
Abstract
Emerging evidence suggests that N6-methyladenosine (m6A) modification significantly influences lung injury, lung cancer, and immune responses. The current study explores the potential involvement of m6A modification in the development of IPF. This research analyzed the GSE93606 dataset of 20 non-IPF and 154 IPF patients, identifying 26 m6A regulators and developing predictive models with RF and SVM, assessed via ROC curves. A nomogram was created with selected m6A factors, including molecular subtyping, PCA for m6A features, immune cell analysis, DEG identification, and functional enrichment. In vitro experiments on MRC-5 cells used RT-qPCR and Western blotting, and virtual drug screening targeted the WTAP protein through molecular docking. Analysis revealed 26 differential m6A regulators in IPF patients, with 16 significant; IGFBP2 and YTHDF2 were overexpressed, while others decreased. RF and SVM models identified predictive m6A regulators, and a nomogram was developed using five factors to predict IPF incidence. Distinct m6A patterns showed changes in RNA levels of specific genes in the BLM-induced group, and five compounds targeting WTAP were identified. This research explored m6A factors’ impact on IPF diagnosis and prognosis, identifying WTAP as a potential biomarker.
Similar content being viewed by others

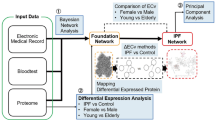

Introduction
Idiopathic pulmonary fibrosis (IPF) is a progressive, chronic, and irreversible diffuse interstitial pulmonary fibrosis disease with an unclear etiology1. Its characteristics include diffuse alveolar inflammation, epithelial-mesenchymal transition (EMT), and destruction of alveolar structure2,3. Although some progress has been made in therapeutic interventions in recent years, such as pirfenidone and nintedanib showing potential to slow disease progression and reduce mortality, the ultimate cure for IPF remains uncertain due to limited understanding of the underlying molecular mechanisms4,5. In this context, having a profound understanding of the pathogenesis of IPF is crucial to identify effective therapeutic targets. With the advancement of multi-omics data analysis technology, it offers a potential avenue to explore the development process of IPF, aiding in early diagnosis and the discovery of potential biomarkers6,7.
m6A methylation plays a crucial role in the mRNA lifecycle, participating in regulating mRNA transcription, output, translation, and degradation processes8. Abnormal m6A levels are linked to various diseases, including cancer, neurological disorders, and fibrosis8. Previous studies have indicated a close association between m6A RNA methylation and the progression of fibrosis in different fibrotic conditions like cardiac, liver, and renal fibrosis9. Reports suggest that m6A writers (METTL3, METTL14, and WTAP) can enhance the stability of pro-fibrotic gene mRNA, thereby promoting fibrosis3,10. However, the precise role of m6A RNA methylation in the onset and advancement of IPF remains poorly understood.
Inflammation is considered a crucial component in numerous autoimmune diseases and plays a significant role in various key pathogenic processes within IPF2,11. Throughout this progression, a significant release of cytokines and other substances, such as interleukin, fibroblast growth factor, tumor necrosis factor, occurs, resulting in fibroblast proliferation and the deposition of extracellular matrix12,13. Jia et al. utilized Mendelian randomization (MR) analysis on two sets of samples to investigate the causal connection between the risk of developing IPF and the serum inflammatory cytokine levels predicted by 24 common genes14. Several preclinical studies have indicated that the development of IPF is closely linked to signaling pathways mediating the inflammatory response associated with transforming growth factor-β (TGF-β)15. METTL14 can mediate the inflammatory response of macrophages in atherosclerosis through the NF-κB/IL-6 signaling pathway16. FTO may influence the translation of ATF4 by regulating its m6A level, thereby increasing the expression of p-STAT3 and inflammatory factors, ultimately leading to uveitis17. HOXA10 promotes metastasis in gastric cancer by regulating the TGFB2/Smad/METTL3 signaling pathway to induce EMT18. However, the precise role of m6A modification in the progression of IPF remains unclear.
In our study, we obtained IPF-related data information from the GEO database. Using a consensus clustering method, we established a gene model to predict IPF based on the expression levels of 26 m6A regulatory factors and categorized patients into two distinct m6A modification patterns19. We identified differentially expressed genes between these two pattern groups and selected genes closely associated with m6A modification and the progression of pulmonary fibrosis. Additionally, we employed virtual screening techniques to select 21 compounds as potential drugs for targeting WTAP in the treatment of IPF.
Methods and materials
Data acquisition
The GSE93606 dataset was obtained from the GEO database and comprises data from 20 non-IPF patients and 154 IPF patients (Supplementary Table S1). Through analysis of the differences between non-IPF and IPF patients, we have identified significant m6A regulators, resulting in the extraction of a total of 26 m6A regulators from the dataset. These regulators consist of 9 writers (METTL3, METTL14, METTL16, WTAP, VIRMA, ZC3H13, RBM15, RBM15B, and CBLL1), 15 readers (YTHDC1, YTHDC2, YTHDF1, YTHDF2, YTHDF3, HNRNPC, FMR1, LRPPRC, HNRNPA2B1, IGFBP1, IGFBP2, IGFBP3, RBMX, ELAVL1, and IGF2BP1), and 2 erasers (FTO and ALKBH5).
Construction of random forest model and support vector machine model
We employed RF and SVM models to construct a prediction model for IPF occurrence. To assess the performance of these models, we generated visualizations including the "reverse cumulative distribution of residuals" plot, residual box plot, and Receiver Operating Characteristic (ROC) curve.
The RF model is a supervised learning method that extends the concept of decision trees. In our study, we utilized the “RandomForest” package from the R to build the RF model20. We selected candidate m6A regulatory proteins from the pool of 26 m6A regulatory factors to predict IPF occurrence. The parameters ntree and mtry were set to 500 and 2, respectively. Additionally, we conducted importance analysis on these m6A regulatory factors and employed tenfold cross-validation to select the optimal model with minimal error for IPF prediction. The SVM model is a supervised machine learning algorithm based on the principle of structural risk minimization in statistical learning theory21. In our study, each data point was represented as a point in an n-dimensional space, where n corresponds to the number of m6A regulators. We then searched for an optimal hyperplane to accurately classify the two types of data (non-IPF and IPF).
Construction of a nomogram model
We constructed a column line chart model using the “rms” and “rmds” packages in R based on the selected candidate m6A regulatory factors to predict the prevalence of IPF patients. The calibration curve was used to assess the consistency between our predicted values and the actual outcomes. We performed decision curve analysis (DCA) and created clinical impact curves to evaluate whether decisions based on this model would be beneficial for patients.
Molecular subtyping based on important m6a regulatory factors
Consensus clustering is a resampling-based algorithm utilized to determine the number of subgroups and validate the clustering results. In our study, we employed the “ConsensusClusterPlus” software package in R to perform consensus clustering based on important m6A regulatory factors22. Furthermore, we created differential expression heatmaps to visually represent the differences in m6A typing. These heatmaps showcase the relative expression levels of m6A regulatory factors across various m6A subgroups, highlighting the contrasting expression patterns.
Estimation of m6A gene features
To quantify m6A patterns, we employed the Principal Component Analysis (PCA) algorithm to calculate the m6A scores for each sample. Initially, PCA was performed to distinguish the m6A patterns. Subsequently, the m6A scores were computed using the following formula: m6A score = PC1i, where PC1 represents the first principal component and i represents the expression of differentially expressed genes (DEGs)23.
Estimation of immune cell infiltration
Single-sample gene set enrichment analysis (ssGSEA) is used to evaluate the abundance of immune cells in IPF samples24. Initially, ssGSEA is employed to rank the gene expression levels within the samples. Then, we search for these genes within the input dataset and calculate their expression levels. Based on this assessment, we obtain the abundance of immune cells in each sample.
Identification of DEGs in m6A patterns and functional enrichment analysis using gene ontology
The “limma” package in R is used to filter DEGs between different m6A patterns. The selection criterion is set as p < 0.01, in accordance with previous studies and standards within the field.To understand the potential mechanisms of DEG involvement in IPF, the “clusterProfiler” package in R is employed for gene ontology (GO) functional enrichment analysis. The results are visualized using enrichment circle plots.
Cell culture
MRC-5 cells were procured from Procell Life Science Technology Co., Ltd. Log-phase MRC-5 cells were collected, and a cell suspension was prepared and seeded into a 6-well plate at a density of 1 × 105 cells. MRC-5 cells were maintained in a complete medium containing 10% fetal bovine serum (FBS) at 37 °C within a humidified incubator with 5% CO2. For the serum starvation experiments, cells at 70–80% confluence were washed twice with phosphate-buffered saline (PBS) and subsequently incubated in a medium devoid of FBS for a duration of 24 h. Following 24 h of cell starvation, complete culture medium and bleomycin culture medium (40 μg/ml, BLM)6 were added separately, and the cells were incubated for an additional 24 h for subsequent experiments.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
According to the manufacturer’s instructions, RNA extraction was performed using the SparkZol Reagent (AC0101) kit. Subsequently, cDNA was synthesized from total RNA using the SPARKscript II RT Plus Kit (with gDNA Eraser, AG0304), and mRNA expression was analyzed via RT-qPCR using 2 × SYBR Green qPCR Mix (with ROX, AH0104). The primer sequences are detailed in Table 1. The relative expression level of mRNA was normalized to the expression level of GAPDH within the same sample. Relative changes in target gene expression compared to the control group were determined using RT-qPCR analysis (2-ΔΔCt).
Western blotting
Cellular proteins were extracted using RIPA buffer and the protease inhibitor benzenesulfonyl fluoride (PMSF), followed by BCA concentration measurement. The proteins were separated through SDS-PAGE and then transferred onto PVDF membranes. Following sealing with 5% skim milk, primary antibodies (GAPDH (Protein, 10494–1-AP); FTO (Bios, Bs-7056R); WTAP (Protein, 60188–1-IG); α-SMA (Bios, Bs-10196R)) were incubated overnight at 4 °C. Subsequently, the membrane was treated at room temperature for one hour, followed by incubation with secondary antibodies coupled with horseradish peroxidase (HRP). The membrane was placed on a chemiluminescence imaging board, excess TBST was removed with filter paper, sufficient chemiluminescence substrate was added to cover the PVDF membrane, and exposure was carried out using a chemiluminescence imaging system.
Virtual screening for potential drugs
Download the electron microscopy structure of the WTAP protein (PDB ID: 7VF2) from the PDB online database (https://www.rcsb.org) and use Schrodinger’s Protein Preparation25 module to remove water molecules and excess ions, repair missing side chains and loop regions, followed by energy minimization. Utilize Schrodinger’s SiteMap module to predict potential binding sites and generate docking grid files using Schrodinger’s Glide Grid module. Download small molecule compound data from the YaTCM database (http://cadd.pharmacy.nankai.edu.cn/yatcm/home)26 and preprocess the molecules in the database using Schrodinger’s LigPrep module, including protonation, desalting, hydrogenation, generating tautomers, generating conformations, and performing energy minimization27. Conduct ADMET screening using Schrodinger’s QikProp module, retaining molecules that comply with Lipinski’s rule of five and do not contain reactive fragments for the next round of docking screening27. Subsequently, perform high-throughput virtual screening (HTVS), standard precision virtual screening (SP), and extra precision virtual screening (XP) using Schrodinger’s Glide module to select candidate molecules for flexible docking, followed by analysis of binding modes for the top five molecules selected25.
Statistical analysis
The “corrplot” package in R is utilized to analyze the correlation between m6A genes and their association with immune cells. Kruskal–Wallis’s test and chi-square test are employed to compare differences between groups. The statistical significance is set at p < 0.05. All statistical analyses are performed using R version 4.30.
Results
Differential expression of m6A regulatory factors in IPF
The differential expression levels of 26 m6A regulatory factors between non-IPF and IPF patients were analyzed using the “limma” package in R. Heatmaps and histograms were used for screening and visualization of 16 important m6A regulatory factors (CBLL1, ELAVL1, FTO, HNRNPC, HNRNPA2B1, IGFBP2, LRPPRC, METTL3, METTL16, RBMX, WTAP, YTHDC1, YTHDC2, YTHDF1, YTHDF2, and ZC3H13). We found that IGFBP2, and YTHDF2 were overexpressed in IPF patients, while other significant m6A regulatory factors showed decreased expression in IPF patients compared to non-IPF patients (Fig. 1a, b, *p < 0.05, **p < 0.01, ***p < 0.001). The chromosome positions of the 26 m6A regulatory factors were visualized using the “RCircos” package (Fig. 1c).

Landscape of N6-methyladenosine (m6A) regulators in IPF. (a) Heatmap illustrating the differential expression of 16 m6A regulators. (b) Histogram depicting the variance in expression levels of 26 m6A regulators between the healthy control group(con) and IPF patients(treat). (c) Utilize the “RCircos” package to visualize the chromosomal locations of 26 m6A regulators. *p < 0.05, ** p < 0.01, and ***p < 0.001.
The correlation analysis of the 26 m6A regulatory factors in IPF
In our analysis, we observed strong positive correlations between the expression levels of ELAVL1, LRPPRC, RBMX, METTL3, and METTL16 with FTO in IPF patients (Fig. 2a–e). We also found a positive correlation between WTAP expression and readers (HNRNPC, HNRNPA2B1, and YTHDC1, Fig. 2f–h). Moreover, when examining the correlation between the writers and readers, we discovered that the expression levels of most writers were positively correlated with the readers (Supplementary Fig. S1). These findings demonstrate diverse correlations among different m6A regulatory factors.

The correlation between m6A regulators in IPF. (a–e) The correlation between FTO and other m6A regulators (ELAVL1, LRPPRC, RBMX, METTL3, and METTL16). (f–h) The correlation between WTAP and other m6A regulators (HNRNPC, HNRNPA2B1, and YTHDC1).
The construction of random forest (RF) and support vector machine (SVM) models
We developed both RF and SVM model to identify potential candidate m6A regulatory factors for predicting the occurrence of IPF. The RF model demonstrated the smallest residuals based on the "reverse cumulative distribution of residuals" (Fig. 3a) and the "box plot of residuals" (Fig. 3b). Furthermore, we evaluated the models using ROC curves, and the AUC value for the RF model (Fig. 3c) indicated higher accuracy compared to the SVM model. The random forest map displayed in Fig. 3d revealed that the model comprising 16 m6A regulatory factors achieved the lowest error. We ranked these genes based on their importance and visualized the top five m6A regulatory factors: FTO, HNRNPA2B1, WTAP, YTHDC1, and ZC3H13 (Fig. 3e). These factors were selected as candidate genes for further investigation.

Construction of Random Forest (RF) and Support Vector Machine (SVM) models. (a) Residual boxplot of the RF and SVM models. (b) Reverse cumulative distribution plots of the RF and SVM models. (c) ROC curves for the RF and SVM models. (d) Evaluation of the importance of 16 m6A regulatory factors using the RF model. (e) Cross-validation curves of the RF model illustrating error levels for the treatment group (red line), control group (green line), and overall sample (black line).
The construction of the nomogram model
A column chart model based on the five candidate m6A regulatory factors was constructed using the “rms” package in R to predict the prevalence of IPF patients (Fig. 4a). The model’s calibration curve demonstrates that its predictive performance is accurate, aligning well with the ideal diagonal line (Fig. 4b). Furthermore, the DCA curve indicates that decisions made based on the column chart model may provide benefits for IPF patients, as demonstrated by the red line remaining consistently above the gray and black reference lines (Fig. 4c). Additionally, the clinical impact curve exhibits a significant predictive ability of the column chart model, indicating its potential practical implications (Fig. 4d).

The construction of nomogram model. (a) A Nomogram model built using 5 candidate m6A regulators. A cumulative score of 33 corresponds to a 10% prevalence rate, and a score of 70 represents a 90% prevalence rate. (b) Assessment of the nomogram model accuracy via calibration curves. (c) Decision curve analysis of the nomogram model. (d) Clinical impact curve of the nomogram model.
Identification of two distinct m6A patterns based on effective m6A regulatory factors
Consensus clustering analysis performed based on 16 significant m6A regulatory factors. The optimal clustering outcome was achieved by categorizing IPF patients into two subgroups, revealing distinct m6A patterns denoted as cluster A and cluster B. These clusters exhibited the highest intra-group correlation and lower inter-group correlation (Fig. 5a–d). More clustering analysis heatmaps can be found in Supplementary Fig. S2. m6AClusterA comprised 87 cases, whereas m6AclusterB consisted of 67 cases (Table 2). To visually represent the variation in expression levels of the 16 m6A regulatory factors between the two clusters, heat maps and histograms were generated. The expression levels of METTL3, METTL16, WTAP, ZC3H13, CBLL1, YTHDC1, YTHDC2, YTHDF1, HNRNPC, LRPPRC, HNRNPA2B1, RBMX, ELAVL1, and FTO were found to be higher in m6AclusterA compared to m6AclusterB, with IGFBP2 showing the opposite trend. YTHDF2 did not exhibit any significant difference between m6AclusterA and m6AclusterB (Fig. 5e, f, *p < 0.05, ***p < 0.001). PCA demonstrated that the 16 effective m6A regulators could completely distinguish between the two m6A patterns (Fig. 5g). Subsequently, 11 DEGs related to m6A were selected from these two m6A patterns. To understand the potential mechanisms of these DEGs in IPF, GO functional enrichment analysis was performed and the results were visualized using enrichment circle plots. The enriched GO terms primarily included immune receptor activity, cytokine binding, cytokine receptor activity, and regulation of cell growth/differentiation, indicating their involvement in immune function regulation (Fig. 5h). Furthermore, KEGG analysis revealed a significant focus on the cytokine-cytokine receptor interaction pathway, indicating its relevance to the m6A patterns observed (Fig. 5i).

Consensus clustering of 16 m6A regulatory factors in IPF. (a) Consensus clustering matrix at k = 2. (b) Consensus clustering Cumulative Distribution Function (CDF) for k values from 2 to 9. (c) Relative change in the area under the CDF curve when k = 2. (d) Item tracking plot for Consensus clustering. (e) Histogram of Cluster A and Cluster B. (f) Heatmap of Cluster A and Cluster B. (g) Principal Component Analysis based on 16 significant m6A regulatory factors showing significant differences between Cluster A and Cluster B. (h) GO enrichment analysis of m6A-related Differentially Expressed Genes (DEGs). (i) KEGG analysis of m6A-related DEGs. *p < 0.05, ** p < 0.01, and *** p < 0.001.
We utilized the ssGSEA method to estimate the abundance of immune cells in IPF samples and examined the correlation between the 16 m6A regulatory factors and immune cells. We observed that, with the exception of IGF2BP2 and YTHDF2, most m6A regulatory factors exhibited a negative correlation with Activated. dendritic. cell, Neutrophil, Macrophage, Mast. cell, Type.17. T. helper. cell, Plasmacytoid. dendritic. cell, Monocyte, Natural. killer. cell, and Activated. CD4. T. cell, while showing a positive correlation with Activated.B.cell, Activated.CD8.T.cell, and Immature.dendritic.cell (Fig. 6a, **p < 0.01, ***p < 0.001). Furthermore, we discovered that IPF patients with higher expression levels of WTAP and FTO showed increased infiltration of adaptive immune cells, such as Activated.B.cell, Activated.CD4.T.cell, Activated.CD8.T.cell, and Activated.dendritic.cell, while exhibiting decreased infiltration of innate immune cells, including Macrophage, Mast.cell, and Neutrophil, compared to IPF patients with lower expression levels of WTAP and FTO (Fig. 6b, c, *p < 0.05, **p < 0.01, ***p < 0.001). The correlation between more m6A regulatory factors and immune cells can be seen in Supplementary Fig. S3. Finally, we performed an analysis of the differential immune cell infiltration between the two m6A patterns. Our results indicated that cluster A was associated with adaptive immunity, while cluster B was associated with Type.17.T.helper.cell, Regulatory.T.cell, and innate immunity (Fig. 6d, *p < 0.05, **p < 0.01, ***p < 0.001).

Single-sample Gene Set Enrichment Analysis (GSEA) for immune infiltration. (a) The difference in immune cell infiltration between the m6Acluster A and m6Acluster B. (b) A heatmap revealing the relationship between immune cells and 16 important m6A regulatory factors. (c) The difference in immune cell infiltration between subgroups with high and low expression of WTAP. (d) The difference in immune cell infiltration between subgroups with high expression and low expression of FTO. *p < 0.05, **p < 0.01, ***p < 0.001.
Identification of two different m6A gene patterns and generation of m6A gene characteristics
In order to enhance the validation of the m6A pattern, patients with IPF were categorized into distinct genomic subtypes through consistent clustering utilizing 11 DEGs related to m6A. Our findings revealed the presence of two distinct m6A gene patterns, namely gene cluster A and geneClusterB (Table 3), which align with the grouping of m6A patterns as depicted in Fig. 7a-d. More clustering analysis heatmaps can be found in Supplementary Fig. S4. The expression levels of the 11 m6A related DEGs in geneclusters A and B are visually represented in Fig. 7e. Additionally, the analysis of Fig. 7f, g reveals a correlation between the differential expression levels of the 16 significant m6A regulatory factors and immune cell infiltration in geneclusters A and B, which aligns with the observed m6A pattern (*p < 0.05, **p < 0.01, ***p < 0.001). In order to quantitatively assess the m6A pattern, we utilized the PCA algorithm to calculate an m6A score for each sample. Subsequently, we compared the m6A scores between the two distinct m6A patterns or m6A gene patterns. The findings indicate that the m6A score was higher in cluster B or geneClusterB in comparison to cluster A or gene cluster A, as depicted in Fig. 7h, i.

Consistent clustering analysis of m6A-related genes in IPF. (a–d) Consistency matrices for cluster numbers ranging from 2 to 5. (e) Expression heatmaps of m6A-related DEGs in geneCluster A and geneCluster B. (f) Differential expression histogram in geneClusterA and geneClusterB. (g) The difference in immune cell infiltration between geneCluster A and geneCluster B. (h) The difference in m6A scores between geneCluster A and geneCluster B. (i) The difference in m6A scores between m6Acluster A and m6Acluster B. *p < 0.05, **p < 0.01, and ***p < 0.001.
The significance of m6A mode in the identification of IPF
The Sankey plot (Fig. 8a) provides a visual representation of the interplay between the m6A pattern, m6A gene pattern, and m6A score. To further elucidate the association between m6A patterns and IPF, we conducted an investigation into the correlation between m6A patterns and cytokines. The findings revealed that the expression levels of IL-27 and IL-18 in m6Acluster B or genecluster B were significantly higher than those in m6Acluster A or geneClusterA Conversely, the expression levels of IL-15, IL-6, and TNF-α in m6Acluster B or genecluster B were notably lower than those in m6Acluster A or genecluster A. These results suggest a strong correlation between m6Acluster B or genecluster B and IPF, which is characterized by a Th17 immune response (Fig. 8b, c, *p < 0.05, **p < 0.01, ***p < 0.001). We also explored the correlation between m6A patterns and EMT-related genes. In IPF, it has been established that epithelial markers are downregulated, while mesenchymal markers show a positive correlation with the severity of fibrosis. Our results revealed lower expression of epithelial markers (CTNNB1 and DSP) in m6Acluster B and genecluster B, whereas higher expression of mesenchymal markers (MMP9, ZEB1, CDH2, and CDH1) was observed (Fig. 8d, e, *p < 0.05, **p < 0.01, ***p < 0.001). These findings suggest a close association between m6Acluster B, genecluster B, and the EMT characteristics of IPF. Furthermore, our previous network pharmacology analysis indicated that Yupingfeng can effectively treat pulmonary fibrosis6. Hence, we investigated the relationship between 18 key target genes and m6A patterns. The outcomes demonstrated that in pulmonary fibrosis, TP63 and AKR1C3 exhibited low expression in m6Acluster B and genecluster B, while CYP1B1, PLAU, PTGS2, and SPP1 showed high expression in m6Acluster B and genecluster B, aligning with our earlier research findings (Fig. 8f, g, *p < 0.05, **p < 0.01, ***p < 0.001).

The role of m6A patterns and gene patterns in distinguishing IPF. (a) A Sankey diagram revealing the correlation between m6A patterns, m6A gene patterns, and m6A scores. (b) Different expression levels of cytokines between m6Acluster A and m6Acluster B. (c) Different expression levels of cytokines between geneCluster A and geneCluster B. (d) Different expression levels of epithelial markers and mesenchymal markers between m6Acluster A and m6Acluster B. (e) Different expression levels of epithelial markers and mesenchymal markers between geneCluster A and geneCluster B. (f) Differences in the expression levels of key genes for treating pulmonary fibrosis with Yùpíngfēng in m6Acluster A and m6Acluster B. (g) Differences in the expression levels of key genes for treating pulmonary fibrosis with Yùpíngfēng between geneCluster A and geneCluster B. *p < 0.05, **p < 0.01, and ***p < 0.001.
Expression of WTAP, FTO, HNRNPA2B1, ZC3H13, and α-SMA
In BLM-induced group, the RNA expression levels of WTAP, FTO, HNRNPA2B1, and ZC3H13 were significantly reduced, while the RNA expression level of α-SMA increased (Fig. 9a–e,*p < 0.05, **p < 0.01, ***p < 0.001). Western Blot analysis confirmed similar findings, demonstrating a notable decrease in WTAP protein expression in the BLM-induced group, along with a significant increase in α-SMA protein expression (Fig. 9f, *p < 0.05).

Expression of key genes in IPF model. (a–e) The mRNA expression of WTAP, FTO, HNRNPA2B1, ZC3H13, and α-SMA. (f) The protein expression of WTAP and α-SMA. *p < 0.05, **p < 0.01, and ***p < 0.001.
Virtual screening for potential drugs targeting WTAP
Predict the potential binding sites of WTAP used Schrodinger. A total of 47,696 compounds were screened from the YaTCM database, and 21 compounds were selected for molecular docking based on ADMET, HTVS, SP, and XP criteria (Supplementary Fig. S5). All these compounds exhibited affinities below -5.5 kcal/mol, as depicted in Table 4. The top 5 compounds (GA17, Chebulic acid, 4,8-Dimethoxy-7-hydroxy-2-oxo-2H-1-benzopyran-5,6-dicarboxylic acid, 7-Hydroxy eucommic acid, Saccharic acid, Fig. 10a–e were chosen as potential drugs for targeting WTAP.

Virtual screening for potential drugs targeting WTAP. (a–e) Represent the top five drugs located, respectively: GA17, Chebulic acid, 4,8-Dimethoxy-7-hydroxy-2-oxo-2H-1-benzopyran-5,6-dicarboxylic acid, 7-Hydroxy eucommic acid, Saccharic acid.
Discussion
IPF is a disease characterized by an unknown etiology, irreversible progression, and fatal consequences2. The occurrence and progression of this disease involve numerous pathological and physiological factors, most of which remain incompletely understood. Currently, drugs used in clinical practice have demonstrated efficacy against many of the pathological and physiological processes associated with IPF, particularly targeting aging, oxidative stress, and EMT2,28. As an epigenetic modification, N6 methyladenosine (m6A) plays a crucial role in various diseases29. Studies have indicated that in cases of As-IPF, AECs increase TGF-β1 secretion levels through the YTHDF1/m6A/NREP pathway, promoting the transition of fibroblasts to myofibroblasts (FMT)30. Nonetheless, the precise role of m6A in IPF remains not entirely understood. The primary aim of this study is to investigate the role of m6A in IPF, identify potential new prognostic markers, and develop novel targeted drugs. In this study, we identified the overexpression of IGFBP2 and YTHDF2 as potential therapeutic targets for idiopathic pulmonary fibrosis (IPF). Previous research has demonstrated that IGFBP2 was highly expressed in gliomas and was significantly associated with poor prognosis31. It facilitated glioma progression by inducing the polarization of M2-type macrophages31. Furthermore, IGFBP2 overexpression can activate the Wnt/β-catenin signaling pathway, expedite the epithelial-mesenchymal transition (EMT), and enhance the metastatic potential of hepatocellular carcinoma (HCC) cells32. Based on these observations, we hypothesize that IGFBP2 may play a crucial role in the pathogenesis of IPF by influencing the EMT process. Additional studies have verified that METTL14 can inhibit the activity of NOVA2 through an m6A-YTHDF2-dependent mechanism, thereby promoting the expression of genes associated with hepatic fibrosis33. YTHDF2 was capable of recognizing and binding to circIRF2 modified by N6-methyladenosine, and it can inhibit the onset and progression of hepatic fibrosis by promoting the nuclear translocation of FOXO334. In conclusion, IGFBP2 and YTHDF2, as principal regulators of m6A modification, exhibit unique mechanisms of action and therapeutic potential across various disease models. These observations indicate that IGFBP2 and YTHDF2 may serve as promising therapeutic targets for IPF. Nonetheless, a comprehensive understanding of their specific mechanisms of action in diverse disease contexts necessitates further detailed investigation. In this study, data from a public database were used. Due to the partial absence of clinical information, we were unable to conduct a comprehensive analysis of confounding variables. The missing data were mainly concentrated in variables such as gender and FVC. Future studies will implement stricter quality control measures during the data collection phase to minimize the impact of missing data on the results.
In our study, we conducted a comprehensive assessment of the impact of m6A regulatory factors on the diagnosis and subtype classification of IPF using the GSE93606 dataset. We developed a five genes model (FTO, HNRNPA2B1, WTAP, YTHDC1, and ZC3H13) for predicting IPF based on 16 candidate m6A regulatory factors. Subsequently, we established a nomogram model to predict the incidence of IPF in patients. DCA revealed that decisions based on the bar graph model could be beneficial for IPF patients. The clinical impact curve demonstrated the significant predictive capability of the bar graph model, indicating its potential practical application significance. The fat mass and obesity-associated protein (FTO) has been identified as the first RNA m6A demethylase, playing a crucial role in tumor initiation, cancer stem cell (CSC) self-renewal, microenvironment regulation, immunity, and metabolism35. Heterogeneous nuclear ribonucleoproteins A2/B1 (HNRNPA2B1) is an m6A reader that binds to newly transcribed RNA, thereby intricately influencing a variety of RNA metabolism processes36. WTAP is a conserved nuclear protein that acts as an associate of Wilms’ tumor 1 (WT1) and participates in various cellular processes including alternative splicing, X chromosome inactivation, and cell cycle regulation37. YTHDC1 is a distinct m6A reader as it holds a prominent position within the nucleus, providing it with post-transcriptional regulatory functions including pre-mRNA splicing, mRNA export, and mRNA stability38. Zinc finger CCCH type-containing 13 (ZC3H13) is an m6A-modified core methyltransferase that plays a significant role in various diseases39. While our model exhibits promising predictive capabilities in clinical settings, several limitations must be acknowledged. Firstly, the dataset utilized in this study originates from a single center. Despite ensuring the model’s robustness through rigorous internal validation, its generalizability necessitates further confirmation via multicenter external validation. Secondly, the model’s features are based on existing clinical parameters and molecular markers, which may not capture all potentially relevant factors, thereby potentially limiting its predictive performance in certain scenarios. Future research should aim to expand data sources, optimize feature selection methods, and explore integration with other emerging technologies to further enhance the generalizability of model.This study seeks to uncover the potential roles of m6A modification and m6A regulatory factors in the pathogenesis of IPF.
In our study, we utilized a consensus clustering algorithm based on 16 key m6A regulatory factors to identify two m6A patterns (m6AClusterA and m6AClusterB). m6AClusterB was found to be strongly associated with Th17 cell infiltration, innate immunity, and Treg cell infiltration, indicating a high correlation between m6AClusterB and IPF. Furthermore, we pinpointed 11 m6A-related genes based on these patterns, with GO enrichment analysis revealing their involvement in immune receptor activity, cytokine binding, cytokine receptor activity, and regulation of cell growth/differentiation. Additionally, KEGG analysis highlighted a significant focus on the cytokine-cytokine receptor interaction pathway. We also computed m6A scores for all IPF patients to quantify the m6A pattern. Results indicated that the majority of m6AClusterB samples fell into the geneClusterB group, with most cases showing high m6A-Score. The m6A-Score exhibited a positive correlation with immune cell infiltration, escalating with the advancement of pulmonary fibrosis. Patients in the high m6A-Score group demonstrated notably more severe fibrosis.
IPF is a fibrotic lung disease characterized by the destruction of alveolar structure and pulmonary interstitial fibrosis, resulting in breathing difficulties and potentially suffocation and death in patients40. EMT is recognized as a key factor in the development of IPF41. Studies have demonstrated that the levels of certain cytokines and chemokines elevate in bronchoalveolar lavage fluid (BALF) and/or lung samples of IPF patients, suggesting their potential role as pro-fibrotic agents42. Our research findings indicate that the expression of epithelial markers (CTNNB1 and DSP) is lower in m6AClusterB and geneClusterB, while the expression of mesenchymal markers (MMP9, ZEB1, CDH2, and CDH1) is higher. We also conducted an analysis of the correlation between cytokine expression and m6A patterns. The results revealed that the expression levels of IL-27 and IL-18 in m6AClusterB or geneClusterB were significantly higher than those in m6ACluster A or geneClusterA. Conversely, IL-15, IL-6, and TNF in m6AClusterB or geneClusterB showed significantly lower expression levels compared to m6AClusterA or geneClusterA. Extracellular histones promoted pulmonary fibrosis through a platelet macrophage loop involving TGF-β1 and IL-272. The upregulation of IL-18 regulated EMT through a Snail-1 dependent pathway, contributed to bleomycin (BLM)-induced pulmonary fibrosis, and enhanced its progression43. These research findings were consistent with our discoveries, further confirming the reliability of our m6A model.
Furthermore, we initially selected 4 prognostic m6A regulatory factors from the GEO dataset for further validation. In BLM group, the RNA expression levels of WTAP, FTO, HNRNPA2B1, and ZC3H13 significantly decreased, while the RNA expression of α-SMA increased. Western blot analysis confirmed similar findings, showing a substantial decrease in WTAP protein expression and a significant increase in α-SMA protein expression in BLM group. Studies have demonstrated that WTAP was associated with cardiac development, and its expression was downregulated in both humans and mice suffering from heart failure44. WTAP played a crucial role in cardiac development and function by maintaining the expression of heart-specific genes44. The downregulation of WTAP may lead to mitochondrial damage by affecting the m6A methylation of NOA145. Additionally, it activated GPX4 expression, reduces Fe2 + levels, and increased the GSH/GSSG ratio, thereby inhibiting lipid peroxidation and suppressing the occurrence and progression of hepatocellular carcinoma (HCC)45. However, the role of WTAP in pulmonary fibrotic diseases remains unclear. We speculate that WTAP may also play a crucial role in pulmonary fibrosis, but this hypothesis requires further research.
In recent years, significant progress has been made in the treatment strategies for IPF. Currently, the drugs approved by the FDA mainly include nintedanib and pirfenidone, which primarily delay disease progression by inhibiting the fibrotic process4. However, existing therapies still have limitations, such as limited efficacy and notable side effects.This study elucidated the role of WTAP in IPF, providing important evidence for the development of new therapeutic targets. WTAP regulated the EMT process through the m6A modification, thereby affecting the fibrotic process. Drugs targeting WTAP can be combined with existing anti-fibrotic drugs, with the potential to produce synergistic effects. Through virtual screening, we selected five compounds targeting WTAP as potential therapeutic agents for IPF26. These compounds predominantly interact with WTAP via various interactions, such as salt bridges, hydrogen bonds, and π–π stacking, demonstrating good binding properties and achieving high molecular docking scores46,47. Nevertheless, further validation is essential before the practical application of these potential drugs.
While WTAP has shown promise as a drug target in in vitro studies, it encounters several challenges during in vivo validation. Comprehensive functional verification of WTAP in both cellular and animal models necessitates further investigation, particularly within disease-relevant physiological and pathological contexts. Moreover, in vivo validation must account for the differential expression of WTAP across various tissues and cell types, as well as its interactions with other signaling pathways. Future research endeavors will concentrate on validating the role of WTAP in idiopathic pulmonary fibrosis (IPF) disease models, employing integrated multi-omics approaches such as transcriptomics, proteomics, and metabolomics to elucidate the molecular mechanisms underlying WTAP’s involvement in disease progression. Upon successful validation of WTAP’s efficacy and safety in animal models, subsequent preclinical and clinical studies will be essential to explore its potential as a therapeutic target. It is important to recognize that, despite the promising therapeutic prospects of targeting WTAP, it presents a double-edged sword, necessitating further research to assess its long-term safety and efficacy.
Conclusion
m6A regulatory factors play a crucial role in the occurrence and development of IPF. Using a nomogram model, we have selected 5 regulatory factors to accurately predict the incidence rate of IPF. Additionally, we have explored two different m6A patterns, m6AClusterB and geneClusterB, which have shown close associations with IPF. Furthermore, we investigated the potential of WTAP as a protective factor and therapeutic target for IPF. We have confirmed the close correlation between m6A modification and immune cell infiltration as well as the progression of IPF. This study provided new insights for the prevention, diagnosis, and targeted drug treatment of IPF.
Data availability
Open databases were analyzed in our study. The GSE93606 dataset was obtained from the GEO database (https://www.ncbi.nlm.nih.gov/geo/).
References
Guo, S. et al. Integrative analysis reveals the recurrent genetic etiologies in idiopathic pulmonary fibrosis. QJM: Monthly J. Assoc. Phys. 116, 983–992. https://doi.org/10.1093/qjmed/hcad206 (2023).
Phan, T. H. G. et al. Emerging cellular and molecular determinants of idiopathic pulmonary fibrosis. Cell. Mol. Life Sci.: CMLS 78, 2031–2057. https://doi.org/10.1007/s00018-020-03693-7 (2021).
Mei, Q., Liu, Z., Zuo, H., Yang, Z. & Qu, J. Idiopathic pulmonary fibrosis: An update on pathogenesis. Front. Pharmacol. 12, 797292. https://doi.org/10.3389/fphar.2021.797292 (2021).
Canestaro, W. J., Forrester, S. H., Raghu, G., Ho, L. & Devine, B. E. Drug treatment of idiopathic pulmonary fibrosis: Systematic review and network meta-analysis. Chest 149, 756–766. https://doi.org/10.1016/j.chest.2015.11.013 (2016).
Huang, G., Huang, S. & Cui, H. Effect of M6A regulators on diagnosis, subtype classification, prognosis and novel therapeutic target development of idiopathic pulmonary fibrosis. Front. Pharmacol. 13, 993567. https://doi.org/10.3389/fphar.2022.993567 (2022).
Chen, X. et al. YuPingFeng (YPF) upregulates caveolin-1 (CAV1) to alleviate pulmonary fibrosis through the TGF-β1/Smad2 pathway. J. Ethnopharmacol. 319, 117357. https://doi.org/10.1016/j.jep.2023.117357 (2024).
Wang, J. et al. Targeting PI3K/AKT signaling for treatment of idiopathic pulmonary fibrosis. Acta Pharmaceutica Sinica. B 12, 18–32. https://doi.org/10.1016/j.apsb.2021.07.023 (2022).
Yu, M. et al. The role of m6A demethylases in lung cancer: Diagnostic and therapeutic implications. Front. Immunol. 14, 1279735. https://doi.org/10.3389/fimmu.2023.1279735 (2023).
Wang, J. et al. Integrative analysis implicates the significance of m6A in the liver fibrosis of biliary atresia by regulating THY1. Hepatol. Commun. 7, e0004. https://doi.org/10.1097/hc9.0000000000000004 (2023).
Li, T. et al. Silencing of METTL3 attenuates cardiac fibrosis induced by myocardial infarction via inhibiting the activation of cardiac fibroblasts. FASEB J.: Off. Publ. Federation Am. Soc. Exp. Biol. 35, e21162. https://doi.org/10.1096/fj.201903169R (2021).
Bringardner, B. D., Baran, C. P., Eubank, T. D. & Marsh, C. B. The role of inflammation in the pathogenesis of idiopathic pulmonary fibrosis. Antioxid. Redox Signal. 10, 287–301. https://doi.org/10.1089/ars.2007.1897 (2008).
Guo, X., Sunil, C. & Qian, G. Obesity and the development of lung fibrosis. Front. Pharmacol. 12, 812166. https://doi.org/10.3389/fphar.2021.812166 (2021).
Fathimath Muneesa, M., Shaikh, S. B., Jeena, T. M. & Bhandary, Y. P. Inflammatory mediators in various molecular pathways involved in the development of pulmonary fibrosis. Int. Immunopharmacol. 96, 107608. https://doi.org/10.1016/j.intimp.2021.107608 (2021).
Jia, Q. et al. Circulating inflammatory cytokines and risk of idiopathic pulmonary fibrosis: A Mendelian randomization study. BMC Pulm. Med. 23, 369. https://doi.org/10.1186/s12890-023-02658-3 (2023).
Park, S. J. et al. Kurarinone attenuates BLM-induced pulmonary fibrosis via inhibiting TGF-β signaling pathways. Int. J. Mol. Sci. 22, https://doi.org/10.3390/ijms22168388 (2021).
Zheng, Y. et al. Mettl14 mediates the inflammatory response of macrophages in atherosclerosis through the NF-κB/IL-6 signaling pathway. Cell. Mol. Life Sci.: CMLS 79, 311. https://doi.org/10.1007/s00018-022-04331-0 (2022).
Tang, S. et al. N6-methyladenosine demethylase FTO regulates inflammatory cytokine secretion and tight junctions in retinal pigment epithelium cells. Clin. Immunol. (Orlando, Fla.) 241, 109080, https://doi.org/10.1016/j.clim.2022.109080 (2022).
Xi, S. et al. Downregulation of N6-methyladenosine-modified LINC00641 promotes EMT, but provides a ferroptotic vulnerability in lung cancer. Cell Death Dis. 14, 359. https://doi.org/10.1038/s41419-023-05880-3 (2023).
Li, R. et al. Pan-cancer prognostic, immunity, stemness, and anticancer drug sensitivity characterization of N6-methyladenosine RNA modification regulators in human cancers. Front. Mol. Biosci. 8, 644620. https://doi.org/10.3389/fmolb.2021.644620 (2021).
Wei, C. et al. Identification and verification of diagnostic biomarkers in recurrent pregnancy loss via machine learning algorithm and WGCNA. Front. Immunol. 14, 1241816. https://doi.org/10.3389/fimmu.2023.1241816 (2023).
Jin, W., Dong, S., Yu, C. & Luo, Q. A data-driven hybrid ensemble AI model for COVID-19 infection forecast using multiple neural networks and reinforced learning. Comput. Biol. Med. 146, 105560. https://doi.org/10.1016/j.compbiomed.2022.105560 (2022).
Hu, X., Ni, S., Zhao, K., Qian, J. & Duan, Y. Bioinformatics-led discovery of osteoarthritis biomarkers and inflammatory infiltrates. Front. Immunol. 13, 871008. https://doi.org/10.3389/fimmu.2022.871008 (2022).
Abdelhafez, O. H. et al. Metabolomics analysis and biological investigation of three Malvaceae plants. Phytochem. Anal.: PCA 31, 204–214. https://doi.org/10.1002/pca.2883 (2020).
Chen, Y. et al. A novel immune-related gene signature to identify the tumor microenvironment and prognose disease among patients with oral squamous cell carcinoma patients using ssGSEA: A bioinformatics and biological validation study. Front. Immunol. 13, 922195. https://doi.org/10.3389/fimmu.2022.922195 (2022).
Rants’o, T. A., Johan van der Westhuizen, C. & van Zyl, R. L. Optimization of covalent docking for organophosphates interaction with Anopheles acetylcholinesterase. J. Mol. Graph. Model. 110, 108054. https://doi.org/10.1016/j.jmgm.2021.108054 (2022).
Li, B. et al. YaTCM: Yet another traditional chinese medicine database for drug discovery. Comput. Struct. Biotechnol. J. 16, 600–610. https://doi.org/10.1016/j.csbj.2018.11.002 (2018).
Afanamol, M. S., Dinesh, A. D., Ali, K. S., Vengamthodi, A. & Rasheed, A. Drug repurposing by in silico prediction of cyclizine derivatives as antihyperlipemic agents. In Silico Pharmacol. 11, 27. https://doi.org/10.1007/s40203-023-00164-2 (2023).
Somogyi, V. et al. The therapy of idiopathic pulmonary fibrosis: what is next? Eur. Respir. Rev.: Off. J. Eur. Respir. Soc. 28, https://doi.org/10.1183/16000617.0021-2019 (2019).
Zhang, J. et al. Genes related to N6-methyladenosine in the diagnosis and prognosis of idiopathic pulmonary fibrosis. Front. Genet. 13, 1102422. https://doi.org/10.3389/fgene.2022.1102422 (2022).
Wang, P. et al. H3K18 lactylation promotes the progression of arsenite-related idiopathic pulmonary fibrosis via YTHDF1/m6A/NREP. J. Hazard. Mater. 461, 132582. https://doi.org/10.1016/j.jhazmat.2023.132582 (2024).
Zhang, X., Sun, X., Guo, C., Li, J. & Liang, G. Cancer-associated fibroblast-associated gene IGFBP2 promotes glioma progression through induction of M2 macrophage polarization. Am. J. Physiol. Cell Physiol. 326, C252-c268. https://doi.org/10.1152/ajpcell.00234.2023 (2024).
Chen, X. et al. IGFBP2 drives epithelial-mesenchymal transition in hepatocellular carcinoma via activating the Wnt/β-catenin pathway. Infectious Agents Cancer 18, 73. https://doi.org/10.1186/s13027-023-00543-6 (2023).
Hou, X. et al. METTL14 reverses liver fibrosis by inhibiting NOVA2 through an m6A-YTHDF2-dependent mechanism. Hepatol. Commun. 7, https://doi.org/10.1097/hc9.0000000000000199 (2023).
Chen, X. et al. N(6)-methyladenosine-modified circIRF2, identified by YTHDF2, suppresses liver fibrosis via facilitating FOXO3 nuclear translocation. Int. J. Biol. Macromol. 248, 125811. https://doi.org/10.1016/j.ijbiomac.2023.125811 (2023).
Li, Y., Su, R., Deng, X., Chen, Y. & Chen, J. FTO in cancer: functions, molecular mechanisms, and therapeutic implications. Trends Cancer 8, 598–614. https://doi.org/10.1016/j.trecan.2022.02.010 (2022).
Jiang, F. et al. HNRNPA2B1 promotes multiple myeloma progression by increasing AKT3 expression via m6A-dependent stabilization of ILF3 mRNA. J. Hematol. Oncol. 14, 54. https://doi.org/10.1186/s13045-021-01066-6 (2021).
Chen, Y. et al. WTAP facilitates progression of hepatocellular carcinoma via m6A-HuR-dependent epigenetic silencing of ETS1. Mol. Cancer 18, 127. https://doi.org/10.1186/s12943-019-1053-8 (2019).
Qiao, Y. et al. Nuclear m6A reader YTHDC1 promotes muscle stem cell activation/proliferation by regulating mRNA splicing and nuclear export. eLife 12, https://doi.org/10.7554/eLife.82703 (2023).
Huang, C. et al. ZC3H13-mediated N6-methyladenosine modification of PHF10 is impaired by fisetin which inhibits the DNA damage response in pancreatic cancer. Cancer Lett. 530, 16–28. https://doi.org/10.1016/j.canlet.2022.01.013 (2022).
Ji, J. et al. Increased expression of OPN contributes to idiopathic pulmonary fibrosis and indicates a poor prognosis. J. Transl. Med. 21, 640. https://doi.org/10.1186/s12967-023-04279-0 (2023).
Katzen, J. & Beers, M. F. Contributions of alveolar epithelial cell quality control to pulmonary fibrosis. J. Clin. Investig. 130, 5088–5099. https://doi.org/10.1172/jci139519 (2020).
Agostini, C. & Gurrieri, C. Chemokine/cytokine cocktail in idiopathic pulmonary fibrosis. Proc. Am. Thorac. Soc. 3, 357–363. https://doi.org/10.1513/pats.200601-010TK (2006).
Zhang, L. M. et al. Neutralization of IL-18 by IL-18 binding protein ameliorates bleomycin-induced pulmonary fibrosis via inhibition of epithelial-mesenchymal transition. Biochem. Biophys. Res. Commun. 508, 660–666. https://doi.org/10.1016/j.bbrc.2018.11.129 (2019).
Shi, L. et al. Downregulation of Wtap causes dilated cardiomyopathy and heart failure. J. Mol. Cell. Cardiol. 188, 38–51. https://doi.org/10.1016/j.yjmcc.2024.01.002 (2024).
Liu, S., Shang, M., Gong, J., Sun, H. & Hu, B. WTAP regulates mitochondrial damage and lipid oxidation in HCC by NOA1 mediated m6A modification. J. Cancer 16, 315–330. https://doi.org/10.7150/jca.102618 (2025).
Zhu, H. L. et al. A porous π–π stacking framework with dicopper(I) sites and adjacent proton relays for electroreduction of CO(2) to C(2+) products. J. Am. Chem. Soc. 144, 13319–13326. https://doi.org/10.1021/jacs.2c04670 (2022).
Pimviriyakul, P., Jaruwat, A., Chitnumsub, P. & Chaiyen, P. Structural insights into a flavin-dependent dehalogenase HadA explain catalysis and substrate inhibition via quadruple π-stacking. J. Biol. Chem. 297, 100952. https://doi.org/10.1016/j.jbc.2021.100952 (2021).
Funding
This work was supported by the Traditional Chinese Medicine Science and Technology Project of Shandong Province (M20240903).
Author information
Authors and Affiliations
Contributions
“GDL, JL and JQF: Writing-original draft, Visualization, Methodology, Data curation, Conceptualization. JYL: Writing-review & editing, Visualization. BZ and XC: Writing – review & editing, Funding acquisition, Conceptualization. All authors reviewed the manuscript.”
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, GD., Li, J., Fan, JQ. et al. Predictive models and WTAP targeting for idiopathic pulmonary fibrosis (IPF). Sci Rep 15, 14622 (2025). https://doi.org/10.1038/s41598-025-98490-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-98490-2
