
Abstract
P2X7 receptor (P2X7R) augments lipopolysaccharide (LPS)-toll-like receptor 4 (TLR4)-mediated neuroinflammation. These roles of P2X7R in neuroinflammation are relevant to nitrosative stress through nuclear factor-κB (NF-κB)-inducible nitric oxide synthase (iNOS) pathway, while the underlying mechanisms are largely unknown. In the present study, we investigated whether protein disulfide isomerase (PDI) is involved in the integration of TLR4-P2X7R functions in response to LPS in vivo. The present study showed that LPS elicited NF-κB-mediated PDI upregulation, iNOS induction and S-nitrosylated PDI (SNO-PDI) level, independent of S-nitrosylation of NF-κB p65 subunit, in P2X7R+/+ mice more than P2X7R−/− mice. SN50 (an NF-κB inhibitor) effectively diminished LPS-induced PDI upregulation in both P2X7R+/+ and P2X7R−/− mice. PDI knockdown attenuated LPS-induced p65 S276 phosphorylation and iNOS induction in both strains. Of interest, S-nitroso-N-acetyl-DL-penicillamine (SNAP, a NO donor) increased SNO-PDI level, surface P2X7R expression and p65 S276 phosphorylation in P2X7R+/+ mice under physiological condition. In P2X7R−/− mice, SNAP was less effective on NF-κB S276 phosphorylation, although SNO-PDI level was similar to that in P2X7R+/+ mice. Taken together, the present data demonstrate that PDI may be an intermediator to integrate TLR4- and P2X7R-mediated signaling pathways in a positive feedback loop, which would exert NF-κB-iNOS-mediated nitrosative stress during LPS-induced neuroinflammation.
Similar content being viewed by others
Introduction
In various neurological diseases, neuroinflammation aggravates the brain lesion by generating reactive oxygen species, proteolytic enzymes and proinflammatory cytokines. Therefore, the inhibition of neuroinflammation is one of the therapeutic interventions for promoting neuroprotection and avoidance of brain injury. P2X7 receptor (P2X7R), an ATP-gated nonselective cationic channel, has attracted much attention as the modulator of inflammatory pathways in the brain by regulating proinflammatory signaling molecules including inducible nitric oxide synthase (iNOS) and nuclear factor-κB (NF-κB)1,2,3,4. P2X7R augments lipopolysaccharide (LPS)-toll-like receptor 4 (TLR4)-mediated neuroinflammation4,5. These roles of P2X7R in neuroinflammation are relevant to iNOS-induced nitrosative stress in a positive feedback manner4,5, while intermediators have been elusive.
Protein disulfide isomerase (PDI) plays an important role as a chaperone in endoplasmic reticulum (ER), which catalyzes formation, reduction and isomerization of disulfide bonds in proteins. PDI has an oxidoreductase activity regulating redox states of cell surface receptors, and also acts as a donor of nitric oxide (NO) and a denitrosylase modulating S-nitrosylation of cytosolic proteins (an oxidative modification of a thiol (-SH) converting to a nitrosothiol (-SNO) between NO and a redox-reactive cysteine)6,7,8,9,10. Interestingly, PDI regulates a P2X7R-dependent activation of prothrombotic tissue factor (TF) that is a coagulation cofactor/receptor expressed in the vessel wall11,12. Furthermore, PDI exerts the trafficking of P2X7R by modulating dynamic redox status and S-nitrosylation of cysteine residues on an extracellular domain forming disulfide bonds, which reinforces microglial activation induced by seizures13. PDI also elicits NF-κB p65-serine (S) 276 phosphorylation that enhances its transactivation potentials and microglial activation13,14. Therefore, it is likely that PDI would function as a cofactor for many transcription factors affected by P2X7R, suggesting the cooperation between P2X7R and PDI in the pathogenesis of NF-κB-mediated neuroinflammation. However, there are controversial in the literatures concerning this: (1) LPS lowers PDI level caused by oxidative stress-induced degradation15, while it enhances PDI induction16,17. (2) PDI suppresses LPS-induced NF-κB transactivation without affecting its DNA-binding activity and phosphorylation18,19, whereas it elicits expressions of proinflammatory factors by increasing NF-κB phosphorylation20,21,22. (3) Reduction of a disulfide bond on NF-κB by thioredoxin (TRX) increases its DNA binding activity23,24, which is suppressed by PDI in a dose-dependent manner18. With respect to these previous reports, notable questions have been raised (1) whether PDI is a mediator for P2X7R-NF-κB-iNOS signal pathway generating LPS-induced nitrosative stress, (2) the S-nitrosylation of PDI distinctly modulates NF-κB transcription activity in response to LPS, which would contribute to the discrepancies in previous studies, and (3) P2X7R ablation changes the action of thiol-modifying oxidoreductase activity of PDI under LPS-induced inflammatory condition. Considering the roles of P2X7R and nitrosative stress in neuroinflammation1,2,3,4,5, it is noteworthy elucidating these questions to understand the novel mechanism of neuroinflammation, which would provide a strategy for inflammation-related neurological diseases.
Here, we demonstrate that LPS elicited NF-κB-mediated PDI upregulation, iNOS induction and S-nitrosylated PDI (SNO-PDI) level, independent of S-nitrosylation of NF-κB p65 subunit, in P2X7R+/+ mice more than P2X7R−/− mice. SN50 (an NF-κB inhibitor) effectively diminished LPS-induced PDI upregulation in both P2X7R+/+ and P2X7R−/− mice. PDI knockdown attenuated LPS-induced p65 S276 phosphorylation and iNOS induction in both strains. Furthermore, S-nitroso-N-acetyl-DL-penicillamine (SNAP, a NO donor) increased SNO-PDI level, surface P2X7R expression and p65 S276 phosphorylation in P2X7R+/+ mice under physiological condition. In P2X7R−/− mice, SNAP was less effective on NF-κB S276 phosphorylation, although SNO-PDI level was similar to that in P2X7R+/+ mice. These findings indicate that PDI may play an important role in a positive feedback loop of TLR4 and P2X7R interactions to regulate NF-κB activation following LPS treatment.
Results
P2X7R ablation inhibits PDI upregulation following LPS treatment
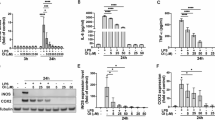
First, we explored whether P2X7R deletion affects PDI expression in the mouse hippocampus following LPS treatment. P2X7R deletion did not affect total PDI protein level in the hippocampus under physiological condition (Fig. 1A). LPS increased total PDI protein level in the hippocampus of P2X7R+/+ mice (χ2(3) = 22.163, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 1A and Supplementary Fig. 1), accompanied by microglial activation (Fig. 1B). P2X7R deletion attenuated LPS-induced microglial activation (Fig. 1B) and PDI upregulation in the hippocampus (p = 0.027, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 1A).

Effects of P2X7R deletion on PDI expression and reactive gliosis in response to LPS. P2X7R deletion does not alter PDI expression level in the hippocampus under physiological condition. P2X7R ablation attenuates PDI upregulation and microglial (not astroglial) activation following LPS treatment. (A) Effects of P2X7R deletion on PDI expression in response to LPS. Representative Western blot images for PDI (left panels) and quantification of the effects of P2X7R deletion on PDI density (right panel) following LPS treatment (#,*p < 0.05 vs. control and P2X7R+/+mice, respectively; Kruskal–Wallis test, n = 7, respectively). (B) Representative photos for Iba-1 positive microglia in the hippocampus following LPS treatment. (C, D) Representative immunofluorescent images for PDI expression in microglia (C) and astrocytes. (E) Quantification of the effects of P2X7R deletion on IB4 (a microglial marker), GFAP (an astroglial marker) and PDI intensities under post-LPS condition (*p < 0.05 vs. P2X7R+/+mice, respectively; Mann–Whitney test, n = 7, respectively).
Immunofluorescent study reveals that LPS increased PDI levels in microglia and astrocytes in P2X7R+/+ mice more than P2X7R−/− mice (z = 2.875, p = 0.004, n = 7, respectively, Mann–Whitney test; Fig. 1C–E). Consistent with a previous study4, P2X7R ablation inhibited LPS-induced microglial activation (z = 3.006, p = 0.003, n = 7, respectively, Mann–Whitney test), but not reactive astrogliosis (z = 0.321, p = 0.748, n = 7, respectively, Mann–Whitney test; Fig. 1C–E). These findings indicate that P2X7R may reinforce PDI upregulation in response to LPS, concomitant with microglial activation.
NF-κB activation elicits PDI upregulation in response to LPS in P2X7R +/+ and P2X7R −/− mice
NF-κB is known as one of key regulators during LPS-induced inflammatory process. In particular, p65 S276 phosphorylation is required for microglial activation under neuroinflammatory condition25,26,27, which is regulated by P2X7R28. Therefore, we investigated the changed p65 S276 phosphorylation induced by LPS. P2X7R deletion did not alter NF-κB expression and p65 S276 phosphorylation under basal condition (Fig. 2A and Supplementary Fig. 2). LPS increased p65 expression level to 1.56-fold of basal level in P2X7R+/+ mice. P2X7R deletion attenuated LPS-induced p65 upregulation (χ2(3) = 22.4, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 2A). p65 S276 phosphorylation was also enhanced in P2X7R+/+ mice more than P2X7R−/− mice following LPS treatment (χ2(3) = 20.555, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 2A). Immunofluorescent study also revealed that P2X7R deletion ameliorated LPS-induced p65 S276 phosphorylation (z = 2.686, p = 0.007, n = 7, respectively, Mann–Whitney test; Fig. 2B, C). To investigate further the role of NF-κB activation in LPS-induced PDI upregulation, we also applied SN50 (an NF-κB inhibitor). SN50 effectively diminished LPS-induced PDI upregulation in both strains (χ2(3) = 17.469, p = 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 2D and Supplementary Fig. 2). These findings indicate that NF-κB may be one of the upstream regulators for PDI upregulation in response to LPS.

Effects of P2X7R-mediated NF-κB activation on PDI upregulation in response to LPS. P2X7R deletion cannot affect p65 expression and its S276 phosphorylation under basal condition. LPS augments p65 expression and p65 S276 phosphorylation in P2X7R+/+ mice. P2X7R deletion attenuates these events. SN50 (a NF-κB inhibitor) ameliorates LPS-induced PDI upregulation in P2X7R+/+ and P2X7R−/− mice. (A) Effects of P2X7R deletion on NF-κB activation in response to LPS. Representative Western blot images for p65 expression and p65 S276 phosphorylation (left panel) and quantification of the effects of P2X7R deletion on p65 and p65 S276 densities (right panel) following LPS treatment (#,*p < 0.05 vs. control and P2X7R+/+mice, respectively; Kruskal–Wallis test, n = 7, respectively). (B) Representative photos for p65 S276 phosphorylation in microglia under post-LPS condition. (C) Quantification of the effects of P2X7R deletion on p65 S276 level in microglia under post-LPS condition (*p < 0.05 vs. P2X7R+/+mice, respectively; Mann–Whitney test, n = 7, respectively). (D) Effects of SN50 on PDI expression in response to LPS. Representative Western blot images for PDI expression (left panel) and quantification of the effects of SN50 on PDI density (right panel) under post-LPS condition (#,*p < 0.05 vs. vehicle and P2X7R+/+mice, respectively; Kruskal–Wallis test, n = 7, respectively).
PDI knockdown attenuates LPS-induced p65 S276 phosphorylation and iNOS induction in P2X7R +/+ mice more than P2X7R −/− mice
In RAW 264.7 cells (a monocyte/macrophage cell line derived from mice), PDI overexpression suppresses the NF-κB transcriptional activity induced by LPS treatment18. In contrast, PDI deletion also inhibits NF-κB activation following LPS treatment20. To confirm the role of PDI in LPS-induced NF-κB activation, thus, we investigated the effects of PDI siRNA on p65 S276 phosphorylation. PDI knockdown effectively inhibited LPS-induced PDI upregulation (χ2(3) = 19.511, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 3A and Supplementary Fig. 3). As compared to control siRNA, PDI knockdown attenuated LPS-induced NF-κB upregulation (χ2(3) = 21.454, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 3A) and p65 S276 phosphorylation in P2X7R+/+ and P2X7R−/− mice (χ2(3) = 19.926, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 3A and Supplementary Fig. 3). These findings indicate that NF-κB and PDI may reciprocally regulate each other following LPS treatment, and P2X7R may intervene in this interaction.

Roles of PDI in NF-κB activation iNOS induction in response to LPS. P2X7R deletion ameliorates PDI upregulation, NF-κB activation and iNOS induction in response to LPS. PDI knockdown effectively inhibits LPS-induced PDI upregulation and attenuates LPS-induced NF-κB activation and iNOS induction in P2X7R+/+ and P2X7R−/− mice. (A) Effects of PDI knockdown on NF-κB activation under post-LPS condition. Representative Western blot images for PDI, p65 and p65 S276 levels (left panel) and quantification of the effects of PDI knockdown on PDI, p65 and p65 S276 densities (right panel) under post-LPS condition (#,*p < 0.05 vs. control siRNA and P2X7R+/+mice, respectively; Kruskal–Wallis test, n = 7, respectively). (B) Effects of PDI knockdown on iNOS induction in response to LPS. Representative Western blot images for iNOS level (left panel) and quantification of the effects of PDI knockdown on iNOS density (right panel) following LPS treatment (#,*p < 0.05 vs. control siRNA and P2X7R+/+mice, respectively; Kruskal–Wallis test, n = 7, respectively).
LPS-induced NF-κB activation leads to iNOS induction, which evokes nitrosative stress29,30,31. To elucidate the role of PDI in NF-κB-mediated iNOS induction, we evaluated the effect of PDI knockdown on iNOS induction following LPS treatment. Under physiological conditions, these was no difference in iNOS protein level between P2X7R+/+ and P2X7R−/− mice. LPS increased iNOS expression in P2X7R+/+ mice more than P2X7R−/− mice (p = 0.017, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 3B and Supplementary Fig. 3). PDI siRNA attenuated LPS-induced iNOS upregulation in both strains (χ2(5) = 32.078, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 3B). These findings indicate that P2X7R may amplify PDI-mediated argumentation of NF-κB-iNOS activation in response to LPS.
P2X7R-mediated NF-κB activation upregulates PDI expression without affecting the yield of S-nitrosylation of PDI following LPS treatment
P2X7R also regulates iNOS-mediated nitrosative stress2,3,4,5. Indeed, P2X7R ablation reduces SNO-cysteine production in microglia and astrocytes under physiological and LPS-induced inflammatory conditions4. In the present study, furthermore, P2X7R ablation inhibited LPS-induced iNOS induction. Since S-nitrosylation of PDI determines its enzyme properties as a denitrosylase or a NO transporter6,32, we investigated whether P2X7R-mediated NF-κB activation affects S-nitrosylation of PDI following LPS treatment.
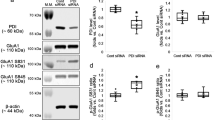
P2X7R ablation did not affect basal PDI level. However, P2X7R ablation attenuated the increased PDI expression in response to LPS. SN50 ameliorated LPS-induced PDI upregulation in both strains (χ2(5) = 35.958, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 4A, B and Supplementary Fig. 4). Under physiological condition, SNO-PDI level in P2X7R−/− mice was lower than that in P2X7R+/+ mice. LPS increased SNO-PDI level in P2X7R+/+ mice more than P2X7R−/− mice, which were attenuated by SN50 (χ2(5) = 36.462, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 4A, C). Under physiological condition, SNO-PDI ratio (SNO-PDI/total PDI ratio) in P2X7R−/− mice was lower than that in P2X7R+/+ mice (p = 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 4A, D). However, LPS increased SNO-PDI ratio in P2X7R−/− mice (p = 0.032, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively), but not in P2X7R+/+ mice (Fig. 4A, D). SN50 did not affect SNO-PDI ratio in both strains following LPS treatment (Fig. 4A, D). These findings indicate that P2X7R may augment LPS-induced neuroinflammation by enhancing basal SNO-PDI ratio and NF-κB-mediated PDI upregulation, although both P2X7R and NF-κB pathways may not affect the yield of S-nitrosylation of PDI following LPS treatment.

The roles of P2X7R-mediated NF-κB activation in PDI expression and its S-nitrosylation following LPS treatment. P2X7R ablation cannot affect basal PDI level, but decreases S-nitrosylated (SNO-) PDI level under physiological condition. P2X7R deletion ameliorates LPS-induced PDI upregulation and SNO-PDI level, which are ameliorated by SN50 in P2X7R+/+ and P2X7R−/− mice. However, LPS increases SNO-PDI ratio only in P2X7R−/− mice, which is unaffected by SN50. (A) Representative Western blot images for PDI and SNO-PDI levels following LPS treatment. (B–D) Quantification of the effects of SN50 on PDI upregulation (B), SNO-PDI level (C) and SNO-PDI ratio (D) in response to LPS (#,*,$p < 0.05 vs. control, P2X7R+/+mice and vehicle, respectively; Kruskal–Wallis test, n = 7, respectively).
P2X7R and PDI do not affect SNO-p65 level following LPS treatment
NF-κB p65 subunit is constitutively S-nitrosylated on cysteine 38 residue, which inhibits NF-κB transcriptional activity33,34,35. LPS exerts a decrease in SNO-p65 levels concomitant with NF-κB activation in initiation of the inflammatory responses36. Therefore, S-nitrosylation of p65 is proposed as a negative feedback mechanism to iNOS induction in response to LPS. Since the present study revealed that LPS increased SNO-PDI level, we validated whether PDI regulates NF-κB activation by affecting S-nitrosylation of p65 subunit following LPS treatment.
P2X7R deletion did not alter basal p65 level. LPS increased p65 expression level in P2X7R+/+ mice more than P2X7R−/− mice, which were ameliorated by PDI siRNA (χ2(5) = 37.425, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 5A, B and Supplementary Fig. 5). Under physiological condition, there was no difference in SNO-p65 level between P2X7R+/+ and P2X7R−/− mice. LPS reduced SNO-p65 levels in both strains, which were unaffected by PDI knockdown (χ2(5) = 27.701, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 5A, C). There was no difference in SNO-p65 ratio (SNO-p65/total p65 ratio) between P2X7R+/+ and P2X7R−/− mice under physiological condition (Fig. 5A, D). LPS also diminished SNO-p65 ratio in both strains, which was unaffected by PDI knockdown (Fig. 5A, D), although P2X7R deletion mitigated the reduced SNO-p65 ratio following LPS treatment (χ2(5) = 34.112, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 5A, D). These findings indicate that P2X7R may decrease LPS-induced SNO-p65 ratio, independent of PDI, and suggest that PDI may regulate the progress of LPS-induced neuroinflammation rather than initiation.

The effects of PDI knockdown on S-nitrosylation of p65 following LPS treatment. P2X7R ablation cannot affect basal p65 and S-nitrosylated (SNO-) p65 levels under physiological condition. LPS increases p65 expression, but reduces SNO-p65 level and its ratio. P2X7R deletion ameliorates LPS-induced p65 upregulation without affecting SNO-p65 level. Thus, P2X7R deletion mitigates LPS-induced downregulation of SNO-p65 ratio. PDI knockdown attenuates LPS-induced p65 upregulation in P2X7R+/+ and P2X7R−/− mice without altering SNO-p65 level and SNO-p65 ratio. (A) Representative Western blot images for p65 and SNO-p65 levels following LPS treatment. (B–D) Quantification of the effects of PDI knockdown on p65 upregulation (B), SNO-p65 level (C) and SNO-p65 ratio (D) in response to LPS (#,*,$p < 0.05 vs. control, P2X7R+/+mice and control siRNA, respectively; Kruskal–Wallis test, n = 7, respectively).
SNO-PDI augments the integration of P2X7R-NF-κB signaling axis following LPS treatment
PDI-mediated S-nitrosylation enhances surface P2X7R expression following status epilepticus (SE)13. Furthermore, SNAP (a NO donor) activates microglia by inducing nuclear p65 translocation37. Therefore, it is likely that SNO-PDI may play an important role in a positive feedback loop of P2X7R-NF-κB interaction. To confirm this, we applied SNAP to confirm this hypothesis.
Under physiological condition, SNAP augmented IB4 signal (χ2(3) = 22.934, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 6A, B) and microglial p65 S276 phosphorylation (χ2(3) = 22.908, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 6A, C) in P2X7R+/+ mice more than P2X7R−/− mice, indicating microglial activation. SNAP also similarly increased total PDI (χ2(3) = 20.317, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 7A, B and Supplementary Fig. 6) and SNO-PDI levels (χ2(3) = 22.852, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 7A, C) in P2X7R+/+ and P2X7R−/− mice. Similar to the case of LPS treatment, SNAP increased SNO-PDI ratio in P2X7R−/− mice, but not in P2X7R+/+ mice (χ2(3) = 16.588, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 7A, D). Although SNAP did not affect total- and SNO-p65 levels in both strains (Fig. 7A, E, F), it enhanced p65 S276 phosphorylation in P2X7R+/+ mice more than P2X7R−/− mice (χ2(3) = 22.901, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 7A, G). In P2X7R+/+ mice, SNAP enhanced P2X7R expression (χ2(3) = 25.719, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 7A, H), SNO-P2X7R level (χ2(3) = 26.055, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 7A, I) and SNO-P2X7R ratio (χ2(3) = 26.601, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 7A, J), concomitant with the increased surface P2X7R expression (χ2(3) = 25.831, p < 0.001, Kruskal–Wallis with Dunn-Bonferroni post-hoc test, n = 7, respectively, Fig. 7A, K). Taken together, our findings indicate that SNAP-induced S-nitrosylation of PDI may activate NF-κB, which may be elicited by P2X7R in a positive feedback manner.

The effects of NO on microglial activation and p65 S276 phosphorylation under physiological condition. SNAP (a NO donor) leads to microglial activation and p65 phosphorylation under physiological condition, which are attenuated by P2X7R deletion. (A) Representative photos for p65 S276 phosphorylation in microglia following SNAP treatment. (B–C) Quantification of the effects of SNAP on IB4 (a microglial marker, B) and p65 S276 intensities (C; #,*p < 0.05 vs. vehicle and P2X7R+/+mice, respectively; Kruskal–Wallis test, n = 7, respectively).

The effects of SNAP on expression and S-nitrosylation of PDI, p65 and P2X7R, and surface P2X7R expression under physiological condition. SNAP upregulates PDI expression and S-nitrosylated (SNO-) PDI level, which are unaffected by P2X7R deletion. SNAP augments p65 S276 phosphorylation in P2X7R+/+ and P2X7R−/− mice without altering p65 and SNO-p65 densities. In addition, SNAP increases total and surface P2X7R expressions, concomitant with enhanced S-nitrosylation of P2X7R in P2X7R+/+ mice. (A) Representative images for expression and S-nitrosylation of PDI, p65 and P2X7R, and surface P2X7R expression following SNAP treatment. (B–K) Quantification of the effects of SNAP on PDI, SNO-PDI, p65, SNO-p65, p65 S276, P2X7R, SNO-P2X7R and surface P2X7R levels (#,*p < 0.05 vs. vehicle and P2X7R+/+ mice, respectively; Kruskal–Wallis test, n = 7, respectively).
Discussion
TLR4-NF-κB-iNOS signaling pathway plays an important role in LPS-induced inflammatory processes29,38. Furthermore, oxidized ATP (OxATP, an irreversible P2X7R inhibitor) attenuates a NF-κB binding activity, iNOS expression and NO production in response to various stimuli including LPS39,40. These reports indicate that P2X7R itself modulates NF-κB activity, independent of TLR4. After exposure to LPS, interestingly, P2X7R contributes microglial hyperactivity, which augments NF-κB-mediated iNOS expression and NO production41,42,43. Therefore, we hypothesized that TLR4 and P2X7R may compose a positive feedback loop in the mouse hippocampus during inflammatory processes, which would activate NF-κB-iNOS signaling pathway in response to LPS in vivo. Considering PDI-mediated trafficking of P2X7R and p65 S276 phosphorylation13, it is likely that PDI may act as an intermediator of this TLR4-P2X7R interaction by regulating NF-κB activity after LPS stimulation. The present data reveal that S-nitrosylation of PDI contributes LPS-induced NF-κB activation, which was attenuated by P2X7R deletion. Thus, these findings suggest that PDI may integrate TLR4- and P2X7R-mediated signaling pathways during inflammation (Fig. 8).

Scheme of roles of PDI in the integration of TLR4- and P2X7R-mediated signaling pathways during inflammation. LPS-induced TLR4 activation led to NF-κB activation by S-nitrosylation and phosphorylation of p65 subunit, which upregulated iNOS and PDI expression in microglia. S-nitrosylation of PDI induced by iNOS further augmented NF-κB activation by a direct interaction as well as the enhanced surface P2X7R expression, independent of S-nitrosylation of p65.
PDI is an oxidoreductase in the ER of all types of cells. Upregulation of PDI is an important part of unfolded protein response (UPR)44. LPS activates the NF-κΒ pathway and enhances PDI induction16,17, which elicits expressions of proinflammatory factors including iNOS20,21,22,45. Compatible with these reports, the present data show that LPS led to PDI upregulation, NF-κB S276 phosphorylation and iNOS induction, which was ameliorated by P2X7R deletion, PDI knockdown and SN50 treatment. Indeed, PDI ablation abrogates the inflammatory function of macrophages20. Therefore, our findings indicate that P2X7R and PDI may contribute to NF-κB activation induced by LPS and provide an evidence for the presence of TLR4-NF-κB-PDI-P2X7R-mediated signaling loop that may be activated by LPS in a positive feedback mechanism.
PDI also acts as a denitrosylase as well as a NO donor to regulate S-nitrosylation of various target proteins in nuclear envelopes, plasma membranes, mitochondria, and other organelles6,18,32,46,47. Therefore, S-nitrosylation changes the properties of PDI from chaperone/isomerase/denitrosylase to a NO transporter6,7,8,9,10. In the present study, SNO-PDI level and its ratio in P2X7R−/− mice were lower than those in P2X7R+/+ mice under physiological condition. LPS increased SNO-PDI level in P2X7R+/+ mice more than P2X7R−/− mice, which were attenuated by SN50. However, LPS increased SNO-PDI ratio in P2X7R−/− mice, but not in P2X7R+/+ mice, which were unaffected by SN50. These findings indicate that both P2X7R and NF-κB pathways may not alter the yield of S-nitrosylation of PDI following LPS treatment. Instead, P2X7R may augment LPS-induced neuroinflammation by enhancing basal SNO-PDI ratio and NF-κB-mediated PDI upregulation.
On the other hand, the present study demonstrates that under physiological condition SNAP increased total PDI level in P2X7R−/− mice similar to that in P2X7R+/+ mice. SNAP induces ER stress in microglia48, which upregulates PDI expression49. Therefore, it is unsurprising that SNAP upregulated PDI expression in both strains. Furthermore, SNAP similarly elevated SNO-PDI level in P2X7R+/+ and P2X7R−/− mice, although SNAP increased SNO-PDI ratio only in the P2X7R−/− mice, indicating that SNAP induced S-nitrosylation of PDI, independent of P2X7R. However, SNAP induced microglial activation in P2X7R+/+ mice more than P2X7R−/− mice. SNAP also elicited p65 S276 phosphorylation in P2X7R+/+ mice more than P2X7R−/− mice. SNAP itself leads to microglial activation through IκB-mediated canonical NF-κB pathway37. These data indicate that P2X7R may contribute to SNO-PDI-mediated p65 S276 phosphorylation. Indeed, PDI siRNA attenuates microglial activation and the increased P2X7R surface expression induced by SE13. Consistent with this report, the present data also reveal that SNAP increased surface P2X7R expression in P2X7R+/+ mice, accompanied by the enhancement of total- and SNO-P2X7R level. Therefore, our findings indicate that SNO-PDI may control p65 S276 phosphorylation by modulating P2X7R as well as the direct interaction. Taken together with the data obtained from LPS trials, the present data suggest that SNO-PDI may activate NF-κB, which may be elicited by P2X7R in a positive feedback manner.
NF-κB transcriptional activity is negatively regulated by S-nitrosylation of p65 subunit33,34,35. Indeed, LPS exerts a decrease in SNO-p65 levels concomitant with NF-κB activation in initiation of the inflammatory responses36. Of interest, PDI suppresses TRX that increases DNA binding activity of NF-κB by denitrosylating SNO-p65 and reducing a disulfide bond on NF-κB18,23,24,50. Since P2X7R activation augments the DNA-binding activity of NF-κB and iNOS expression following LPS treatment4,51, it is plausible that P2X7R would reinforce LPS-induced NF-κB transactivation by regulating S-nitrosylation of p65 subunit. Unlike PDI, the present data show that no difference in SNO-p65 levels between P2X7R+/+ and P2X7R−/− mice was observed following LPS treatment. The present data also demonstrate that PDI knockdown did not affect SNO-p65 level in P2X7R+/+ and P2X7R−/− mice under physiological- and post-LPS conditions. These findings indicate that S-nitrosylation of p65 level may be independent of PDI. Furthermore, SNAP did not affect total- and SNO-p65 levels in both strains under physiological condition. NF-κB p65 subunit is constitutively S-nitrosylated on cysteine 38 residue, which inhibits NF-κB transcriptional activity33,34,35. LPS elicits a decrease in SNO-p65 level concomitant with NF-κB activation. This process is mediated by TRX. Indeed, inhibition of Trx activity attenuates LPS-induced SNO-p65 denitrosylation and NF-κB activation36. Therefore, our findings indicate that TLR4-NF-κB-PDI-P2X7R signaling loop may modulate LPS-induced neuroinflammation, independent of S-nitrosylation of p65 subunit.
The present study demonstrates that under physiological condition SNAP increased surface P2X7R expression in P2X7R+/+ mice, accompanied by the enhancement of total- and SNO-P2X7R level. An extracellular domain of P2X7R has ten cysteine residues forming five intrasubunit disulfide bonds (SS1–SS5)52, which are needed for its trafficking onto the cell surface53. S-nitrosylation facilitates disulfide bond formation (oxidation) between cysteine residues54, which is required for the trafficking of P2X7R53. Regarding that PDI exerts the trafficking of P2X7R by modulating dynamic redox status and S-nitrosylation of cysteine residues on an extracellular domain forming disulfide bonds13, SNAP may facilitate surface P2X7R trafficking by increasing SNO-PDI mediated S-nitrosylation of P2X7R.
P2X7R is an ATP-gated nonselective cationic channel. In microglia, P2X7R agonists rapidly activate AMP-activated kinase (AMPK). P2X7R antagonist can abolish agonist-induced AMPK activation, but not that induced by nigericin (K+ ionophore)55. Similarly, P2X7R agonist-induced interleukin-1β (IL-1β) release is inhibited by P2X7R antagonists, while IL-1β release mediated by nigericin is not affected by the addition of P2X receptor antagonists56. In addition, LPS-induced macrophage activation is strongly attenuated by the P2X7R antagonists. However, P2X7R antagonists do not affect macrophage stimulation by Ca2+ ionophore A2318757. Considering these previous reports, it is likely that P2X7R-mediated K+ efflux and/or Ca2+ influx may affect LPS-induced S-nitrosylation of PDI. Further studies are needed to validate the effects of ionophore on TLR4-P2X7R interactions in response to LPS-induced nitrosative stress.
In the present study, we provided a novel PDI-mediated positive feedback loop between TLR4- and P2X7R-mediated signaling pathway. LPS-induced TLR4 activation led to NF-κB p65 S276 phosphorylation, which upregulated iNOS and PDI expression in microglia. In turn, iNOS-induced S-nitrosylation of PDI further augmented p65 S276 phosphorylation by a direct interaction as well as the enhanced surface P2X7R expression, independent of S-nitrosylation of p65. These findings serve as the first comprehensive description indicating that PDI may integrate TLR4- and P2X7R-mediated signaling pathway in a positive feedback manner under neuroinflammatory condition. Therefore, our findings suggest the targeting of PDI may be one of the important therapeutic strategies of neuroinflammation.
Methods
Experimental animals and chemicals
Animal experiments were designed and performed in accordance with the relevant guidelines and regulations. For all the procedures of the study we followed the ARRIVE guidelines. Animal protocols were approved by the Institutional Animal Care and Use Committee of Hallym University (Chuncheon, South Korea, Code number: (#Hallym 2021–30, approval date: May 17, 2021). All reagents were obtained from Sigma-Aldrich unless otherwise indicated. Male C57Bl/6 J (Wild type, P2X7R+/+) and P2X7R knockout (KO, P2X7R−/−) mice (60–90-day-old, 25–30 g, The Jackson Laboratory, USA) were used in the present study. Mice were given a diet and water ad libitum under controlled conditions (22 ± 2 °C, 55% ± 5% humidity, and 12-h light/12-h dark cycle).
Surgery, PDI knockdown, drug infusions and LPS treatment
To avoid the limitation of the permeability of the chemicals and siRNA across blood–brain barrier in vivo and maintain their constant concentration in the brain, we applied intracerebroventricular administration. Mice were anesthetized with Isoflurane (3% induction, 1.5–2% for surgery and 1.5% maintenance in a 65:35 mixture of N2O:O2). A brain infusion kit 3 (Alzet, USA) was implanted into the right lateral ventricle (1 mm posterior; 1.5 mm lateral; 3.5 mm depth from bregma) and connected to an osmotic pump (1007D, Alzet, USA) containing a vehicle, SN50 (20 μM), S-nitroso-N-acetyl-D,L-penicillamine (SNAP) (0.1 μM), non-silencing RNA (control siRNA) or mouse PDI siRNA (CCUUUGCUAGCGAAUCUCAGAGCC), which were continuously infused over 6-day period58. Three days after surgery, mice were given a single dose of LPS (5 mg/kg) or an equal volume of normal saline instead of LPS. Saline-treated mice were used as controls4.
Western blot
Three days after LPS treatment, animals were sacrificed by decapitation under urethane anesthesia (1.5 g/kg, i.p.). The hippocampus was dissected out and homogenized in lysis buffer (50 mM Tris containing 50 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (pH 7.4), ethylene glycol tetraacetic acid (pH 8.0), 0.2% Tergitol type NP-40, 10 mM ethylenediaminetetraacetic acid (pH 8.0), 15 mM sodium pyrophosphate, 100 mM β-glycerophosphate, 50 mM NaF, 150 mM NaCl, 2 mM sodium orthovanadate, 1 mM phenylmethylsulfonyl fluoride, and 1 mM dithiothreitol). Total protein content was measured by BCA protein assay kit. Western blot was performed according to standard procedures (Table 1). The signals were scanned and analyzed by ImageQuant LAS4000 system (GE health). The values of each sample were normalized with the corresponding amount of anti-β-actin (input). The phosphoprotein/total protein ratio was represented as the phosphorylation density1,4,5,9,10,13,58.
Measurement of S-nitrosylation
Modified biotin switch assay was performed with the S-nitrosylation Western Blot Kit (ThermoFisher, USA) according to the manufacturer’s protocol. Briefly, lysates were reacted with ascorbate in HENS buffer for specific labeling with iodoTMTzero reagents with MMT pretreatment. Protein labeling can be confirmed by Western blot using TMT antibody. Thereafter, TMT-labeled proteins were purified by Anti-TMT Resin, eluted by TMT elusion buffer, and identified by Western blot according to standard procedures as aforementioned. For technical controls, we omitted ascorbate for each sample. The ratio of SNO-protein to total protein was described as S-nitrosylation ratio10,13,58.
Membrane fraction
To analyze membrane expressions of P2X7R, we used subcellular Protein Fractionation Kit for Tissues (Thermo Scientific, USA), according to the manufacturer’s instructions. Next, Western blot was performed according to standard procedures13.
Immunohistochemistry
Three days after LPS injection, animals were anesthetized with urethane anesthesia (1.5 g/kg, i.p.) and perfused transcardially with 4% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4). Brains were post-fixed in the same fixative overnight and then cryoprotected and sectioned at 30 μm with a cryostat. Free-floating sections were washed 3 times in PBS (0.1 M, pH 7.3) and incubated with 3% bovine serum albumin in PBS for 30 min at room temperature. Later, sections were incubated with a cocktail solution containing primary antibodies (Table 1) in PBS containing 0.3% Triton X-100 overnight at room temperature. Thereafter, sections were visualized with appropriate Cy2- and Cy3-conjugated secondary antibodies. Some tissues were incubated in biotinylated IgG and avidin-peroxidase complex and developed in 3,3′-diaminobenzidine in 0.1 M Tris buffer. Immunoreaction was observed using an AxioScope microscope (Carl Zeiss Korea, Seoul, South Korea). To establish the specificity of the immunostaining, a negative control test was carried out with preimmune serum instead of the primary antibody. All experimental procedures in this study were performed under the same conditions and in parallel. To measure fluorescent intensity, five areas/animals (1 × 104 μm2/area) were randomly selected within the hippocampus (5 sections from each animal, n = 7 in each group). Thereafter, the mean intensity of each section was measured by using AxioVision Rel. 4.8 and ImageJ software. Intensity measurements were represented as the number of a 256 grayscale. The intensity of each section was standardized by setting the threshold level (mean background intensity obtained from five image inputs). Fluorescent intensity was performed by two different investigators who were blind to the classification of tissues. Manipulation of the images was restricted to threshold and brightness adjustments to the whole image1,4,5,9,10,13.
Statistical analysis
Comparisons of data among groups were analyzed by Mann–Whitney test or Kruskal–Wallis test followed by Dunn-Bonferroni post-hoc test. A p < 0.05 is considered to be statistically different.
Data availability
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
References
Kim, J. E. et al. The effect of P2X7 receptor activation on nuclear factor-κB phosphorylation induced by status epilepticus in the rat hippocampus. Hippocampus 23, 500–514. https://doi.org/10.1002/hipo.22109 (2013).
Feng, L. et al. P2X7R blockade prevents NLRP3 inflammasome activation and brain injury in a rat model of intracerebral hemorrhage: Involvement of peroxynitrite. J. Neuroinflammation 12, 190. https://doi.org/10.1186/s12974-015-0409-2 (2015).
Huang, C. et al. Inhibition of P2X7 receptor ameliorates nuclear factor-kappa B mediated neuroinflammation induced by status epilepticus in rat hippocampus. J. Mol. Neurosci. 63, 173–184. https://doi.org/10.1007/s12031-017-0968-z (2017).
Lee, D. S. & Kim, J. E. P2X7 receptor augments LPS-induced nitrosative stress by regulating Nrf2 and GSH levels in the mouse hippocampus. Antioxidants (Basel) 11, 778. https://doi.org/10.3390/antiox11040778 (2022).
Park, H. & Kim, J. E. Deletion of P2X7 receptor decreases basal glutathione level by changing glutamate-glutamine cycle and neutral amino acid transporters. Cells 9, 995. https://doi.org/10.3390/cells9040995 (2020).
Zai, A., Rudd, M. A., Scribner, A. W. & Loscalzo, J. Cell-surface protein disulfide isomerase catalyzes transnitrosation and regulates intracellular transfer of nitric oxide. J. Clin. Invest. 103, 393–399. https://doi.org/10.1172/JCI4890 (1999).
Turano, C., Coppari, S., Altieri, F. & Ferraro, A. Proteins of the PDI family: Unpredicted non-ER locations and functions. J. Cell Physiol. 193, 154–163. https://doi.org/10.1002/jcp.10172 (2002).
Ramachandran, N., Root, P., Jiang, X. M., Hogg, P. J. & Mutus, B. Mechanism of transfer of NO from extracellular S-nitrosothiols into the cytosol by cell-surface protein disulfide isomerase. Proc. Natl. Acad. Sci. USA 98, 9539–9544. https://doi.org/10.1073/pnas.171180998 (2001).
Kim, J. Y., Ko, A. R., Hyun, H. W., Min, S. J. & Kim, J. E. PDI regulates seizure activity via NMDA receptor redox in rats. Sci. Rep. 7, 42491. https://doi.org/10.1038/srep42491 (2017).
Lee, D. S. & Kim, J. E. PDI-mediated S-nitrosylation of DRP1 facilitates DRP1-S616 phosphorylation and mitochondrial fission in CA1 neurons. Cell Death Dis. 9, 869. https://doi.org/10.1038/s41419-018-0910-5 (2018).
Furlan-Freguia, C., Marchese, P., Gruber, A., Ruggeri, Z. M. & Ruf, W. P2X7 receptor signaling contributes to tissue factor-dependent thrombosis in mice. J. Clin. Invest. 121, 2932–2944. https://doi.org/10.1172/JCI46129 (2011).
Ruf, W. Role of thiol pathways in TF procoagulant regulation. Thromb. Res. 129, S11-12. https://doi.org/10.1016/j.thromres.2012.02.020 (2012).
Lee, D. S. & Kim, J. E. Protein disulfide isomerase-mediated S-nitrosylation facilitates surface expression of P2X7 receptor following status epilepticus. J. Neuroinflamm. 18, 14. https://doi.org/10.1186/s12974-020-02058-y (2021).
Furusawa, J. et al. Licochalcone A significantly suppresses LPS signaling pathway through the inhibition of NF-kappaB p65 phosphorylation at serine 276. Cell Signal. 21, 778–785. https://doi.org/10.1016/j.cellsig.2009.01.021 (2009).
Gadgil, H. S. et al. Proteome of monocytes primed with lipopolysaccharide: Analysis of the abundant proteins. Proteomics 3, 1767–1780. https://doi.org/10.1002/pmic.200300532 (2003).
Triantafyllou, E. A., Mylonis, I., Simos, G. & Paraskeva, E. Hypoxia induces pro-fibrotic and fibrosis marker genes in hepatocellular carcinoma cells independently of inflammatory stimulation and the NF-κΒ pathway. Hypoxia (Auckl) 7, 87–91. https://doi.org/10.2147/HP.S235967 (2019).
Sang, A. et al. Quercetin attenuates sepsis-induced acute lung injury via suppressing oxidative stress-mediated ER stress through activation of SIRT1/AMPK pathways. Cell Signal. 96, 110363. https://doi.org/10.1016/j.cellsig.2022.110363 (2022).
Higuchi, T., Watanabe, Y. & Waga, I. Protein disulfide isomerase suppresses the transcriptional activity of NF-kappaB. Biochem. Biophys. Res. Commun. 318, 46–52. https://doi.org/10.1016/j.bbrc.2004.04.002 (2004).
Zhou, M. et al. Downregulation of protein disulfide isomerase in sepsis and its role in tumor necrosis factor-alpha release. Crit. Care 12, R100. https://doi.org/10.1186/cc6977 (2008).
Xiao, Y. et al. Protein disulfide isomerase silence inhibits inflammatory functions of macrophages by suppressing reactive oxygen species and NF-κB pathway. Inflammation 41, 614–625. https://doi.org/10.1007/s10753-017-0717-z (2018).
Wang, W. T., Sun, L. & Sun, C. H. PDIA3-regulted inflammation and oxidative stress contribute to the traumatic brain injury (TBI) in mice. Biochem. Biophys. Res. Commun. 518, 657–663. https://doi.org/10.1016/j.bbrc.2019.08.100 (2019).
Kamarehei, M. et al. Inhibition of protein disulfide isomerase has neuroprotective effects in a mouse model of experimental autoimmune encephalomyelitis. Int. Immunopharmacol. 82, 106286. https://doi.org/10.1016/j.intimp.2020.106286 (2020).
Matthews, J. R., Wakasugi, N., Virelizier, J. L., Yodoi, J. & Hay, R. T. Thioredoxin regulates the DNA binding activity of NF-kappa B by reduction of a disulphide bond involving cysteine 62. Nucleic Acids Res. 20, 3821–3830. https://doi.org/10.1093/nar/20.15.3821 (1992).
Hayashi, T., Ueno, Y. & Okamoto, T. Oxidoreductive regulation of nuclear factor kappa B. Involvement of a cellular reducing catalyst thioredoxin. J. Biol. Chem. 268, 11380–11388 (1993).
Roy, A., Fung, Y. K., Liu, X. & Pahan, K. Up-regulation of microglial CD11b expression by nitric oxide. J. Biol. Chem. 281, 14971–14980. https://doi.org/10.1074/jbc.M600236200 (2006).
Zhong, L. M. et al. Resveratrol inhibits inflammatory responses via the mammalian target of rapamycin signaling pathway in cultured LPS-stimulated microglial cells. PLoS ONE 7, e32195. https://doi.org/10.1371/journal.pone.0032195 (2012).
Lee, S. K., Kim, J. E., Kim, Y. J., Kim, M. J. & Kang, T. C. Hyperforin attenuates microglia activation and inhibits p65-Ser276 NFκB phosphorylation in the rat piriform cortex following status epilepticus. Neurosci. Res. 85, 39–50. https://doi.org/10.1016/j.neures.2014.05.006 (2014).
Kim, W. I. et al. Differential nuclear factor-kappa B phosphorylation induced by lipopolysaccharide in the hippocampus of P2X7 receptor knockout mouse. Neurol. Res. 35, 369–381. https://doi.org/10.1179/1743132812Y.0000000137 (2013).
Xie, Q. W., Kashiwabara, Y. & Nathan, C. Role of transcription factor NF-kappa B/Rel in induction of nitric oxide synthase. J. Biol. Chem. 269, 4705–4708 (1994).
Zhao, J. et al. Neuroinflammation induced by lipopolysaccharide causes cognitive impairment in mice. Sci. Rep. 9, 5790. https://doi.org/10.1038/s41598-019-42286-8 (2019).
Ju, C. et al. Inhibition of Dyrk1A Attenuates LPS-Induced Neuroinflammation via the TLR4/NF-κB P65 Signaling Pathway. Inflammation 45, 2375–2387. https://doi.org/10.1007/s10753-022-01699-w (2022).
Kallakunta, V. M., Slama-Schwok, A. & Mutus, B. Protein disulfide isomerase may facilitate the efflux of nitrite derived S-nitrosothiols from red blood cells. Redox Biol. 1, 373–380. https://doi.org/10.1016/j.redox.2013.07.002 (2013).
Kelleher, Z. T., Matsumoto, A., Stamler, J. S. & Marshall, H. E. NOS2 regulation of NF-kappaB by S-nitrosylation of p65. J. Biol. Chem. 42, 30667–30672. https://doi.org/10.1074/jbc.M705929200 (2007).
Perkins, N. D. Cysteine 38 holds the key to NF-kappaB activation. Mol. cell. 1, 1–3. https://doi.org/10.1016/j.molcel.2011.12.023 (2012).
Sen, N. et al. Hydrogen sulfide-linked sulfhydration of NF-κB mediates its antiapoptotic actions. Mol. Cell. 45, 13–24. https://doi.org/10.1016/j.molcel.2011.10.021 (2012).
Kelleher, Z. T. et al. Thioredoxin-mediated denitrosylation regulates cytokine-induced nuclear factor κB (NF-κB) activation. J. Biol. Chem. 289, 3066–3072. https://doi.org/10.1074/jbc.M113.503938 (2014).
Sudo, K., Takezawa, Y., Kohsaka, S. & Nakajima, K. Involvement of nitric oxide in the induction of interleukin-1 beta in microglia. Brain Res. 1625, 121–134. https://doi.org/10.1016/j.brainres.2015.08.030 (2015).
Barbierato, M. et al. Astrocyte-microglia cooperation in the expression of a pro-inflammatory phenotype. CNS Neurol. Disord. Drug Targets 12, 608–618. https://doi.org/10.2174/18715273113129990064 (2013).
Hu, Y. et al. Purinergic receptor modulation of lipopolysaccharide signaling and inducible nitric-oxide synthase expression in RAW 264.7 macrophages. J. Biol. Chem. 273, 27170–27175. https://doi.org/10.1074/jbc.273.42.27170 (1998).
Tung, H. C. et al. The beneficial effects of P2X7 antagonism in rats with bile duct ligation-induced cirrhosis. PLoS ONE 10, e0124654. https://doi.org/10.1371/journal.pone.0124654 (2015).
Guerra, A. N. et al. Purinergic receptor regulation of LPS-induced signaling and pathophysiology. J. Endotoxin Res. 9, 256–263. https://doi.org/10.1179/096805103225001468 (2003).
Friedle, S. A., Brautigam, V. M., Nikodemova, M., Wright, M. L. & Watters, J. J. The P2X7-Egr pathway regulates nucleotide-dependent inflammatory gene expression in microglia. Glia 59, 1–13. https://doi.org/10.1002/glia.21071 (2011).
Dang, R. et al. Fish oil supplementation attenuates neuroinflammation and alleviates depressive-like behavior in rats submitted to repeated lipopolysaccharide. Eur. J. Nutr. 57, 893–906. https://doi.org/10.1007/s00394-016-1373-z (2018).
Walker, A. K. et al. Protein disulphide isomerase protects against protein aggregation and is S-nitrosylated in amyotrophic lateral sclerosis. Brain 133, 105–116. https://doi.org/10.1093/brain/awp267 (2010).
Wang, H., Wang, P. & Zhu, B. T. Mechanism of erastin-induced ferroptosis in MDA-MB-231 human breast cancer cells: Evidence for a critical role of protein disulfide isomerase. Mol. Cell Biol. 42, e0052221. https://doi.org/10.1128/mcb.00522-21 (2022).
Akagi, S. et al. Distribution of protein disulfide isomerase in rat hepatocytes. J. Histochem. Cytochem. 36, 1533–1542. https://doi.org/10.1177/36.12.3192937 (1988).
Rigobello, M. P., Donella-Deana, A., Cesaro, L. & Bindoli, A. Distribution of protein disulphide isomerase in rat liver mitochondria. Biochem. J. 356, 567–570. https://doi.org/10.1042/0264-6021:3560567 (2001).
Kawahara, K. et al. Induction of CHOP and apoptosis by nitric oxide in p53-deficient microglial cells. FEBS. Lett. 506, 135–139. https://doi.org/10.1016/s0014-5793(01)02898-8 (2001).
Tanaka, S., Uehara, T. & Nomura, Y. Up-regulation of protein-disulfide isomerase in response to hypoxia/brain ischemia and its protective effect against apoptotic cell death. J. Biol. Chem. 275, 10388–10393. https://doi.org/10.1074/jbc.275.14.10388 (2000).
Hashemy, S. I. & Holmgren, A. Regulation of the catalytic activity and structure of human thioredoxin 1 via oxidation and S-nitrosylation of cysteine residues. J. Biol. Chem. 283, 21890–21898. https://doi.org/10.1074/jbc.M801047200 (2008).
Aga, M. et al. Evidence for nucleotide receptor modulation of cross talk between MAP kinase and NF-kappa B signaling pathways in murine RAW 264.7 macrophages. Am. J. Physiol Cell Physiol. 286, C923–C930. https://doi.org/10.1152/ajpcell.00417.2003 (2004).
Coddou, C., Yan, Z., Obsil, T., Huidobro-Toro, J. P. & Stojilkovic, S. S. Activation and regulation of purinergic P2X receptor channels. Pharmacol. Rev. 63, 641–683. https://doi.org/10.1124/pr.110.003129 (2011).
Jindrichova, M., Kuzyk, P., Li, S., Stojilkovic, S. S. & Zemkova, H. Conserved ectodomain cysteines are essential for rat P2X7 receptor trafficking. Purinergic Signal. 8, 317–325. https://doi.org/10.1007/s11302-012-9291-x (2012).
Lipton, S. A. et al. Cysteine regulation of protein function–as exemplified by NMDA-receptor modulation. Trends Neurosci. 25, 474–480. https://doi.org/10.1016/s0166-2236(02)02245-2 (2002).
Sekar, P., Huang, D. Y., Hsieh, S. L., Chang, S. F. & Lin, W. W. AMPK-dependent and independent actions of P2X7 in regulation of mitochondrial and lysosomal functions in microglia. Cell Commun. Signal. 16, 83. https://doi.org/10.1186/s12964-018-0293-3 (2018).
Bockstiegel, J., Engelhardt, J. & Weindl, G. P2X7 receptor activation leads to NLRP3-independent IL-1β release by human macrophages. Cell Commun. Signal. 21, 335. https://doi.org/10.1186/s12964-023-01356-1 (2023).
Balboa, M. A., Balsinde, J., Johnson, C. A. & Dennis, E. A. Regulation of arachidonic acid mobilization in lipopolysaccharide-activated P388D(1) macrophages by adenosine triphosphate. J. Biol. Chem. 274, 36764–36768. https://doi.org/10.1074/jbc.274.51.36764 (1999).
Lee, D. S., Kim, T. H., Park, H. & Kim, J. E. PDI augments kainic acid-induced seizure activity and neuronal death by inhibiting PP2A-GluA2-PICK1-mediated AMPA receptor internalization in the mouse hippocampus. Sci. Rep. 13, 13927. https://doi.org/10.1038/s41598-023-41014-7 (2023).
Acknowledgements
This study was supported by a grant of National Research Foundation of Korea (NRF) grant (No. 2021R1A2C4002003). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
J-EK designed and supervised the project. D-SL, T-HK and SYW performed the experiments described in the manuscript with J-EK and analyzed the data. J-EK wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
Animal protocols were approved by the Institutional Animal Care and Use Committee of Hallym University, Chuncheon, South Korea, Code number: (#Hallym 2021-30, approval date: May 17, 2021).
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kim, JE., Wang, S.H., Lee, DS. et al. Protein disulfide isomerase integrates toll-like receptor 4 and P2X7 receptor signaling pathways during lipopolysaccharide-induced neuroinflammation. Sci Rep 15, 7906 (2025). https://doi.org/10.1038/s41598-025-92780-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-92780-5
