
Abstract
This study was to explore how exosomal circRNA_43350 secreted by bone marrow MSCs (BMSCs) influences the development of chronic obstructive pulmonary disease (COPD). Pulmonary microvascular endothelial cells (PMVECs) and animal models of COPD were constructed in this study. Lung tissue morphometry was observed by hematoxylin and eosin staining. Apoptosis related indicators were measured by flow cytometry, western blot assay and TUNEL staining. The expression of circRNA_43350 and miR-342-5p was analyzed by real-time quantitative polymerase chain reaction. Dual luciferase reporter gene assay was used to investigate the relationship between circRNA_43350 and miR-342-5p. After intervention with BMSCs exosomes from normal mice, the cell apoptosis of PMVECs induced by CSE and emphysema in COPD mice were alleviated. Mmu_circRNA_43350 was highly expressed in normal BMSCs exosomes, while decreased significantly in BMSCs exosomes of COPD. After overexpression of circRNA_43350 in the CSE-induced PMVECs and COPD mice, the apoptosis-related indicators were decreased and the changes of emphysema were alleviated. Additionally, circRNA_43350 acted as a miRNA-342-5p sponge in PMVECs. Further study, we found that circRNA_43350 attenuated CSE-induced apoptosis in PMVECs by regulating the expression of miRNA-342-5p, and overexpression of miRNA-342-5p partially reversed the apoptosis-inhibiting effect of circRNA_43350. CircRNA_43350 derived from BMSCs exosomes attenuated CSE-induced apoptosis in PMVECs by regulating the expression of miRNA-342-5p.
Similar content being viewed by others

Introduction
Chronic obstructive pulmonary disease (COPD) is a common heterogeneous lung condition characterized by chronic, progressive airflow obstruction due to abnormalities in the airways and/or alveoli, resulting in bronchitis, bronchiolitis, or emphysema1,2. Cigarette smoke is the most significant single risk factor for the development of COPD. Prolonged exposure to cigarette smoke damages the structure of the airway walls, impairs the septa of the alveoli, leading to airway remodeling and changes consistent with emphysema3. Numerous studies have found that inflammation, oxidative stress, cell apoptosis, imbalance of proteases/anti-proteases, autoimmune reactions, and changes in the microbiome all contribute to the onset and progression of COPD4,5. The previous study has demonstrated that apoptosis of pulmonary microvascular endothelial cells (PMVECs) plays a significant role in the development of COPD, and apoptosis of these cells can be mitigated by specific antagonists, reversing or partially reversing the severity of emphysema in COPD mice6,7.
Our previous research find that patients with COPD have reduced stem cell numbers and impaired function compared with normal people8. Mesenchymal stem cells (MSCs) are also a type of stem cell with biological characteristics such as differentiation and regeneration. Evidence suggests that bone marrow MSCs (BMSCs) alleviate lung inflammation and damage in moderate experimental emphysema9. Furthermore, two doses of MSCs enhanced lung repair and partially reverse cardiorespiratory dysfunction in experimental emphysema10. Even though there are not differences in the baseline structure, migration, and proliferation of MSCs between COPD and controls, MSCs from COPD patients have abnormal functional capacity, like secreting more interleukin-6 and unable to reverse cardiorespiratory dysfunction11,12,13. However, it remains unknown what components or mechanisms lead to abnormal functions of COPD-derived MSCs.
MSCs has demonstrated immunomodulatory and anti-inflammatory response in COPD by secreting soluble trophic factors and extracellular vesicles13,14. Nevertheless, one research shows that extracellular vesicles derived from emphysematous donors demonstrate no amelioration in cardiorespiratory dysfunction for emphysematous mice13. Therefore, the potential effects of extracellular vesicles from COPD-MSCs should be further characterized which is important for further cell therapy. Exosomes are extracellular vesicles with diameters ranging from 30 to 200 nm and a density of 1.1 to 1.2 g/ml. They carry a plethora of bioactive substances, including specific proteins, lipids, nucleic acids, and non-coding ribonucleic acids (ncRNAs)15. NcRNAs comprise long non-coding RNAs (lncRNAs), microRNAs (miRNAs), and circular RNAs (circRNAs)16.
CircRNAs are a new research hotspot in the field of RNAs and have a variety of cellular functions. Unlike the typical splicing process of linear RNAs, circRNAs are generated through back-splicing, forming closed-loop structures via covalent bonds, devoid of free ends17. CircRNAs are abundant in various cell types such as macrophages, endothelial cells, and are present in extracellular fluids (e.g., cerebrospinal fluid, saliva, blood) as well as within exosomes18,19. They exhibit numerous biological properties, functioning as competitive endogenous RNAs that sequester miRNAs, interact with proteins, and directly regulate transcription20. For example, Qian et al.21 have discovered that circ_0044235 acts as a sponge for miR-375 and regulate interleukin-induced apoptosis and cellular inflammation.
Therefore, we propose the following hypothesis: The mechanism of impaired function of BMSCs in COPD patients may be caused by the interaction between certain circRNAs and miRNAs by BMSCs exosomes.
Materials and methods
Patient samples
The human lung tissues were obtained from patients with COPD and control patients who underwent pneumonectomy for lung nodules or tumors at the Department of Thoracic Surgery in Second Xiangya Hospital of Central South University. Patients with COPD were diagnosed according to Global Initiative for Chronic Obstructive Lung Disease 2019 (A post-bronchodilator forced expiratory volume in 1 s/forced vital capacity < 0.70)1. The study was approved by the Ethics Committee of the Second Xiangya Hospital of Central South University (approval number: 2020090, date: 07-04-2020). All participants signed informed consent. This study is performed in accordance with relevant guidelines and regulations. All methods are reported in accordance with ARRIVE guidelines.
Preparation of cigarette smoke extract (CSE)
CSE was prepared as previous research methods22. Smoke from five cigarettes (Furong, Changde Cigarette Company, China; each containing 12 mg tar, 1.1 mg nicotine and 14 mg carbon monoxide) was dissolved sequentially into 10 ml of phosphate-buffered saline (PBS) using a vacuum pump and a modified suction tube. Subsequently, the solution was filtered through a 0.22 μm pore-size filter and was adjusted power of hydrogen to obtain 100% CSE. Finally, the 100% CSE solution was diluted with PBS to achieve the required concentration for animal experiments. For cellular experiments, half a cigarette was burnt and melted into 10 ml of serum-free medium, then diluted into the medium in the proportion required for the experiment.
Mouse model of COPD and intratracheal intervention
Male C57BL/6 mice (4–6 weeks old) were purchased from Hunan Slyke Jingda Laboratory Animal corporation (Hunan, China). According to the previous method of constructing a mouse model of COPD23, the mice were randomly divided into control group and COPD group. The COPD group mice and the control group mice were injected intraperitoneally with 0.3 ml/20 g of 100% CSE or 0.3 mL/20 g of PBS on the 1st, 12th, 23rd, and 32nd days. Animals were housed in specific pathogen-free conditions on a 12-h light/dark cycle with ad libitum access to chow (Animal Laboratory Center of Hunan Provincial People’s Hospital, Changsha, China) and acidified reverse osmosis water. All experiments and protocol about animals were approved by the Institutional Animal Care and Use Committee (IACUC) of the Second Xiangya Hospital of Central South University (approval number: 2020145, date: 03-04-2020). The animal study is performed in accordance with relevant guidelines and regulations.
After anesthesia with 1% pentobarbital sodium (Sinoparm, Beijing, China) through intraperitoneal injection, mice were injected intratracheally of exosome 50 µL/each or PBS 50 µL/each on day 28 of intraperitoneal injection of CSE or PBS according to the different subgroups, and the concentration of diluted exosomes was 6.35 × 108 exosomes/50 µL. This is the same with injection of overexpression of circRNA_43350 lentivirus by intratracheal. At the end of all treatments, lung tissues were collected after general anesthesia and then the mice were killed by dislocation.
Cell isolation and culture
BMSCs from mice bone marrow were obtained according to previously described protocol24. Briefly, bone marrow was harvested from the leg bones of mice after anesthesia with 1% pentobarbital sodium. After centrifuged and resuspended, cell suspension was cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco Life Technologies, NY, USA) supplemented with 100 U/ml penicillin, 100 µg/ml streptomycin and 10% fetal bovine serum (FBS; Gibco Life Technologies, NY, USA) at 37 °C in a 5% CO2 culture chamber.
The mice primary PMVECs were isolated as previously described25. In short, lung tissue was isolated from mice after anesthesia with 1% pentobarbital sodium and digested with type I collagenase solution (Sigma, USA). Secondly, cells were passed through an LS sorting column (Mildtenny, Germany) and centrifuged to obtain PMVECs. Finally, PMVECs were cultured in endothelial cell medium (ECM; Sciencell, USA) with 5% FBS and cell growth factors at 37 °C in a 5% CO2 culture chamber.
Isolation and characterization of exosomes
BMSCs derived-exosomes were isolated by ultracentrifugation according to the previously described protocol26. Briefly, the culture medium of BMSCs were collected and centrifuged at 300 g at 4℃ for 20 min, followed by 2,000 g for 20 min and 12,000 g for 30 min. Resulting supernates were filtered by a 0.22 μm filter film (Merck Millipore, USA) and then ultracentrifuged at 120,000 g at 4℃ for 70 min. The sediments were collected and resuspended in PBS. Then, they were purified by centrifugation at 120,000 g at 4℃ for 70 min. Finally, the sediments were resuspended in 200 µL PBS and stored at -80 °C.
Exosomes were characterized by detecting the expression of its specific markers, such as CD9 (Abcam, UK) and TSG101 (Abcam, UK); its concentration was detected by Flow NanoAnalyzer (NanoFCM Inc., China), and its size distribution and morphology were detected by transmission electron microscopy (Hitachi, Japan).
Lung tissue morphometry
After fixation with 4% paraformaldehyde, the lung tissue of the mice was embedded in paraffin wax and then cut into 3.5 μm thick sections which were stained with hematoxylin and eosin (HE). Emphysematous changes were quantified by the values of mean linear intercept (MLI) and destructive index (DI) measured as previously described27. MLI was the ratio of the total length of the crosshair to the number of alveolar septa in the field of view intersecting with the crosshair at 100 × magnification. DI was calculated by dividing the defined destructive alveoli by the total number of alveoli.
Measurement of the apoptotic index
Terminal-deoxynucleotidyl transferase mediated dUTP Nick-End Labeling (TUNEL; Roche, Switzerland) was performed to estimate the apoptosis level in the lung tissue or cells following the manufacturer’s instructions. The apoptotic index (AI) was calculated as the percentage of TUNEL-positive nuclei in the lung tissue or cells at 400 × magnification.
Assessment of the apoptotic rate
After treatment and/or transfection, PMVECs were harvested, washed and resuspended in PBS. Annexin V and Propidium Iodide (PI) staining (Invitrogen, USA) was used to measure the cell apoptosis in PMVECs. According to the manufacturer’s instructions, PMVECs were incubated with 1 × binding buffer, annexin V-FITC and PI for 10 min away from light. Finally, flow cytometry (BD Bioscience, USA) was applied to quantify apoptotic cells. The apoptotic rate is calculated by the early and late cell apoptosis.
Western blot (WB) analysis
Proteins from lung tissue, cells and exosomes were extracted using RIPA lysis buffer (Beyotime, China) and protease inhibitor cocktail (Cwbio, China). The protein concentrations were determined using a BCA protein assay kit (Thermo Fisher Scientific, USA). Equal amounts of proteins were separated using SDS-polyacrylamide gel (Cwbio, China) and transferred to a polyvinylidene fluoride (PVDF) membrane (Amersham, Germany), then blocked with 5% nonfat milk (Beyotime, China) for 1 h. Primary antibodies including antibodies against TSG101 (Abcam, USA), CD9 (Abcam, USA), Bax (CST, USA), Bcl-2 (CST, USA), Cleaved caspase-3 (CST, USA), β-actin (CST, USA) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (CST, USA) were incubated at 4 °C for > 12 h. Afterwards, they were incubated with respective secondary antibodies, HRP-conjugated goat anti-rabbit IgG (Proteintech, USA) or HRP-conjugated goat anti-mouse IgG (Proteintech, USA) for 1 h. The bands were viewed using an enhanced chemiluminescence detection system (BIO-RAD, USA) and analyzed by image J software (National Institutes of Health, USA).
Real-time quantitative polymerase chain reaction (RT-qPCR)
Total RNA from exosomes, lung tissue and cells were extracted by using Trizol reagent (Invitrogen, USA) as previously described28. RNA concentration was measured by UV spectrophotometer. Subsequently, isolated RNA was converted into cDNA using a RevertAid First Strand cDNA Synthesis Kit (Thermo, USA). To detect mature miRNA, a stem-loop method was used. Random primers were used to analyze the construction of circRNAs. Then, qPCR for detecting circRNA and mRNA expression levels was performed with SYBR green master mix (Yeasen, China). U6 was used as the internal reference for miRNA-342-5p, GAPDH was used as the internal reference for circRNAs. Each PCR analysis was performed in triplicate and the fold changes of gene expression were calculated by means of relative quantification (2−△△Ct method). Sanger sequencing was used to detect the binding sites of circRNA_43350 in the RT–qPCR product. Specially designed divergent and convergent primers were designed for a RT–qPCR assay using to verify the circle construction of circRNA_43350. All of the primers (Table 1) were designed and constructed by Beijing Tsingke (Beijing, China) and RiboBio corporation (Guangzhou, China).
Cell transfection
PMVECs were cultured into 6-well plates and conducted transfection when cell confluence reached 60% to 70%. We used Lipofectamine 3000 (Invitrogen, USA) to transfer over-expression of circRNA_43350 plasmid (RiboBio, China), miR-342-5p inhibitor (RiboBio, China), over-expression of miR-342-5p plasmid (Genechem, China) and their negative controls including circRNA-NC, inhibitor-NC and miR-NC into PMVECs, respectively, according to the manufacturer’s instructions. Then, the PMVECs were treated with 2.5% CSE for 24 h. After all of the treatments, the PMVECs were collected for the next experiment.
Dual luciferase reporter gene assay
A wild-type and mut-circRNA_43350 fragment was synthesized and integrated into psiCHECK-2 dual-luciferase vector (HonorGene, China) to construct psiCHECK-2-circRNA_43350-WT with predicted binding site for miR-342-5p and psiCHECK-2-circRNA_43350-MUT with mutation sequence, respectively. Subsequently, the reporter plasmids were co-transfected into PMVECs with miR-342-5p mimics or mimic control using Lipofectamine 3000 (Invitrogen, USA). At last, the firefly and renilla luciferase activities were detected by a Dual-Glo Luciferase Assay Kit (Promega, USA).
Statistical analysis
All data from three independent duplicate tests were analyzed using PASW 18.0 software (PASW, Inc., Chicago, Illinois) and shown as mean ± standard deviation (SD). One-way ANOVA or t-tests were performed to analyze the differences between groups as appropriate. P value < 0.05 was considered statistically significant.
Results
Isolation and identification of exosomes from BMSCs
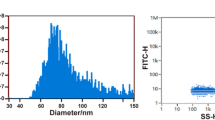
Ultracentrifugation was used to isolate exosomes from BMSCs. Electron microscopy showed that the exosomes of BMSCs were typical round shapes (Fig. 1A). Results of Nanotechnology showed that the average particle size of exosomes was 76.88 nm with a concentration of 1.59 × 109 particles/ml (Fig. 1B). WB analysis confirmed that the expression levels of CD9 and TSG101 in exosomes were higher than those in control cell lysates (Fig. 1C).

Isolation and identification of exosomes from BMSCs. (A) Transmission electron microscopy. (B) Particle size analysis and concentration detection by Flow NanoAnalyzer. (C) The expression level of CD9/TSG101 in cell lysates and exosomes by WB. The experiments were repeated three times. The original blots/gels are presented in Supplementary Fig. 1C.
BMSCs exosomes attenuate cell apoptosis and reverse the pathological changes of emphysema
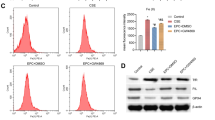
To examine whether exosomes from BMSCs can be taken in PMVECs, BMSCs exosomes was stained with PKH67 and then cultured with PMVECs, the results showed that exosomes can be taken up by PMVECs (Fig. 2A). Then, we used BMSCs exosomes from normal mice to intervene the PMVECs induced by CSE. The expression of bcl-2 was increased, while the expression of bax was decreased after exosomes intervention (Fig. 2B). To further investigate the function of exosomes in COPD mice, BMSCs exosomes from normal mice were injected intratracheally into mice. As a result, we noticed that emphysematous changes were reduced with lower MLI and DI after exosomal intervention with BMSCs by HE staining (Fig. 2C). Furthermore, the number of TUNEL-stained positive cells was significantly reduced after exosomal intervention (Fig. 2D). The WB results showed that the expression of bcl-2 was increased, while the expression of bax and cleaved caspase-3 were decreased after exosomes intervention (Fig. 2E). Collectively, these primitive results suggested that BMSCs exosomes from normal mice reduce the apoptosis of PMVECs induced by CSE stimulation, and partially reverse the pathological changes of emphysema and lung tissue apoptosis in mice with cigarette-induced COPD.

BMSCs exosomes attenuate cell apoptosis and reverse the pathological changes of emphysema in mice with COPD. (A) Internalization of PKH67 in PMVECs, DAPI staining (in blue) was performed to visualize the nuclei of PMVECs. (B) The expression of bax and bcl-2 after exosomal intervention in PMVECs induced by CSE. The experiments were repeated three times. (C) HE staining in lung tissues from control, COPD and COPD with exosomes mice (n = 10). Emphysematous changes were quantified by the values of MLI and DI, shown as mean ± SD. (D) TUNEL staining in lung tissues from different groups. (E) The expression of bax, bcl-2 and cleaved caspase-3 after exosomal intervention in different groups of mice (n = 10). The experiments were repeated three time. The original blots/gels are presented in Supplementary Fig. 2B and 2E. Error bars, SEM. ns, no significant; * P < 0.05, ** P < 0.01. (One-way ANOVA analysis for B, C and E)
Detection and identification of circRNA_43350 in COPD mice
To explore which circRNA is involved in BMSCs exosome dysfunction in COPD, we used circRNA microarray to test the circRNAs expression in exosomes of BMSCs from COPD and control mice. The results showed that a total of 13,987 circRNAs were detected. Among them, there were 14 circRNAs up-regulated and 11 down-regulated circRNAs with significant differential expression (|FC|>1.30, P < 0.05) passing volcanic map and heat map (Fig. 3A-B). Then, we selected four of the most significant different circRNAs (mmu_circRNA_32223, mmu_circRNA_37502, mmu_circRNA_43350 and mmu_circRNA_44658) for further validation analyses. Three of them (except mmu_circRNA_44658) were expressed significantly in exosomes of BMSCs from normal and COPD mice. Subsequently, results from RT-qPCR indicated that only mmu_circRNA_43350 was highly expressed in normal lung tissue and decreased in COPD lung tissue with significance (Fig. 3C-D). In addition, further searches through the CIRCpedia v2 database (http://yang-laboratory.com/circpedia/search) and the UCSC Genome Browser (https://genome.ucsc.edu/) revealed that mmu_circRNA_43350 is located on chromosome 8 with a full length of 929 bp, and the gene is derived from Ciapin1 (NM_134141.4), which is cyclised from exon 2 and exon 3 of this gene (Fig. 3E). Besides, the reverse splice site AG-TA was successfully detected by Sanger sequencing and simultaneously verified to be a cyclic RNA in BMSCs exosomes via PCR using divergent and polymeric primers (Fig. 3F-G). We detected mmu_circRNA_43350 in cDNA with divergent and convergent primers, while only in gDNA with convergent primer (Fig. 3G). Put together, these findings suggested that circRNA_43350 was down-regulated in both BMSCs exosomes and lung tissues from mice with COPD compared to normal mice.

Detection and identification of circRNA_43350. (A) All circRNAs were detected in BMSCs exosomes by circRNA microarray. (B) The circRNAs differentially expressed in BMSCs exosomes. (C) The expression of four of the most significant different circRNAs in BMSCs exosomes from control and COPD mice (n = 5). The experiments were repeated three times. (D) The expression of mmu_circRNA_37502 and mmu_circRNA_43350 in lung tissues from control and COPD mice (n = 5). The experiments were repeated four times. (E) CircRNA_43350 is located on chromosome 8 and generated from the second and third exons of Ciapin1. (F) The reverse splice site of circRNA_43350 was AG-TA detected by Sanger sequencing. (G) We detected the expression levels of the back-splice of circRNA_43350 in cDNA and gDNA from BMSCs exosomes by PCR and an agarose gel electrophoresis assay. The original gels are presented in Supplementary Fig. 3G. Error bars, SEM. ns, no significant; * P < 0.05, ** P < 0.01. (Unpaired two-tailed Student’s t-test for C and D)
CircRNA_43350 overexpression protected PMVECs and lung tissues from CSE-induced damage
To determine the function of circRNA_43350 in COPD, intervening PMVECs with 2.5%CSE was to build a COPD cell model. Overexpression plasmid of circRNA_43350 was transferred into PMVECs prior to 2.5% CSE treatment. As displayed in Fig. 4A, circRNA_43350 abundance was obviously restored via transfection of circRNA_43350 overexpression plasmid in CSE-induced cells. Moreover, circRNA_43350 overexpression attenuated CSE-induced cell apoptosis by flow cytometry and WB assay (Fig. 4B-C). Further in vivo animal experiments were performed to clarify this effect, overexpressing circRNA_43350 virus was constructed and injected into mice via endotracheal tubes to observe the effects on histopathological alterations and apoptosis in the lungs of mice with COPD. The results of RT-qPCR indicated that the overexpressing circRNA_43350 virus was successfully injected into mouse lung tissue (Fig. 4D). Furthermore, overexpression of circRNA_43350 significantly reduced emphysematous changes and decreased MLI and DI in mice with COPD (Fig. 4E). Additionally, overexpression of circRNA_43350 reduce the number of TUNEL-positive cells which were mainly in endothelial cells, decreased the expression of bax and cleaved caspase-3 and increased expression of bcl-2 (Fig. 4F-G). To sum up, these findings manifested that circRNA_43350 overexpression attenuates CSE-induced apoptosis in PMVECs, partially reverses lung histopathological changes and ameliorates lung tissue apoptosis in mice with COPD.

Overexpression of circRNA_43350 protected PMVECs and lung tissues from CSE-induced damage. (A) The expression of circRNA_43350 in PMVECs under different intervention conditions, including control group, CSE group, CSE with negative control (NC) and CSE with overexpression of circRNA_43350. The experiments were repeated three times. (B) Effect of overexpression of circRNA_43350 on CSE-induced apoptosis rate in PMVECs detected by flow cytometry. The experiments were repeated three times. (C) Effect of overexpression of circRNA_43350 on CSE-induced apoptosis-associated protein expression levels in PMVECs detected by WB. The experiments were repeated three times. (D) The expression of circRNA_43350 in lung tissues of different groups (n = 3/group). The experiments were repeated three times. (E) Overexpression of circRNA_43350 attenuates lung histopathological changes in COPD mice (n = 3/group). (F) The number of apoptotic cells in lung tissues of different groups of mice (n = 3/group) were detected by TUNEL+CD31 fluorescent staining. (G) The expression of apoptosis-related proteins in lung tissues of different groups (n = 3/group) detected by WB. The experiments were repeated three times. The original blots/gels are presented in Supplementary Fig. 4C and 4G. Error bars, SEM. ns, no significant; * P < 0.05, ** P < 0.01, *** P < 0.001. (One-way ANOVA analysis for A, B, C, D and E)
MiR-342-5p was bound to circRNA_43350 and its role in COPD
CircRNA microarray showed that 103 miRNAs have interacting targets with the circRNA_43350 sequence (Fig. 5A). Based on previous studies and review of the literature29,30, miRNA-342-5p was screened for further experiments. The binding site of miRNA-342-5p to circRNA_43350 was predicted to be CACCTC (Fig. 5B). Subsequently, dual-luciferase reporter assay showed that miRNA-342-5p mimic reduced the luciferase activity of wild-type reporter plasmid of circRNA_43350 but not the luciferase activity of mutant reporter plasmid (Fig. 5C). To explore the role of miRNA-342-5p in the development of COPD, RT-qPCR was performed to examined the expression of miRNA-342-5p in patients with COPD, mice with COPD and CSE-induced PMVECs. The results showed that miRNA-342-5p expression was increased in lung tissues of COPD and CSE-induced PMVECs (Fig. 5D). Furthermore, the expression of miRNA-342-5p was significantly reduced in PMVECs after use of miRNA-342-5p inhibitor (Fig. 5E). More importantly, by combining flow cytometry and WB assay, cell apoptosis was significantly reduced, the expression of bax and cleaved caspase-3 were significantly decreased and the expression of bcl-2 was significantly increased with miRNA-342-5p inhibitors (Fig. 5F-G). In a word, circRNA_43350 acted as a miRNA-342-5p sponge in PMVECs. Meanwhile, miRNA-342-5p depletion protected PMVECs from CSE-induced cell apoptosis.

MiR-342-5p was bound to circRNA_43350 and its role in COPD. (A) Potential target miRNAs of circRNA_43350 by RT-qPCR. (B-C) Binding sites and mutation sites of circRNA_43350 and miRNA-342-5p. (D) The expression of miRNA-342-5p in lung tissues of human subjects (non-COPD and COPD patients, n = 3/group), mice (control and COPD, n = 3/group) and in PMVECs induced by CSE. The experiments were repeated three times. (E) Expression levels of miRNA-342-5p after using miRNA-342-5p inhibitor. The experiments were repeated three times. (F) Effect of inhibition of miRNA-342-5p on CSE-induced apoptosis rate in PMVECs detected by flow cytometry. The experiments were repeated six times. (G) Effect of inhibition of miRNA-342-5p on CSE-induced apoptosis-associated protein expression levels in PMVECs detected by WB. The experiments were repeated three times. The original blots/gels are presented in Supplementary Fig. 5G. Error bars, SEM. ns, no significant; * P < 0.05, ** P < 0.01, *** P < 0.001. (One-way ANOVA analysis for D, E, F and G; Unpaired two-tailed Student’s t-test for C and D)
CircRNA_43350 modulated CSE-induced cell apoptosis by interacting with miRNA-342-5p
To clarify the regulatory relationship between circRNA_43350 and miRNA-342-5p, overexpression circRNA_43350 virus was constructed and injected into the trachea of mice. As showed in Fig. 6A, overexpression of circRNA_43350 inhibited the expression of miRNA-342-5p. For further confirmation, an overexpression circRNA_43350 plasmid and an overexpression miRNA-342-5p plasmid were constructed and transfected into CSE-intervened PMVECs. The results of RT-qPCR showed that mutual constraint existed between circRNA_43350 and miRNA-342-5p (Fig. 6B). Then, we examined how circRNA_43350 and miRNA-342-5p were involved in regulating cell apoptosis. As a result, overexpression of circRNA_43350 ameliorates apoptosis-related changes induced by CSE. Comparatively, overexpression of circRNA_43350 and overexpression of miRNA-342-5p could weaken the anti-apoptotic effect (Fig. 6C-E). Taken together, circRNA_43350 attenuated CSE-induced apoptosis in PMVECs by regulating the expression of miRNA-342-5p, and overexpression of miRNA-342-5p partially reversed the apoptosis-inhibiting effect of circRNA_43350.

CircRNA_43350 modulated CSE-induced cell apoptosis by interacting with miRNA-342-5p. (A) Effect of overexpression of circRNA_43350 on miRNA-342-5p expression in lung tissues of mice with COPD (n = 5/group). The experiments were repeated three times. (B) Expression of circRNA_43350 and miRNA-342-5p in PMVECs after CSE intervention. The experiments were repeated three times. (C) The number of TUNEL positive staining cells induced by CSE in PMVECs after transfection of circRNA_43350 overexpression and miRNA-342-5p overexpression. The experiments were repeated three times. (D) The apoptotic rate in PMVECs induced by CSE after transfection of circRNA_43350 overexpression and miRNA-342-5p overexpression. The experiments were repeated three times. (E) The expression levels of CSE-induced apoptosis-associated protein in PMVECs after transfection of circRNA_43350 overexpression and miRNA-342-5p overexpression detected by WB. The experiments were repeated three times. The original blots/gels are presented in Supplementary Fig. 6E. Error bars, SEM. ns, no significant; * P < 0.05, ** P < 0.01, *** P < 0.001. (One-way ANOVA analysis for A, B, C, D and E)
Discussion
To our knowledge, this is the first study to explore the differences of BMSCs exosomes between COPD and normal mice. In this study, the results demonstrated that there was a reduction of circRNA_43350 in BMSCs exosomes from COPD, which participated in the mechanism of CSE-induced PMVECs apoptosis by interacting with miRNA-342-5p.
Exosomes derived from MSCs have the similar function with MSCs, such as anti-inflammatory effect and protective response against the CSE-altered mitochondrial respiration parameters31,32. Ultracentrifugation is the most common method to isolate the exosomes33. In present study, exosomes were also extracted by this method and identified by electron microscopy, particle size and particle concentration detection and WB detection of exosome-associated proteins, which were consistent with the characteristics of exosomes.
In recent years, more and more studies have focused on the therapeutic role of MSCs exosomes in various diseases, such as autoimmune diseases34, cartilage defects35, cardiovascular diseases36, and inflammation of the lungs37. One study shows that human umbilical cord MSCs-derived exosomes effectively rescued the papain-induced emphysema injury and also attenuate endothelial cell apoptosis. In current study, we also found that BMSCs-derived exosomes from normal mice attenuate CSE-induced apoptosis of lung microvascular endothelial cells and partially inverse emphysematous changes in mice with COPD. Antunes et al.13 found that MSCs extracellular vesicles from COPD patients have reduced mRNA expression of anti-inflammatory, antioxidant, and immunomodulatory mediators as well as dysfunctional mitochondria, which do not yield the same therapeutic benefits as those of healthy people. In this study, there were 25 circRNAs significantly expressing in the exosomes of BMSCs from COPD and control mice. Among them, circRNA_43350 was reduced in MSCs-derived exosomes and lung tissue from COPD mice compared to control mice. It is reasonable to believe that circRNA_43350 is an important factor in the impaired exosome function of BMSCs in COPD.
CircRNAs are one type of long non-coding RNAs that can be found in exosomes38. Multiple studies have shown that circRNAs are involved in the development of COPD, but the specific mechanisms still need to be further explored39. In present study, the results revealed that CSE could inhibit the expression of circRNA_43350 in vitro cellular experiments and in vivo animal experiments. Overexpression of circRNA_43350 reduced CSE-induced apoptosis in PMVECs, and also partially reversed lung histopathological changes as well as attenuated apoptosis in lung tissues of COPD mice.
CircRNAs can perform their biological functions through a variety of mechanisms, such as acting as miRNA sponges, protein sponges and post-transcriptional regulation of gene expression. To date, most circRNAs in humans and animals have been found to be miRNA sponges40. For example, ciRS-7 has 73 binding sites for miR-7 and has a high affinity for miR-7, thereby inhibiting its activity and leading to increased expression levels of miR-7 targets41. Guo et al.42 investigated that silencing of circRNA BBS9 reduces the proportion of apoptotic cells in CSE-induced PMVECs, and up-regulation of miRNA-103a-3p inhibits CSE-induced apoptosis. Herein, we confirmed that circRNA_43350 can act as a sponge of miR-342-5p by dual luciferase reporter gene experiments. There was a mutually restraining effect between circRNA_43350 and miR-342-5p, which was that overexpression of circRNA_43350 inhibited the expression of miR-342-5p and attenuated CSE-induced apoptosis, whereas overexpression of miR-342-5p also decreased the expression of circRNA_43350 and promoted CSE-induced apoptosis. Hence, circRNA_43350 is involved in the dysfunction of BMSCs exosomes from COPD, which regulates CSE-induced PMVEC apoptosis by sponging miR-342-5p.
However, this research just explored the role of circRNA_43350 in BMSCs exosomes and did not explore the role of other components in exosomes, such as proteins. It needs to be explored further in the future. Moreover, this is only a preliminary study on the circRNA_43350/miR-342-5p pathway in COPD, and the specific mechanism has not been studied in depth. For example, whether circRNA_43350 acts on the downstream target genes of miR-342-5p to regulate apoptosis is still unknown, and this is also a subject for future research. More detailed and in-depth experiments need to be designed to explore the targeted pathways in the impaired protective function of BMSCs exosomes from COPD.
Conclusion
Our study demonstrated that BMSC-derived exosomes from normal mice can mitigate CSE-stimulated apoptosis in PMVECs, partially reversing cigarette-induced pathological changes in emphysema and the impact of pulmonary tissue apoptosis in COPD mice. Further study found that circRNA_43350 expression is downregulated in BMSC-derived exosomes from COPD mice. Decreased circRNA_43350 was involved in the CSE-induced PMVECs apoptosis by sponging miR-342-5p, which may be one of the reasons for the weakened exosomal protection of BMSCs in COPD. This study provides new insights underlying the effects and regulatory mechanisms of circRNA_43350 in the function of BMSCs exosomes from COPD and provides preclinical data supporting the potential application of circRNA_43350 overexpression as a therapeutic approach to COPD.
Data availability
The datasets generated and/or analysed during the current study are not publicly available due to keeping data security but are available from the corresponding author on reasonable request.
Abbreviations
- COPD:
-
Chronic obstructive pulmonary disease
- BMSCs:
-
Bone marrow mesenchymal stem cells
- ncRNAs:
-
Non-coding ribonucleic acids
- miRNAs:
-
MicroRNAs (miRNAs)
- circRNAs:
-
Circular RNAs
- CSE:
-
Cigarette smoke extract
- PBS:
-
Phosphate buffered saline
- PMVECs:
-
Pulmonary microvascular endothelial cells
- FBS:
-
Fetal bovine serum
- HE:
-
Hematoxylin and eosin
- MLI:
-
Mean linear intercept
- DI:
-
Destructive index
- TUNEL:
-
Terminal-deoxynucleotidyl transferase mediated dUTP Nick-End Labeling
- AI:
-
Apoptotic index
- WB:
-
Western blot
- RT-qPCR:
-
Real-time quantitative polymerase chain reaction
- GAPDH:
-
Glyceraldehyde-3-phosphate dehydrogenase
References
GE, C. Global strategy for the diagnosis, management and prevention of chronic obstructive pulmonary disease (2023 report). (2022). https://goldcopd.org/. (Accessed 29 December 2022).
Chen, L. L. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat. Rev. Mol. Cell. Biol. 21, 475–490. https://doi.org/10.1038/s41580-020-0243-y (2020).
Agusti, A. & Hogg, J. C. Update on the pathogenesis of chronic obstructive pulmonary disease. N Engl. J. Med. 381, 1248–1256. https://doi.org/10.1056/NEJMra1900475 (2019).
Barnes, P. J. et al. Chronic obstructive pulmonary disease. Nat. Rev. Dis. Primers. 1 https://doi.org/10.1038/nrdp.2015.76 (2015).
Bagdonas, E., Raudoniute, J., Bruzauskaite, I. & Aldonyte, R. Novel aspects of pathogenesis and regeneration mechanisms in COPD. Int. J. Chron. Obstruct Pulmon Dis. 10, 995–1013. https://doi.org/10.2147/COPD.S82518 (2015).
Chen, Y. et al. Protective effect of Beraprost sodium, a stable Prostacyclin analog, in the development of cigarette smoke extract-induced emphysema. Am. J. Physiol. Lung Cell. Mol. Physiol. 296, L648–656. https://doi.org/10.1152/ajplung.90270.2008 (2009).
Chen, Y. et al. Endothelin-1 receptor antagonists prevent the development of pulmonary emphysema in rats. Eur. Respir J. 35, 904–912. https://doi.org/10.1183/09031936.00003909 (2010).
Yang, Y. et al. Decreased and dysfunctional Circulating endothelial progenitor cells in patients with chronic obstructive pulmonary disease. Chin. Med. J. (Engl). 126, 3222–3227 (2013).
Antunes, M. A. et al. Effects of different mesenchymal stromal cell sources and delivery routes in experimental emphysema. Respir Res. 15, 118. https://doi.org/10.1186/s12931-014-0118-x (2014).
Poggio, H. A. et al. Impact of one versus two doses of mesenchymal stromal cells on lung and cardiovascular repair in experimental emphysema. Stem Cell. Res. Ther. 9, 296. https://doi.org/10.1186/s13287-018-1043-6 (2018).
Broekman, W. et al. Functional characterisation of bone marrow-derived mesenchymal stromal cells from COPD patients. ERJ Open. Res. 2 https://doi.org/10.1183/23120541.00045-2015 (2016).
Jahn, A. et al. Bone marrow-derived MSCs from patients with COPD have abnormal functional capacity. European Respiratory Journal 42, P250, https://doi.org/10.1183/13993003/erj.42.Suppl_57(2013).
Antunes, M. A. et al. Mesenchymal stromal cells from emphysematous donors and their extracellular vesicles are unable to reverse cardiorespiratory dysfunction in experimental severe emphysema. Front. Cell. Dev. Biol. 9, 661385. https://doi.org/10.3389/fcell.2021.661385 (2021).
Kusuma, G. D., Carthew, J., Lim, R. & Frith, J. E. Effect of the microenvironment on mesenchymal stem cell paracrine signaling: opportunities to engineer the therapeutic effect. Stem Cells Dev. 26, 617–631. https://doi.org/10.1089/scd.2016.0349 (2017).
Kalluri, R. & LeBleu, V. S. The biology, function, and biomedical applications of exosomes. Science 367 https://doi.org/10.1126/science.aau6977 (2020).
Yan, H. & Bu, P. Non-coding RNA in cancer. Essays Biochem. 65, 625–639. https://doi.org/10.1042/EBC20200032 (2021).
Jeck, W. R. et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 19, 141–157. https://doi.org/10.1261/rna.035667.112 (2013).
Vea, A., Llorente-Cortes, V. & de Gonzalo-Calvo, D. Circular RNAs in blood. Adv. Exp. Med. Biol. 1087, 119–130. https://doi.org/10.1007/978-981-13-1426-1_10 (2018).
Li, Y. et al. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell. Res. 25, 981–984. https://doi.org/10.1038/cr.2015.82 (2015).
Qu, S. et al. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 365, 141–148. https://doi.org/10.1016/j.canlet.2015.06.003 (2015).
Qian, W. et al. Circ_0044235 regulates the development of osteoarthritis by the modulation of miR-375/PIK3R3 axis. J. Orthop. Surg. Res. 19, 241. https://doi.org/10.1186/s13018-024-04694-z (2024).
Cui, Y. et al. MFG-E8 stabilized by deubiquitinase USP14 suppresses cigarette smoke-induced ferroptosis in bronchial epithelial cells. Cell. Death Dis. 14 https://doi.org/10.1038/s41419-022-05455-8 (2023).
He, Z. H. et al. Comparison between cigarette smoke-induced emphysema and cigarette smoke extract-induced emphysema. Tob. Induc. Dis. 13, 6. https://doi.org/10.1186/s12971-015-0033-z (2015).
Boregowda, S. V., Krishnappa, V. & Phinney, D. G. Isolation of mouse bone marrow mesenchymal stem cells. Methods Mol. Biol. 1416, 205–223. https://doi.org/10.1007/978-1-4939-3584-0_11 (2016).
Lin, L. et al. MiR-216a reduces apoptosis of pulmonary microvascular endothelial cells in COPD by targeting DNMT1. Tob. Induc. Dis. 21, 130. https://doi.org/10.18332/tid/171357 (2023).
Yu, L. et al. Exosomes derived from microRNA-199a-overexpressing mesenchymal stem cells inhibit glioma progression by down-regulating AGAP2. Aging (Albany NY). 11, 5300–5318. https://doi.org/10.18632/aging.102092 (2019).
Zeng, H. et al. Oxidative stress mediates the apoptosis and epigenetic modification of the Bcl-2 promoter via DNMT1 in a cigarette smoke-induced emphysema model. Respir Res. 21, 229. https://doi.org/10.1186/s12931-020-01495-w (2020).
Rio, D. C., Ares, M. Jr., Hannon, G. J. & Nilsen, T. W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb Protoc pdb prot5439, (2010). https://doi.org/10.1101/pdb.prot5439 (2010).
Long, Y. J. et al. miR-34a is involved in CSE-induced apoptosis of human pulmonary microvascular endothelial cells by targeting Notch-1 receptor protein. Respir Res. 19, 21. https://doi.org/10.1186/s12931-018-0722-2 (2018).
Wen, T. et al. miR-342-5p promotes vascular smooth muscle cell phenotypic transition through a negative-feedback regulation of Notch signaling via targeting FOXO3. Life Sci. 326, 121828. https://doi.org/10.1016/j.lfs.2023.121828 (2023).
Maremanda, K. P., Sundar, I. K. & Rahman, I. Protective role of mesenchymal stem cells and mesenchymal stem cell-derived exosomes in cigarette smoke-induced mitochondrial dysfunction in mice. Toxicol. Appl. Pharmacol. 385, 114788. https://doi.org/10.1016/j.taap.2019.114788 (2019).
Kim, S. H. et al. Exosomes derived from genetically modified DC expressing FasL are anti-inflammatory and immunosuppressive. Mol. Ther. 13, 289–300. https://doi.org/10.1016/j.ymthe.2005.09.015 (2006).
Zhu, L. et al. Isolation and characterization of exosomes for cancer research. J. Hematol. Oncol. 13, 152. https://doi.org/10.1186/s13045-020-00987-y (2020).
Shen, Z. et al. Effects of mesenchymal stem Cell-Derived exosomes on autoimmune diseases. Front. Immunol. 12, 749192. https://doi.org/10.3389/fimmu.2021.749192 (2021).
Zhang, S. et al. Mesenchymal stem cell exosomes promote functional osteochondral repair in a clinically relevant Porcine model. Am. J. Sports Med. 50, 788–800. https://doi.org/10.1177/03635465211068129 (2022).
Hade, M. D., Suire, C. N. & Suo, Z. Mesenchymal stem Cell-Derived exosomes: applications in regenerative medicine. Cells 10 https://doi.org/10.3390/cells10081959 (2021).
Harrell, C. R., Jovicic, N., Djonov, V., Arsenijevic, N. & Volarevic, V. Mesenchymal stem Cell-Derived exosomes and other extracellular vesicles as new remedies in the therapy of inflammatory diseases. Cells 8 https://doi.org/10.3390/cells8121605 (2019).
Braicu, C. et al. Exosomes as divine messengers: are they the Hermes of modern molecular oncology? Cell. Death Differ. 22, 34–45. https://doi.org/10.1038/cdd.2014.130 (2015).
Liu, X., Ali, M. K., Dua, K., Mao, Y. & Liu, J. Circular rnas: emerging players in asthma and COPD. Front. Cell. Dev. Biol. 11, 1267792. https://doi.org/10.3389/fcell.2023.1267792 (2023).
Chen, L. L. The biogenesis and emerging roles of circular RNAs. Nat. Rev. Mol. Cell. Biol. 17, 205–211. https://doi.org/10.1038/nrm.2015.32 (2016).
Hansen, T. B. et al. Natural RNA circles function as efficient MicroRNA sponges. Nature 495, 384–388. https://doi.org/10.1038/nature11993 (2013).
Guo, P., Lu, J. & Lei, Y. Significant role of circrna BBS9 in chronic obstructive pulmonary disease via miRNA-103a-3p/BCL2L13. BMC Pulm Med. 23, 257. https://doi.org/10.1186/s12890-023-02540-2 (2023).
Acknowledgements
Not applicable.
Funding
This research was supported by a grant of the National Natural Science Foundation of China (NSFC; Grant 81770046), NSFC (Grant 81970044), Xiangya Mingyi grant (2013) and the Hunan Provincial Natural Science Foundation (Grant 2024JJ6561).
Author information
Authors and Affiliations
Contributions
Conceptualization, YQZ, QS, LYY, SC, PC; Methodology, YQZ, QS, LYY, WC, CL, LL; Investigation, YQZ, LYY, WC, CL, LL; Writing, YQZ, QS, WC, CL, LL, SC, PC; Funding Acquisition, YQZ, PC; Supervision, SC, PC. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
The human lung tissues were obtained at the Department of Thoracic Surgery in Second Xiangya Hospital of Central South University. The study was approved by the Ethics Committee of the Second Xiangya Hospital of Central South University (approval number: 2020090, date: 07-04-2020) and was conducted in compliance with the Declaration of Helsinki. All participants signed informed consent. All animal experiments were conducted in accordance with the ARRIVE guidelines (Animal Research: Reporting of In Vivo Experiments) and approved by the Institutional Animal Care and Use Committee (IACUC) of the Second Xiangya Hospital of Central South University (approval number: 2020145, date: 03-04-2020).
Consent for publication
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zeng, Y., Song, Q., Yang, L. et al. BMSCs exosomes regulate pulmonary microvascular endothelial apoptosis via circRNA_43350/miR-342-5p in COPD. Sci Rep 16, 8183 (2026). https://doi.org/10.1038/s41598-026-39678-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-39678-y
