fetal bovine serum (absin, abs993, China) for 48 h. Following an initial centrifugation at 2000 × g for 30 min, the supernatant underwent additional concentration at 10,000 × g for 30 min to eliminate cells and debris. The supernatant was then subjected to further concentration via centrifugation at 100,000 × g for 90 min at 4 °C using an ultrafiltration tube with a 0.22 μm pore filter to extract the exosomes. The extracted exosomes were resuspended in phosphate-buffered saline (PBS) (Seven, SW132-01, Beijing) and stored at − 80 °C until needed.
Transmission electron microscopy (TEM)
PBS-suspended exosomes were deposited onto carbon-coated copper grids at room temperature for 5 min. The staining solution containing phosphotungstic acid was applied for 2 min before being absorbed with filter paper. The resulting images were observed and captured using TEM (Hitachi, Japan).
Nanoparticle tracking analysis (NTA)
Exosomes were introduced into the ZetaView PMX 110 NTA analyzer with appropriate dilutions of 1× PBS. The particle size and concentration of the samples were assessed by selecting 11 random positions within the analyzer. Measurement data were analyzed using ZetaView software version 8.04.02. System calibration was performed with 110 nm polystyrene particles, while temperature was monitored between 23 °C and 30 °C throughout the detection procedure.
Western blot
Total protein was extracted from exosomes resuspended in RIPA buffer (Applygen, Beijing, China). The protein concentration was determined using a Micro BCA Protein Assay Kit (Applygen, Beijing, China). Subsequently, the proteins were subjected to SDS-PAGE (NCM Biotech, P2013, China) and transferred to PVDF membranes (Immobilon™-PSQ Membranes, ISEQ00005, USA). The membranes were blocked with 5% nonfat milk for 2 h and incubated with primary antibodies against TSG101 (Abcam, ab125011, 1:1000; Britain), CD9 (Immunoway, YT0782; 1:1000; USA), LC3B (Proteintech, 18725-1-AP; 1:1000; USA), and GAPDH (Beyotime, AF2823; 1:1000; China) at 4 °C for 12 h. Following this incubation period, the PVDF membranes were treated with secondary antibodies (Beyotime A0192 and A0208; both at 1:1000; China) for one hour. Blots were imaged using a Tanon Imaging System (Tanon-5200, Shanghai, China), and density values were analyzed using ImageJ software.
Identification and quantification of Exosomal RNAs
The original images and data analyzed through high-throughput RNA sequencing were transformed into sequenced reads (Raw Data or Raw Reads). The statistical power of this experiment was calculated using the Illumina HiSeq™ 2500 at Qphred = − 10log10(e). Subsequently, the CircRNA Raw Reads were filtered to obtain Clean Reads, with the genome of these filtered Clean Reads being processed using HISAT2 (version 2.0.4) with parameters --dta --rna-strandness RF. Find_circ (version 1.1) was employed to identify circRNAs; its principle involves extracting a 20 nt anchor sequence from each end of reads that do not align with the reference sequence, followed by realigning each pair of anchor sequences with the reference sequence. If the 5’ end of an anchor sequence aligns with the reference sequence (with A3 and A4 indicating start and termination sites), while its 3’ end aligns upstream of this site (with A1 and A2 indicating start and termination sites), and if there is a splicing site (GT-AG) between A2 and A3 in the reference sequence, then this read is considered a candidate circRNA. The sRNA Raw Reads were filtered into Clean Reads by removing low-quality reads, those with a ratio of N bases greater than 10%, contamination at the 5’ joint, absence of a 3’ joint sequence, insert fragments, and polyA/T/G/C sequences. Bowtie version 0.12.9 was used for statistical analysis with parameters -v 0 -k 1 to analyze small RNA distribution across the reference sequence after length screening; subsequently aligning it within miRBase’s specified range to acquire secondary structures for confirmed miRNAs based on their sequences, lengths, and frequencies in each sample. Potential rRNA, tRNA, snRNA, snoRNA, YRNA sequences as well as repeat sequences were removed; sRNAs were aligned against exons and introns of total mRNA to eliminate short reads originating from introns. Novel miRNAs were identified using miREvo (version 1.1) and mirdeep2_0_0_5 software packages; their respective statistical powers included parameters -i-r-M-m-k-p10-g50000 for miREvo and quantifier.pl-p-m-r-y-g0-T10 for mirdeep.
Analysis of differentially expressed circrna and MiRNA
CircRNAs are derived from the splicing of exons or introns, and circRNA expression measurements were normalized to TPM, calculated as follows: expression normalization is performed using the formula (readCount × 1,000,000)/libsize (where libsize represents the sum of circRNA read counts across samples). Furthermore, based on their chromosomal locations, all differentially expressed circRNAs were classified. These circRNAs are distributed across 10 chromosomes, including the X chromosome. The RIES (Internal Ribosome Entry Site) is an important regulatory element in 5′ UTR cap-independent translation that mediates the translation of polypeptides and proteins. IRESfinder was utilized to predict whether a given circRNA contains an IRES. The differential expression data for circRNAs and miRNAs were obtained from read count data identified during circRNA and miRNA expression level analyses. Analysis of samples with biological replicates was conducted using DESeq2 (version 1.10.1) at a significance threshold of padj < 0.05, which is based on a negative binomial distribution.
Predication of target MiRNAs of circrnas
To predict the targeted miRNAs of circRNAs, binding sites were identified using MiRanda (version 3.3a) and TargetFinder (version 1.0) with parameters -sc 140 -en -10 -scale 4 –strict and -c 6 -t 6 -p table. A higher score for binding sites indicates a greater probability of targeting; conversely, a lower energy value suggests a more stable binding site. Differentially expressed circRNAs and miRNAs were also annotated with Gene Ontology (GO) terms (http://www.geneontology.org/) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways (https://www.genome.jp/kegg/pathway.html).
Analysis of the correlation of TOB1 and circrnas
To investigate the potential correlation between TOB1 expression and AKT2, which serves as the host gene for the up-regulated hsa_circ_0008719, we utilized GEPIA (gepia.cancer-pku.cn) and CCLE (https://sites.broadinstitute.org/ccle) to evaluate gene expression correlations, including expression patterns across different gastric cell lines.
qRT-PCR
Total RNA was extracted from cells and tissues using TRIzol Reagent (Ambion, 420811, USA). Total exosomal RNAs were obtained with the miRNeasy Serum/Plasma Kit (Qiagen, 217184, USA). The quantification of RNAs was performed using a NanoDrop 2000 spectrophotometer (Thermo Scientific, USA). The Transcription First Strand cDNA Synthesis Kit (Roche, 67112920, Switzerland) was utilized for the reverse transcription of total RNA into cDNA. For miRNAs, the Reverse Transcription Kit (RiboBio, Guangzhou, C11027-2, China) was employed to convert total RNA into cDNA. FastStart Essential DNA Green Master (ROX) (Roche, 71795000, Switzerland) was used in the quantitative PCR system (LightCycler 480, Roche, Switzerland). The expression levels of circRNAs and miRNAs were normalized to internal references ß-actin (GENERAL BIOL, Anhui, China) and U6 (RiboBio, Guangzhou, China), respectively. Primers used in this study are listed in Supplementary Table 1. The 2−△△CT value reflects relative expression levels. All validation assays were conducted in triplicate across a minimum of three independent experiments.
Validation of circrna specificity
A 2% agarose gel was used to separate PCR products amplified with hsa_circ_0008719 primers. Sanger sequencing was carried out to validate the back-splicing junction site of hsa_circ_0008719 (RuiBiotech, Beijing, China).
RNase R resistance assay
The RNA extracted from AGS or HGC-27 cells underwent treatment with RNase R (1 U/µg, Bio Search), followed by a 10-minute incubation at 37 °C. The processed RNAs were then mixed with the Transcriptor First Strand cDNA Synthesis Kit to synthesize cDNA and subsequently analyzed by quantitative reverse transcription PCR (qRT-PCR).
Nuclear and cytoplasmic extraction
1 × 106 AGS or HGC-27 cells were harvested and washed with pre-chilled PBS. The suspension was centrifuged at 4 °C for 5 min at 500 g. The supernatant, representing the cytoplasmic fraction, was carefully aspirated away from the nuclear pellet. Cytoplasmic and nuclear RNA were separated and purified using the PARIS kit (Thermo Fisher, AM1921, USA) according to the manufacturer’s protocol.
Dual-luciferase reporter gene assay
To verify the binding site between circRNA and miRNA, pmiR-RB-Report™-hsa_circ_0008719-WT/MUT and miR-3615-mimic/NC (Ribobio, Guangzhou, China) were co-transfected into 293T cells. After 48 h, the luciferase activities of the cells were quantified using the Dual-Glo® Luciferase Assay System (Promega, E2920, USA).
Lentivirus infection
The lentivirus for overexpressing and knocking down hsa_circ_0008719 was obtained from Ribobio (Guangzhou, China). Following viral infection, AGS-Vector and AGS-TOB1 cells were selected in medium containing G418 at a concentration of 100 mg/mL to establish a cell line stably expressing hsa_circ_0008719. The cells were subsequently subjected to functional analyses.
CCK-8 assay
Cells were inoculated into 96-well plates at a density of 1,000 cells per well, with 5 replicate wells for each cell group. Ten microliters of CCK-8 reagent (Seven Biotech, SC119, China) was added to each well, and after a 2-hour incubation period, the absorbance at 450 nm was measured daily for 6 days to construct the growth curve.
Colony formation assay
Cells were inoculated into 6-well plates at a density of 400 cells per well, with 3 replicate wells established for each group. The cells were incubated in 5% CO2 at 37 °C for 15 days, followed by washing with PBS and fixation using fixative solution (Seven, SI101-01, Beijing) for 15 min. Finally, the cells were stained with crystal violet (Beyotime, C0121, Shanghai) for an additional 15 min.
Statistical analysis
Graphs were generated using GraphPad Prism 8.0 and Adobe Illustrator 2021 software, while analyses were conducted with SPSS Statistics 17.0 software. Normally distributed samples were analyzed using a paired t-test, whereas non-normally distributed samples were assessed using the Wilcoxon rank-sum test. The correlation between hsa_circ_0008719 in gastric cancer tissues and the clinicopathological characteristics of gastric cancer patients was evaluated through chi-square test correlation analysis. Results were considered statistically significant at p < 0.05 (*p < 0.05, **p < 0.01, ***p < 0.001).
Results
Identification of exosomes derived from gastric cancer cells with overexpressing of TOB1
Exosomes were extracted from the culture medium of gastric cancer cell lines and categorized into three groups: exo-AGS-TOB1 (experimental group) with ectopic overexpression of TOB1, exo-AGS-Vector (control group) with low endogenous TOB1 expression, and exo-HGC-27 exhibiting high endogenous TOB1 expression. The morphological structure of the exosomes was observed using TEM (Fig. 1A–C), revealing approximately spherical lipid bilayer vesicles with diameters ranging from 30 to 150 nm. The transparency of these vesicles varied based on their internal contents. Size distribution analysis of the exosomes was performed using NTA. The highest peak for purified exosomes was approximately 100 nm in diameter (Fig. 1D–F), which falls within the expected range for exosome diameters. Based on NTA analysis of isolated nanoparticles, purified exosomal particles displayed irregular Brownian motion. Finally, as demonstrated by western blotting analysis (Fig. 1G), marker proteins TSG101 and CD9 were enriched in cell-derived exosomes.

Identification of exosomes released by gastric cancer cells. (A–C) Morphological structures of exo-AGS-TOB1, exo-AGS-Vector, and exo-HGC-27 (scale bar = 100 nm). (D–F) Peak diameters and Brownian motion characteristics of purified exosomal particles from exo-AGS-TOB1, exo-AGS-Vector, and exo-HGC-27. (G) Expression levels of the exosomal markers TSG101 and CD9 proteins as determined by western blot analysis.
Differentially expressed circrnas and miRNAs in exosomes of gastric cancer cells with overexpressing of TOB1
Compared to exo-AGS-Vector (control), 89 circRNAs and 9 miRNAs were differentially expressed in exo-AGS-TOB1 (TOB1 overexpression). Among these, 87 circRNAs were significantly upregulated and 2 circRNAs were significantly downregulated in exo-AGS-TOB1 (supplementary Table 2), while one miRNA was significantly upregulated and 8 miRNAs were significantly downregulated (Table 1). Of the 89 differentially expressed circRNAs, 12 were identified as novel circRNAs that have not been functionally annotated in current circRNA databases (Table 2). Among these novel circRNAs, 10 exhibited significant upregulation, whereas 2 showed significant downregulation. Detailed information regarding all of these specific circRNAs, including their source genes and chromosomal locations, is presented in supplementary Table 3.
Hierarchical clustering analysis revealed distinct expression patterns of circRNAs (Fig. 2A) and miRNAs (Fig. 2B) between the two groups. The volcano plot provided a more intuitive representation of significantly differentially expressed circRNAs (Fig. 2C) and miRNAs (Fig. 2D). Based on the circRNA information databases Find_circ (version 1.1) and circBase (http://www.circbase.org/), it was shown that nearly half of the significantly differentially expressed circRNAs are transcribed from mRNA exons. According to the results of GO and KEGG pathway analyses for the source gene AKT2, hsa_circ_0008719, which is upregulated with TOB1 overexpression, was selected for subsequent analysis. It was found that exosomal hsa_circ_0008719 is spliced from the AKT2 gene (intron10) located at chr19:40255269–40,256,925(-), with a final length of 371 nt (Table 3). Interestingly, AKT2, as the host gene for upregulated hsa_circ_0008719, was determined to be primarily associated with gastric cancer pathways and autophagy-related pathways based on KEGG PATHWAY Database analysis21,22,23(Fig. 2E).

Differentially expressed circRNAs and miRNAs in exosomes of TOB1 overexpressing gastric cancer cells. (A, B) The overall hierarchical clustering diagram was generated based on the log10 (TPM + 1) values. Red, upregulated circRNAs and miRNAs; blue, downregulated circRNAs and miRNAs. (C) Volcano plot revealing altered circRNAs. Green, downregulated circRNAs; red, upregulated circRNAs. (D) Volcano plot revealing altered miRNAs. Green, downregulated miRNAs; red, upregulated miRNAs. (E) The mainly KEGG pathways of the host gene of hsa_circ_0008719 (by Figdraw).
Functional analysis of hsa_circ_0008719
In this study, upregulated and downregulated circRNAs were considered potential tumor suppressors and oncogenes associated with TOB1 overexpression, respectively. To gain a deeper understanding of the functions of these circRNAs, the host genes of differentially expressed circRNAs were analyzed using GO and KEGG pathway analyses. The most significantly enriched GO terms for circRNAs in the biological process, cellular component, and molecular function categories included organelle organization, non-membrane-bounded organelle, and protein binding, respectively (Fig. 3A). In the analysis of the top 20 KEGG pathways, the host genes were primarily linked to two gastric cancer-related pathways: RNA degradation and lysine degradation (Fig. 3B; Table 4). GO analysis indicated that AKT2 was highly correlated with cell proliferation and migration. According to CC classification results, hsa_circ_0008719 may play a functional role in the cytoplasm (supplementary Table 4).
With TOB1 overexpression, it was preliminarily hypothesized that the upregulated hsa_circ_0008719 derived from AKT2 may have a positive correlation with TOB1. Consequently, a Pearson correlation test was conducted using the GEPIA web server (http://gepia.cancer-pku.cn/) to examine the relationship between AKT2 and TOB1 in 408 TCGA (STAD) tumor samples, 36 TCGA (STAD) normal tissues, and 175 GTEx (stomach) samples. The results indicated that the expression level of TOB1 is positively correlated with the expression level of AKT2 (R = 0.13, **P = 0.0013) (Fig. 3C). A similar conclusion was also supported by data from the Cancer Cell Line Encyclopedia (CCLE) (https://sites.broadinstitute.org/ccle), which includes gene expression profiles, chromosomal copy number variations, and massively parallel sequencing data from 1019 human cancer cell lines (Fig. 3D). The expression of AKT2 in AGS and HGC-27 cell lines was comparatively higher than in other stomach adenocarcinoma cell lines. Furthermore, TOB1 knockdown enhances the cytotoxic effects of SYMPK and AKT2 in gastric cancer cell lines (Fig. 3E).

Functional analysis of hsa_circ_0008719. (A) Bar plot showing circRNAs GO analysis. (B) Scatter plot of circRNAs KEGG enrichment analysis. The rich factor shows the proportion of enriched genes in the entire pathway’s gene pool. Gene number represents the number of genes enriched in this pathway. (C) The expression levels of TOB1 and AKT2 in GEPIA. (D) The expression levels of TOB1 and AKT2 in CCLE. (E) The expression levels of TOB1 knockdown and AKT2 in CCLE.
Hsa_circ_0008719 targets and interacts with miR-3615, leading to decreased expression in gastric cancer cells
The binding sites of differentially expressed circRNAs and miRNAs were predicted using miRanda (version 3.3a) and TargetFinder (version 1.0). Among these, only the downregulated miR-3615 was found to potentially bind to the upregulated hsa_circ_0008719 (Fig. 4A), suggesting their involvement in the autophagy process of gastric cancer. Subsequently, a dual-luciferase reporter assay was performed, demonstrating the combination between miR-3615 and the 3’ UTR of hsa_circ_0008719 (p < 0.01; Fig. 4B, C).
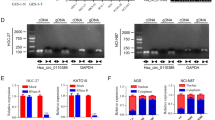
Additionally, Sanger sequencing was performed to confirm the junction site formed by the back-splicing event of hsa_circ_0008719 (Fig. 4D). It was observed that hsa_circ_0008719 is a circular RNA spliced from intron 10 of AKT2. qRT-PCR analysis confirmed that hsa_circ_0008719 exhibits resistance to RNase R, while AKT2 mRNA levels were significantly reduced after RNase R treatment (p < 0.01) (Fig. 4E). Furthermore, nuclear and cytoplasmic separation experiments demonstrated that hsa_circ_0008719 preferentially localizes within the cytoplasm in AGS and HGC-27 cells. These results suggest that hsa_circ_0008719 is a stable cytoplasmic circRNA acting as a molecular sponge for miR-3615 to exert its functions (Fig. 4F, G).
To investigate the potential function and clinical significance of hsa_circ_0008719 in gastric cancer, the expression levels of hsa_circ_0008719 were compared in 52 paired gastric cancer tissues using qRT-PCR. The results indicated that hsa_circ_0008719 was significantly downregulated in gastric cancer tissues (Fig. 4H) (p < 0.01). Furthermore, low expression of hsa_circ_0008719 was significantly associated with moderate to poor differentiation (p = 0.035) (Table 5).

Characteristics of hsa_circ_0008719. (A) Hsa_circ_0008719-miRNA regulatory network, with red indicating the target miR-3615. (B, C) Dual-luciferase reporter assays illustrates the interaction between miR-3615 and hsa_circ_0008719. (D) Hsa_circ_0008719 is generated by the circularization of intron 10 of the AKT2 gene; verification of the back-splicing junction site was performed using Sanger sequencing. (E) qRT-PCR analysis of hsa_circ_0008719 and linear AKT2 in AGS cells treated with RNase R. (F, G) qRT-PCR analysis of hsa_circ_0008719 expression level in different subcellular fractions of AGS and HGC-27 cells. (H) Expression level of hsa_circ_0008719 in gastric cancer tissues.
Verification of differentially expressed hsa_circ_0008719 and its target miR-3615 in exosomes and cells of gastric cancer
To support our hypothesis, a verification experiment was conducted for hsa_circ_0008719 and miR-3615, selected based on the aforementioned RNA sequencing data analysis. Using qRT-PCR, we determined the expression levels of TOB1, hsa_circ_0008719, and its target miR-3615 in exosomes derived from gastric cancer cell lines AGS and HGC-27 with exogenous and endogenous overexpression of TOB1, respectively. The results indicated that compared to exosomes from control cells (exo-AGS-Vector), TOB1 expression was significantly upregulated in exosomes from AGS cells overexpressing TOB1 (exo-AGS-TOB1) but downregulated in exosomes from HGC-27 cells (exo-HGC-27). Furthermore, hsa_circ_0008719 exhibited higher expression levels in exo-AGS-TOB1 than in exo-AGS-Vector, while lower levels were observed in exo-HGC-27 compared to exo-AGS-Vector. Conversely, miR-3615 displayed an opposite trend, it was downregulated in exo-AGS–TOB1 but upregulated in exo-HGC-27 relative to the control groups (Fig. 5A–C). Subsequently, we verified the expression levels of hsa_circ_0008719 and its target miR-3615 in AGS and HGC-27 cells. The results showed that TOB1 was significantly upregulated in AGS-TOB1 compared to AGS-Vector; similarly, HGC-27 cells also expressed higher levels of TOB1 than the control groups. Moreover, hsa_circ_0008719 exhibited elevated expression levels in AGS-TOB1 compared to AGS-Vector; conversely, miR-3615 demonstrated reduced expression levels under similar conditions. Likewise, hsa_circ_0008719 showed higher expression levels in HGC-27 cells than those seen with AGS-Vector while exhibiting lower expressions for miR-3615 within HGC-27 cells (Fig. 5D–F). These findings demonstrate that TOB1 expression is positively correlated with hsa_circ_0008719 expression while negatively correlating with miR-3615 expression.

The expression of TOB1, circRNAs and miRNA in exosomes and cells of gastric cancer. (A–C) The expression of TOB1, hsa_circ_0008719 and miR-3615 in exo- AGS-vector, exo-AGS-TOB1, and exo-HGC-27. (D–F) The expression of TOB1, hsa_circ_0008719 and miR-3615 in gastric cancer cell lines AGS-vector, AGS-TOB1, and HGC-27.
The hsa_circ_0008719 is delivered via exosomes to exhibit the proliferation and promote autophagy in gastric cancer cells
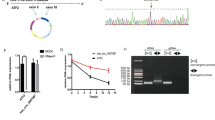
To better observe the delivery of hsa_circ_0008719 and its effects on gastric cancer progression, lentiviruses carrying hsa_circ_0008719 interference sequences (LV-RNAi-123305-2, LV-RNAi-123304-2) significantly knocked down hsa_circ_0008719 in AGS-TOB1 cells (Fig. 6A). Figure 6B shows a higher expression of miR-3615 in AGS-TOB1-si-1 and AGS-TOB1-si-2 with hsa_circ_0008719 knockdown compared to the control group (AGS-TOB1-si-NC), while there was no significant change in TOB1 expression (Fig. 6C). Next, we demonstrated that exosomal hsa_circ_0008719 was downregulated in the hsa_circ_0008719-depletion group (Fig. 6D). Additionally, we stably overexpressed has_circ_0008719 in AGS-Vector cells using a lentiviral vector (LV-94100–1) targeting hsa_circ_0008719 (AGS-Vec-8719), with an empty vector as a control (AGS-Vec-NC) (Fig. 6E). Notably, miR-3615 was downregulated in the hsa_circ_0008719-overexpression group; however, TOB1 also showed no significant alteration across groups (Fig. 6F, G), and level of hsa_circ_0008719 were upregulated in exosomes derived from AGS-Vec-8719 cells (Fig. 6H). Collectively, these results suggest that TOB1 can positively regulate the expression of hsa_circ_0008719 while negatively regulating miR-3615 expression; furthermore, it appears that hsa_circ_0008719 may competitively adsorb miR-3615 without affecting the expression of its upstream regulator TOB1. The consistency of hsa_circ_0008719 expression between cellular and exosomal contexts suggests that exosomes can serve as effective delivery vectors for transporting hsa_circ_0008719.
To further investigate the effects of exosomal hsa_circ_0008719 on the malignant phenotype of gastric cancer cells, we cultured HGC-27-TOB1-6E12 cells in exosome-free conditioned medium and purified exosomes from AGS-Vec-8719 and AGS-Vec-NC, respectively. The growth rate and colony numbers of cells co-cultured with exo-AGS-Vec-8719 were significantly lower than those in the control group, as assessed by CCK-8 (Fig. 6I) and colony formation assays (Fig. 6J). Western blot analysis revealed that LC3-I levels were significantly reduced, while LC3-II levels were significantly increased; both the LC3-II/LC3-I ratio and the LC3-II/GAPDH ratio showed significant increases in HGC-27-TOB1-6E12 treated with exo-AGS-Vec-8719 compared to the control group (Fig. 6K). These findings suggest that exosomal hsa_circ_0008719 can effectively inhibit the malignant phenotype and promote autophagy of gastric cancer cells.

Exosome delivery of hsa_circ_0008719 to decelerates gastric cancer cells growth while inducing autophagy. (A–D) The expression of hsa_circ_0008719, miR-3615, TOB1 and exo- hsa_circ_0008719 in AGS-TOB1 cells after knockdown hsa_circ_0008719. (E–H) The expression of hsa_circ_0008719, miR-3615, TOB1 and exo- hsa_circ_0008719 in AGS-Vector cells after overexpression hsa_circ_0008719. (I) The growth curves (J) and colony formation (K) and expression levels of the autophagy protein LC3 of HGC-27-TOB1-6E12 treated with exo- AGS-Vec-8719 and AGS-Vec-NC respectively.
Discussion
Currently, a substantial body of evidence suggests that exosomal circRNAs can regulate autophagy activation in gastric cancer24. Several studies indicate that autophagy and exosome biogenesis share common elements, proposing a link between these two processes25. Autophagy, as a degradative pathway, and exosome secretion play synergistic roles in maintaining cellular homeostasis26. The correlation between autophagy and exosomes was also demonstrated in Jiang’s study. Specifically, PTEN deficiency in cholangiocarcinoma impairs lysosomal biogenesis, thereby disrupting the autophagy-lysosome pathway essential for maintaining cellular homeostasis. Conversely, this deficiency enhances exosome secretion by cholangiocarcinoma cells27. And it is worth noting that the overexpression of TOB1 induced autophagy10 by secreting exosomes in our previous studies11. Therefore, it is essential to elucidate the intricate mechanisms underlying the interaction between exosomes and autophagy in tumor cells, including those in gastric cancer. CircRNAs are abundant within exosomes and exhibit greater stability than their parental linear RNAs. They are widely distributed and perform diverse roles in cellular functions28. Exosomal circPACRGL has been found to have an oncogenic role in enhancing the proliferation and metastasis of colorectal cancer29. And exosome-shuttled circSHOC2 from ischemic-preconditioned astrocyte could also regulate neuronal autophagy30.
Through RNA sequencing analysis, we identified 89 differentially expressed circRNAs and 9 miRNAs in exosomes derived from gastric cancer AGS cells with TOB1 overexpression. Among the total of 89 circRNAs, 87 were significantly upregulated and 2 were significantly downregulated following TOB1 overexpression. Furthermore, we classified 12 circRNAs (including 10 upregulated and 2 downregulated) that are not functionally annotated in current circRNA databases as novel circRNAs. These results suggest that the significantly dysregulated circRNAs may be implicated in TOB1-related gastric cancer, warranting further confirmation.
We conducted GO and KEGG pathway analyses on the host genes of significantly dysregulated circRNAs and revealed that hsa_circ_0008719 are derived from AKT2. Our analysis indicated that AKT2 may be involved in the biological process of autophagy according to GO analysis. Similarly, KEGG analysis determined that AKT2 is primarily associated with gastric cancer pathways and autophagy-related pathways, which further influence the tumorigenesis of gastric cancer. Moreover, based on Pearson correlation tests using data from the GEPIA web server and CCLE database, we found that both AKT2 and TOB1 are positively correlated across various gastric cancer cell lines, suggesting that TOB1 may regulate downstream hsa_circ_0008719.
The host gene AKT2 has been identified to trigger secretory autophagy by reducing lysosomal function in retinal pigment epithelial (RPE) cells31. However, the role of hsa_circ_0008719 driven by AKT2 has not been previously recognized in tumors, including gastric cancer. Sanger sequencing and RNase R treatment assay confirmed the circular structure of hsa_circ_0008719, and dual-luciferase reporter assays and nucleocytoplasmic separation experiments demonstrated that hsa_circ_0008719 plays a significant role in the cytoplasm by interacting with miR-3615, which is believed to be involved in autophagy related to gastric cancer through its host gene AKT2. For the cytoplasmic-localized hsa_circ_0008719, the criteria used to evaluate its coding potential include the presence of open reading frames (ORFs) with an internal ribosome entry site (IRES)32. However, analysis using IRESfinder (Supplementary Table 2) revealed that hsa_circ_0008719 lacks an internal ribosome entry site (IRES), suggesting that hsa_circ_0008719 has no potential to encode peptides. Additionally, exosomal miRNAs play a crucial role in tumor development; for instance, exosomal miR-552 has been found to promote the malignant progression of laryngeal carcinoma cells via the PTEN/TOB1 axis33. The role of exosomal miR-3615 is not clear in gastric cancer. The expression of miR-3615 in patients with HCC was inversely associated with their overall survival time and positively correlated with high TNM stage, and serum alpha-fetoprotein level34. In pancreatic ductal adenocarcinoma, hsa_circ_0008719 has been shown to be downregulated compared to adjacent normal tissues (circRNADisease v2.0), suggesting that hsa_circ_0008719 may function as a tumor suppressor. Consistent with this finding, our results also indicated a significant upregulation of hsa_circ_0008719 in exosomes derived from gastric cancer cells overexpressing TOB1. Most importantly, we observed that the expression of hsa_circ_0008719 in 52 gastric cancer tissues was significantly lower than that in corresponding normal gastric tissues, and the reduced expression of hsa_circ_0008719 in gastric cancer tissues was associated with poor differentiation, which may indicate an unfavorable prognosis for gastric cancer patients.
It has been found that not only intracellular circRNAs, but also exosomal circRNAs could exert their effects to regulate autophagy. Hsa_circ_0001658 modulates apoptosis and autophagy in AGS and HGC-27 cells through miRNA-18235. Additionally, exosome-derived circ-PVT1 regulates autophagy through miR-30a-5p, thereby promoting cisplatin resistance in AGS and HGC-27 cells36. Thus, a series of follow-up qRT-PCR assays were used for the first time to elucidate the potential relationship of TOB1, hsa_circ_0008719 and its target miR-3615. Primarily, we identified that hsa_circ_0008719 are remarkably upregulated in AGS and HGC-27 cell lines and their exosomes with TOB1 overexpression, while miR-3615 exhibited the opposite trend. These results indicate that the expression of hsa_circ_0008719 positively associated with TOB1 in gastric cancer cells and exosomes, establishing the premise that TOB1 overexpression can reprogram the expression of both hsa_circ_0008719 and its target miR-3615. Furthermore, this conclusion highlights that hsa_circ_0008719 may be linked to autophagy through the secretion of numerous exosomes in gastric cancer cells with TOB1 overexpression10,11. Wang et al. also identified differentially expressed circRNAs in exosomes derived from liver cancer cells via RNA sequencing. They demonstrated that circTGFBR2, functioning as a competitive endogenous RNA (ceRNA), promotes ATG5 expression and enhances autophagy in liver cancer cells by binding to miR-205-5p. Consequently, circTGFBR2 represents a novel tumor-promoting circRNA in liver cancer cell-derived exosomes37.
The specific function of exosomal hsa_circ_0008719 in gastric cancer was further investigated through the knockdown and overexpression of hsa_circ_0008719 in AGS-TOB1 and AGS-Vector cells, respectively. This manipulation resulted in the upregulation and downregulation of miR-3615, with corresponding changes observed in exosomal hsa_circ_0008719 compared to the control group. Previous studies have reported that exosomal circRNAs can induce autophagy; for instance, circATG4B promotes autophagy in colorectal cancer cells via exosomes, thereby inducing resistance to oxaliplatin38. Additionally, exosomes delivering circFOXP1 can upregulate autophagy and promote HCC progression through the p196 protein encoded by circFOXP139. We subsequently co-cultured HGC-27-TOB1-6E12 cells with exosomes derived from AGS-Vec-8719 and AGS-Vec-NC cells separately, followed by a series of functional experiments. Our findings indicate that TOB1 acts as an upstream regulator of hsa_circ_0008719 and miR-3615; hsa_circ_0008719 can be stably delivered via exosomes, which in turn inhibit gastric cancer cell proliferation and enhance autophagy levels, thereby exerting a tumor suppressor effect (Fig. 7).

Schematic representation of how TOB1 overexpression in gastric cancer induces the differential expression of hsa_circ_0008719 and miR-3615 in both cells and exosomes (by Figdraw).
In conclusion, our study identifies 98 differentially expressed noncoding RNAs associated with TOB1 overexpression. Hsa_circ_0008719 can competitively bind to miR-3615. Exosomes transport hsa_circ_0008719, thereby reducing the growth and proliferation of gastric cancer cells while promoting autophagy to function as a tumor suppressor. We elucidate for the first time that exosomal hsa_circ_0008719 represents a novel target for gastric cancer treatment. Further investigation into the specific mechanisms by which TOB1-hsa_circ_0008719-miR-3615-induced autophagy suppresses gastric cancer progression is warranted.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request. We have submitted the raw data to the SRA database(accession number PRJNA1243057).
References
Thrift, A. P., Wenker, T. N. & El-Serag, H. B. Global burden of gastric cancer: epidemiological trends, risk factors, screening and prevention. Nat. Reviews Clin. Oncol. 20 (5), 338–349 (2023).
Otani, K. et al. Epigenetic-mediated tumor suppressor genes as diagnostic or prognostic biomarkers in gastric cancer. Expert Rev. Mol. Diagn. 13 (5), 445–455 (2014).
Lee, H. S. et al. Transducer of ERBB2.1 (TOB1) as a tumor suppressor: A mechanistic perspective. Int. J. Mol. Sci. 16 (12), 29815–29828 (2015).
Wang, D. et al. Phosphorylation of TOB1 at T172 and S320 is critical for gastric cancer proliferation and progression. Am. J. Translational Res. 11 (8), 5227–5239 (2019).
Helms, M. W. et al. TOB1 is regulated by EGF-dependent HER2 and EGFR signaling, is highly phosphorylated, and indicates poor prognosis in node-negative breast cancer. Cancer Res. 69 (12), 5049–5056 (2009).
Yu, J. C. et al. Human gastric adenocarcinoma allelotype on chromosomes 17 and 18. J. Int. Med. Res. 36 (2), 279–288 (2008).
Yu, J. et al. Three distinct regions of allelic deletion on chromosome 17 involved in sporadic gastric cancer. Hepatogastroenterology 55 (85), 1487–1491 (2008).
Guan, R. et al. Decreased TOB1 expression and increased phosphorylation of nuclear TOB1 promotes gastric cancer. Oncotarget 8 (43), 75243–75253 (2017).
Guo, H. et al. Decreased expression levels of DAL-1 and TOB1 are associated with clinicopathological features and poor prognosis in gastric cancer. Pathol. Res. Pract. 215 (6), 152403 (2019).
Wang, D. et al. Involvement of TOB1 on autophagy in gastric cancer AGS cells via decreasing the activation of akt/mtor signaling pathway. PeerJ 10, e12904 (2022).
Wang, Y. et al. The overexpression of TOB1 induces autophagy in gastric cancer cells by secreting exosomes. Dis. Markers 2022, 7925097 (2022).
Zhang, Y. et al. Exosome: A review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int. J. Nanomed. 15, 6917–6934 (2020).
Yue, B. et al. Exosome biogenesis, secretion and function of Exosomal MiRNAs in skeletal muscle myogenesis. Cell. Prolif. 53 (7), e12857 (2020).
Li, Y. et al. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell. Res. 25 (8), 981–984 (2015).
Shi, Y., Jia, X. & Xu, J. The new function of circrna: translation. Clin. Transl Oncol. 22 (12), 2162–2169 (2020).
Cao, B. et al. Role of circular RNAs and long non–coding RNAs in the clinical translation of gastric cancer (Review). Int. J. Mol. Med. 47 (1), 77–91 (2021).
Xia, H., Green, D. R. & Zou, W. Autophagy in tumour immunity and therapy. Nat. Rev. Cancer. 21 (5), 281–297 (2021).
Debnath, J., Gammoh, N. & Ryan, K. M. Autophagy and autophagy-related pathways in cancer. Nat. Rev. Mol. Cell. Biol. 24 (8), 560–575 (2023).
Villarroya-Beltri, C. et al. ISGylation controls exosome secretion by promoting lysosomal degradation of MVB proteins. Nat. Commun. 7, 13588 (2016).
Wang, Y. et al. Exosome-autophagy crosstalk in enveloped virus infection. Int. J. Mol. Sci. 24 (13) (2023).
Kanehisa, M. et al. KEGG: biological systems database as a model of the real world. Nucleic Acids Res. 53 (D1), D672–D677 (2025).
Kanehisa, M. Toward Understanding the origin and evolution of cellular organisms. Protein Sci. 28 (11), 1947–1951 (2019).
Ogata, H. et al. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 27 (1), 29–34 (1999).
Sang, H. et al. Exosomal circRELL1 serves as a miR-637 sponge to modulate gastric cancer progression via regulating autophagy activation. Cell. Death Dis. 13 (1), 56 (2022).
Babuta, M. et al. Dysregulated autophagy and lysosome function are linked to exosome production by micro-RNA 155 in alcoholic liver disease. Hepatology 70 (6), 2123–2141 (2019).
Hessvik, N. P. & Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 75 (2), 193–208 (2018).
Jiang, T. Y. et al. PTEN deficiency facilitates exosome secretion and metastasis in cholangiocarcinoma by impairing TFEB-mediated lysosome biogenesis. Gastroenterology 164 (3), 424–438 (2023).
Wang, Y. et al. Exosomal circrnas: biogenesis, effect and application in human diseases. Mol. Cancer. 18 (1), 116 (2019).
Shang, A. et al. Exosomal circpacrgl promotes progression of colorectal cancer via the miR-142-3p/miR-506-3p- TGF-beta1 axis. Mol. Cancer. 19 (1), 117 (2020).
Chen, W. H. et al. Exosome-shuttled circSHOC2 from IPASs regulates neuronal autophagy and ameliorates ischemic brain injury via the miR-7670-3p/SIRT1 axis. Mol. Therapy-Nucleic Acids. 22, 657–672 (2020).
Ghosh, S., Hose, S. & Sinha, D. AKT2-mediated lysosomal dysfunction promotes secretory autophagy in retinal pigment epithelium (RPE) cells. Autophagy 1–2 (2024).
Chen, C. K. et al. Structured elements drive extensive circular RNA translation. Mol. Cell. 81 (20), 4300–4318 (2021).
Liu, L. et al. Exosome miR-552 promotes laryngocarcinoma cells malignant progression via the PTEN/TOB1 axis. Pharmacology 108 (4), 321–330 (2023).
Yuan, X., Zhang, Y. & Yu, Z. Expression and clinical significance of miR-3615 in hepatocellular carcinoma. J. Int. Med. Res. 49 (1), 300060520981547 (2021).
Duan, X., Yu, X. & Li, Z. Circular RNA hsa_circ_0001658 regulates apoptosis and autophagy in gastric cancer through microRNA-182/Ras-related protein Rab-10 signaling axis. Bioengineered 13 (2), 2387–2397 (2022).
Yao, W. et al. Exosome-derived Circ-PVT1 contributes to cisplatin resistance by regulating autophagy, invasion, and apoptosis via miR-30a-5p/YAP1 axis in gastric cancer cells. Cancer Biother Radiopharm. 36 (4), 347–359 (2021).
Wang, X. et al. Exosomal circTGFBR2 promotes hepatocellular carcinoma progression via enhancing ATG5 mediated protective autophagy. Cell. Death Dis. 14 (7), 451 (2023).
Pan, Z. et al. A novel protein encoded by Exosomal CircATG4B induces oxaliplatin resistance in colorectal cancer by promoting autophagy. Adv. Sci. (Weinh). 9 (35), e2204513 (2022).
Zhao, P. et al. Exosome-delivered circFOXP1 upregulates autophagy and promotes hepatocellular carcinoma progression through its encoded p196 protein targeting the KHDRBS3/ULK1 axis. Int. J. Nanomed. 20, 5247–5265 (2025).
Funding
This work was supported by National Natural Science Foundation of China (81372174).
Author information
Authors and Affiliations
Contributions
Na Qian and Tongtong Huang: Methodology, Validation, Formal analysis, Writing—original draft. Lulu Zhang: Methodology, Validation. Li Zhang: Validation, Formal analysis. Lijie Zhou: Resources, Writing—review & editing. Dong Wang: Investigation, Formal analysis. Xiao Liang: Validation. Songbin Fu: Investigation, Supervision, Data curation. Yingwei Xue: Resources. Jingcui Yu: Conceptualization, Methodology, Resources, Writing—review & editing, Funding acquisition.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Qian, N., Huang, T., Zhang, L. et al. Reprogramming of hsa_circ_0008719 and its target miR-3615 induced by TOB1 in exosomes of gastric cancer cells: a contribution to inhibit gastric cancer progression. Sci Rep 15, 27089 (2025). https://doi.org/10.1038/s41598-025-12789-8
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-12789-8
