
Abstract
Despite advances in understanding the morphological disruptions that lead to defects in palate formation, the precise perturbations within the signaling microenvironment of palatal clefts remain poorly understood. To explore in greater depth the genomic basis of palatal clefts, we designed and implemented the first single cell spatial RNA-sequencing study in a cleft palate model, utilizing the Pax9−/− murine model at multiple developmental timepoints, which exhibits a consistent cleft palate defect. Visium HD, an emerging platform for true single-cell resolution spatially resolved transcriptomics, was employed using custom bins of 2 × 2 μm spatial gene expression data. Validation of spatial gene expression was then validated using custom designed Xenium In Situ mRNA spatial profiling and RNAscope Multiplex assays. Functional enrichment analysis revealed a palate cell-specific perturbation in Wnt signaling effector function in tandem with disrupted expression of extracellular matrix genes in developing mesenchyme. As a key step toward laying the framework for identifying key molecular targets these data can be used for translational studies aimed at developing effective therapies for human palatal clefts.
Similar content being viewed by others
Introduction
Home to some of the most intricate structures in the human body, the craniofacial complex requires tightly controlled molecular crosstalk between proliferating and differentiating cell networks to properly develop1. During embryogenesis, perturbations in the growth environment as well as genetic aberrations can substantially impact this tenuous molecular milieu2. While considerable progress has been made in studying isolated genetic mutations leading to syndromic and non-syndromic craniofacial disorders3,4 the molecular-genetic mechanisms driving morphogenetic events specific to craniofacial structures remain poorly understood. Such gaps in our understanding have restricted clinical treatment options for patients affected by common developmental anomalies, such as cleft lip and palate5. Thus, there is a strong biologic rationale for a thorough investigation of basic molecular mechanisms driving craniofacial structure morphogenesis, which may pave the way toward translatable therapeutic developments for patients.
Among the critical structures of the craniofacial complex, the palate (an anatomic bridge separating oral and nasal cavities) is of particular interest – given its key functional impact in speech and swallowing6. Palatogenesis is a highly regulated and tightly controlled process involving cascades of key genes and signaling pathways which are expressed in an orchestrated manner7. For the palate to properly form in utero, several key developmental milestones must be achieved during the first trimester of gestation. Starting around the fourth week of human development, cranial neural crest cells (cNCCs) migrate from the dorsal edge of the rostral neural tube to form paired maxillary prominences as well as the frontonasal prominence surrounding the primate oral cavity8. The nasal pits then take form, developing into paired medial and lateral nasal processes from the frontonasal prominence by the fifth week. The sixth week then brings about the merging of medial nasal processes with the maxillary processes to form the upper lip and primary palate; simultaneously, bilateral outgrowths from the maxillary processes, termed the palatal shelves, grow vertically along either side of the tongue. Mandibular growth in week seven further allows for descent of the tongue and the elevation of the palatal shelves to a more horizontal position. Proliferation and migration of the palatal shelf progenitor cell populations (epithelium and mesenchyme) leads to the formation of the midline epithelial seam – triggering the onset of the palatal fusion event. By the tenth week, the secondary palate fuses with the primary palate and nasal septum, allowing palatal mesenchyme to differentiate into bony and muscular elements. These fusion processes of primary and secondary palate components are complete by week 12, by which time the secondary palate is divided along the anterior-posterior axis into the bony (hard) palate and the muscular (soft) palate.
To better understand the molecular basis of these key developmental milestones during palatogenesis, genome-wide association studies (GWAS) have identified several transcription factors which influence risk to cleft palate9,10. Among these identified transcription factors is PAX9 (paired homeobox domain-9), a master orchestrator of developmental patterning associated with proper morphogenesis of the axial skeleton, limbs, and craniofacial complex11. Mechanistic studies have pointed to the importance of PAX9-dependent Wnt signaling during palatal development, including its connection to palatal osteogenesis12,13,14. Meanwhile, PAX9’s role in the developmental events immediately preceding elevation and fusion of the palatal shelves has remained relatively underexplored. The aim of the present study was to test the hypothesis that PAX9 regulates the differentiation and patterning of progenitor cell populations prior to the osteogenic switch observed at the stage of palatal fusion. We hypothesized that PAX9’s upstream regulatory role as a patterning transcription factor extends beyond the development of palatal bone, including other mesenchyme-derived cell types and extracellular components.
To address this hypothesis and to better understand the key effector-ligand signaling interactions within the palatal shelves during embryogenesis, we performed the first high-resolution whole-transcriptome spatial RNA-sequencing study focused on the palate. Then, we further dissected the spatial transcriptomic expression profiles of target signaling networks via highly multiplex targeted single cell spatial profiling analyses (Xenium in situ, 10x Genomics, Inc.) using a custom designed panel of 350 unique genes specific to the developing palate. Finally, the key genetic targets identified from these high scale gene expression assays were validated via high-resolution multiplex RNAscope immunofluorescent in situ hybridization. Together, these analyses unveil a novel role of the transcription factor PAX9 in the embryonic palate.
Results and discussion
Whole transcriptome single cell Spatial sequencing in cleft palate model reveals disruption in Wnt signaling and collagen organization
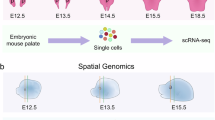
To dissect the spatial gene expression landscape of the embryonic palate at its highest resolution to-date, we profiled embryonic day (E) 12.5 and E13.5 murine palate tissues in both normal (wild-type, WT) and cleft (Pax9−/−) palate model organisms. We generated the first whole transcriptome spatial map of palate development at a single-cell level via Visium HD spatial RNA-seq (10x Genomics, Inc.). Embryos from both time points (WT n = 2 and Pax9−/− n = 2 embryos; each palatal shelf was isolated as its own biological replicate for differential analysis) were processed and sequenced together to avoid batch effect in differential analysis (Fig. 1A).
First, pseudo-bulk analysis was performed to determine general transcriptomic similarity between comparison groups (Fig. 1B). To differentially analyze the molecular impact of the loss of the transcription factor, Pax9, we ran functional enrichment analysis (FEA) for both E12.5 (Fig. 1C) and E13.5 (Fig. 1D) showing an over representation test of the top 50 significant Biological Processes-Gene Ontology (BP-GO) terms. Upon differential comparison of the WT E12.5 to the Pax9−/− palate of the same age, the top biological processes of significant change included: autophagy, synapse assembly; negative regulation of Wnt, Fgf; chondrocyte, cranial skeleton development; macrophage, neurogenesis; and neural crest cell, bone mineralization, and osteoblast differentiation (Fig. 1C). In the E13.5 differential analysis, the loss of Pax9 in the palate exposed significant alterations in collagen fibril organization, Bmp signaling, actin filament assembly; negative regulation of Wnt signaling, cell growth; endothelial cell, muscle cell proliferation; apoptosis, stem cell down-regulation; and metabolic regulation (Fig. 1D).
Spatial mapping of key genes associated with Wnt signaling regulation (Sfrp2) and collagen fibril organization (Col1a1) allowed for quantitative spatial comparison of expression patterns between WT and Pax9−/− palatal shelves (Fig. 1E and F). Each of these marker genes were identified to be downregulated along the most medial regions of the palatal shelves as early as E12.5 in development.
While the most sensitive phenotype in humans carrying heterozygous PAX9 variants is oligontia (congenital absence of six or more permanent teeth), a growing body of literature supports the functional interplay of PAX9 in the development of non-tooth oral structures of neural crest-derived mesenchyme15,16. As the palatal shelves are similarly derived from cranial neural crest cells, the functional role of Pax9 in the setting of failed palatal mesenchymal patterning is likely preeminent.

Whole transcriptome single cell spatial sequencing in cleft palate model reveals disruption in Wnt signaling and collagen organization.(a) Schematic representation of workflow for first whole transcriptome spatial sequencing in normal and abnormal palate development model. (b) Pseudo-bulk principial component analysis (PCA) of WT and Pax9−/− samples. (c-d) Functional enrichment analysis (FEA) showing an over representation test of the top 50 significant Biological Processes-Gene Ontology (BP-GO) terms of (c) E12.5 and (d) E13.5 wild type vs. KO embryonic palate tissues from Visium HD whole transcriptome spatial analysis. e-f) Spatial distribution maps (e) and violin plots (f) of differentially expressed genes, Sfrp2 (Wnt agonist) and Col1a1 (critical extracellular matrix structural component) both in the presence (WT) and absence (KO) of the functional protein, PAX9.
Highly multiplex Spatial transcriptomic analysis of murine embryonic palate using novel computational pipeline
To further analyze the expression patterns of these significant biological processes identified in our whole transcriptome single cell spatial analysis, we designed a highly multiplex spatial mRNA in situ assay for embryonic palate tissues using a custom genetic panel and a novel computational pipeline (Figs. 2A, S4). Xenium In Situ (10x Genomics, Inc.) was employed on a separate batch of age matched and genotype matched samples from our spatial RNA-seq analysis described above. Sub-regional analysis of each palatal shelf included in this targeted validation of spatial sequencing. Relative antero-posterior (AP) depth of section was compared between replicate sections via expression confirmation of Shh (Figure S1a). Furthermore, Ptch1 expression was utilized to highlight the oral palatal domain versus the nasal domain (Fig. 2B) as described previously17. First, the morphological transition from proliferation to elongation characteristic of normal palatal development was visualized with hematoxylin & eosin (H&E) staining and 10X Genomics Xenium Explorer v3.0 on WT embryos in the mid-coronal plane of E12.5 and E13.5 (Fig. 2C). Based on our subregional comparison between developmental stages, volcano plots of differential gene expression were used to highlight statistically significant (P-value < 0.05, |LG2FC| > 1) upregulation (red) and downregulation (blue) of genes identified in transcriptomic comparison of the whole palatal shelves (48 upregulated, 6 downregulated) (Fig. 2D), nasal domains (48 upregulated, 15 downregulated) (Fig. 2E), and oral domains (26 upregulated, 1 downregulated) (Fig. 2F) from E12.5 and E13.5. This spatial transcriptomic roadmap of palatal development from E12.5 to E13.5 allowed for the verification of normal genetic expression trajectories, specifically with the up-regulated expression of extracellular matrix component genes (e.g., Col1a1, Eln, Bgn, etc.) as well as patterning molecules (e.g., Prrx1, Alx1, Dlx1, etc.), allowing us to compare this trajectory with that of the Pax9−/− palatal shelves (see Figure S1b-d).
The Pax9 −/− cleft palate demonstrates disrupted Spatiotemporal genetic programs across oral and nasal domains
Next, we differentially analyzed the spatial transcriptomic profiles of abnormal (cleft, Pax9−/−) across both E12.5 and E13.5 developmental stages via sub-regional highly multiplex mRNA localization (Figure S2A). Aberrancies in the morphological transition from proliferation to elongation in the Pax9−/− cleft palate at both E12.5 and E13.5 were visualized via H&E and Xenium Explorer, highlighting the lack of proper extension of the mesial palatal shelves bilaterally compared to the WT palate of the same age. Volcano plots of differential gene expression highlight statistically significant (P-value < 0.05, |LG2FC| > 1) upregulation (red) and downregulation (blue) of genes identified in transcriptomic comparison of the whole palatal shelves (48 upregulated, 8 downregulated) (Figure S2B), nasal domains (35 upregulated, 9 downregulated) (Figure S2C), and oral domains (60 upregulated, 5 downregulated) (Figure S2D) from E12.5 to E13.5.

Highly multiplex spatial transcriptomic analysis of murine embryonic palate using novel computational pipeline.(a, c) Murine embryos were explanted on embryonic days 12.5 and 13.5 – key stages of palatogenesis – then formalin fixed, paraffin embedded prior to coronal sectioning for spatial transcriptomic pipelines. (b) Expression of Ptch1 clearly defines sub-regions (oral versus nasal domains) of palatal shelves for refined spatial analyses using 10X Genomics Xenium In Situ. (c) Morphological transition from proliferation to elongation characteristic of normal palatal development visualized with H&E staining and 10X Genomics Xenium Explorer v3.0 on E12.5 and E13.5. PR right palatal shelf, PL left palatal shelf, T tongue, TBMand mandibular tooth bud, TBMax maxillary tooth bud. d – f) Volcano plots of differential gene expression highlight statistically significant (P-value < 0.05, |LG2FC| > 1) upregulation (red) and downregulation (blue) of genes identified in transcriptomic comparison of the whole palatal shelves (d), nasal domains (e), and oral domains (f) from E12.5 and E13.5. All spatial transcriptomic analyses derived from n = 4 palatal shelf biological replicates Not pictured: (d) Tnn (p-value = 0.008112, LG2FC = 8.369539); (e) Tnn (p-value = 0.003351, LG2GC = 8.942666); (f) Fmod (p-value = 0.00019, LG2FC = 7.12479).
See also Figure S1.
Palatal shelves in Pax9 −/− model characterized by aberrant expression of homeobox genes, Wnt modulators & effectors, and ECM proteins
Unsurprisingly, where we identified the greatest transcriptomic shift upon differential expression analysis was the comparison between WT and Pax9−/− palatal shelves of the same stage in development (Fig. 3A). Our analysis identified differential gene expression in four key gene categories: homeobox genes (Alx1, Prrx1, Msx1, and Gsc); Wnt signaling modulators (Dkk1, Rspo1, Lgr5, Wif1, Sparc, Prickle1, Tnn, and Jag1); Wnt signaling effectors (Wnt5a and Wnt16); and extracellular matrix proteins (Lum, Ogn, Fmod, Dcn, Eln, Col1a1, Col12a1, Col2a1, Col23a1, Postn, Adamts17, and Palld).
Differential analysis comparing transcription in the whole palatal shelves at E12.5 revealed six differentially expressed genes (Fig. 3B). Of the six differentially expressed genes, one (Dkk1) was upregulated and five (Mapk13, Wif1, Lum, Cntn4, and Nos3) were downregulated in the Pax9−/− embryos relative to WT controls. The nasal palatal domain subregional analysis revealed only one differentially expressed gene, Flt1, to be downregulated in the Pax9−/− model relative to WT control (Fig. 3C). The oral palatal domain subregional analysis at E12.5 revealed 10 differentially expressed genes; one (Dkk1) was upregulated and nine (Tgfb1, Lum, Rspo1, Jag1, Sparc, Wif1, Cntn4, Postn, Col1a1) were downregulated in Pax9−/− cleft (Fig. 3D).
At E13.5, our differential analysis revealed 27 differentially expressed genes within the whole palatal shelves of Pax9−/− embryos compared to WT controls (Fig. 3E). Of the 27 differentially expressed genes, five (Prickle, Phactr1, Chodl, Lgr5, and Wnt16) were upregulated and 25 (Msx1, Lgr5, Gdf10, Col12a1, Adamts17, Igf1, Pdpn, Ogn, Angptl1, Fmod, Fgf7, Dio3, Foxc1, Dcn, Creb3l1, Ndrg2, Palld, Prickle1, Gfra2, Wnt5a, Wif1, Eln, Alx1, and Prrx1) were downregulated. In the nasal subregion at E13.5, 18 differentially expressed genes were identified; three (Prickle1, Enpp2, and Wnt16) were upregulated and 15 (Wnt5a, Tnn, Mapk13, Gsc, Gdf10, Gfra2, Ogn, Foxc1, Fgf7, Eln, Palld, Alx1, Ndrg, Bmp6, and Prrx1) were downregulated (Fig. 3F). In the oral sub-domain, 10 differentially expressed genes were identified, three of which were upregulated (Col2a1, Erg, and Bmp5) while 7 (Col23a1, Wif1, Prrx1, Creb3l1, Fmod, Alx1, Wnt5a) were downregulated in the Pax9−/− cleft palate (Fig. 3G). Taken together, this targeted validation of spatial RNA expression profiles between normal and cleft palate samples identified significant alterations to homeobox gene expression, Wnt signaling modulation, and extracellular matrix structural components (Fig. 3H).
Canonical Wnt/β-catenin signaling has been implicated downstream of Pax9 in the craniofacial complex, perhaps best exemplified by pivotal work demonstrating reduced Wnt activity in palatal shelves in the absence of functional Pax912,13,14. This pathway has been shown to be crucial for intramembranous bone development, which may explain the reversal of cleft palate phenotype with genetic and pharmacological modulation of Wnt signaling in the setting of Pax9 deficiency13,14. However, these studies also helped delineate that Pax9 is involved in other overlapping pathways and processes in addition to Wnt signaling, as observed by the prevailing phenotype of tooth agenesis despite genetic and pharmacologic intervention18. Thus, the identification of homeobox gene and ECM network aberrations in the present study provide additional insights to the expansive role of Pax9 in the developing palate.

Palatal shelves in Pax9−/− model characterized by aberrant expression of homeobox genes, Wnt modulators & effectors, and ECM proteins.(a) Side-by-side visual comparison using H&E staining and 10x Genomics Xenium Explorer v3.0 highlights morphological differences in palatal development from E12.5 to E13.5 in wildtype versus Pax9−/− models. PR right palatal shelf, PL left palatal shelf, T tongue, TBMMand mandibular tooth bud, TBMax maxillary tooth bud. b – d) Volcano plots of differential gene expression highlight statistically significant (P-value < 0.05, |LG2FC| > 1) upregulation (red) and downregulation (blue) of genes identified in transcriptomic comparison of the whole palatal shelves (b), nasal domains (c), and oral domains (d) in E12.5 wildtype versus Pax9−/− models. All spatial transcriptomic analyses derived from n = 4 palatal shelf biological replicates per stage of development. e – g) Volcano plots representing the same sequence of transcriptomic comparisons at E13.5. All spatial transcriptomic analyses derived from n = 4 palatal shelf biological replicates per stage of development.
Xenium in situ RNA localization of differentially expressed genetic markers in Pax9 −/− cleft palate
To better understand the spatial profiles of the quantified transcriptomic findings in this novel computational pipeline, we generated spatial plots and density maps of gene expression to visualize transcript localization within the palatal shelves in both WT and Pax9−/− embryos at E12.5 and E13.5. Spatial maps of homeobox genes (Alx1 and Prrx1) (Fig. 4A), Wnt modulator genes (Tnn and Wif1) (Fig. 4B), Wnt effector genes (Wnt5a and Wnt16) (Fig. 4C), as well as ECM component small leucine-rich proteoglycans (SLRPs) (Lum and Ogn) (Fig. 4D), each delineate aberrant expression patterns in the absence of Pax9 in the embryonic palate. Of note, morphological comparison between samples analyzed in each group was performed using H&E overlaid images of serial tissue sections alongside Xenium In Situ slides, allowing for proper interpretation of the AP axis of expression patterns observed (Fig. 4E). While slight differences in AP depth of sectioning between samples may be present, the region of the posterior secondary palate in the coronal plane remains consistent among all groups compared.
Together, these overlapping signaling networks’ aberrant spatial gene expression patterns in the Pax9−/− palate likely point to the complex interplay and requirement for crosstalk between patterning transcription factors (such as those homeobox proteins identified herein) and Wnt signaling molecules to produce a viable ECM framework. Given the fully penetrant phenotype of cleft secondary palate observed in the Pax9−/− model, this crosstalk may be required to undergo requisite vertical elongation and eventual midline growth to achieve palatal fusion. While the genetic data demonstrated in the present study hint at this potential mechanism (Fig. 4F), further functional studies will be prudent to verify this intrinsic interaction between Pax9, Wnts, and the palatal ECM framework during embryogenesis.
Validation of high throughput Spatial transcriptomic xenium in situ gene expression via RNAscope multiplex
As this study represents one of the very first studies to apply the Xenium In Situ spatial RNA localization technology to-date, it was important for us to further validate our key transcriptomic targets by way of an established RNA profiling platform, as we did in our recent work12,19. To that end, we performed a targeted multiplex RNAscope immunofluorescence in situ hybridization to validate in sub-cellular resolution the expression profiles of key genes in our differential analysis for both developmental stages and genotypes (Figures S3A-C). This included the homeobox genes (Alx1, Prrx1), Wnt modulators and effector genes (Rspo1, Wif1, Sparc, Wnt5a), fibrous ECM protein genes (Eln, Col1a1) and the SLRP family gene, Bgn. All these genes, which were first identified from whole transcriptome and highly multiplex in situ analyses, were validated via RNAscope with concordant expression patterns in all palatal shelves analyzed.
As observed in multiple platforms of spatial transcriptomic analysis, Pax9 may be involved in mesenchymal patterning and organization, with likely contribution to Wnt signaling and ECM structural organization in the developing palate (Fig. 4F)20. As a patterning transcription factor involved early in development, Pax9’s role in recruiting proliferating and differentiating neural crest-derived mesenchyme to form intramembranous bone and its surrounding ECM architecture likely contributes to the observed cleft phenotype in its absence21.

Xenium in situ mRNA localization of differentially expressed genetic pathways in Pax9−/− cleft palate. (a – d) Spatial resolution of homeobox (a), Wnt modulator and effector (b, c), and ECM protein (d) gene transcripts (above) and corresponding density maps (below) validates prior differential gene expression analysis suggesting spatiotemporal disruption of genetic pathways in E12.5 and E13.5 Pax9−/−palatal shelves; (e) Morphological relationship between wide view H&E images (above) and more narrowly focused Xenium transcription maps (below, Alx1 expression shown here as example) of the developing palate at E12.5 and E13.5.; (f) Schematic summary of proposed signaling pathway in the presence of functional Pax9 in embryonic palatal shelves, contributing to the cleft palate phenotype observed.
By employing these next-generation spatial sequencing platforms in the embryonic palate, we hope to empower the craniofacial biology research community with abundant access to these novel datasets. With these tools and the technologies available for downstream analysis, we hope to further identify developmental signaling pathways which go awry in the setting of cleft palate disorders. Together, the results from this study compiling novel statistical and multimodal spatiotemporal transcriptomic analysis approaches reveal a central role for the transcription factor, Pax9, in proper patterning and organization of ECM in the embryonic palate. As a key step toward laying the framework for identifying key molecular targets these data can be used for translational studies aimed at developing effective therapies for human palatal clefts.
Methods
Animals
All animal procedures were approved by the National Institutes of Health, National Institute of Child Health and Human Development Animal Care and Use Committee (ACUC), under Animal Study Protocol (ASP) #21–031. Furthermore, all experiments were performed in accordance with relevant guidelines and regulations. C57BL/6J mice were obtained from the Jackson Laboratory. Inbred strain of female C57BL/6 mice were utilized for all experiments. Pax9+/− mice were provided by Dr. Rulang Jiang (Cincinnati Children’s Hospital) and generated as described previously (Zhou et al. 2013). Timed pregnancies were conducted via vaginal plug identification, with day 0.5 indicating date of identification. To assure accurate developmental stage comparison, embryos comparatively analyzed were of the same litter, with the same C57BL/6 background. Genotyping of all mice included in this study was performed by Transnetyx, Inc, as well as sex determination for inclusion of both male and female sex (at least one biological male and one biological female per comparison group). This study conforms to the ARRIVE 2.0 guidelines.
Visium HD spatial RNA-sequencing
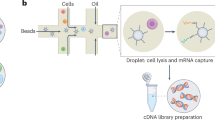
All steps from 10X Genomics’ FFPE Visium HD Spatial workflow were followed. In brief, high-definition spatial RNA sequencing (Visium HD) slides are coated with an array of poly-T primers, which encode unique spatial barcodes. These barcodes contain thousands of encoded oligonucleotides within a catchment frame of 6.5 × 6.5 mm. The oligonucleotides are selectively hybridized with the 3’ end of mRNA eluted upon tissue permeabilization, enabling true single-cell-seq-like mRNA sequencing when binned to 2 × 2 micron resolution. FFPE tissue was sectioned directly onto the barcoded slide, H&E stained, and then underwent enzymatic permeabilization, which allowed for mRNA release and subsequent capture by primer-coated slides. These mRNA molecules are visualized through the incorporation of fluorescent nucleotides into the complementary DNA (cDNA) synthesis process. The resolution of unique spatial barcoding in situ allows the matching of RNA abundance with the original spatial location in the tissue section, providing a whole-transcriptome RNA-sequencing at single-cell resolution with 2-dimensional spatial relation.
Xenium in situ mRNA localization and analysis
Xenium In Situ sub-cellular mRNA detection technology was employed as previously reported following all manufacturer guidelines and specifications22. We custom designed a 350-plex targeted gene panel as previously reported12 to detect mRNA expression for cell type identification as well as signaling pathway interactions selected and curated primarily based on single cell atlas data we generated in our previous work on palate development19. For this assay, we formalin fixed and paraffin embedded 2 embryos of Pax9−/− genotype and 2 embryos of Pax9+/+ genotype (each palatal shelf was isolated as its own biological replicate for differential analysis) at embryonic days 12.5 and 13.5. Whole heads were processed, embedded, and sectioned (4 μm) to the level of the 1 st molar tooth bud. All 4 samples were assayed in a single batch under identical conditions. The post-Xenium H&E staining followed Demonstrated Protocol CG000160 from 10x Genomics, Inc.
Computational analysis of visium HD data
We created custom bins of 2 × 2 μm Visium HD spatial gene expression data based on the identified cell nuclei in the microscope image of the tissue used in the Visium HD assay. The cell nuclei were automatically segmented using the StarDist AI model, and improperly segmented nuclei were filtered by assessing nuclei size and unique molecular identifier (UMI) counts. We directly removed mitochondrial (mt)RNA reads to avoid data skewing, simplify analysis, and focus on specific nuclear RNA transcript populations as previously exemplified23,24. Gene expression was quantified at single-cell resolution by mapping spatial transcriptomic data to the segmented nuclei. Downstream analysis was conducted using Seurat V5 in R, including normalization and scaling based on UMI counts via the SCTransform (v2) algorithm. We identified highly variable genes across the segmented single cells, performed principal component analysis (PCA) with pseudo-bulk function, uniform manifold approximation and projection (UMAP), and unsupervised clustering, and identified differentially expressed cell-type-specific markers. Specifically, the differentially expressed markers were identified using the “roc” method within Seurat V5, which evaluates each gene’s ability to distinguish between two cell groups using receiver operating characteristic (ROC) curve analysis, returning a ranked matrix of predictive power based on area under the curve (AUC) values.
Differentially expressed genes (DEGs) in E12.5 WT vs. E13.5 WT, E12.5 WT vs. E12.5 KO, and E13.5 WT vs. E13.5 KO were identified using Model-based Analysis of Single-cell Transcriptomics (MAST) in R, with the criteria of |LG2FC| > 1 and false discovery rate (FDR) < 0.01. The identified DEGs were subsequently used for over-representation analysis (ORA) to identify enriched biological functions through the Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) databases using the clusterProfiler package, with criteria of P-value < 0.05 and gene counts > 1.
Computational analysis of xenium in situ data
At the time of this analysis, no publicly available open-access software would allow us to perform the statistical tests necessary to achieve our project aims. Thus, all analyses were generated using a custom designed data pipeline in Microsoft Excel.
Our analysis began in the 10x Genomics open-access software, Xenium Explorer v3.0.
First, we sought out to develop a method for reliably dividing the embryonic palatal shelves into nasal palatal (NP) and oral palatal (OP) regions for more precise region-specific analysis. We were able to define a reproduceable oronasal axis in both our WT and Pax9−/−mouse models by isolating the co-expression of Shh and Ptch1 on the Xenium Explorer v3.0 density map. During embryonic development, Shh expression is restricted to the epithelium of the oral side of the palatal shelves27 while known target gene, Ptch1, is localized to the nearby mesenchyme.28 Thus, by isolating the co-expression of Shh and Ptch1, we were able to successfully distinguish the oral region of the palatal shelves from the nasal region.
Next, we defined our regions of interest (ROIs). ROIs were standardized by surface area (6400 um2 +/- 100 μm). Three ROIs were selected from each side of the oronasal axis, per palatal shelf; thus, a total of 12 ROIs were generated per section. The ROIs were exported as.csv files and opened using Microsoft Excel. Raw data sets were produced using the transcript density data from these files.
To perform differential gene expression (DGE) analysis, we calculated both p-values and Log2 fold-change (LG2FCs) from our transcript density data sets. While the p-value is a measure of the statistical significance of the expression difference between comparison groups, the LG2FC is an estimate of the log2 ratio of the change in expression12. Genes that displayed a p-value < 0.05 and LG2FC > 1 were considered differentially expressed for each of our comparison groups.
LG2FCs were calculated using raw data. By contrast, to obtain p-values, we first normalized our data using a LG2(x + 1) transformation to reduce variance. We then tested for equal variance using an F-test (F-critical = 0.05). Based on the F-statistic, a student’s (equal variance) or Welch’s (unequal variance) T-test was deployed. The Benjamini-Hochberg procedure was performed to obtain p-values adjusted for false discovery rates. Finally, the LG2FC and -Log10(p-value) were mapped onto volcano plots to visually represent the differentially expressed genes.
Data availability
All results and analysis data are available in the main text or the supplementary file. Additional information and materials are available from the corresponding author upon reasonable request. All raw data from spatial RNA-sequencing is accessible open-access at the accession: GSE284271. The codes for analysis of 10x Visium HD Spatial RNA-sequencing were deposited into the GitHub at https://github.com/LabOnoM/GSE284271.
Materials availability
All materials used for this study have been reported in the Methods section; any further questions regarding materials can be directed to the first author of correspondence, Dr. Jeremie Oliver Piña (jeremie.pina@nih.gov).
References
Vanneste, M. et al. Syndrome-informed phenotyping identifies a polygenic background for achondroplasia-like facial variation in the general population. Nat. Commun. 15, 10458. https://doi.org/10.1038/s41467-024-54839-1 (2024).
Zhang, W. et al. Proteomic analysis illustrates the potential involvement of dysregulated ribosome-related pathways and disrupted metabolism during retinoic acid-induced cleft palate development. BMC Med. Genomics. 17 https://doi.org/10.1186/s12920-024-02054-8 (2024).
Xu, X. et al. Genetic variants in METTL16 affect the risk of non-syndromic orofacial clefts. Birth Defects Res. 116, e2403. https://doi.org/10.1002/bdr2.2403 (2024).
Haller, J., Abedi, N., Hafedi, A., Shehab, O. & Wietecha, M. S. Spatial transcriptomics unravel the tissue complexity of oral pathogenesis. J. Dent. Res. 103, 1331–1339. https://doi.org/10.1177/00220345241271934 (2024).
Oliver, J. D. et al. Molecular diagnostics and in utero therapeutics for orofacial clefts. J. Dent. Res. 99, 1221–1227. https://doi.org/10.1177/0022034520936245 (2020).
Best, D. L., Yau, V., Manon, V., Noto, S. & Perry, R. J. Management of persistent hypernasality after pharyngeal flap: A novel V-to-Y revision procedure to tighten the lateral ports. Cleft Palate Craniofac. J. 10556656241305227 https://doi.org/10.1177/10556656241305227 (2024).
Reynolds, K. et al. Wnt signaling in orofacial clefts: crosstalk, pathogenesis and models. Dis Model Mech 12. (2019). https://doi.org/10.1242/dmm.037051
Oliver, J. D. et al. Innovative molecular and cellular therapeutics in cleft palate tissue engineering. Tissue Eng. Part. B Rev. 27, 215–237. https://doi.org/10.1089/ten.TEB.2020.0181 (2021).
Siewert, A., Hoeland, S., Mangold, E. & Ludwig, K. U. Combining genetic and single-cell expression data reveals cell types and novel candidate genes for orofacial clefting. Sci. Rep. 14, 26492. https://doi.org/10.1038/s41598-024-77724-9 (2024).
Beaty, T. H., Marazita, M. L. & Leslie, E. J. Genetic factors influencing risk to orofacial clefts: today’s challenges and tomorrow’s opportunities. F1000Res 5, 2800. (2016). https://doi.org/10.12688/f1000research.9503.1
Marazita, M. L. et al. Genome scan, fine-mapping, and candidate gene analysis of non-syndromic cleft lip with or without cleft palate reveals phenotype-specific differences in linkage and association results. Hum. Hered. 68, 151–170. https://doi.org/10.1159/000224636 (2009).
Pina, J. O. et al. Spatial Multi-omics reveals the role of the Wnt modulator, Dkk2, in palatogenesis’. J. Dent. Res. 103, 1412–1420. https://doi.org/10.1177/00220345241256600 (2024).
Jia, S., Zhou, J. & D’Souza, R. N. Pax9’s dual roles in modulating Wnt signaling during murine palatogenesis. Dev. Dyn. 249, 1274–1284. https://doi.org/10.1002/dvdy.189 (2020).
Jia, S. et al. Small-molecule Wnt agonists correct cleft palates in Pax9 mutant mice in utero. Development 144, 3819–3828. https://doi.org/10.1242/dev.157750 (2017).
Pak, B. et al. Pax9 is essential for granulopoiesis but dispensable for erythropoiesis in zebrafish. Biochem. Biophys. Res. Commun. 534, 359–366. https://doi.org/10.1016/j.bbrc.2020.11.077 (2021).
Swartz, M. E., Sheehan-Rooney, K., Dixon, M. J. & Eberhart, J. K. Examination of a palatogenic gene program in zebrafish. Dev. Dyn. 240, 2204–2220. https://doi.org/10.1002/dvdy.22713 (2011).
Cobourne, M. T. et al. Sonic Hedgehog signalling inhibits palatogenesis and arrests tooth development in a mouse model of the nevoid basal cell carcinoma syndrome. Dev. Biol. 331 (1), 38–49. https://doi.org/10.1016/j.ydbio.2009.04 (2009).
Li, C., Lan, Y., Krumlauf, R. & Jiang, R. Modulating Wnt signaling rescues palate morphogenesis in Pax9 mutant mice. J. Dent. Res. 96, 1273–1281. https://doi.org/10.1177/0022034517719865 (2017).
Pina, J. O. et al. Multimodal Spatiotemporal transcriptomic resolution of embryonic palate osteogenesis. Nat. Commun. 14, 5687. https://doi.org/10.1038/s41467-023-41349-9 (2023).
Gaur, T. et al. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 280, 33132–33140. https://doi.org/10.1074/jbc.M500608200 (2005).
Zhou, J., Gao, Y., Lan, Y., Jia, S. & Jiang, R. Pax9 regulates a molecular network involving Bmp4, Fgf10, Shh signaling and the Osr2 transcription factor to control palate morphogenesis. Development 140, 4709–4718. https://doi.org/10.1242/dev.099028 (2013).
Janesick, A. et al. High resolution mapping of the tumor microenvironment using integrated single-cell, Spatial and in situ analysis. Nat. Commun. 14, 8353. https://doi.org/10.1038/s41467-023-43458-x (2023).
Naraine, R. et al. Evolutionary conservation of maternal RNA localization in fishes and amphibians revealed by TOMO-Seq. Dev. Biol. 489, 146–160. https://doi.org/10.1016/j.ydbio.2022.06.013 (2022).
Iegorova, V., Naraine, R., Psenicka, M., Zelazowska, M. & Sindelka, R. Comparison of RNA localization during oogenesis within acipenser ruthenus and xenopus laevis. Front. Cell. Dev. Biol. 10, 982732. https://doi.org/10.3389/fcell.2022.982732 (2022).
Xu, J., Iyyanar, P. P. R., Lan, Lan. & Jiang, R. Sonic Hedgehog signaling in craniofacial development. Differentiation 133, 60–76. https://doi.org/10.1016/j.diff.2023.07.002 (2023).
Lan, Y. & Jiang, R. Sonic Hedgehog signaling regulates reciprocal epithelial-mesenchymal interactions controlling palatal outgrowth. Development 136, 1387–1396. https://doi.org/10.1242/dev.028167 (2009).
Acknowledgements
We thank Dr. Sergey Leiken (NIH/NICHD) for his technical and analytical support in establishing our independent next-generation sequencing workflow and Xenium in situ assays; Dr. Blake Warner (NIH/NIDCR) for his support in Xenium instrument access; Dr. James Iben (NIH/NICHD) and Dr. Fabio R. Faucz (NIH/NICHD) for their bench support for Visium HD experiments; Dr. Vincent Schram (NIH/NICHD) for immunofluorescence imaging support. We also thank Dr. Shawn Hallett (University of Michigan) and Mr. Caris Smith (University of Alabama, Birmingham) for their scientific review and feedback on this work.
Funding
Open access funding provided by the National Institutes of Health. ZIA HD009015 “Wnt Signaling Pathway: Genes in Craniofacial Development: Palate and Tooth”, National Institutes of Health, National Institute of Dental and Craniofacial Research (RDS); National Institutes of Health, National Institute of Child Health and Human Development, Intramural Research Training Award (JOP).
Author information
Authors and Affiliations
Contributions
Conceptualization, JOP, RR, ES, RDS; Methodology, JOP, RR, ES, RDS; Software, JOP, RR, ES, ZW, MO; Validation, JOP, RR, ACM, PC; Formal Analysis, JOP, RR, ES, ZW, MO; Investigation, JOP, RR, ES, ACM, ZW, MO, PC, MF, RDS; Resources, JOP, RDS; Data Curation, JOP, RR, ES, ZW, MO; Writing – Original Draft, JOP; Writing – Review & Editing, JOP, RR, ES, ACM, ZW, MO, PC, MF, RDS; Visualization, JOP, RR, ES; Supervision, RDS; Project Administration, JOP, PC, RDS; Funding Acquisition, JOP, RDS.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Piña, J.O., Raju, R., Stipano, E. et al. Single cell spatial transcriptomics links Wnt signaling disruption to extracellular matrix development in a cleft palate model. Sci Rep 15, 29639 (2025). https://doi.org/10.1038/s41598-025-14807-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-14807-1
