
Abstract
Temporal interference (TI) stimulation is a popular non-invasive neurostimulation technique that utilizes the following salient neural behavior: pure sinusoid (generated in off-target brain regions) appears to cause no stimulation, whereas modulated sinusoid (generated in target brain regions) does. To understand its effects and mechanisms, we examine responses of different cell types, excitatory pyramidal (Pyr) and inhibitory parvalbumin-expressing (PV) neurons, to pure and modulated sinusoids, in intact network as well as in isolation. In intact network, we present data showing that PV neurons are much less likely than Pyr neurons to exhibit TI stimulation. Remarkably, in isolation, our data shows that almost all Pyr neurons stop exhibiting TI stimulation. We conclude that TI stimulation is largely a network phenomenon. Indeed, PV neurons actively inhibit Pyr neurons in the off-target regions due to pure sinusoids (in off-target regions) generating much higher PV firing rates than modulated sinusoids in the target regions. Additionally, we use computational studies to support and extend our experimental observations.
Similar content being viewed by others
Introduction
Transcranial electrical stimulation (TES) is widely employed for stimulating target brain regions in neuroscientific studies and clinical treatments1,2,3. A particular drawback of existing TES techniques is substantial off-target stimulation. The off-target stimulation is especially high when attempting to stimulate deeper brain regions due to currents traveling a considerable distance from the electrodes to the target region. “Temporal Interference” (TI) stimulation4 is a recent advancement in the field of TES related to earlier work on interferential stimulation5,6. Rodent studies on TI stimulation4,7 showed significantly reduced off-target stimulation for targets in the deep brain. Consequently, the concept of TI stimulation has received and continues to receive substantial attention across several works4,7,8,9,10,11,12,13,14,15,16,17,18,19, including many that have exposed limitations of this technique in being able to stimulate the deep-brain without shallow effects9,12.
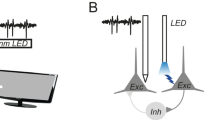
Nevertheless, TI stimulation appears to utilize the following striking neural behavior: a high-frequency sinusoidal input (pure sinusoid) does not elicit firing, but a similar amplitude high-frequency sinusoid modulated by a low-frequency sinusoid (modulated sinusoid) can cause neural firing at the modulation frequency4. TI stimulation uses carefully designed electrode configurations with sinusoidal inputs at slightly different frequencies, such that the resultant spatial interference of the sinusoidal inputs generates modulated sinusoids in the target region and pure sinusoids in regions close to the electrodes (off-target regions), aiming to elicit firing only in the target region (Fig. 1; left).

(Left) Simplified schematic of the current understanding of TI stimulation: neurons in the target region are preferentially activated by modulated sinusoids, whereas pure sinusoids do not activate neurons in the off-target regions. (Middle) Our results show that under isolation from the network, two common neuron types fire in both off-target regions (by pure sinusoids) and target regions (by modulated sinusoids), implying isolated neurons rarely exhibit TI stimulation. (Right) Simplified schematic of the revised understanding of TI: pure sinusoids make PV neurons fire at high firing rates, thereby actively inhibiting Pyr neurons from firing in the off-target region. In the target region, modulated sinusoids make PV and Pyr fire at similar firing rates, thereby causing lower inhibition in Pyr neurons and allowing Pyr neurons to fire at lower thresholds.
Despite the interest in TI stimulation, its underlying neural mechanisms remain unclear. Grossman et al.4 and related efforts20 provide useful insights into the phenomenon but do not completely explain the underlying neural mechanisms. Some computational works9,10,12,21 attempt to obtain an understanding, but our results show that the obtained understanding without rigorous experimental testing is incomplete. Understanding the neural mechanisms underlying TI stimulation is relevant as these mechanisms can help optimize TI stimulation parameters9,20 and can lead to new stimulation techniques that harness the underlying neural dynamics more effectively than TI stimulation to reduce off-target stimulation further.
In this work, we focus our attention on understanding the neural mechanisms of supra-threshold TI stimulation by analyzing the neural responses produced by pure and modulated sinusoids. Thus, we use the following definition for the exhibition of TI stimulation: a neuron exhibits TI stimulation if a current amplitude (defined as its peak value) exists for which the neuron responds to a modulated sinusoid but not to a pure sinusoid. Hence, the activation threshold of the neuron is lower with a modulated sinusoid than with a pure sinusoid. This simplistic definition was used earlier in Cao & Grover9 and is further discussed in the Results section. Note that the spatial interference of sinusoids leads to complex phenomena, such as field vector rotation21, of which the pure and modulated sinusoids form the two extremes. We use computational studies to extrapolate the insights gained from our experimental studies to the general case of the spatial interference of sinusoids.
Main contributions: Using extracellular stimulation and intracellular whole-cell patch-clamp recordings of an intact brain network (in an ex vivo setting), we observe that the exhibition of TI stimulation in a neuron depends on its type. Regular-spiking pyramidal (Pyr) neurons (excitatory) frequently exhibit TI stimulation, whereas fast-spiking parvalbumin (PV) neurons (inhibitory) rarely exhibit TI stimulation. Furthermore, a statistical comparison of the PV and Pyr stimulation thresholds reveals: a) PV neurons’ stimulation thresholds for both pure and modulated sinusoids tend to be lower than Pyr neurons’ thresholds with modulated sinusoids; b) PV and Pyr neurons have similar firing rates with modulated sinusoids which is much smaller than PV neurons’ firing rates with pure sinusoids. Together, these observations have important implications in revising the current understanding of TI stimulation (Fig. 1; right):
-
1.
In off-target regions, while existing understanding suggests no stimulation, our results suggest that PV neurons are expected to have high firing rates.
-
2.
In target regions, in agreement with existing understanding, Pyr and PV neurons are expected to be stimulated at similar firing rates.
To further understand the difference in the exhibition of TI stimulation among different cell types, we performed additional experiments where we used synaptic blockers to examine the responses of isolated neurons (Fig. 1; middle). Surprisingly, all but 2 of the 10 Pyr neurons that did exhibit TI stimulation in network settings stopped exhibiting TI stimulation in isolation. This result contradicts the dominant thought in the literature4,12 that isolated single neurons exhibit TI stimulation by simply performing “envelope-demodulation”. Our results provide strong evidence that the exhibition of TI stimulation in Pyr neurons is primarily due to network effects. Our experimental results suggest the following hypothesis for Pyr neurons exhibiting TI stimulation: PV neurons fire at a high spike rate for pure sinusoids, suggesting that Pyr neurons’ excitation is actively inhibited by PV neurons in the off-target region. For modulated sinusoidal inputs, PV neurons fire at a lower spiking rate, causing less inhibition, thereby allowing Pyr neurons to fire in the target region.
We further test our hypothesis of increased inhibition of Pyr neurons in off-target regions with computational studies. It is well-known that the spiking behavior of neurons changes according to the spatial geometry of the applied electric field22. Consequently, we simulate the firing rates of PV and Pyr neurons using biophysically realistic neuron models under different applied extracellular electric fields. We observe that the PV firing rates continue to be significantly larger for pure sinusoids than modulated sinusoids (similar to our experimental results for isolated neurons) for various electric fields observed in transcranial stimulation. One question still remains: why 2 cells of 10 continue to exhibit TI stimulation in isolation. We address this question using a reductionist approach. Through simplified computational models, we observe that the speed of a feedback control mechanism—an integral control mechanism23 present in the dynamics of neurons—explains this behavior.
Results
Before describing our results, we formally state our definition of a neuron exhibiting TI stimulation for a particular base frequency f and modulation frequency Δf. We say that a neuron exhibits TI stimulation if there exists a current amplitude A that satisfies the following two conditions: 1) The modulated sinusoid \(A\cos (\pi \Delta ft)\sin (2\pi (\, f+\Delta f/2)t)\) (which is equal to the sum of the two sinusoids, namely, \(1/2A\sin (2\pi ft)+1/2A\sin (2\pi (\,f+\Delta f)t)\)) causes the neuron to spike consistently; 2) The pure sinusoid \(A\sin (2\pi ft)\) does not cause consistent spiking (for the same A). Note that the maximum amplitude of the two signals (namely, A) is the same. A higher stimulation threshold in the off-target region (i.e., under pure sinusoidal stimulation) is desirable to reduce off-target stimulation. Our definition of TI exhibition reflects this property of a higher off-target stimulation threshold. However, the specific ratio of amplitudes between the target and off-target regions that cause off-target stimulation (with pure sinusoids) will depend on electrode configuration and locations, head size, and other factors. A neuron is said to spike consistently in response to a given sinusoid when spikes are observed throughout the stimulation duration across multiple trials.
Heterogeneous cell type response to TI stimulation
To investigate the effect of TI stimulation on different neuron types, we performed whole-cell patch recordings from cortical layer 2–3 (L2-3) Pyr and PV neurons in mouse brain slices delivering pure and modulated sinusoids with an external stimulation probe (Fig. 2a). We approximate the effect of TI stimulation in the target region as stimulation of neurons by modulated sinusoids. Correspondingly, we approximated the effect of TI stimulation in the off-target region as stimulation of the neurons by pure sinusoids. The evoked firing responses for both neuron types were carefully examined for modulated and pure sinusoidal inputs at two different base frequencies, f = 2kHz, and f = 4kHz, maintaining the modulation frequency Δf at 20 Hz.

a Schematic of the experimental set up (from left to right): cell-types recorded, recording electrode and bipolar stimulation probe in cortical L2-3 of mouse coronal brain slices, along with applied waveforms. b Variation in responses and whole-cell patch clamp representative recordings from a L2-3 Pyr neuron. (i) Percentage of L2-3 Pyr neurons that exhibit TI stimulation. (ii) Regular-spiking firing pattern in response to intracellular current injection and morphology of a L2-3 Pyr neuron. (iii) Consistent firing response to an extracellular modulated sinusoid at 90 μA amplitude, base frequency of 2 kHz and modulation frequency of 20Hz. (iv) For the same amplitude (90 μA) and base frequency (2 kHz), no consistent firing response to an extracellular pure sinusoid. c Variation in responses and whole-cell patch clamp representative recordings from a L2-3 PV neuron. (i) Percentage of L2-3 PV neurons that exhibit TI stimulation. (ii) Fast-spiking firing pattern in response to intracellular current injection and morphology of a L2-3 PV neuron. (iii) Consistent firing response to an extracellular modulated sinusoid at 40 μA amplitude, base frequency of 2 kHz and modulation frequency of 20Hz. (iv) For the same amplitude (40 μA) and base frequency (2 kHz), consistent firing response to an extracellular pure sinusoid.
Our experimental results show a striking difference in the exhibition of TI stimulation between the two neuron types: 73% ± 13%(n = 30) of the L2-3 Pyr neurons exhibited TI stimulation, while only 24% ± 14%(n = 25) of the L2-3 PV neurons exhibited TI stimulation. For this analysis, we considered a neuron to exhibit TI stimulation if it exhibited TI stimulation for any of the two base frequencies. Representative responses of Pyr and PV neurons exhibiting and not exhibiting TI stimulation are shown in Fig. 2b and c, respectively.
We observe that Pyr neurons consistently fired for the modulated sinusoid, with roughly one spike at each beat (see Fig. 2b iii; Firing Rate (FR): 17.74 ± 2.34Hz; Inter-Spike Interval (ISI): 0.06 ± 0.01s), while it did not consistently fire for the pure sinusoid (see Fig. 2b iv; FR: 5.61 ± 3.60Hz; ISI: 0.20 ± 0.18s). While Pyr neurons mostly did not consistently fire for pure sinusoids, we often observed a spike at the beginning of the applied waveform. As reported in Grossman et al.4 (and also observed in neural conduction block24,25,26), bursts of spikes at the beginning and the end of the stimulation are common, caused by sudden changes in the waveform. In contrast, PV neurons consistently fired for both modulated and pure sinusoids (see Figs. 2c iii and iv; FR: 21.05 ± 11.03Hz; ISI: 0.05 ± 0.02s for modulated sinusoids, and FR: 54.95 ± 23.94Hz; ISI: 0.03 ± 0.02s for pure sinusoids). Additional representative electrophysiological traces of both neuron types exhibiting and not exhibiting TI stimulation are provided in Supplementary Information (SI) Figs. 1 and 2.
Of note is the difference in the FR and ISI for the PV neurons: the modulated sinusoid has a lower FR and higher ISI than the pure sinusoid. In addition, PV and Pyr neurons share similar FR and ISI values for the modulated sinusoid, suggesting synchronized firing from both neuron types to the modulated sinusoid. We study these differences more deeply in the next subsection.
Differences in firing rates, inter-spike interval, and activation thresholds of PV and Pyr neurons under TI stimulation
We examine the activation thresholds, FR, and ISI for L2-3 Pyr and PV with pure and modulated sinusoids for each base frequency (Fig. 3; additional data on current densities is provided in SI Fig. 3). We refer to activation thresholds with pure or modulated sinusoids as the minimum amplitude required for each neuron to consistently spike with the respective sinusoidal input. In comparing activation thresholds, we normalize the threshold amplitudes to account for differences in the distance of the stimulating electrodes from the stimulation site in our brain-slice experiments. Specifically, we emulate the amplitudes each neuron would receive if the distance between the recorded neuron and the external stimulation probe was fixed to a nominal 100μm. Lastly, in Fig. 3 a and b ii-iv, data for L2-3 Pyr neurons with pure sinusoids are not shown since most of these neurons did not spike with this waveform at any of the tested amplitudes.

a Examination of (i) TI exhibition, (ii) activation thresholds, (iii) FR, and (iv) ISI of L2-3 Pyr and PV neurons at 2 kHz base frequency. b Examination of (i) TI exhibition, (ii) activation thresholds, (iii) FR, and (iv) ISI of L2-3 Pyr and PV neurons at 4 kHz base frequency. Red cross indicates the datapoints in the dataset that are outliers. Asterisk indicates statistical significance (p < 0.05).
We first discuss the results for the 2 kHz base frequency. Fig. 3a i shows that the TI exhibition percentages for Pyr and PV neurons for each base frequency remain similar to the overall percentages reported in the previous subsection. Furthermore, the average normalized activation thresholds of pure and modulated sinusoids are similar for the PV neurons and are ~ 36% of the average Pyr normalized activation threshold with modulated sinusoids, as shown in Fig. 3a ii. These differences between PV and Pyr activation thresholds are statistically significant (see SI Table 1). The normalized activation threshold differences indicate that when Pyr neurons are consistently firing in the target region, PV neurons in off-target and target regions are also consistently firing.
We further observe a distinct difference in the FR and ISI of PV neurons under pure and modulated sinusoidal stimulation. Figs. 3aiii and iv show that the average FR and ISI for PV neurons with pure sinusoids is ~ 190% and ~ 75% of the average FR and ISI for PV neurons with modulated sinusoids, respectively. Thus, PV neurons fire at a higher firing rate in the off-target region with pure sinusoids, than in the target region with modulated sinusoids. Furthermore, PV and Pyr neurons in the target region have similar average FR and ISI (SI Table 2). In Fig. 3b, we observe that at 4 kHz base frequency, the qualitative observations for 2 kHz continue to hold: PV neurons have a lower activation threshold than Pyr neurons, and PV neurons have a significantly higher FR and lower ISI for the pure sinusoid in comparison to the modulated sinusoid.
Understanding contributions of the network using synaptic blockers and inferred mechanisms of TI stimulation
Why do Pyr and PV neurons respond differently to TI stimulation? Broadly, there are two potential contributing factors: intrinsic dynamics of each neuron’s membrane potential in response to the applied current and the synaptic effects of the surrounding network. To better understand the effect of the network, we isolate the cell from its network by blocking glutamatergic (excitatory), and GABAergic (inhibitory) synaptic transmissions using synaptic blockers (namely, CNQX, D-AP5, and Gabazine that will block AMPA, NMDA, and GABAA receptors, respectively). The same steps specified in Fig. 2a were followed. We examined the evoked firing responses for each cell before and after synaptic blocking, but only at a fixed base frequency f = 2 kHz and a modulation frequency Δf = 20Hz.
In the intact network, before synaptic blockers’ intervention, 70% ± 24% (n = 10) and 25% ± 25% (n = 8) of the L2-3 Pyr and PV neurons exhibited TI stimulation, respectively (Fig. 4a and b ii). Surprisingly, after isolation from the network through the synaptic blocker intervention, the percentages of TI exhibition in both neuron types reduced significantly. After synaptic blocking, 80 ± 21% (n = 10) of the L2-3 Pyr neurons did not exhibit TI stimulation compared to the 30 ± 24% prior to synaptic blocking (Fig. 4a iii). Similarly, all 100 ± 0%(n = 8) of the L2-3 PV neurons did not exhibit TI stimulation after synaptic blocking compared to the 75 ± 25% prior to synaptic blocking (Fig. 4b iii). Overall, our results suggest a major contribution of network effects in causing the TI phenomenon: TI exhibition is almost nonexistent in isolated neurons, except for two Pyr neurons.

a Whole-cell patch clamp representative recordings from a L2-3 Pyr neuron to TI stimulation before and after pharmacological isolation from the network. (i) Regular-spiking firing pattern in response to intracellular current injection and morphology of a L2-3 Pyr neuron. (ii) Consistent firing response to an extracellular modulated sinusoid at 70 μA amplitude, base frequency of 2 kHz and modulation frequency of 20Hz. Percentage of TI exhibition before pharmacology application. For the same amplitude (70 μA) and frequency (2 kHz), no consistent firing response to an extracellular pure sinusoid. (iii) After pharmacology application: percentage of TI exhibition, and illustrative plots of responses showing lack of TI exhibition. b Whole-cell patch clamp representative recordings from a L2-3 PV neuron to TI stimulation before and after pharmacological isolation from the network. (i) Fast-spiking firing pattern in response to intracellular current injection and morphology of a L2-3 PV neuron. (ii) No consistent firing response to an extracellular modulated sinusoid at 30 μA amplitude, base frequency of 2 kHz and modulation frequency of 20Hz. Percentage of TI exhibition before pharmacology application. For the same amplitude (30 μA) and base frequency (2 kHz), consistent firing response to an extracellular pure sinusoid. (iii) After pharmacology application: percentage of TI exhibition after pharmacology application, and illustrative plots of responses showing lack of TI exhibition.
The firing behavior of isolated PV and Pyr neurons further illuminates the role of network in exhibition of TI stimulation in Pyr neurons in intact network. We first make two important observations: i) The FR of isolated PV neurons is significantly larger under pure sinusoids as compared to modulated sinusoids (pure sinusoid FR: 49.7 ± 36.8Hz, and modulated sinusoid FR: 19.7 ± 16.33Hz); ii) The FR of isolated Pyr neurons is roughly similar for both pure and modulated sinusoids (pure sinusoid FR: 14.3 ± 8.31Hz, and modulated sinusoid FR: 16 ± 6.83Hz). These two observations, together with the observation that intact network Pyr neurons tend to exhibit TI stimulation, lead us to the following proposed mechanism for TI stimulation in Pyr neurons (in intact network): Pyr neurons in off-target regions are strongly inhibited by PV neurons due to their high FR in response to pure sinusoidal stimulation, thereby raising Pyr activation threshold. In contrast, Pyr neurons in the target regions experience decreased synaptic inhibition due to PV neurons as a result of lower PV FR under modulated sinusoidal stimulation, thereby allowing Pyr activation. We hypothesize that this selective inhibition is the primary reason Pyr neurons exhibit TI stimulation.
Computational results: understanding the spiking behavior of isolated PV and Pyr neurons under different electrical field shapes and orientations
It is well known that the neuronal spiking behavior under extracellular electrical stimulation depends upon the shape and orientation of the applied electric field (the shape of the electric field refers to the spatial distribution of the electric field at the neuron’s location, and orientation refers to the direction of the electric field relative to the neural axon.)22,27,28. Our proposed mechanism of TI stimulation relies on the differences in the spiking behaviors of isolated Pyr and PV neurons under pure and modulated sinusoidal stimulation. Thus, it is essential to understand the spiking behavior of isolated PV and Pyr neurons under different applied electric fields to assess if our proposed TI stimulation mechanism generalizes to different settings, particularly TES. Towards that goal, we simulated spiking patterns of isolated PV and Pyr neurons under different applied electric fields for pure and modulated sinusoids, focusing more on transcranial electric fields. We further analyze the isolated PV and Pyr firing rate patterns under spatial interference of sinusoids observed in TI stimulation, where the waveform also changes in addition to the electric field shape21.
These simulations aimed to assess if the isolated PV and Pyr spiking patterns are qualitatively similar to the isolated PV and Pyr spiking patterns observed experimentally in the Results section. Consistent with our experimental observations, the difference in the isolated PV and Pyr firing rates under modulated sinusoidal stimulation tends to be much smaller than in pure sinusoidal stimulation across various applied electric fields.
Our simulation results provide evidence that pure sinusoids continue to cause stronger Pyr inhibition than modulated sinusoids by stimulating PV neurons at higher firing rates. We next detail our simulation studies.
Computational Study 1: understanding spiking behavior of PV and Pyr neurons under different electrical fields
We simulated 210 different applied electric fields using point electrodes (previously used in Mirzakhalili et al.12) and uniform fields (previously used in Wang et al.21). For each electric field, we calculated the activation thresholds (defined in the methods section) and the corresponding firing rates for both PV and Pyr neuron models under pure and modulated sinusoidal stimulation. For point electrode stimulation, the point electrode was placed at 6 different distances: {0.5mm, 1mm, 2mm, 4mm, 8mm, 16mm}, interpolating invasive electric field shapes to non-invasive (uniform) fields with increasing electrode-neuron distance. We generated 30 orientations for each electrode-neuron distance by placing the electrode at 30 approximately uniformly spaced locations on a sphere centered at the neuron’s soma, with the radius being the corresponding electrode-neuron distance. We similarly generated 30 orientations for uniform field stimulation by choosing 30 different directions as follows:
-
1.
We choose the origin as the center of the neuron’s soma, and then choose 30 different uniformly spaced locations on a unit sphere centered at the origin. Same locations were chosen for the uniform stimulation and the point electrode stimulation corresponding to 1mm electrode-neuron distance, thereby generating the same orientations for uniform and the point electrode electric fields.
-
2.
Each chosen individual direction corresponds to the direction of the vector starting at the origin and ending at one of these 30 locations.
Figure 5a shows a schematic of the different point electrode locations for the 1mm electrode-neuron distance. Fig. 5c and d show that, on average, the activation thresholds for Pyr neuron continues to be much larger than the activation thresholds for PV neuron for all 7 electric field shapes (6 shapes from point electrodes and 1 from uniform stimulation), consistent with our experimental results (see Fig. 3). For all electric field shapes, the Pyr activation thresholds’ median (among the orientations) is > 40% larger than the corresponding PV activation thresholds’ median. The decreased spread across box plots in Fig. 5c implies that the dependence of Pyr and PV thresholds on electric field orientations decreases from invasive to non-invasive fields. Although uncommon, there are electric field shapes and orientations having lower activation thresholds for Pyr neurons than PV neurons, suggesting existence of electric field shapes and orientations for which Pyr neurons (in network) may not exhibit TI stimulation.

Results of computational study 1: a provides a representative figure of the 30 different point electrode positions (denoted by orange dots) relative to the neuron model for 1mm electrode-neuron distance. b shows the box plot of the difference in PV firing rate with Pyr firing rate at the Pyr activation threshold for the 7 electric field shapes with dots representing different orientations. c shows the relative increase in the Pyr activation threshold with respect to the PV activation threshold, with each blue dot corresponding to a particular orientation, and d shows the corresponding median (among the orientations) of the relative increase in the Pyr activation threshold. Results of computational study 2: e provides a schematic of the non-invasive simulation setup used in computational study 2. The black dot denotes the cathode (placed at C4), and the red dot denotes the anode (placed at C3). The blue dots represent the spatial locations where PV and Pyr firing rates are evaluated for pure and modulated sinusoids. f and g show the spatial map of PV-Pyr firing rate, and Pyr firing rate, respectively, for the C3-C4 stimulation at 500mA injected current. h shows the box-plot of PV-Pyr firing rate among the spatial locations having a non-zero Pyr firing rate for all injected currents for the C3-C4 configuration. Results of computational study 3: i shows the schematic of simulating a traditional TI arrangement. The two electrode pairs are denoted by the red and green dots, respectively. The target and off-target regions are denoted by orange and blue dots, respectively. j and k show the PV and Pyr firing rates in the target and off-target regions, respectively, when stimulated with TI arrangement at 1100mA injected current. l shows the PV and Pyr firing rates along a 1-D line directly parallel to the line joining the electrodes at a depth of 1.5cm.
Figure 5b shows the median (among the 30 orientations) of the difference in the PV and Pyr firing rates at the Pyr activation threshold for all electric field shapes. This difference is larger for pure sinusoids than modulated sinusoids, suggesting increased inhibition under pure sinusoidal stimulation. Notably, the difference in PV and Pyr firing rates for pure sinusoids tend to be twice the corresponding difference for modulated sinusoids in the invasive (electrode-neuron distance: 0.5mm) and non-invasive (electrode-neuron distance: 8mm, 16mm, and uniform) fields’ case. For electric fields corresponding to intermediate electrode-neuron distances (namely, 1mm, 2mm, and 4mm), the PV and Pyr rates’ differences for pure sinusoids, although still higher, are more similar to the corresponding modulated sinusoids’ differences, compared to the invasive and non-invasive electric fields’ counterparts. This behavior suggests a complicated relationship between the neural response and the electric field shape and orientation. More results are provided in SI Sec. 3.
Computational Study 2: Understanding spiking behavior of PV and Pyr neuron under transcranial fields
We simulated the spiking behavior of isolated PV and Pyr neuron models under a standard transcranial electrode arrangement29,30. The cathode and anode are respectively placed at C4 and C3 locations based on the EEG 10-20 system31. The resultant electric field is simulated using a 4-sphere head model (previously used in Forssell & Goswami et al.32 and Goswami & Grover33). Figure 5e provides a schematic of the study. We evaluated the firing rate of PV and Pyr neurons placed at 250 uniformly randomly chosen locations (shown as blue dots in Fig. 5e) across 10 levels of injected current ranging from 100mA to 1000mA. The Pyr and PV neurons were oriented perpendicular to the brain sphere surface, representing the common orientation of these neuron types found in cortical gyri34.
Figure 5f and g, respectively, show the PV and Pyr firing rates’ differences and the corresponding Pyr firing rates with pure and modulated sinusoidal stimulation across the spatial locations denoted by blue dots in Fig. 5e (at 400mA). From both these figures, we observe: i) Pyr neurons fire at roughly similar firing rates under both pure and modulated sinusoidal stimulation; ii) The Pyr and PV firing rates’ difference continues to be larger for pure sinusoids (similar to our experimental results). These two observations suggest that pure sinusoids continue to inhibit Pyr neurons for transcranial fields strongly. Figure 5h shows that the median of the differences between PV and Pyr firing rates (computed across the locations with non-zero Pyr firing rates for modulated sinusoids) continues to be significantly larger for pure sinusoids across all the amplitudes tested. We repeated this study for two more electrode arrangements and found the results remain qualitatively the same (see SI Sec. 4). These results provide further evidence that under transcranial fields, pure sinusoids should continue to cause increased inhibition in Pyr neurons due to higher PV firing rates for common transcranial electrode arrangements.
Computational Study 3: Understanding spiking behavior of PV and Pyr neurons under spatial interference of sinusoids
The spatial interference of sinusoids creates complex spatiotemporal phenomena such as rotating electric fields21, of which the pure and modulated sinusoids form the two extremes. We contend that the exhibition of TI stimulation in Pyr neurons (with synaptic inputs) is primarily a network effect, as most isolated Pyr neurons’ individual ion-channels dynamics do not allow for TI exhibition (See SI Fig. 7 and Fig. 4). Consequently, regardless of the exact spatiotemporal profile of the electric field resulting from the spatial interference of sinusoids, we expect Pyr neurons to be activated at a lower threshold in the target region if the following two conditions hold: i) PV and Pyr neurons fire at similar firing rate in the target region (resulting in lower inhibition mimicking modulated sinusoids’ network response); ii) PV neurons fire at much higher firing rate than Pyr neurons in the off-target region (resulting in higher inhibition mimicking pure sinusoids’ network response).
We simulated spatial interference under a traditional TI arrangement and examined the induced firing rates in PV and Pyr neurons at different spatial locations. Figure 5i provides a schematic of our simulation setup. We evaluated the firing rates of PV and Pyr neurons at 100 target and 250 off-target locations, uniformly randomly sampled. The target and the off-target regions are represented by orange and blue dots, respectively, in Fig. 5i. The target region is a 1cm radius disc centered at coordinates (0cm, 0cm, 7.7cm), where the spatial interference of sinusoids results in waveforms closely resembling modulated sinusoids. Eight different amplitudes of injected currents ranging from 500mA-1200mA were tested. Figure 5j shows that the firing rates of PV and Pyr neurons in the target region are similar. Figure 5k shows that PV neurons have much higher firing rates in the off-target region than Pyr neurons, providing evidence for stronger inhibition of Pyr neurons in the off-target region. This stronger Pyr inhibition in the off-target region may allow for preferential activation of Pyr neurons in the target region. Additional Results are in SI Sec. 5.
Computational Study 4: understanding how and why isolated Pyr neurons can show TI
As observed in our experimental work, a few Pyr cells continue to exhibit TI stimulation in isolation. This raises a natural question: what about the single-cell dynamics allows these cells to exhibit TI stimulation? In this section, we explore this question through computational models. We utilize a reductionist approach, simplifying models in order to isolate the phenomenon that causes TI stimulation in these isolated neurons. Specifically, we examine a condition on a neurons’ ion-channel dynamics that allows exhibition of TI stimulation. We borrow insights from dynamical system analysis and control theory35 to propose a simple mechanism (discussed in detail in SI Sec. 6) by which isolated neurons can continue to exhibit TI stimulation, namely, integral control. We use simplistic neuron models to test our proposed mechanism.
Our computational results suggest that the exhibition of TI stimulation in isolated neurons requires perturbing a delicate neural “current balance”. For pure sinusoid input (of moderate amplitudes), the inward and outward ionic currents balance each other perfectly over each period of the sinusoid, preventing a neuron from firing. Instead, the neuron is maintained in a subthreshold periodic orbit (the high-frequency current balance phenomenon of interest here can be thought of as an “envelope accommodation”, and is discussed in more detail in the Discussion section.) This current balance, as is shown in SI Fig. 14, is present in sophisticated models of spiking neurons (e.g., FitzHugh-Nagumo and Hodgkin-Huxley models). However, what separates neurons that exhibit TI and those that do not is how fast the dynamics of an integral-control feedback (caused by inward currents) mechanism are. If these dynamics are sufficiently slow, as can be the case for regular-spiking neurons (e.g. Pyr neurons), then a modulated envelope of sufficiently high slope can allow the neuron to escape the subthreshold periodic orbit and fire. If these dynamics are fast, as is the case for fast-spiking neurons, a very high slope is required for the neuron to escape this orbit. Producing high slopes can require high amplitudes at which even the pure sinusoid may cause firing.
Why is the nature of the control mechanism, namely that it is an integral control, important? This is because integral control ensures zero residual “error” between inward and outward currents23. The inward current is automatically adjusted to balance the outward current, creating the current balance we observe. In SI Fig. 14, we observe that this integral control can indeed be observed to be present in FitzHugh-Nagumo and Hodgkin-Huxley models by linearizing them about an operating point. Hence, we propose that a minority of Pyr neurons’ ion-channel dynamics are such that a modulated sinusoid has a sufficiently high slope to escape the stable sub-threshold periodic orbit.
Discussion
This work provides neuron-type selective insights on how TI stimulation influences neural activity. We discover that pure and modulated sinusoids elicit different firing rate patterns in excitatory Pyr and inhibitory PV neurons, summarized as follows: i) Pure sinusoids induce much higher firing rates in isolated PV neurons compared to modulated sinusoids; ii) Pure and modulated sinusoids produce similar firing rates in isolated Pyr neurons. We further observe that most isolated PV and Pyr neurons, i.e., without synaptic inputs, do not exhibit TI stimulation both experimentally (Fig. 4) and in realistic biophysical neuron models (see SI Fig. 7). These results are contrary to the dominant thought in the literature, where neurons are thought to perform envelope demodulation. Based on our results, we propose a revised understanding of TI stimulation (Fig. 1; right): i) In the off-target regions, where the injected waveforms resemble pure sinusoids, the activation thresholds of Pyr neurons are higher due to increased synaptic inhibition resulting from higher firing rates of PV neurons; ii) In the target region, where the injected waveforms resemble modulated sinusoids, the activation thresholds of Pyr neurons are lower due to decreased synaptic inhibition resulting from lower firing rates of PV neurons, allowing preferential activation of Pyr neurons in the target region. We note that, although uncommon, some neurons’ intrinsic dynamics (without synaptic influence) also enable them to exhibit TI stimulation, which we explain using computational observations on neural models.
Our insight into the role of the network in TI stimulation for cortical circuits provides a new lens for interpreting and predicting its effects. Our results suggest that the current design philosophy behind TI stimulation, i.e., ensuring that the waveforms resemble modulated and pure sinusoids in the target and off-target regions, respectively, does not guarantee preferential activation of Pyr neurons in the target region. Successful utilization of TI stimulation requires designing the correct spatiotemporal distribution of the currents which causes high inhibition of Pyr neurons in the off-target region and low inhibition of Pyr neurons in the target region. Lastly, with TI stimulation, PV neurons are expected to inhibit the endogenous activity in the off-target region, and any resultant side-effect should be studied in subsequent studies.
Our cell-type selective insights into the TI mechanism also open up novel avenues for stimulus design. Traditional TES strategies9,33,36,37,38,39 have not explicitly considered using cell-type selective stimulation to reduce off-target stimulation. Recent computational works40,41 have designed waveforms that selectively activate PV and Pyr neurons with much higher selectivity than sinusoids. Such waveform shapes can be used to design more effective alternative strategies to TI stimulation, where PV-selective and Pyr-selective waveforms can inhibit and activate Pyr neurons in the off-target and target regions, respectively. These aspects deserve further study.
Scalp pain is a significant issue for translating TI stimulation to humans and, generally, TES42. Furthermore, our simulation study in the Results section and other simulation studies9,12,21 suggest that supra-threshold TI stimulation would require substantially larger currents (2 orders of magnitude) than the ones attempted in preliminary human TI stimulation studies13,17,43, which could likely lead to significant scalp pain. Multi-electrode arrangements9 could lead to lower scalp pain. Furthermore, methodologies such as applying local anesthesia (which has successfully reduced scalp pain in TDCS44) can reduce scalp pain during TI stimulation. Our proposed network insight could also be beneficial in performing TI stimulation through Transcranial Magnetic Stimulation (TMS)45. TMS is known to cause significantly lower or no scalp pain compared to TES42,46. Our proposed mechanism can be used to optimize TMS-TI further to mimic the desired network effect in the cortex.
The study has several limitations. The experimental results here show a causal effect of network effects on the exhibition of TI stimulation but do not show a direct causal influence of PV activation on Pyr inhibition. E.g., other inhibitory neuron types could be involved. Nevertheless, it seems clear that network inhibition plays a role in the TI stimulation exhibition. Additional neuron types involved in inhibiting Pyr neurons need to be studied further.
Our brain-slice paradigm has inherent limitations. In making inferences about activation in target brain region vs off-target brain region, we assume that models of neurons are the same (i.e., the PV and Pyr neurons used here have similar dynamics in target and off-target tissue). This assumption could be closer to reality, e.g., when trying to activate sulcal neurons while not activating the off-target gyri. However, the results will need to be tested carefully when the target region is not cortical (e.g., hippocampal regions), in which case the dynamics and the networks could be very different. Furthermore, it is infeasible to experimentally test PV and Pyr firing rates under various electric field configurations for our experimental setup. We used simulation studies to supplement our existing experimental results and further understand the TI stimulation mechanisms. Note that simulation studies have been widely used for understanding mechanisms of TI12,21). Our brain-slice experiments used a bipolar concentric probe for stimulating neurons which is more akin to a model of invasive stimulation. While our simulation studies suggest that our mechanistic explanations are likely to hold for non-invasive stimulation, a uniform electric field setup can be used to better model non-invasive stimulation experimentally.
The electric field simulation in our computational studies uses a simplified spherical head geometry for modeling TES. While spherical head models have been used for studying TES32,33, more accurate simulations can be performed using more realistic MRI head models. We chose spherical head models due to the voltage and electric fields produced by an electrode arrangement having closed-form expressions32, providing us with arbitrary spatial precision in voltage. Most MRI head models have a ~ 1mm3 spatial precision in the resultant voltage, which is not sufficiently accurate for simulating the extracellular stimulation.
The simpler neuron models used for understanding why and how few isolated Pyr neurons exhibit TI stimulation have different ion-channel dynamics than typical cortical neurons. Most relevant models of isolated cortical neurons do not exhibit TI stimulation, as expected from our experimental studies, rendering them unusable for understanding why and how isolated Pyr neurons can exhibit TI stimulation. Consequently, we decided to find what conditions on ion-channel dynamics could allow isolated neurons to exhibit TI stimulation in simpler models rather than trying to find a suitable cortical neuron model.
The suggested mechanism of TI stimulation in isolated neurons seems intimately linked with that of “nerve conduction block”24,25,26, where pure sinusoidal currents are applied at kHz frequencies, commonly locally on long axons of peripheral nerves, to create a block of nerve conduction. Action potentials, generated at or closer to the soma, are arrested as they pass the axon location affected most by the blocking electrode. Some of the proposed mechanisms of conduction block25,47,48 have connections with our observations. A slow membrane depolarization causes sodium channel inactivation on a pure sinusoidal input25. Further, the “onset response”49 (also discussed in Kilgore & Bhadra26) observed in the conduction block is also observed in our simulations as well as experiments. This connection also deserves further study.
Methods
TI exhibition criteria
For simplicity, in this work (as also in Cao & Grover10), we focus on modulated sinusoids of the form \(A\cos (\pi \Delta ft)\sin (2\pi (\, f+\Delta f/2)t)\), comparing them with pure sinusoids of the form \(A\sin (2\pi ft)\), so that the peak amplitudes of the modulated and the pure sinusoid are the same. f and Δf are the base and modulation frequencies, respectively.
A neuron is deemed to exhibit TI when, for the same peak amplitude: i) the modulated sinusoid causes the neuron to fire consistently, and ii) the pure sinusoid does not cause a consistent firing neural response. Firing is considered consistent when repeated ( > 7 spikes) are distributed roughly uniformly through the stimulation duration. For example, a neural response of an initial spike without subsequent firing is considered to be inconsistent, and the corresponding neuron is deemed to have not been stimulated.
Experimental methods
Animals
Experiments were performed in C57/Bl6 and PV-tdt(-) mice (n=20) (Jackson Laboratory Stock No: 000664) when targeting Pyr neurons and PV-tdt(+) mice (n=13) (Jackson Laboratory Stock No: 032301) when targeting PV neurons. Animals from both sexes were indistinctly selected and were sacrificed for electrophysiology experiments at P18-P30. Brief isoflurane anesthesia was applied before decapitation. All experimental procedures were conducted in accordance with the NIH guidelines and were approved by the Institutional Animal Care and Use Committee at Carnegie Mellon University. We have complied with all relevant ethical regulations for animal use.
Electrophysiology
Coronal brain slices, 350 μm thick, were obtained on a VT1200S vibrating blade microtome (Leica Biosystems Inc., Buffalo Grove, IL) submerged into 95% CO2 and 5% O2 oxygenated ice-cold artificial cerebrospinal fluid consisting of (in mM): 119 NaCl, 26.2 NaHCO3, 11 D-glucose, 2.5 KCl, 2.5 CaCl2, 1.3 MgSO4 and 1 NaH2PO4. After 30 min of recovery at room temperature, the slices were transferred into the recording chamber for whole-cell current-clamp experiments. L2-3 Pyr and PV neurons from the primary somatosensory cortex were targeted under Olympus BX43 Light Microscope (Tokyo, Japan) and recorded from 6-10MΩ resistance borosilicate glass electrodes filled with internal solution consisting of (in mM): 125 K-gluconate, 10 HEPES, 2 KCl, 0.5 EGTA, 4 Mg-ATP, 0.3 Na-GTP (pH 7.21 and 281mOsm) with 2 μL/ml of Alexa Fluor 594 dye added just before use. The electrophysiological data was acquired using MultiClamp 700B amplifier (Axon Instruments, Foster City, CA), digitized with National Instruments acquisition interface (National Instruments, Austin, TX), and collected using IgorPro6.0 software (Wavemetrics, Lake Oswego, Oregon) at 10 kHz digitizer sampling rate. L2-3 Pyr neurons’ identities were confirmed by pyramidal-shape morphology with thick apical dendrite, resting membrane potential around -70mV, and a regular-spiking firing pattern characterized by sequentially increasing intracellular depolarizing DC currents from 25-300pA and 500ms duration. Similarly, L2-3 PV neurons’ identities were confirmed using fluorescence overlay between pre-recording reporter image and post-recording filled neuron image, resting membrane potential around -65mV, and a fast-spiking firing pattern characterized using the same protocol applied in Pyr neurons.
Electrical stimulation
All waveforms were generated from a custom script controlling a Rigol DG1062Z arbitrary waveform generator (TEquipment, Long Branch, NJ), interfacing with a custom voltage-to-current converter implemented using a modified Howland current source and delivered using a Pt/Ir CEA-200 concentric electrode (MicroProbes for Life Science, Gaithersburg, MD). The stimulation electrode was placed in cortical layers 2–3 at an angle relative to the vertical axis perpendicular to the layer. All tested stimulations had a duration of 0.5s and were applied for 3 trials, with 10s inter-trial spacing. Pure and modulated sinusoids were applied with amplitude increased in steps of 10 μA until one of the waveforms caused the neuron to fire consistently. The waveform type (pure and modulated sinusoid) and base frequency (2 kHz and 4 kHz) were randomized to avoid confounding factors. Finally, an intracellular depolarizing current above the rheobase was injected to check for any changes in the neuron excitability throughout the recording.
Pharmacology
We bath-applied the following synaptic blockers to asses network effects: 10 μM CNQX (Sigma) AMPA/kainate receptor antagonist, 50 μM D-AP5 (Tocris) NMDA receptor antagonist, and 5 μM SR 95531 hydrobromide (Tocris) GABAA receptor antagonist. The same TI stimulation protocol was applied first in the absence and then in the presence of the synaptic blockers to each neuron. The protocols were spaced by 15 min to allow the synaptic blockers to take effect. The order of the pharmacological conditions was not randomized to avoid undesired pharmacological wash-out consequences.
Data analysis
The data from a neuron was discarded unless it satisfied the following conditions: i) the baseline membrane potential was consistently maintained at less than -50mV and -45mV for Pyr and PV neurons, respectively, and ii) no confounding spontaneous action potentials were observed. The collected data was analyzed using MATLAB software (MathWorks). Action potential spikes were detected after filtering the recording traces using a 6th-order Butterworth bandpass filter of 10-1000Hz cutoff frequencies. A spike was counted when its amplitude was > 15mV above the baseline and was confirmed through visual examination. All reported spike counts were averaged across 3 trials. A neuron’s TI stimulation exhibition was determined according to criteria defined in the Results section.
We normalize the activation threshold amplitude to account for different electrode-neuron distances for comparing activation thresholds across different neurons. The normalization procedure is as follows: let the experimentally observed current amplitude at the threshold be A, and the distance between the neuron and the electrode tip be d. The normalized current amplitude then is calculated as: Normalized Amplitude = A × ∥J(d)∥/∥J(100)∥, where J(d) (Am−2) is the simulated current density function at distance d from the bipolar electrode, and ∥ ⋅ ∥ denotes its magnitude. The current density function (J(d)) was simulated using finite element modeling in COMSOL, where the bipolar probe was placed in an infinite homogeneous medium of conductivity 1Sm−1 and injected with 1μA of current. The simulated current density plots are shown in SI Fig. 4. The distance d was measured using the measurement tool from the Retiga ELECTRO camera software (Teledyne Photometrics, Tucson, AZ). This normalization procedure tries to emulate the current amplitude that would have been required if the electrode-neuron distance was 100μm.
Statistics
All statistical analysis was performed using R (Rstudio). In the first subsection of Results and Fig. 2, we jointly examine all neuron samples across all base frequencies, including the samples, before applying a synaptic blocker. For this analysis, we excluded the same neuron samples after the presence of synaptic blockers. In the third subsection and Fig. 4, we only examine the neuron samples that undergo synaptic blocking and consider both their conditions, i.e., before and after synaptic blocking. In the second subsection and Fig. 3, we split neuron samples for each base frequency and analyze them separately. Throughout the manuscript, we report averages with their standard deviations and percentages with their 95% confidence intervals. To compare activation thresholds, FR, and ISI, we use non-parametric tests listed in SI Table 1 and SI Table 2. Particularly, for amplitude threshold comparisons, we applied a Kruskal-Wallis using Mann-Whitney rank sum test with Holm correction because it was a comparison between multiple samples and, even after a log transformation of the data, normal distribution was a poor approximation. Similar corrections like Bonferroni or Tukey were equally appropriate and supported the same statistical significance reported. Statistical significance is reported at p < 0.05 (with p-values explicitly provided). Unless specified, all statistical tests are two-sided.
Computational models
The first three computational studies described in the Results were performed using python50, and all the corresponding figures were made using matplotlib51. The final computational study in the Results section on understanding why and how few isolated Pyr neuron exhibit TI stimulation was performed using MATLAB52.
Neuron models
We use realistic biophysical neuron models for the first three computational studies described in the “Results” section, originally published in Markram et al.53. We use models of a layer 2–3 cortical Pyr neuron and a layer 2–3 inhibitory classical non-accommodating firing-type PV-positive large basket cell neuron (the same neuron types from our experimental study) from the rat somatosensory cortex. The rat neurons tend to be slightly larger than the mouse neurons used in our experimental studies, but the inferences obtained from rat neurons should largely correspond to mice neurons. The neuron models were modified according to the algorithm presented in Aberra et al.54 for simulating extracellular stimulation. The neuron models were simulated using the python environment of NEURON55,56. Similar models were earlier used by Wang et al.21 for studying TI stimulation. For the fourth and final computational study, simpler single-compartment models, i.e., integrate-and-fire, FitzHugh-Nagumo, and Hodgkin-Huxley models, were used to examine the question of what ion channel dynamics allow isolated neurons to exhibit TI stimulation. Note that the realistic neuron models used in the first three computational studies do not exhibit TI stimulation and cannot be used for understanding when isolated neurons exhibit TI stimulation. Furthermore, none of the neuron models exhibit spontaneous firing activity under no electrical stimulation. Additional model parameter choices are detailed in SI.
Electric field simulation
To simulate the extracellular stimulation of the bio-physically realistic neuron models used in the first three computational studies, we need to specify the corresponding extracellular voltage induced along the morphology of the neuron models. We briefly describe the details of different extracellular voltages induced by different extracellular electric fields used for simulating extracellular stimulation of neurons in the first three computational studies. The extracellular voltage at location (x, y, z), denoted as Vpoint(x, y, z) and Vuniform(x, y, z), due to a point electrode placed at (x0, y0, z0) and uniform electric field, respectively, is specified as follows:
where I is the current injected at the electrode and σ is the conductivity of the medium, chosen as 0.33 Sm−1. The vectors \(\hat{{{\bf{i}}}},\hat{{{\bf{j}}}}\), and \(\hat{{{\bf{k}}}}\) denote the corresponding standard vectors of the x-, y-, and z-directions, respectively. \({E}_{unif}^{x},{E}_{unif}^{y}\,\,{\mbox{and}}\,\,{E}_{unif}^{z}\) are the respective x, y, and z components of the unit vector denoting the direction of the uniform electric field, and Aunif denotes the electric field amplitude. The symbol ⋅ denotes the dot product in (2). Note that since the amplitude of \({\overrightarrow{{{\bf{E}}}}}_{unif}\) is Aunif, we have:
The voltage in the 4-sphere head model used in our computational studies was calculated using the closed-form expression described in theorem 1 of Forssell et al.32. The parameters of the 4-sphere head models are as follows: overall radius of the 4-sphere head model was 9.2cm, with the thickness of scalp, skull, and cerebro-spinal fluid (CSF) being 6mm, 5mm, and 1mm, respectively, taken from57. The conductivity values for the scalp, skull, CSF, and brain layers were chosen as 0.3Sm−1, 0.006Sm−1, 1.79Sm−1, and 0.33Sm−1, respectively, taken from Fernandez-Corazza et al.36. Each electrode was modeled as a disc electrode with a diameter of 0.5cm.
Activation threshold
For point electrode stimulation, the activation threshold for each neuron type was chosen as the smallest input current amplitude at which the chosen neuron type fires at 20Hz with modulated sinusoids. The value of 20Hz was chosen to maintain consistency with experimental results, where both neuron types (with and without synaptic blockers) have ~ 20Hz firing rates at their respective activation threshold. The increase in the threshold between Pyr and PV neurons, shown in Figs. 5g and j, is quantified as: \({\,{\mbox{Th}}\,}_{{{\rm{PV}}}\to \,{\mbox{Pyr}}}^{inc}=({I}_{th}^{Pyr}-{I}_{th}^{PV})/ { {\mbox{Median}}}\,({{\mbox{I}}}_{th}^{PV})\times 100\), where \({I}_{th}^{Pyr}\) and \({I}_{th}^{PV}\) are the activation thresholds for a particular orientation of a given electric field shape and \(\,{\mbox{Median}}\,({I}_{th}^{PV})\) is the median of the activation thresholds across all the orientations for that particular electric field shape. For uniform field stimulation, the activation threshold is similarly defined as the point electrode stimulation case, where we use the smallest electric field amplitude at the neuron instead of the smallest input current amplitude.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The source data for generating all the experimental figures is provided in the Github repository58 located at https://github.com/chaitanyagoswami/Cell-Specific-Effects-of-Temporal-Interference-Stimulation-on-Cortical-Function-Code/tree/main. The raw electrophysiological data is available at the following https://doi.org/10.1184/R1/26517562.v159.
Code availability
The code for generating all the computational results as well as the experimental figures present in this work is available at https://github.com/chaitanyagoswami/Cell-Specific-Effects-of-Temporal-Interference-Stimulation-on-Cortical-Function-Code/tree/main58.
References
Nitsche, M. A. & Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 527, 633–639 (2000).
Bennabi, D. & Haffen, E. Transcranial direct current stimulation (tDCS): a promising treatment for major depressive disorder? Brain Sci. 8, 81 (2018).
Nitsche, M. A. et al. Transcranial direct current stimulation: state of the art 2008. Brain Stimul. 1, 206–223 (2008).
Grossman, N. et al. Noninvasive deep brain stimulation via temporally interfering electric fields. Cell 169, 1029–1041 (2017).
Goats, G. Interferential current therapy. Br. J. Sports Med. 24, 87 (1990).
Savage, B. Interferential Therapy (Faber And Faber, 1984).
Song, X., Zhao, X., Li, X., Liu, S. & Ming, D. Multi-channel transcranial temporally interfering stimulation (tTIS): application to living mice brain. J. Neural Eng. 18, 036003 (2021).
Rampersad, S. et al. Prospects for transcranial temporal interference stimulation in humans: a computational study. NeuroImage 202, 116124 (2019).
Cao, J. & Grover, P. STIMULUS: noninvasive dynamic patterns of neurostimulation using spatio-temporal interference. IEEE Trans. Biomed. Eng. 67, 726–737 (2019).
Cao, J. & Grover, P. Do single neuron models exhibit temporal interference stimulation? (2018). In IEEE Biomedical Circuits and Systems Conference (BioCAS), pp. 1-4 (2018).
Dmochowski, J. & Bikson, M. Noninvasive neuromodulation goes deep. Cell 169, 977–978 (2017).
Mirzakhalili, E., Barra, B., Capogrosso, M. & Lempka, S. F. Biophysics of temporal interference stimulation. Cell Syst. 11, 557–572 (2020).
Zhu, Z. et al. Temporal interference (TI) stimulation boosts functional connectivity in human motor cortex: a comparison study with transcranial direct current stimulation (tDCS). Neural Plast. 2022, 7605046 (2022).
Zhang, Z. et al. Designing and pilot testing a novel transcranial temporal interference stimulation device for neuromodulation. IEEE Trans. Neural Syst. Rehabil. Eng. 30, 1483–1493 (2022).
Howell, B. & McIntyre, C. C. Feasibility of interferential and pulsed transcranial electrical stimulation for neuromodulation at the human scale. Neuromodulation 24, 843–853 (2021).
TI Solutions. https://temporalinterference.com/ (2022).
von Conta, J. et al. Benchmarking the effects of transcranial temporal interference stimulation (ttis) in humans. Cortex 154, 299–310 (2022).
Zhang, Y. et al. Temporal interference stimulation targeting right frontoparietal areas enhances working memory in healthy individuals. Front. Hum. Neurosci. 16, 918470 (2022).
Acerbo, E. et al. Focal non-invasive deep-brain stimulation with temporal interference for the suppression of epileptic biomarkers. Front. Neurosci. 16, 945221 (2022).
Huang, Y., Datta, A. & Parra, L. C. Optimization of interferential stimulation of the human brain with electrode arrays. J. Neural Eng. 17, 036023 (2020).
Wang, B., Aberra, A. S., Grill, W. M. & Peterchev, A. V. Responses of model cortical neurons to temporal interference stimulation and related transcranial alternating current stimulation modalities. J. Neural Eng. 19, 066047 (2023).
Rattay, F. Analysis of models for external stimulation of axons. IEEE Trans. Biomed. Eng. BME-33, 974–977 (1986).
Ogata, K. Modern Control Engineering (Prentice hall, 2010).
Ackermann, D. M., Bhadra, N., Gerges, M. & Thomas, P. J. Dynamics and sensitivity analysis of high-frequency conduction block. J. Neural Eng. 8, 065007 (2011).
Kilgore, K. L. & Bhadra, N. Nerve conduction block utilising high-frequency alternating current. Med. Biol. Eng. Comput. 42, 394–406 (2004).
Kilgore, K. L. & Bhadra, N. Reversible nerve conduction block using kilohertz frequency alternating current. Neuromodulation 17, 242–255 (2014).
Rattay, F. The basic mechanism for the electrical stimulation of the nervous system. Neuroscience 89, 335–346 (1999).
Fellner, A., Heshmat, A., Werginz, P. & Rattay, F. A finite element method framework to model extracellular neural stimulation. J. Neural Eng. 19, 022001 (2022).
Calancie, B., Klose, K. J., Baier, S. & Green, B. A. Isoflurane-induced attenuation of motor evoked potentials caused by electrical motor cortex stimulation during surgery. J. Neurosurg. 74, 897–904 (1991).
Szelényi, A., Kothbauer, K. F. & Deletis, V. Transcranial electric stimulation for intraoperative motor evoked potential monitoring: stimulation parameters and electrode montages. Clin. Neurophysiol. 118, 1586–1595 (2007).
Homan, R. W., Herman, J. & Purdy, P. Cerebral location of international 10–20 system electrode placement. Electroencephalogr. Clin. Neurophysiol. 66, 376–382 (1987).
Forssell, M., Goswami, C., Krishnan, A., Chamanzar, M. & Grover, P. Effect of skull thickness and conductivity on current propagation for noninvasively injected currents. J. Neural Eng. 18, 046042 (2021).
Goswami, C. & Grover, P. Hingeplace: focused transcranial electrical current stimulation that allows subthreshold fields outside the stimulation target (2021). In 43rd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), pp. 1577–1583 (2021).
Kandel, E. R. et al. Principles Of Neural Science, Vol. 4 (McGraw-hill New York, 2000).
Izhikevich, E. M. Dynamical Systems In Neuroscience (The MIT Press, 2007).
Fernandez-Corazza, M., Turovets, S. & Muravchik, C. H. Unification of optimal targeting methods in transcranial electrical stimulation. Neuroimage 209, 116403 (2020).
Dmochowski, J. P., Datta, A., Bikson, M., Su, Y. & Parra, L. C. Optimized multi-electrode stimulation increases focality and intensity at target. J. Neural Eng. 8, 046011 (2011).
Guler, S. et al. Optimization of focality and direction in dense electrode array transcranial direct current stimulation (tDCS). J. Neural Eng. 13, 036020 (2016).
Huang, Y. & Parra, L. C. Can transcranial electric stimulation with multiple electrodes reach deep targets? Brain Stimul. 12, 30–40 (2019).
Goswami, C. & Grover, P. Automated electrical waveform design for cell-type selective stimulation (2023). In 11th International IEEE/EMBS Conference on Neural Engineering (NER). pp. 1–6 (2023).
Goswami, C. & Grover, P. Pathfinder: designing stimulus for neuromodulation through data-driven inverse estimation of non-linear functions. IEEE Trans. Biomed. Eng. 70, 3300–3311 (2023).
Hallett, M. Transcranial magnetic stimulation: a primer. Neuron 55, 187–199 (2007).
Ma, R. et al. High gamma and beta temporal interference stimulation in the human motor cortex improves motor functions. Front. Neurosci. 15, 800436 (2022).
McFadden, J. L., Borckardt, J. J., George, M. S. & Beam, W. Reducing procedural pain and discomfort associated with transcranial direct current stimulation. Brain Stimul. 4, 38–42 (2011).
Zhu, Z. & Yin, L. A mini-review: recent advancements in temporal interference stimulation in modulating brain function and behavior. Front. Hum. Neurosci. 17, 1266753 (2023).
Klomjai, W., Katz, R. & Lackmy-Vallée, A. Basic principles of transcranial magnetic stimulation (TMS) and repetitive tms (rTMS). Ann. Phys. Rehabil. Med. 58, 208–213 (2015).
Williamson, R. P. & Andrews, B. J. Localized electrical nerve blocking. IEEE Trans. Biomed. Eng. 52, 362–370 (2005).
Tanner, J. Reversible blocking of nerve conduction by alternating-current excitation. Nature 195, 712–713 (1962).
Bhadra, N. & Kilgore, K. L. High-frequency electrical conduction block of mammalian peripheral motor nerve. Muscle Nerve 32, 782–790 (2005).
Van Rossum, G. & Drake, F. L. Python 3 Reference Manual (2009).
Hunter, J. D. Matplotlib: A 2d graphics environment. Comput. Sci. Eng. 9, 90–95 (2007).
The MathWorks Inc. Matlab version: 9.13.0 (R2022b) (2022). https://www.mathworks.com (2022).
Markram, H. et al. Reconstruction and simulation of neocortical microcircuitry. Cell 163, 456–492 (2015).
Aberra, A. S., Peterchev, A. V. & Grill, W. M. Biophysically realistic neuron models for simulation of cortical stimulation. J. Neural Eng. 15, 066023 (2018).
Carnevale, N. T. & Hines, M. L. The NEURON book (Cambridge University Press, 2006).
Hines, M., Davison, A. P. & Muller, E. Neuron and python. Front. Neuroinf. 3, 391 (2009).
Nunez, P. L. & Srinivasan, R. Electric Fields of the Brain: The Neurophysics of EEG (Oxford University Press, 2006).
Goswami, C. et al. Source code for cell-specific effects of temporal interference stimulation on cortical function Zenodo https://zenodo.org/doi/10.5281/zenodo.12797409 (2024).
Caldas-Martinez, S., Barth, A. L. & Grover, P. Cell-Specific Effects of Temporal Interference Stimulation on Cortical Function – Raw Data (Carnegie Mellon University, 2024).
Acknowledgements
We thank Brent Doiron for helpful discussions, and Jay Reddy for technical help with recordings. Figures 1 and 2 abstract representations of neurons, brains, and instrumentation were created with BioRender.com. This material is based upon work supported by the Naval Information Warfare Center (NIWC) Atlantic and the Defense Advanced Research Projects Agency (DARPA) under Contract No. N65236-19-C-8017. Any opinions, findings and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the NIWC Atlantic and DARPA. Chaitanya Goswami was also supported by a fellowship from the Center for Machine Learning and Health at Carnegie Mellon University.
Author information
Authors and Affiliations
Contributions
S.C.M and C.G. have equally contributed to this work. S.C.M. conducted the biological experiments and the statistical analyses. S.C.M., C.G., and A.L.B. designed all biological experiments. A.L.B. supervised the data interpretation. M.F. contributed to the hardware and scripts of the biological experiments. C.G., J.C., and P.G. obtained the simulation results. S.C.M., C.G., and P.G. conceived the study and wrote the manuscript. All authors edited and approved the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editors: Karli Montague-Cardoso and Benjamin Bessieres. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Caldas-Martinez, S., Goswami, C., Forssell, M. et al. Cell-specific effects of temporal interference stimulation on cortical function. Commun Biol 7, 1076 (2024). https://doi.org/10.1038/s42003-024-06728-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-024-06728-y
