
Abstract
The flame-like reddish to orange-pigmented planctomycetal strain SH139T was isolated from brackish water obtained from Fjord Schlei in Northern Germany as part of an isolation effort led by Heinz Schlesner at Kiel University between the 1980s and early 2000s. Single gene- and whole genome-based analyses employing five established phylogenetic markers indicate that the novel isolate constitutes a novel genus and species within the family Pirellulaceae in the phylum Planctomycetota. The strain is aerobic, heterotrophic, mesophilic, and exhibits neutrophilic to slight alkaliphilic properties. Its genome size and DNA G + C content of 7.8 Mb and 55.8%, respectively, are comparable to the current closest relatives. The results of the phenotypic and phylogenetic analyses are supported by comprehensive comparative genomic data, including pangenome reconstruction and profiling of carbohydrate-active enzymes and biosynthetic gene clusters. We thus introduce Kothea flammea gen. nov., sp. nov., that is constituted by the here characterized isolate SH139T (= KCTC 102020T = DSM 116129T) as type strain. The novel genus is named after Prof. Erika Kothe (Friedrich Schiller University Jena, Germany) in recognition of her pioneering work on microbial communication.
Similar content being viewed by others

Introduction
At present, available axenic cultures encompass less than 1% of the suspected bacterial diversity1, while the remaining 99% of elusive microorganisms are classified as ‘microbial dark matter’2. Various cultivation strategies aiming at isolating fastidious bacterial strains in undersampled bacterial phyla have been developed3. However, the natural co-existence of yet untapped bacteria with complex communities of easily cultivable and rapidly growing strains often hinders enrichment and purification to yield axenic cultures of the former. Consequently, the targeted cultivation and deep-cultivation methodologies are essential for the isolation of novel, elusive microbes belonging to understudied bacterial phyla, such as Planctomycetota. Initially discovered almost a century ago, the first strain belonging to this phylum was misidentified as a floating fungus, leading to its name4. Strains of the phylum are ubiquitous and garner significant interest due to their distinctive cell biological characteristics. Prior research indicates that model strains within this phylum can import intact high molecular-weight polysaccharides, such as dextran, into an expanded periplasmic space, in which they are subsequently degraded into oligo- and monosaccharides5. This ‘selfish’ behavior, in contrast to the secretion of polysaccharide-degrading enzymes, may confer a competitive advantage in oligotrophic environments in which readily degradable sugars are scarce, and carbon must be derived from the breakdown of recalcitrant complex polysaccharides, such as those of plant or algal origin. Additional uncommon cellular characteristics include an enigmatic form of asymmetric cell division referred to as “budding” and the absence of otherwise essential bacterial cell division proteins including FtsZ, the hallmark protein for binary fission6,7,8. In silico genomic analyses of Planctomycetota strains highlight their potential to synthesize novel small molecules with prospective antimicrobial and health-promoting properties9,10. Recent investigations have further confirmed that metabolite extracts from this phylum exhibit antimicrobial activities7,11,12,13, positioning Planctomycetota as a promising, yet underexplored, reservoir of novel bioactive compounds. Till today, only a handful of compounds have been isolated from the phylum14,15,16,17,18,19.
The phylum Planctomycetota demonstrates remarkable metabolic versatility, facilitating the colonization of a broader range of ecological niches7 and the degradation of various polysaccharides20,21. Although predominantly characterized as aerobic, mesophilic, and heterotrophic, these bacteria can thrive over a spectrum of environmental conditions, such as pH, oxygen availability and temperature. Notably, members of the class “Candidatus Brocadiia” exhibit the unique ability to perform anaerobic ammonium oxidation (anammox)22,23. Strains belonging to the phylum have been documented in or isolated from a wide array of terrestrial habitats, including soils24,25,26 and peat bogs27, as well as from various aquatic environments, such as marine waters28,29,30,31,32,33, freshwater systems26,34,35,36, sediments37,38, non-natural plastic surfaces39,40 and deep-sea deposits41. Planctomycetota also exhibit adaptability to extreme environments, including hot springs42 and desert soils43. Numerous strains within this phylum have been isolated from biotic surfaces of potential symbiotic interaction partners, such as sponges23,30,44,45,46, macroalgae47,48, and also from the gastrointestinal tract49 or surface50 of other organisms, highlighting their ecological significance.
In this study, we report a novel isolate that phylogenetically affiliates with the family Pirellulaceae within the order Pirellulales. The order Pirellulales, belonging to the class Planctomycetia, is currently the best-characterized lineage within the phylum in terms of cultured representatives. With twenty described genera, Pirellulaceae represents the family with the highest number of described taxa in the entire phylum. Within the family, the current genus Rosemaritima comprises three species: R. ulvae, R. sediminicola, and R. multifibrata29,48,51. A closely related provisional genus, “Candidatus Laterigemmans baculatus”, is characterized on the basis of genomic data and exhibits a distinctive lateral budding mode of cell division52. The two genera are the closest neighbors of the here investigated novel isolate and are thus considered during comparison of analysed phenotypic and genomic features.
Materials and methods
Isolation, cultivation and initial identification of the strain
Strain SH139T was originally isolated by Heinz Schlesner (Institute for Microbiology, Christian Albrechts University, Kiel, Germany) from surface water of Fjord Schlei, an estuary of the Baltic Sea close to Kiel in Northern Germany. The strain was originally isolated using M13(3x) medium and was re-inoculated from the cryogenic stock and routinely cultivated at 21 °C in the same medium. M13(3x) medium was composed of (per litre double distilled water; pH 7.8–8.0): peptone, 0.75 g; yeast extract, 0.75 g; artificial seawater (ASW), 250 mL (see below); Tris-HCl, 5 mL (final concentration of 5 mM); Hutner’s Basal Salts, 20 mL (see below). After autoclaving, the following sterile solutions were added: D-Glucose, 3 mL (of 250 g/L stock solution); vitamin solution, 5 mL (see below). The following supplemental solutions were prepared for the medium: 1 L vitamin solution contained p-aminobenzoic acid (0.01 g), biotin (0.004 g), pyridoxine hydrochloride (0.02 g), thiamine hydrochloride (0.01 g), sodium pantothenate (0.01 g), folic acid (0.004 g), riboflavin (0.01 g), nicotinamide (0.01 g) and vitamin B12 (0.0002 g). 1 L Mineral Salt Solution (Hutner’s Basal Salts) contained nitrilotriacetic acid (10 g), MgSO4 × 7 H2O (29.7 g), CaCl2 × 2 H2O (3.34 g), FeSO4 × 7 H2O (0.099 g) and Metal Salt Solution 44 (50 mL). 1 L of Metal Salt Solution 44 contained Na2-EDTA (0.25 g), ZnSO4 × 7 H2O (1.095 g), FeSO4 × 7 H2O (0.5 g), MnSO4 x H2O (0.154 g), CuSO4 × 5 H2O (0.0395 g), CoCl2 × 6 H2O (0.0203 g), and Na2B4O7 × 10 H2O (0.0177 g). 1 L ASW contained NaCl (24 g), MgCl2 × 6 H2O (5 g), Na2SO4 (4 g), CaCl2 × (1.1 g), KCl (0.7 g), NaHCO3 (0.2 g), KBr (0.1 g), H3BO3 (0.026 g), SrCl2 (0.024 g), and NaF (0.003 g). Solidified medium was prepared from 15 g/L agar in 200 mL double distilled H2O that was autoclaved separately and added prior to pouring of the plates. The 16S rRNA gene sequence of the isolate was amplified via polymerase chain reaction (PCR), purified using the NucleoSpin Gel and PCR Clean-up kit (Machery-Nagel), and subsequently sequenced at Macrogen Europe (Amsterdam, The Netherlands).
Physiological analyses
The growth rate of strain SH139T was assessed in three biological replicates with 5 mL culture each incubated in M13(3x) medium at 21 °C under shaking conditions (100 rpm). Optical density measurements at 600 nm (OD600) were recorded every 12 h for up to two weeks using a Biochrom UV-Vis spectrophotometer. To ascertain the optimal temperature for microbial growth, 100 µL of supernatant from an exponentially growing culture, devoid of visible aggregates, was spread on agar plates. These plates were incubated in duplicates at temperatures ranging from 4 °C to 42 °C (4, 10, 18, 21, 24, 28, 32, 37, 42 °C). Daily inspections were conducted, and growth was assessed by determining the time required for the formation of visible colonies or lawn. The temperature at which colonies or lawn appeared first was identified as the optimal growth temperature. The optimal pH for growth was determined in a 96-well cultivation experiment with continuous shaking over a cultivation time of two weeks. M13(3x) medium was supplemented with 100 mM of one of the following buffering agents: 2-(N-morpholino)ethanesulfonic acid (MES) for pH 5.0 and 6.0, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) for pH 7.0, 7.5, and 8.0, or N-cyclohexyl-2-aminoethanesulfonic acid (CHES) for pH 9.0 and 10.0. Growth was quantified by measuring the OD600 in a BioTek Epoch2 microplate spectrophotometer (Agilent) at 21 °C. Each condition was tested in duplicates. Each measurement cycle lasted 30 min and was comprised of two shaking phases of 15 min interrupted by the OD600 measurements of the entire 96-well plate (Brand Plate pureGrade™ S, transparent sterile 96-well plates). The shaking regime of the 15 min interval was changed between linear to orbital to double orbital every 5 min. To mitigate condensation on the plate lid, a temperature gradient was established, maintaining the liquid at 21 °C and the lid temperature was maintained 2 °C above the cultivation temperature. For data analysis, the mean values of each time point were calculated, and the mean of the medium blank was subtracted. The growth rate at each pH was calculated from a selection of data points with the maximal slope of the natural logarithm of OD600 plotted against the cultivation time.
Light microscopy and cell size determination
Light microscopy was performed following the methodology outlined in a previous study53. In brief, cells harvested from liquid cultures at the half-maximal OD600 were mounted on a 1% (w/v) agarose cushion prepared in deionized water (dH2O). Once the culture had dried on the agarose cushion, a coverslip was applied and secured at the edges with VLAP (a 1:1:1 mixture of vaseline, lanolin, and paraffin by weight) to ensure stability. Imaging was conducted using an inverted Nikon Ti2 microscope equipped with a Nikon Plan Apo λ 100x immersion oil objective, configured with a phase ring for phase-contrast (PhC) imaging or without for differential interference contrast (DIC) imaging3. The system included a Nikon DS-Ri2 camera and NIS-Elements software (version 5.30). Three-channel RGB images were processed in FIJI54 to generate single-channel RGB images. TIFF files were subsequently analyzed in BacStalk55, with cell segmentation performed using thresholds of 25 pixels for cell size and 15 pixels for minimum cell size. A total of three biological replicates, each comprising 150 cells, were evaluated. For data visualization, results were uploaded to SuperPlotsOfData56. To enhance visualization, brightness and contrast adjustments were manually applied to PhC and DIC images.
Genomic DNA isolation, genome sequencing, annotation and analysis
Genomic DNA extraction, sequencing, assembly, and polishing were conducted following established protocols53. De novo genome assembly was performed using long-read data from Oxford Nanopore sequencing, with subsequent polishing utilizing short-read data from Illumina sequencing. For this, the sequencing reads were uploaded to the Galaxy web platform and the server available under the public domain usegalaxy.eu was used for the processing of the data using a customized workflow57. Details on the sequencing chemistry and bioinformatic workflow as well as tools, tool versions and optional parameters are provided in the Supplementary Materials in Table S1. Illumina sequencing was performed by Eurofins Genomics (Ebersberg, Germany). The genome completeness was assessed with BUSCO (version 5.8.2), while coding density and DNA G + C content were evaluated using CheckM (version 1.2.3). Following initial annotation with Prokka (version 1.14.5), the chromosome was re-oriented to the start codon of the dnaA gene, encoding the replication initiator protein, and subjected to final re-annotation using PGAP (version 2025-05-06, build 7983).
Nucleotide sequence accession numbers
The 16S rRNA gene sequence of strain SH139T has been deposited in the GenBank database under accession number PV955757. The genome sequence is available from NCBI under the accession number CP197414.
Phylogenetic and genome-based analyses
The full-length 16S rRNA gene sequence of the novel isolate was retrieved from the genome annotated with Prokka and employed to identify the closest relatives via NCBI BLAST. Maximum likelihood phylogenetic trees were constructed based on 16S rRNA gene sequences and multi-locus sequence analysis (MLSA) for the novel strain and type strains of all species in the phylum Planctomycetota. The 16S rRNA gene sequences of the type strains of Opitutus terrae (NCBI accession no. AJ229235), Kiritimatiella glycovorans (accession no. NR_146840), and Lentisphaera araneosa (accession no. NR_027571), representing strains of the Planctomycetota-Verrucomicrobiota-Chlamydiota (PVC) superphylum outside of the phylum Planctomycetota, were used as outgroup in the 16S rRNA gene sequence-based tree. The sequence alignment was conducted using ClustalW58, and the phylogenetic tree were reconstructed with FastTree v2.259 employing 1000 bootstrap replicates. The MLSA-based phylogeny was inferred using the autoMLST tool60 with 500 bootstrap replicates, including the genomes of Planctopirus limnophila DSM 3776T (GenBank acc. no. CP001744.1), Gimesia maris DSM 8797T (GenBank acc. no. CP042910.1) and Rubinisphaera brasiliensis DSM 5305T (GenBank acc. no. CP002546.1), all from the family Planctomycetaceae, as outgroup. Phylogenetic trees were visualized using iTOL v661. A 16S rRNA gene sequence similarity matrix was generated using TaxonDC62 based on the ClustalW alignment used for phylogenetic tree construction. Average amino acid identities (AAI) and average nucleotide identities (ANI) were calculated using scripts from the enveomics collection63. Additional phylogenetic markers, including sequence similarity of a 1298 bp partial sequence of the rpoB gene and percentage of conserved proteins (POCP), were determined following established methods64,65. The pangenome of selected strains was constructed using anvi’o v.8 with default parameters66. The same tool was also used for to assign the most closely related isolate or metagenome-assembled genome according to the Genome Taxonomy Database (GTDB). Biosynthetic gene clusters (BGCs) were predicted with antiSMASH v.8.067 in relaxed mode with all extra features activated, and carbohydrate-active enzymes (CAZymes) were identified using dbCAN368. The genomes of Roseimaritima spp. and “Ca. Laterigemmans sp.” were analysed in the same manner to ensure comparability of the data.
Results and discussion
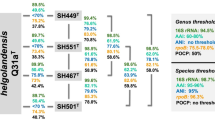
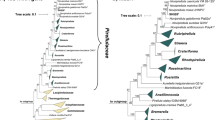
Initial nucleotide BLAST analyses of the 16S rRNA gene sequence of strain SH139T against 16S rRNA gene sequences of cultivated strains resulted in a maximal similarity of 95.4% with “Ca. Laterigemmans baculatus” CH01T and 91.1% with Roseimaritima spp. The similarity to “Ca. Laterigemmans baculatus” CH01T slightly exceeds the 94.5% threshold typically used to define a new genus, but it is significantly below this threshold when compared with Roseimaritima spp.65(Fig. 1). The provisional taxon “Ca. Laterigemmans baculatus” retains its candidatus designation because the type strain failed to survive beyond a few subcultures, and it was not successfully deposited in any culture collection. Phylogenetic analyses and the clustering pattern in both phylogenetic trees (Fig. 2), corroborated the low sequence similarity, indicating a genus-level relationship with Roseimaritima spp., but showed species-level relationship with “Ca. Laterigemmans baculatus” CH01T. Furthermore, comparisons of AAI and rpoB values between strain SH139T and Roseimaritima spp. revealed maximum similarities of 55.8% and 77.8%, respectively. These values fall below the recognized thresholds of 60–80% for AAI and in range of 75.5–78.0% for rpoB used for genus delineation64,65,69(Fig. 1). The comparison of POCP values between strain SH139T and Roseimaritima spp. revealed maximum similarities of 51.5%, which is slightly above the genus threshold value of 50%. In summary, three out of four phylogenetic markers applicable for the delineation of genera support the conclusion of the introduction of a novel genus in case that this is also supported by differences in phenotypic and genomic characteristics of strain SH139T. In the past several novel genera were reported, although they had POCP values well above the threshold of 50% when compared with their closest described relatives70. NCBI BLAST analysis of the 16S rRNA gene sequence of the novel isolate (query coverage > 90%, sequence identity > 94.5%) and (meta)genomes of close relatives listed in the GTDB revealed that uncultured close relatives have been detected in a wide variety of environments, including microbial mats from hot water lakes, hypersaline mats, and wastewater treatment plants (Table S2). These results suggest that additional strains belonging to the proposed genus are likely to be cultured from diverse environmental sources in the future.

Comparison of phylogenetic markers for genus and species delineation. Markers used: 16S rRNA gene sequence identity (16S rRNA), average amino acid identity (AAI), average nucleotide identity (ANI), sequence similarity of a partial sequence of the rpoB gene (rpoB), percentage of conserved proteins (POCP).

Phylogenetic placement. (A) Maximum likelihood phylogenetic tree based on 16S rRNA gene sequences showing the phylogenetic relationship of the novel isolate within the family Pirellulaceae. Bar, 0.1 substitutions per nucleotide position. (B) Multi-locus sequence analysis (MLSA)-based phylogenetic tree constructed with the genomes of characterized members in the family Pirellulaceae. The tree was computed based on a set of at least 30 single-copy gene-encoding proteins in a maximum likelihood approach with 500 bootstrap replications. Bar, 0.1 substitution per amino acid position. Bootstrap values for both trees are given at the nodes (in %). Phylogenetic trees were visualized with iTOL v6.
Genomic characteristics
The genome size of the novel isolate SH139T is 7.84 Mbp, which aligns with the range of genome sizes reported for Roseimaritima spp. that vary between 6.25 and 8.21 Mbp. Conversely, with 5.76 Mbp “Ca. Laterigemmans baculatus” CH01T possesses the smallest genome within this group (Table 1). Despite phylogenetic markers indicating a species-level relationship between SH139T and “Candidatus Laterigemmans baculatus” CH01T, the significant difference in their genome sizes suggests potential divergence. The DNA G + C content of strain SH139T (55.8%) lies within the range observed for Roseimaritima spp. genomes (54.5–62.4%). Consistent with other species of the genus Roseimaritima, the novel isolate SH139T is devoid of extrachromosomal elements. Its genome harbours ca. 724 protein-coding genes per Mbp, includes 1,391 genes encoding hypothetical proteins, and possesses the highest coding density (89%) and highest number of tRNA genes (96) among the five closely related strains. Like R. multifibrata FF011LT, strain SH139T possesses duplicate copies of the 5S, 16S, and 23S ribosomal RNA genes, while the other Roseimaritima spp. possess single copies.
Pangenome reconstruction and evaluation of genome-encoded functions
To visualize the genome-based similarity among the compared strains, a pangenome was constructed and analyzed. This pangenome, encompassing the genomes of all five compared species including strain SH139T, comprises 14,234 clusters, of which 1,459 are conserved across all five strains (core genome). The remaining clusters are either unique to individual strains (singletons) or not conserved across all strains. The singleton gene count for strain SH139T is 2,943. Within the pangenome visualization, beyond the core genome shown at the 9–10 o’clock segment, strain SH139T and “Ca. Laterigemmans baculatus” CH01T exhibit shared accessory genes, as evidenced at the 11 o’clock position. Correspondingly, each of the three Roseimaritima species possesses distinct supplementary genes, similarly represented at the 11 o’clock position in the pangenome visualization (Fig. 3). These findings reflect the species-level affiliation between the novel isolate SH139T and “Ca. Laterigemmans baculatus” CH01T, while confirming a more divergent evolutionary relationship with the three Roseimaritima species.

Pangenome reconstruction. Each open circle represents the pangenome of all strains but is colored darker when the gene is present in the respective genome. The heatmap in the upper right corner indicates the degree of relationship based on ANI values (ANI ≤ 70%, pale burgundy red to ANI = 100%, bright burgundy red).
Genome mining for secondary metabolite-associated biosynthetic gene clusters (BGCs) using antiSMASH identified 14 BGCs in the genome of strain SH139T (Table 1). Four of these BGCs are likely involved in the biosynthesis of terpenoids or their precursors. Other predicted BGCs include genes encoding putative type I or type III polyketide synthases (PKSs) or are linked to the synthesis of short peptides formed by non-ribosomal peptide synthetase (NRPS)-like enzymes or heterocyst glycolipid synthase-like PKSs (hglE-KS). Across all five compared strains, the identification of CAZyme-encoding genes resulted in 134–193 hits per genome, with comparable numbers of hits in the classes of glycoside hydrolases and glycosyltransferases, though these numbers were slightly higher in R. ulvae UC8T (Table 1). This is not surprising since the genome of R. ulvae UC8T is 0.5 Mbp larger than that of the strain SH139T. The counts of genes encoding polysaccharide lyases or proteins with auxiliary activities fall below ten per genome for all five genomes analyzed. This indicates that all five strains can in principle degrade complex carbohydrates into simpler sugars, however, the exact substrate spectrum needs to be determined in future cultivation experiments based on the axenic cultures and cannot be reliably predicted from the genome sequence. The genome of strain SH139T indicates its ability to produce cyanophycinase enzymes, which are essential for breaking down cyanophycin, a compound of biotechnological interest as a source of polyaspartic acid. The genome also encodes multiple genes potentially conferring resistance to heavy metals, including cobalt, zinc, and cadmium. This may further underscore the environmental importance of the novel planctomycetal isolate.
Physiological and phenotypic characterization
Strain SH139T displays a maximal growth rate of 0.017 h⁻¹ which corresponds to a doubling time of ca. 41 h (Fig. 4). Hence, the novel strain grows relatively slowly compared to most of the characterized planctomycetal isolates (typical growth rates range from 0.01 to 0.07 h− 1)71,72. In both, agar plate and liquid cultures, the novel strain displays an intense flame-like reddish-orange pigmentation, different from that of its closest relatives, “Ca. Laterigemmans baculatus” CH01T, and Roseimaritima spp. Colonies of the novel strain show either circular or irregular shapes with a more mucoid texture (Fig. 5A). Strain SH139T grows at temperatures between 18 and 28 °C, with optimal growth at 21 °C, and tolerates a narrow pH range of 7.0–8.0, with an optimum at pH 7.5 (Table 2). Like its closest relatives, it also exhibits an aerobic and heterotrophic lifestyle. On the microscopic level, cells appear pear-shaped and have a mean cell length and width of 2.1 ± 0.2 μm and 1.5 ± 0.1 μm, respectively (Fig. 5B, C). This observation renders the cells of strain SH139T smaller than those of the closest relative “Ca. Laterigemmans baculatus” CH01T, which have a length of 3.0 to 4.0 μm and a width of 0.5 to 0.8 µm52. Strain CH01T was also observed to divide via lateral budding, a form of planctomycetotal cell division in which the daughter cell is formed on the lateral side instead of the pole of the mother cell52. Along with Kolteria novifilia Pan216T, Alienimonas chondri LzC2T and some others7,73,74, strain CH01T thus shows variations from bud formation exactly at the cell pole. In contrast to this observation, we could observe cells of strain SH139T to divide via polar cell division (polar budding), the common form of cell division in the class Planctomycetia as also observed for the genus Roseimaritima. Here, the daughter cell emerges on one of the cellular poles, elongates (Fig. 5B) and subsequently pinches off from the mother cell to start a new cell cycle.

Growth curve. The graph shows a typical growth curve of strain SH139T. Data points at each time represent the average of three biological replicates. Doubling times were derived from optical density (OD600) measurements in the exponential phase. The error bars indicate the standard deviation.

Colony appearance, cell morphology and cell size of strain SH139T. Appearance of colonies on plates, and cell morphology of strain SH139T using phase contrast (PhC) and differential interference contrast (DIC) were examined. On agar plates, colonies appear big, irregular and have a slimy consistency (A). An individual cell and a cell during cell division are depicted with a larger mother and smaller daughter cell still connected to each other (B). Cell size of strain SH139T obtained from three replicates (visualized by different shades of blue), mean values of each replicate are depicted by larger circles (C). Scale bars represent 2 μm.
Conclusion
Based on the analysis of phylogenetic markers and supported by phenotypic and genomic differences, we conclude that the analyzed isolate belongs to a novel genus and species of the family Pirellulaceae. Thus, we propose the name Kothea flammea gen. nov., sp. nov., with the novel species being represented by SH139T as the type strain. Since strain SH139T does not divide by lateral budding, it was decided to assign it to a separate genus and not to assign it to the provisional genus “Ca. Laterigemmans”.
Description of Kothea gen. nov.
Ko.the’a. N.L. fem. n. Kothea, named in honor of Prof. Dr. Erika Kothe, Friedrich Schiller University Jena, for her outstanding contributions in microbial communication.
Strains belonging to the genus are heterotrophic, aerobic, mesophilic and neutrophilic. Cells divide by polar budding. Laterally budding cells were not observed. The DNA G + C content is around 56%. The genus belongs to the family Pirellulaceae, order Pirellulales, class Planctomycetia, phylum Planctomycetota. The type species of the genus is Kothea flammea.
Description of Kothea flammea sp. nov.
L. fem. adj. flammea, flame-colored, fiery red, referring to the reddish to orange pigmentation of the type strain.
Cells produce flame-like reddish-orange pigmented colonies and are pear-shaped, measuring approximately 2.1 × 1.5 μm. The type strain is SH139T (= KCTC 102020T = DSM 116129T). It was isolated from Fjord Schlei, part of the Baltic Sea in Northern Germany. The type strain grows optimally at a pH of 7.5 (range 7.0–8.0) and a temperature of 21 °C (range 18–28 °C). The doubling time of the type strain in M13(3x) medium is around 41 h. The type strain genome is 7.84 Mb in size and has a DNA G + C content of 55.8%. The type strain lacks extrachromosomal elements.
Data availability
The 16S rRNA gene sequence of strain SH139T has been deposited in the GenBank database under accession number PV955757. The genome sequence is available from NCBI under the accession number CP197414.
References
van Teeseling, M. C. F. & Jogler, C. Cultivation of elusive microbes unearthed exciting biology. Nat. Commun. 12, 75. https://doi.org/10.1038/s41467-020-20393-9 (2021).
Jiao, J. Y. et al. Microbial dark matter coming to light: challenges and opportunities. Natl. Sci. Rev. 8, nwaa280. https://doi.org/10.1093/nsr/nwaa280 (2021).
Haufschild, T. et al. An untargeted cultivation approach revealed Pseudogemmatithrix spongiicola gen. nov., sp. nov., and sheds light on the gemmatimonadotal mode of cell division: binary fission. Sci. Rep. 14, 16764. https://doi.org/10.1038/s41598-024-67408-9 (2024).
Gimesi, N. Hydrobiologiai Tanulmanyok (Hydrobiologische Studien). I: Planctomyces bekefii gim. nov. gen. et sp. Kiadja Magyar Ciszterci Rend. Budapest Hungary, 1–8 (1924).
Boedeker, C. et al. Determining the bacterial cell biology of Planctomycetes. Nat. Commun. 8, 14853. https://doi.org/10.1038/ncomms14853 (2017).
Rivas-Marin, E. et al. Non-essentiality of canonical cell division genes in the planctomycete Planctopirus limnophila. Sci. Rep. 10, 66. https://doi.org/10.1038/s41598-019-56978-8 (2020).
Wiegand, S. et al. Cultivation and functional characterization of 79 planctomycetes uncovers their unique biology. Nat. Microbiol. 5, 126–140. https://doi.org/10.1038/s41564-019-0588-1 (2020).
Jogler, C. et al. Identification of proteins likely to be involved in morphogenesis, cell division, and signal transduction in Planctomycetes by comparative genomics. J. Bacteriol. 194, 6419–6430. https://doi.org/10.1128/JB.01325-12 (2012).
Kallscheuer, N. & Jogler, C. The bacterial phylum Planctomycetes as novel source for bioactive small molecules. Biotechnol. Adv. 53, 107818. https://doi.org/10.1016/j.biotechadv.2021.107818 (2021).
Calisto, R., Godinho, O., Devos, D. P. & Lage, O. M. Genome-based in silico assessment of biosynthetic gene clusters in Planctomycetota: Evidences of its wide divergent nature. Genomics 117, 110965. https://doi.org/10.1016/j.ygeno.2024.110965 (2025).
Belova, S., Saltykova, V. & Dedysh, S. Antimicrobial activity of a novel freshwater planctomycete Lacipirellula parvula PX69T. Microbiology 89, 503–509 (2020).
Kumar, G. et al. Gemmata algarum, a Novel Planctomycete Isolated from an Algal Mat, Displays Antimicrobial Activity. Mar. Drugs. 22 https://doi.org/10.3390/md22010010 (2023).
Jeske, O., Jogler, M., Petersen, J., Sikorski, J. & Jogler, C. From genome mining to phenotypic microarrays: Planctomycetes as source for novel bioactive molecules. Antonie Van Leeuwenhoek. 104, 551–567. https://doi.org/10.1007/s10482-013-0007-1 (2013).
Panter, F. et al. Production of a Dibrominated Aromatic Secondary Metabolite by a Planctomycete Implies Complex Interaction with a Macroalgal Host. ACS Chem. Biol. 14, 2713–2719. https://doi.org/10.1021/acschembio.9b00641 (2019).
Sauer, M. et al. Investigations into the biosynthesis of stieleriacines and related N-acyl tyrosines by comparative genomics, knock-out studies and total synthesis of epi-stieleriacine C. Commun. Chem. 8, 283. https://doi.org/10.1038/s42004-025-01654-4 (2025).
Kallscheuer, N. et al. The planctomycete Stieleria maiorica Mal15T employs stieleriacines to alter the species composition in marine biofilms. Commun. Biol. 3, 303. https://doi.org/10.1038/s42003-020-0993-2 (2020).
Milke, L. et al. A type III polyketide synthase cluster in the phylum Planctomycetota is involved in alkylresorcinol biosynthesis. Appl. Microbiol. Biotechnol. 108, 239. https://doi.org/10.1007/s00253-024-13065-x (2024).
Sandargo, B. et al. Stieleriacines, N-Acyl Dehydrotyrosines From the Marine Planctomycete Stieleria neptunia sp. nov. Front. Microbiol. 11, 1408. https://doi.org/10.3389/fmicb.2020.01408 (2020).
Vitorino, I. R. et al. Alichondrichlorin, a Novel Chlorohydrin-Containing Natural Product With Tumoral Cytotoxic Activity Isolated From the Planctomycetota Bacterium Alienimonas chondri LzC2T. Microb. Biotechnol. 18, e70076 (2025). https://doi.org/10.1111/1751-7915.70076
Kündgen, M., Jogler, C. & Kallscheuer, N. Substrate utilization and secondary metabolite biosynthesis in the phylum Planctomycetota. Appl. Microbiol. Biotechnol. 109, 123. https://doi.org/10.1007/s00253-025-13514-1 (2025).
Pérez-Cruz, C. et al. Mechanisms of recalcitrant fucoidan breakdown in marine Planctomycetota. Nat. Commun. 15, 10906. https://doi.org/10.1038/s41467-024-55268-w (2024).
Kuenen, J. G. Anammox bacteria: from discovery to application. Nat. Rev. Microbiol. 6, 320–326. https://doi.org/10.1038/nrmicro1857 (2008).
Kumar, G., Radha, V., Jagadeeshwari, U. & Sasikala, C. & Venkata Ramana, C. Bacterial communities of sponges from the wetland ecosystem of Little Rann of Kutch, India with particular reference to Planctomycetes. 3 Biotech 10, 478 (2020).
Buckley, D. H., Huangyutitham, V., Nelson, T. A., Rumberger, A. & Thies, J. E. Diversity of Planctomycetes in soil in relation to soil history and environmental heterogeneity. Appl. Environ. Microbiol. 72, 4522–4531. https://doi.org/10.1128/aem.00149-06 (2006).
Kaushik, R. et al. Paludisphaera soli sp. nov., a new member of the family Isosphaeraceae isolated from high altitude soil in the Western Himalaya. Antonie Van Leeuwenhoek. 113, 1663–1674 (2020).
Lhingjakim, K. L. et al. Paludisphaera rhizosphaereae sp. nov., a new member of the family Isosphaeraceae, isolated from the rhizosphere soil of Erianthus ravennae. Antonie Van Leeuwenhoek. 115, 1073–1084 (2022).
Kulichevskaia, I. S., Pankratov, T. A. & Dedysh, S. N. [Detection of representatives of the Planctomycetes in Sphagnum peat bogs by molecular and cultivation methods]. Mikrobiologiia 75, 389–396 (2006).
Gaurav, K. et al. Phylo-taxogenomics of the genus Tautonia with descriptions of Tautonia marina sp. nov., Tautonia rosea sp. nov., and emended description of the genus. Syst. Appl. Microbiol. 44, 126229 (2021).
Kumar, D. et al. Roseimaritima sediminicola sp. nov., a new member of Planctomycetaceae isolated from Chilika lagoon. Int. J. Syst. Evol. Microbiol. 70, 2616–2623 (2020).
Kallscheuer, N. et al. Cultivation-Independent Analysis of the Bacterial Community Associated With the Calcareous Sponge Clathrina clathrus and Isolation of Poriferisphaera corsica gen. nov., sp. nov., Belonging to the Barely Studied Class Phycisphaerae in the Phylum Planctomycetes. Front. Microbiol. 11, 602250. https://doi.org/10.3389/fmicb.2020.602250 (2020).
Godinho, O. et al. Bremerella alba sp. nov., a novel planctomycete isolated from the surface of the macroalga Fucus spiralis. Syst. Appl. Microbiol. 44, 126189. https://doi.org/10.1016/j.syapm.2021.126189 (2021).
Kallscheuer, N. et al. Blastopirellula retiformator sp. nov. isolated from the shallow-sea hydrothermal vent system close to Panarea Island. Antonie Van Leeuwenhoek. 113, 1811–1822. https://doi.org/10.1007/s10482-019-01377-2 (2020).
Peeters, S. H. et al. Three marine strains constitute the novel genus and species Crateriforma conspicua in the phylum Planctomycetes. Antonie Van Leeuwenhoek. 113, 1797–1809. https://doi.org/10.1007/s10482-019-01375-4 (2020).
Kumar, G. et al. Aquisphaera insulae sp. nov., a new member in the family Isosphaeraceae, isolated from the floating island of Loktak lake and emended description of the genus Aquisphaera. Antonie Van Leeuwenhoek. 114, 1465–1477 (2021).
Kohn, T. et al. Planctopirus ephydatiae, a novel Planctomycete isolated from a freshwater sponge. Syst. Appl. Microbiol. 43, 126022. https://doi.org/10.1016/j.syapm.2019.126022 (2020).
Kallscheuer, N. et al. Analysis of bacterial communities in a municipal duck pond during a phytoplankton bloom and isolation of Anatilimnocola aggregata gen. nov., sp. nov., Lacipirellula limnantheis sp. nov. and Urbifossiella limnaea gen. nov., sp. nov. belonging to the phylum Planctomycetes. Environ. Microbiol. 23, 1379–1396. https://doi.org/10.1111/1462-2920.15341 (2021).
Lage, O. M. & Bondoso, J. Bringing Planctomycetes into pure culture. Front. Microbiol. 3, 405. https://doi.org/10.3389/fmicb.2012.00405 (2012).
Sreya, P., Gaurav, K., Ahmed, S., Sasikala, C. & Ramana, C. V. Blastopirellula sediminis sp. nov. a new member of Pirellulaceae isolated from the Andaman and Nicobar Islands. Antonie Van Leeuwenhoek. 116, 463–475 (2023).
Peeters, S. H. et al. Description of Polystyrenella longa gen. nov., sp. nov., isolated from polystyrene particles incubated in the Baltic Sea. Antonie Van Leeuwenhoek. 113, 1851–1862. https://doi.org/10.1007/s10482-020-01406-5 (2020).
Kallscheuer, N. et al. Rhodopirellula heiligendammensis sp. nov., Rhodopirellula pilleata sp. nov., and Rhodopirellula solitaria sp. nov. isolated from natural or artificial marine surfaces in Northern Germany and California, USA, and emended description of the genus Rhodopirellula. Antonie Van Leeuwenhoek. 113, 1737–1750. https://doi.org/10.1007/s10482-019-01366-5 (2020).
Storesund, J. E. & Øvreås, L. Diversity of Planctomycetes in iron-hydroxide deposits from the Arctic Mid Ocean Ridge (AMOR) and description of Bythopirellula goksoyri gen. nov., sp. nov., a novel Planctomycete from deep sea iron-hydroxide deposits. Antonie Van Leeuwenhoek. 104, 569–584. https://doi.org/10.1007/s10482-013-0019-x (2013).
Elcheninov, A. G. et al. Thermogemmata fonticola gen. nov., sp. nov., the first thermophilic planctomycete of the order Gemmatales from a Kamchatka hot spring. Syst. Appl. Microbiol. 44, 126157. https://doi.org/10.1016/j.syapm.2020.126157 (2021).
Andrew, D. R. et al. Abiotic factors shape microbial diversity in Sonoran Desert soils. Appl. Environ. Microbiol. 78, 7527–7537. https://doi.org/10.1128/aem.01459-12 (2012).
Kallscheuer, N. et al. Description of Stieleria mannarensis sp. nov., isolated from a marine sponge, and proposal to include members of the genus Roseiconus in the genus Stieleria. Antonie Van Leeuwenhoek. 118, 111. https://doi.org/10.1007/s10482-025-02106-8 (2025).
Kumar, G. et al. A genomic overview including polyphasic taxonomy of Thalassoroseus pseudoceratinae gen. nov., sp. nov. isolated from a marine sponge, Pseudoceratina sp. Antonie Van Leeuwenhoek. 115, 843–856 (2022).
Kumar, G. et al. Crateriforma spongiae sp. nov., isolated from a marine sponge and emended description of the genus Crateriforma. Antonie Van Leeuwenhoek. 114, 341–353. https://doi.org/10.1007/s10482-020-01515-1 (2021).
Lage, O. M. & Bondoso, J. Planctomycetes and macroalgae, a striking association. Front. Microbiol. 5, 267–267. https://doi.org/10.3389/fmicb.2014.00267 (2014).
Wiegand, S. et al. Analysis of Bacterial Communities on North Sea Macroalgae and Characterization of the Isolated Planctomycetes Adhaeretor mobilis gen. nov., sp. nov., Roseimaritima multifibrata sp. nov., Rosistilla ulvae sp. nov. and Rubripirellula lacrimiformis sp. nov. Microorganisms 9, 1494 (2021). https://doi.org/10.3390/microorganisms9071494
Cayrou, C., Sambe, B., Armougom, F., Raoult, D. & Drancourt, M. Molecular diversity of the Planctomycetes in the human gut microbiota in France and Senegal. Apmis 121, 1082–1090. https://doi.org/10.1111/apm.12087 (2013).
Kallscheuer, N. et al. Aureliella helgolandensis gen. nov., sp. nov., a novel Planctomycete isolated from a jellyfish at the shore of the island Helgoland. Antonie Van Leeuwenhoek. 113, 1839–1849. https://doi.org/10.1007/s10482-020-01403-8 (2020).
Bondoso, J. et al. Roseimaritima ulvae gen. nov., sp. nov. and Rubripirellula obstinata gen. nov., sp. nov. two novel planctomycetes isolated from the epiphytic community of macroalgae. Syst. Appl. Microbiol. 38, 8–15. https://doi.org/10.1016/j.syapm.2014.10.004 (2015).
Kumar, D., Kumar, G., Jagadeeshwari, U., Sasikala, C. & Ramana, C. V. Candidatus Laterigemmans baculatus gen. nov. sp. nov., the first representative of rod shaped planctomycetes with lateral budding in the family Pirellulaceae. Syst. Appl. Microbiol. 44, 126188. https://doi.org/10.1016/j.syapm.2021.126188 (2021).
Wurzbacher, C. E. et al. Planctoellipticum variicoloris gen. nov., sp. nov., a novel member of the family Planctomycetaceae isolated from wastewater of the aeration lagoon of a sugar processing plant in Northern Germany. Sci. Rep. 14, 5741. https://doi.org/10.1038/s41598-024-56373-y (2024).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods. 9, 676–682. https://doi.org/10.1038/nmeth.2019 (2012).
Hartmann, R., van Teeseling, M. C. F., Thanbichler, M., Drescher, K. & BacStalk A comprehensive and interactive image analysis software tool for bacterial cell biology. Mol. Microbiol. 114, 140–150. https://doi.org/10.1111/mmi.14501 (2020).
Goedhart, J. SuperPlotsOfData-a web app for the transparent display and quantitative comparison of continuous data from different conditions. Mol. Biol. Cell. 32, 470–474. https://doi.org/10.1091/mbc.E20-09-0583 (2021).
The Galaxy platform. for accessible, reproducible and collaborative biomedical analyses: 2022 update. Nucleic Acids Res. 50, W345–w351. https://doi.org/10.1093/nar/gkac247 (2022).
Thompson, J. D., Gibson, T. J. & Higgins, D. G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinf. (Chap. 2), Unit23. https://doi.org/10.1002/0471250953.bi0203s00 (2002).
Price, M. N., Dehal, P. S. & Arkin, A. P. FastTree: computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 26, 1641–1650. https://doi.org/10.1093/molbev/msp077 (2009).
Alanjary, M., Steinke, K. & Ziemert, N. AutoMLST: an automated web server for generating multi-locus species trees highlighting natural product potential. Nucleic Acids Res. 47, W276–w282. https://doi.org/10.1093/nar/gkz282 (2019).
Letunic, I. & Bork, P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 49, W293–w296. https://doi.org/10.1093/nar/gkab301 (2021).
Tarlachkov, S. & Starodumova, I. TaxonDC: calculating the similarity value of the 16S rRNA gene sequences of prokaryotes or ITS regions of fungi. J. Bioinform. Genom. 2017, 3–5. https://doi.org/10.18454/jbg.2017.3.5.1 (2017).
Rodriguez-R, L. M. & Konstantinidis, K. T. The enveomics collection: a toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Preprints, 2167–9843 (2016).
Bondoso, J., Harder, J. & Lage, O. M. rpoB gene as a novel molecular marker to infer phylogeny in Planctomycetales. Antonie Van Leeuwenhoek. 104, 477–488 (2013).
Qin, Q. L. et al. A proposed genus boundary for the prokaryotes based on genomic insights. J. Bacteriol. 196, 2210–2215 (2014).
Delmont, T. O. & Eren, A. M. Linking pangenomes and metagenomes: the Prochlorococcus metapangenome. PeerJ 6, e4320 (2018).
Blin, K. et al. antiSMASH 8.0: extended gene cluster detection capabilities and analyses of chemistry, enzymology, and regulation. Nucleic Acids Res. https://doi.org/10.1093/nar/gkaf334 (2025).
Zheng, J. et al. dbCAN3: automated carbohydrate-active enzyme and substrate annotation. Nucleic Acids Res. 51, W115–W121. https://doi.org/10.1093/nar/gkad328 (2023).
Kallscheuer, N. et al. Description of three bacterial strains belonging to the new genus Novipirellula gen. nov., reclassificiation of Rhodopirellula rosea and Rhodopirellula caenicola and readjustment of the genus threshold of the phylogenetic marker rpoB for Planctomycetaceae. Antonie Van Leeuwenhoek. 113, 1779–1795. https://doi.org/10.1007/s10482-019-01374-5 (2020).
Kumar, G. et al. Stratiformator vulcanicus gen. nov., sp. nov., a marine member of the family Planctomycetaceae isolated from a red biofilm in the Tyrrhenian Sea close to the volcanic island Panarea. Antonie Van Leeuwenhoek. 116, 995–1007 (2023).
Devos, D. P., Lage, O. M. & Sutcliffe, I. C. Bringing the diversity of Planctomycetes into the light: Introduction to papers from the special issue on novel taxa of Planctomycetes. Antonie Van Leeuwenhoek. 113, 1715–1726. https://doi.org/10.1007/s10482-020-01499-y (2020).
Wiegand, S., Jogler, M. & Jogler, C. On the maverick Planctomycetes. FEMS Microbiol. Rev. 42, 739–760. https://doi.org/10.1093/femsre/fuy029 (2018).
Kallscheuer, N. et al. Kolteria novifilia, a novel planctomycetotal strain from the volcanic habitat of Panarea divides by unusual lateral budding. J. Bacteriol. 207, e00337–e00324. https://doi.org/10.1128/jb.00337-24 (2025).
Vitorino, I. et al. Alienimonas chondri sp. nov., a novel planctomycete isolated from the biofilm of the red alga Chondrus crispus. Syst. Appl. Microbiol. 43, 126083. https://doi.org/10.1016/j.syapm.2020.126083 (2020).
Acknowledgements
The authors thank Vera Thiel (DSMZ, Braunschweig, Germany) and the staff from the Korean Collection for Type Cultures (KCTC) for help during deposition of the strains and the Jena School for Microbial Communication (JSMC) for continuous support. Help from Emanuel Petre and Hanan Alderzy during genomic DNA extraction, DNA quality control and library preparation is greatly acknowledged. The authors also sincerely thank Aharon Oren for checking the etymology for the names of the novel genus and species.
Funding
Open Access funding enabled and organized by Projekt DEAL. This study was funded by the German Research Foundation (DFG, Deutsche Forschungsgemeinschaft) under Project-ID 239748522 (ChemBioSys) and Germany´s Excellence Strategy under Project-ID 390713860 - EXC 2051. Funded by the Landesgraduiertenstipendium of the Free State of Thuringia awarded by the Friedrich Schiller University Jena. Funding from the Carl Zeiss Stiftung is greatly acknowledged.
Author information
Authors and Affiliations
Contributions
GK revived, identified and cultivated the strain, took care of the deposition and performed the determination of the temperature and pH range and optimum for growth and wrote the manuscript; NK performed genome annotation, analyses of phylogeny and genome-encoded features and revised the manuscript; DA performed the determination of the growth rate; JH performed genome sequencing and assembly; TH performed light microscopy and cell size determination. CJ supervised the study and contributed to data analysis. All authors have worked on the final version of the manuscript and agreed to the submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kumar, G., Kallscheuer, N., Appiah, D. et al. Characterisation of Kothea flammea gen. nov., sp. nov., a planctomycete of the family Pirellulaceae isolated from Fjord Schlei in the Baltic Sea. Sci Rep 16, 15751 (2026). https://doi.org/10.1038/s41598-026-52476-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-52476-w
