
Abstract
Processing bodies (P-bodies) are membrane-less organelles composed of condensed mRNAs and proteins that play essential role in mRNAs decay and storage, contributing to the translational control of cellular proteostasis. Regulation of P-body assembly/disassembly by signaling events, cellular stress or specific environmental conditions shapes the rate of RNA turnover and protein synthesis, controlling cell growth, differentiation and survival. Deregulation of protein translation is an important factor for tumor development and progression and cancer cells benefit from P-bodies to reshape their proteome to support specific metabolic needs and promote tumor development, progression and metastasis. Hence, understanding the composition and the regulation of P-bodies, both under physiological and pathological conditions, will define the mechanisms underlying cancer cell plasticity and develop novel therapeutic strategies to inhibit cancer growth and metastasis. Here, we will discuss the principal mechanisms of P-body regulation and function, with special focus on the role of these ribonucleoprotein condensates in cancer.
Similar content being viewed by others
Introduction
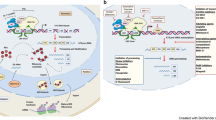
The regulation of gene expression is an important mechanism that cells adopt to modulate their proteome composition under specific growth and environmental conditions, thus playing a fundamental role in cell proliferation, differentiation, metabolism, and survival. Gene expression is controlled at multiple levels: epigenetic, transcriptional, and post-transcriptional, each involving spatially and timely regulated distinct molecular mechanisms. The first level of regulation involves epigenetic modifications of chromatin that include methylation of DNA and post-translational modifications (PTM) of histones that deeply influence chromatin structure and gene expression1. The second level is mediated by transcription factors that bind to DNA sequences located within promoter genes and to components of the transcription machinery and regulate the rate of gene transcription2,3. The third level of regulation takes place after mRNA synthesis and includes processing of newly synthetized transcripts, their export from the nucleus to the cytoplasm, association with ribosomes, and degradation by ribonucleases. These distinct regulatory layers, even if spatially and mechanistically different, are essential to define the final cellular proteome, and their deregulation is causally linked to the development of a variety of human disorders, including cancer4. Recently, a regulatory role of gene expression has been attributed to the mechanism of liquid–liquid phase sepration (LLPS), a physical process of condensation of macromolecular complexes at distinct intracellular sites5. At the epigenetic level, the formation of chromatin-associated condensate allows the organization of regulatory factors in the nuclear space6. The transcriptional control is often modulated by nuclear transcriptional condesates, that concentrate transcription machinery components at active gene loci7. At the post-transcriptional level, the formation of cytoplasmatic ribonucleoprotein (RNP) condensates, including mRNAs and proteins, namely P-bodies, allows a fine regulation of mRNA transcripts fate8 (Fig. 1). At P-bodies, mRNAs can be temporarily stored or processed by the RNA decay machinery. This is a complex regulatory system of RNA translational repression and turnover that dynamically shapes the cellular proteome9. Current evidence indicates that the dynamic assembly/disassembly of P-bodies constitutes an efficient mechanism that cells adopt to control the rate of protein translation during cell cycle progression or under specific cellular needs and environmental conditions10,11,12,13,14. Given the importance of P-bodies in the control of RNA metabolism, decay, and translation, these condensates are attracting great interest not only in the field of cell biology but also in translational oncology. This minireview summarizes the most relevant and recent findings on the role of P-bodies in cancer biology, highlighting the possibility of targeting the RNPs condensates for the treatment of aggressive malignant tumors.

The first level of regulation of gene expression is represented by modifications and remodeling of chromatin structure that occur within the nucleus. This mechanism is tightly controlled by chromatin-associated condensates (blue) that assemble regulatory factors including DNA methyltransferases (DNMTs), histone deacetylase (HDAC), histone acetyltransferase (HAT), and heterochromatin protein 1 (HP1). The second level of regulation concerns the transcription of unmethylated genes by the RNA polymerase machinery. Transcription factors (TFs), transcriptional co-activators (TF-A), and repressors (TF-R), included in the nuclear condensates (grey), contribute to the regulation of gene transcription. The third level of regulation of gene expression is represented by the control of mRNA stability and translation that occurs within the cytoplasm. Here, mature mRNA molecules exported from the nucleus accumulate within membrane-less condensates, namely P-bodies (green) and stress granules (yellow). In these compartments, ribonucleoprotein complexes (RBPs) tightly control the translation and turnover of mRNAs in response to specific cellular needs.
Membrane-less organelles and liquid–liquid phase separation (LLPS)
An important control mechanism of protein translation is represented by the binding of mRNA molecules to RNA binding-proteins (RBPs), leading to the formation of RNP complexes15. Under specific cellular conditions, different RNPs can physically condensate to form a variety of membrane-less macromolecular complexes, including processing bodies (P-bodies) and stress granules16. These condensates form through the LLPS process driven by weak interactions between proteins and nucleic acids, playing a fundamental role in localizing and concentrating untranslated mRNAs at specific intracellular compartments17,18,19,20,21,22,23. Besides P-bodies and stress granules, other mRNPs granules have been identified in eukaryotic cells, including the TIS granules composed of mRNA with an AU-rich element associated with the endoplasmic reticulum24, the Cajal bodies that are subnuclear structures linked with the nucleolus25 and the U snRNP (Uridine-rich small nuclear RNP) bodies, also named U-bodies, that associate with P-bodies and are sites of assembly of snRNPs in the cytoplasm before their transport to the nucleus26,27. In Drosophila, alterations of P-body structure impact on the composition of U-bodies, suggesting a cooperation between both types of organelles in the regulation of snRNPs26. Moreover, GW bodies are cytoplasmatic foci that regulate the storage, the processing, and the stability of mRNAs, and their dynamic is closely connected to cell cycle phase28,29. GW bodies were originally identified as granules containing the GW182 protein involved in miRNA-mediated gene silencing and were considered similar to P-bodies30. Subsequent studies in Drosophila revealed that GW bodies have different dynamics and protein composition compared to P-bodies, suggesting that they are distinct mRNPs granules31. However, this conclusion has not been formally demonstrated in human cells.
P-bodies are constitutively present in cells, whereas the assembly of stress granules is primed by stress conditions. Although morphologically and biochemically distinct, P-bodies and stress granules are functionally and dynamically interconnected. In particular, RBPs and RNA molecules located at the surface of these condensates mediate the docking and the consequent shuttling of mRNAs and RBPs between both compartments21,32. DDX6, a principal component of P-bodies, plays a fundamental role in this process by limiting stress granule formation and negatively regulating the docking between the two types of mRNP aggregates33.
P-body background and historical context
The complexity and composition of P-bodies have been well characterized through studies in lower eukaryotes. The principal constituents of P-bodies are proteins involved in the translational repression and mRNA decay that include deadenylating (CCR4-Not) and decay (Lsm1-7) protein complexes, decapping enzymes (DCP1/DCP2), activators of decapping enzymes (EDC3/EDC4 and PAT1), the RNA helicase (DDX6), and ribonuclease (XRN1)34,35,36. P-bodies also contain microRNAs (miRNAs), mRNA-binding proteins, and regulators involved in translational repression, such as CPEB337,38. The translational initiation factor eIF4E and the related factor eIF4E-transporter (eIF4E-T), both involved in the translational repression, interact with DDX6 and localize at P-bodies39. Interestingly, eIF4E is the only component of the translation machinery localized at P-bodies, where its interaction with 4E-T represses protein translation40. At first, P-bodies were considered sites of mRNA turnover, since the principal components of P-bodies in yeast were proteins involved in mRNA decay41. Subsequent studies demonstrated that P-bodies are also sites for the temporary storage of untranslated transcripts. In fact, reporter mRNAs can be temporarily localized in P-bodies when the translation is inhibited. The removal of protein synthesis inhibitors promotes a rapid exit of mRNAs from the mRNP aggregates and resumption of protein synthesis42. Moreover, cells arrested in the stationary phase have larger P-bodies, mostly enriched of mRNAs that become available for translation when cells re-enter the growth phase42. In metazoans, the translation of the cationic amino acid transporter (CAT-1) transcript, that is repressed by miR-122, is restored after its release from P-bodies and consequent recruitment by polysomes43.
The majority of early studies on P-bodies were conducted in yeast cells, which exhibit several differences from mammalian P-bodies, most notably the regulation of assembly. In particular, in these cells, P-bodies are not present under normal growth conditions, and their assembly can be primed by different environmental conditions, such as glucose deprivation, osmotic stress, and DNA replication stress44. However, these events have not been confirmed in all eukaryotes, where P-bodies are constitutively present. Accordingly, since most of the studies and findings made in yeast cells cannot be extended to mammalian systems, this review will primarily focus on the structure and regulation of mammalian P‑bodies.
Key players of mammalian P-bodies
Although the number of resident and regulatory proteins that localize at P-bodies is still growing, in mammals, only a few proteins are essential for their assembly, such as DDX6 and its partners 4-ET and LSM14A45,46. Genetic knockdown experiments demonstrated that DDX6, 4-ET, CCR4, and LSM1 are necessary for their mutual accumulation in P-bodies39. Other proteins, although playing an essential role in the regulation of mRNAs within P-bodies, are not required for the assembly of these condensates. This is the case of DCP2 and EDC3, whose silencing in mammalian cells does not affect the number of P-bodies39,45. An important regulatory element in P-body assembly is the physical interaction between various components of these granules, as demonstrated by a two-hybrid fluorescence-based assay47. The N-terminal portion of EDC4 directly interacts with LSM14A, PATL1, XRN1 and the Negative Regulator Of P-Body Association (NBDY), while only indirectly with DDX6, whereas the C-terminal portion of the protein binds to DCP1A and CCHR1 and indirectly interacts with EDC3. In cells lacking P-bodies, as a consequence of genetic deletion of DDX6 or LSM14A, overexpression of the N-terminal portion of EDC4, but not wild-type form, is sufficient to form cytoplasmic dots that are structurally indistinguishable from P-bodies, but lack LSM14A and DDX6, respectively47. The interaction between EDC4 and XRN1, an mRNA decay enzyme, is also essential for P-body dynamics and for the regulation of mRNA decapping and decay. Thus, in human cells depleted of XRN1, P-bodies are enlarged. Re-expression of a catalytic mutant of XRN1 that retains the mRNA decay activity in XRN1-depleted cells is not sufficient to restore the normal size of P-bodies. Instead, the effects of XRN1 depletion on P-bodies size can be reversed by overexpressing both the catalytic and EDC4-interacting domains of the protein. This indicates that EDC4-XRN1 interaction is essential for P-body dynamics and mRNA decay activity in human cells48. Conversely, the C-terminal portion of EDC4 is required for EDC4 self-association and condensation, and its overexpression is sufficient to promote the assembly of P-bodies49. Altogether, these results suggest that the direct interaction among components of RNP complexes is, indeed, a driving force for P-body assembly.
Regulation of P-body dynamics
Although in mammals P-bodies are constitutively present within cells, different conditions can impact on their assembly. In fact, the availability of mRNAs represents an important element for the formation of P-bodies, as demonstrated by the dissolvement of mRNP aggregates treating the cells with actinomycin-D, a potent and selective inhibitor of RNA transcription50. Moreover, experiments aimed to reconstitute bioengineered condensates in human cells demonstrated that the recruitment of RNAs influences the composition and size of RNP aggregates, and the effects of RNAs on granule size inversely correlates with its surface density51.
Metabolic changes can also impact on the assembly of P-bodies. Thus, treatment with 2-deoxiglucose and oligomycin, an experimental condition that reduces intracellular ATP levels and induces energy stress, increases the number of P-bodies52. Similarly, treatment with arsenite, an inducer of oxidative stress, promotes P-body assembly32. In this context, EDC3 plays a fundamental role. Thus, the mutagenesis of tyrosine 475 on EDC3 alters the binding to DDX6 and DCP1A and markedly increases P-body number in response to arsenite treatment, supporting a critical role of EDC3 in P-body dynamics in response to stress53. Cell cycle progression also influences the number and size of RNP condensates. In cycling cells, P-bodies dissolve during mitosis, assemble in G1, and enlarge at the interfase, with no major changes in the number of these condensates11.
As firstly observed in yeast, also in mammals has been proved that P-bodies are temporary stores of untranslated mRNAs, and not only sites of RNA turnover. Thus, in human cells, during amino acid starvation, mRNAs accumulate within P-bodies. Amino acid supplementation promotes the clearance of P-bodies and degradation of selected transcripts54. Similarly, under glucose deprivation, mRNAs condensate in P-bodies. Here, a group of transcripts is safely stored, whereas others undergo to degradation55. In summary, P-bodies represent a dynamic and rapid mechanism to control mRNA translation and turnover, explaining the variability in the RNA/protein composition of P-bodies in different cell types or under specific biological conditions.
Control of P-body dynamics by post-translational modifications
Post-translational modifications (PTMs) of proteins play a relevant role in the formation of P-bodies. The decapping enzyme DCP1a is ubiquitylated by TRAF6 and subsequently phosphorylated by c-Jun N-terminal kinase (Jnk) on serine 315. Phosphorylation primes DCP1a binding to EDC4 and Xrn1 and assembly of P-bodies56,57. Conversely, the decapping protein complex is negatively regulated by Trim24-mediated ubiquitylation of EDC458. Similarly, ubiquitylation of 4-ET by Trim56 or its deubiquitylation by Otud4 promotes P-body assembly or disassembly, respectively59. Ubiquitylation is not the only PTM that regulates P-body dynamics. Dimethylation of two arginine residues within the RGG domain of Lsm4 primes P-body assembly60. Under stress conditions, phosphorylation of 4E-T by JNK kinase promotes P-body condensation61.
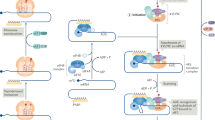
cAMP-dependent protein kinase (PKA) also plays a relevant role in P-body dynamics. Thus, in yeast, activation of cAMP signaling inhibits P-body formation through PKA-mediated phosphorylation of Pat1 and consequent inhibition of DDX6 recruitment to the complex62. Conversely, in mammals, activation of PKA induces phase-separation of regulatory subunits of PKA (PKARIα) and consequent formation of biomolecular condensates enriched of PKA and cAMP levels, contributing to compartmentalized cAMP signaling63. Similarly, the rise of cAMP levels drastically increases the number of P-bodies, confirming the positive role of PKA in the assembly of the cytoplasmic mRNP granules64 (Fig. 2).

P-bodies are intracellular condensates composed of mRNAs and RNA-binding proteins (RBPs), including deadenylating protein complex CC4-Not, decay complex Lsm 1-7, decapping enzymes DCP1/2, activators of decapping enzymes EDC3/4 and PAT1, RNA helicase DDX6, and ribonuclease XRN1. A Under stress conditions, Jnk phosphorylates 4-ET and promotes P-body assembly. Ubiquitylation of DCP1a by TRAF6 and subsequent phosphorylation by Jnk induces its binding to EDC4 and XRN1. Dimethylation of Lsm4 and ubiquitylation of 4-ET by Trim56 promotes P-body assembly. B Ubiquitylation of EDC4 by Trim24 inhibits the assembly of the decapping protein complex. Otud4-mediated deubiquitylation of 4E-T promotes P-body disassembly.
A role of P-bodies in cancer development and progression
Derangement of translational regulation is one of the main aspects of cancer cell plasticity, a mechanism that allows tumoral cells to rapidly adapt to changes of microenvironmental conditions65. Given the contribution of P-bodies in different aspects of RNA metabolism, decay, and translation, they exert a multifaceted role in cancer growth and dissemination. In fact, by modulating the expression levels of oncogenes, oncosuppressors, and growth regulators, these RNP condensates support cancer cell adaptation to stress or to specific metabolic needs66. The importance of P-bodies in tumor growth derives from the observation that, in tumoral tissues, the expression of fundamental components of these mRNP granules is often deregulated, even in the absence of major changes in P-body dynamics. This is the case of EDC4, an important component of P-bodies, that has been found mutated in BRACA1/2 mutation-negative breast cancer tissues67. Also, DCP1a expression is increased in colorectal cancer tissues, and its levels correlate with patients’ survival68. Furthermore, the metastatic lymph node 51 (MLN51) protein, an RNA-binding protein of the exon junction complex that shuttles mRNA molecules between nucleus and cytoplasm, has been recently identified as a component of P-bodies in human cells, where its overexpression promotes the disassembly of these RNP condensates. Interestingly, MLN51 is overexpressed in HER2+ breast cancer cells, and its expression levels correlate with the abundance of P-bodies. In about 50% of HER2+ breast cancers, MLN51 is co-amplified with c-erbB2, a gene encoding for the HER2 receptor. In breast cancers overexpressing both HER2 and MLN51 proteins, the number of P-bodies is notably decreased, compared to HER2+ breast cancer tissues that do not overexpress MLN51 or to normal tissue counterparts. Although a formal proof of a causal link between P-body disassembly and stages of tumorigenesis or tumor aggressiveness is missing, this study strongly suggests a correlation between P-body dynamics and malignant tumor behavior69.
A link between P-bodies and tumor progression has been proved in myeloid leukemia (AML) cells that possess an increased number of P-bodies that are essential for tumor progression. In AML cells, overexpression of DDX6 induces sequestration and translational repression of mRNAs encoding for tumor suppressors. Dissolution of P-bodies induces a resumption of mRNA translation and consequent inhibition of AML cell proliferation70. This finding demonstrated that, at least in part, the role of P-bodies in cancer cells relies on the translational regulation of selected mRNAs. Similarly, in different types of human cancer, the disassembly of P-bodies, induced by treating cells with an NBDY synthetic peptide that prevents EDC4 condensation, activates the p53 pathway and increases the stability of associated transcripts, thus inhibiting tumor proliferation49. These findings indicate that the reversible storage of mRNAs within P-bodies, thus, represents an important homeostatic control mechanism of gene expression underlying cell differentiation and tumorigenesis.
Regulation of P-body dynamics by signaling pathways in cancer
Alterations of signaling pathways and related PTMs are common features of cancer cells71. In this context, derangement of signaling events controlling P-body dynamics and protein translation can be causally linked to cancer development. Thus, Pim1/3 kinases phoshorylate EDC3 on serine 161, originally identified as an AKT phosphorylation site72,73,74. Phoshorylation by Pim1/3 kinases inhibits EDC3 localization at P-bodies, markedly impacting on tumor growth72. In prostate cancer cells, as well as in multiple breast cancer cell lines, EDC3 protein is hyperphosphorylated compared to normal cells. In a xenograft mouse model of prostate cancer, overexpression of a phosphorylation-deficient mutant of EDC3, carrying a substitution of serine 161 to alanine, reduces the growth, motility, and invasiveness behavior of cancer cells. As expected, treatment with a small kinase inhibitor that prevents EDC3 phosphorylation increases the number of P-bodies and negatively impacts on tumor growth72. The TGFβ pathway has also been correlated to the regulation of P-bodies in cancer. TGFβ promotes the epithelial-to-mesenchimal transition (EMT) in different types of human tumors, including breast cancer75,76. Treatment of breast cancer cells with TGFβ or overexpression of Twist, a transcription factor and downstream target of the TGFβ pathway, markedly increases the number of P-bodies. These effects are reversed by removing the growth factor from the medium and require the activation of the autophagic machinery. The induction of EMT by TGFβ relies, in part, on P-body formation. Thus, in orthotopic models of breast cancer, downregulation of DDX6 prevents P-body assembly and inhibits the formation of pulmonary metastasis, thus highlighting a fundamental role of these RNP condensates in the invasiveness potential of breast cancer cells77.
The Hippo pathway is an oncosuppressive signaling cascade that negatively controls cancer cell growth. In particular, activation of the Hippo pathway promotes phosphorylation and proteolytic degradation of transcriptional regulators YAP and TAZ. In proliferating cells, the inactivation of the Hippo pathway leads to YAP/TAZ translocation to the nucleus and consequent transcription of genes underlying cell growth and survival78. YAP/TAZ have a pro-tumorigenic role in different types of tumors, including colorectal cancer (CRC), and are therapeutic targets for cancer treatment79,80,81. The pro-tumorigenic role of YAP and TAZ depends, at least in part, on their capacity to regulate P-body dynamics. Specifically, YAP/TAZ transcriptionally regulate the levels of mRNAs encoding for SMAD4A, AJUBA, and PNRC1 that accumulate within P-bodies. By controlling the levels of these transcripts, YAP/TAZ promote the assembly of P-bodies. In particular, YAP negatively regulates the expression of PNRC1, a tumor suppressor gene that inhibits P-body assembly. Disassembly of P-bodies induced by the downregulation of DDX6 or LSM14A attenuates the oncogenic role of YAP in colon cancer cells, affecting both proliferation and migration, indicating that P-body dynamics is crucial for tumorigenesis82.
A role of G protein-coupled receptor signaling cascade in P-body dynamics has been recently identified. Thus, activation of the cAMP-PKA pathway induces a non-proteolytic ubiquitylation of DDX6 by praja2, an E3 ligase involved in different biological processes, both under physiological and pathological conditions, including cancer83,84,85,86,87,88. Ubiquitylation of DDX6 by praja2 markedly impacts on P-body formation and protein translation in human glioblastoma (GBM) cells. In xenograft models of GBM, overexpression of a DDX6-defective ubiquitylation point mutant impairs the formation of P-bodies and accelerated tumor growth64. This study points to a relevant link between P-body dynamics and GBM growth, highlighting P-bodies as potential therapeutic targets for the treatment of this aggressive brain tumor.
Ubiquitylation of P-bodies resident proteins plays a relevant role also in CRC. As previously mentioned, in CRC cells, energy stress induced by low ATP levels promotes P-body formation and sustains tumor growth. Mechanistically, ATP depletion induces Trim23-mediated non-proteolytic ubiquitylation of HCLS1-associated protein X-1 (HAX1), a P-body component that binds the 3′UTR of mRNAs89,90. Ubiquitylation primes liquid–liquid phase separation of HAX1 that, in turn, induces condensation of DDX6 and LSM14A complexes and P-body assembly with consequent inhibition of protein synthesis. Depleting Trim23 inhibits P-body formation and restrains the proliferation of CRC cells52. Altogether, these studies point to a pathogenic role of signaling pathways and PTMs operating on P-body dynamics in the control of cancer growth (Fig. 3).

Under energy stress conditions, Trim23 ubiquitylates HAX1 that induces the binding between DDX6 and Lsm14A, favoring the assembly of P-bodies and colon rectal cancer growth. TGF-β promotes P-body assembly that positively correlates with breast cancer growth. cAMP induces the praja2-mediated ubiquitylation of DDX6 that stimulates P-body assembly and regulates glioblastoma. Phosphorylation of EDC3 by Pim1/3 kinases inhibits its localization at P-bodies, preventing their assembly and favoring the growth of prostate cancer. P-body disassembly causes protein translation of ADAR1 mRNA that, in turn, supports gastric cancer growth. Treatment with MEK inhibitors (MEKi) induces P-body dissolution and consequent translation of RAS mRNA that supports chemoresistance.
P-bodies, hypoxia, and metabolic reprogramming in cancer
Hypoxia is one of the main characteristics of solid tumors, and it is associated with a high grade of malignancy. Hypoxia is caused by a limited vascular oxygen supply within the tumoral tissue that is not proportional to the high rate of cell proliferation91. Under hypoxic conditions, cancer cells reprogram their metabolism by adopting aerobic glycolysis and lactic acid fermentation to produce ATP and biosynthetic intermediates. To promote the metabolic rewiring during hypoxia, cancer cells also modify their translational landscape92. In this context, P-bodies may play a relevant role, representing an important proteostatic mechanism for cancer cells to survive and proliferate under extreme environmental conditions.
Although a link between hypoxia, P-bodies, and cancer has still not well established, different studies suggest an involvement of mRNPs granules in hypoxia-induced translational and metabolic rewiring of tumor cells. The first functional connection with hypoxia derives from the identification of P-bodies as regulators of HIF1α mRNA fate. HIF1α is the master transcriptional regulator activated during hypoxia. The ubiquitin-specific protease 52 (UPS52), also known as poly(A) nuclease 2 (PAN2), is a component of P-bodies that interacts with- and targets HIF1α mRNA within the RNPs condensates. Localization at P-bodies prevents HIF1α mRNA degradation and promotes accumulation of HIF1A protein, leading to transcriptional upregulation of hypoxia-induced target genes. The disassembly of P-bodies by genetic downregulation of GW182 or LSM1 reduces the levels of HIF1α mRNA and attenuates the hypoxia-mediated translational reprogramming93. On the other hand, hypoxia can negatively regulate protein synthesis through a biphasic inhibition of mRNA translation by two distinct mechanisms. In the acute phase, hypoxia induces phosphorylation and inhibition of the eukaryotic translation initiation factor 2 A (eIF2a). Under prolonged hypoxia, disruption of cap-binding complex eIF4F and sequestration of eIF4E by its inhibitor 4E-BP1 and transporter 4E-T within P-bodies contributes to its reduced translation and regulation of hypoxic gene expression94. The biological effects of P-bodies on the translational reprogramming induced by hypoxia have also been demonstrated in cancer cells. In MCF7 breast cancer cells, DDX6 controls the translation of VEGF, a proangiogenic factor that promotes the formation of new vessels during hypoxia. Under normoxic conditions, DDX6 binds to- and localizes VEGF mRNA within DCP1a-containing granules, most likely P-bodies, thus inhibiting its translation. In the course of hypoxia, the decrease of DDX6 levels removes the inhibitory constraint and promotes VEGF translation95.
A link between hypoxia and P-bodies in the control of the metastatic spreading of cancer cells has been proposed. LIN28A is an RNA-binding protein upregulated in many tumors, and its levels correlate with a poor prognosis. In colon cancer cells, hypoxia increases LIN28A mRNA levels, rather than protein levels, and promotes its localization at P-bodies. Within these condensates, the translation of LIN28A mRNA is inhibited. Interestingly, proteomic analysis revealed that non-coding LIN28A mRNA upregulates the expression of methionyl aminopeptidase 2 (METAP2), a positive regulator of cancer metastasis. Concomitant downregulation of DICER, a ribonuclease involved in the production of miRNAs and siRNAs molecules, abolished the effects of non-coding LIN28A on METAP2 protein expression and cancer metastasis. These findings support a role of P-bodies in the control of a miRNA-mediated mechanism operating in metastatic cancer cells under hypoxia96. A relevant role for tumor growth under hypoxia is played by EXOSC9, a component of the RNA exosome complex involved in mRNA decay. Depletion of EXOSC9 in breast cancer cells inhibits proliferation under stress conditions97. Although not a resident of P-bodies, EXOSC9 regulates P-body dynamics since its genetic downregulation decreases the number of these condensates. At the mechanistic level, EXOSC9 regulates the decay of APOBEC34 mRNA that is located at P-bodies. Accumulation of this mRNA has a negative effect on P-body assembly, whereas its depletion promotes P-body formation. The assembly of P-bodies regulated by EXOSC9 positively correlates with breast cancer growth. Thus, EXOSC9 depletion attenuates P-body formation and drastically reduces tumor growth in xenograft models of breast cancer. Re-expressing wild-type EXOSC9, but not its mRNA binding mutant, unable to promote P-body condensation, restores cell proliferation and tumor growth97. These findings strongly support the role of P-bodies in the dynamic regulation of protein translation underlying growth and dissemination of cancer cells under stress conditions. Nevertheless, additional studies are needed to provide formal proof of the involvement of P-bodies in the metabolic rewiring of cancer cells under hypoxic or stress conditions and to mechanistically dissect the contribution of P-bodies in the control of global or selective mRNA translation.
A role of P-bodies in cancer stemness
Cancer stemness is one of the hallmarks of tumor development98. Cancer stem cells (CSCs) are responsible for tumor initiation, self-renewal, differentiation, and plasticity99. The RNA editing process has been identified as a mechanism of CSCs regulation and, in this context, a relevant role is played by the adenosine deaminase acting on RNA 1 (ADAR1), a member of the double-stranded RNA-specific adenosine deaminase family that converts adenosine to inosine in a double-stranded RNA substrate. ADAR1 exists in two isoforms, both involved in the editing of dsRNA: p100 isoform that is prevalently located in the nucleus, and the p150 principally located in the cytoplasm and inducible by interferon100. ADAR1, carrying out its activity in CSCs, plays a pro-tumorigenic role in chronic myeloid leukemia, glioblastoma, and liver cancer101,102,103. An oncogenic activity of ADAR1 has been identified in gastric cancer, where its overexpression promotes tumor growth. Therefore, inhibition of ADAR1 activity has been proposed as a therapeutic strategy for this type of malignant tumor104,105. Cytoplasmic ADAR1 protein was originally identified as a resident of stress granules106. Recent protein–protein interaction (PPI) network analysis demonstrated that ADAR1 p150 variant forms multimeric complexes with proteins mostly enriched in P-bodies, suggesting a role of this RNA editing enzyme also in P-bodies107. The translation of ADAR1 is regulated through the localization of its mRNA within P-bodies. The cytoplasmic polyadenylation element-binding protein 3 (CPEB3) is downregulated in gastric cancer tissue, and its expression correlates with a better prognosis for patients. The oncosuppressive effect of CPEB3 is related to its capacity to promote the localization and translational repression of ADAR1 mRNA within P-bodies. In mouse models of gastric cancer, overexpression of CPEB3 by adeno-associated vectors inhibits RNA editing by ADAR1 and restrains tumor growth108 (Fig. 3). Considering the critical role of ADAR1 in different types of CSCs, these findings support an involvement of P-bodies in the regulation of cancer stemness through the control of ADAR1 mRNA metabolism. In P-bodies, DDX6 binds to- and translationally represses mRNAs encoding for transcription factors that control the exit from the pluripotency state of CSCs109. Moreover, a transcriptomic profiling of coding and non-coding RNAs that accumulate in P-bodies shows differentially enriched transcripts at different stage of development in multiple vertebrate species. Importantly, this study revealed the essential role of P-bodies not much in the regulation of the global gene expression, but rather in controlling the accumulation and translational repression of transcripts relative to the previous developmental stage in a miRNA-dependent mechanism. In this context, the cell fate transition in pluripotent stem cells can be induced by regulating the selective sequestration of RNA in biomolecular condensates at P-bodies110. Currently, not enough studies have been conducted to provide formal evidence of the role of P-bodies in cancer stemness. However, the data available so far suggest that P-bodies, through the regulation of the stability and/or translation of specific transcripts, indeed play a crucial role in maintaining the stem-like properties characteristic of cancer cells.
P-bodies in cancer chemoresistance
Chemoresistance represents one of the most difficult challenges for cancer treatment, demanding the necessity of a deep understanding of the molecular basis of its insurgence that might contribute to develop new drugs that sensitize cancer cells to chemotherapeutic agents. Chemoresistance relies on different molecular mechanisms underlying DNA repair, regulation of oncogenes or oncosuppressors, EMT and cancer stemness111. The development of chemoresistance is linked to genetic and epigenetic mechanisms, and it is strictly dependent on post-transcriptional regulation of RNA binding proteins, miRNAs and components of the translational machinery66. To counteract the effects of chemotherapeutics, cancer cells take advantage from these mechanisms to regulate the translation rate of selected mRNAs involved in cell survival, EMT, drug metabolism and proliferation. In this context, given their capacity to regulate the stability and translation efficiency of specific mRNAs, P-bodies have a direct role in tumor drug resistance. Protein kinases MEK1/2 act as oncogenes in different types of cancer since they promote cell proliferation and survival, supporting tumor expansion and metastasis. About 30% of human cancers show uncontrolled activation of Ras/Raf/MEK/ERK pathway. Accordingly, potent and selective inhibitors of MEK (MEKi) are currently being used for the treatment of different types of tumors112,113. However, many of these tumors develop resistance to the chemotherapeutics and this is mostly associated with high levels of K-Ras and N-Ras proteins113,114. P-bodies play a fundamental role in the development of chemoresistance. In fact, mRNAs encoding for both Ras genes are located within P-bodies and in breast, lung and melanoma cancer cells, MEKi treatment induces the resumption of their translation by promoting P-body disassembly. Similar effects are obtained by genetic silencing of DDX6 that promotes P-body dissolution, and expression analysis in cancer tissue samples shows an inverse correlation between DDX6 and Ras protein levels115. The molecular mechanism by which MEKi induces P-body disassembly is unclear; however, considering the importance of PTMs in regulating P-body dynamics, we can assume that the MAPK pathway targets essential components of P-bodies, impacting on the dynamics of these mRNP condensates. Indeed, a phosphoproteomic analysis of ERK substrates identified 4E-T as an indirect target of MAPK116. Although the mechanisms through which MAPKi leads to drug resistance via P-bodies remain to be addressed, these findings are consistent with the model whereby translational regulation of mRNA at P-bodies in cancer cells controls the expression levels of Ras proteins which, in turn, support chemoresistance115 (Fig. 3). Therefore, understanding the mechanisms operating at P-bodies that control protein translation represents an important goal for the design of novel therapeutic strategies able to overcome chemoresistance of otherwise aggressive tumors.
Conclusions and perspectives
P-bodies are RNP condensates that play a fundamental role in the translational control of gene expression. By regulating mRNA storage or decay, P-bodies exert important biological effects underlying key cellular activities. Although P-bodies are constitutively present in cells, the assembly/disassembly of these condensates can be regulated by various signaling pathways and post-translational events that rapidly modify the rate of mRNA translation under different microenvironmental conditions. Cancer cells need to rapidly modify their proteome in response to metabolic changes or modifications of the tumor microenvironment that would otherwise limit cancer cell growth and dissemination. The regulation of P-body dynamics, thus, represents a fundamental mechanism for cancer cells to rapidly change their translational asset. In the last years, many studies have focused on the role of P-body dynamics in cancer growth and development, demonstrating the relevance of these condensates in cancer cell plasticity. This allows adaptation of proliferating cancer cells to hormone/growth factor stimulation or metabolic needs, promoting expansion of tumor mass and resistance to chemotherapeutics.
The role of P-bodies in regulating tumor growth, metastasis, and response to therapy is a relatively new field, and cumulative evidence supports the existence of a dynamic intersection between these consendates and oncogenic pathways that shapes the proteome of cancer cells. The link between P-bodies and tumors has been established in different types of human cancer. However, it is still debating whether the RNP condensates impact on the behavior of specific types of tumors or they have a more general implication in malignant cell transformation. A rapidly growing tumor is often associated with changes in the levels of RNPs condensates, but these alterations do not always reflect significant modifications of protein translation. Furthermore, the number of P-bodies do not reflect the aggressiveness potential or the type of malignant tumors, since breast cancer growth has been linked to both an increase and a decrease in the number of RNP condensates. Instead, it is most likely a consequence of the highly dynamic nature of P-bodies, which can rapidly assemble/disassemble in response to changes in the tumor microenvironment or nutrient availability. Also, the importance of P-bodies in tumor growth depends on the type of mRNAs they regulate, rather than the abundance of RNPs granules. In fact, although modifications of P-body dynamics in tumor cells are linked to changes of global protein translation, the regulation of specific mRNAs encoding for oncogenes or tumor suppressors by the RNPs condensates is likely more important. As example, studies investigating the role of the hypoxia-P-bodies axis in tumor cells revealed that, under hypoxic conditions, these biomolecular condensates regulate the fate of selected transcripts encoding for proteins involved in cancer cell growth. Similarly, the control of certain transcripts within P-bodies underlies the development of cancer chemoresistance. However, the precise biological role of RNPs condensates in drug resistance, and the molecular mechanisms involved, needs further in-depth investigation.
The current model points to an important role of P-bodies in various aspects of tumor cell biology, such as initiation, proliferation, stemness, development of metastases, stress response, and chemoresistance. However, it is still difficult to determine whether, and on which aspect, they have a greater impact. The main characteristic of P-bodies is the ability to dynamically regulate mRNAs, both through degradation or storage, rapidly and reversibly impacting on protein translation. For this reason, we believe that the role of P-bodies in tumor biology most likely relies on their ability to confer cell plasticity, a characteristic that supports adaptation of cancer cells to rapid changes in the tumor microenvironment, promoting genetic heterogeneity, cell growth, immune evasion, resistance to chemotherapeutics, and metastatic spreading. These aspects are of particular relevance in tumors where no recurrent mutations or molecular alterations driving carcinogenesis have been identified. In this context, P-body dynamics, by conferring plasticity to cancer cells, may contribute to tumor growth and expansion, thus representing relevant therapeutic targets.
In conclusion, this review highlighted the emerging role of P-bodies in cancer. Further studies will elucidate the molecular mechanisms underlying the P-body impact on cancer cell plasticity and will likely contribute to identify and target components and regulators of P-bodies, thus providing novel strategies for the treatment of otherwise aggressive malignant tumors.
References
Kim, S. & Kaang, B.-K. Epigenetic regulation and chromatin remodeling in learning and memory. Exp. Mol. Med. 49, e281–e281 (2017).
Wang, G. et al. Understanding transcription factor regulation by integrating gene expression and DNase I hypersensitive sites. BioMed. Res. Int. 2015, 1–7 (2015).
Reynolds, N., O’Shaughnessy, A. & Hendrich, B. Transcriptional repressors: multifaceted regulators of gene expression. Development 140, 505–512 (2013).
Corbett, A. H. Post-transcriptional regulation of gene expression and human disease. Curr. Opin. Cell Biol. 52, 96–104 (2018).
Peng, L., Li, E.-M. & Xu, L.-Y. From start to end: phase separation and transcriptional regulation. Biochim. Biophys. Acta Gene Regul. Mech. 1863, 194641 (2020).
Yu, Z. et al. Decoding the genomic landscape of chromatin-associated biomolecular condensates. Nat. Commun. 15, 6952 (2024).
Demmerle, J., Hao, S. & Cai, D. Transcriptional condensates and phase separation: condensing information across scales and mechanisms. Nucleus 14, 2213551 (2023).
Luo, Y., Na, Z. & Slavoff, S. A. P-bodies: composition, properties, and functions. Biochemistry 57, 2424–2431 (2018).
Eulalio, A., Behm-Ansmant, I. & Izaurralde, E. P bodies: at the crossroads of post-transcriptional pathways. Nat. Rev. Mol. Cell Biol. 8, 9–22 (2007).
Nsengimana, B. et al. Processing body (P-body) and its mediators in cancer. Mol. Cell Biochem. 477, 1217–1238 (2022).
Safieddine, A. et al. Cell-cycle-dependent mRNA localization in P-bodies. Mol. Cell 84, 4191–4208.e7 (2024).
Ren, Z., Zhao, S., Tang, W. & Zou, P. Spatially resolved multibait mapping of stress granule and processing body transcriptome. Anal. Chem. 97, 12767–12775 (2025).
Shan, T. et al. m6A modification negatively regulates translation by switching mRNA from polysome to P-body via IGF2BP3. Mol. Cell 83, 4494–4508.e6 (2023).
Fang, L. et al. Pooled CRISPR screening identifies P-bodies as repressors of cancer epithelial-mesenchymal transition. Cancer Res. 84, 659–674 (2024).
Mitchell, S. F. & Parker, R. Principles and properties of eukaryotic mRNPs. Mol. Cell 54, 547–558 (2014).
Guan, H. et al. Liquid-liquid phase separation of membrane-less condensates: from biogenesis to function. Front. Cell Dev. Biol. 13, 1600430 (2025).
Lavut, A. & Raveh, D. Sequestration of highly expressed mRNAs in cytoplasmic granules, P-bodies, and stress granules enhances cell viability. PLoS Genet. 8, e1002527 (2012).
Lee, M. et al. Optogenetic control of mRNA condensation reveals an intimate link between condensate material properties and functions. Nat. Commun. 15, 3216 (2024).
Khong, A. et al. The stress granule transcriptome reveals principles of mRNA accumulation in stress granules. Mol. Cell 68, 808–820.e5 (2017).
Sugawara, K. et al. Nanoscale dynamics and localization of single endogenous mRNAs in stress granules. Nucleic Acids Res. 52, 8675–8686 (2024).
Moon, S. L. et al. Multicolour single-molecule tracking of mRNA interactions with RNP granules. Nat. Cell Biol. 21, 162–168 (2019).
Liu, Z. et al. Liquid‒liquid phase separation: roles and implications in future cancer treatment. Int. J. Biol. Sci. 19, 4139–4156 (2023).
Dai, Z. & Yang, X. The regulation of liquid-liquid phase separated condensates containing nucleic acids. FEBS J. 291, 2320–2331 (2024).
Ma, W. & Mayr, C. A membraneless organelle associated with the endoplasmic reticulum enables 3’UTR-mediated protein-protein interactions. Cell 175, 1492–1506.e19 (2018).
Cioce, M. & Lamond, A. I. Cajal bodies: a long history of discovery. Annu Rev. Cell Dev. Biol. 21, 105–131 (2005).
Liu, J.-L. & Gall, J. G. U bodies are cytoplasmic structures that contain uridine-rich small nuclear ribonucleoproteins and associate with P bodies. Proc. Natl. Acad. Sci. USA 104, 11655–11659 (2007).
Maita, H. & Nakagawa, S. Balancing RNA processing and innate immune response: Possible roles for SMN condensates in snRNP biogenesis. Biochim. Biophys. Acta Gen. Subj. 1869, 130764 (2025).
Yang, Z. et al. GW182 is critical for the stability of GW bodies expressed during the cell cycle and cell proliferation. J. Cell Sci. 117, 5567–5578 (2004).
Lian, S. et al. GW Bodies, MicroRNAs and the Cell Cycle. Cell Cycle 5, 242–245 (2006).
Eystathioy, T. et al. A phosphorylated cytoplasmic autoantigen, GW182, associates with a unique population of human mRNAs within novel cytoplasmic speckles. Mol. Biol. Cell 13, 1338–1351 (2002).
Patel, P. H., Barbee, S. A. & Blankenship, J. T. GW-bodies and P-bodies constitute two separate pools of sequestered non-translating RNAs. PLoS ONE 11, e0150291 (2016).
Kedersha, N. et al. Stress granules and processing bodies are dynamically linked sites of mRNP remodeling. J. Cell Biol. 169, 871–884 (2005).
Ripin, N., Macedo de Vasconcelos, L., Ugay, D. A. & Parker, R. DDX6 modulates P-body and stress granule assembly, composition, and docking. J. Cell Biol. 223, e202306022 (2024).
Parker, R. & Sheth, U. P bodies and the control of mRNA translation and degradation. Mol. Cell 25, 635–646 (2007).
Hubstenberger, A. et al. P-body purification reveals the condensation of repressed mRNA regulons. Mol. Cell 68, 144–157.e5 (2017).
Youn, J.-Y. et al. High-density proximity mapping reveals the subcellular organization of mRNA-associated granules and bodies. Mol. Cell 69, 517–532.e11 (2018).
Liu, J., Valencia-Sanchez, M. A., Hannon, G. J. & Parker, R. MicroRNA-dependent localization of targeted mRNAs to mammalian P-bodies. Nat. Cell Biol. 7, 719–723 (2005).
Ford, L., Ling, E., Kandel, E. R. & Fioriti, L. CPEB3 inhibits translation of mRNA targets by localizing them to P bodies. Proc. Natl. Acad. Sci. USA 116, 18078–18087 (2019).
Andrei, M. A. et al. A role for eIF4E and eIF4E-transporter in targeting mRNPs to mammalian processing bodies. RNA 11, 717–727 (2005).
Ferraiuolo, M. A. et al. A role for the eIF4E-binding protein 4E-T in P-body formation and mRNA decay. J. Cell Biol. 170, 913–924 (2005).
Sheth, U. & Parker, R. Decapping and decay of messenger RNA occur in cytoplasmic processing bodies. Science 300, 805–808 (2003).
Brengues, M., Teixeira, D. & Parker, R. Movement of eukaryotic mRNAs between polysomes and cytoplasmic processing bodies. Science 310, 486–489 (2005).
Bhattacharyya, S. N., Habermacher, R., Martine, U., Closs, E. I. & Filipowicz, W. Relief of microRNA-mediated translational repression in human cells subjected to stress. Cell 125, 1111–1124 (2006).
Teixeira, D., Sheth, U., Valencia-Sanchez, M. A., Brengues, M. & Parker, R. Processing bodies require RNA for assembly and contain nontranslating mRNAs. RNA 11, 371–382 (2005).
Ayache, J. et al. P-body assembly requires DDX6 repression complexes rather than decay or Ataxin2/2L complexes. Mol. Biol. Cell 26, 2579–2595 (2015).
Kamenska, A. et al. The DDX6-4E-T interaction mediates translational repression and P-body assembly. Nucleic Acids Res. 44, 6318–6334 (2016).
Bloch, D. B., Sinow, C. O., Sauer, A. J. & Corman, B. H. P. Assembly and regulation of the mammalian mRNA processing body. PLoS ONE 18, e0282496 (2023).
Brothers, W. R., Ali, F., Kajjo, S. & Fabian, M. R. The EDC4-XRN1 interaction controls P-body dynamics to link mRNA decapping with decay. EMBO J. 42, e113933 (2023).
Cheng, Y.-H. et al. EDC4 C-terminal domain scaffolds P-body assembly and links P-body dynamics to p53-mediated tumor suppression. RNA 31, 1176–1194 (2025).
Cougot, N., Babajko, S. & Séraphin, B. Cytoplasmic foci are sites of mRNA decay in human cells. J. Cell Biol. 165, 31–40 (2004).
Cochard, A. et al. RNA at the surface of phase-separated condensates impacts their size and number. Biophys. J. 121, 1675–1690 (2022).
Zhan, W. et al. Energy stress promotes P-bodies formation via lysine-63-linked polyubiquitination of HAX1. EMBO J. 43, 2759–2788 (2024).
Ciancone, A. M. et al. Global profiling identifies a stress-responsive tyrosine site on EDC3 regulating biomolecular condensate formation. Cell Chem. Biol. 29, 1709–1720.e7 (2022).
Aizer, A. et al. Quantifying mRNA targeting to P bodies in living human cells reveals a dual role in mRNA decay and storage. J. Cell Sci. jcs.152975 https://doi.org/10.1242/jcs.152975.(2014)
Wang, C. et al. Context-dependent deposition and regulation of mRNAs in P-bodies. Elife 7, e29815 (2018).
Rzeczkowski, K. et al. c-Jun N-terminal kinase phosphorylates DCP1a to control formation of P bodies. J. Cell Biol. 194, 581–596 (2011).
Tenekeci, U. et al. K63-ubiquitylation and TRAF6 pathways regulate mammalian P-body formation and mRNA decapping. Mol. Cell 62, 943–957 (2016).
Wei, W. et al. TRIM24 is an insulin-responsive regulator of P-bodies. Nat. Commun. 13, 3972 (2022).
Kedia, S. et al. Ubiquitination and deubiquitination of 4E-T regulate neural progenitor cell maintenance and neurogenesis by controlling P-body formation. Cell Rep. 40, 111070 (2022).
Arribas-Layton, M., Dennis, J., Bennett, E. J., Damgaard, C. K. & Lykke-Andersen, J. The C-terminal RGG domain of human Lsm4 promotes processing body formation stimulated by arginine dimethylation. Mol. Cell Biol. 36, 2226–2235 (2016).
Cargnello, M. et al. Phosphorylation of the eukaryotic translation initiation factor 4E-transporter (4E-T) by c-Jun N-terminal kinase promotes stress-dependent P-body assembly. Mol. Cell. Biol. 32, 4572–4584 (2012).
Ramachandran, V., Shah, K. H. & Herman, P. K. The cAMP-dependent protein kinase signaling pathway is a key regulator of p body foci formation. Mol. Cell 43, 973–981 (2011).
Zhang, J. Z. et al. Phase separation of a PKA regulatory subunit controls cAMP Compartmentation and oncogenic signaling. Cell 182, 1531–1544.e15 (2020).
Senatore, E. et al. Praja2 controls P-body assembly and translation in glioblastoma by non-proteolytic ubiquitylation of DDX6. EMBO Rep. 26, 2347–2377 (2025)
Pérez-González, A., Bévant, K. & Blanpain, C. Cancer cell plasticity during tumor progression, metastasis and response to therapy. Nat. Cancer 4, 1063–1082 (2023).
Fabbri, L., Chakraborty, A., Robert, C. & Vagner, S. The plasticity of mRNA translation during cancer progression and therapy resistance. Nat. Rev. Cancer 21, 558–577 (2021).
Hernández, G. et al. Decapping protein EDC4 regulates DNA repair and phenocopies BRCA1. Nat. Commun. 9, 967 (2018).
Wu, C. et al. Overexpression of mRNA‑decapping enzyme 1a affects survival rate in colorectal carcinoma. Oncol. Lett. https://doi.org/10.3892/ol.2018.8730 (2018).
Cougot, N. et al. MLN51 triggers P-body disassembly and formation of a new type of RNA granules. J. Cell Sci. 127, 4692–701 (2014).
Kodali, S. et al. RNA sequestration in P-bodies sustains myeloid leukaemia. Nat. Cell Biol. 26, 1745–1758 (2024).
Sanchez-Vega, F. et al. Oncogenic signaling pathways in the Cancer Genome Atlas. Cell 173, 321–337.e10 (2018).
Bearss, J. J. et al. EDC3 phosphorylation regulates growth and invasion through controlling P-body formation and dynamics. EMBO Rep. 22, e50835 (2021).
Gnawali, G. R. et al. Synthesis of 2-oxoquinoline derivatives as dual pim and mTORC protein kinase inhibitors. Med. Chem. Res. 31, 1154–1175 (2022).
Larance, M. et al. Global phosphoproteomics identifies a major role for AKT and 14-3-3 in regulating EDC3. Mol. Cell Proteom. 9, 682–694 (2010).
Xu, J., Lamouille, S. & Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 19, 156–172 (2009).
Pang, M.-F. et al. TGF-β1-induced EMT promotes targeted migration of breast cancer cells through the lymphatic system by the activation of CCR7/CCL21-mediated chemotaxis. Oncogene 35, 748–760 (2016).
Hardy, S. D., Shinde, A., Wang, W.-H., Wendt, M. K. & Geahlen, R. L. Regulation of epithelial-mesenchymal transition and metastasis by TGF-β, P-bodies, and autophagy. Oncotarget 8, 103302–103314 (2017).
Fu, M. et al. The Hippo signalling pathway and its implications in human health and diseases. Signal Transduct. Target Ther. 7, 376 (2022).
Xu, Z., Wang, H., Gao, L., Zhang, H. & Wang, X. YAP levels combined with plasma CEA levels are prognostic biomarkers for early-clinical-stage patients of colorectal cancer. Biomed. Res. Int. 2019, 2170830 (2019).
Yang, W. et al. Influence of the Hippo-YAP signalling pathway on tumor associated macrophages (TAMs) and its implications on cancer immunosuppressive microenvironment. Ann. Transl. Med. 8, 399 (2020).
Piccolo, S., Panciera, T., Contessotto, P. & Cordenonsi, M. YAP/TAZ as master regulators in cancer: modulation, function and therapeutic approaches. Nat. Cancer 4, 9–26 (2022).
Shen, X. et al. YAP/TAZ enhances P-body formation to promote tumorigenesis. eLife 12, RP88573 (2024).
Lignitto, L. et al. Control of PKA stability and signalling by the RING ligase praja2. Nat. Cell Biol. 13, 412–422 (2011).
Lignitto, L. et al. Proteolysis of MOB1 by the ubiquitin ligase praja2 attenuates Hippo signalling and supports glioblastoma growth. Nat. Commun. 4, 1822 (2013).
Rinaldi, L. et al. praja2 regulates KSR1 stability and mitogenic signaling. Cell Death Dis. 7, e2230 (2016).
Rinaldi, L. et al. Downregulation of praja2 restrains endocytosis and boosts tyrosine kinase receptors in kidney cancer. Commun. Biol. 7, 208 (2024).
Senatore, E. et al. The TBC1D31/praja2 complex controls primary ciliogenesis through PKA-directed OFD1 ubiquitylation. EMBO J. 40, e106503 (2021).
Delle Donne, R. et al. Targeted inhibition of ubiquitin signaling reverses the metabolic reprogramming and suppresses glioblastoma growth. Commun. Biol. 5, 780 (2022).
Grzybowska, E. A. et al. HAX-1 is a nucleocytoplasmic shuttling protein with a possible role in mRNA processing. FEBS J. 280, 256–272 (2013).
Zayat, V. et al. HAX-1: a novel p-body protein. DNA Cell Biol. 34, 43–54 (2015).
Zhuang, Y. et al. Hypoxia signaling in cancer: implications for therapeutic interventions. MedComm 4, e203 (2023).
Al Tameemi, W., Dale, T. P., Al-Jumaily, R. M. K. & Forsyth, N. R. Hypoxia-modified cancer cell metabolism. Front. Cell Dev. Biol. 7, 4 (2019).
Bett, J. S. et al. The P-body component USP52/PAN2 is a novel regulator of HIF1A mRNA stability. Biochem. J. 451, 185–194 (2013).
Koritzinsky, M. et al. Gene expression during acute and prolonged hypoxia is regulated by distinct mechanisms of translational control. EMBO J. 25, 1114–1125 (2006).
de Vries, S. et al. Identification of DEAD-box RNA helicase 6 (DDX6) as a cellular modulator of vascular endothelial growth factor expression under hypoxia. J. Biol. Chem. 288, 5815–5827 (2013).
Weng, M. et al. Hypoxia-induced LIN28A mRNA promotes the metastasis of colon cancer in a protein-coding-independent manner. Front. Cell Dev. Biol. 9, 642930 (2021).
Yoshino, S. et al. EXOSC9 depletion attenuates P-body formation, stress resistance, and tumorigenicity of cancer cells. Sci. Rep. 10, 9275 (2020).
Jing, N., Gao, W.-Q. & Fang, Y.-X. Regulation of formation, stemness and therapeutic resistance of cancer stem cells. Front. Cell Dev. Biol. 9, 641498 (2021).
Loh, J.-J. & Ma, S. Hallmarks of cancer stemness. Cell Stem Cell 31, 617–639 (2024).
Rehwinkel, J. & Mehdipour, P. ADAR1: from basic mechanisms to inhibitors. Trends Cell Biol. 35, 59–73 (2025).
Jiang, Q. et al. ADAR1 promotes malignant progenitor reprogramming in chronic myeloid leukemia. Proc. Natl. Acad. Sci. USA 110, 1041–1046 (2013).
Jiang, L. et al. ADAR1-mediated RNA editing links ganglioside catabolism to glioblastoma stem cell maintenance. J. Clin. Investig. 132, e143397 (2022).
Luo, J. et al. Enhanced mitophagy driven by ADAR1-GLI1 editing supports the self-renewal of cancer stem cells in HCC. Hepatology 79, 61–78 (2024).
Chan, T. H. M. et al. ADAR-mediated RNA editing predicts progression and prognosis of gastric cancer. Gastroenterology 151, 637–650.e10 (2016).
Dou, N. et al. Aberrant overexpression of ADAR1 promotes gastric cancer progression by activating mTOR/p70S6K signaling. Oncotarget 7, 86161–86173 (2016).
Weissbach, R. & Scadden, A. D. J. Tudor-SN and ADAR1 are components of cytoplasmic stress granules. RNA 18, 462–471 (2012).
Vukić, D. et al. Distinct interactomes of ADAR1 nuclear and cytoplasmic protein isoforms and their responses to interferon induction. Nucleic Acids Res. 52, 14184–14204 (2024).
Chen, J. et al. CPEB3 suppresses gastric cancer progression by inhibiting ADAR1-mediated RNA editing via localizing ADAR1 mRNA to P bodies. Oncogene 41, 4591–4605 (2022).
Di Stefano, B. et al. The RNA helicase DDX6 controls cellular plasticity by modulating P-body homeostasis. Cell Stem Cell 25, 622–638.e13 (2019).
Pessina, P. et al. Selective RNA sequestration in biomolecular condensates directs cell fate transitions. Nat. Biotechnol. https://doi.org/10.1038/s41587-025-02853-z. Published online (2025).
Zheng, H.-C. The molecular mechanisms of chemoresistance in cancers. Oncotarget 8, 59950–59964 (2017).
Wu, P.-K. & Park, J.-I. MEK1/2 inhibitors: molecular activity and resistance mechanisms. Semin. Oncol. 42, 849–862 (2015).
Wang, C. et al. Research progress of MEK1/2 inhibitors and degraders in the treatment of cancer. Eur. J. Med. Chem. 218, 113386 (2021).
Tsai, Y. S. et al. Rapid idiosyncratic mechanisms of clinical resistance to KRAS G12C inhibition. J. Clin. Investig. 132, e155523 (2022).
Vidal-Cruchez, O. et al. KRAS and NRAS translation is increased upon MEK inhibitors-induced processing bodies dissolution. Cancers 15, 3078 (2023).
Ünal, E. B., Uhlitz, F. & Blüthgen, N. A compendium of ERK targets. FEBS Lett. 591, 2607–2615 (2017).
Acknowledgements
This work was supported by Fondazione AIRC per la Ricerca sul Cancro (IG2023-29124), the Italian Ministry of University and Research (National Center for Gene Therapy and Drugs based on RNA Technology, PNRR-CN3: E63C22000940007; PRIN2022: E53D23009690006 and E53D23021760001, MNESYS-A multiscale integrated approach to the study of the nervous system in health and disease’ PE0000006, DN.1553), European Regional Development Fund (POR Campania FESR 2021-2027), grant “RARE.Glials” to AF. The authors apologize to those whose work was not cited owing to space limitations.
Author information
Authors and Affiliations
Contributions
E.S. and A.F. wrote the article with contributions from L.R., F.C., and A.G.B. E.S. prepared the figures.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Leyla Fahim, Patrick Brest and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Johannes Stortz. [A peer review file is available].
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Senatore, E., Rinaldi, L., Chiuso, F. et al. Reshaping cancer cell plasticity by P-body dynamics and protein translation. Commun Biol 9, 160 (2026). https://doi.org/10.1038/s42003-025-09439-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-025-09439-0
