
Abstract
Functional anorectal pain (FAP) is a chronic condition with unclear pathophysiological mechanisms that is often linked to psychological distress. This resting-state functional magnetic resonance imaging (rs-fMRI) study investigated aberrant interhemispheric connections in 30 FAP patients versus 21 matched healthy controls (HC) via seed-based functional connectivity (FC) and voxel-mirrored homotopic connectivity (VMHC). Compared with HC, FAP patients presented enhanced FC between the left middle frontal gyrus (MFG.L) and regions such as the right MFG (MFG.R) and left superior temporal gyrus (STG.L). VMHC analysis revealed increased patterns in the MFG.L and left superior medial frontal gyrus (SFGmed.L) in FAP patients. Moreover, the strength of FC between the MFG.L and MFG.R was negatively correlated with age, indicating that this heightened connection may diminish with age. These findings indicate that FAP involves aberrant interhemispheric hyperconnectivity, which may play crucial roles in pain perception and emotional processing. The age-dependent decline in FC highlights the eroding of neuroplasticity in aging patients. These neural alterations could serve as diagnostic biomarkers and provide targets for therapeutic interventions. Our work positions FAP within a brain-gut axis dysregulation framework and suggests circuit-specific therapeutics to restore neural homeostasis.
Similar content being viewed by others
Introduction
Functional anorectal pain (FAP) is a chronic pain condition characterized by persistent discomfort in the anorectal region, with no clear organic or structural findings upon standard clinical examination such as anoscopy and palpation1. Despite its prevalence, the pathophysiological mechanisms underlying FAP remain largely unclear. This condition is often associated with significant psychological distress, including symptoms of depression and anxiety, which can further exacerbate the pain experience2,3. Although direct evidence in FAP is evolving, features of personality disorders or maladaptive personality traits may also be present in some patients, potentially influencing clinical presentation and treatment response4,5,6,7. The pain symptoms can be significantly alleviated by paroxetine, as identified in our previous findings3. The complex interplay between physical symptoms and psychological factors suggests that FAP is not solely a result of peripheral abnormalities but also involves central nervous system processes. Recent research has indicated that FAP is closely associated with neurobiological factors, such as alterations in brain structure and function, as well as psychological factors, including mood disorders and cognitive dysfunction, particularly in domains such as executive function, attention, and memory8. Understanding these multifactorial contributors is essential for developing effective treatment strategies and improving the quality of life for patients with FAP.
Advancements in neuroimaging techniques, particularly resting-state functional magnetic resonance imaging (rs-fMRI), have provided valuable insights into the brain alterations associated with chronic pain conditions9,10. Research utilizing fMRI has consistently suggested that regions such as the superior frontal gyrus (SFG), orbitofrontal cortex (OFC), middle frontal gyrus (MFG), and cerebellum display notable changes in chronic pain patients11,12,13. These brain areas play key roles in pain processing, emotion regulation, and cognitive control12,14,15,16,17,18. Previous research has also shown that altered functional connectivity (FC) in these regions is associated with chronic pain conditions, suggesting a potential link between inter-regional functional connections and pain perception10. Kim et al. reported that fibromyalgia patients have altered structural covariance networks involving the dorsolateral prefrontal cortex (DLPFC) and cerebellum, which are associated with heightened pain sensitivity19. In addition, the cerebellum receives nociceptive input and is activated during both acute and chronic pain20. The connectivity between the cerebellum and other brain regions indicates its important role in integrative functions related to pain perception and emotional responses21.
Our previous multimodal MRI investigations of FAP8 utilized structural magnetic resonance imaging (sMRI), diffusion tensor imaging (DTI), and rs-fMRI to investigate brain alterations in FAP patients, revealing complex alterations in gray matter regions such as the superior frontal gyrus (SFG), fusiform gyrus, middle occipital gyrus, and compromised white matter integrity in the superior longitudinal fasciculus, forceps minor, and anterior thalamic radiation, as well as increased neural activity in the right cerebellum and right MFG (MFG.R). These findings indicate that FAP is linked to extensive brain changes, with certain regions possibly being key contributors to symptoms. Given the prominent functional alterations observed in the right hemisphere, we hypothesized that communication and coordination between the hemispheres might be disrupted. Although lateralized activity suggests an imbalance, it does not fully capture the functional integration between homotopic regions. Voxel-mirrored homotopic connectivity (VMHC) measures the correlation between blood-oxygen-level-dependent time series of symmetrical brain regions, thereby reflecting interhemispheric information exchange and integration. This method provides a direct assessment of interhemispheric synchrony, allowing us to distinguish whether the observed lateralization co-occurs with pathological hypersynchrony, which may reflect impaired inhibition, or hyposynchrony, suggestive of functional disconnection. As such, VMHC offers a novel and necessary level of insight that extends beyond what can be inferred from regional activation changes alone22. As a rs-fMRI method for analyzing interhemispheric functional connections, it is vital for studying brain information integration and has been widely used to assess interhemispheric homotopic connectivity in various neuropsychiatric disorders23,24,25 and pain research26,27. This research direction is crucial for understanding whether the observed right-hemisphere-predominant changes reflect a broader pattern of brain function lateralization in FAP patients. By exploring interhemispheric homotopic connectivity, we can determine whether there is an imbalance in communication between brain hemispheres and how it might contribute to the pathophysiology of FAP.
Given the complex interplay between pain perception, emotional distress, and cognitive control in FAP, it is essential to investigate the functional patterns in these regions to better understand the neural mechanisms underlying this condition. This study aims to explore the imbalanced resting-state FC (rs-FC) in FAP patients, focusing on the MFG, cerebellum, and SFG, and to determine how these changes correlate with pain symptoms. Additionally, this research explores interhemispheric homotopic connectivity with the VMHC approach. By elucidating the above issues in FAP, we hope to provide new insights into the pathophysiology of this condition and identify potential targets for therapeutic interventions.
Materials and methods
Participants
The study was conducted in accordance with the principles of the Helsinki Declaration, and approval was obtained from the Research Ethics Committee of the Affiliated Hospital of Jining Medical University. A total of 30 patients diagnosed with FAP and 21 healthy controls (HC) were recruited from the Department of Anorectal Surgery and the Health Management Center of the Affiliated Hospital of Jining Medical University. All participants provided informed consent and underwent comprehensive medical evaluations, including medical history review, physical examination, and blood biochemical analysis. FAP patients, aged between 18 and 65 years, were diagnosed and classified on the basis of the Rome IV criteria for disorders of gut-brain interaction (DGBI) by experienced colorectal specialists. The exclusion criteria were severe physical illnesses that prevented participation, current or past mental/neurological disorders or brain trauma, previous anal or rectal surgeries, distinct secondary anal or rectal pain, severe gastrointestinal conditions, medications affecting gastrointestinal function, a history of substance abuse or dependence, pregnancy or lactation, and MRI contraindications such as heart stents, pacemakers, or metal dentures. Healthy controls were matched to FAP patients in terms of age, sex, and exclusion criteria, with all the subjects being right-handed.
Clinical data collection
All the FAP subjects were asked to complete a clinical data questionnaire before undergoing MRI scanning. The questionnaire collected information on disease progression and used the short-form McGill Pain Questionnaire (SF-MPQ) for pain assessment. The SF-MPQ includes 11 pain - intensity assessments (PRI: S), 4 pain emotion items (PRI: A), a Visual Analogue Scale (VAS), and Present Pain Intensity (PPI) to measure total pain intensity. Moreover, all subjects underwent assessment of depressive and anxiety symptoms and sleep quality by completing the Hamilton Depression Scale (HAMD), Hamilton Anxiety Scale (HAMA), and Pittsburgh Sleep Quality Index (PSQI) under psychiatrist guidance.
MRI data acquisition
MRI data were collected using the Philips Ingenia CX 3.0T MRI scanner. Participants were positioned supine with their heads secured in an advanced position on the examination table. Sponge pads were placed symmetrically on both sides of the head to minimize movement. Before the scan, the participants were instructed to relax their entire body, keep their eyes closed but remain awake, avoid falling asleep, and refrain from engaging in specific mental activities. After the scan, participants were asked whether they had fallen asleep during the process. High-resolution T1-weighted images were obtained using three-dimensional Brain Volume Imaging (3D-TFE) with the following parameters: TR/TE = 6.6 ms/3.0 ms, FOV = 256 mm × 256 mm, slice thickness = 1.0 mm, no gap, voxel size = 1 mm × 1 mm × 1 mm, flip angle = 12°, scan matrix = 256 × 256, pixel bandwidth = 271 Hz, number of slices = 180, and scan time = 4 min and 20 s. Diffusion-weighted imaging (DWI) was performed using a single-shot echo planar imaging (EPI) sequence with the following parameters: TR/TE = 3800 ms/76 ms, FOV = 256 mm × 256 mm, acquisition matrix = 128 × 128, slice thickness = 2 mm, no gap, and a total of 62 slices covering the entire brain. Each participant underwent 64 non-collinear diffusion-weighted scans (b = 1000 s/mm²) and one unweighted (b = 0 s/mm²) scan. The acquisition of DWI data was part of the overarching multimodal imaging protocol for this cohort. The analysis of white matter integrity from this DWI data has been reported previously8. The present study focuses specifically on the analysis of rs-fMRI. Rs-fMRI data were acquired using a gradient-echo planar imaging sequence with the following parameters: TR/TE = 2000 ms/30 ms, FOV = 220 mm × 220 mm, slice thickness/gap = 3.0 mm/1.0 mm, scan matrix = 80 × 80, voxel size = 2.75 mm × 2.75 mm × 3 mm, number of slices = 36, and flip angle = 65°. The scan duration for each rs-fMRI session was 6 min and 12 s, with a total of 180 time points acquired. Additionally, routine MRI scans including T1-weighted imaging (T1WI), T2-weighted imaging (T2WI), and T2-fluid-attenuated inversion recovery (FLAIR) were performed to exclude participants with brain structural abnormalities.
MRI data processing
MRI data were processed and analyzed via the GRETNA toolbox v2.0.028 in Matlab 2018a (MathWorks, Natick, MA, USA). The preprocessing pipeline included discarding the first five time points, followed by slice-timing correction, head motion correction, spatial normalization to a standard space with 3 × 3 × 3 mm3 voxel dimensions using the structural image, spatial smoothing with a 6 mm full-width at half-maximum Gaussian kernel, removal of linear trends, regression of 24 Friston head motion parameters and white matter and cerebrospinal fluid signals, and application of a [0.01, 0.08] Hz temporal bandpass filter. For FC analysis, six predefined ROIs were selected a priori based on regions identified as structurally or functionally altered in our previous study of this cohort8. The seeds included the left SFG (SFG.L) [x = − 18, y = 8, z = 71], right SFG (SFG.R) [x = 18, y = 8, z = 71], left cerebellum [x = − 15, y = − 66, z = − 15], right cerebellum [x = 15, y = − 66, z = − 15], left MFG (MFG.L) [x = − 39, y = 51, z = 21], MFG.R [x = 39, y = 51, z = 21], were selected as the seed regions. Spherical ROIs with an 8 mm diameter were created for each seed region, and Pearson correlation coefficients were calculated between the average signal of each ROI and the signals of all the brain voxels. Fisher Z-transformation was applied to the correlation coefficients for statistical analysis, which was restricted to gray matter regions defined by the Automated Anatomical Labeling (AAL) atlas (116 regions). The results were visualized by using the BrainNet Viewer toolbox29.
VMHC analysis was performed using the right hemisphere mask from the AAL atlas applied to the preprocessed functional data. To ensure exact voxel-wise correspondence, all functional images were spatially normalized to a symmetric MNI template prior to the analysis. This was achieved by creating a study-specific symmetric template from the normalized T1 images of all participants and then registering both structural and functional images to this template. This method calculates the FC between symmetric voxels, reflecting the symmetry of FC between the brain hemispheres. The VMHC map was generated by calculating the Pearson’s correlation coefficient between the residual time series of each voxel and its mirrored counterpart in the opposite hemisphere, with lower VMHC values indicating higher asymmetry in functional activity. The resulting correlation coefficients underwent Fisher z-transformation to enhance normality, yielding the VMHC values for subsequent statistical analysis.
Statistical analysis
Statistical analysis was performed using SPSS version 21.0. Independent samples t-tests were used to compare demographic data, clinical symptoms, and psychological scale scores between FAP patients and HC. The chi-square test was used to compare the distribution of sex assigned at birth between the two groups. All tests were two-tailed, with statistical significance set at P < 0.05.
Statistical analysis was performed using the Statistical Parametric Mapping (SPM) version 12 (Release 7771, released on January 13, 2020). Two-sample t-tests were applied to compare the FC and VMHC differences between FAP patients and HC, with sex assigned at birth and age as covariates of no interest to isolate the group effect. For both the FC and VMHC results, statistical significance was determined using Alphasim correction for multiple comparisons, which provides a well-established and validated balance between statistical sensitivity and Type I error control for fMRI data analysis. The voxel-wise threshold was P < 0.001, and clusters with more than 10 voxels were considered significant.
Once statistically significant differences in imaging indices were observed between FAP patients and HC, post-hoc correlation analyses were conducted to explore the relationships between these indices and clinical variables. These exploratory correlations were performed using Pearson’s correlation without including additional covariates. All clinical correlation analyses were restricted to the FAP patient group, as the relevant clinical data (e.g., pain measures) were not collected from the HC. The threshold for statistical significance was set at P < 0.05.
Results
Clinical data results
No statistically significant differences were found in terms of age and sex distribution between FAP patients and HC. Compared with HC, FAP patients showed significantly higher HAMD, HAMA, and PSQI scores (P < 0.001). The detailed demographic data is presented in Table 1.
Seed-based FC between groups
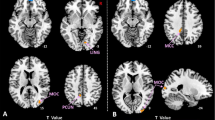
increased FC was observed between the MFG.L and three other brain regions, including FC between the MFG.L and two MFG.R regions, and one FC between the MFG.L and the left superior temporal gyrus (STG.L), with Alphasim-corrected P < 0.001. These findings indicate enhanced functional integration in these regions in FAP. Refer to Table 2; Fig. 1A for details.

Altered functional connectivity (FC) in functional anorectal pain (FAP). (A) Increased functional connectivity in FAP patients versus healthy controls (HC). Significant clusters (Alphasim-corrected P < 0.001) show enhanced connectivity between the left middle frontal gyrus (MFG.L; MNI: − 39, 51, 21) and two regions of the right middle frontal gyrus (MFG.R; MNI: 45, 54, 18, and MNI: 33, 3, 54), and between MFG.L and left superior temporal gyrus (STG.L; MNI: − 54, − 6, − 3). (B) Negative correlation between MFG.L-MFG.R connectivity strength and age in FAP patients (R = − 0.478, P = 0.009).
VMHC differences between groups
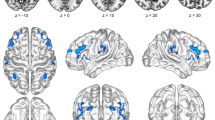
As shown in Table 3; Fig. 2, compared with HC, FAP patients exhibited significantly increased VMHC in the MFG.L and left superior medial frontal gyrus (SFGmed.L) at Alphasim-corrected P < 0.001. No regions showed decreased VMHC in the patients relative to the controls.

Voxel-mirrored homotopic connectivity (VMHC) alterations in functional anorectal pain (FAP). Regions showing increased VMHC in FAP patients versus HC (Alphasim-corrected P < 0.001). The color bar indicates t-values. Sagittal and axial views are shown.
Correlation analysis results
in FAP patients, the enhanced FC between the MFG.L and one cluster of the MFG.R was negatively correlated with age (R = − 0.478, P = 0.009; Fig. 1b). This finding indicates that as patient age increases, the strength of this connection tends to decrease. No other correlations were detected between brain functional alterations and demographic or clinical variables (Supplementary Table S1 and Table S2 in the Supplementary Material.
Discussion
Our study identified increased rs-FC in FAP patients, particularly between the MFG.L and regions such as the MFG.R and STG.L. Additionally, VMHC analysis suggested enhanced connectivity in the MFG.L and SFGmed.L compared with HC. These results suggest a pattern of heightened interhemispheric functional integration in FAP patients. Notably, the negative correlation between age and FC between the left and right MFG indicates that this heightened connection may diminish with age. Overall, these findings indicate alteration in interhemispheric communication in FAP patients, which may contribute to the complex pathophysiology of the syndrome.
The significantly enhanced rs-FC between the MFG.L and both the MFG.R and STG.L in FAP patients reflects a distinct neuroadaptive reorganization within prefrontal-executive and sensory-affective networks. This hyperconnectivity likely reflects compensatory top-down modulation by the DLPFC, which plays a critical role in cognitive control of pain and emotion regulation30,31,32. Specifically, heightened MFG.L-MFG.R connectivity may represent an effort to enhance interhemispheric coordination for inhibitory control over limbic hyperactivity (e.g., the amygdala) and maladaptive nociceptive signaling from the anorectal region33,34. This is consistent with established models of prefrontal engagement in chronic pain conditions, where MFG hyperactivity attempts to restore inhibitory balance in disrupted pain-processing networks35,36,37,38. Concurrently, the increased MFG.L-STG.L connectivity implicates dysregulated sensory-affective integration, as the STG processes visceral sensory input and assigns emotional valence to interoceptive signals39,40. This aligns with evidence from irritable bowel syndrome (IBS) research41,42, where MFG-STG hyperconnectivity correlates with visceral hypersensitivity and symptom severity, suggesting a shared neural substrate for central sensitization across functional gastrointestinal disorders involving aberrant attentional bias toward visceral stimuli43. Collectively, these FC alterations highlight an MFG-centered pathophysiology in FAP. Failed compensatory efforts to control pain via prefrontal networks coexist with amplified sensory-limbic processing, perpetuating the cycle of chronic pain and psychological distress characteristic of this disorder.
The heightened VMHC in the MFG.L and SFGmed.L in FAP patients corresponds with our prior findings of predominantly right-sided functional changes8. The elevated synchrony in the MFG.L/SFGmed.L may represent a futile compensatory attempt to regulate right-sided pain signals through top-down inhibition. Typically, the DLPFC plays a role in pain suppression via corticolimbic circuits. However, excessive interhemispheric coupling in FAP might reduce neural adaptability, contributing to central sensitization and psychological issues such as anxiety and depression. the elevated VMHC observed in the MFG.L and SFGmed.L may stem from impaired transcallosal inhibition44. GABAergic interneurons manage this inhibition, which normally maintains hemispheric functional segregation45. When this mechanism is compromised, it might result in excessive interaction between homotopic prefrontal regions in the two hemispheres46. This disruption may interfere with the normal role of each hemisphere in pain processing. For instance, the right hemisphere is typically dominant in sensory-discriminative pain processing, whereas the left hemisphere is more involved in cognitive-affective appraisal47. The over-synchronization between homotopic prefrontal regions might lead to unhealthy integration of visceral nociception and psychological distress, potentially amplifying maladaptive pain processing48. Importantly, the increased VMHC in the SFGmed.L, a key part of the DMN, hints at disrupted self-referential processing49. This might lead to a maladaptive focus on internal threats such as rectal discomfort, thereby intensifying symptom perception. This aligns with the clinical presentation of FAP, where psychological distress worsens visceral hypersensitivity and pelvic floor dysfunction2,8. The distinct pattern of VMHC increase in FAP highlights its unique neuropathology within the brain-gut axis, which differs from the classic “disconnection” models observed in chronic pain or psychiatric disorders24,50. Future research is needed to determine whether this VMHC pattern can serve as a biomarker for neuromodulation targeting the MFG.L and SFGmed.L to restore inhibitory balance.
The simultaneous increase of both FC and VMHC in FAP patients highlights distinctive frontal hyperactivity coupled with enhanced interhemispheric equilibrium. This pattern suggests a failure in transcallosal inhibition—a GABAergic-mediated process that normally maintains hemispheric segregation during sensory-affective processing51,52. In healthy brains, the right hemisphere predominantly processes sensory-discriminative aspects of pain (e.g., intensity and location), while the left prefrontal cortex regulates cognitive-affective appraisal (e.g., threat evaluation and emotional response)53,54. In FAP, however, the heightened FC between the MFG.L and MFG.R, alongside increased VMHC in the MFG.L and SFGmed.L, indicates pathological hyper-synchronization that disrupts this lateralization. This susceptibility amplifies right-hemisphere nociceptive signaling (e.g., from the anorectal region) while overloading left prefrontal attempts to exert top-down control, ultimately perpetuating pain chronification53,55. Our previous multimodal MRI study revealed decreased gray matter volume in the SFG.R, MFG.L, left middle temporal gyrus (MTG.L) and other brain regions8. These findings provide a critical context for the current functional observations: the left frontal FC/VMHC elevations likely represent a maladaptive compensatory response to right-sided nociceptive dominance. This asymmetric reorganization aligns with the clinical features of FAP, such as peripheral treatment resistance and comorbid psychological distress2,3. Such hemispheric dyscoordination exacerbates central sensitization through disrupted corticolimbic integration56.
The observed negative correlation between MFG.L-MFG.R FC strength and patient age reveal a critical decline in neuroplasticity in FAP, where the compensatory capacity of the brain diminishes with aging. Younger individuals may sustain stronger MFG.L-MFG.R connectivity to gate right-lateralized visceral pain processing, but this compensatory mechanism might decline with advancing age owing to reduced GABAergic neurotransmission and gray matter atrophy in dorsolateral prefrontal circuits57,58,59. Consequently, older FAP patients might exhibit heightened vulnerability to pain chronicity despite comparable psychological comorbidity severity, potentially explaining their poorer response to conventional therapies such as pelvic floor physical training. The absence of correlations between FC alterations and clinical pain/psychological scores further indicates that age is an independent modulator of prefrontal network efficiency, whereas FAP-related brain changes represent trait-like neural signatures rather than symptom-dependent fluctuations. Clinically, our integrated model suggests that cognitive-behavioral therapy (CBT), including mindfulness-based approaches, should be considered a foundational intervention for all FAP patients, given its established efficacy in managing chronic pain and associated psychological distress, and its evidence for inducing beneficial neurofunctional changes60,61,62. Building on this foundation, our findings tentatively suggest that treatment could be further stratified. Younger patients, who may possess greater neuroplastic potential, might derive particular benefit from the adjunctive use of neuromodulation techniques targeting the identified prefrontal circuitry to amplify therapeutic effects63,64,65. For older patients, or those who may not fully respond to first-line approaches, the combination of CBT with pharmacotherapy such as paroxetine might represent a particularly effective multimodal strategy3,66,67. These suggestions are hypothetical and intended to generate future research into personalized treatment algorithms for FAP. Collectively, these findings position age as a pivotal biomarker for precision therapeutics in FAP, suggesting that neural-resilience-adapted strategies might be used to restore disrupted interhemispheric homeostasis. Critically, the lack of correlation between neural alterations and clinical pain measures must be interpreted with caution. These findings may indicate that the observed interhemispheric hyperconnectivity is not a direct neural correlate of momentary pain perception. Instead, it might represent a trait-like feature of the broader FAP syndrome, reflecting a stable maladaptation in brain networks that underpins the complex interplay between chronic pain and affective comorbidity.
Taken together, our findings suggest that FAP involves aberrant interhemispheric hyperconnectivity. The heightened FC between the MFG.L and both MFG.R and STG.L, coupled with increased VMHC in the MFG.L and SFGmed.L, reveals maladaptive neural reorganization within prefrontal regulatory networks. This pattern suggests maladaptive overcompensation efforts to modulate right-lateralized nociceptive signaling through top-down inhibition, while simultaneously amplifying the sensory-affective processing of visceral stimuli. Critically, the age-dependent decline in MFG.L-MFG.R FC underscores eroding neuroplasticity in aging patients, explaining their poorer treatment response. These neural alterations represent trait-like signatures of FAP pathophysiology rather than symptom-dependent fluctuations, supporting their use as diagnostic biomarkers. The neural circuits identified in this study could potentially inform future therapeutic development, although this requires rigorous validation in interventional studies.
several limitations should be noted in this current study. Our findings should be interpreted considering methodological constraints. First, the moderate sample size (30 FAP patients vs. 21 HC) limits the statistical power for subgroup analyses such as age stratification and may affect generalizability. Second, the cross-sectional design precludes causal inferences about whether the observed FC/VMHC alterations drive or result from chronic pain. Therefore, the derivative discussion of potential clinical applications should be interpreted cautiously. Finally, recruitment exclusively from a Chinese population restricts the extrapolation of findings to other ethnic groups; validation in diverse populations is essential to confirm the cultural/biological generalizability of the identified interhemispheric imbalance pattern.
Conclusion
This study establishes altered interhemispheric communication as a core mechanism in FAP, characterized by aberrant hyperconnectivity in the frontal networks. Key findings include heightened FC between the MFG.L and bilateral pain-processing regions, elevated homotopic synchrony in the medial prefrontal areas, and an age-dependent decline in interhemispheric FC. These alterations reflect maladaptive overcompensation to regulate right-lateralized nociception through impaired transcallosal inhibition. Our work positions FAP within a brain-gut axis dysregulation framework and suggests circuit-specific therapeutics to restore neural homeostasis.
Data availability
The datasets analyzed during this study are available from the corresponding author on reasonable request.
References
Rao, S. S. et al. Anorectal disorders. Gastroenterology https://doi.org/10.1053/j.gastro.2016.02.009 (2016).
Dong, L., Sun, T., Tong, P., Guo, J. & Ke, X. Prevalence and correlates of depression and anxiety in patients with functional anorectal pain. J. Pain Res. 16, 225–232. https://doi.org/10.2147/JPR.S400085 (2023).
Fan, M. et al. Psychological status of patients with functional anorectal pain and treatment efficacy of paroxetine in alleviating the symptoms: A retrospective study. Sci. Rep. 13, 18007. https://doi.org/10.1038/s41598-023-45401-y (2023).
Tanum, L. & Malt, U. F. Personality and physical symptoms in nonpsychiatric patients with functional gastrointestinal disorder. J. Psychosom. Res. 50, 139–146. https://doi.org/10.1016/s0022-3999(00)00219-1 (2001).
McKernan, L. C. et al. Posttraumatic stress disorder in interstitial cystitis/bladder pain syndrome: Relationship to patient phenotype and clinical practice implications. Neurourol. Urodyn. 38, 353–362. https://doi.org/10.1002/nau.23861 (2018).
Liu, R.-L., Song, Q.-C., Liu, L.-M., Yang, Y.-F. & Zhu, W.-H. Mood instability and risk of gastrointestinal diseases—A univariable and multivariable Mendelian randomization study. Ann. Gen. Psychiatry 23, 50. https://doi.org/10.1186/s12991-024-00537-7 (2024).
Ulgen, A., Bourtzoni, S. & Reed, P. Pain perception in personality disorders. PLoS ONE 20, e0323004. https://doi.org/10.1371/journal.pone.0323004 (2025).
Wan, Y. et al. Exploring psychological factors and brain alterations in functional anorectal pain patients: Insights from multimodal magnetic resonance imaging investigations. Neurogastroenterol. Motil. https://doi.org/10.1111/nmo.15017 (2025).
Barkhof, F., Haller, S. & Rombouts, S. A. R. B. Resting-state functional MR imaging: A new window to the brain. Radiology 272, 28–48. https://doi.org/10.1148/radiol.14132388 (2014).
Wang, X. et al. Structural and functional changes of insula subregions in migraine without aura and their relationships with pain perception. Sci. Rep. 15, 13342. https://doi.org/10.1038/s41598-025-98193-8 (2025).
Seminowicz, D. A. & Moayedi, M. The dorsolateral prefrontal cortex in acute and chronic pain. J. Pain 18, 1027–1035. https://doi.org/10.1016/j.jpain.2017.03.008 (2017).
Zeng, X., Sun, Y., Zhiying, Z., Hua, L. & Yuan, Z. Chronic pain-induced functional and structural alterations in the brain: A multi-modal meta-analysis. J. Pain https://doi.org/10.1016/j.jpain.2024.104740 (2024).
Prati, J. M. & Gianlorenço, A. C. A new vision of the role of the cerebellum in pain processing. J. Neural Transm. https://doi.org/10.1007/s00702-024-02872-1 (2025).
Agoalikum, E., Wu, H., Klugah-Brown, B. & Maes, M. Brain structural differences between fibromyalgia patients and healthy control subjects: a source-based morphometric study. Sci. Rep. 15, 17446. https://doi.org/10.1038/s41598-025-01070-7 (2025).
Del Mauro, G. et al. Chronic pain is associated with greater brain entropy in the prefrontal cortex. J. Pain 32, 105421. https://doi.org/10.1016/j.jpain.2025.105421r (2025).
Patel, M., Hasoon, J., Diez Tafur, R., Lo Bianco, G. & Abd-Elsayed, A. The impact of chronic pain on cognitive function. Brain Sci. 15, 559. https://doi.org/10.3390/brainsci15060559 (2025).
Zhao, Y. et al. The role of prefrontal cortex in chronic low back pain with comorbid depression: A resting-state fMRI study. Brain Res. Bull. 229, 111408. https://doi.org/10.1016/j.brainresbull.2025.111408 (2025).
Fiúza-Fernandes, J. et al. Common neural correlates of chronic pain—A systematic review and meta-analysis of resting-state fMRI studies. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 138, 111326. https://doi.org/10.1016/j.pnpbp.2025.111326 (2025).
Kim, H. et al. Fibromyalgia is characterized by altered frontal and cerebellar structural covariance brain networks. NeuroImage Clin. 7, 667–677. https://doi.org/10.1016/j.nicl.2015.02.022 (2015).
Han, X. et al. Neural activation during tonic pain and interaction between pain and emotion in bipolar disorder: An fMRI study. Front. Psychiatry 9, 555. https://doi.org/10.3389/fpsyt.2018.00555 (2018).
Moulton, E. A., Schmahmann, J. D., Becerra, L. & Borsook, D. The cerebellum and pain: Passive integrator or active participator?. Brain Res. Rev. 65, 14–27. https://doi.org/10.1016/j.brainresrev.2010.05.005 (2010).
Zuo, X. N. et al. Growing together and growing apart: Regional and sex differences in the lifespan developmental trajectories of functional homotopy. J. Neurosci. 30, 15034–15043. https://doi.org/10.1523/JNEUROSCI.2612-10.2010 (2010).
Deng, K. et al. Reduced interhemispheric functional connectivity in obsessive-compulsive disorder patients. Front. Psych. 10, 418. https://doi.org/10.3389/fpsyt.2019.00418 (2019).
Deng, K. et al. Impaired robust interhemispheric function integration of depressive brain from REST-meta-MDD database in China. Bipolar Disord. 24, 400–411. https://doi.org/10.1111/bdi.13139 (2022).
Liu, Y. et al. Exploring brain asymmetry in early-stage Parkinson’s disease through functional and structural MRI. CNS Neurosci. Ther. 30, e14874. https://doi.org/10.1111/cns.14874 (2024).
Yang, Z. et al. Hippocampal functional radiomic features for identification of the cognitively impaired patients from low-back-related pain: A prospective machine learning study. J. Pain Res. 18, 271–282. https://doi.org/10.2147/jpr.S484680 (2025).
Cao, Z.-M. et al. Abnormalities of thalamic functional connectivity in patients with migraine: A resting-state fMRI study. Pain Ther. 11, 561–574. https://doi.org/10.1007/s40122-022-00365-1 (2022).
Wang, J. et al. GRETNA: A graph theoretical network analysis toolbox for imaging connectomics. Front. Hum. Neurosci. 9, 386. https://doi.org/10.3389/fnhum.2015.00386 (2015).
Xia, M., Wang, J. & He, Y. BrainNet Viewer: A network visualization tool for human brain connectomics. PLoS ONE 8, e68910. https://doi.org/10.1371/journal.pone.0068910 (2013).
Gianola, M., Llabre, M. M. & Losin, E. A. Does pain hurt more in Spanish? The neurobiology of pain among Spanish-English bilingual adults. Soc. Cogn. Affect. Neurosci. 19, nsad074. https://doi.org/10.1093/scan/nsad074 (2023).
Sun, J. et al. A Study of differential resting-state brain functional activity in males and females with recurrent depressive disorder. Brain Sci. 12, 1508. https://doi.org/10.3390/brainsci12111508 (2022).
Amaoui, S. et al. Intrinsic neural network dynamics underlying the ability to down-regulate emotions in male perpetrators of intimate partner violence against women. Brain Struct. Funct. 228, 2025–2040. https://doi.org/10.1007/s00429-023-02696-x (2023).
Neugebauer, V. et al. Amygdala, neuropeptides, and chronic pain-related affective behaviors. Neuropharmacology 170, 108052. https://doi.org/10.1016/j.neuropharm.2020.108052 (2020).
Kiritoshi, T. et al. Cells and circuits for amygdala neuroplasticity in the transition to chronic pain. Cell Rep. https://doi.org/10.1016/j.celrep.2024.114669 (2024).
Ong, W.-Y., Stohler, C. S. & Herr, D. R. Role of the prefrontal cortex in pain processing. Mol. Neurobiol. 56, 1137–1166. https://doi.org/10.1007/s12035-018-1130-9 (2018).
White, L. K., Makhoul, W., Teferi, M., Sheline, Y. I. & Balderston, N. L. The role of dlPFC laterality in the expression and regulation of anxiety. Neuropharmacology 224, 109355. https://doi.org/10.1016/j.neuropharm.2022.109355 (2023).
De Groote, S. et al. Functional magnetic resonance imaging: cerebral function alterations in subthreshold and suprathreshold spinal cord stimulation. J. Pain Res. 11, 2517–2526. https://doi.org/10.2147/jpr.S160890 (2018).
Kang, D. et al. Disruption to normal excitatory and inhibitory function within the medial prefrontal cortex in people with chronic pain. Eur. J. Pain 25, 2242–2256. https://doi.org/10.1002/ejp.1838 (2021).
Sandoval, G. M. et al. Neuroanatomical abnormalities in fragile X syndrome during the adolescent and young adult years. J. Psychiatr. Res. 107, 138–144. https://doi.org/10.1016/j.jpsychires.2018.10.014 (2018).
Jin, Z. et al. Increased resting-state interhemispheric functional connectivity of posterior superior temporal gyrus and posterior cingulate cortex in congenital amusia. Front. Neurosci. 15, 653325. https://doi.org/10.3389/fnins.2021.653325 (2021).
Su, C. et al. Abnormal resting-state local spontaneous functional activity in irritable bowel syndrome patients: A meta-analysis. J. Affect. Disord. 302, 177–184. https://doi.org/10.1016/j.jad.2022.01.075 (2022).
Shuai, Y. et al. Aberrant intrinsic brain activities in functional gastrointestinal disorders revealed by seed-based d mapping with permutation of subject images. Front. Neurosci. 18, 1452216. https://doi.org/10.3389/fnins.2024.1452216 (2024).
Barbara, G. et al. Mechanisms underlying visceral hypersensitivity in irritable bowel syndrome. Curr. Gastroenterol. Rep. 13, 308–315. https://doi.org/10.1007/s11894-011-0195-7 (2011).
Chen, C., Lidstone, D., Crocetti, D., Mostofsky, S. H. & Nebel, M. B. Increased interhemispheric somatomotor functional connectivity and mirror overflow in ADHD. NeuroImage Clin. 31, 102759. https://doi.org/10.1016/j.nicl.2021.102759 (2021).
Schuster, J. et al. ZEB2 haploinsufficient Mowat-Wilson syndrome induced pluripotent stem cells show disrupted GABAergic transcriptional regulation and function. Front. Mol. Neurosci. 15, 988993. https://doi.org/10.3389/fnmol.2022.988993 (2022).
Tan, Y., Zhou, C. & He, L. Altered structural and functional abnormalities of hippocampus in classical trigeminal neuralgia: A combination of DTI and fMRI study. J. Healthc. Eng. 2022, 8538700. https://doi.org/10.1155/2022/8538700 (2022).
Ryu, S., Gwon, D., Park, C., Ha, Y. & Ahn, M. Resting-state frontal electroencephalography (EEG) biomarkers for detecting the severity of chronic neuropathic pain. Sci. Rep. 14, 20188. https://doi.org/10.1038/s41598-024-71219-3 (2024).
Lan, L. et al. Acupuncture modulates the spontaneous activity and functional connectivity of calcarine in patients with chronic stable angina pectoris. Front. Mol. Neurosci. 15, 842674. https://doi.org/10.3389/fnmol.2022.842674 (2022).
Zhang, H. et al. Topographical information-based high-order functional connectivity and its application in abnormality detection for mild cognitive impairment. J. Alzheimer’s Disease JAD 54, 1095–1112. https://doi.org/10.3233/jad-160092 (2016).
Jiang, H. et al. Aberrant resting-state voxel-mirrored homotopic connectivity in major depressive disorder with and without anxiety. J. Affect. Disord. 368, 191–199. https://doi.org/10.1016/j.jad.2024.08.099 (2025).
Keitel, A., Øfsteng, H., Krause, V. & Pollok, B. Anodal transcranial direct current stimulation (tDCS) over the right primary motor cortex (M1) impairs implicit motor sequence learning of the ipsilateral hand. Front. Hum. Neurosci. 12, 289. https://doi.org/10.3389/fnhum.2018.00289 (2018).
Vlcek, P. & Bob, P. Schizophrenia, bipolar disorder and pre-attentional inhibitory deficits. Neuropsychiatric Disease Treat. 18, 821. https://doi.org/10.2147/ndt.S352157 (2022).
Cho, J. & Huh, Y. Astrocytic calcium dynamics along the pain pathway. Front. Cell. Neurosci. 14, 594216. https://doi.org/10.3389/fncel.2020.594216 (2020).
Robayo, L. E. et al. Neurometabolite alterations in traumatic brain injury and associations with chronic pain. Front. Neurosci. 17, 1125128. https://doi.org/10.3389/fnins.2023.1125128 (2023).
Qu, Y. et al. Acupuncture-related therapy for knee osteoarthritis: A narrative review of neuroimaging studies. J Pain Res 17, 773–784. https://doi.org/10.2147/jpr.S450515 (2024).
Werthman, E. H., Colloca, L. & Oswald, L. M. Adverse childhood experiences and burn pain: a review of biopsychosocial mechanisms that may influence healing. Pain Rep. 7, e1013. https://doi.org/10.1097/pr9.0000000000001013 (2022).
Stockmeier, C. A. & Rajkowska, G. Cellular abnormalities in depression: evidence from postmortem brain tissue. Dialogues Clin. Neurosci. 6, 185–197. https://doi.org/10.31887/DCNS.2004.6.2/cstockmeier (2004).
Heyes, S. et al. Genetic disruption of voltage-gated calcium channels in psychiatric and neurological disorders. Prog. Neurobiol. 134, 36–54. https://doi.org/10.1016/j.pneurobio.2015.09.002 (2015).
Medina, A. M. et al. Neurotransmission-related gene expression in the frontal pole is altered in subjects with bipolar disorder and schizophrenia. Transl. Psychiatry 13, 118. https://doi.org/10.1038/s41398-023-02418-1 (2023).
Ehde, D. M., Dillworth, T. M. & Turner, J. A. Cognitive-behavioral therapy for individuals with chronic pain: Efficacy, innovations, and directions for research. Am. Psychol. 69, 153–166. https://doi.org/10.1037/a0035747 (2014).
Tang, N. K. Y. Cognitive behavioural therapy in pain and psychological disorders: Towards a hybrid future. Prog. Neuropsychopharmacol. Biol. Psychiatry 87, 281–289. https://doi.org/10.1016/j.pnpbp.2017.02.023 (2018).
Sanabria-Mazo, J. P. et al. A systematic review of cognitive behavioral therapy-based interventions for comorbid chronic pain and clinically relevant psychological distress. Front. Psychol. 14, 1200685. https://doi.org/10.3389/fpsyg.2023.1200685 (2023).
Regenold, W. T., Deng, Z.-D. & Lisanby, S. H. Noninvasive neuromodulation of the prefrontal cortex in mental health disorders. Neuropsychopharmacology 47, 361–372. https://doi.org/10.1038/s41386-021-01094-3 (2021).
Gazerani, P. The neuroplastic brain: current breakthroughs and emerging frontiers. Brain Res. 1858, 149643. https://doi.org/10.1016/j.brainres.2025.149643 (2025).
Ferreira, M. et al. Evidence for neuroplasticity in the human brain in health and disease: A systematic review focusing on molecular imaging using PET. Neural Plast. https://doi.org/10.1155/np/9423232 (2025).
Kolasa, M. et al. Paroxetine and low-dose risperidone induce serotonin 5-HT(1A) and dopamine D2 receptor heteromerization in the mouse prefrontal cortex. Neuroscience 377, 184–196. https://doi.org/10.1016/j.neuroscience.2018.03.004 (2018).
Nakayama, K. Effect of paroxetine on extracellular serotonin and dopamine levels in the prefrontal cortex. Naunyn Schmiedebergs Arch. Pharmacol. 365, 102–105. https://doi.org/10.1007/s00210-001-0497-7 (2002).
Funding
This work was supported by the Graduate Education and Teaching Reform Research Project of Shandong Province (SDYJSJGC2024066), the Medical and Health Science and Technology Development Plan of Shandong Province (202304011343), the Key Research and Development Plan of Jining City (2021YXNS024, 2021YXNS127), and the Cultivation Plan of High-level Scientific Research Projects of Jining Medical University (JYGC2021KJ006).
Author information
Authors and Affiliations
Contributions
X.X. conducted the statistical analysis and wrote the entire manuscript. W.P, S.G, S.W, and M.F. performed participant recruitment and collected demographic data. H.Y and C.S. acquired the MRI data and conducted preprocessing. Y.W. and C.Z. collected the clinical assessment and processed the MRI datasets. Y.W. implemented the MRI quality control. C.Z. and Y.J. designed the study framework and revised the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xu, X., Pei, W., Gao, S. et al. Enhanced interhemispheric functional connectivity in patients with functional anorectal pain. Sci Rep 15, 42489 (2025). https://doi.org/10.1038/s41598-025-26490-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-26490-3
