
Abstract
Bael [Aegle marmelos (L.) Correa] is widely consumed for its therapeutic benefits having significant medicinal and nutritional properties. However, limited data exists regarding the compositional diversity among bael genotypes in the ranges of North Western Himalayan regions. This research focused on characterizing the quality attributes and sensory landscapes of selected bael genotypes. This study evaluates the physicochemical traits of eighty wild bael genotypes to identify promising candidature for selective breeding when compared with commercial cultivars (NB-5, NB-9). The wild genotype JMU-Bael (Sel-27) exhibited the highest TSS of mucilage (49.60 0brix), TSS of pulp (36.77 0brix), total sugars (21.17%), reducing sugars (9.58%), non-reducing sugars (11.59%) and ascorbic acid (25.45 mg/100 g). Principal component analysis (PCA) highlighted significant genetic diversity, with the first component accounting for 81.17% of the total variation. Cluster analysis grouped the genotypes into two main clusters, offering insights for breeding programs aimed at enhancing bael fruit quality.
Similar content being viewed by others


Introduction
Bael [Aegle marmelos (L.) Correa] belonging to the Rutaceae family, is a significant yet underutilized fruit crop in India, renowned for its medicinal and nutritional benefits1. Bael is primarily found in tropical and subtropical regions and is highly adaptable to challenging soil and environmental conditions2,3. Bael can tolerate temperatures ranging from − 7 to 50 °C and thrives in alkaline, stony and shallow soils.
Bael is an excellent choice for regions with limited water resources due to its rich vitamin and mineral content. With the growing global demand for natural antioxidants and functional foods, bael fruit stands out as a valuable source of riboflavin, providing therapeutic benefits for conditions such as beriberi4. Almost every part of the bael tree is used in different ayurvedic and traditional medicines for its antidysentery, antidiarrheal, antipyretic, antidiabetic, antibacterial, antifungal, antiviral, anticancer, analgesic, antimicrobial and antihelminthic properties. The fruit pulp and leaves are particularly rich in bioactive chemicals, including total carotenoids, tannins, flavonoids, phenolics, terpenoids and coumarins, which are primarily responsible for these therapeutic qualities5,6. Several bioactive compounds have been isolated from various parts of the tree, including marmelosin, aegelin, citral, dictamnine, luvangetin, cineole, citronellol, rutin, lupeol, skimmianine, xanthotoxin, scopoletin, psoralen and tembamide5,7. The fruit is also an excellent source of protein, carbohydrates, minerals and vitamins, particularly vitamin B2 (riboflavin)8,9. The ripened fruit pulp contains higher amounts of marmelosin compared to other parts of the bael tree10.
In the Indian subcontinent, mature bael fruits are primarily consumed fresh, with the ripened pulp being used to make beverages such as sharbat or shakes. The pulp is also commercially processed to produce a variety of products including murabba, toffees, jams, powders squash and fruit bars11. Due to its capacity to treat constipation and its calming, laxative and fragrant qualities, bael is highly valued in tropical and desert regions. Recent technological advancements have led to the development of dehydrated bael powder, which is particularly noted for its ability to retain pharmacological ingredients and for its longer shelf life compared to fresh fruit pulp10. Nowadays, several processing companies have introduced ready-to-serve (RTS) beverages made from bael fruit pulp as a component of mixed fruit juice. The processing industry requires bael cultivars that can provide desirable fruit quality characteristics, such as higher high total soluble solids (TSS), fewer seeds, pulp recovery, thinner shells and higher medicinal and nutritional content for commercial purposes.
However, the species is currently categorized as “Near Threatened” by the Interna-tional Union for Conservation of Nature and Natural Resources (IUCN) and “Vulnerable” under Criterion A, as well as a Rare, Endangered, and Threatened (RET) species by the Foundation for the Revitalization of Local Health Traditions (FRLHT), Bangalore, India, due to manmade threats to its natural habitat12,13. This highlights the need for concrete strategies for the cultivation and conservation of bael14.
The primary goal of the study was to investigate in situ variability, characterize and quantify the variation in various physicochemical traits and identify promising selections from a diverse range of bael genotypes in the North Western Himalayas. Additionally, fruit quality characteristics were evaluated to pinpoint potential accessions with valuable attributes for Himalayan regions. The biochemical properties of fruits from various bael accessions were analysed to offer insights into their genetic enhancement and commercial potential. This information is intended to improve conservation management and facilitate future utilization in bael breeding programs.
Results
Physico-chemical traits and sensory attributes
The maximum total soluble solids (TSS) of mucilage (49.60 °Brix) was observed in JMU-Bael (Sel-27) followed by JMU-Bael (Sel-9) at 49.47 °Brix and JMU-Bael (Sel-19) at 49.40 °Brix, with the lowest (40.10 °Brix) in JMU-Bael (Sel-63) among all the bael genotypes. For the pulp, the highest TSS (36.77 °Brix) was recorded in JMU-Bael (Sel-27), followed by JMU-Bael (Sel-41) at 36.67 °Brix and the lowest (25.90 °Brix) in JMU-Bael (Sel-63). The highest titratable acidity (0.33%) was recorded in JMU-Bael (Sel-63), while the lowest (0.20%) was observed in JMU-Bael (Sel-27). Regarding total sugars, JMU-Bael (Sel-27) exhibited the highest content (21.17%) followed by JMU-Bael (Sel-32) at 21.11% and JMU-Bael (Sel-9) at 21.09%, with the lowest (8.11%) in JMU-Bael (Sel-63). In terms of reducing sugars, the highest value (9.58%) was observed in JMU-Bael (Sel-27) followed by JMU-Bael (Sel-9) (9.55%) and JMU-Bael (Sel-32) (9.54%), while the lowest (2.51%) was found in JMU-Bael (Sel-63). For non-reducing sugars, the highest content (11.59%) was recorded in JMU-Bael (Sel-27), followed by JMU-Bael (Sel-32) (11.57%) and JMU-Bael (Sel-9) (11.54%), while the lowest (5.60%) was found in JMU-Bael (Sel-63). The maximum ascorbic acid content (25.45 mg/100 g) was recorded in JMU-Bael (Sel-27), followed by JMU-Bael (Sel-20) (25.38 mg/100 g), while the minimum (14.38 mg/100 g) was found in JMU-Bael (Sel-63) (Table 1).
Among the eighty wild bael genotypes evaluated alongside two commercial cultivars, pulp color was pale yellow in sixty-one genotypes including NB-5, yellow in ten genotypes, and dark yellow in eleven genotypes, including NB-9 (Supplementary Fig. S1). The pulp taste varied, with thirty genotypes having less sweet pulp, forty genotypes, including NB-9, having medium sweet pulp and twelve genotypes, including NB-5, having sweet pulp. Pulp flavor was mild in thirty bael genotypes, moderate in forty bael genotypes, including NB-9 and strong in twelve bael genotypes, including NB-5. Pulp acridity was absent in fifty-two bael genotypes, including NB-5 and NB-9, while it was present in thirty bael genotypes (Table 2).
Genetic parameters
Environmental coefficient of variation (ECV), genotypic coefficient of variation (GCV), phenotypic coefficient of variation (PCV), broad-sense heritability (h²b), genetic advance (GA) and genetic advance as a percentage of the mean (GAM) were calculated for seven physico-chemical traits (Table 3). The highest values for h²b were observed in titratable acidity (99.99%) and total sugars (99.99%); GA was observed in total sugars (6.30%), while the highest ECV (2.78%) was recorded in the titratable acidity. Reducing sugars showed the highest GCV (37.02%), PCV (37.03%) and GAM (76.26%). The lowest values for ECV was found in total sugars (0.10%); GCV (6.65%) and PCV (6.66%) were found in TSS mucilage; GA in titratable acidity (0.06%) and GAM in TSS Mucilage (13.69%). All seven physico-chemical traits exhibited high broad-sense heritability values (> 98%), indicating their potential for improvement through selective breeding.
Correlation matrices
Correlations at both genetic and phenotypic levels play a significant role in the selection process for parental plants in breeding programs. For instance, TSS mucilage exhibited highly strong correlations with a multitude of traits including TSS Pulp, total sugars, reducing sugars, non reducing sugars and ascorbic acid. Similarly, TSS pulp demonstrated strong associations with several traits, including total sugars, reducing sugars, non reducing sugars and ascorbic acid. However, total sugars exhibited highly strong correlations with reducing sugars, non reducing sugars and ascorbic acid. Conversely, titratable acidity showed highly strong negative correlations with certain traits such as total sugars, reducing sugars, non reducing sugars and ascorbic acid (Tables 4 and 5).
Principal components analysis
In this study, principal component analysis (PCA) was recorded in 7 physico-chemical traits. The eigenvalues of the first principal components was found to be greater than 1, accounting for a cumulative contribution rate of 81.17%. This indicates that first principal components effectively represent the majority of the information related to the 7 traits analyzed (Table 6; Fig. 1). The first principal component (PC1) showed significant factor loadings for traits such as total sugars (0.41), reducing sugars (0.39), TSS pulp (0.39), TSS mucilage (0.38), ascorbic acid (0.36) and non reducing sugars (0.34), explaining 81.17% of the total variance.

2-Dimensional scatter plot for the studied of bael genotypes based on PC1/PC2.
Cluster analysis
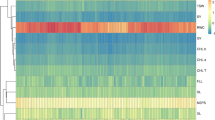
In this study, cluster analysis was performed using the Ward method on seven physico-chemical traits from eighty wild bael genotypes along with two commercial cultivars, NB-5 and NB-9. The analysis grouped the bael genotypes and cultivars into two primary clusters, each further divided into sub-clusters. Cluster I contained sixty genotypes and Cluster II comprised twenty two bael genotypes (Fig. 2).

Ward cluster analysis of the studied bael genotypes based on physico-chemical traits using Euclidean distances. The results show that the populations were divided into 2 categories, which was indicated by red and blue colors.
Discussion
The variations recorded in physico-chemical traits and sensory attributes among the eighty wild bael genotypes and two commercial cultivars highlight the rich genetic diversity within the bael population. This diversity offers significant potential for breeding programs aimed at improving biochemical composition and organoleptic attributes. The high TSS of mucilage and pulp observed in JMU-Bael (Sel-27) and similar genotypes indicates their potential for producing sweeter fruits, which are generally preferred by consumers. The significant variation in TSS, highlights the scope for selecting and breeding genotypes with enhanced sweetness. The variation in TSS (Total Soluble Solids) observed in bael fruits can likely be attributed to differences in their genetic backgrounds. Since TSS is a critical determinant of fruit sweetness and overall quality, it is essential for breeders to focus on this trait when selecting elite bael types. By prioritizing TSS content, breeders can develop cultivars that meet consumer preferences for sweeter fruits, which are increasingly important in both fresh and processed fruit markets13. The variation in titratable acidity, with JMU-Bael (Sel-63) having the highest acidity, suggests that some genotypes may be more suitable for processing into products like preserves and beverages, where a higher acid content is desirable. The low acidity observed in JMU-Bael (Sel-27) could make it more suitable for fresh consumption, catering to consumers who prefer less acidic fruits. The high levels of total sugars, reducing sugars, and non-reducing sugars in JMU-Bael (Sel-27) further enhance its appeal as a cultivar with superior sweetness. The high ascorbic acid content in JMU-Bael (Sel-27) is particularly noteworthy, as ascorbic acid (vitamin C) is an important antioxidant that contributes to the nutritional value of bael fruits. Similarly, consumers tend to favor fruits that have higher total soluble solids content and lower acidity levels. This finding aligns with earlier research showed that considerable variations in physico-chemical traits among different bael genotypes provide ample opportunities for breeding of desirable hybrids13,15,16,17,18.
The evaluation of eighty wild bael genotypes alongside two commercial cultivars revealed a wide range of variability in organoleptic parameters, which is consistent with findings from other studies on bael fruit diversity. The pulp colour, one of the most notable visual traits, ranged from pale yellow to dark yellow. The diversity in pulp sweetness also aligns with the variability in sugar content reported in bael germplasm collections. The sweet pulp of JMU-Bael (Sel-27), indicate that this genotype may possess favourable sugar profiles, making them candidates for further selection. Pulp flavor, an important organoleptic characteristic, showed similar trends. The strong flavor of JMU-Bael (Sel-27) could be a result of higher concentrations of these aromatic compounds makes it a balanced option for consumer preference. Pulp acridity was absent in a majority of genotypes, which is a desirable trait for fresh consumption and processing purposes. The absence of acridity in JMU-Bael (Sel-27) boosts its potential as a high-quality genotype for breeding and commercialization, aligning with consumer preferences and consistent with Parihar19 and Singh et al.20, which used SSR markers on the same 80 wild genotypes and NB-5/NB-9 to confirm genetic divergence and support our physicochemical clustering. This molecular data reinforces JMU-Bael (Sel-27)’s genetic superiority for future breeding.
The genetic coefficient of variance (GCV) and heritability are crucial parameters for enhancing crops through selective breeding. High genotypic coefficient of variance (GCV) and phenotypic coefficient of variance (PCV) indicate significant genetic variation that can be effectively utilized for developing improved cultivars, whereas low values suggest limited potential for selection-driven improvement. In vegetatively propagated crops such as bael, broad-sense heritability estimates are particularly valuable as they account for both additive and non-additive genetic components inherited by offspring. These findings are consistent with Debbarma and Hazarika13, who investigated ECV, GCV and PCV in various fruit traits of thirty bael accessions. The pronounced heritable variation reflected by high GCV and PCV values underscores the potential for employing targeted selection and breeding strategies to improve desirable traits in this nutritionally and economically important fruit crop.
These correlations are essential for breeders to identify and choose parents with desired traits. In this study, highly significant correlations (p < 0.01) were observed among all physico-chemical traits examined. The correlation coefficients among all traits were further analyzed to delineate genotypic and phenotypic relationships. These analyses, based on error variance and covariance matrices of physico-chemical traits, revealed highly significant associations among all traits examined. Specifically, TSS mucilage, TSS pulp, total sugars, reducing sugars and non reducing sugars exhibited significant positive correlations with other traits at both the genotypic and phenotypic levels. However, it was observed that titratable acidity exhibited negative correlations with other traits. Moreover, certain traits displayed particularly strong associations. In present study revealed that for most traits, the genotypic correlation coefficients were of a higher magnitude than the phenotypic ones, indicating the predominant role of heritable factors. Genetic relation of traits may result from pleotropic effects of a gene, linkage of two genes, chromogema and regimental affiliation or due to the environmental influences. These results are in conformity to the findings of Debbarma and Hazarika13 who studied the genotypic and phenotypic correlation of morphological characteristics in thirty bael accessions.
Plotting relationships among individuals in two or more dimensions using PCA enables a more realistic interpretation of those relationships21. PCA reduces dimensionality, which looks at data to find relationships between objects, in order to estimate the correlation structure of variables and find the minimum number of components (a linear combination of the original features) required to explain the majority of the variance with the least amount of information loss22,23. For the purpose of developing higher-quality bael genotypes, it is essential to identify and characterize the genetic variability present in bael. According to PCA, the genetic diversity among genotypes may be influenced by elements such as heterogeneity, population genetic architecture, and developmental features24. In the current study, genotype similarities and differences were assessed using PCA with physico-chemical variables. PCA generally verified that the physico-chemical features were arranged into species-specific clusters. The results revealed that the initial first principal components collectively contributed 81.17% of the total variance, suggesting they could effectively represent the majority of bael traits, potentially indicating the integration of these traits. Individuals with higher scores from comprehensive evaluation were subsequently chosen through this analysis. For instance, prior investigations unveiled that in bael, the initial eight principal components collectively accounted for 94.80% of the total variance, indicating a comparable concentration of variability within that germplasm17. Similarly, in bael, the first ten principal components elucidated a striking 85.15% of the observed variation, showcasing a tightly packed distribution of variability within this particular bael genotypes13. Furthermore, in the case of bael, the total variance of the first principal components amounted to 47.12%, signifying a substantial portion of variability captured within a limited set of dimensions15. Similarly, in bael, the first principal components accounted for an impressive 84.60% of the total variation, highlighting a concentrated distribution of variability among 213 bael accessions16. Consequently, the findings of this study offer potential applications in the selection of bael individuals with outstanding traits.
Cluster analysis revealed significant diversity among bael genotypes. In this study, sixty genotypes were grouped into Cluster I, while the remaining twenty two genotypes were classified into Cluster II. Similar studies on bael have identified two primary clusters among thirty accessions, with five accessions in the first cluster and twenty-five in the second cluster13. Another analysis of seventy five bael genotypes also revealed two primary clusters, with twelve genotypes in the first cluster and sixty-three genotypes in the second cluster17. This study involving twenty one bael accessions, where statistical analysis led to their classification into three groups: the first group comprising seven accessions, the second group including two accessions and the third group consisting of twelve accessions15. Similar study on two hundred thirteen bael genotypes, where cluster analysis resulted in the classification of the genotypes into six groups: the first group consist One hundred thirty-one genotypes, the second group consist Seventy three genotypes, the third group consist six genotypes and cluster fourth, cluster fifth and cluster sixth each consist of one genotype16.
Conclusions
Bael is cultivated and consumed globally for its exceptional nutritional benefits, appealing taste and valuable health-promoting properties. The wide range of physico-biochemical and sensory attributes observed among the bael genotypes underscores the importance of conserving and utilizing this genetic diversity. The genotype JMU-Bael (Sel-27), with their superior physico-chemical traits and pulp quality, could serve as valuable resources for future breeding programs aimed at developing high-quality bael cultivars. Moreover, the identification of genotypes with specific traits, such as TSS, Sugars or high vitamin C content, provides opportunities for targeted breeding to meet specific consumer and market needs.
Materials and methods
Plant materials
In the present investigation, a total of 80 wild bael genotypes, were selected from major bael growing districts in Jammu, Samba and Kathua within the Jammu province of the Jammu and Kashmir Union Territory in India (Fig. 3). These 80 genotypes were compared with two commercial cultivars, NB-5 and NB-9. The geographic coordinates for each region, encompassing latitude, longitude and altitude, were recorded using a Global Positioning System (GPS) (Supplementary Table S1).The collection of plant material complied with all relevant institutional, national and international regulations and guidelines. Voucher specimens of the collected bael (Aegle marmelos) germplasm were identified by Prabhdeep Singh under the supervision of Dr. Akash Sharma, as part of a breeding program aimed at developing new cultivars. These specimens have been deposited in the Division of Fruit Science, Faculty of Horticulture, SKUAST-Jammu, under the voucher ID AUJ/FS/23–24/121.

Collection sites of bael genotypes in the Jammu and Kashmir districts of India.
Physico-chemical traits and sensory attributes
Total Soluble Solids (TSS) of mucilage were measured by collecting mucilage from the fruit and using a digital refractometer (0–53 °Brix, Pocket PAL-1, ATAGO Corporation, Tokyo, Japan) to determine the TSS in degrees Brix. To determine the Total Soluble Solids (TSS) of the pulp, 20 g of fruit pulp were blended for 3 min, then wrapped in cheesecloth, hand-squeezed, and the juice extracted was measured using a digital refractometer (0–53 °Brix, Pocket PAL-1, ATAGO Corporation, Tokyo, Japan). Titratable acidity (%) was assessed by titrating 2 ml of bael fruit juice with a standardized N/10 Sodium Hydroxide (NaOH) solution, using phenolphthalein dye (two drops) as an indicator until the end point was reached, marked by a light pink color persisting for at least 2 s, and expressed as a percentage of citric acid25.
For sugar analysis, 5 ml of bael juice extract and placed in a 100 ml beaker after that add 2 ml of 45% lead acetate. The solution was kept at room temperature for 10 min, after which 5 ml of 22% potassium oxalate solution was added. Distilled water was then added to adjust the volume to 100 ml. The mixture was filtered using Whatman No. 1 filter paper. The amounts of total sugars and reducing sugars were analyzed using the Lane-Eynon titration technique. To determine reducing sugars, 5 ml of 60% hydrochloric acid was mixed with 25 ml of the filtrate from the prepared solution. This mixture was left undisturbed overnight to ensure complete hydrolysis. The hydrolyzed sample was heated for 10 min in a water bath at 68 °C and then neutralized with 10% and 0.1% sodium hydroxide solutions. Phenolphthalein was used as an indicator to determine the neutralization point. Reducing sugars were calculated with the formula: Reducing sugars = (0.05/V1) × (V2/V3) × 100, where V1 is the volume of filtrate used, V2 is the dilution factor, and V3 is the volume of juice. For total sugars, an aliquot of the filtrate was placed in a burette. A 5 ml portion of each of the preheated Fehling solutions A and B was combined with the filtrate, and the mixture was heated on a hot plate. Methylene blue served as an indicator, with the endpoint indicated by the appearance of a brick-red precipitate. The percentage of total sugars was calculated using the formula: Total sugars = (0.05/V1) × (V2/V3) × (V4/V1) × 100, where V1 is the volume of filtrate, V2 is the dilution factor, V3 is the volume of juice, and V4 is the final volume. Non-reducing sugars were determined by subtracting the amount of reducing sugars from the total sugars, expressed as a percentage: Non-reducing sugars (%) = (Total sugars - Reducing sugars).
The ascorbic acid (Vitamin C) content was determined using the 2,6-dichlorophenol indophenol (DCPIP) titration method. For the analysis, 5 g of the fruit sample were macerated with approximately 25 ml of 4% oxalic acid solution. The mixture was filtered through Whatman No. 4 filter paper, and the filtrate was collected into a 50 ml volumetric flask. The obtained solution was titrated against a standard DCPIP dye solution until a stable rose-pink color appeared and persisted for at least 5 s. The ascorbic acid content was calculated and expressed in mg per 100 g of the sample25.
The observations on various traits such as pulp colour, pulp taste, pulp flavour and pulp acridity were recorded following the Guidelines for the Conduct of Tests for Distinctiveness, Uniformity and Stability of Bael (Aegle marmelos Correa) by the Protection of Plant Varieties and Farmers’ Rights Authority (PPV&FRA), Government of India26 and the bael descriptor provided by the National Bureau of Plant Genetic Resources27.
Statistical analyses
Variability among bael genotypes was assessed using one-way analysis of variance (ANOVA) and Least Significant Differences (LSD) with OriginPro 9.1 software, where the LSD test was conducted at a significance level of p < 0.05.
Genetic parameters
Genetic parameters, including Genotypic Coefficient of Variation (GCV), Phenotypic Coefficient of Variation (PCV), Heritability, Genetic Advance (GA) and Genetic Advance as a Percentage of Mean (GAM) were analyzed using R Software. The genotypic and phenotypic variances of the traits were determined using the formulas denoted. Furthermore, the covariance components encompassing genotypic, phenotypic and environmental aspects between the two traits were estimated utilizing the formulas indicated as.
GCV (%) = \(\:\frac{\sqrt{{\sigma\:}_{G}^{2}}}{\stackrel{-}{X}}\)x 100.
PCV (%) = \(\:\frac{\sqrt{{\sigma\:}_{p}^{2}}}{\stackrel{-}{X}}\)x 100.
ECV (%) = \(\:\frac{\sqrt{{\sigma\:}_{E}^{2}}}{\stackrel{-}{X}}\)x 100
.
Where, GV = Genotypic variance, PV = Phenotypic variance, \(\:{\sigma\:}_{E}^{2}\) = Error variance, GCV = Genotypic coefficient of variation, PCV = Phenotypic coefficient of variation, ECV = Environmental coefficient of variation, MSG = Genotypic mean square value, MSE = Error mean square value, r = No. of replication, = \(\:{\sigma\:}_{E}^{2}=\)Environmental variance (Error mean square from the ANOVA), \(\:\stackrel{-}{X\:}\) or \(\:\stackrel{-}{Y}\) = Mean value of the trait or variable X or Y being evaluated.
Broad sense heritability \(\:{\varvec{h}}_{\varvec{b}\varvec{s}}^{2}\) (%): in percentage (%): Broad sense heritability was calculated for all traits employing the formula as follows:
\(\:{h}_{bs}^{2}\left(\%\right)=\frac{\sqrt{{\sigma\:}_{G}^{2}}}{\sqrt{{\sigma\:}_{P}^{2}}}\:\)x 100
Genetic advance (GA) and genetic advance in percentage of mean (GAM)
The Estimate GA for physico-chemical traits under determined utilizing the formula as follows.
GA=\(\:\frac{\sqrt{{\sigma\:}_{G}^{2}}}{\sqrt{{\sigma\:}_{P}^{2}}}\:x\:\)K x σp.
Where: K = Constant selection differential at 5% level intensity (= 2.06), σp = Phenotypic standard deviation, h2 = Heritability in broad sense. Genetic advance in percentage of mean (GAM) was calculated as:
GA% M =\(\:\frac{GA}{\stackrel{-}{X}}\)x 100.
Where, \(\:\stackrel{-}{X}\) = grand mean of the character.
Correlation coefficient analysis
Correlation coefficient analysis, covering both Phenotypic and Genotypic correlation coefficients, was also performed with R Software. Correlations at the genotypic level (rGXY) and phenotypic level (rPXY) were estimated by considering the variables X and Y. The variance and covariance among the traits were estimated using the formulae. Following this, genotypic correlations (rGXY) and phenotypic correlations (rPXY) for all characters were derived utilizing the covariance components and the equations.
1) Phenotypic correlation [{rxy (p)}] =\(\:\frac{Cov.xy\left(p\right)\:}{\left[Vx\left(p\right)Vy\left(p\right)\right]}\)12.
2) Genotypic correlation [{rxy (g)}] =\(\:\frac{\text{C}\text{o}\text{v}.\text{x}\text{y}\left(\text{g}\right)\:}{\left[\text{V}\text{x}\left(\text{p}\right)\text{V}\text{y}\left(\text{p}\right)\right]}\)12
Where,
Cov.xy (p) = phenotypic covariance among traits x and y which was calculated as follows.
Cov.xy (p) = genotypic covariance among traits x and y which was estimated as follows:
Cov.xy (g) = Cov.xy (p) - Conv.xy (e).
Vx (P) and Vy (p) = phenotypic variances for the traits x and y, respectively.
Vx (g) and Vy (g) = genotypic variances for the traits x and y, respectively.
Principle component analysis (PCA)
The relationship among bael genotypes was evaluated through Principal Component Analysis (PCA) using OriginPro 9.1 software. Additionally, a scatter plot depicting the first and second principal components (PC1 and PC2) was generated using OriginPro 9.1 software. Interpreting extensive datasets in research can prove challenging. However, Principal Component Analysis (PCA) alleviates these challenges by enhancing data comprehensiveness while mitigating information loss. This is achieved by generating fresh, uncorrelated variables that progressively maximize variance. The process involves identifying these novel variables and principal components, specific to the dataset. It ultimately involves solving an eigenvalue/eigenvector problem through PCA, as exemplified in the study by Jolliffea and Cadima28.
Cluster analysis
Cluster analysis was conducted using Euclidean distance coefficients and Ward’s method, with distance coefficients standardized using the Z scale. The transformed and uncorrelated means of characters were employed to compute Mahalanobis’ distance (D) values. The mean deviation, denoted as \(\:{Y}_{i}^{1}\)– \(\:{Y}_{i}^{2}\), where i = 1, 2 … p, was evaluated, and subsequently, the D2 value was computed as the sum of squares of these deviations, specifically, \(\:\sum\:{(Y}_{i}^{1}\,-\,{Y}_{i}^{2}\:{)}^{2}\:\). The D2 values were determined for all conceivable pairs of combinations among bael genotypes.
Data availability
The original contributions presented in the study are included in the article/Supplementary Files, further inquiries can be directed to the corresponding authors.
References
Amulya, R. N., Shivakumar, A. N., Satish, K. M. & B. S., & Studies on genetic variability and relationship of Bael (Aegle Marmelos (L) Correa) using morphological and molecular markers. J. Hortic. Sci. 17 (1), 88–94. https://doi.org/10.24154/jhs.v17i1.846 (2022).
Uddin, M. S., Islam, M. S., Alam, M. A. & Hossain, M. M. Study on Physico-Morphological characteristics of 14 Bael (Aegle Marmelos Corr.) genotypes grown at chapainawabganj, Bangladesh. Int. J. Minor. Fruits Med. Aromat. Plants. 2 (2), 29–33 (2016).
Khanal, A., Dall’acqua, S. & Adhikari, R. Bael (Aegle marmelos), an underutilized fruit with enormous potential to be developed as a functional food product: A review. J. Food Process. Preserv. 1–11. https://doi.org/10.1155/2023/8863630 (2023).
Meena, V. S. et al. Underutilized fruit crops of Indian arid and Semi-Arid regions: importance, conservation and utilization strategies. Horticulturae 8 (2), 171. https://doi.org/10.3390/horticulturae8020171 (2022).
Maity, P., Dhananjay, H., Bandopadhyay, U. & Mishra, D. K. Biological activities of crude extracts and chemical constituents of Bael (Aegle Mamelos Correa). Indian J. Exp. Biol. 47, 849–861 (2009).
Lambole, V. B., Murti, K., Kumar, U. & Sandipkumar, B. P. Gajera V. Phytopharmacological properties of Aegle marmelos, a potential medicinal tree: an overview. Int. J. Pharm. Sci. Rev. Res. 5 (2), 67–72 (2010).
Sarkar, T., Salauddin, M. & Chakraborty, R. In-Depth Pharmacological and nutritional properties of Bael (Aegle marmelos): A critical review. J. Agric. Food Res. 2, 1–23. https://doi.org/10.1016/j.jafr.2020.100081 (2020).
Manandhar, B., Paudel, K. R., Sharma, B. & Karki, R. Phytochemical profile and Pharmacological activity of Aegle marmelos Linn. J. Integr. Med. 16 (3), 153–163. https://doi.org/10.1016/j.joim.2018.04.007 (2018).
Pathirana, C. K. et al. Assessment of the elite accessions of Bael [Aegle marmelos (L.) Corr.] in Sri Lanka Based on Morphometric, Organoleptic, and Elemental Properties of the Fruits and Phylogenetic Relationships. PLoS One. 15 (5), 1–20. https://doi.org/10.1371/journal.pone.0233609 (2020).
Gurjar, P. S., Bhattacherjee, A. K., Singh, A., Dikshit, A. & Singh, V. K. Characterization of nutraceuticals in Bael powder prepared from fruits harvested at different developmental stages. Indian J. Tradit Knowl. 18 (4), 724–730 (2019). http://op.niscair.res.in/index.php/IJTK/article/view/29006
Sharma, N. et al. Aegle marmelos (L.) correa: an underutilized fruit with high nutraceutical values: A review. Int. J. Mol. Sci. 23 (18), 10889. https://doi.org/10.3390%2Fijms231810889 (2022).
Plummer, J. Aegle marmelos. IUCN Red List. Threatened Species. https://doi.org/10.2305/IUCN.UK.2020-3.RLTS.T156233789A156238207.en (2020).
Debbarma, P. & Hazarika, T. K. Genetic diversity of Bael [Aegle marmelos (L.) Corr.] Accessions from North-East India Based on Principal Component and Cluster Analysis. Genet. Resour. Crop Evol. 1–25. https://doi.org/10.1007/s10722-023-01619-3 (2023).
Chaturvedi, K. et al. Exploring the genetic diversity of Aegle marmelos (L.) Correa populations in India. Plant. Genet. Resour. 21 (2), 107–114. https://doi.org/10.1017/S1479262123000485 (2023).
Kannaujia, P. K. et al. Diversity in Morpho-logical and some chemical traits of Aegle marmelos (L.) Correa germplasm explored from Achanakmar-Amarkantak biosphere reserve, chhattisgarh, India. Genet. Resour. Crop Evol. 1–14. https://doi.org/10.1007/s10722-024-02118-9 (2024).
Singh, A. K. et al. Descriptors for characterization and evaluation of Bael (Aegle Marmelos (L.) Correa ex Roxb.) germplasm for utilization in crop improvement. Genet. Resour. Crop Evol. 1–30. https://doi.org/10.1007/s10722-024-01903-w (2024).
Dhakar, M. K., Das, B., Nath, V., Sarkar, P. K. & Singh, A. K. Genotypic diversity for fruit characteristics in Bael [Aegle marmelos (L.) Corr.] Based on Principal Component Analysis. Genet. Resour. Crop Evol. 66, 951–964. https://doi.org/10.1007/s10722-019-00757-x (2019).
Singh, A. K., Singh, S., Singh, R. S., Joshi, H. K. & Sharma, S. K. Characterization of Bael (Aegle marmelos) varieties under rainfed hot Semi-Arid environment of Western India. Indian J. Agric. Sci. 84 (10), 1236–1242. https://doi.org/10.56093/ijas.v84i10.44206 (2014).
Parihar, N. Variability Studies in Bael (Aegle marmelos Correa) Gene Pool of Kymore Plateau and Satpura Hill Region. M.Sc. Thesis, Jawaharlal Nehru KrishiVishwaVidyalaya, Jabalpur, India (2015).
Singh, P. et al. Genetic diversity and population structure of Bael [Aegle marmelos (L.) Correa] genotypes using molecular markers in the North-Western plains of India. Sci. Rep. 14 (1), 18032. https://doi.org/10.1038/s41598-024-69030-1 (2024).
Crovello, T. J. Analysis of character variation in ecology and systematics. Annu. Rev. Ecol. Evol. Syst. 1, 55–98. https://doi.org/10.1146/annurev.es.01.110170.000415 (1970).
Rodriguez-Delgado, M. A. et al. Principal component analysis of the polyphenol content in young red wines. Food Chem. 78 (4), 523–532. https://doi.org/10.1016/S0308-8146(02)00206-6 (2002).
Samec, D. et al. Assessment of the differences in the physical, chemical and phytochemical properties of four strawberry cultivars using principal component analysis. Food Chem. 194, 828–834. https://doi.org/10.1016/j.foodchem.2015.08.095 (2016).
Murty, B. R. & Arunachalam, V. The nature of genetic divergence in relation to breeding system in crop plants. Indian J. Genet. Plant. Breed. 26, 316–321 (1966).
AOAC. Official Methods of Analysis. The Association of Official Analytical Chemists, 17th edition; Gaithersburg, MD, USA. (2000).
Patel, A. et al. Guidelines for the conduct of test for distinctiveness, uniformity and stability of Bael (Aegle Marmelos Correa). Protection of Plant Varieties and Farmer’s Right Authority. Government India 1–25 (2011).
Mahajan, R. K. et al. Minimal Descriptors of Agri-Horticultural Crops. Part III: Fruit Crops27–31 (National Bureau of Plant Genetic Resources, 2002).
Jolliffe, I. T. & Cadima, J. Principal component analysis: a review and recent developments. Philos Trans A Math Phys Eng Sci 374 20150202. (2065). https://doi.org/10.1098/rsta.2015.0202 (2016).
Acknowledgements
The authors from SKUAST-Jammu are thankful to the Division of Fruit Science, Faculty of Horticulture & Forestry/Faculty of Basic Sciences and Institute of Biotechnology for providing the lab facilities for conducting this research.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
P.S. performed the experiment and wrote the main manuscript; A.S. and R.K.S. designed the experiment; M.S. analysed the statistical data; V.G. S.K.G. and A.J. reviewed the manuscript. All authors contributed to the article.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Singh, P., Sharma, A., Gupta, S.K. et al. Genotypic diversity and nutritional profiling of Bael [Aegle marmelos (L.) Correa] genotypes in India. Sci Rep 15, 31231 (2025). https://doi.org/10.1038/s41598-025-12986-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-12986-5
