
Abstract
Stand-replacing wildfire is the primary natural disturbance in jack pine-dominated boreal forests; but clearcut harvest also emulates this natural renewal process. We used a 60-year clearcut harvest chronosequence to assess whether soil communities became more similar to those in wildfire-origin stands over time. We assessed convergence across disturbance types at each stand development stage and recovery compared to the wildfire mature stand development stage (~ 85 years). To evaluate cumulative effects, we also assessed a 20-year salvage harvest chronosequence where wildfire was followed by salvage logging of fire-killed trees. Beta diversity analyses showed different recovery times among soil taxa. Following clearcut harvest, bacteria converged to wildfire reference conditions more quickly, followed by arthropods, whereas fungi did not converge within the study period. Soil communities in salvage-logged sites diverged from clearcut harvest and wildfire references suggesting compounded disturbance effects. This work showcases how highly-scalable DNA metabarcoding and bioinformatic tools can be applied to simultaneously monitor a diverse array of soil biota. In future work, tracking fungal and arthropod soil communities may provide more insights into the longer-term effects of current forest management practices and provide guidance when comparing alternative approaches.
Similar content being viewed by others

Introduction
In Canada’s eastern boreal forest, jack pine (Pinus banksiana Lamb.) is a commonly occurring species that grows in pure to nearly pure stands on coarse-textured, glaciofluvial outwash deposits and is a staple species that provides both sawlog and pulpwood feedstock1. It is an intolerant, pioneer species well adapted to stand-replacing wildfire, due in large part to having serotinous cones that release seed following high temperatures associated with wildfire resulting in high density pine-dominated stands. Silvicultural systems that emulate these natural disturbance processes are used as a model for sustainable forest management2.
Forest management strategies that integrate stand-replacing disturbance like full-tree clearcut harvest may emulate certain aspects of a stand-replacing event such as natural wildfire (e.g., landscape pattern emulation). Clearcut harvest removes the overstory, but retains the existing downed woody debris and forest floor3. Wildfire, on the other hand, is a pyrogenic process that kills trees but retains an overstory structure and consumes a portion of the forest floor as well as existing downed woody debris. Another forest operation currently used is post-wildfire salvage harvest that results in the partial consumption of the forest floor and downed woody debris by the stand-replacing wildfire followed by the removal of the fire-killed overstory trees.
The motivation for salvage harvesting following wildfire is to ameliorate economic losses by salvaging wood volumes that would have otherwise been lost through decomposition or blowdown following the wildfire event4,5. Salvage harvesting is controversial, however, because studies have shown that post-wildfire salvage may act more like a cumulative disturbance4,6,7,8,9,10. Specifically, salvage harvesting has been shown to: (1) reduce the ecosystem benefits provided by natural disturbance, (2) have negative impacts on many taxa, especially saproxylic species that require dead or dying wood for some part of their life cycle, (3) impair ecosystem recovery, and possibly alter ecosystem stability (i.e., not returning to the pre-disturbance condition), and (4) may result in additive effects associated with two disturbance events happening within 2–3 years4,10.
The successional dynamics of boreal forest below-ground communities such as bacteria, fungi, and arthropods in response to different disturbance types remain poorly understood. Understanding the below-ground impacts of above-ground harvest and silviculture relative to wildfire is needed to more fully evaluate the sustainable management of boreal forest systems3. Examples of post-disturbance recovery dynamics that occur below ground include nutrient cycling processes such as decomposition, mycorrhizal symbioses, and soil respiration11,12,13,14. Reference information on soil biotic succession after natural wildfire disturbance can then be used to evaluate and compare the convergence and recovery trajectories after anthropogenic disturbances such as clearcut harvesting or salvage logging. Though there have been many studies that have examined the effects of fire and harvest disturbances on specific above-ground communities such as plants, animals, and insects10,15,16,17,18,19,20,21,22much less is known about below-ground communities largely because they tend to be difficult to study using conventional methods23,24,25,26. Advances in molecular biodiversity monitoring, high throughput sequencing of signature DNA regions, also known as DNA metabarcoding, provides a rapid means for surveying the diverse soil organisms from bulk soil samples such as bacteria, fungi, and soil arthropods, that play important roles in nutrient cycling and soil remediation following disturbance and have well-developed molecular tools for their detection and identification27,28,29,30.
In our previous study of soil biotic succession following stand-replacing wildfires, we used DNA metabarcoding to show that each successional stage had unique biotic communities31. Here, we extend our original study by comparing our post-wildfire soil successional reference with soil bacterial, fungal, and arthropod succession following full tree clearcut harvest (60 year chronosequence) and salvage harvest (20 year chronosequence). We used the chronosequence approach, where space is substituted for time, to evaluate soil community succession along the successional sequence (i.e., stand development stages). The objectives of this study were to: (1) assess whether soil communities become more similar over time by systematically analyzing community composition at each stand development stage, between the natural wildfire reference versus clearcut harvest or salvage harvest sites, (2) determine if/when the soil communities recover and return to the post-wildfire mature stand development stage, the natural disturbance endpoint in stand development, and (3) assess the effect of salvage logging (i.e., cumulative disturbance) on early soil community development. Further, we examined how various soil chemical properties correlate with these community changes.
Methods
Study site descriptions
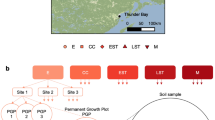
In this paper, we compare the below-ground successional patterns in post-wildfire and post-harvest sites. A total of 31 sites were included in this study (Fig. 1) including 14 wildfire origin stands, 11 full-tree clearcut harvest sites, and 6 post-wildfire salvage harvest stands. All sites are in the Lake Nipigon ecoregion, a cold, continental climate region, with annual mean temperatures of 1.0–1.8 °C, Growing Degree-Days (GDD) from 1112 to 1263, and annual precipitation ranging from 68 to 82 cm yr−1 (growing season precipitation from 41 to 49 cm yr−1)32. Soils for all sites had comparable parent soil materials, well-drained, outwash sands, with well-developed Orthic Humo-Ferric Podzolic profiles33. Stands were jack pine-dominated (Pinus banksiana) with many having minor amounts of black spruce (Picea mariana), trembling aspen (Populus tremuloides), and white birch (Betula paperifera). Wildfire and harvested sites generally had understories dominated by mosses, while salvage harvesting sites had litter and lichen dominated understories. Most stands are classified as Site Class I34 with estimated site index values ranging between 18 and 22 m at breast height age 50.

Sites and experimental design. (a) Inset map shows the Great Lakes Region, Ontario, Canada. Main map shows the distribution of sampling sites by disturbance type and stand development stage. Scale bar shows 50 km increments. Adjacent table summarizes the years since disturbance and the number of sites sampled for each disturbance type. (b) Summary of community composition comparisons at each stand development stage to assess whether harvest and wildfire sites become more similar over time (convergence) and comparisons with the reference wildfire condition at the mature stand development stage to assess whether sites return to the natural post-wildfire successional endpoint (recovery). (c) Summary of community composition comparisons with the wildfire reference condition and comparison of harvest types at each stand development stage. Abbreviations: E = establishment, CC = crown closure, EST = early self-thinning, LST = late self-thinning, M = mature. © Stadia Maps © OpenMapTiles © OpenStreetMap © Stamen Design.
Each of the wildfire and salvage sites experienced spring, stand-replacing fires with Fire Weather Index (FWI) values ranging from 17.8 to 44.6 (exceeds extreme) in wildfire sites and 12.5 to 44.6 (exceeds extreme) in salvage sites. These sites were grouped along a time-since-disturbance sequence that correspond to stand development stages (establishment: 0–15 years, crown closure: 16–30, early self-thinning: 31–45, late self-thinning: 46–70, mature: 71+) for jack pine that were comparable between the three disturbance types. Clearcut harvesting has only been operational for the past approximately 70 years, so our sites only have establishment to late self-thinning stages for this disturbance type. Post-wildfire salvage operations have only become commonplace in the last 20 years in Ontario’s northwest region, so our sites only have establishment to crown closure stages for this disturbance type.
A chronosequence is a space for time substitution approach that is often used when the time frame of the study question exceeds the life span of investigators. This approach is suitable for studying succession across decadal timescales when sites at different ages are following the same successional trajectory35. The underlying assumption of this approach is that the primary differentiating factor between samples within a chronosequence is time since disturbance35. To conform to the assumption noted above, our principal selection criterion was based on similarity in the basic site characteristics (e.g., climate, soils, tree species composition, and productivity) described previously in more detail31. Sites were jack pine-dominated (Pinus banksiana) separated by at least 1 km and represent independent stands, though some were the result of the same large-scale wildfire events.
Soil sampling and laboratory analysis
The original field sampling design, including a detailed description of our sampling approach, are available31. Briefly, in 2017, within each sampling site shown in Fig. 1, three replicate bulk soil samples were collected and pooled to help account for heterogeneity. The bryophyte, organic LFH (litter, fermentation, and humus), and upper mineral soil layers were kept separate at this stage. Samples were immediately frozen at −20˚C after sampling and held at that temperature until analysis. Additional samples of the LFH and mineral soil horizons (Ae, Bf collected separately) were also collected for pH and chemical determinations as previously described (Table S1).
Molecular biology methods and sequencing
The molecular biology and sequencing methods were previously described in detail31. Briefly, DNA extractions performed by the Natural Resources Canada lab at the Great Lakes Forestry Centre used the MoBio PowerSoil DNA Isolation Kit following the manufacturer’s instructions except that 200 ul of 100 mM AlNH4(SO4)2 was added to the tube along with soil and Solution C1 followed by a 10 min incubation at 70 °C to help lyse cells36. The bacterial community was enriched using PCR by targeting the 16S v4-v5 region by prefixing the 515F-Y and 926R primers with Illumina adapters37; the fungal community was enriched by targeting the ITS2 region by prefixing the ITS9 and ITS4 primers with Illumina adapters38,39. Invertebrate communities were enriched using two sets of primers targeting portions of the COI DNA barcode region to help account for known primer bias to detect a broader array of arthropods40. The F230R_modN marker was targeted by prefixing the LCO1490 and the 230R_modN primers with Illumina adapters41,42; and the BE marker was targeted by prefixing the B and E primers with Illumina adapters43. For each sample a PCR cocktail of 5 µl template DNA (5 ng/µl in 10 mM Tris pH 8.5), 0.5 µl each of 10 µM forward and reverse primer, 25 µl HotStar Taq plus master mix kit 2x, and 19 µl of sterile water. PCR cycling conditions were as follows: 95 °C for 5 min followed by 30 cycles (16S) or 35 cycles (ITS) or 40 cycles (COI) of 95 °C for 30 s, 50 °C for 30 s, and 72 °C for 1 min, followed by a final extension of 72 °C for 5 min (10 min for COI) and hold at 4 °C. PCR products were visualized then purified using a magnetic bead solution (Agencourt AMPure XP, Beckman Coulter Life Science, Indianapolis, IN, USA) according to Illumina’s protocol44. Sample indexes were added by amplifying 5 µl of the purified PCR product with 25 µl of KAPA HIFI HotStart Ready Mix, 5 µl of each Nextera XT Index Primer (Illumina Inc., San Diego, CA, USA) and 10 µl of UltraPure DNase/RNase-Free Distilled Water for a total volume of 50 µl. Thermal cycling conditions were as follows: 3 min at 98 °C, 8 cycles of 30 s at 98 °C, 30 s at 55 °C, 30 s at 72 °C, and a final elongation step of 5 min at 72 °C. Indexed amplicons were purified, quantified using a Qubit dsDNA BR Assay Kit (Life Technologies), and combined at equimolar concentration. Paired-end sequencing (2 × 250 bp) of the pools was carried out on an Illumina MiSeq at the National Research Council Canada, Saskatoon. 15% PhiX was added to help compensate for low sequence heterogeneity on the plate.
Bioinformatic methods
Bioinformatic methods were previously described in detail31. Briefly, demultiplexed, paired-end Illumina reads were analyzed on the General Purpose Science Cluster at Shared Services Canada. Metabarcodes were processed using the MetaWorks v1.9.3 multi-marker metabarcode pipeline available from https://github.com/terrimporter/MetaWorks using the standard sequence variant workflow45. Reads were paired using SEQPREP v1.3.2 using the default parameters except that the Phred score quality cutoff was set to 20 and the minimum overlap was set to 25 bp46. Primers were trimmed using CUTADAPT v3.2 using the default parameters except that the minimum sequence length was set to 150 bp, Phred quality score cutoffs at the ends was set to 20, and the maximum number of N’s was set to 347. Reads were dereplicated and denoised using VSEARCH v2.15.2, setting the minimum cluster size to retain to 3, so clusters with only 1 or 2 reads were removed as noise48. The 16S metabarcodes were taxonomically assigned using the RDP Classifier v2.13 using the built-in 16S reference set49. The ITS metabarcodes were taxonomically assigned using the RDP Classifier with a custom-trained reference set based on the UNITE v8.2, QIIME release for Fungi v 04.02.2020, available from https://github.com/terrimporter/UNITE_ITSClassifier50. The COI metabarcodes were taxonomically assigned using the RDP classifier with a custom-trained COI classifier v4 reference set available from https://github.com/terrimporter/CO1Classifier51. In total we sequenced 49,721,154 paired-end reads for this study. After read pairing, primer trimming, and other quality control steps to remove poor quality and artefactual sequences, we retained 10,077,185 reads clustered into a total of 114,968 sequence variants (SVs) (Table S2).
Data analysis
All data analyses was conducted in RStudio 2021.09.0 build351 using R v4.1.152,53. Diversity analyses were carried out with the vegan 2.6-2 package54. For each sample, sequence reads were rarefied using the ‘rarecurve’ function to ensure that sequencing depth was sufficient across samples (Figure S1). In subsequent analyses, additional rare SVs that comprise less than 0.01% of reads were removed to account for the possibility of tag switching events55. Also in all subsequent analyses, for each sample, sequences from each soil layer were pooled together so that we could assess changes in composite diversity. After this filtering step we retained 1,671 SVs comprised of 6,642,061 reads. Although each SV was associated with a taxonomic assignment to the species (fungi, arthropods) or genus (bacteria) level, all subsequent analyses were conducted at the sequence variant level associated with an alphanumeric label that generally represents a finer-level of resolution than an operational taxonomic unit (OTU) or species56. Alpha diversity was measured by sequence richness, defined as the number of unique sequences detected at each site. We did not calculate any measure of evenness or abundance as metabarcoding read counts do not necessarily reflect abundance for taxa where there is a large variation in body-size such as for fungi and arthropods and due to primer bias57,58. SV richness was assessed using the ‘specnumber’ function. Normality was assessed visually using the ‘ggqqplot’ function from the ggpubr 0.4.0 library and the base R Shapiro-Wilk test ‘shapiro.test’ function, and either Wilcox or t-tests were used to compare samples against the reference condition59. “Holm” adjusted p-values are shown for multiple comparisons. For each taxon, a ‘global’ comparison of beta diversity for samples across all stand development stages and site characteristics were conducted as follows: Binary Bray Curtis (Sorensen) dissimilarity matrices were created using the ‘vegdist’ function with binary = TRUE54. Non-metric multidimensional scaling (NMDS) plots were calculated using the ‘metaMDS’ function using 3 dimensions after running ‘dimcheckNMDS’ from the goeveg 0.5.1 package and setting trymax = 10060. Stress was assessed using the ‘stressplot’ function. Dispersion was assessed using the ‘betadisper’ function and the base R ‘anova’ function. Environmental variables were fit to the ordination using the ‘envfit’ function with 999 permutations, and only variables with a p-value < 0.05 were plotted. To facilitate readability, samples in each ordination were faceted (subset) by stand development stage while keeping the same axis and ‘global’ correlation (arrow) scaling across each subset plot. We checked for significant interactions between disturbance type and stand development stage using permutational analysis of variance (PERMANOVA) using the ‘adonis2’ function with 999 permutations. When checking for convergence, at each stand development stage, we conducted pairwise comparisons of clearcut harvest disturbance with the wildfire reference condition using the ‘pairwise.adonis’ function with the pairwiseAdonis 0.4 wrapper for multi-level pairwise comparisons using adonis2 with 999 permutations and ‘Holm’ corrected p-values61. When checking for recovery, for each major group of soil taxa, we conducted pairwise comparisons of each stand development stage with respect to the mature wildfire reference condition. All plots were created using ggplot2 3.4.4 and the map was created using ggmap 4.0.062,63.
Results
In general, the soil community was taxonomically diverse, especially for bacteria and fungi that tended to be dominated by a handful of high-level taxa (Figure S2). Across all sites, common taxa included bacterial Alphaproteobacteria, Acidobacteria, Chintinophagia, Gammaproteobacteria, and Planctomycetacia; fungal Agaricomycetes and Leotiomycetes; and arthropod Insecta and Arachnida.
Establishing the reference condition: succession following wildfire
Soil bacterial, fungal, and arthropod succession as well as community assembly patterns following natural wildfire disturbance were previously described in detail31. In that study, bacterial communities were the first to recover after wildfire, returning to a community structure similar to the wildfire reference condition at the mature stand development stage by the crown closure stage (~ 20 years). Bacterial communities were found to have a large core community with many shared taxa across each stand development stage. Moreover, bacterial community shifts were significantly correlated with changes in pH, total nitrogen, and total organic carbon. Fungi and arthropods, however, shared smaller core communities, i.e. had fewer taxa in common across stand development stages. In contrast with bacterial communities, fungal and arthropod communities had more taxon-replacement from one stand development stage to the next, i.e., each stand development stage supported more unique biodiversity compared with bacterial communities. Soil fungi and arthropod community shifts were significantly correlated with changes in total nitrogen and total organic carbon. Bacterial, fungal, and arthropod stand development stage bioindicator taxa were detected.
Assessing convergence and recovery between wildfire and clearcut harvest stands
Convergence was compared between full tree clearcut harvest stands with the wildfire reference condition at the same stand development stage, from establishment to late self-thinning (Fig. 2 (a)). At the establishment and early self-thinning stages, the bacterial, fungal, and arthropod communities had statistically significant higher richness in clearcut versus wildfire sites (adjusted p-value < 0.05). We also assessed recovery of clearcut harvest sites with respect to the mature, wildfire reference condition (Fig. 2 (b)). Richness recovery patterns were not clear and varied by taxonomic group. For bacteria, richness in clearcut harvest sites were similar with the mature wildfire reference condition across all stand development stages except late self-thinning (adjusted p-values > = 0.05). For fungi and arthropods, richness appeared to recover by the late self-thinning stand development stage.

Analysis of richness did not reveal clear patterns of convergence or recovery. Sequence variant richness was compared between the wildfire reference condition and full tree clearcut harvest sites to check for (a) convergence and (b) recovery across stand development stages. Wilcox or t-tests were used to compare soil communities and an asterisk is shown if the Holm corrected p-value was < 0.05. For part (a), statistical comparisons were grouped by taxon and stand development stage to compare richness between clearcut harvest and wildfire. For part (b), statistical comparisons were grouped by taxon to compare wildfire richness at each stand development stage with wildfire richness at the mature stand development stage. Abbreviations: E = establishment, CC = crown closure, EST = early self-thinning, LST = late self-thinning, M = mature.
Using beta diversity, instead of richness, allowed us to detect clearer patterns of convergence. Indirect gradient analysis was used to visualize convergence of beta diversity patterns following full tree clearcut harvest. NMDS plots show how community composition at each stand development stage shifted for each organismal group (bacteria stress = 0.08 and linear R2 = 0.98; fungi stress = 0.11 and linear R2 = 0.94; arthropod stress = 0.13 and linear R2 = 0.92) (Fig. 3 (a)). For each panel, an asterisk in the top right corner indicates when communities are significantly different between the disturbance types (adjusted p-value < 0.05). The lack of statistical significance between successional stages (adjusted p-value > = 0.05), signifies when putative convergence in beta diversity occurred for each taxon. Based on this analysis, soil bacteria beta diversity appeared to converge at the crown closure stage, and this was correlated by changes in pH. For instance, at the establishment stage, wildfire bacterial communities were correlated with higher pH, whereas clearcut harvest bacterial communities were correlated with lower pH. By the crown closure stage onwards, bacterial communities from wildfire and clearcut harvest sites appeared to be more similar and wildfire bacterial communities correlated with lower pH. Soil fungal beta diversity from wildfire and clearcut harvest sites remained distinct across all stand development stages from establishment to late self-thinning (p-values < 0.05) indicating a that a longer time may be needed to observe convergence. For soil fungi at the establishment stage, wildfire communities were correlated with higher pH, whereas clearcut harvest communities were correlated with lower pH, higher total organic carbon and total nitrogen. By the crown closure stage onwards, fungal communities from wildfire and clearcut harvest sites appeared to be more similar with wildfire fungal communities correlated with lower pH and higher total organic carbon (TOC) over time. Soil arthropod beta diversity appeared to converge at the early self-thinning stage. Arthropod communities at the establishment stage following wildfire were correlated with higher pH, whereas clearcut harvest communities were correlated with higher total organic carbon and total nitrogen as well as lower pH. By the early self-thinning stage, soil arthropod communities in both wildfire and clearcut harvest stands appeared to be similar, with both being correlated with lower pH, lower TN, and higher total organic carbon.

Analysis of beta diversity reveals clear patterns of convergence and shows a trajectory of recovery. Binary Bray Curtis dissimilarities (Sorensen distances) were compared between full-tree clearcut harvest sites and the wildfire reference condition. (a) Non-metric multi-dimensional scaling plots (NMDS) were used to assess community convergence at each stand development stage. For each taxon, the scale of the axes are consistent across each stand development stage. Arrows indicate significantly correlated environmental variables with correlations > 0.30 and p-values < 0.05. Holm-corrected p-values < 0.05 from pairwise permutational analysis of variance (PERMANOVA) tests are marked with an asterisk. (b) NMDS plots were used to assess recovery trajectory, significantly correlated environmental variables, and the corresponding Holm-corrected p-values from pairwise PERMANOVA tests are shown below each plot. Abbreviations: E = establishment, CC = crown closure, CCH = clear cut harvest; EST = early self-thinning, LST = late self-thinning, M = mature, TN = total nitrogen; TOC = total organic carbon, W = wildfire.
Recovery patterns are shown in Fig. 3 (b) for each taxon (bacteria stress = 0.08 and linear R2 = 0.98; fungi stress = 0.12 and linear R2 = 0.93; arthropod stress = 0.14 and linear R2 = 0.89). Holm-corrected pairwise PERMANOVA p-values are shown below the recovery plots. Bacterial communities were distinct from the mature, wildfire reference condition at both the crown closure and late self-thinning stages. Fungal and arthropod communities at each stand development stage appeared to be distinct from the mature, wildfire reference condition but the trajectory appeared to approach that of the mature, wildfire reference condition. All taxa appeared to correlate with lower pH over time, fungi and arthropods were correlated with higher total organic carbon over time, and bacteria and arthropods were correlated with lower total nitrogen over time. It is worth highlighting that because the variances in beta diversity were not homogenous, the significant differences found in the PERMANOVA could be due to this dispersion effect, especially for the fungal and arthropod communities as the clustering in the NMDS plots do suggest a recovery trajectory over time. For all taxa, variance in beta diversity was largely explained by stand development stage (28%, 22%, and 20%, respectively), followed by the interaction between stand development stage and disturbance type (full tree clearcut harvest and wildfire) (16%, 12%, and 11%) followed by disturbance type alone (4%, 5%, 4%) (p-value = 0.001).
Comparison of full tree clearcut and salvage harvest versus natural wildfire disturbance
Soil diversity across two stand development stages, establishment and crown closure, were compared between wildfire, full tree clearcut harvest, and post-wildfire salvage harvest. In this case, the wildfire-origin sites were used as the reference condition at each stand development stage to assess the impact of harvest type. All taxa converged on a similar SV richness by the crown closure stage (Fig. 4 (a)). Bacterial richness was not significantly different between harvest types (p-value > 0.05), but both were significantly elevated (p-value < 0.05) compared with the reference condition at the establishment stage. By the crown closure stage, bacterial richness was no longer significantly different from the reference condition (p-value > 0.05). Fungal richness was significantly elevated in clearcut and even more elevated in salvage harvest sites compared with the reference condition (p-value < 0.05) at the establishment stage. By the crown closure stage, fungal richness was no longer significantly different from the reference condition (p-value > 0.05). Arthropoda richness was not significantly different (p-value > 0.05) between harvest types but were significantly elevated (p-value < 0.05) compared with the reference condition at the establishment stage. By crown closure, arthropod richness was no longer significantly different from the reference condition (p-value > 0.05).

Salvage harvest appears to have a greater impact on soil community convergence than full tree clearcut harvest. The reference condition for clearcut and salvage harvest disturbances are wildfire samples in the establishment and crown closure stages. Convergence patterns are shown for (a) SV richness, (b) indirect gradient analysis using NMDS with fitted environmental variables, and (c) beta diversity comparisons among disturbance types using pair-wise PERMANOVA. Abbreviations: * = p < 0.05, ** = p < 0.01, *** = p < 0.001, A = arthropoda, B = bacteria, F = fungi, NMDS = non-metric multi-dimensional scaling, TN = total nitrogen, TOC = total organic carbon.
Using SV richness, all sites appeared to converge by the crown closure stage. When examining beta diversity patterns, it was more difficult to detect convergence with the reference condition (bacteria stress = 0.07, R2 linear = 0.97; fungi stress = 0.11, R2 linear = 0.93; arthropoda stress = 0.13, R2 linear = 0.90) (Fig. 4 (b)). At the establishment stage, higher pH in wildfire sites, and higher total organic carbon in clearcut harvest and salvage sites resulted in distinct bacterial, fungal, and arthropod groups. At both the establishment and crown closure stages, lower total nitrogen in salvage sites resulted in distinct bacterial, fungal, and arthropod groups. For all taxa, by the crown closure stage, wildfire and full tree clearcut harvest sites visually clustered more closely together compared to the salvage sites. Even after ~ 20 years following disturbance, salvage sites clustered together for all taxa and appeared to form their own distinct group. For bacteria, these results were supported by PERMANOVA pair-wise tests shown in Fig. 4 (c). For fungi and arthropods, PERMANOVA pair-wise tests showed that even at the crown closure stage, wildfire and fulltree clearcut harvest sites were significantly different but we feel this is largely due to significant dispersion in the variances across disturbance types for fungi (p-value = 0.004) and arthropods (p-value < 0.001). For all taxa, significant interactions were found between stand development stage and disturbance type with respect to variance in beta diversity (PERMANOVA, p = 0.001).
Discussion
We assessed whether the soil biotic richness in stands disturbed by full-tree clearcut harvesting converged with wildfire disturbed stands and showed signs of recovery (i.e., tracking towards the mature, wildfire reference condition) on similar trajectories as naturally disturbed stands following wildfire. Assessment of convergence involved the comparison of post-wildfire and harvest origin stands at comparable stand development stages. We found that richness was numerically higher for bacteria and significantly higher for fungi in full tree clearcut harvest sites compared with wildfire sites during the initial stand establishment stage (0–10 years following disturbance). This result corresponds with a previous study that assessed microbial diversity in post-wildfire and post-harvest jack-pine stands and found lower microbial diversity following wildfire64. This is also consistent with a meta-analysis of 139 published studies that assessed soil bacterial and fungal biomass following fire and harvest, and found that biomass was also lower following fire compared with harvest65. These initially higher richness values may be due to a legacy effect of retaining a larger portion of the pre-harvest bacterial and fungal communities suggesting some level of resilience or the consequence of high severity wildfires that cause heat-induced mortality, consumes a high proportion of the organic horizon that harbours the majority of microbial diversity, and may reduce soil carbon quantity and quality66. For ectomycorrhizal fungi, higher organic horizon consumption and higher tree mortality has been shown to lead to lower legacy ectomycorrhizal fungi and higher potential of colonization by spores67.
In contrast with richness, bacteria showed beta diversity convergence with the post-fire community composition by the crown closure stage that was sustained across subsequent successional stages. The relatively early/rapid convergence of the bacterial communities suggests that bacteria are not overly sensitive to the demonstrated differences between clearcut harvesting and wildfire, again, consistent with previous studies that looked at the recovery of microbial biomass following fire in boreal habitats65. This lack of sensitivity contrasts strongly with the other taxa. In a previous study following wildfire, we showed great variability in fungal community composition that shifted across stand development stages following wildfire, less so with arthropoda31. In study study following clearcut harvest, arthropod communities did not converge until the late self-thinning stage and the fungal communities did not show convergence at any stage. These results indicate that harvesting disturbances to soil fungal and arthropod communities are not directly analogous to those experienced in typical fire successional dynamics and that clearcut harvesting of mature forests does create different habitat conditions across most of the stand development stages. These differences in responses between taxa are likely driven, in part, by the provision of different habitat conditions created by these two disturbance types that could represent distinct successional processes between these two disturbances. This is consistent a study of beetles in the Canadian boreal forest that observed that post fire and post harvest communities had begun to converge by 30 years68.
As soil biotic communities have been demonstrated to be highly responsive to changes in physicochemical conditions (e.g., moisture, temperature)69 these soil biotic community differences are likely reflecting real differences in site conditions. For example, wildfire consumes a substantial portion of the pre-disturbance forest floor and downed woody debris (DWD) complex, volatilizes N, raises pH via ash deposition, and the blackened soil surface absorbs more incoming solar radiation70,71. In contrast, forest floors and existing DWD following harvesting largely remain intact, although some mechanical disruption does occur along wood extraction trails72,73,74,75. As highlighted in Fig. 3, in wildfire stands at the establishment stage, all taxa were correlated with higher pH. Additionally, fungi and arthropods in these stands were associated with lower TOC and lower TN compared to those in clearcut harvest sites.
We also assessed the recovery of soil biotic communities back to their pre-harvest, mature, fire-origin condition (i.e., natural succession endpoint). This assessment involved the comparison of harvested stands at each stand development stage against the mature, wildfire reference condition. The rapid recovery of bacterial communities in full tree clearcut harvested sites further supports the likelihood of a legacy effect where more organisms from the ‘mature’ (pre-disturbance) condition were retained through harvesting when compared against their wildfire disturbance counterparts. This legacy effect is also likely a function of retaining the existing forest floor and DWD material. This result is consistent with a previous study of boreal forest soil bacteria that showed similar co-occurrence networks at 1 year and 5 years post fire as well as communities that became more similar to unburned controls at 5 years post fire76.
The observed greater richness following harvesting at the establishment stage may have implications for organic matter decomposition and soil carbon storage if these taxa contained more saprotrophs from the mature condition which might increase organic matter decomposition and reduce soil organic carbon stores. In Canadian old boreal forests, it was shown that soil carbon pools began to decrease, or at least showed no notable increases, in mature forests77. They also showed that while early successional stages accumulate the most tree carbon, many of the organisms that are plant pathogens or saprotrophs present in mature soils could potentially limit carbon accumulation if they are more abundant in early successional clearcuts. Canadian boreal forests in northern Ontario, despite measured declines in soil C pools following clearcut harvesting in jack pine and black spruce-dominated stands, it was found that these pools had returned to pre-disturbance levels within 20 years after disturbance78.
In this study, soil biotic community composition across all three taxa in salvage-logged sites appeared distinct from either the wildfire or clearcut harvest sites and remain so through the crown closure stage of stand development (out to 20 years following disturbance). As highlighted in Fig. 4 (b), these differences are largely correlated with low soil N concentrations on salvage logged sites. These differences may have implications for soil function including nutrient cycling in these coarse-textured, nutrient poor soils. A previous study that assessed the impacts of wildfire and salvage harvest suggested that soil nutrients in jack pine and black spruce stands would not likely return to their pre-burn level even in a 110-year rotation79. In a review comparing wildfire and harvest systems, differences in nutrient pools between wildfire and other stand-replacing disturbances such as clearcut harvest, or salvage harvest following wildfire were also highlighted3. Previous work has indicated that harvesting of trees from sites before they have recovered from a natural disturbance, may represent a second disturbance resulting in compounded effects that, in turn, result in longer-term alteration of the affected biotic communities8,75,80 which is consistent with what we observed here. Post-fire salvage logging has been shown to reduce the habitat preferred by pyrophilous species such as certain arthropods and fungi7,81,82. However, responses of fungi may be less severe than reductions in arthropods, as ectomycorrhizal fungi can persist on the roots of surviving trees or as resistant structures in the soil, after a severe fire or salvage logging where most of the trees are killed or removed11. In the context of forest operations, the ecological consequences of a reduction of fungi are not always perceived as negative though, as some fungi cause plant disease83. From an ecological perspective, however, fungi are also important recyclers of organic material, plant mutualists, and are natural components of healthy forests84,85. Where this legacy fungal inoculum is reduced, by salvage logging for example, development of soil communities may be slow, or altered, as recolonization proceeds primarily from spores dispersed by air, animal vectors, or from resistant propagules in soil67,86,87. Slow spore dispersal rates might explain the lack of fungal convergence between wildfire and salvage sites out to 20 years post-disturbance. Similarly, the removal of fire-killed trees during salvage harvest, reduces the downed woody debris pool necessary for some elements of the arthropod community such as beetles and flat bugs, contributing to the observed community differences81,88,89. The removal of these tree residues can also impact soil conditions by changing site microtopographic characteristics like temperature and moisture that can impact soil microbial communities90. It has been proposed that the retention of standing trees and deadwood can moderate observed changes in the fungal community by preserving habitat for ectomycorrhizal and saprotrophic fungi91,92. There is also evidence that compounding disturbances in forest management practices can contribute to impacts to the arthropod community88,93. Our results suggest that salvage logging can extend the time for convergence of the forest soil biotic community towards the wildfire disturbance condition and creates unique communities that are not part of the natural successional sequence. Although our timelines were limited here, salvage logging appears to alter the successional pathways of the soil communities which undermines the assumptions of the natural disturbance emulation approach.
In many jurisdictions, including Ontario, Canada, forest sustainability includes reference to using forest practices that emulate natural disturbances (END) to conserve biodiversity and their associated ecological processes (i.e., Ontario’s Crown Forest Sustainability Act of 1994). In the boreal forest, END has generally focused on landscape pattern emulation (e.g., the size and distribution of harvest relative to wildfire). However, at the stand/site scale, wildfire and harvest are not the same disturbance mechanism. One is a pyrochemical process that kills, but retains, the overstory tree stems, but consumes a substantial portion of the existing forest floor and downed woody debris. In contrast, clearcut harvesting is a mechanical process that removes the merchantable overstory trees but leaves the forest floor and DWD complex largely intact. Even though both are stand-replacing disturbances that reset the disturbed areas to the establishment stage of stand development, the conditions immediately after disturbance are different. Within the context of END, the underlying assumption is that there will be convergence over a reasonable period of time and will eventually resemble (recover) the pre-disturbance condition, that, in turn, would define the “ecological” rotation92,94.
The lack of convergence of arthropod and fungal communities seen in this study does raise a concern with the END paradigm. Our results suggest that soil communities in harvested sites can remain significantly different through successional stages up to the late self-thinning stage (46–70 years following disturbance). Given the spatial extent of clearcut harvesting, these results suggest potential large-scale changes to the distribution of distinct soil communities across Ontario’s boreal forest. The use of prescribed fire, as a site preparation tool, may be one approach that could be used to ameliorate these effects, but in general they are difficult to implement and not commonly done. Another option might be to introduce some of the aspects of fire disturbance (e.g., selective additions of high pH wood ash from local bioenergy production facilities) to reduce the gap between harvest and fire origin soil communities95.
The lack of recovery in both the fungal and arthropod communities out to the late self-thinning stage would also suggest that timber extraction rotations, for at least a portion of our managed stands, need to be longer than 70 years, and not based primarily on the culmination of MAI (mean annual increment), to allow for the full recovery of the soil biotic community. This is consistent with previous forest management recommendations and would ensure representation of old forest conditions through both retention and succession92. Unfortunately, we did not have any harvest origin stands beyond 66 years to confirm when full recovery is achieved. We do know from our previous study of post-wildfire soil biota succession that soil communities in fire origin stands remained unique out to at least age 8031.
Results for both convergence and recovery highlight the value of using beta diversity rather than just species richness to assess successional trajectories as it provides more information relevant to decision making for forest management regarding community composition. In addition, the beta diversity results demonstrate clear distinctions between the response of individual taxa highlighting the value of a multi-taxonomic approach to sustainability assessments31,96. Beyond beta diversity, however, we also need to understand how these shifts in community composition affect soil function.
Adaptive management is an approach that recognizes that policy should be treated as a testable hypothesis and its effectiveness should be measured to support policy refinement. This will be increasingly important for adapting forest management to climate change97. Biodiversity is an essential indicator of forest management policy to achieve sustainability. Our work highlights the capacity for DNA metabarcoding to inform forestry-related questions about below-ground biodiversity that are generally challenging to study using more conventional methods. The metabarcoding approach also supports the inclusion of a wide diversity of taxa in the assessment. Our results clearly demonstrate the need for multi-taxonomic assessments due to the taxonomic variation in response to disturbances. The results also highlight that a wider array of taxa provides more insight at functional levels that should be examined in the future. Because a major limitation when working with such a broad array of organisms is lack of representation in reference sequence databases at a fine taxonomic level, this work focused on the analysis of sequence variants. Future studies using similar techniques could be used to further assess the potential ecological implications of the differences observed in this study. One potential focus could be investigating the aboveground-belowground linkages facilitated by organism exchange and, potential functional activities identified with enzyme assays or functional databases. This is an important avenue for future research and could involve the use of functional databases such as FUNGuild, Faprotax, BETSI, GLoBi that can map trophic interactions, enzyme assays that measure changes in potential nutrient use, or shotgun sequencing that can provide information on the functional genes that are present98,99,100,101,102,103. Additional analysis of these data can be used to assess the functional dynamics of recovery and understand which components of the community are responsive or resilient to the changes represented by the different forest age classes, and even which organisms can colonize from external sources, versus those that require in-situ regeneration.
Tests of the effectiveness of forest management strategies and practices aimed at emulating natural disturbance are rare and difficult. This study offers a unique comparison of biodiversity response to harvest relative to fire. The chronosequence approach provides unique information about the contrast between harvest and fire over time frames that are not achievable in long-term studies. Efforts to establish and maintain chronosequence studies should be a focus of effectiveness research, especially as the information necessary to establish chronosequence studies is being lost over time. Also, in the Ontario boreal forest, we are just now seeing post industrially-harvested stands reaching 70 years and so there is significant potential to extend these chronosequences over longer timeframes to provide additional insights with respect to full recovery and ecological rotations.
Conclusions
This chronosequence study combined with DNA metabarcoding of soil biodiversity has provided some unique insights into the effectiveness of natural disturbance emulation as a forest management paradigm. Soil fungi and arthropods were the most responsive taxa indicating that there are significant differences in soil community composition through all examined stand development stages, and that some taxa do not recover to the mature, fire-origin reference condition over a 60-year timeframe. A comprehensive look at potential functional differences is now needed to better understand the ecological implications of these community differences. Our results do, however, suggest that innovative management approaches should be considered to support soil communities that, in turn, support resilient forests. For example, we recommend consideration of use of prescribed burns as a site preparation tool, where feasible, or the selective addition of wood ash from bioenergy production to improve the emulation of wildfire. We also recommend allowing for longer rotations and balancing age class distributions for the provision of habitat for the full range of soil biota. Lastly, we recommend only limited use of salvage logging after fire because it shifts soil communities away from a natural fire-origin composition. Where conducted, salvage-logged areas should have extended rotations to account for the lag in convergence and recovery.
Data availability
Sequences have been deposited to the NCBI SRA under the GRDI-Ecobiomics project accession PRJNA565010 (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA565010) for the BioSample accessions SAMN26926703 - SAMN26926795 used in this study. The MetaWorks v1.9.3 multi-marker metabarcode bioinformatic pipeline is available from https://github.com/terrimporter/MetaWorks. The ITS classifier was based on the ITS UNITE+INSD full dataset v8.2 and trained to work with the RDP Classifier (https://github.com/terrimporter/UNITE_ITSClassifier). The COI classifier v4 is available from https://github.com/terrimporter/CO1Classifier. The code used to produce figures, including infile and metadata files, are available on https://github.com/terrimporter/Chronosequence_HarvestType.
References
Rudolph, T. D. & Laidly, P. R. Pinus Banksiana lamb. Jack pine. Silvics North. Am. 1, 280–293 (1990).
O’Hara, K. L. & Ramage, B. S. Silviculture in an uncertain world: utilizing multi-aged management systems to integrate disturbance. Forestry 86, 401–410 (2013).
McRae, D. J., Duchesne, L. C., Freedman, B., Lynham, T. J. & Woodley, S. Comparisons between wildfire and forest harvesting and their implications in forest management. Environ. Rev. 9, 223–260 (2001).
Lindenmayer, D. B. et al. Salvage harvesting policies after natural disturbance. Science 303, 1303–1303 (2004).
Sessions, J., Bettinger, P., Buckman, R., Newton, M. & Hamann, J. Hastening the return of complex forests following fire. J. Forest. 102, 38–45 (2004).
Donato, D. C. et al. Post-wildfire logging hinders regeneration and increases fire risk. Science 311, 352–352 (2006).
Lindenmayer, D. B. & Noss, R. F. Salvage logging, ecosystem processes, and biodiversity conservation: overview of salvage logging. Conserv. Biol. 20, 949–958 (2006).
D’Amato, A. W., Fraver, S., Palik, B. J., Bradford, J. B. & Patty, L. Singular and interactive effects of blowdown, salvage logging, and wildfire in sub-boreal pine systems. For. Ecol. Manag. 262, 2070–2078 (2011).
Lindenmayer, D. B., Burton, P. J. & Franklin, J. F. Salvage Logging and its Ecological Consequences (Island, 2012).
Thorn, S. et al. Impacts of salvage logging on biodiversity: A meta-analysis. J. Appl. Ecol. 55, 279–289 (2018).
Visser, S. Ectomycorrhizal fungal succession in Jack pine stands following wildfire. New Phytol. 129, 389–401 (1995).
LeDuc, S. D. & Rothstein, D. E. Initial recovery of soil carbon and nitrogen pools and dynamics following disturbance in Jack pine forests: A comparison of wildfire and clearcut harvesting. Soil Biol. Biochem. 39, 2865–2876 (2007).
Yermakov, Z. & Rothstein, D. E. Changes in soil carbon and nitrogen cycling along a 72-year wildfire chronosequence in Michigan Jack pine forests. Oecologia 149, 690–700 (2006).
LeDuc, S. D., Lilleskov, E. A., Horton, T. R. & Rothstein, D. E. Ectomycorrhizal fungal succession coincides with shifts in organic nitrogen availability and canopy closure in post-wildfire Jack pine forests. Oecologia 172, 257–269 (2013).
Schieck, J. & Song, S. J. Changes in bird communities throughout succession following fire and harvest in boreal forests of Western North america: literature review and meta-analyses. Can. J. For. Res. 36, 1299–1318 (2006).
Zwolak, R. A meta-analysis of the effects of wildfire, clearcutting, and partial harvest on the abundance of North American small mammals. For. Ecol. Manag. 258, 539–545 (2009).
Clark, J. A. & Covey, K. R. Tree species richness and the logging of natural forests: A meta-analysis. For. Ecol. Manag. 276, 146–153 (2012).
Gerstner, K., Dormann, C. F., Stein, A., Manceur, A. M. & Seppelt, R. Effects of land use on plant diversity – A global metaanalysis. J. Appl. Ecol. 51, 1690–1700 (2014). Editor’s chiice: Review.
Michał, B. & Rafał, Z. Responses of small mammals to clear-cutting in temperate and boreal forests of europe: a meta-analysis and review. Eur. J. For. Res. 133, 1–11 (2014).
Vasconcelos, H. L., Maravalhas, J. B. & Cornelissen, T. Effects of fire disturbance on ant abundance and diversity: a global meta-analysis. Biodivers. Conserv. 26, 177–188 (2017).
Lee, D. E. Spotted owls and forest fire: a systematic review and metaanalysis of the evidence. Ecosphere 9, e02354 (2018).
Carbone, L. M., Tavella, J., Pausas, J. G. & Aguilar, R. A global synthesis of fire effects on pollinators. Glob. Ecol. Biogeogr. 28, 1487–1498 (2019).
Dove, N. C. & Hart, S. C. Fire reduces fungal species richness and in situ mycorrhizal colonization: a meta-analysis. Fire Ecol. 13, 37–65 (2017).
Certini, G., Moya, D., Lucas-Borja, M. E. & Mastrolonardo, G. The impact of fire on soil-dwelling biota: A review. For. Ecol. Manag. 488, 118989 (2021).
Kudrin, A., Perminova, E., Taskaeva, A., Ditts, A. & Konakova, T. A Meta-analysis of the effects of harvesting on the abundance and richness of soil fauna in boreal and temperate forests. Forests 14, 923 (2023).
Liu, W. et al. Effects of fire on the soil microbial metabolic quotient: A global meta-analysis. CATENA 224, 106957 (2023).
Claridge, A. W., Trappe, J. M. & Hansen, K. Do fungi have a role as soil stabilizers and remediators after forest fire? For. Ecol. Manag. 257, 1063–1069 (2009).
Neary, D. G., Klopatek, C. C., DeBano, L. F. & Ffolliott, P. F. Fire effects on belowground sustainability: a review and synthesis. For. Ecol. Manag. 122, 51–71 (1999).
Menta, C. & Remelli, S. Soil health and arthropods: from complex system to worthwhile investigation. Insects 11, 54 (2020).
Filialuna, O. & Cripps, C. Evidence that pyrophilous fungi aggregate soil after forest fire. For. Ecol. Manag. 498, 119579 (2021).
Porter, T. M., Smenderovac, E., Morris, D. & Venier, L. All boreal forest successional stages needed to maintain the full suite of soil biodiversity, community composition, and function following wildfire. Sci. Rep. 13, 7978 (2023).
Crins, W. J. Ecosystems of Ontario. Part 1, Ecozones and ecoregionsOntario, ministry of natural resources, inventory, monitoring and assessement section, Toronto, Ont.,. (2009).
Soil classification working group. the canadian system of soil classification. National Research Council of Canada, Ottawa, (1998).
Plonski, W. L. Normal Yield Tables (metric) for Major Forest Species of Ontario (Ontario Ministry of Natural resources, 1981).
Dyck, W. J. & Cole, D. W. Strategies for determining consequences of harvesting and associated practices on long-term productivity. In Impacts of Forest Harvesting on long-term Site Productivity (eds. Dyck, W.J., Cole, D.W., & Comerford, N.B.) 13–40 (Springer, 1994).
Braid, M. D., Daniels, L. M. & Kitts, C. L. Removal of PCR inhibitors from soil DNA by chemical flocculation. J. Microbiol. Methods. 52, 389–393 (2003).
Parada, A. E., Needham, D. M. & Fuhrman, J. A. Every base matters: assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 18, 1403–1414 (2016).
Menkis, A. et al. Occurrence and impact of the root-rot biocontrol agent phlebiopsis gigantea on soil fungal communities in Picea abies forests of Northern Europe. FEMS Microbiol. Ecol. 81, 438–445 (2012).
White, T. J., Bruns, T., Lee, S. & Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. in PCR Protocols: A Guide To Methods and Applications (eds. Innis, M.A., Gelfand, D.H., Sninsky, J.J., & White, T.J.) 315–322 (Academic Press, Inc., 1990).
Hajibabaei, M., Porter, T. M., Wright, M. & Rudar, J. COI metabarcoding primer choice affects richness and recovery of indicator taxa in freshwater systems. PLoS ONE. 14, e0220953 (2019).
Folmer, O., Black, M., Hoeh, W., Lutz, R. & Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3, 294–299 (1994).
Gibson, J. et al. Large-scale biomonitoring of remote and threatened ecosystems via high-throughput sequencing. PLos One. 10, e0138432 (2015).
Hajibabaei, M., Spall, J. L., Shokralla, S. & van Konynenburg, S. Assessing biodiversity of a freshwater benthic macroinvertebrate community through non-destructive environmental barcoding of DNA from preservative ethanol. BMC Ecol. 12, 28 (2012).
Illumina. 16S metagenomic sequencing library preparation - Preparing 16S ribosomal RNA gene amplicons for the Illumina MiSeq System. (2013). https://support.illumina.com/downloads/16s_metagenomic_sequencing_library_preparation.html
Porter, T. M., Hajibabaei, M. & MetaWorks A flexible, scalable bioinformatic pipeline for high-throughput multi-marker biodiversity assessments. PLoS ONE. 17, e0274260 (2022).
St. John, J. SeqPrep. (2016). https://github.com/jstjohn/SeqPrep/releases
Martin, M. & EMBnet. Cutadapt removes adapter sequences from high-throughput sequencing reads. Journal 17, pp–10 (2011).
Rognes, T., Flouri, T., Nichols, B., Quince, C. & Mahé, F. VSEARCH: a versatile open source tool for metagenomics. PeerJ 4, e2584 (2016).
Wang, Q., Garrity, G. M., Tiedje, J. M. & Cole, J. R. Naive bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267 (2007).
Abarenkov, K. et al. UNITE QIIME release for Fungi. Version 04.02.2020. UNITE Community. (2020).
Porter, T. M. & Hajibabaei, M. Automated high throughput animal CO1 metabarcode classification. Sci. Rep. 8, 4226 (2018).
RStudio Team. RStudio: integrated development environment for R. (2016). http://www.rstudio.com/
R Core Team. R: A language and environment for statistical computing. (2018). https://www.R-project.org/
Oksanen, J. et al. vegan: Community Ecology Package. R package version 2.5-2. (2018). https://CRAN.R-project.org/package=vegan
Elbrecht, V., Vamos, E. E., Meissner, K., Aroviita, J. & Leese, F. Assessing strengths and weaknesses of DNA metabarcoding-based macroinvertebrate identification for routine stream monitoring. Methods Ecol. Evol. 8, 1265–1275 (2017).
Callahan, B. J., McMurdie, P. J. & Holmes, S. P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 11, 2639–2643 (2017).
Amend, A. S., Seifert, K. A. & Bruns, T. D. Quantifying microbial communities with 454 pyrosequencing: does read abundance count? Quantitative pyrosequencing of microbes. Mol. Ecol. 19, 5555–5565 (2010).
Lamb, P. D. et al. How quantitative is metabarcoding: A meta-analytical approach. Mol. Ecol. 28, 420–430 (2019).
Kassambara, A. & ggpubr ‘ggplot2’ based publication ready plots. (2020).
Goral, F., Schellenberg, J. & goeveg Functions for Community Data and Ordinations. (2018). https://CRAN.R-project.org/package=goeveg
Martinez Arbizu, P. & pairwiseAdonis Pairwise multilevel comparison using adonis. R package version 0.0.1. (2017). https://github.com/pmartinezarbizu/pairwiseAdonis
Wickham, H. Ggplot2: Elegant Graphics for Data Analysis (Springer-, 2009).
Kahle, D. & Wickham, H. Ggmap: Spatial visualization with ggplot2. R J. 5, 144–161 (2013).
Staddon, W. J., Duchesne, L. C. & Trevors, J. T. Impact of clear-cutting and prescribed burning on microbial diversity and community structure in a Jack pine (Pinus Banksiana Lamb.) clear-cut using biolog Gram-negative microplates. World J. Microbiol. Biotechnol. 14, 5 (1998).
Holden, S. R. & Treseder, K. K. A meta-analysis of soil microbial biomass responses to forest disturbances. Front. Microbiol. 4, 163 (2013).
Dooley, S. R. & Treseder, K. K. The effect of fire on microbial biomass: a meta-analysis of field studies. Biogeochemistry 109, 49–61 (2012).
Dahlberg, A. Effects of fire on ectomycorrhizal fungi in Fennoscandian boreal forests. Silva Fenn. 36, 69–80 (2002).
Hammond, H. E. J., Langor, D. W. & Spence, J. R. Changes in saproxylic beetle (Insecta: Coleoptera) assemblages following wildfire and harvest in boreal Populus forests. For. Ecol. Manag. 401, 319–329 (2017).
Wilhelm, R. C. et al. Biogeography and organic matter removal shape long-term effects of timber harvesting on forest soil microbial communities. ISME J. 11, 2552–2568 (2017).
Johnson, D. B. et al. Effects of fire and fire-induced changes in soil properties on post-burn soil respiration. Fire Ecol. 20, 90 (2024).
Zhao, J. et al. Forest fire size amplifies postfire land surface warming. For. Fire Size Amplifies Postfire Land. Surf. Warming. 633, 828–834 (2024).
Thiffault, E. et al. Chemical composition of forest floor and consequences for nutrient availability after wildfire and harvesting in the boreal forest. Plant. Soil. 308, 37–53 (2008).
Norris, C. E., Quideau, S. A., Bhatti, J. S., Wasylishen, R. E. & MacKenzie, M. D. Influence of fire and harvest on soil organic carbon in Jack pine sites. Can. J. For. Res. 39, 642–654 (2009).
Löf, M., Dey, D. C., Navarro, R. M. & Jacobs, D. F. Mechanical site Preparation for forest restoration. New Forest. 43, 825–848 (2012).
Maynard, D. G. et al. How do natural disturbances and human activities affect soils and tree nutrition and growth in the Canadian boreal forest? Environ. Rev. 22, 161–178 (2014).
Whitman, T., Woolet, J., Sikora, M., Johnson, D. B. & Whitman, E. Resilience in soil bacterial communities of the boreal forest from one to five years after wildfire across a severity gradient. Soil Biol. Biochem. 172, 108755 (2022).
Gao, B. et al. Carbon storage declines in old boreal forests irrespective of succession pathway. Ecosystems 21, 1168–1182 (2018).
Morris, D. M. et al. Effects of biomass removal levels on soil carbon and nutrient reserves in conifer-dominated, coarse‐textured sites in Northern Ontario: 20‐year results. Soil Sci. Soc. Amer. J. 83, S116–S132 (2019).
Brais, S., David, P. & Ouimet, R. Impacts of wild fire severity and salvage harvesting on the nutrient balance of Jack pine and black Spruce boreal stands. For. Ecol. Manag. 1–3, 231–243 (2000).
Paine, R. T., Tegner, M. J. & Johnson, E. A. Compounded Perturbations Yield Ecol. Surprises Ecosystems 1, 535–545 (1998).
Cobb, T. P. et al. Effects of postfire salvage logging on Deadwood-Associated beetles: salvaging logging and Deadwood-Associated beetles. Conserv. Biol. 25, 94–104 (2011).
Pérez-Izquierdo, L. et al. Crown‐fire severity is more important than ground‐fire severity in determining soil fungal community development in the boreal forest. J. Ecol. 109, 504–518 (2021).
Loo, J. A. Ecological impacts of non-indigenous invasive fungi as forest pathogens. in Ecological impacts of non-native invertebrates and fungi on terrestrial ecosystems 81–96Springer, Dordrecht, (2009).
Ostry, M. E. & Laflamme, G. Fungi and diseases — natural components of healthy forests This article is one of a collection of papers based on a presentation from the stem and shoot fungal pathogens and parasitic plants: the values of biological diversity session of the XXII international union of forestry research Organization world congress meeting held in Brisbane, Queensland, Australia, in 2005. Botany 87, 22–25 (2009).
Tomao, A., Antonio Bonet, J., Castaño, C. & de-Miguel, S. How does forest management affect fungal diversity and community composition? Current knowledge and future perspectives for the conservation of forest fungi. For. Ecol. Manag. 457, 117678 (2020).
Baar, J., Horton, T. R., Kretzer, A. M. & Bruns, T. D. Mycorrhizal colonization of Pinus muricata from resistant propagules after a stand-replacing wildfire. New Phytol. 143, 409–418 (1999).
Glassman, S. I., Levine, C. R., DiRocco, A. M., Battles, J. J. & Bruns, T. D. Ectomycorrhizal fungal spore bank recovery after a severe forest fire: some like it hot. ISME J. 10, 1228–1239 (2016).
Cobb, T. P., Langor, D. W. & Spence, J. R. Biodiversity and multiple disturbances: boreal forest ground beetle (Coleoptera: Carabidae) responses to wildfire, harvesting, and herbicide. Can. J. Res. 37, 1310–1323 (2007).
Heikkala, O., Martikainen, P. & Kouki, J. Prescribed burning is an effective and quick method to conserve rare pyrophilous forest-dwelling flat Bugs. Insect Conserv. Divers. 10, 32–41 (2017).
Pereira, P., Francos, M., Brevik, E. C., Ubeda, X. & Bogunovic I. Post-fire soil management. Curr. Opin. Environ. Sci. Health. 5, 26–32 (2018).
Mayer, M. et al. Surviving trees and Deadwood moderate changes in soil fungal communities and associated functioning after natural forest disturbance and salvage logging. Soil Biol. Biochem. 166, 108558 (2022).
Prescott, C. E. & Perspectives Regenerative forestry – Managing forests for soil life. For. Ecol. Manag. 554, 121674 (2024).
Work, T. T. et al. Cumulative effects of biomass harvesting and herbicide application on litter-dwelling arthropod communities in Jack pine-dominated forests: 7th year postharvest assessment. Can. J. Res. 53, 931–952 (2023).
Kimmins, J. P. Sustained yield, timber mining, and the concept of ecological rotation: a British Columbian view. Forestry Chron. 50, 27–31 (1974).
Hannam, K. D., Fleming, R. L., Venier, L. & Hazlett, P. W. Can bioenergy Ash applications emulate the effects of wildfire on upland forest soil chemical properties? Soil Sci. Soc. Am. J. 83, S201–S217 (2019).
Venier, L. A. et al. Effects of natural resource development on the terrestrial biodiversity of Canadian boreal forests. Environ. Rev. 22, 457–490 (2014).
Keenan, R. J. Climate change impacts and adaptation in forest management: a review. Ann. For. Sci. 72, 145–167 (2015).
Tringe, S. G. et al. Comparative metagenomics of microbial communities. Science 308, 554–557 (2005).
Jackson, C. R., Tyler, H. L. & Millar, J. J. Determination of microbial extracellular enzyme activity in waters, soils, and sediments using high throughput microplate assays. JoVE 50399 https://doi.org/10.3791/50399 (2013).
Poelen, J. H., Simons, J. D. & Mungall, C. J. Global biotic interactions: an open infrastructure to share and analyze species-interaction datasets. Ecol. Inf. 24, 148–159 (2014).
Louca, S., Parfrey, L. W. & Doebeli, M. Decoupling function and taxonomy in the global ocean Microbiome. Science 353, 1272–1277 (2016).
Nguyen, N. H. et al. FUNGuild: an open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 20, 241–248 (2016).
Joimel, S. et al. FAO, Rome, Italy,. A large database on functional traits for soil ecologists: BETSI. in Global Symposium on Soil Biodiversity 523–528 (2021).
Acknowledgements
TMP, ES, EJSE, and LV were funded by the Government of Canada through the Genomics Research and Development Initiative (GRDI) Metagenomics-Based Ecosystem Biomonitoring (Ecobiomics) project. We are grateful for the soil chemical analysis performed by the Ontario Forest Research Inorganic Laboratory Services (ORFRILS) in Sault Ste. Marie; Derek Chartrand, Kerrie Wainio-Keizer and Susan Bowman at the Great Lakes Forestry Centre for soil sample processing and conducting DNA extractions; Marie-Josée Morency and the Seguin lab for amplicon and sequencing library preparation; as well as to the Natural Research Council Canada sequencing facility in Saskatoon for Illumina MiSeq sequencing services.
Author information
Authors and Affiliations
Contributions
DM and LV conceived of the project and conducted site selection. DM, LV and EE obtained funding for the project. LV and EE provided the laboratory services for sample processing. TMP did the bioinformatics, data analysis, and wrote the manuscript. ES, DM, and LV assisted with data analysis. All authors contributed to editing the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Porter, T.M., Morris, D.M., Smenderovac, E. et al. Soil communities following harvest have different early successional dynamics compared with post-wildfire patterns. Sci Rep 15, 43413 (2025). https://doi.org/10.1038/s41598-025-16373-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-16373-y
