
Abstract
This research focuses on enhancing the extraction efficiency of Phylloporia ribis and assessing its biological functions. Key parameters including extraction temperature, duration, and ethanol-to-water ratio were optimized through both Response Surface Methodology (RSM) and an integrated Artificial Neural Network–Genetic Algorithm (ANN-GA) approach. The extracts obtained via ANN-GA exhibited greater antioxidant activity and higher concentrations of phenolic constituents such as gallic acid, quercetin, and vanillic acid. Compared to RSM-optimized samples, ANN-GA extracts demonstrated superior free radical scavenging, stronger ferric reducing power, and a more potent dose-dependent inhibition of cell proliferation. In addition, P. ribis extracts showed enzyme-inhibitory properties against acetylcholinesterase and butyrylcholinesterase, suggesting their potential utility in pharmaceutical and biotechnological applications. The ANN-GA method appears to be a promising tool for maximizing both the yield of phenolic compounds and the biological efficacy of extracts. Further advanced biotechnological optimization studies are advised to unlock the full therapeutic potential of P. ribis.
Similar content being viewed by others



Introduction
Medicinal mushrooms are natural organisms that can have multifaceted effects on human health thanks to the bioactive components they contain. They attract attention with their properties of balancing the immune system, supporting cellular functions and reducing oxidative stress1,2. These mushrooms, which have been used in traditional treatment approaches for centuries, are now better understood through scientific research and are evaluated in various pharmacological applications. They offer potential support in the prevention and treatment of many chronic diseases, especially thanks to their antioxidant and anti-inflammatory effects. Increasing clinical studies further reinforce the importance of medicinal mushrooms in modern health3.
In order to obtain the highest yield from medicinal mushrooms, the extraction process must be carefully planned. Variables such as the method used, solvent type, temperature, time and pressure have a direct effect on the concentration and bioavailability of the obtained bioactive components4. Selecting appropriate extraction techniques is of great importance for the effective release of targeted active components. Polysaccharides, phenolic compounds and terpenoids found in mushrooms can be obtained with higher purity by using water-based, alcohol-based or combined extraction methods. Determining the optimum extraction conditions is an important step that both increases therapeutic efficacy and supports sustainability in industrial production5,6,7.
In our study, Phylloporia ribis (Schumach.) Ryvarden was used as material. Phylloporia (Hymenochaetaceae) is a polypore fungus genus. It has been reported that there are 30 species belonging to the genus. Fungi belonging to the Phylloporia genus are generally known for their species developing as parasites on living leaves and branches. However, some species, such as P. minutispora and P. spathulata, can be found on fallen plant material or decomposing wood under the soil. Fungi belonging to this genus have monomitic or dimitic hyphal structures, and their tissue structures can be homogeneous or heterogeneous. Their basidiocarps can develop in resupinate, capped or petiolate forms and can have an annual or perennial life cycle. Their feeding strategies may differ as detritivores or parasites. However, the common feature of Phylloporia species is that they produce small-sized, colorful and thick-walled basidiospores8,9.
Despite the growing interest in medicinal mushrooms, P. ribis remains a largely understudied species with limited data on its bioactive profile and extraction efficiency. To date, no systematic optimization study has been reported for this species. Therefore, the present work addresses this critical knowledge gap by performing, for the first time, a comparative optimization of P. ribis extraction using both classical statistical modeling (RSM) and artificial intelligence-driven approaches (ANN-GA). Establishing this premise highlights the novelty and significance of our research in bridging traditional methodologies with modern computational techniques.
Material and methods
The mushroom specimens utilized in this study were collected from the Ankara region of Türkiye. Voucher specimens were deposited at the Department of Biology, Osmaniye Korkut Ata University. Taxonomic identification of the fungal material was performed by Dr. Ilgaz Akata. After collection, the mushroom specimens were cleaned of debris using a sterile brush and gently rinsed with distilled water. They were then air-dried at room temperature for 48 h to remove surface moisture. Subsequently, the dried samples were stored in airtight containers and kept at –20 °C until extraction to preserve their biochemical integrity.
Extraction protocols
To develop the extraction procedure, a full factorial experimental design was implemented, considering three key parameters: extraction temperature, duration, and the ethanol-to-water ratio. Each variable was tested at three levels, resulting in a total of 27 different extraction trials using a Soxhlet extraction system. Soxhlet extraction was selected because it provides continuous solvent cycling and exhaustive extraction, which are essential for reliable quantitative comparisons. This method also minimizes variability between replicates and ensures reproducibility, thereby making it particularly suitable for systematic optimization studies such as the present work. The solvent-to-solid ratio was kept constant across all experiments at 100 mL solvent per 10 g of dried mushroom material (equivalent to a 10:1 mL/g ratio) to isolate the effects of temperature, time, and solvent composition on extraction performance. To ensure precise temperature control, we used an automated Gerhardt SOX-414 Soxhlet apparatus. Unlike conventional Soxhlet systems, this instrument is equipped with a programmable heating module and reflux monitoring, allowing the extraction process to be carried out at defined temperatures (45 °C, 55 °C, 65 °C) independent of the solvent’s boiling point. In practice, the heating bath temperature was regulated electronically, and the condensation/reflux cycle was automatically adjusted by the system. For aqueous (0%) and ethanol/water (50%) mixtures, the same system ensured that the extraction thimble chamber remained at the specified temperature, thereby allowing reproducible operation across different solvent polarities. The selection of temperature levels (45 °C, 55 °C, and 65 °C) was based on previous literature indicating that phenolic compounds and antioxidants in mushrooms remain stable and bioavailable within this range, while minimizing thermal degradatio4,5. Extraction durations (5, 10, and 15 h) were chosen to capture both early and extended extraction profiles, as bioactive component recovery often reaches equilibrium between 10 and 15 h depending on solvent polarity and matrix complexity. Ethanol concentrations (0%, 50%, 100%) were selected to represent a full spectrum of polarity—from purely aqueous to purely organic conditions—allowing evaluation of hydrophilic and lipophilic compound extraction efficiencie5,10. The solvent-to-solid ratio was kept constant across all experiments to isolate the effects of temperature, time, and solvent composition on extraction performance. This ratio was fixed based on preliminary trials and standard practices in Soxhlet-based optimization studies, and will be evaluated in detail in future multifactorial models including additional variables such as pH or particle size. Temperature settings included 45 °C, 55 °C, and 65 °C; durations were 5, 10, and 15 h; and ethanol concentrations were adjusted to 0%, 50%, and 100%. The experimental results were then analyzed and optimized using Response Surface Methodology (RSM) and an advanced artificial intelligence approach integrating Artificial Neural Networks (ANN) with Genetic Algorithms (GA).
Response surface methodology (RSM)
In this research, the optimization process was carried out using Response Surface Methodology (RSM), with emphasis on three independent parameters: extraction temperature, duration, and ethanol-to-water ratio. The total antioxidant status (TAS) of the extract was selected as the response variable for modeling purposes. The TAS value was selected as the primary response variable because it provides a comprehensive measure of the total antioxidant capacity of the extract, accounting for both known and unknown antioxidant constituents. Unlike individual assays that target specific radicals or compounds, TAS reflects the cumulative antioxidant potential of the entire extract in biologically relevant conditions. This makes it an ideal integrative parameter for optimization purposes in both RSM and ANN-GA modeling frameworks, particularly in studies where the goal is to enhance the overall bioactivity of complex natural matrices such as mushroom extract11,12.
Optimization was performed with the aid of Design Expert version 13, utilizing a second-order polynomial regression model, as presented below:
In the model, Yk represented the response variable (with Yi denoting the TAS value of the extract), while xi corresponded to the coded independent variables—specifically, x1 for extraction temperature, x2 for extraction time, and x3 for the ethanol-to-water ratio. The term βk0 referred to the predicted response at the central point of the experimental design.
The appropriateness of the model was assessed using the coefficient of determination (R2), analysis of variance (ANOVA), and corresponding p-values. To ascertain the ideal reaction circumstances, critical points were identified by computing the derivatives of the polynomial equation. Furthermore, three-dimensional surface plots were created to demonstrate the interactions between the independent variables and their impact on the response variable, facilitating a more profound comprehension of the system’s behavior.
Artificial neural network–genetic algorithm (ANN–GA)
This study involved the development of an Artificial Neural Network (ANN) model to simulate the extraction process, utilizing extraction temperature, duration, and ethanol-to-water ratio as input variables, with total antioxidant status (TAS) as the target output. The dataset was divided into training (80%), validation (10%), and testing (10%) subsets. The training phase utilized the Levenberg–Marquardt (LM) optimization technique. The LM algorithm was chosen because of its efficiency and rapid convergence in training moderately sized feedforward networks, offering a balance between computational speed and accuracy. To determine the best ANN architecture, twenty distinct configurations with differing quantities of hidden neurons (from 1 to 20) were assessed. The hidden layer used the tansig (hyperbolic tangent sigmoid) activation function, while the output layer employed purelin (linear) transfer function, as these combinations are widely applied in regression-oriented biological modeling. The learning rate and momentum were established at 0.5, the maximum number of iterations was capped at 500, the validation check limit was set to 50, and the error tolerance level was defined as 1 × 10⁻5. Each configuration underwent training for 1000 iterations to guarantee robustness.
Model accuracy was quantified using two standard metrics: mean square error (MSE) and mean absolute percentage error (MAPE). These were calculated according to the formulas provided in Eqs. 1 and 2.
where e is the experimental result, p is the prediction result, and n is the number of samples.
During the optimization phase, the Genetic Algorithm (GA) was utilized. Various population sizes were tested to assess their effect on performance, and the selection of individuals was carried out using the roulette wheel approach. A single-point crossover strategy was employed to facilitate genetic recombination. The crossover probability was set to 0.8, and the mutation rate was set to 0.01, which provided stable convergence while avoiding premature stagnation. The optimal number of iterations was determined by analyzing the convergence curves. In order to enhance the likelihood of reaching a global optimum, each optimization experiment was conducted a minimum of 60 times13.
Extraction for bioactivity
Optimal manufacturing parameters were determined to yield extracts with maximal biological activity from mushroom samples. The optimization performed using Response Surface Methodology (RSM) identified the optimal extraction parameters as a temperature of 54.202 °C, an extraction duration of 13.414 h, and an ethanol-to-water ratio of 42.320%. The Artificial Neural Network–Genetic Algorithm (ANN-GA) method produced ideal settings of 57.473 °C, 9.418 h, and a 45.114% ethanol/water ratio. Utilizing these parameters, the extraction procedure was conducted with computational assistance via the Gerhardt SOX-414 system, yielding optimal results. The biological activity of the extracts produced under these improved circumstances was rigorously assessed using comprehensive experimental investigations.
Phenolic analysis
The phenolic composition of the optimized mushroom extracts was thoroughly examined utilizing LC–MS/MS technology. A total of 24 typical phenolic compounds were screened using commercial standards (Sigma-Aldrich), and the quantification was carried out via external calibration curves. The analysis was performed on a Shimadzu Nexera LC–MS/MS system consisting of an LC-30AD binary pump, DGU-20A3R degasser, SIL-30AC autosampler, and CTO-10AS vp column oven. Chromatographic separation was achieved using a reverse-phase C18 Intersil ODS-4 column (3.0 mm × 100 mm, 2 µm), maintained at 40 °C. The mobile phases consisted of water containing 0.1% formic acid (Phase A) and methanol with 0.1% formic acid (Phase B). The flow rate was 0.3 mL/min and the injection volume was 2 μL. A gradient elution program was applied as follows: 0–1 min, 5% B; 1–8 min, linear increase to 90% B; 8–10 min, held at 90% B; 10–12 min, returned to 5% B; and 12–15 min, re-equilibration. Total run time was 15 min. Detection was performed in negative electrospray ionization mode (ESI⁻), and compound identification was based on retention time, molecular ion [M–H]⁻, and MS/MS fragmentation patterns. All data acquisition, peak identification, and quantification procedures were performed using LabSolutions software (Shimadzu, Japan). The list of screened compounds included acetohydroxamic acid, kaempferol, fumaric acid, gallic acid, protocatechuic acid, 4-hydroxybenzoic acid, caffeic acid, naringenin, quercetin, luteolin, catechinhydrate, vanillic acid, syringic acid, thymoquinone, resveratrol, myricetin, salicylic acid, phloridzin dihydrate, 2-hydroxycinnamic acid, oleuropein, 2-hydroxy-1,4-naphthoquinone, silymarin, alizarin, and curcumin. However, only gallic acid, quercetin, vanillic acid, 4-hydroxybenzoic acid, and syringic acid were consistently detected at quantifiable levels across all extract types14. The remaining compounds were either not detected or were below the quantification limit, and thus are not included in Table 2. All phenolic content analyses were performed in triplicate (n = 3), and results are expressed as mean ± standard deviation. Statistical analysis (independent t-test) showed that the differences observed between RSM and ANN-GA extracts were significant at p < 0.05 for compounds marked with different superscript letters in Table 2. This confirms that the reported variations are not only descriptive but also statistically supported.
Antioxidant activity tests
Total antioxidant and oxidant tests
The total antioxidant status of the optimized mushroom extracts was evaluated using the Rel Assay TAS kit, with results reported as mmol Trolox equivalents per liter. The Rel Assay TOS kit was utilized to assess the total oxidant status, with results expressed in μmol hydrogen peroxide equivalents per liter11,12. The oxidative stress index (OSI) was determined by dividing the total oxidant status (TOS) value by the total antioxidant status (TAS) value, with the outcome given as a percentage15.
DPPH free radical scavenging activity test
Stock solutions of the mushroom extracts were formulated at a concentration of 1 mg/mL with DMSO as the solvent. One milliliter of each solution was amalgamated with 160 µl of 0.267 mM DPPH solution, which was produced in four milliliters of 0.004% methanol. The combination was incubated for 30 min at ambient temperature in a dark setting to avert light-induced deterioration. After incubation, absorbance was measured at 517 nm, and antioxidant capacity was quantified as mg Trolox equivalents per gram of extract16.
Ferric reducing antioxidant power assay
A 100 µL stock solution was created from the optimized mushroom extracts and mixed with 2 mL of FRAP reagent. The FRAP reagent consisted of 300 mM acetate buffer (pH 3.6), 40 mM HCl, 20 mM FeCl₃·6H₂O, and 10 mM 2,4,6-tris(2-pyridyl)-S-triazine (TPTZ) in a 10:1:1 ratio. The solution was incubated at 37 °C for 4 min, followed by absorbance measurement at 593 nm. The antioxidant ability was quantified as milligrams of Trolox equivalents per gram of extract16.
Anticholinesterase activity tests
The anticholinesterase activity of the optimized mushroom extracts was assessed using the procedure established by Ellman et al.17. Galantamine functioned as the reference standard in these tests. Solutions at varying concentrations (from 200 to 3.125 μg/mL) were produced. In the assay, 130 μL of 0.1 M phosphate buffer (pH 8) was initially introduced to the wells, succeeded by 10 μL of extract solution and 20 μL of either acetylcholinesterase (AChE) or butyrylcholinesterase (BChE) enzyme solution. The solution was incubated at 25 °C in darkness for 10 min. To commence the enzymatic reaction, 20 μL of DTNB (5,5’-dithiobis-(2-nitrobenzoic acid)) and 20 μL of substrate (acetylcholine iodide or butyrylcholine iodide) were included. Absorbance was recorded at 412 nm, and the percentage of enzyme inhibition along with IC₅₀ values (μg/mL) were determined and documented.
Antiproliferative activity
The antiproliferative effects of the optimized mushroom extracts were assessed using the A549 human lung cancer cell line. Solutions were produced at concentrations of 25, 50, 100, and 200 μg/mL. Upon achieving around 70–80% confluency, the cells were detached using 3 mL of Trypsin–EDTA solution (Sigma-Aldrich, MO, USA) and subsequently transferred to appropriate culture plates. After cell seeding, the plates were incubated for 24 h. Subsequent to this duration, the extract solutions were introduced, and the cells were cultured for an additional 24 h. Upon completion of the treatment, the medium was extracted and substituted with fresh growth medium. Subsequently, a 1 mg/mL MTT solution was added, and the plates were incubated at 37 °C until purple formazan crystals formed. In the concluding stage, dimethyl sulfoxide (DMSO) was introduced to solubilize the formazan, and absorbance was quantified at 570 nm utilizing an Epoch spectrophotometer (BioTek Instruments, Winooski, VT18.
Statistical analyses
All statistical analyses in this study were conducted using the SPSS 21.0 software program for Windows. The independent samples t-test was utilized to compare two groups, whereas one-way ANOVA was employed for comparisons among more than two groups. A post-hoc analysis was conducted utilizing Duncan’s multiple range test at a significance level of α = 0.05 to discern significant differences among groups.
Results and discussions
Optimization of extraction conditions
In this study, the effects of three key factors—extraction temperature (45, 55, 65 °C), extraction time (5, 10, 15 h), and ethanol/water ratio (0%, 50%, 100%) were investigated, and their impacts on total antioxidant status (TAS) values were evaluated. TAS values of the extracts obtained in this study are presented in Table 1.
The experimental findings indicated that the highest total antioxidant status (TAS) value (6.068 ± 0.039 mmol/L) was achieved at 55 °C with a 10-h extraction time and a 50% ethanol-to-water ratio. In contrast, the lowest TAS measurement (3.144 ± 0.008 mmol/L) was observed under conditions of 65 °C, 15 h, and 100% ethanol. These results suggest that 55 °C and 10 h of extraction constitute the most effective parameters for maximizing TAS. Notably, utilizing a 50% ethanol/water mixture substantially enhanced antioxidant capacity, whereas pure ethanol diminished it. These outcomes highlight the critical role of extraction parameters in determining antioxidant activity.
Two distinct optimization methodologies were employed in the study based on the experimental dataset. Within the Response Surface Methodology (RSM) framework, several regression models—including linear, two-factor interaction (2FI), quadratic, and cubic—were assessed. Among these, the quadratic model was chosen due to its superior coefficient of determination (R2) value. R2 reflects the proportion of the total variance in the response variable that is explained by the model’s predictors. A high R2 value implies a good fit to the data. In this study, the selected model yielded an R2 of 0.891, indicating that it reliably describes the interaction among variables and provides robust predictions for optimization. In addition to the coefficient of determination (R2), further statistical indicators confirmed the robustness of the quadratic model. The Adjusted R2 and Predicted R2 values were both high and in close agreement, indicating that the model was not overfitted and retained predictive power. The overall regression model was highly significant according to ANOVA (p < 0.001), and the lack-of-fit test was non-significant (p > 0.05), confirming that the quadratic model adequately described the experimental data.
The second-degree polynomial equation derived from the regression analysis to predict the TAS values of P. ribis is given below:
In the equation \(X_{1}\), \(X_{2}\) ve \(X_{3}\) correspond to the extraction temperature, extraction time, and ethanol/water ratio, respectively.
The response surface plots illustrating the TAS of P. ribis are presented in Fig. 1, showing that TAS increased with temperature up to a mid-level before declining, reached maximum values around 50% ethanol due to polarity balance, and was less influenced by extraction time. The curved surfaces also highlight strong interactions, particularly between temperature and solvent ratio.

Response surface plots of TAS of Phylloporia ribis.
During the optimization phase utilizing artificial intelligence approaches, the experimental dataset was modeled with an Artificial Neural Network (ANN). The optimal ANN design was then enhanced using a Genetic Algorithm (GA). The optimum model architecture was determined to be 3–4–1, consisting of four hidden neurons. This model demonstrated exceptional prediction accuracy, with a mean square error (MSE) of 0.002, a mean absolute percentage error (MAPE) of 0.414%, and a correlation coefficient (R) of 0.999.
The Genetic Algorithm was applied to enhance the predictions of the best-performing ANN model. Following the evaluation of different population sizes, a size of 10 was determined to be the most suitable. A population size of 10 was selected because it provided the optimal balance between high-quality solutions (highest TAS values) and computational efficiency (fast convergence). As shown in Fig. 2, the convergence curve confirms that 20 iterations were adequate for achieving stable results. In terms of predictive accuracy, the ANN-GA model exhibited superior performance, with a higher predictive R2 and lower error values (RMSE and MAE) compared to RSM, confirming its stronger generalization ability. These results indicate that while both approaches identified similar trends, ANN-GA provided more precise predictions and achieved a slightly different but biologically more favorable optimum.

Convergence graph of optimization process.
Phenolic contents
Mushrooms are recognized for their rich content of phenolic compounds, which play a crucial role in their many biological activities. These phenolics serve as essential natural bioactive compounds that account for numerous therapeutic actions found in mushrooms19. This study evaluated the phenolic composition of P. ribis extracts prepared under optimal circumstances. The relevant results are displayed in Table 2.
In our study, phenolic compounds detected in RSM and ANN-GA extracts produced under optimum conditions of P. ribis were determined. The results obtained reveal that the extracts obtained with the ANN-GA method generally provide higher yields in phenolic compound concentrations. This suggests that artificial intelligence-based optimization techniques can play a more effective role in extraction processes compared to traditional statistical methods. Gallic acid was determined as 10,491.72 mg/kg by the ANN-GA method and showed a significant increase compared to the value of 7270.54 mg/kg obtained by the RSM method. This finding shows that due to the hydrophilic nature of gallic acid20,21, the capacity of ANN-GA to determine optimal solvent ratios and temperature parameters provides higher extraction efficiency. The amount of quercetin was also measured as 9960.53 mg/kg by ANN-GA method and showed a significant increase compared to the value of 8542.81 mg/kg obtained by RSM method. Quercetin is a flavonoid with lipophilic character22, and it is thought to be extracted more efficiently thanks to the ability of ANN-GA to optimize the solvent selection and extraction temperature. These results support that ANN-GA can separate compounds with different polarity with higher efficiency. However, it was observed that some phenolic compounds such as 4-hydroxybenzoic acid and vanillic acid were extracted with higher efficiency by ANN-GA method. Vanillic acid was determined as 1144.93 mg/kg by ANN-GA method, while it was measured as 525.87 mg/kg by RSM method. This significant increase shows that ANN-GA can increase the extraction efficiency of phenolic acids by optimizing the solvent concentration and pH level. However, the syringic acid concentration was determined as 2374.52 mg/kg by the RSM method and 2152.67 mg/kg by the ANN-GA method, and a statistically significant decrease (p < 0.05) was observed in the ANN-GA extract compared to the RSM extract. This may be due to the fact that ANN-GA may have created non-ideal conditions for syringic acid at certain optimization parameters or that this compound has a more unstable structure. The bioavailability and biological activities of phenolic compounds vary depending on the extraction techniques and solvents used10. Although the RSM method is effective in determining the optimum extraction conditions based on statistical modeling, artificial intelligence-based optimization techniques such as ANN-GA can increase the extraction efficiency by better modeling the nonlinear relationships between parameters. Studies show that the ANN-GA method can minimize the loss of phenolic compounds by adjusting parameters such as extraction temperature, time, and solvent ratio more precisely23. In this study, ANN-GA provided higher efficiency in the extraction of phenolic compounds, particularly gallic acid, quercetin, and vanillic acid, emphasizing its potential as a powerful tool for biotechnological optimization. These compounds are of high industrial relevance: gallic acid is used in the food and nutraceutical sectors as a natural antioxidant and preservative21, quercetin is applied in supplements and functional foods for its anticancer and anti-inflammatory effects24, and vanillic acid is utilized in cosmetics and skincare products for its free-radical scavenging and anti-aging properties25. However, the fact that the RSM method yielded higher recovery for certain compounds (e.g., syringic acid) indicates that optimization should be evaluated on a compound-specific basis. Therefore, while ANN-GA demonstrates strong potential to enhance the extraction efficiency of phenolic compounds, complementary optimization strategies may be required for individual molecules. Future studies combining ANN-GA with different solvent systems, temperature gradients, and extraction times may further maximize the bioavailability of phenolic compounds and support their industrial applications in food, pharmaceutical, and cosmetic sectors.
Antioxidant activity
Mushrooms possess potent antioxidant capabilities. Compounds including polyphenols, flavonoids, and ergothioneine safeguard the body from the detrimental effects of free radicals by diminishing oxidative stress at the cellular level. These substances may mitigate the risk of inflammation, aging, and certain diseases induced by free radicals in the body26. Moreover, substances like beta-glucans included in mushrooms enhance immune system fortification. Mushrooms are a vital component of a nutritious diet and promote cellular health due to their antioxidant capabilities27,28. The antioxidant capacity of the optimized extract of P. ribis was assessed in our investigation. The results are presented in Table 3.
In this research, the antioxidant potential of P. ribis extracts obtained under optimized conditions was assessed, and the performance of RSM and ANN-GA optimization techniques was comparatively analyzed. All assays were conducted in triplicate (n = 3), expressed as mean ± SD, and statistical testing (p < 0.05) confirmed that the observed differences between RSM and ANN-GA were significant. Overall, extracts optimized through the ANN-GA approach demonstrated superior antioxidant properties. The total antioxidant status (TAS) of the ANN-GA extract was measured at 6.092 mmol/L, which was higher than the 5.997 mmol/L observed in the RSM-optimized sample, indicating a more efficient enhancement of antioxidant compounds using ANN-GA. For total oxidant status (TOS), the ANN-GA method yielded a higher value (13.389 μmol/L) compared to RSM (12.701 μmol/L). The oxidative stress index (OSI) was also higher in the ANN-GA group (0.220). While ANN-GA successfully enhanced the TAS, these results indicate that it also led to a statistically significant increase in TOS and OSI, suggesting a potential rise in oxidative stress. This implies that the optimized conditions, while favorable for extracting antioxidants, may also promote the extraction or formation of oxidant compounds. Therefore, the net biological benefit should be interpreted with caution, considering both the positive antioxidant gains and the undesirable increase in oxidant markers. This dual outcome may be due to the co-extraction of pro-oxidant molecules alongside antioxidants under the same solvent and temperature conditions, a phenomenon also noted in other mushroom extraction studies. Regarding ferric reducing antioxidant power (FRAP), the ANN-GA extract reached 102.033 mg Trolox equivalents/g, outperforming the RSM counterpart, which showed 95.803 mg Trolox equivalents/g. This result indicates a stronger iron-reducing potential and enhanced antioxidant function when using the ANN-GA model. Higher FRAP and DPPH values suggest stronger radical scavenging potential, which translates into practical advantages for food preservation, nutraceutical formulations with enhanced antioxidant efficacy, and cosmetic applications targeting oxidative stress and skin aging29. Similarly, the DPPH radical scavenging activity was significantly higher in the ANN-GA extract (124.320 mg Trolox equivalents/g) compared to the RSM extract (109.140 mg Trolox equivalents/g), further supporting the effectiveness of ANN-GA in improving radical neutralization capacity. Collectively, these findings suggest that the ANN-GA optimization approach is more successful in enhancing the antioxidant performance of P. ribis extracts. Its ability to handle nonlinear interactions and model complex variable relationships likely contributes to its superior outcomes. Thus, ANN-GA stands out as a promising tool for optimizing antioxidant-rich extracts, facilitating their more efficient use in biotechnological applications.
No studies on the antioxidant potential of P. ribis were found in the literature. However, TAS, TOS and OSI values of different wild mushroom species were reported in the literature. From these studies, TAS values of Lactarius deliciosus, Laeticutis cristata, Hericium erinaceus and Cantharellus cibarius were reported as 7.468, 3.623, 5.426 and 5.511 mmol/L. TOS values were reported as 13.161, 27.476, 6.621 and 7.289 μmol/L, respectively. OSI values were reported as 0.176, 0.765, 0.122 and 0.132, respectively30,31,32,33. Compared to these studies, the TAS values of both RSM and ANN-GA optimized extracts of P. ribis used in our study were determined to be lower than L. deliciosus and higher than H. erinaceus, L. cristata and C. cibarius. The TAS value reflects the cumulative presence of antioxidant compounds naturally occurring in biological materials34. Based on this, P. ribis appears to possess higher levels and greater efficacy of phenolic compounds, flavonoids, and other antioxidant constituents compared to some species, while remaining lower than others. These comparative patterns are consistent with our phenolic analysis, where ANN-GA optimization enhanced the recovery of compounds such as gallic acid and quercetin, which likely contributed to the higher FRAP and DPPH activities observed. TOS, on the other hand, represents the overall concentration of oxidant molecules in natural products34. In this study, the TOS value of the P. ribis extract optimized via the RSM method was found to be lower than that of Lactarius deliciosus and L. cristata, but higher than that of Hericium erinaceus and Cantharellus cibarius. Meanwhile, the TOS value of the ANN-GA optimized P. ribis extract was lower than L. cristata, yet higher than L. deliciosus, H. erinaceus, and C. cibarius. The OSI value, which indicates the ratio of oxidants relative to antioxidants, also serves as a useful comparative measure34. In our findings, OSI values of both RSM and ANN-GA optimized P. ribis extracts were lower than those of L. cristata and higher than L. deliciosus, H. erinaceus, and C. cibarius.
These differences could be explained by possible chemical mechanisms. Solvent polarity may influence the extent to which hydrophilic and lipophilic antioxidant compounds are extracted; for instance, ethanol–water mixtures may favor the solubilization of phenolic molecules, whereas purely aqueous or organic solvents might under- or over-extract certain classes of compounds. Extraction temperature may also play a role by facilitating the release of bound antioxidants while simultaneously causing partial degradation of thermally sensitive compounds. Taken together, the observed TAS, TOS, and OSI patterns may reflect a balance between enhanced extraction efficiency and the potential co-extraction or degradation of oxidant molecules. These findings indicate that optimization strategies can substantially influence the antioxidant–oxidant equilibrium in mushroom extracts. The ANN-GA approach proved more effective in enhancing antioxidant potential, as shown by elevated FRAP and DPPH values; however, the accompanying increases in TOS and OSI emphasize the importance of maintaining a balance between antioxidant and oxidant components. Thus, while the primary goal of optimizing P. ribis extracts is to maximize antioxidant capacity, it is equally important to monitor and control oxidant levels. Considering that ANN-GA outperformed RSM in improving biological activity, this approach can be regarded as a promising strategy for the development of natural products with enhanced antioxidant properties.
Anticholinesterase activity
This study provides the first report on the anti-Alzheimer potential of P. ribis, highlighting the novelty of evaluating its cholinesterase inhibitory properties under optimized extraction conditions. Mushrooms are known to be rich in natural compounds with diverse biological effects, including anticholinesterase activity. Anticholinesterase agents inhibit the enzyme acetylcholinesterase, thereby increasing the levels of acetylcholine available for neurotransmission in the brain, a mechanism particularly relevant in Alzheimer’s disease where reduced acetylcholine levels are linked to cognitive decline35,36. Through such bioactive molecules, mushrooms may contribute to maintaining nervous system health and potentially delay neurodegeneration, offering neuroprotective benefits. Thus, they serve not only as nutritional resources but also as valuable therapeutic sources37. In this study, the anticholinesterase activity of optimized P. ribis extracts was evaluated, and the IC₅₀ values are presented in Table 3. As shown in Table 3, the IC₅₀ values for AChE inhibition were 47.20 ± 0.87 µg/mL for the ANN-GA extract and 52.57 ± 1.42 µg/mL for the RSM extract, while BChE inhibition values were 67.05 ± 1.00 µg/mL and 72.58 ± 1.13 µg/mL, respectively. These results demonstrate that ANN-GA optimization produced significantly stronger cholinesterase inhibition.
Currently, there is no published data on the anticholinesterase activity of P. ribis. In this study, the inhibitory effects of optimized P. ribis extracts were assessed, and the activities of extracts produced via RSM and ANN-GA methods against acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) enzymes were compared. The IC₅₀ values obtained indicate that both optimization methods yielded extracts capable of inhibiting these enzymes to a certain extent. Notably, the ANN-GA optimized extract exhibited lower IC₅₀ values for both AChE and BChE, suggesting a more pronounced inhibitory potential via this method. However, when compared with galantamine, a clinically used reference compound, the IC₅₀ values of P. ribis extracts were found to be higher. This implies that while P. ribis does not match the potency of synthetic or semi-synthetic inhibitors, it could still serve as a promising natural source of cholinesterase inhibitors. The ANN-GA approach may have facilitated the enhancement of bioactive components responsible for AChE and BChE inhibition, thereby improving overall biological activity. Enzyme inhibition, particularly of those implicated in disease pathogenesis, plays a crucial role in therapeutic strategies38. Cholinesterases are central to neurotransmission and are targeted by inhibitors in the treatment of neurodegenerative diseases like Alzheimer’s disease39. The superior activity of ANN-GA extracts can be mechanistically explained by their higher phenolic content, particularly quercetin and gallic acid, both of which are reported inhibitors of acetylcholinesterase. Thus, the optimization process not only enhanced overall phenolic recovery but also enriched compounds directly relevant to cholinesterase inhibition, providing a rationale for the stronger anti-Alzheimer potential of ANN-GA extracts. Within this framework, advancing pharmacological and biotechnological optimization techniques to enhance the anticholinesterase capacity of P. ribis may contribute to unlocking its therapeutic potential among natural compound sources.
Antiproliferative activity
Mushrooms represent a valuable natural reservoir of compounds with antiproliferative properties. These effects can suppress or halt the progression of diseases such as cancer by inhibiting abnormal cell proliferation. Bioactive constituents including polysaccharides, triterpenoids, and phenolic compounds found in mushrooms have been shown to impede cancer cell growth and influence cell cycle regulation40. Due to these attributes, mushrooms are increasingly explored as potential anticancer agents. Moreover, their ability to modulate and enhance the immune response may offer supportive benefits both before and during cancer therapy. As such, mushrooms are gaining attention as promising natural candidates for cancer prevention and treatment strategies41,42. In the present study, the antiproliferative effect of the optimized P. ribis extract was evaluated against the A549 human lung cancer cell line. The experimental results are presented in Fig. 3.
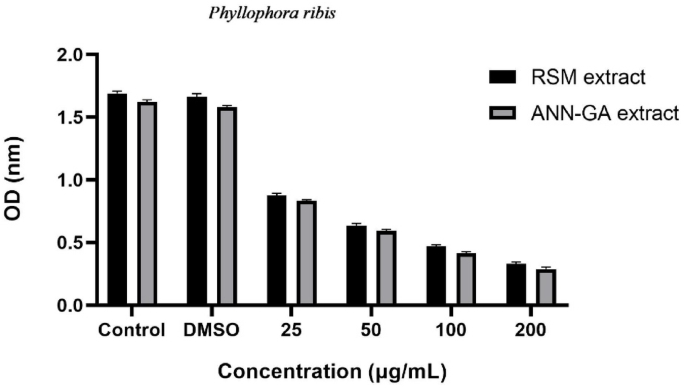
Antiproliferative effects of the optimized Phylloporia ribis extract on the A549 cell line. Note: Control refers to the untreated group maintained only in culture medium. DMSO indicates the group exposed to the solvent (DMSO) without extract. Other groups represent cells treated with the extract at concentrations of 25, 50, 100, and 200 µg/mL.
In our study, the antiproliferative effects of optimized extracts of P. ribis were evaluated and their effects on cellular proliferation were determined at different concentrations (Fig. 3). The control group and DMSO group were used to evaluate the natural growth process and the solvent effect of the cells. It was observed that the extracts obtained with both optimization methods (RSM and ANN-GA) had a concentration-dependent inhibitory effect on cell proliferation. While a more limited decrease in proliferation was observed at low concentrations (25 and 50 µg/mL), a significant decrease in cell viability was detected at concentrations of 100 and 200 µg/mL. It was determined that the extract optimized with the ANN-GA method generally showed a stronger antiproliferative effect compared to that obtained with the RSM method. This suggests that the ANN-GA method increases the biological activity of the extract and provides higher efficacy of the antiproliferative components. The dose-dependent inhibitory effect of the extract indicates that the bioactive compounds it contains can suppress cellular growth and division mechanisms. The results of this study suggest that optimized extracts of P. ribis possess notable antiproliferative potential, with the extract processed via the ANN-GA method exhibiting particularly enhanced activity. Previous research has shown that numerous wild mushroom species display antiproliferative effects, and their actions on various cancer cell lines have been widely explored43,44,45. These studies indicate that bioactive molecules isolated from mushrooms may serve as promising anticancer candidates by suppressing tumor cell proliferation. The enhanced antiproliferative effect of the ANN-GA extract can be mechanistically linked to its higher content of bioactive phenolics such as gallic acid and quercetin, both of which are reported to induce apoptosis and inhibit cancer cell proliferation. Thus, the stronger activity observed is consistent with the phenolic composition profile obtained under ANN-GA optimization. In our investigation, the optimized P. ribis extracts demonstrated substantial antiproliferative activity against the A549 lung cancer cell line. The enhancement of biological activity through extract optimization, especially with the ANN-GA method, highlights the potential of P. ribis as a natural source capable of inhibiting cancer cell growth and contributing to the development of novel therapeutic agents for lung cancer treatment.
Conclusion
This study aimed to optimize the extraction conditions of P. ribis utilizing Response Surface Methodology (RSM) and Artificial Neural Network–Genetic Algorithm (ANN-GA) techniques, then assessing the biological activities of the resultant extracts. The ANN-GA approach exhibited superior efficacy in augmenting phenolic content, antioxidant capacity, and overall biological activity. Extracts generated through this approach demonstrated enhanced efficacy in free radical scavenging and ferric reducing antioxidant capacity relative to those optimized via RSM. Furthermore, the ANN-GA optimized extract demonstrated a more significant antiproliferative impact against the A549 lung cancer cell line. The data suggest that P. ribis is a promising natural source of pharmacologically and biotechnologically significant chemicals. Future research should do comprehensive biological evaluations employing diverse cancer cell lines and models of neurological disorders. Moreover, the utilization of different solvent systems and sophisticated extraction Technologies-such as enzymatic hydrolysis-may augment the bioactivity of the resultant molecules. The integration of artificial intelligence-driven optimization techniques may significantly enhance the therapeutic use of fungal-derived natural compounds and aid in the advancement of next-generation pharmaceuticals.
Data availability
Data are available from the corresponding author on reasonable request.
References
Comlekcioglu, U., Ozkose, E., Akyol, İ & Ekİncİ, M. S. Fatty acid analysis of anaerobic ruminal fungi Neocallimastix, Caecomyces and Orpinomyces. Int. J. Agric. Biol. 12, 635–637 (2010).
Rizzo, G., Goggi, S., Giampieri, F. & Baroni, L. A review of mushrooms in human nutrition and health. Trends Food Sci. Technol. 117, 60–73 (2021).
Sevindik, M., Bal, C., Eraslan, E. C., Uysal, I. & Mohammed, F. S. Medicinal mushrooms: A comprehensive study on their antiviral potential. Prospect. Pharmac. Sci. 21, 42–56 (2023).
Milovanovic, I., Zengin, G., Maksimovic, S. & Tadic, V. Supercritical and ultrasound-assisted extracts from Pleurotus pulmonarius mushroom: Chemical profiles, antioxidative, and enzyme-inhibitory properties. J. Sci. Food Agric. 101, 2284–2293 (2021).
Roselló-Soto, E. et al. Application of non-conventional extraction methods: Toward a sustainable and green production of valuable compounds from mushrooms. Food Eng. Rev. 8, 214–234 (2016).
Umaña, M., Eim, V., Garau, C., Rosselló, C. & Simal, S. Ultrasound-assisted extraction of ergosterol and antioxidant components from mushroom by-products and the attainment of a β-glucan rich residue. Food Chem. 332, 127390 (2020).
Gürgen, A. & Sevindik, M. Application of artificial neural network coupling multiobjective particle swarm optimization algorithm to optimize Pleurotus ostreatus extraction parameters. J. Food Process. Preserv. 46, e16949 (2022).
Zhou, L.-W. & Dai, Y.-C. Phylogeny and taxonomy of Phylloporia (Hymenochaetales): New species and a worldwide key to the genus. Mycologia 104, 211–222 (2012).
Zhou, L.-W. Four new species of Phylloporia (Hymenochaetales, Basidiomycota) from tropical China with a key to Phylloporia species worldwide. Mycologia 107, 1184–1192 (2015).
Alara, O. R., Abdurahman, N. H. & Ukaegbu, C. I. Extraction of phenolic compounds: A review. Curr. Res. Food Sci. 4, 200–214 (2021).
Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 37, 277–285 (2004).
Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 38, 1103–1111 (2005).
Sevindik, M., Bal, C., Krupodorova, T., Gürgen, A. & Eraslan, E. C. Extract optimization and biological activities of Otidea onotica using artificial neural network-genetic algorithm and response surface methodology techniques. BMC Biotechnol. 25, 25 (2025).
Gürgen, A., Sevindik, M., Krupodorova, T., Uysal, I. & Unal, O. Biological activities of Hypericum spectabile extract optimized using artificial neural network combined with genetic algorithm application. BMC Biotechnol. 24, 83 (2024).
Sevindik, M. The novel biological tests on various extracts of Cerioporus varius. Fresenius Environ. Bull. 28, 3713–3717 (2019).
Sevindik, M., Gürgen, A., Krupodorova, T., Uysal, İ & Koçer, O. A hybrid artificial neural network and multi-objective genetic algorithm approach to optimize extraction conditions of Mentha longifolia and biological activities. Sci. Rep. 14, 31403 (2024).
Ellman, G. L., Courtney, K. D., Andres, V. Jr. & Featherstone, R. M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 7, 88–95 (1961).
Bal, C., Sevindik, M., Krupodorova, T. & Eraslan, E. Total carbohydrate and protein contents and some biological activities of edible Macrolepiota mastoidea mushroom. Acta Aliment 54, 337–346 (2025).
Yahia, E. M., Gutiérrez-Orozco, F. & Moreno-Pérez, M. A. Identification of phenolic compounds by liquid chromatography-mass spectrometry in seventeen species of wild mushrooms in Central Mexico and determination of their antioxidant activity and bioactive compounds. Food Chem. 226, 14–22 (2017).
Hu, X. et al. Hydrophilic gallic acid–imprinted polymers over magnetic mesoporous silica microspheres with excellent molecular recognition ability in aqueous fruit juices. Food Chem. 179, 206–212 (2015).
Şabik, A. E., Mohammed, F. S., Sevindik, M., Uysal, I. & Bal, C. Gallic acid: derivatives and biosynthesis, pharmacological and therapeutic effect, biological activity. Bulletin of University of agricultural sciences and veterinary medicine Cluj-Napoca. Food Sci. Technol. 81(1), 18–27 (2024).
Deepika, & Maurya, P. K. Health benefits of quercetin in age-related diseases. Molecules 27, 2498 (2022).
Amalia, A., Suryono, S., Suseno, J. E. & Kurniawati, R. in Journal of Physics: Conference Series. 012020 (IOP Publishing).
Mohammed, F. S., Sevindik, M., Uysal, İ & Sabik, A. E. Quercetin: Derivatives, biosynthesis, biological activity, pharmacological and therapeutic effects. Prospect. Pharmac. Sci. 21, 49–56 (2023).
Sharma, N. et al. An overview of therapeutic effects of vanillic acid. Plant Arch. 20, 3053–3059 (2020).
Podkowa, A., Kryczyk-Poprawa, A., Opoka, W. & Muszyńska, B. Culinary–medicinal mushrooms: A review of organic compounds and bioelements with antioxidant activity. Eur. Food Res. Technol. 247, 513–533 (2021).
Baba, H., Sevindik, M., Dogan, M. & Akgül, H. Antioxidant, antimicrobial activities and heavy metal contents of some Myxomycetes. Fresenius Environ. Bull. 29, 7840–7846 (2020).
Bolesławska, I. et al. Wild mushrooms as a source of bioactive compounds and their antioxidant properties—Preliminary studies. Foods 13, 2612 (2024).
Birch-Machin, M. & Bowman, A. Oxidative stress and ageing. Br. J. Dermatol. 175, 26–29 (2016).
Sevindik, M. & Bal, C. Antioxidant, antimicrobial, and antiproliferative activities of wild mushroom, Laeticutis cristata (Agaricomycetes), from Turkey. Int. J. Med. Mushrooms 23 (2021).
Sevindik, M., Gürgen, A., Khassanov, V. T. & Bal, C. Biological activities of ethanol extracts of Hericium erinaceus obtained as a result of optimization analysis. Foods 13, 1560 (2024).
Gürgen, A., Unal, O. & Sevindik, M. Biological activities of the golden chantarelle mushroom Cantharellus cibarius (Agaricomycetes) extracts obtained as a result of single and multi-objective optimization studies. Int. J. Med. Mushrooms 26, 63–74 (2024).
Gürgen, A. & Sevindik, M. Single and multi-objective optimization of the red pine mushroom Lactarius Deliciosus (Agaricomycetes) extraction conditions using artificial intelligence methods and biological activities of optimized extracts. Int. J. Med. Mushrooms 27, 59–73 (2025).
Eraslan, E. C. et al. Some biological activities and element contents of ethanol extract of wild edible mushroom Morchella esculenta. Sigma J. Eng. Nat. Sci. 39, 24–28 (2021).
Rai, S. N., Mishra, D., Singh, P., Vamanu, E. & Singh, M. Therapeutic applications of mushrooms and their biomolecules along with a glimpse of in silico approach in neurodegenerative diseases. Biomed. Pharmacother. 137, 111377 (2021).
Jiang, X. et al. Mushroom polysaccharides as potential candidates for alleviating neurodegenerative diseases. Nutrients 14, 4833 (2022).
Phan, C.-W., David, P., Naidu, M., Wong, K.-H. & Sabaratnam, V. Therapeutic potential of culinary-medicinal mushrooms for the management of neurodegenerative diseases: Diversity, metabolite, and mechanism. Crit. Rev. Biotechnol. 35, 355–368 (2015).
Verma, G. et al. Pharmacological strategies for enzyme inhibition in disease therapeutics: A comprehensive review. Curr. Enzym. Inhib. 20, 96–108 (2024).
Haake, A., Nguyen, K., Friedman, L., Chakkamparambil, B. & Grossberg, G. T. An update on the utility and safety of cholinesterase inhibitors for the treatment of Alzheimer’s disease. Expert Opin. Drug Saf. 19, 147–157 (2020).
Nowakowski, P. et al. Mushrooms as potential therapeutic agents in the treatment of cancer: Evaluation of anti-glioma effects of Coprinus comatus, Cantharellus cibarius, Lycoperdon perlatum and Lactarius deliciosus extracts. Biomed. Pharmacother. 133, 111090 (2021).
Wasser, S. P. Medicinal mushrooms in human clinical studies. Part I. Anticancer, oncoimmunological, and immunomodulatory activities: A review. Int. J. Med. Mushrooms 19 (2017).
Fonseca, J., Vaz, J. A. & Ricardo, S. The potential of mushroom extracts to improve chemotherapy efficacy in cancer cells: A systematic review. Cells 13, 510 (2024).
Elhusseiny, S. M. et al. Antiviral, cytotoxic, and antioxidant activities of three edible agaricomycetes mushrooms: Pleurotus columbinus, Pleurotus sajor-caju, and Agaricus bisporus. J. Fungi 7, 645 (2021).
Kaur, A. et al. Valorization of polypore mushroom phellinus fastuosus by analyzing antioxidative, antiproliferative and apoptosis induction potential. Waste Biomass Valorization 14, 2659–2672 (2023).
Park, G. S., Shin, J., Hong, S., Gopal, J. & Oh, J.-W. Anticarcinogenic potential of the mushroom polysaccharide lentinan on gastric and colon cancer cells: Antiproliferative, antitumorigenic, Mu-2-related death-inducing gene, MUDENG ramifications. J. Ind. Eng. Chem. 135, 122–130 (2024).
Author information
Authors and Affiliations
Contributions
Conceptualization, A.F.K., A.G., T.K. M.S.; methodology, A.F.K., A.G., T.K. M.S., I.A; validation, A.F.K., A.G., T.K. M.S.; investigation, A.F.K., A.G., T.K. M.S.; resources, A.F.K., A.G., T.K. M.S., I.A; data curation, A.F.K., A.G., T.K. M.S.; writing—original draft preparation, A.F.K., A.G., T.K. M.S., I.A.; writing—review and editing, A.F.K., A.G., T.K. M.S. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Consent for publication
All authors have approved the fnal article and consented to publication.
Informed consent
This study did not involve human participants.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Korkmaz, A.F., Gürgen, A., Krupodorova, T. et al. Artificial intelligence-assisted optimization of extraction enhances the biological activity of Phylloporia ribis. Sci Rep 15, 41206 (2025). https://doi.org/10.1038/s41598-025-25130-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-25130-0
