
Abstract
European badgers (Meles meles) are reservoirs for animal tuberculosis (TB) in some European countries, complicating TB control in cattle. Badger vaccination and a deeper understanding of the subsequent protection mechanisms are necessary for effective TB control. In a previous study, two of eight badgers immunized with the heat-inactivated Mycobacterium bovis (HIMB) vaccine exhibited an unusual immune response (divergent), developing exacerbated lesions. The present study aimed to describe the local immune response in divergent badgers (those with severe disease progression), with respect to that observed in standard (where the vaccine showed efficacy) and control badgers. Immunohistochemistry was performed to evaluate immune cells (macrophages, T and B lymphocytes, plasma cells), and proteins (TGF-β, IL-10, Fox-P3) within TB granulomas in the lung and bronchial lymph node (LN), after TB challenge. Lung lesion volume, bacterial load and immunological response were also evaluated. The divergent immune response was characterized by elevated IL-10 and Fox-P3, few macrophages and high B lymphocytes (mainly in lungs), suggesting a Th1/Th2 imbalance with reduced Th1 cellular immunity leading to severe TB. In contrast, vaccinated badgers with a standard immune response showed a balanced response, with significantly lower bacterial loads (85.5% LN and 99.9% lung) than control group. This study provides new insights into the immune mechanisms in HIMB-vaccinated badgers, to improve TB control strategies.
Similar content being viewed by others

Introduction
Animal tuberculosis (TB) is a chronic infectious disease caused by Mycobacterium bovis and other related Mycobacterium tuberculosis complex (MTC) bacteria with major social, economic, and public health impacts1,2. It is a multi-host disease that can affect many wildlife and livestock species, even humans3,4,5.
The European badger (Meles meles) has been recognized as a significant reservoir of TB in certain regions of Europe including the United Kingdom, Ireland, and northern Spain, playing a crucial role in its epidemiology by participating in the persistence and spread of the disease6,7,8. The transmission of TB between badgers and cattle has raised concerns due to its impact on animal health, agricultural economics, and disease control policies, frustrating the extensive control and eradication efforts in cattle and incurring in significant costs7. The vaccination of cattle against TB is forbidden under EU legislation (Council Directive 78/52/EEC), as vaccination is considered not compatible with the provisions for testing and herd qualification (Council Directive 64/432/EEC). However, wildlife vaccination is a promising tool for controlling the spread of TB in these populations and minimizing its potential as a reservoir for livestock populations. The immunization of wildlife has been demonstrated to reduce the incidence of TB in species such as wild boar (Sus scrofa), badger, white-tailed deer (Odocoileus virginianus), and brushtail possum (Trichosurus vulpecula), among others, providing a sustainable long-term control strategy9,10,11,12,13,14,15. Specifically, the use of a heat-inactivated Mycobacterium bovis (HIMB) vaccine administered via bait has been used, as it is expected to be more stable and safer than Bacillus Calmette-Guérin (BCG) due to being based on dead bacteria16,17, thus overcoming the main BCG limitations. Recent studies have demonstrated the effects of HIMB vaccination in several wild species, i.e., captive wild boar17,18,19, Molokai-origin wild pig20, red deer (Cervus elaphus)21, European badger12, and African buffalo (Syncerus caffer)22. This approach aims to be included in integrated and innovative control schemes which combine existing control measures, such as culling and biosecurity, offering a sustainable alternative.
Understanding the immune responses developed by vaccinated badgers is crucial for both assessing the efficacy of vaccines against TB and elucidating mechanisms of protection. In addition, the characterization of granulomas using immunohistochemical approaches, by identifying cell composition and cytokine expression, allows to detect differences in the composition of TB granulomas among individuals and their subsequent singularities in the host immune response against the pathogen23,24. Immunization with HIMB, besides inducing specific cellular and humoral adaptative responses, can also activate innate immune mechanisms of the trained immunity (TRAIM) type, thus providing protection not only against mycobacteria but also against other pathogens which share the same pathogen-associated molecular patterns (PAMPs)21,25,26,27,28. TRAIM has been defined as the functional feature of the innate immune system that enhances secondary responses to the same or different pathogens through non-specific memory29. When the innate immune response, in which macrophages play an important role, is unable to efficiently eliminate the invading pathogen due to its burden, route or virulence, the adaptative response is activated, along with lymphocytes (T and B lymphocytes, and differentiated matured B cells into plasma cells under antigenic activation), allowing for the development of a more specific response following pathogen recognition. Since the latter takes up to two weeks to establish, the innate response partially mediates the protection against the pathogen, developing a certain immunological memory. This has been demonstrated in a recent study in which 15 badgers were vaccinated with HIMB using edible baits30. In the mentioned study, vaccination acted according to the expected TRAIM model, resulting in partial protection against disease progression among the vaccinated group. However, two vaccinated badgers exhibited an exacerbated or divergent response (severe disease progression). Based on those results, in the current study we attempt to take advantage of this unexpected finding to deepen in the description of relevant variables in badgers showing a divergent immune response compared to standard responding vaccinated (in which vaccine showed efficacy) and control badgers.
Materials and methods
Experimental design
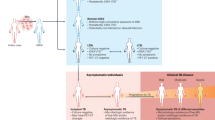
This study is a continuation of Juste et al30. where all procedures are reported in detail. Briefly, fifteen young adult badgers, seven of which were male and eight females, were trapped between March and September 2018 in Asturias (north-western Spain). Afterwards, eight badgers were individually given five baits containing a HIMB dose of heat-inactivated 5 × 107 colony-forming units (CFU) M. bovis. In parallel, a control group of seven badgers did not receive the vaccine. One month and a half later, a second dose of the vaccine was offered to the vaccinated group. Ninety-four days after the second dose, all badgers were challenged with M. bovis (103 CFU/animal) delivered endobronchially to the right middle lung lobe. Clinical, immunological, pathological and bacteriological variables were measured throughout the whole study to assess the efficacy of the vaccine30. Two vaccinated animals showed higher bacterial load of M. bovis and more severe lesions than the control group. Those badgers were defined as a “divergent” HIMB-vaccinated badgers’ type. Conversely, the other six vaccinated animals showed an improvement in bacterial load and pathology with respect to the control group30. Those badgers were defined as “standard” HIMB-vaccinated badgers’ type. In the present study we performed immunohistochemistry (IHC) to evaluate the local immune response in lungs and lymph nodes (LNs) of the 15 badgers and correlated the results obtained at the last time-point (date killing and post-mortem examination). We selected relevant lesional, immunological and bacteriological variables influenced by the vaccine and challenge (Supplementary Material Raw Data). The lesion variable was the total volume of lung TB lesions measured by magnetic resonance imagery (MRI)30. The immune variables were the cellular-immune response by the ELISPOT technique (specifically with cocktail CFP-10/ESAT-6) and humoral immune response in a P22 antigen ELISA test. Colony-forming unit equivalents were calculated from readings in the Mycobacteria Growth Indicator Tube (MGIT)30. Only readings from the MGIT tubes were used, because the dynamic range in liquid culture (MGIT tubes) is greater than in solid culture, being more accurate for bacterial load assessment.
Immunohistochemistry (IHC)
Counts of specifically immune-marked cells (namely macrophages, T lymphocytes, B lymphocytes and plasma cells), and protein levels (cytokines; namely TGF-β, IL-10 and the transcription factor Fox-P3) were made on the tuberculous granulomas in the right middle lung lobe and right bronchial LN. The proteins were selected because of their association with immune depletion31,32,33. The right middle lung lobe was selected for the analysis because it was consistently affected with visible TB lesions, as expected following experimental challenge targeting this lobe of the lung. In addition, the right bronchial LN was selected because it was the most severely affected LN in both vaccinated and control animals. The specific details of the IHC protocols are summarized in Supplementary Table 1. Briefly, formalin fixed lung lobe and LN samples were cut into 3 μm sections, dewaxed, rehydrated and rinsed with tap water at room temperature (RT). Afterwards, epitope demasking was performed microwaving in sodium citrate buffer [1.48 g sodium citrate (Merck KGaA, Darmstadt, Germany) in 500 mL distilled water]. Then, the sections were washed twice with tris buffered saline 1X [TBS 1X, 5 mM Tris (Merck KGaA, Darmstadt, Germany)/HCl (Panreac Química, SLU, Barcelona, Spain) pH 7.6, 136 mM NaCl (Merck KGaA, Darstadt, Germany)]. Afterwards, the slides were placed in distilled water at 0.5% of hydrogen peroxide to block endogenous peroxidase activity for 30 min at RT and washed twice with TBS 1X. The samples were then treated to prevent primary antibody cross-reactivity with tissue constituents with a 20-min incubation period at RT, with 10% normal horse serum (Vector Laboratories, Newark, CA, USA) for CD3 (T lymphocytes) and TGF-β protocols or with 10% normal goat serum (Vector Laboratories, Newark, CA, USA) for IBA-1 (macrophages), CD20 (B lymphocytes), lambda light chain (plasma cells), IL-10 and Fox-P3 protocols, and 3% bovine serum albumin (BSA, Sigma-Aldrich, Saint Louis, MO, USA) in TBS for all proteins. Then, sections were incubated overnight at 4 °C with commercial monoclonal and polyclonal antibodies diluted 1:200 (Supplementary Table 1), and washed twice with TBS 1X. Afterwards, samples were incubated 30 min with the appropriate biotinylated secondary antibody (anti-mouse biotinylated in horse or anti-rabbit biotinylated in goat; Vector Laboratories, Newark, CA, USA) (Supplementary Table 1) at RT, before being washed twice with TBS 1X. The sections were incubated for 30 min at RT with Avidin Biotin Complex (ABC kit Peroxidase Standard, Vector Laboratories, Newark, CA, USA). Finally, the sections were washed twice with TBS 1X and the signal was detected using Vector®NovaRED peroxidase substrate kit SK-4800 (Vector Laboratories, Newark, CA, USA) for a 1 min incubation. Samples were washed twice with TBS 1X for 5 min, before being placed in Mayer’s hematoxylin (MerckKGaA, Darmstadt, Germany) for 30 s. Afterwards, the slides were rinsed with tap water, rehydrated, and mounted with DPX (MerckKGaA, Darmstadt, Germany). Controls were included in each IHC run. Those included positive controls (LNs from a healthy badger where the target antigen was present in the section and the specific antibody was used) and negative controls (slides with omission of the primary antibody to evaluate possible nonspecific labelling).
Immunohistochemical analyses and score assignement
A total of 120 (i.e., 15 individuals x 2 tissues x 4 types) and 90 (i.e., 15 individuals x 2 tissues x 3 proteins) records of immunostained cells and proteins, respectively, were obtained by optical microscopy. Cell immunolabelled sections were scanned at 400x using an Olympus BX51 microscope (Olympus, Tokyo, Japan). For each slide (both cells and proteins), five randomly selected areas were sourced as regions of interest (RI) and analyzed at 200x magnification.
For cell counting, an Olympus XC10 camera was used (Olympus, Tokyo, Japan). Scanned images and RI were analyzed with digital image analysis software (Nikon NIS-Elements Ar, Nikon, Japan). After setting the thresholds, the area with an immunohistochemical-positive reaction within RI was calculated by the Nikon NIS-Elements software. The results were expressed as the proportion of the positively immunolabelled area within the total area of the selected RI. The score assigned to each slide corresponded to the sum of positively immunolabelled percent area in the five RI.
For protein assessment, the slides were visually analyzed, and the results were expressed as the number of cells with an immunohistochemical-positive reaction within RI. These counts were categorized as following: 0 (absence of positive reaction), 1 (< 5 positive cells), 2 (5–10 positive cells), and 3 (> 10 positive cells). The score assigned to each slide corresponded to the sum of the positively immunolabelled markers in the five RI.
Statistical analysis
An exploratory factor analysis (EFA) by Principal Component Analysis (PCA) was performed to assess the underlying grouping of the individual’s immune response type and disease severity by identifying patterns of co-variation among variables. The total immunological scores of cells and proteins, the antibody ELISA P22 optical density (OD), the ELISPOT cocktail CFP-10/ESAT-6 (C/E), the MRI lesion volume and the M. bovis MGIT results were included in the analysis. Some data generated in LN were excluded from the PCA due to the absence of information for some of these variables. The estimating procedure was maximum likelihood. The resulting factors with eigenvalues > 1 were selected. Factor loadings were examined to interpret the relationships between variables and factors. The FactoExtra library in R software was used.
Generalized linear models were carried out to assess the differences in the immune response variables (individual badger sum of cell counts or protein scores for its five RI) between types and tissues. These models were carried out for the outcomes observed post-hoc in the previous experimental design (control, standard and divergent)30. For each estimated variable, a model was constructed in which tissue (lung and LN), response type group (standard, divergent and non-vaccinated/controls) and their interaction were introduced as explanatory variables. The significance of the differences among groups was explored through multiple pairwise comparison post-hoc analyses with the Tukey test. Statistical significance was accepted at p < 0.05.
In addition, a generalized linear model tested the differences in the mycobacterial loads between response type and tissues, including these variables and their interaction as explanatory variables based on a Poisson distribution model. From the marginal means of the model, the percentage reduction in bacterial load observed in the standard HIMB group compared to the control group was calculated using the following formula:
Results
Cell counts
The mean scores of cells, considering the tissue, response type, and vaccination response are displayed in Fig. 1. Representative histological sections are shown in Fig. 2.

Average cell scores detected in tuberculous granulomas from badgers. Scores were obtained from the right bronchial lymph node (LN) and right middle lung lobe (Lung) of 15 badgers according to the group (control, standard HIMB and divergent HIMB). Error bars represent the standard error of the mean. HIMB: heat-inactivated Mycobacterium bovis vaccine.

Immunohistochemical characterization of cell populations forming the tuberculous granulomas in badgers. Cell population is shown in the right bronchial lymph node (a-f, l) and right middle lung lobe (j-k) in control, standard heat-inactivated M. bovis (HIMB)-vaccinated and divergent HIMB-vaccinated badger groups: macrophages (a-c), T lymphocytes (d-f), B lymphocytes (g-i), and plasma cells (j-l). Positive immunolabelling is shown in red color. ABC complex technique, bars = 100 microns. The images from tissues (lung or lymph node) were selected based on the most representative results. Arrows in the image j indicate the immunopositivity.
For macrophages, the divergent response type group displayed significantly lower levels compared to the other groups (p < 0.01) (Fig. 3a & Supplementary Table 2a). In terms of tissue distribution, macrophage levels in TB lesions were lower in the LN than in the lung in all types (p = 0.02; Fig. 3a & Supplementary Table 2b).

Predicted cell scores in tuberculous granulomas depending on the tissue and group. (a) Macrophages, (b) T lymphocytes, (c) B lymphocytes, and (d) plasma cells. Note that the significance of the differences between groups and tissues is displayed in Supplementary Tables 2–5. HIMB: heat-inactivated Mycobacterium bovis vaccine. LN: lymph node.
For T lymphocytes, overall, individuals with a divergent response also showed significantly lower levels than the control group (p = 0.02; Fig. 3b), and the LN scores were notably higher than the lung scores (p < 0.01; Supplementary Table 3a). The marked differences observed among response groups were mainly attributed to the LN scores. In the controls and standard, the scores were lower in the lung than in the LN, whereas no difference between tissues was detected for the divergent type (p < 0.01; Fig. 3b & Supplementary Table 3b).
The lymphocyte B distribution of the divergent type significantly differed from that exhibited by the other groups (p < 0.01) (Fig. 3c). Scores observed in lung were significantly lower than those detected in the LN for standard and control groups (p standard HIMB = 0.02 and p control = 0.01), while the opposite occurred for the divergent (p = 0.05), showing significant higher values in the lung than the standard and control groups (p < 0.01 and p = 0.01, respectively; Figs. 2 and 3c & Supplementary Tables 4a and b).
For plasma cells, individuals with a divergent response exhibited significantly higher cell levels than the other groups (p to standard HIMB = 0.06 and p to control < 0.01), and the lung scores were marginally higher than the LN scores (p = 0.06) (Figs. 2 and 3d & Supplementary Table 5a). Differences between groups were particularly pronounced for the LN, with the control group showing significantly lower plasma cell values than both the divergent (p < 0.01; Fig. 2) and the standard groups (p = 0.05), being these LN scores lower than those observed in the lung in this control group (p < 0.01, Fig. 3d & Supplementary Table 5b).
Protein scores
Figure 4 display the mean scores of proteins considering the tissue, treatment group, and type of response to vaccination. Representative histological sections are shown in Fig. 5.

Average protein/cytokine scores detected in tuberculous granulomas in badgers. Protein scores were obtained from the right bronchial lymph node (LN) and right middle lung lobe (lung) of 15 badgers according to the group (control, standard HIMB and divergent HIMB). Error bars represent the standard error of the mean. HIMB: heat-inactivated Mycobacterium bovis vaccine.

Immunocharacterization of protein expression in tuberculous granulomas of badgers. Results are shown in the right bronchial lymph node (a,d) and right middle lung lobe (b,c) in standard heat-inactivated M. bovis (HIMB)-vaccinated (b) and divergent HIMB-vaccinated (a,c,d) badger groups. (a) TGF-β in the divergent HIMB-vaccinated group, (b) IL-10 in the standard HIMB-vaccinated group, (c) IL-10 in the divergent HIMB-vaccinated group, and (d) Fox-P3 in the divergent HIMB-vaccinated group. Positive immunolabelling is shown in red color. ABC complex technique, bars = 100 microns. The images from tissues (lung or lymph node) were selected based on the most representative results.
For TGF-β, the HIMB individuals (divergent and standard response) showed significantly higher cytokine levels than the control group (p < 0.01; Figs. 5 and 6a & Supplementary Table 6a). However, when examining the distribution of TGF-β between tissues, individuals with a standard response behaved completely differently from both the divergent response group and control group, displaying significantly lower TGF-β scores in the LN than in the lung (p = 0.01) (Supplementary Table 6b). The divergent response group and control group showed a similar distribution pattern, with no difference in TGF-β levels between tissues (Fig. 6a & Supplementary Table 6b).

Predicted score of proteins depending on the tissue and group. (a) TGF-β, (b) IL-10, and (c) Fox-P3. Note that the significance of the differences between groups and tissues is displayed in Supplementary Tables 6–8. HIMB: heat-inactivated Mycobacterium bovis vaccine. LN: lymph node.
For IL-10, badgers with a divergent response exhibited statistically higher levels of IL-10 than standard and control group in both tissues (p < 0.01; Figs. 5 and 6b & Supplementary Tables 7a and b).
For Fox-P3, the divergent type also exhibited statistically higher levels for this protein than the standard and control groups (p < 0.01 for both comparisons; Supplementary Table 8a). The significant differences between tissues observed were primarily attributed to the divergent group (Fig. 6c), which exhibited significantly higher levels of Fox-P3 in the LN compared to the lung. However, no significant differences in Fox-P3 scores were detected in the lung among the groups (Fig. 6c & Supplementary Table 8b).
M. bovis counts
In all three groups, M. bovis counts were consistently higher in the LN than in the lung (p < 0.01) (Fig. 7). Moreover, the divergent type response group showed significantly higher M. bovis counts compared to the other groups (p < 0.01), with LN values still exceeding the counts obtained in the lung. A significant reduction in bacterial load in the standard response type group compared to control (85.5% and 99.9%) was observed in LN and lung, respectively (p < 0.01) (Supplementary Table 9).

Predicted Mycobacterium bovis counts (colony-forming units per gram) depending on tissue and group. HIMB: heat-inactivated Mycobacterium bovis vaccine. LN: lymph node.
Factorial analysis
Two uncorrelated factors accounted for 60.5% of co-variation among parameters measured in the right middle lung lobe. Elevated levels of IL-10, Fox-P3 and TGF-β, M. bovis counts, MRI volume lung lesions (results published in Juste et al.30), and low macrophages were associated with higher factor 1 values, while higher factor 2 values were indicative of increased B lymphocytes and higher responses to ELISPOT cocktail C/E levels, as well as reduced plasma cells.
The factorial analyses revealed four different patterns of immune response according to how the measured parameters were associated with each other: (i) B lymphocytes were linked to T lymphocytes and ELISPOT cocktail C/E; (ii) proteins (IL-10, Fox-P3 and TGF-β) were grouped together with M. bovis counts and volume lesions; (iii) plasma cells were positively related to the ELISA P22 results; and (iv) macrophages were separated and opposite to protein expression and disease severity levels (Fig. 8).

Projections of the 15 badgers on the two-dimensional space. The space was defined by principal component axes 1 and 2 (Factor 1 and 2) according to their type of response to treatment. Arrows reflect the direction and contribution to the axes of each of the seven parameters included in the principal component analysis (PCA). Confidence 95% ellipses are displayed. Post-mortem C/E cocktail refers to CFP-10 and ESAT-6 cellular immune response at post-mortem measured by ELISPOT. Post-mortem P22 refers to humoral immune response at post-mortem measured by ELISA P22. CFU: colony-forming units. HIMB: heat-inactivated Mycobacterium bovis vaccine.
Discussion
This study describes the local cell and protein response pattern in badgers following the administration of the HIMB vaccine via edible baits and subsequent challenge with 103 CFU of M. bovis delivered endobronchially. We identified significantly differential outcomes between vaccinated individuals who developed exacerbated TB lesions (divergent response HIMB vaccinated type) and those for whom the vaccine granted partial protection (standard response HIMB vaccinated type), thus confirming a model of HIMB vaccination response with an unknown factor greatly affecting the pathological and microbiological vaccine effects. In that regard, the local immune response in the divergent response group was primarily characterized by elevated IL-10 levels, fewer macrophages in both LN and lung tissues and higher scores of Fox-P3 and B lymphocytes in the LN and right middle lung lobe, respectively. Overall, the PCA analysis provided valuable insights into the underlying grouping of the individuals studied, facilitating a deeper understanding of the different types of immune responses to the HIMB vaccination and subsequent infection outcomes.
In addition, the mycobacterial load slightly increased in the vaccinated group if all the vaccinated badgers were considered together. However, if the different response groups were segregated, the standard response type presented a bacterial load reduction that reached 99% in the lung, whereas the vaccinated badgers with a divergent response presented very high bacterial loads in the lung and draining LNs (3.54 × 108 CFU/mL vs. 1.78 × 106 controls and 2.4 × 105 vaccinated with a standard response). Reductions of the bacterial load observed in the badgers with a standard response compared to controls revealed that HIMB vaccination induced a greater protective response in the lungs (99% reduction) than in the LN (84% reduction), suggesting two functional compartments and indicating that TB spread from the lung can be effectively hampered by vaccine12,23. In this regard, the lung is the pathogenetically and epidemiologically critical organ of M. bovis entrance/exit34. The interaction between vaccine and tissue has been reported in humans by Silver et al. who showed that BCG oral vaccination induced limited antigen specific systemic immunity, but distinct pulmonary responses in bronchoalveolar lavage35. In addition, distinct gene expression signatures in blood and bronchoalveolar lavage comparing latent TB infection with different routes of BCG vaccination were observed35. Differential pathogenesis and immune responses between tissues (nasal mucosa and blood) were also recently observed in SARS-CoV-2-challenged humans resulting in either sustained or transient viral replication36. This would be in agreement with the results of several experimental and field studies assessing the efficacy of oral BCG and HIMB vaccines in badgers where it was concluded that while vaccination did not confer complete protective immunity, it generally reduced mycobacterial excretion and volume of lesions, thereby reducing the spread of the disease12,30,37,38,39. However, vaccination can sometimes exacerbate manifestations of disease rather than mitigate them30,37,40, in line with the more severe lesions and greater weight loss observed in badgers with a divergent response30 and in BCG-vaccinated badgers35. Several hypotheses have been proposed to explain this particular type of response: (i) a shift of the immune response from Th1 to Th2, since the Th1/Th2 balance is critical for determining TB disease progression and outcomes41, (ii) an enhanced lymphatic spread of the bacteria, which has been observed associated with lymphoma in other studies involving oral BCG in badgers42, and (iii) depletion of T-cells unable to eliminate the mycobacteria due to persistent antigen and chronic T-cell stimulation43. Additionally, the possibility of the bacteria entering the bloodstream during the bacillus inoculation procedure has been considered30.
Our PCA results confirmed that the responses observed in this experiment concurred with what could be expected under the TRAIM model44,45. Factor 1, the one carrying the best part of variability (60%) would represent the TRAIM non-specific mechanisms where bacterial load and lesions were positively associated with immune activity dampening proteins IL-10, Fox-P3 and TGF-β on one end, while macrophage activation was maximum in the opposite end. Transversal factor 2 carrying the variability associated to B and T lymphocytes and ELISPOT C/E-specific response on the positive side and plasma cells and ELISA P22-specific humoral response on the negative end would represent the adaptive component of the immune response. The overall picture would therefore suggest that both bacterial load and lesions do not correlate well with the specific immune response, but rather with the non-specific TRAIM. This gives an important clue on TB pathogenesis as it confirms that the adaptive immune response would play a secondary, if not rather collateral, role in the defense against mycobacteria load and lesions caused by them. It also suggests that specific immunity has a limited utility in terms of diagnosis. Ultimately, this prompts to further study on whether any of the immune mediators associated with damage and bacterial load in this study or a combination of them might be a good indicator of TB pathogenesis and, therefore, have a use for vaccine protection and infection diagnostic assessment.
For an effective response to vaccination, adequate control of the inflammatory response is necessary to develop immunological memory without causing excessive tissue damage46,47. In this line, the standard response type displayed a balanced Th1/Th2 immune response. The macrophage activation was elevated, suggesting a robust and effective innate immune response, while the levels of IL-10 and TGF-β cytokines were moderate, thus preventing excessive tissue damage. Although plasma cell and B lymphocyte levels were only marginally higher in the lungs than in the LN, this group showed effective antibody production primarily focused on lymphatic tissues.
On the contrary, the divergent response type, characterized by exacerbated TB lesions and higher bacterial load, showed reinforced protein and humoral (Th2) responses, with elevated scores of IL-10, Fox-P3, B lymphocytes and plasma cells, particularly in the lung, compared to standard vaccinated and control groups. These badgers also displayed lower T lymphocyte (Th1) and macrophage levels. According to our results, three general characteristics of this divergent immune response could be behind these TB outcomes:
-
(i)
Anti-inflammatory immune mediators´ activation: the elevated scores of IL-10, TGF-β, and Fox-P3 mirror an increased regulatory immune response, which might potentially inhibit an effective immune response against TB48, leading to higher M. bovis loads and more severe lesions31,49,50. Specifically, IL-10 has been recognized as a promising cytokine biomarker of M. bovis infection and disease severity in both domestic and wildlife species32,51,52. While IL-10 initially helps control inflammation and tissue damage, sustained high levels are related to greater disease severity. In the early stages of infection, IL-10 limits inflammation and the subsequent tissue damage by deactivating macrophages and reducing its production of reactive nitrogen and oxygen species, as well as IFN-γ levels32,33. However, constantly high IL-10 levels can hamper the Th1 response, being associated with increased susceptibility to M. bovis infection and more severe disease progression53,54, as observed in studies where animals with higher IL-10 activation exhibited higher lesion scores and bacterial loads50,52,55,56,57. The PCA results concurred with this mechanism, clustering bacterial load, IL-10 scores and MRI lesion volume contrary to macrophage scores in factor 1 (innate immune response). TGF-β was also linked to lesion volume and bacterial loads in the PCA, i.e., it was significantly higher in badgers with a divergent response compared to individuals with standard response and controls. This cytokine suppresses protective immune responses against M. bovis by promoting the elimination of post-activated T-cells49, thus increasing TB persistence and chronicity. The same effect was reported for M. tuberculosis58. TGF-β can also reduce the antimicrobial function of macrophages, promoting fibrosis, especially in the lung31. Finally, Fox-P3 is a transcription factor for T regulatory cells (Fox-P3 + Tregs), which is relevant in regulating/moderating the immune response, balancing effective defense against M. bovis and, preventing excessive tissue damage through the promotion of anti-inflammatory cytokines such as IL-10 and TGF-β, and the suppression of T and other immune cells, thus maintaining immunological tolerance59,60. Previous studies suggested that M. bovis and other pathogens can exploit this regulation capability to evade the immune response61,62,63. The elevated Fox-P3 + Tregs activity, along with lower levels of T cells observed in this study, may be associated with the reduced efficacy of the HIMB vaccine in the vaccinated group with a divergent response.
-
(ii)
A Th1/Th2 imbalanced immune response: the lower levels of T cells and macrophages (debilitated Th1 response) together with marked regulatory response driven by cytokines such as IL-10 may limit an effective defense33.
-
(iii)
Enhanced but localized humoral immune response: the increased activation of B lymphocytes and plasma cells in the lungs suggested a strong humoral immune response44, although effective TB control depends largely on cellular immunity64,65, which seemed to be limited in the divergent type. This marked but localized humoral immune response might not have been sufficient to effectively control TB dissemination, leading to the exacerbated lesions observed. Indeed, differences in B lymphocyte levels confirmed discrepancy in the efficacy of the HIMB vaccine, since the vaccinated group with a standard response had significantly lower B lymphocyte scores than controls in both tissues. However, the differences between tissues in badgers with standard and divergent responses highlighted the stronger humoral Th2 response of the badgers with a divergent response, also observed in the PCA, and characterized by exacerbated lesions with high lytic activity and tissue necrosis. Furthermore, lower T cell levels in this group suggested a reduced Th1 cellular response. Therefore, assuming that regulatory and effector responses have a genetic and environmental background, extraordinary care should be taken in selecting closely related individuals at their early days of life for experimental purposes, to minimize the genetic variability noise and to avoid exposure to environmental TRAIM-inducing agents. On the other hand, attention must also be given to the route and dose of vaccine and challenge administration as both variables might be relevant in overwhelming the TRAIM response and triggering the inefficient, potentially damaging, adaptive immune response.
Finally, it should be taken into consideration that this study has several limitations. An important constraint was the random and variable conditions (individual and behavioral traits, health status, as well as environmental and climatic factors) of free-ranging trapped badgers. In addition, the intake of bait by the badgers, which is critical in the vaccination process, may have varied among individuals, leading to different levels of exposure to the vaccine. These factors might lead to potential variability in the immune responses observed. Given the random nature of badger acquisition and the lack of standardization or homogenization, it is possible that individuals with inherently different immune characteristics regarding TB response were captured, suggesting the presence of two distinct response types rather than a single homogeneous group. In this regard, the small sample size in each experimental group may limit the statistical power of the findings. However, rather than attributing the observed variability in the immune response exclusively to individual differences, this study leveraged the unexpected divergence in the immune response to describe and assess differences in the response to vaccination. In addition, the study demonstrates that these differences exist based on statistically significant variations in immunological and pathological variables.
Conclusions
The badgers with a divergent response showed an immune response characterized by increased local activity of regulatory immune mediators and enhanced humoral immune response, which may have limited the protective capacity of responses against M. bovis challenge. Given the complexity of immune responses to M. bovis, it is highly likely that other parameters will play a role. However, the hypothesis of unbalanced immunomodulation (Th1/Th2) should be considered as a bias to responses to HIMB (and BCG) vaccination programs in the future. Therefore, further studies should focus on optimizing the balance between regulatory and effector responses to enhance protective immunity against M. bovis.
Data availability
Data is provided within the manuscript or supplementary information files.
References
Thoen, C. O., Steele, J. H. & Gilsdorf, M. J. in Mycobacterium Bovis Infection in Animals and Humans. 1–315 (eds Thoen, C. O., Steele, J. H. & Gilsdorf, M. J.) (Wiley, Iowa State University, 2006).
Zinsstag, J. et al. Cost-effective control strategies for animal and zoonotic diseases in pastoralist populations. Rev. Sci. Tech. L’OIE. 35, 673–681 (2016).
Santos, N. et al. Quantification of the animal tuberculosis Multi-Host community offers insights for control. Pathogens 9, 421 (2020).
Barasona, J. A., Gortázar, C., de la Fuente, J. & Vicente, J. Host richness increases tuberculosis disease risk in game-managed areas. Microorganisms 7, 182 (2019).
Gortázar, C., Che Amat, A. & O’Brien, D. J. Open questions and recent advances in the control of a multi-host infectious disease: animal tuberculosis. Mamm. Rev. 45, 160–175 (2015).
Acevedo, P. et al. Tuberculosis epidemiology and Badger (Meles meles) Spatial ecology in a Hot-Spot area in Atlantic Spain. Pathogens 8, 292 (2019).
Allen, A. R., Skuce, R. A. & Byrne, A. W. Bovine tuberculosis in Britain and Ireland – A perfect storm?? The confluence of potential ecological and epidemiological impediments to controlling a chronic infectious disease. Front. Vet. Sci. 5, 109 (2018).
Byrne, A. W., White, P. W., McGrath, G., O′Keeffe, J. & Martin, S. W. Risk of tuberculosis cattle herd breakdowns in ireland: effects of Badger culling effort, density and historic large-scale interventions. Vet. Res. 45, 109 (2014).
Chambers, M. A. et al. Bacillus Calmette-Guerin vaccination reduces the severity and progression of tuberculosis in badgers. Proc. R. Soc. B Biol. Sci. 278, 1913–1920 (2011).
Carter, S. P. et al. BCG vaccination reduces risk of tuberculosis infection in vaccinated badgers and unvaccinated Badger Cubs. PLoS One. 7, e49833 (2012).
Gormley, E. et al. Protective immunity against tuberculosis in a free-living Badger population vaccinated orally with Mycobacterium Bovis Bacille Calmette–Guérin. Transbound. Emerg. Dis. 69, e10–e19 (2022).
Balseiro, A. et al. Protective effect of oral BCG and inactivated Mycobacterium Bovis vaccines in European badgers (Meles meles) experimentally infected with M. Bovis. Front. Vet. Sci. 7, 41 (2020).
Díez-Delgado, I. et al. Parenteral vaccination with Heat-Inactivated Mycobacterium Bovis reduces the prevalence of Tuberculosis-Compatible lesions in farmed wild Boar. Transbound. Emerg. Dis. 64, e18–e21 (2017).
Pandey, A. et al. Wildlife vaccination strategies for eliminating bovine tuberculosis in white-tailed deer populations. PLOS Comput. Biol. 20, e1011287 (2024).
Tompkins, D. M. et al. Oral vaccination reduces the incidence of tuberculosis in free-living brushtail possums. Proc. R. Soc. B Biol. Sci. 276, 2987–2995 (2009).
Jones, G. J. et al. Oral vaccination of cattle with heat inactivated Mycobacterium Bovis does not compromise bovine TB diagnostic tests. Vet. Immunol. Immunopathol. 182, 85–88 (2016).
Díez-Delgado, I. et al. Impact of piglet oral vaccination against tuberculosis in endemic free-ranging wild Boar populations. Prev. Vet. Med. 155, 11–20 (2018).
Garrido, J. M. et al. Protection against tuberculosis in Eurasian wild Boar vaccinated with Heat-Inactivated Mycobacterium Bovis. PLoS One. 6, e24905 (2011).
Beltrán-Beck, B. et al. Oral vaccination with heat inactivated Mycobacterium Bovis activates the complement system to protect against tuberculosis. PLoS One. 9, e98048 (2014).
Nol, P. et al. Effects of inactivated Mycobacterium Bovis vaccination on Molokai-Origin wild pigs experimentally infected with virulent M. Bovis. Pathogens 9, 199 (2020).
Thomas, J. et al. The response of red deer to oral administration of heat-inactivated Mycobacterium Bovis and challenge with a field strain. Vet. Microbiol. 208, 195–202 (2017).
Hewlett, J., Juste, R. A., Rutten, V., Gortázar, C. & Michel, A. Control and management of bovine tuberculosis in African buffalo: cell-mediated immune responses following vaccination and Mycobacterium bovis challenge in: Seventh International Conference on Mycobacterium bovisGalway, Ireland, (2022).
Blanco Vázquez, C. et al. Local lung immune response to Mycobacterium Bovis challenge after BCG and M. Bovis Heat-Inactivated vaccination in European Badger (Meles meles). Pathogens 9, 456 (2020).
Canfield, P. J., Day, M. J., Gavier-Widen, D., Hewinson, R. G. & Chambers, M. A. Immunohistochemical characterization of tuberculous and Non-tuberculous lesionsin naturally infected European badgers (Meles meles). J. Comp. Pathol. 126, 254–264 (2002).
Juste, R. A. et al. Increased lytic efficiency of bovine macrophages trained with killed mycobacteria. PLoS One. 11, e0165607 (2016).
Risalde, M. A. et al. Control of mycobacteriosis in zebrafish (Danio rerio) mucosally vaccinated with heat-inactivated Mycobacterium Bovis. Vaccine 36, 4447–4453 (2018).
Contreras, M. et al. Oral vaccination with a formulation combining rhipicephalus Microplus Subolesin with heat inactivated Mycobacterium Bovis reduces tick infestations in cattle. Front. Cell. Infect. Microbiol. 9, 45 (2019).
Byrne, K. A., Tuggle, C. K. & Loving, C. L. Differential induction of innate memory in Porcine monocytes by β -glucan or Bacillus Calmette-Guerin. Innate Immun. 27, 448–460 (2021).
Netea, M. G. et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 20, 375–388 (2020).
Juste, R. A. et al. Efficacy of heat-inactivated Mycobacterium Bovis vaccine delivered to European badgers (Meles meles) through edible bait. Heliyon 9, e19349 (2023).
Walton, K. L., Johnson, K. E. & Harrison, C. A. Targeting TGF-β mediated SMAD signaling for the prevention of fibrosis. Front. Pharmacol. 8, 461 (2017).
Widdison, S. et al. Cytokine expression profiles of bovine lymph nodes: effects of Mycobacterium Bovis infection and Bacille Calmette–Guérin vaccination. Clin. Exp. Immunol. 144, 281–289 (2006).
Welsh, M. D. et al. Influence of pathological progression on the balance between cellular and humoral immune responses in bovine tuberculosis. Immunology 114, 101–111 (2005).
Behr, M. A. & Waters, W. R. Is tuberculosis a lymphatic disease with a pulmonary portal? Lancet Infect. Dis. 14, 250–255 (2014).
Silver, R. F. et al. Distinct gene expression signatures comparing latent tuberculosis infection with different routes of Bacillus Calmette-Guérin vaccination. Nat. Commun. 14, 8507 (2023).
Lindeboom, R. G. H. et al. Human SARS-CoV-2 challenge uncovers local and systemic response dynamics. Nature 631, 189–198 (2024).
Lesellier, S. et al. Bioreactor-Grown Bacillus of Calmette and Guérin (BCG) Vaccine Protects Badgers against Virulent Mycobacterium bovis When Administered Orally: Identifying Limitations in Baited Vaccine Delivery. Pharmaceutics 12, 782 (2020).
Gormley, E. et al. Oral vaccination of Free-Living badgers (Meles meles) with Bacille calmette Guérin (BCG) vaccine confers protection against tuberculosis. PLoS One. 12, e0168851 (2017).
Chambers, M. A. et al. The effect of oral vaccination with Mycobacterium Bovis BCG on the development of tuberculosis in captive European badgers (Meles meles). Front. Cell. Infect. Microbiol. 7, 6 (2017).
Tizard, I. R. Adverse consequences of vaccination. in Vaccines for Veterinarians (ed. Tizard, I. R.) 115–130 (Elsevier, 2021).
Blanco, F. C. et al. Study of the immunological profile towards Mycobacterium Bovis antigens in naturally infected cattle. Microbiol. Immunol. 53, 460–467 (2009).
Bianco, C. et al. Subclinical BCG-osis in a captive Badger (Meles meles) with lymphoma. J. Comp. Pathol. 176, 76–80 (2020).
Blank, C. U. et al. Defining ‘T cell exhaustion’. Nat. Rev. Immunol. 19, 665–674 (2019).
Marshall, J. S., Warrington, R., Watson, W. & Kim, H. L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 14, 49 (2018).
Ochando, J., Mulder, W. J. M., Madsen, J. C., Netea, M. G. & Duivenvoorden, R. Trained immunity — basic concepts and contributions to immunopathology. Nat. Rev. Nephrol. 19, 23–37 (2023).
Crișan, T. O., Netea, M. G. & Joosten, L. A. B. Innate immune memory: implications for host responses to damage-associated molecular patterns. Eur. J. Immunol. 46, 817–828 (2016).
Siegrist, C. A. Vaccine immunology in Vaccines (eds. Plotkin, S. A., W.A., O. & P.A., O.) 17–36 (Elsevier Health Sciences, 2008).
Cooper, A. M. & Khader, S. A. The role of cytokines in the initiation, expansion, and control of cellular immunity to tuberculosis. Immunol. Rev. 226, 191–204 (2008).
Hernández-Garay, M. & Méndez-Samperio, P. Transforming growth Factor-β decreases survival of Mycobacterium bovis-Activated T cells. Arch. Med. Res. 34, 20–25 (2003).
Gonzalez-Juarrero, M. et al. Experimental aerosol Mycobacterium Bovis model of infection in goats. Tuberculosis 93, 558–564 (2013).
Blanco, F. C., Bigi, F. & Soria, M. A. Identification of potential biomarkers of disease progression in bovine tuberculosis. Vet. Immunol. Immunopathol. 160, 177–183 (2014).
Smith, K., Kleynhans, L., Warren, R. M., Goosen, W. J. & Miller, M. A. Cell-Mediated immunological biomarkers and their diagnostic application in livestock and wildlife infected with Mycobacterium Bovis. Front. Immunol. 12, 639605 (2021).
Palmer, M. V., Thacker, T. C. & Waters, W. R. Analysis of cytokine gene expression using a novel chromogenic In-situ hybridization method in pulmonary granulomas of cattle infected experimentally by aerosolized Mycobacterium Bovis. J. Comp. Pathol. 153, 150–159 (2015).
Jacobs, M., Fick, L., Allie, N., Brown, N. & Ryffel, B. Enhanced immune response in Mycobacterium Bovis Bacille calmette Guerin (BCG)-Infected IL-10-Deficient mice. Clin. Chem. Lab. Med. 40, 9 (2002).
Wong, E. A. et al. IL-10 impairs local immune response in lung granulomas and lymph nodes during early Mycobacterium tuberculosis infection. J. Immunol. 204, 644–659 (2020).
Moreira-Teixeira, L. et al. T Cell–Derived IL-10 impairs host resistance to Mycobacterium tuberculosis infection. J. Immunol. 199, 613–623 (2017).
Canal, A. M. et al. Immunohistochemical detection of pro-inflammatory and anti-inflammatory cytokines in granulomas in cattle with natural Mycobacterium Bovis infection. Res. Vet. Sci. 110, 34–39 (2017).
Gern, B. H. et al. TGFβ restricts expansion, survival, and function of T cells within the tuberculous granuloma. Cell. Host Microbe. 29, 594–606 (2021).
Sakaguchi, S., Yamaguchi, T., Nomura, T. & Ono, M. Regulatory T cells and immune tolerance. Cell 133, 775–787 (2008).
Josefowicz, S. Z., Lu, L. F. & Rudensky, A. Y. Regulatory T cells: mechanisms of differentiation and function. Annu. Rev. Immunol. 30, 531–564 (2012).
Shevach, E. M. Mechanisms of Foxp3 + T regulatory Cell-Mediated suppression. Immunity 30, 636–645 (2009).
Scott-Browne, J. P. et al. Expansion and function of Foxp3-expressing T regulatory cells during tuberculosis. J. Exp. Med. 204, 2159–2169 (2007).
Shafiani, S., Tucker-Heard, G., Kariyone, A., Takatsu, K. & Urdahl, K. B. Pathogen-specific regulatory T cells delay the arrival of effector T cells in the lung during early tuberculosis. J. Exp. Med. 207, 1409–1420 (2010).
Krocova, Z. et al. The role of B cells in an early immune response to Mycobacterium Bovis. Microb. Pathog. 140, 103937 (2020).
Carpenter, E., Fray, L. & Gormley, E. Cellular responses and Mycobacterium Bovis BCG growth Inhibition by bovine lymphocytes. Immunol. Cell. Biol. 75, 554–560 (1997).
Acknowledgements
The authors thank Natalia García Álvarez for her collaboration in this study.
Funding
This work is the result of the I + D + i research project RTI2018-096010-B-C21, funded by the Spanish MCIN/AEI/10.13039/501100011033/Ministry of Science, Innovation and the European Regional Development Funds (FEDER Una manera de hacer Europa). This work was also partially funded by PID2022-141906OB-C22 funded by MCIN/AEI/10.13039/50110 0 011033/FEDER, UE; the Principality of Asturias, PCTI 2024–2027 (GRUPIN: IDI/2024/000764) and European Regional Development Fund. The cost of the running MRI was financed by the European Commission in the context of Horizon 2020 – Vetbionet Transnational Access Activities (TNA) call. DEFRA also funded APHA staff for their participation in the study. Patricia Barroso was supported by a Juan de la Cierva contract 495 [DC2022-049103-I]. Beatriz Sañudo was supported by a ULE-Santander fellowship funded by the University of León and Banco de Santander. Cristina Blanco Vázquez was granted with a predoctoral fellowship CPD2016-0142 funded by MCIN/AEI/10.13039/501100011033 and FSE. Gloria Herrero-García was granted with a predoctoral fellowship by Junta de Castilla y León and FSE (LE036-20).
Author information
Authors and Affiliations
Contributions
RJ, MB and AB performed the fieldwork. BS, DB, GHG, CBV, MB, SL, DD, JG, IAS, RS and AB performed the laboratory analysis. PB performed the statistical analysis. AB conceptualized the study. MB and AB obtained the funding. PB, RJ and AB wrote the first draft of the manuscript. All authors contributed critically to the final draft and approved for publication.
Corresponding author
Ethics declarations
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Ethical approval
The study is reported in accordance with ARRIVE guidelines. All methods were performed following relevant guidelines and regulations. All experimental protocols were approved by Ethical Committees from the Government of Principality of Asturias and the Diputación Foral de Bizkaia with license reference numbers: PROAE 47/2018 and 201944.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Barroso, P., Juste, R., Sañudo, B. et al. Differences in local immune response of bait Mycobacterium bovis heat-inactivated vaccinated badgers showing exacerbated tuberculous lesions after challenge. Sci Rep 15, 36329 (2025). https://doi.org/10.1038/s41598-025-20278-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-20278-1
