
Abstract
Obesity increases the risk of male infertility, primarily attributable to reduced testosterone levels. Exosomes, which facilitate intercellular communication during reproduction, may influence this process. However, the relation between exosomal cargo changes and high-fat diet (HFD)-induced testosterone decrease remains unclear. Here, we show that exosomes influence testosterone synthesis and spermatogenesis in HFD mice. Transferring exosomes derived from the serum of HFD mice to mice fed a normal diet decreases testosterone levels and sperm counts. Treatment with inhibitors of exosomes (GW4869) and ferroptosis (Ferrostatin-1) rescue HFD-induced impaired spermatogenesis. Additionally, elevated miR-122-5p levels in serum exosomes from HFD mice is partially attributed to increased hepatic miR-122-5p expression. Exosomal miR-122-5p induces ferroptosis in Leydig cells by inhibiting stearyl-CoA desaturase 2 expression, reducing testosterone synthesis and impairing spermatogenesis. Collectively, these findings highlight the impact of liver-derived exosomal alterations on testosterone production in HFD, revealing a regulatory pathway in the liver-testes axis.
Similar content being viewed by others

Introduction
The prevalence of obesity has grown exponentially over the last 50 years, reaching global epidemic levels. Obesity negatively impacts organs and physiological functions1, increasing the risk of cardiovascular diseases, reproductive disorders, cancers.By 2025, global obesity prevalence will reach 18% in men and 21% in women2. Notably, obesity-related male infertility is also on the rise3, prompting increased research interest on the impact of obesity on the male reproductive system.
Obesity contributes to several reproductive issues, including decreased sperm quality, erectile dysfunction, and prostatitis4. Although the underlying mechanisms are complex, reduced testosterone levels appear to be the primary consequence of obesity on the male reproductive system5. Although growing evidence links obesity to lower testosterone levels6, the underlying mechanisms have not been fully elucidated.
Exosomes are extracellular vesicles secreted by various cell types and are present in the body fluids of most organisms7. In disease, the composition and concentration of circulating exosomes may be altered8. Consequently, exosomes represent a potential modality for diagnosing and treating male infertility. Our previous research demonstrated that enteritis-induced dysbiosis of the gut microbiome increases the levels of intestine-derived miR-211-5p in circulating exosomes, disrupting meiotic processes in the testes and impairing spermatogenesis9. However, it remains unclear whether exosomes contribute to the occurrence and progression of obesity-associated male infertility and which molecules and target cells are involved in these pathogenic effects.
The liver is a key hub for endocrine control and lipid and cholesterol homeostasis10. Hepatic steatosis and chronic low-grade inflammation caused by obesity are risk factors for liver disease11. Furthermore, liver disorders, such as nonalcoholic fatty liver disease, toxic cirrhosis, and cholestatic disorders, are known to reduce testosterone levels12. While the molecular mechanisms through which the liver regulates testicular function remain unclear. Additionally, it is unknown whether hepatic exosomes contribute to testosterone regulation in obesity.
The aim of this study was to investigate the mechanisms through which exosomes affect testosterone synthesis and spermatogenesis in high-fat diet (HFD) fed mice. Collectively, this study highlights the importance of liver-derived exosomes in obesity-related spermatogenesis disorders and identifies targets for developing effective pharmaceuticals and therapies for the targeted treatment of male infertility.
Results
HFD-circulating exosomes impair testosterone synthesis and spermatogenesis
A mouse model of HFD-induced obesity was successfully established (Fig. 1A), as indicated by increased body weight (Supplementary Fig. 1A), insulin resistance (Supplementary Fig. 1B, C), and glucose intolerance (Supplementary Fig. 1D, E) in HFD mice compared to ND controls. The sperm count, diameter of seminiferous tubules, number of seminiferous tubules, and serum testosterone level were significantly lower in HFD mice than in ND mice (Fig. 1B–E). To analyze the effects of an HFD on testicular cells, serine/threonine kinase 31 (STK31), antizyme inhibitor 2 (AZIN2), cytochrome P450 17A1 (CYP17A1), hydroxysteroid dehydrogenase, and steroidogenic acute regulatory protein (STAR) were assessed as markers of spermatogonia, round spermatids, Leydig cells, and testosterone synthesis, respectively13,14. Hematoxylin-eosin staining (HE) and immunohistochemistry (IHC) revealed reduced numbers of Leydig cells, spermatogonia, and round spermatids in HFD mice compared with ND mice (Fig. 1F). Additionally, the expression of STK31, AZIN2, CYP17A1, STAR, hydroxy-delta-5-steroid dehydrogenase, 3 beta- and steroid delta-isomerase 6 (Hsd3b6), and hydroxysteroid 17-beta dehydrogenase 7 (Hsd17b7) were downregulated in the testes of the HFD group (Fig. 1G, H). These findings indicate that HFD-induced obesity impairs testosterone synthesis and spermatogenesis.

A Schematic diagram of the HFD model (n = 8). B Sperm count in the HFD model (n = 8). C Diameter of seminiferous tubules in the HFD model (n = 30). D Number of seminiferous tubules in the HFD model (n = 5). E Relative testosterone concentration in serum of the HFD model (n = 6). F HE and IHC staining of testes in the HFD model (Scale bar = 200 or 50 µm). G Relative mRNA levels of Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, and Star in the testes of mice from the HFD model (n = 3). H Relative protein levels of STK31, AZIN2, CYP17A1, and STAR in the testes of mice from the HFD model (n = 3). I Sperm count (n = 7). J Diameter of seminiferous tubules (n = 30). K Number of seminiferous tubules (n = 5). L Relative testosterone concentration in serum (n = 8). M Relative mRNA levels of Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, and Star in the testes of mice (n = 3). N Relative protein levels of STK31, AZIN2, CYP17A1, and STAR in the testes of mice from different exosome treatment groups (n = 3). O HE and IHC staining of testes in different exosome treatment groups (Scale bar = 200 or 50 µm). P Sperm count (n = 6). Q Diameter of seminiferous tubules in GW4869-treated HFD mice (n = 30). R Number of seminiferous tubules in GW4869-treated HFD mice (n = 5). S Relative testosterone concentration in serum (n = 3). T HE and IHC staining of testes in GW4869-treated HFD mice (Scale bar = 200 or 50 µm). U Relative mRNA levels of Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, and Star in the testes of GW4869-treated HFD mice (n = 3). V Relative protein levels of STK31, AZIN2, CYP17A1, and STAR in the testes of GW4869-treated HFD mice (n = 3). Data information: Results are expressed as mean values ± SD. Data were analyzed using an unpaired Student’ s t-test or one-way ANOVA. *p < 0.05, **p < 0.01.
To investigate the effects of circulating exosomes from HFD mice on spermatogenesis, exosomes were isolated from the serum of HFD mice (HFD-EXO) and ND mice (ND-EXO) by differential centrifugation. Exosomes were characterized using transmission electron microscopy (TEM), nanoflow, and western blotting (WB) (Supplementary Fig. 1G–I). Exosomes (ND-EXO or HFD-EXO) were injected into ND mice via the tail vein. Those receiving HFD-EXO exhibited glucose intolerance and insulin resistance (Supplementary Fig. 1L–O), mimicking the metabolic disorder phenotype of HFD-fed mice. Moreover, sperm count, diameter of seminiferous tubules, number of seminiferous tubules, and serum testosterone levels (Fig. 1I–L) were significantly decreased in the HFD-EXO group compared with the Control and ND-EXO groups. The results of RT-qPCR (Fig. 1M), WB (Fig. 1N), and HE & IHC (Fig. 1O) further revealed reduced STK31, AZIN2, CYP17A1, STAR, Hsd3b6, and Hsd17b7 expression in the testes of HFD-EXO mice, indicating that circulating exosomes from HFD mice induced abnormal testosterone synthesis and spermatogenesis.
To determine whether the HFD-induced impairment of spermatogenesis depends on circulating exosomes, HFD mice were injected with a noncompetitive sphingomyelinase inhibitor (GW4869) to inhibit exosome release. Mice in the HFD supplemented with GW4869 (HFD + GW) group exhibited significantly reduced body weight and testicular weight compared to the HFD group (Supplementary Fig. 1P, Q). Sperm count, diameter of seminiferous tubules, number of seminiferous tubules, and serum testosterone levels were significantly increased in HFD + GW mice compared to the HFD group (Fig. 1P–S). Moreover, GW4869 treatment rescued the HFD-induced loss of testicular Leydig cells, spermatogonia, and round spermatids (Fig. 1T), and reversed the downregulation of STK31, AZIN2, CYP17A1, STAR, Hsd3b6, and Hsd17b7 in the testes (Fig. 1U, V). These findings suggest that circulating exosomes play a key role in mediating the detrimental effects of HFD on reproductive function, and their inhibition can alleviate these effects.
Altered miRNA expression in HFD-circulating exosomes targets Leydig cells
To identify which cell types in the testes are affected by HFD, we analyzed scATAC-seq datasets of testes from ND and HFD mice, integrating this data with our previously published scRNA-seq data15 to establish the transcriptomic landscape and intercellular communication network (Fig. 2A). Using known marker genes, 12 distinct types were identified: spermatogonia (SPG), early spermatocytes (ESC), late spermatocytes (LSC), round sperm cells (RS), elongated sperm (ES), concentrate sperm (CS), Leydig cells (LC), Sertoli cells, macrophages, innate lymphoid cells, endothelial cells, and myoid cells (Fig. 2B, D, Supplementary Fig. 2A, B). In the HFD group, the number of SPG, RS, ES, LC, and Sertoli cells were decreased compared to the ND group (Fig. 2C, E). Moreover, our previous study demonstrated that testicular Leydig cells in HFD mice exhibited the highest proportion of differentially expressed genes (94.8%) and significantly elevated transcriptional noise compared to ND mice15. These results suggest that Leydig cells are the major cell type affected by HFD. Furthermore, differential analysis of Leydig cells in scRNA-seq and ATAC-seq revealed significant enrichment in several KEGG pathways, including steroid hormone biosynthesis, fatty acid metabolism, biosynthesis of unsaturated fatty acids, and fatty acid biosynthesis (Supplementary Fig. 2C, D).

A Schematic diagram of the experimental flow. B UMAP plot of testicular cell populations determined by scRNA-seq analysis. C Number of testicular cell types determined by scRNA-seq analysis. D UMAP plot of testicular cell populations identified by scATAC-seq analysis. E Number of testicular cell types determined by scATAC-seq analysis. F Venn diagrams comparing miRNA transcriptome profiling of ND-EXO and HFD-EXO. G Correlation analysis of miRNA transcriptomic profiling. H GO analysis of exosomal differential miRNA target genes. I Relative ferrous ion levels in the testes of the HFD model (n = 6). J Relative MDA levels in the testes of the HFD model (n = 3). K Relative ferrous ion levels in the testes of the exosome treatment mice (n = 3). L Relative MDA levels in the testes of the exosome treatment mice (n = 4) M Relative ferrous ion levels in the testes of GW4869-treated HFD mice (n = 3). N Relative MDA levels in the testes of GW4869-treated HFD mice (n = 3). Data information: Results are expressed as mean values ± SD. Data were analyzed using an unpaired Student’s t-test or one-way ANOVA. *p < 0.05, **p < 0.01.
To assess HFD-induced changes in the miRNA cargo of circulating exosomes, we compared the miRNA expression profiles of ND-EXO and HFD-EXO. This analysis revealed 26 upregulated miRNAs and 132 downregulated miRNAs in HFD-EXO compared to ND-EXO (Fig. 2F, G). Of these, 55 miRNAs with high and significant differential expression (p < 0.001, |log2FC| > 1) were selected for target gene prediction. Gene Ontology (GO) analysis revealed that these miRNAs were primarily enriched in lipid metabolism processes, ion transport, mitochondria, and metal ion binding (Fig. 2H). Based on these findings, we hypothesized that the altered miRNA composition of HFD-EXO could induce ferroptosis in Leydig cells, impairing testosterone synthesis and spermatogenesis.
To test this hypothesis, we measured malonaldehyde (MDA) and Fe2+ levels in the testes. The concentrations of both MDA and Fe2+ were significantly increased in the testes of the HFD group compared with the ND group. Treatment with GW4869 significantly reduced these levels (Fig. 2I, J, M, N). Similarly, MDA and Fe2+ levels were significantly higher in the testes of HFD-EXO mice compared with the Control and ND-EXO groups (Fig. 2K, L). These findings indicate that the testes of HFD and HFD-EXO mice exhibit characteristics of ferroptosis.
To identify the downstream targets of the miRNAs impacting Leydig cell function in HFD-EXO, a joint analysis of the miRNA target genes was conducted using the scRNA-seq and scATAC-seq data. The molecular interaction network (Supplementary Fig. 2E, F) revealed an association between Scd2 and pathways related to fatty acid biosynthesis, steroid synthesis, and mitochondria. Moreover, HFD decreased the 0 Cluster of Leydig cells (L0) (Supplementary Fig. 2G), which showed high Scd2 expression (Supplementary Fig. 2H). This suggests that L0 Leydig cells may be particularly sensitive to reduced Scd2 expression. However, the precise mechanism by which HFD affects Scd2 expression and ferroptosis in Leydig cells remains unclear, prompting us to focus on the regulatory pathways of ferroptosis.
HFD and HFD-circulating exosomes promote Leydig cell ferroptosis
To further verify the association between HFD-induced impaired spermatogenesis and ferroptosis, mice were injected with a ferroptosis inhibitor (ferrostatin-1, Fer-1) while being fed an HFD (Fig. 3A). Fer-1 supplementation did not affect body weight (Supplementary Fig. 3A). However, sperm count, diameter of seminiferous tubules, number of seminiferous tubules, and serum testosterone levels were significantly improved in the HFD supplemented with Ferrostatin-1 (HFD+Fer-1) group compared with the HFD group (Fig. 3B–E). Fer-1 treatment prevented the HFD-induced reduction in the number of Leydig cells, spermatogonia, and round spermatids (Fig. 3F), and alleviated the decrease in the expression of STK31, AZIN2, CYP17A1, STAR, Hsd3b6, and Hsd17b7 in the testes (Fig. 3G, H). Additionally, the concentration of MDA and Fe2+ were significantly lower in the testes of the HFD+Fer-1 group compared with the HFD group (Fig. 3I, J). TEM analysis revealed that HFD caused mitochondrial damage, including the presence of numerous vacuoles and loss of cristae. However, Fer-1 treatment partially rescued these mitochondrial abnormalities (Fig. 3K). Thus, administering Fer-1 alleviated HFD-induced ferroptosis and restored the testosterone level, sperm count, and Leydig cell, spermatogonia, and round spermatid abundances in the testes.

A Schematic diagram of HFD mice treated with Fer-1 (n = 6). B Sperm count of HFD mice treated with Fer-1 (n = 6). C Diameter of seminiferous tubules (n = 30). D Number of seminiferous tubules (n = 5). E Relative testosterone concentration in serum of HFD mice treated with Fer-1 (n = 3). F HE and IHC staining of testes in HFD mice treated with Fer-1 (Scale bar = 200 or 50 µm). G Relative protein levels of STK31, AZIN2, CYP17A1, and STAR in the testes of HFD mice treated with Fer-1 (n = 3). H Relative Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, and Star mRNA levels in the testes of HFD mice treated with Fer-1 (n = 3). I Relative MDA levels in the testes of HFD mice treated with Fer-1 (n = 3). J Relative ferrous ion levels in the testes of HFD mice treated with Fer-1 (n = 3). K TEM of mitochondria in the testes from HFD mice treated with Fer-1 (Scale bar = 500 nm). L Leydig cell viability (n = 6). M Relative ferrous ion levels of Leydig cells (n = 3). N-a MMP of Leydig cells (n = 3), N-b Fluorescence intensity of the sample (Scale bar = 100 µm). O Relative MDA levels of Leydig cells (n = 3). P Relative testosterone concentration in Leydig cell supernatants (n = 3). Q-a Relative ROS levels of Leydig cells (n = 3), Q-b Fluorescence intensity of the sample (Scale bar = 100 µm). Data information: Results are expressed as mean values ± SD. Data were analyzed using one-way ANOVA. *p < 0.05, **p < 0.01. Significance between groups was indicated by labeling with different letters; data labeled with the same letter indicate no significant difference.
Subsequently, to determine whether HFD-EXO directly promotes ferroptosis in Leydig cells, primary Leydig cells and TM3 cells were treated with HFD-EXO and Fer-1 in vitro. HFD-EXO significantly decreased Leydig cell viability, which was prevented in the HFD-EXO+Fer-1 group (Fig. 3L, Supplementary Fig. 3D), suggesting that Fer-1 exerts a protective against HFD-EXO-induced cell death. Additionally, the Fe2+ concentration, MDA concentration, and ROS levels were significantly increased in Leydig cells of the HFD-EXO group compared to the Control and ND-EXO groups, while Fer-1 treatment suppressed these effects (Fig. 3M, O, Q, Supplementary Fig. 3E, G, I). HFD-EXO also caused a decrease in mitochondrial membrane potential (MMP) (Fig. 3N, Supplementary Fig. 3F) and testosterone levels (Fig. 3P, Supplementary Fig. 3H), which were restored HFD-EXO+Fer-1 group.
Overall, these results demonstrated that Fer-1 alleviates HFD-EXO-induced ferroptosis in Leydig cells, and that ferroptosis plays a key role in reducing testosterone synthesis and impairing spermatogenesis caused by HFD-altered circulating exosomes.
Elevated miR-122-5p expression in HFD-circulating exosomes promotes Leydig cell ferroptosis
Differential miRNA expression profiling and RT-qPCR results revealed a significant upregulation of miR-122-5p in circulating exosomes from HFD mice (HFD-EXO) compared to those from the ND mice (ND-EXO) (Supplementary Fig. 4A). This increase in miR-122-5p expression was also observed in the testes of HFD mice compared with ND mice, with GW4869 treatment suppressing this upregulation (Supplementary Fig. 4B). miR-122-5p was also significantly elevated in the testes of HFD-EXO mice compared with the Control and ND-EXO groups (Supplementary Fig. 4C).
To further investigate the role of miR-122-5p in regulating ferroptosis in Leydig cells, these cells were treated with a mimics-miR-122-5p. Overexpression of miR-122-5p significantly decreased cell viability (Supplementary Fig. 4D, J), MMP (Supplementary Fig. 4F, L), and testosterone levels in the culture supernatant (Supplementary Fig. 4H, N) compared with mimics-NC. In addition, mimics-miR-122-5p treatment induced significant increases in the levels of Fe2+ (Supplementary Fig. 4E, K), MDA (Supplementary Fig. 4G, M), and ROS (Supplementary Fig. 4I, O) in Leydig cells.
To further elucidate the role of miR-122-5p in exosomes during Leydig cell ferroptosis, HFD-EXO treated with an miR-122-5p antagomir were added to the cell culture medium. Treatment with the miR-122-5p antagomir improved cell viability (Fig. 4A, Supplementary Fig. 4P), MMP (Fig. 4C, Supplementary Fig. 4R), and testosterone levels (Fig. 4E, Supplementary Fig. 4T) in the culture supernatant compared with the NC antago-HE group. The antagomir also alleviated the HFD-EXO-induced increase in Fe2+, MDA, and ROS in Leydig cells (Fig. 4B, D, F, Supplementary Fig. 4Q, S, U), highlighting the importance of miR-122-5p in HFD-EXO-promoted Leydig cell ferroptosis.

A Leydig cell viability (n = 6). B Relative ferrous ion levels of Leydig cells (n = 3). C-a MMP of Leydig cells (n = 3), C-b Fluorescence intensity of the sample (Scale bar = 100 µm). D Relative MDA levels of Leydig cells (n = 3). E Relative T concentration in Leydig cell supernatants (n = 3). F-a Relative ROS levels of Leydig cells (n = 3), F-b Fluorescence intensity of the samples (Scale bar = 100 µm). G Schematic representation of the 122-5p antago-HE-treated mice (n = 5). H Sperm count (n = 5). I Diameter of seminiferous tubules (n = 30). J Number of seminiferous tubules (n = 5). K Relative T concentration in serum (n = 3). L Relative Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, and Star mRNA levels in the testes of mice from different treatment groups (n = 3). M HE and IHC staining of testes in different treatment groups (Scale bar = 200 or 50 µm). N Relative STK31, AZIN2, CYP17A1, and STAR protein levels in the testes of mice from different treatment groups (n = 3). O Relative ferrous ion levels in the testes (n = 3). P Relative MDA levels in the testes (n = 3). Q TEM of mitochondria in the testes from different treatment groups (Scale bar = 1 µm). Data information: Results are expressed as mean values ± SD. Data were analyzed using an unpaired Student’s t-test or one-way ANOVA. *p < 0.05, **p < 0.01.
Thereafter, the role of exosomal miR-122-5p in regulating testosterone synthesis and spermatogenesis was assessed in vivo (Fig. 4G). As shown in Supplementary Fig. 4X, Y, 122-5p antago-HE mice still exhibited insulin resistance and impaired glucose tolerance. However, the miR-122-5p antagomir-treated exosomes alleviated the HFD-EXO-induced decrease in sperm count, diameter of seminiferous tubules, number of seminiferous tubules, and serum testosterone in mice (Fig. 4H–K). Compared with the NC antago-HE group, 122-5p antago-HE mice exhibited increased numbers of Leydig cells, SPG, and RS (Fig. 4M), and increased expression of STK31, AZIN2, CYP17A1, STAR, Hsd3b6, and Hsd17b7 in the testes (Fig. 4L, N). Additionally, the Fe2+ and MDA levels were reduced in the testes of 122-5p antago-HE mice compared with the NC antago-HE group (Fig. 4O, P), and mitochondrial morphology showed improvements, including reduced swelling, increased mitochondrial cristae, and fewer vacuoles (Fig. 4Q). These findings indicate that suppression of miR-122-5p in HFD-EXO did not significantly alleviate systemic metabolic disorders in mice, but improved testosterone secretion and spermatogenesis by mitigating ferroptosis in testicular Leydig cells.
miR-122-5p in HFD-circulating exosomes is primarily derived from the liver
To explore the origin of the HFD-induced increase in miR-122-5p in circulating exosomes, the expression levels of miRNAs were assessed in various organs of ND and HFD mice. miR-122-5p was found to be most abundantly expressed in the liver of HFD mice (Supplementary Fig. 5A). Liver-derived exosomes (LExo) of HFD mice via differential centrifugation were verified for purity using TEM, nanoflow, and WB (Supplementary Fig. 5B–D). The expression of miR-122-5p was significantly higher in HFD-LExo than ND-LExo (Supplementary Fig. 5E). This suggests that the increased expression of miR-122-5p observed in circulating exosomes from HFD mice may be attributed to increased miR-122-5p transcription in the liver and its subsequent incorporation into HFD-LExo and distribution to the circulation. To test this hypothesis, we employed a lentiviral vector specifically expressing CD63-mCherry targeted to the liver. The fluorescence derived from the vector was detected in both the liver and testes (Supplementary Fig. 5F), indicating that liver-derived exosomes with CD63-mCherry reached the testes through the circulation.
To investigate the impact of HFD-LExo on spermatogenesis and ferroptosis, HFD-LExo was administered to ND mice via tail vein injection (Fig. 5A). Mice receiving HFD-LExo significantly decreased sperm count, diameter of seminiferous tubules, number of seminiferous tubules, and serum testosterone levels (Fig. 5B–E). Moreover, HFD-LExo treatment downregulated the expression of STK31, AZIN2, CYP17A1, STAR, Hsd3b6, and Hsd17b7 in the testes (Fig. 5F, H). There was also a significant decrease in the number of Leydig cells, SPG, and RS in the testes of HFD-LExo mice compared with the Control group (Fig. 5G). Furthermore, Fe2+ (Fig. 5I) and MDA (Fig. 5J) levels were significantly increased in the testes of HFD-LExo mice compared with the Control group. Mitochondrial morphology was also adversely affected by this treatment, with reduced mitochondrial volume, loss of cristae, and abnormal morphology (Fig. 5K).

A Schematic diagram of HFD-LExo-treated mice (n = 6). B Sperm count (n = 6). C Diameter of seminiferous tubules (n = 30). D Number of seminiferous tubules (n = 5). E Relative T concentration in serum (n = 3). F Relative Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, and Star mRNA levels in the testes of mice (n = 3). G HE and IHC staining of testes (Scale bar = 200 or 50 µm). H Relative protein levels of STK31, AZIN2, CYP17A1, and STAR in the testes of mice (n = 3). I Relative ferrous ion levels in the testes (n = 3). J Relative MDA levels in the testes (n = 3). K TEM of mitochondria in the testes (Scale bar = 1 µm). L Schematic representation of liver-specific overexpression of miR-122-5p in mice (n = 6). M Sperm count (n = 6). N Diameter of seminiferous tubules (n = 30). O Number of seminiferous tubules (n = 5). P Relative T concentration in serum (n = 3). Q Relative Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, and Star mRNA levels in the testes of mice (n = 3). R Relative STK31, AZIN2, CYP17A1, and STAR protein levels in the testes of mice (n = 3). S HE and IHC staining of testes (Scale bar = 200 or 50 µm). T TEM of mitochondria in the testes (Scale bar = 1 µm). U Relative ferrous ion levels in the testes (n = 3). V Relative MDA levels in the testes (n = 3). Data information: Results are expressed as mean values ± SD. Data were analyzed using an unpaired Student’s t-test or one-way ANOVA. *p < 0.05, **p < 0.01.
The effects of HFD-LExo on Leydig cells were further validated in vitro. Confocal microscopy revealed that LExo were taken up by Leydig cells (Supplementary Fig. 5I). HFD-LExo treatment decreased cell viability (Supplementary Fig. 5J, P), MMP (Supplementary Fig. 5L, R), and testosterone levels in Leydig cell culture supernatant (Supplementary Fig. 5N, T). Additionally, HFD-LExo treatment also increased the concentrations of Fe2+ (Supplementary Fig. 5K, Q), MDA (Supplementary Fig. 5M, S), and ROS (Supplementary Fig. 5O, U) in Leydig cells, further suggesting that HFD-LExo promotes Leydig cell ferroptosis.
To further demonstrate the effect of hepatic miR-122-5p expression on spermatogenesis in vivo, a mouse model was constructed with specific hepatic miR-122 overexpression using AAV-TBG-122 injection (Fig. 5L, Supplementary Fig. 5W). The expression of miR-122-5p in the liver and testes was significantly upregulated by AAV-TBG-122 treatment as verified by RT-qPCR (Supplementary Fig. 5Y, Z). Meanwhile, sperm count, diameter of seminiferous tubules, number of seminiferous tubules, and serum testosterone levels were significantly lower in the AAV-TBG-122 mice compared to WT and AAV-TBG-WT mice (Fig. 5M–P). The expressions of STK31, AZIN2, CYP17A1, STAR, Hsd3b6, and Hsd17b7 were markedly downregulated in AAV-TBG-122 mice (Fig. 5Q, R). Leydig cell, SPG, and RS abundance were significantly lower in the AAV-TBG-122 mice compared to WT and AAV-TBG-WT mice (Fig. 5S). Moreover, the mitochondria in cells of the testis of AAV-TBG-122 mice showed swelling and vacuolation, while the mitochondrial cristae disappeared (Fig. 5T). Additionally, the concentrations of Fe2+ and MDA were significantly increased in the testes of AAV-TBG-122 mice compared to WT and AAV-TBG-WT groups (Fig. 5U, V).
miR-122-5p induces Leydig cell ferroptosis by targeting Scd2
Targeting relationship between miR-122-5p and Scd2 in Leydig cells was predicted (Supplementary Fig. 6A) and demonstrated (Fig. 6A). Additionally, miR-122-5p overexpression significantly downregulated Scd2 expression in Leydig cells in vitro (Fig. 6B, C). Further validation in a mouse model supported these findings. In the testes of HFD-EXO mice, SCD2 levels were significantly reduced compared to the Control and ND-EXO groups (Supplementary Fig. 6B, C, J). Treatment with GW4869 alleviated HFD-induced SCD2 downregulation (Supplementary Fig. 6D, E, J). Additionally, miR-122-5p antagomir treatment of HFD-EXO rescued the HFD-EXO-induced SCD2 downregulation (Supplementary Fig. 6F, G, J). Both HFD-LExo and AAV-TBG-122 treatments significantly reduced SCD2 expression in the testes (Supplementary Fig. 6H–L).

A Relative luciferase activity in different treatment groups (n = 3). B Relative miR-122-5p expression in Leydig cells with different treatments (n = 3). C Relative Scd2 mRNA expression in Leydig cells (n = 3). D Leydig cell viability of si-NC- or si-SCD2-treated groups (n = 6). E Relative ferrous ion levels of Leydig cells. F-a MMP of Leydig cells (n = 3), F-b Fluorescence intensity of the sample (Scale bar = 100 µm). G Relative MDA levels of Leydig cells (n = 3). H Relative testosterone concentration in Leydig cell supernatants (n = 3). I-a Relative ROS levels of Leydig cells treated with si-NC or si-SCD2 (n = 3), I-b Fluorescence intensity of the sample (Scale bar = 100 µm). J Cell viability of Leydig cells transfected with mimics-NC, mimics-miR-122-5p, OE-NC, or OE-SCD2 (n = 6). K Relative ferrous ion levels of Leydig cells transfected with mimics-NC, mimics-miR-122-5p, OE-NC, or OE-SCD2 (n = 3). L Relative MDA levels of Leydig cells transfected with mimics-NC, mimics-miR-122-5p, OE-NC, or OE-SCD2 (n = 3). M-a MMP of Leydig cells transfected with mimics-NC, mimics-miR-122-5p, OE-NC, or OE-SCD2 (n = 3), M-b Fluorescence intensity of the sample (Scale bar = 100 µm). N Relative testosterone concentration in Leydig cell supernatants from groups treated with mimics-NC, mimics-miR-122-5p, OE-NC, or OE-SCD2 (n = 3). O-a Relative ROS levels of Leydig cells transfected with mimics-NC, mimics-miR-122-5p, OE-NC, or OE-SCD (n = 3), O-b Fluorescence intensity of the sample (Scale bar = 100 µm). Data information: Results are expressed as mean values ± SD. Data were analyzed using an unpaired Student’s t-test or one-way ANOVA. *p < 0.05, **p < 0.01. Significance between groups was indicated by labeling with different letters; data labeled with the same letter indicate no significant difference.
To further verify the role of Scd2 in Leydig cell ferroptosis, its expression was knocked down using siRNA. This knockdown (si-SCD2-treated group) resulted in decreased cell viability (Fig. 6D, Supplementary Fig. 6M), MMP (Fig. 6F, Supplementary Fig. 6O), and testosterone levels in the culture supernatant (Fig. 6H, Supplementary Fig. 6Q) compared with the si-NC group. Additionally, si-SCD2 treatment significantly increased Fe2+ (Fig. 6E, Supplementary Fig. 6N), MDA (Fig. 6G, Supplementary Fig. 6P), and ROS (Fig. 6I, Supplementary Fig. 6R) levels in Leydig cells, confirming that SCD2 is involved in regulating ferroptosis in Leydig cells.
To further determine whether miR-122-5p regulates ferroptosis in Leydig cells by negatively regulating Scd2 expression, SCD2-overexpression plasmids (OE-SCD2) (Supplementary Fig. 6S) were transfected into the Leydig cells after treatment with mimics-miR-122-5p. OE-SCD2 significantly counteracted the detrimental effects of mimics-122-5p-induced overexpression, including the decrease in cell viability (Fig. 6J, Supplementary Fig. 6T), MMP (Fig. 6M, Supplementary Fig. 6V), and lower testosterone levels (Fig. 6N, Supplementary Fig. 6X). OE-SCD2 also alleviated miR-122-5p-induced increases in Fe2+ (Fig. 6K, Supplementary Fig. 6U), MDA (Fig. 6L, Supplementary Fig. 6W), and ROS (Fig. 6O, Supplementary Fig. 6Y).
Downregulation of Scd2 promotes Leydig cell ferroptosis in HFD mice
To explore the impact of reduced Scd2 expression on spermatogenesis, an AAV mouse model was constructed with specific Scd2 knockdown in Leydig cells (Fig. 7A). Scd2 was successfully knocked down in Leydig cells (Fig. 7F–H, Supplementary Fig. 7B), leading to lower sperm counts, diameter of seminiferous tubules, number of seminiferous tubules and testosterone levels compared to the WT and AAV-INSL3-WT groups (Fig. 7B–E). Moreover, STK31, AZIN2, CYP17A1, and STAR levels and Hsd3b6 and Hsd17b7 expression were also significantly downregulated in the testes of AAV-INSL3-shScd2 mice (Fig. 7F, G). HE and IHC staining of testes further revealed significantly reduced number of Leydig cells, SPG, and RS in the AAV-INSL3-shScd2 group compared with the WT and AAV-INSL3-WT groups (Fig. 7H). Mitochondrial analysis showed numerous vacuoles and the absence of cristae in the mitochondria of AAV-INSL3-shScd2 mice (Fig. 7I). Additionally, compared with the WT and AAV-INSL3-WT groups, the Fe2+ (Fig. 7J) and MDA (Fig. 7K) levels were significantly higher in the testes of the AAV-INSL3-shScd2 group.

A Schematic illustration of Scd2-specific knockdown in Leydig cells of mouse testes (n = 6). B Sperm count of mouse testes (n = 6). C Diameter of seminiferous tubules (n = 30). D Number of seminiferous tubules (n = 5). E Relative testosterone concentration in serum (n = 3). F Relative mRNA levels of Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, Star and Scd2 in the testes (n = 3). G Relative protein levels of STK31, AZIN2, CYP17A1, STAR, and SCD2 in the testes of mice (n = 3). H HE and IHC staining of testes (Scale bar = 200 or 50 µm). I TEM of mitochondria in the testes (Scale bar = 1 µm). J Relative ferrous ion levels in the testes (n = 3). K Relative MDA levels in the testes (n = 3). L Schematic illustration of the HFD mouse model with specific Scd2 overexpression in Leydig cells (n = 6). M Sperm count (n = 6). N Diameter of seminiferous tubules (n = 30). O Number of seminiferous tubules (n = 5). P Relative testosterone concentration in serum (n = 3). Q Relative mRNA levels of Stk31, Azin2, Cyp17a1, Hsd3b6, Hsd17b7, Star and Scd2 in the testes (n = 3). R Relative protein levels of STK31, AZIN2, CYP17A1, STAR, and SCD2 in the testes (n = 3). S HE and IHC staining of testes (Scale bar = 200 or 50 µm). T TEM of mitochondria in the testes (Scale bar = 1 µm). U Relative ferrous ion levels in the testes (n = 3). V Relative MDA levels in the testes (n = 3). Data information: Results are expressed as mean values ± SD. Data were analyzed using one-way ANOVA. *p < 0.05, **p < 0.01. Significance between groups was indicated by labeling with different letters; data labeled with the same letter indicate no significant difference.
Finally, to confirm that downregulated Scd2 in Leydig cells is critical to HFD-induced spermatogenesis disorders, an AAV vector that induces Leydig cells to specifically overexpress Scd2 (AAV Scd2) was constructed (Fig. 7L, Supplementary Fig. 7E). Scd2 was successfully overexpressed in Leydig cells (Fig. 7Q–S) and this treatment increased sperm counts, diameter of seminiferous tubules, number of seminiferous tubules and testosterone levels in the HFD + AAV Scd2 group compared with the HFD and HFD + AAV CON groups (Fig. 7M–P). Furthermore, HFD + AAV Scd2 treatment reversed the HFD-induced downregulation of STK31, AZIN2, CYP17A1, STAR, Hsd3b6, and Hsd17b7 in the testes (Fig. 7Q, R). HFD + AAV Scd2 treatment also alleviated the reduction in Leydig cells, SPG, and RS in HFD mice (Fig. 7S). HFD + AAV Scd2 treatment partially recovered the HFD-induced abnormal mitochondrial morphology in the testes (Fig. 7T). Additionally, levels of Fe2+ (Fig. 7U) and MDA (Fig. 7V) in the testes of the HFD + AAV Scd2 group were significantly lower than those in the HFD and HFD + AAV CON groups.
Discussion
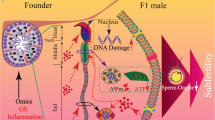
Leydig cells are essential for sexual development and testicular function as they synthesize testosterone and insulin-like 3 to regulate Sertoli cell function and support spermatogenesis16. Decreased testosterone is an important cause of obesity-induced infertility; however, the underlying mechanism has remained unclear. In this study, we uncovered that increased liver-derived miR-122-5p in circulating exosomes impairs testosterone synthesis by inducing Leydig cell ferroptosis via Scd2 downregulation in HFD mice, ultimately impairing spermatogenesis (Fig. 8).

Increased liver-derived miR-122-5p in circulating exosomes of HFD mice promotes ferroptosis by downregulating Scd2 expression in Leydig cells, leading to reduced testosterone synthesis and impaired spermatogenesis. SFAs, saturated fatty acids. MUFAs, mono-unsaturated fatty acids. PUFAs, polyunsaturated fatty acids.
Exosomes participate in intercellular signaling during gametogenesis, sperm capacitation, and embryo implantation17. However, the mechanisms through which exosomes influence obesity-related infertility remain unreported. We found that HFD-EXO reduced Leydig cell abundance and impaired spermatogenesis, similar to the effects of obesity. Treatment with GW4869, an inhibitor of exosome release, ameliorated HFD-induced reproductive impairments, suggesting a direct link between HFD-induced infertility and exosome alterations.
miRNAs in exosomes contribute to gene expression regulation18. Our miRNA sequencing revealed abnormal upregulation of miR-122-5p in HFD-EXO, consistent with the previously reported role of miR-122-5p in metabolic disorders19. Integrating miRNA sequencing of exosomes with scRNA-seq and ATAC-seq data from testicular Leydig cells, we demonstrated that HFD-EXO may impair testicular function through the promotion of ferroptosis in Leydig cells. Zhao et al. demonstrated that phthalates induce Sertoli cell ferroptosis by targeting the transferrin receptor, causing blood–testes barrier dysfunction20. However, the role of ferroptosis in HFD-induced reproductive disorder had not been previously reported. Our study shows that both HFD and HFD-EXO induce ferroptosis in Leydig cells, with Fer-1 mitigating Leydig cell death and restoring normal spermatogenesis.
Mechanistically, we identified a strong relationship between miR-122-5p and Scd2, a key enzyme in the conversion of saturated fatty acids (SFAs) to mono-unsaturated fatty acids (MUFAs)21. Studies have shown the close relationship between lipid metabolism and ferroptosis22,23,24. MUFAs can inhibit ferroptosis by suppressing ROS accumulation and reducing the phospholipid levels of oxidizable polyunsaturated fatty acids (PUFAs)25. Therefore, we speculated that alterations in the expression of Scd2 might affect cell sensitivity to ferroptosis. In vitro and in vivo, an HFD diet reduced testosterone synthesis and impaired spermatogenesis via miR-122-5p-mediated downregulation of Scd2 in Leydig cells, promoting ferroptosis. These findings demonstrate the important role of exosomes in HFD-induced abnormal spermatogenesis, providing theoretical support for miR-122-5p and Scd2 as candidate biomarkers and potential therapeutic targets for HFD-associated infertility. Notably, in addition to miRNAs, exosomes contain proteins, lipids, and other RNA species. A study demonstrated that HFD-induced upregulation of exosomal phosphatidylcholine suppresses insulin signaling26. This indicates broader HFD-driven changes to exosomal components beyond miRNAs. Future research should investigate how these alterations impact spermatogenesis and their role in miR-122-5p-induced ferroptosis in Leydig cells.
Another key finding in our study is the key role of the liver in driving changes in circulating exosomes in HFD mice. miR-122-5p was markedly increased in the liver of HFD mice compared with the intestine, epididymal fat, and kidneys. Additionally, miR-122-5p was significantly elevated in liver-derived exosomes from HFD mice compared to those from ND mice. A previous study reported that miR-122-5p is highly specifically expressed in the liver compared with other tissues27. The liver is an important organ for nutrient metabolism28; however, studies on the targeted relationship of liver-derived exosomes with other organs and the mechanisms of their interactions are limited. Furthermore, the specific functions of liver-derived exosomes in disease progression remain to be elucidated. Our results demonstrate that liver-derived exosomes can reach the testes, and treatment with liver-derived exosomes from HFD mice with liver-specific miR-122 overexpression vectors induces ferroptosis in Leydig cells and promotes abnormalities in testosterone synthesis and spermatogenesis. However, the mechanism underlying the HFD-induced abnormal elevation of miR-122-5p remains unclear. Previous studies have demonstrated that dysregulation of miR-122-5p expression is associated with the development of hepatic pathologies, including fatty liver disease, liver fibrosis, cancer, and infection29,30,31. Circulating miR-122-5p levels are positively correlate with body weight, sagittal height, and particularly visceral adipose tissue32. Additionally, miR-122-5p levels exhibit a positive correlation with very-low-density lipoprotein (VLDL), intermediate-density lipoprotein (IDL), and low-density lipoprotein (LDL) levels33. Our previous study revealed significantly elevated serum LDL and oxidized LDL levels in HFD-fed mice15, suggesting that hepatic miR-122-5p transcriptional upregulation may be a consequence of HFD-induced hepatic lipid accumulation and increased serum LDL levels.
Although this study has demonstrated that liver-derived exosomal miR-122-5p plays a pivotal role in the pathogenesis of obesity-associated spermatogenesis impairment by downregulating Scd2 expression in Leydig cells. However, there are still some limitations. First, additional human sample data are required to validate the potential of exosomal miR-122-5p as a diagnostic biomarker for obesity-related male infertility. Several studies have identified associations between miR-122-5p and male infertility, particularly oligospermia34,35,36. However, whether this association between miR-122-5p and spermatogenesis impairment is indirectly mediated by obesity-induced metabolic disorders remains unexplored and requires further investigation. Second, further investigation is needed to determine whether dietary interventions can reduce miR-122-5p levels in circulating exosomes of obese individuals. Furthermore, we further analyzed the proportion of Leydig cell subtypes and transcriptomic data from scRNA-seq. This revealed that HFD reduced the L0 subtype of Leydig cells, indicating a decrease in the number of primitive Leydig cells. Notably, Scd2 was identified as a key marker for differential expression of L0 cells. This suggests that different Leydig cell subtypes may have varying sensitivities to ferroptosis triggered by HFD-induced Scd2 downregulation. Therefore, the regulatory mechanisms by which HFD-EXO primarily induce ferroptosis in primitive Leydig cells, compared to other subtypes, remain to be elucidated.
In summary, this study uncovers the insights on the liver-testes axis in HFD-induced spermatogenesis disorders. HFD increases liver-derived miR-122-5p in circulating exosomes, which induces Leydig cell ferroptosis by downregulating Scd2, leading to decreased testosterone synthesis and abnormal spermatogenesis. These findings reveal a previously unrecognized role of exosomes in HFD-induced infertility and suggest that targeting ferroptosis and miR-122-5p could offer potential therapeutic avenues for treating obesity-related infertility.
Methods
Animal models
All animal experiments were approved by the Institutional Animal Care and Use Committee of Jilin University and adhered to welfare of laboratory animals (Permit Number: SY202304300). Eight-week-old male C57BL/6 mice (20 ± 2 g) were purchased from Changsheng Biotechnology Co. Ltd., China, and housed in a specific pathogen-free animal lab of Jilin University with standard environmental conditions (22 ± 2 °C, with a relative humidity 40–60% and a 12/12 light/dark cycle). We have complied with all relevant ethical guidelines for animals use. Mice were euthanised via cervical dislocation to ensure instantaneous loss of consciousness and minimize distress during the procedure. The mouse’s head was firmly held with thumb and index finger, while the tail base was gently pulled to dislocate the cervical vertebrae. Death was confirmed by the absence of vital signs.
To evaluate the effect of HFD on spermatogenesis, mice were randomly divided into ND and HFD groups (n = 8 per group). Mice in the normal diet (ND) group were fed a normal diet (Supplementary Table 1), while mice in the HFD group were fed an HFD (Supplementary Table 2) for 8 weeks.
To investigate the effect of serum exosomes on spermatogenesis, mice were divided into three groups: control (Control), ND-mice serum exosome (ND-EXO), and HFD-mice serum exosome (HFD-EXO) (n = 8). Control mice were injected with normal saline (NS) via tail vein. Mice in the ND-EXO and HFD-EXO groups were injected via tail vein with exosomes isolated from the serum of ND mice or HFD mice, respectively. Exosomes from ND or HFD mice were injected weekly (30 μg) for 4 weeks.
To assess whether exosomes play a role in HFD-induced spermatogenesis disorders, mice were divided into three groups: the ND, HFD, and HFD supplemented with GW4869 (Sigma-Aldrich, Saint Louis, USA) (HFD + GW) groups (n = 6). The ND group was fed a normal diet, and the HFD and HFD + GW groups were fed an HFD for 8 weeks. Mice in the HFD + GW, ND, and HFD groups were injected intraperitoneally with GW4869 (2.5 mg kg−1) or control solvent every 3 days.
To determine whether ferroptosis contributes to HFD-induced spermatogenesis disorders, mice were divided into three groups: the ND, HFD, and HFD supplemented with Ferrostatin-1 (MCE, New Jersey, USA) (HFD+Fer-1) groups (n = 6). The ND group was fed a normal diet, while the HFD and HFD+Fer-1 groups were fed an HFD for 8 weeks. Mice in the HFD+Fer-1, ND, and HFD groups were injected intraperitoneally with 0.8 mg kg−1 Fer-1 (MCE, New Jersey, USA) or control solvent every 3 days.
To investigate whether miR-122-5p in exosomes contributes to HFD-EXO-induced spermatogenesis disorders and Leydig cell ferroptosis, HFD-EXO were processed and treated with miR-122-5p antagomir or antagomir NC (RiboBio, Canton, China) according to previously published methods37. Mice were divided into three groups: control (Control), NC antagomir-treated HFD-EXO (NC antago-HE), and miR-122-5p antagomir-treated HFD-EXO (122-5p antago-HE) groups (n = 5). Mice in the Control, NC antago-HE, and 122-5p antago-HE groups were injected with NS, NC antago-HE, or 122-5p antago-HE (30 μg), respectively, via tail vein once per week for 4 weeks.
To determine whether liver-derived exosomes (LExo) are the main source of serum exosomal miR-122-5p and the effect of LExo on spermatogenesis, mice were divided into Control and HFD-LExo groups (n = 6). Mice were injected with NS or HFD-LExo (30 μg), respectively, via the tail vein once per week for 4 weeks.
To assess the role of liver-derived miR-122-5p in HFD-induced spermatogenesis disorders, mice were divided into three groups: WT, AAV-TBG-WT, and AAV-TBG-122 groups (n = 6). Mice were injected intraperitoneally with NS, rAAV-TBG-EGFP-WPRE-pA, or rAAV-TBG-pre-miR-122-EGFP-pA, respectively. AAV was provided by Wuhan BrainVTA Science and Technology Co., Ltd., Wuhan, China, and injected at a dose of 1 × 1011 vg, injected twice, once every 4 weeks.
To assess the effect of Scd2 on Leydig cells and spermatogenesis, mice were divided into three groups: WT, AAV-INSL3-WT, and AAV-INSL3-shScd2 groups (n = 6). Mice were intraperitoneally injected with NS, rAAV-INSL3-EGFP-5′miR-30a-shRNA (scramble)-3′-miR-30a-WPREs, or rAAV-INSL3-EGFP-5′miR-30a-shRNA2(Scd2)-3′-miR-30a-WPREs, respectively. AAV was injected at a dose of 1 × 1011 vg, injected twice, once every 4 weeks.
To evaluate the in vivo role of liver-derived miR-122-5p and Leydig cell Scd2 expression on HFD-induced spermatogenic disorders and ferroptosis, the mice were divided into four groups: ND, HFD, HFD supplemented with AAV-INSL3-CON (HFD + AAV CON), and HFD supplemented with AAV-INSL3-Scd2 (HFD + AAV Scd2) groups (n = 6). Mice in the ND group were fed normal diets, while the other three groups were fed an HFD. Mice in the ND and HFD groups were injected intraperitoneally with NS. Mice in the HFD + AAV CON and HFD + AAV Scd2 groups were intraperitoneally injected with rAAV-INSL3-EGFP-WPREs or rAAV-INSL3-Scd2-EGFP-WPREs at a dose of 1 × 1011 vg, injected twice, once every 4 weeks.
Insulin and glucose tolerance test
For insulin tolerance, mice were fasted for 4 h with free access to water. Blood was collected from the tail, and blood glucose was assessed at 0 min using the Accu-Chek blood glucose monitor (Roche Diagnostics Corp., Basel, Germany). The mice were then injected intraperitoneally with insulin (1 U kg−1), and blood glucose was measured after 15, 30, 60, 90, and 120 min.
For glucose tolerance, mice were fasted for 12 h with free access to water. Blood glucose was detected at 0 min, and then mice were injected intraperitoneally with Deoxy-glucose (2 g kg−1). Blood glucose was then measured after 15, 30, 60, 90, 120 min.
Sperm count
Intact epididymides were collected after euthanasia. The unilateral epididymis was placed in a 35 mm plate containing 1 mL of modified Tyrode’s solution preheated to 37 °C. It was then dissected using ophthalmic scissors and incubated at 37 °C with 5% CO2 for 30 min to release the sperm. Sperm counts were quantified under a light microscope using a hemacytometer.
Cell culture
Testicular Leydig cells were isolated as previously described38. Briefly, the abdominal cavity was opened, testes were collected and placed in a 6-cm cell culture dish with 5 mL of phosphate-buffered saline (PBS; Thermo Fisher, Waltham, MA, USA). The testicle tunica albuginea was dissected, and testes were gently removed. Subsequently, 0.05% type I collagenase was added to 5 mL of PBS and incubated with the testes at 37 °C in a shaking incubator for 5 min. After incubation, 5 mL of Dulbecco’s modified eagle medium (DMEM)F-12 (Sigma-Aldrich, Saint Louis, USA) supplemented with 10% fetal bovine serum (FBS; ABW, Shanghai, China) and 10 mL of PBS were added to terminate digestion. The liquid was collected and centrifuged at 300 × g for 5 min, the supernatant was discarded, and the cells were resuspended in 12 mL of cell culture medium. The cells were then cultured on 10-cm cell culture dishes and incubated at 37 °C with 5% CO2 and full humidity. After 24 h, the cells were washed with PBS, placed in fresh media, and cultured for another 48 h. Finally, the Leydig cells were digested, inoculated onto cell culture plates, incubated with a rabbit anti-CYP17A1 polyclonal antibody overnight at 4 °C, and coupled with a FITC-conjugated secondary antibody for 1 h at room temperature. Cells were characterized by immunofluorescence (Supplementary Fig. 3C).
The TM3 Leydig cell line (American Type Culture Collection, Cat #CRL-1714; Rockville, MD) was cultured in DMEMF-12 supplemented with 10% FBS.
Cell treatments and transfection
To assess the effect of Fer-1 on Leydig cells, they were divided into the Control, ND-EXO, HFD-EXO, and HFD-EXO+Fer-1 groups. After reaching 70% confluence, the cells were treated with a DMSO solvent, 30 µg of EXO, or 2 μM Fer-1.
To investigate the effects of miR-122-5p on ferroptosis in Leydig cells, transfection of mimics-NC or mimics-miR-122-5p (JTS Scientific, Wuhan, China) was performed with Lipofectamine 3000 according to the manufacturer’s instructions.
To investigate the effects of Scd2 on ferroptosis in Leydig cells, transfection with si-NC or si-SCD2 (RiboBio, Canton, China) was performed with Lipofectamine 3000 according to the manufacturer’s instructions by adding.
To assess the effect of miR-122-5p on Scd2, transfection of mimics-NC, mimics-miR-122-5p, OE-NC, or SCD2-overexpression plasmids (OE-SCD2) (Mingsheng Biotech Co., Ltd., Shenyang, China) was performed with Lipofectamine 3000.
The role of miR-122-5p in Leydig cell ferroptosis was assessed by supplementing cell cultures with control solvent, NC antago-HE (30 µg), or 122-5p antago-HE (30 µg).
To further verify the effect of HFD-LExo on Leydig cells, 30 µg of HFD-LExo was added to the cell culture medium to treat the cells.
Testosterone measurement
Testosterone levels in serum or the cell culture supernatant were measured using a mouse T ELISA kit (Shanghai Langdon Biotechnology Co., Ltd, Shanghai, China) following to the manufacturer’s instructions. In brief, add 50 μL of the standard or sample to each well, followed by 50 μL of the biotin-antigen working solution. Seal the plate with adhesive film and incubate at 37 °C for 30 min. After incubation, wash the five times. Next, add 50 μL of affinity-HRP conjugate to all wells except the blank, incubate at 37 °C for another 30 min, and wash the plate five times again. Subsequently, add 50 μL of chromogen A to each well, immediately followed by 50 μL of chromogen B. Incubate the plate at 37 °C in the dark for 10 min. Finally, terminate the reaction by adding 50 μL of stop solution to each well and measure the absorbance at 450 nm.
Exosome purification and characterization
Exosomes were isolated as previously described39,40. Briefly, blood samples were collected from ND and HFD mice, left at room temperature for 30 min, and centrifuged for 15 min (4500 × g) to collect the supernatant. A 30 mL serum was re-centrifuged for 60 min (10,000 × g, 4 °C) to remove cellular debris and large vesicles. The supernatant was ultracentrifuged at 150,000 × g for 2 h at 4 °C. The precipitate was resuspended in PBS and ultracentrifuged again (150,000 × g, 4 °C, 2 h) to obtain the exosomes. Exosomes were resuspended in PBS, collected after filtration through a 0.22-µm membrane, and aliquoted for storage at −80 °C.
For liver-derived exosomes, the livers of mice from the ND and HFD groups were collected, and the large blood vessels, gallbladder, and bile ducts were removed. The liver cell suspension was obtained by shearing and grinding, and the cells were cultured in Roswell Park Memorial Institute (RPMI)-1640 medium (Sigma-Aldrich, Saint Louis, USA) at 37 °C for 24 h. A 50 mLsupernatant was centrifuged at 10,000 × g for 60 min at 4 °C to remove cellular debris and large vesicles, and then ultracentrifuged (150,000 × g, 4 °C, 2 h). The precipitate was resuspended in PBS and centrifuged again at 150,000 × g for 2 h at 4 °C. Exosomes were resuspended in PBS, collected after filtration through a 0.22-µm membrane, and aliquoted for storage at −80 °C.
The size distribution of exosomes was measured with a Flow NanoAnalyzer, and the morphology of exosomes was observed by TEM as previously described37. Exosomes were detected using WB. All antibodies are listed in the structured methods. Exosomes were observed under a laser confocal microscope (Leica, Wetzlar, Germany) using DiD (Thermo Fisher, Massachusetts, USA) to label exosomes, FITC-phalloidin (Abcam, Cambridge, UK) to label microfilaments, and DAPI (Beyotime, Shanghai, China) to label nuclei according to the manufacturer’s instructions. Pitstop 2 (Sigma-Aldrich, Saint Louis, USA) was applied to inhibit clathrin-mediated endocytosis.
A lentiviral vector specifically expressing CD63-mCherry in the liver (Wuhan BrainVTA Science and Technology Co., Ltd, Wuhan, China) was constructed. Mice were injected intraperitoneally with 2 × 108 Tu lentivirus vector to observe the biodistribution of CD63-mCherry-labeled exosomes in vivo.
RNA extraction and RT-qPCR
Total RNA was extracted from tissues using TRIzol (Takara, Kyoto, Japan). miRNAs of serum and liver exosomes were isolated using the miRNeasy Serum/Plasma Kit (QIAGEN, Dusseldorf, Germany). cDNA was reverse transcribed from the RNA using the Reverse Transcription Reagents (mRNA: the PrimeScript RT reagent Kit, Takara Bio, Kyoto, Japan; miRNA: miRNA First Strand cDNA Synthesis, Sangon Biotech, Shanghai, China). mRNA expression was assessed via real-time fluorescence quantitative PCR (Mx3005P; Agilent, California, USA) using the SYBR Premix Ex Taq II kit (Takara Bio, Kyoto, Japan) according to the manufacturer’s protocol. The relative expression of target mRNA was analyzed by the 2−ΔΔCT method using Gapdh or peptidylprolyl isomerase A (Ppia) as the reference genes. All primers used for the study are detailed in Supplementary Table 3.
Protein extraction and WB
Testicular tissue was homogenized using the Radio Immunoprecipitation Assay buffer (RIPA) (Beyotime, Shanghai, China) containing a protease/phosphatase inhibitor (Solarbio Life Sciences, Beijing, China). The homogenate was transferred to centrifuge tubes and lysed at 4 °C for 30 min, then centrifuged at 13,000 × g for 15 min at 4 °C. The supernatant was collected, and total protein content was detected using a BCA Protein Assay Kit (Beyotime, Shanghai, China). Subsequently, equal quantities of protein per well were loaded onto a 12% SDS-PAGE gel for electrophoresis and then transferred to a PVDF membrane. The membrane was blocked with 5% bovine serum albumin (Sigma, USA) in PBS containing 0.1% Tween-20 at room temperature for 1 h, followed by overnight incubation with the primary antibody at 4 °C. After three washes with PBST, the membrane was incubated at room temperature with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (H + L) (Bioworld, China). Following PBST washes, protein signals were detected using a Tanon-5200 fully automated gel imaging analysis system (Shanghai Tianneng, China) with an enhanced chemiluminescence detection kit (Beyotime Biotechnology, China). Gray value scanning was performed using ImageJ. All antibodies are listed in Supplementary Table 4.
Preparation of single-cell suspensions
Testicular tissue was placed in ice-cold PBS, and the tunica albuginea was carefully removed. The tissue was then minced into 0.5 mm² pieces and enzymatically digested in dissociation buffer (0.35% collagenase IV 5, 2 mg/mL papain, 120 Units/mL DNase I) at 37 °C in a water bath shaker (100 rpm) for 20 min. The cell suspension was filtered through a 40μm cell strainer and centrifuged at 300 × g for 5 min at 4 °C. Next, 1 mL of erythrocyte lysis solution (MACS 130-094-183, 10×) was added to the cell suspension and incubate at room temperature for 2–10 min. Cells were centrifuged at 300 × g for 5 min at 4 °C, and the resulting pellet was collected. 100 μl Dead Cell Removal Reagent (MACS 130-090-101) was added to the pellet, followed by incubation at room temperature for 15 min. The reagent and dead cells were removed by adding Binding Buffer to the suspension and loading it onto MS Columns (130-042-201). Cells were centrifuged again at 300 × g for 5 min at 4 °C, washed twice with PBS (0.04% BSA), and centrifuged at 300 × g for 5 min at 4 °C. Cell viability was assessed using the trypan blue staining method, and cell counts were determined using a haemocytometer.
Single-cell RNA analysis
The single-cell suspension was added to the 10×Chromium chip in accordance with the protocol for the 10X Genomics Chromium Single-Cell 3′ Library Preparation Kit (V3). cDNA amplification and library preparation were performed following the standard protocol. Results from Illumina sequencing offline were converted to FASTQ format using bcl2fastq software (version 5.0.1). The scRNA-seq sequencing data were compared to reference genome using CellRanger software, and cellular and individual cellular 3′ end transcripts were identified and counted in the sequenced samples. (https://support.10xgenomics.com/single-cell-gene expression/software/pipelines/latest/what-is cell-ranger, version 7.0.0). The output CellRanger expression profile matrix was loaded into Seurat (version 4.1.0) for filtering of low-quality cells from scRNA-seq data, and the filtered data were downscaled and clustered. Filtering low cell quality thresholds: number of genes expressed per cell >500, mitochondrial genes expressed in <25% of cells. Cells were projected into 2D space using t-SNE or UMAP.
Single-cell ATAC-seq analysis
The cell concentration was adjusted to an optimal level, and gel beads containing barcodes and primers were combined with a mixture of cells and enzymes to form GEMs (Gel Bead in Emulsion) using the Chromium™ Single Cell 3′ Solution platform. Within the GEMs, the gel beads were lysed, mRNA was released from the cells, and reverse transcription produced barcoded cDNA for sequencing. After disrupting the liquid oil layer, cDNA amplification was performed. Following purification, quality control testing was conducted. Library construction was initiated upon passing quality control. The libraries were sequenced on an Illumina NovaSeq 6000 system using paired-end sequencing runs. Sequencing analyses were performed using a 10X Genomics Cellranger-atac. The library preparation and sequencing were performed by LC Bio. Raw sequencing data were filtered and aligned, followed by analysis of barcode statistics. Transposase cleavage sites were identified, and chromatin accessibility peaks were called. Following cell type classification, cellular clustering was performed to assess chromatin accessibility variation across distinct cell lineages. Bioinformatic analysis was performed using the OmicStudio tools (https://www.omicstudio.cn/tool).
miRNA sequencing
Sequencing of exosomal miRNA was conducted according to Illumina protocols, including library preparation and sequencing. The small RNA sequencing library was prepared using the TruSeq Small RNA Sample Prep Kits (Illumina, California, USA). Briefly, 3′ and 5′ RNA adapters were ligated to total RNA, followed by reverse transcription and PCR amplification. The small RNA library was then purified by gel electrophoresis. Following quality validation, sequence was performed on Illumina HiSeq 2000/2500 with a single-end reading length of 1 × 50 bp. miRNA data were analyzed using ACGT101-miR (v4.2) software.
Luciferase activity assay
HEK293 cells were cultured to 90% confluence, digested with 0.25% trypsin, and centrifuged at 800 × g for 3 min. The cells were then seeded into 12-well plates and perform transfection at approximately 80% density. Four transfection conditions were used: pmirGLO-Scd2-wt + miRNA mimics-NC, pmirGLO-Scd2-wt + miRNA mimics, pmirGLO-Scd2-mt + miRNA mimics-NC, pmirGLO-Scd2-mt + miRNA mimics. At 24 h post-transfection, 1 μg/ml lipopolysaccharide was added to the designated treatment groups. After 15 min of incubation at room temperature for cell lysis, the lysates were collected. 100 μL of LAR II and 20 μL of each lysate were mixed per tube to measure the Firefly luciferase value. 100 μL of Stop Glo Reagent solution is then added to each tube to measure the Renilla luciferase value. The luciferase ratio is calculated based on these results.
Morphological and immunohistochemical analysis
Immediately after dissection, the testes were fixed in 4% formaldehyde solution, dehydrated, and embedded in paraffin for serial sectioning. Sections were dewaxed and stained with HE for general morphological examination. Other sections were incubated for 10 min at 95 °C using 0.01 M sodium citrate buffer after dewaxing. They were then blocked with BDT (3% BSA, 10% 29 normal donkey serum in TBS) for 30 min. Immunochemical staining was performed using an IHC kit (MXb, Fujian, China) and a DAB kit (MXb, Fujian, China) according to the manufacturer’s instructions. Finally, the testes sections were observed under an E100 light microscope (Nikon, Tokyo, Japan), and images were obtained using a 600D Photo Imaging System (Canon Inc., Tokyo, Japan). The number of seminiferous tubules in the testicular sections was quantified. Subsequently, the minor and major axes of 10 seminiferous tubules exhibiting a round or nearly round morphology were measured, and the mean diameter was calculated.
Cell proliferation assay
Cell viability was assessed using the Cell Counting Kit-8 assay (APExBIO, Houston, USA). Leydig cells and TM3 cells were seeded in 96-well microplate and treated at 50% cell density. After a 24 h treatment, 10 μL of CCK-8 solution was added, and incubated at 37 °C for 1 h. Optical density (OD) was measured at 450 nm using an enzyme labeler (BioTek, Vermont, USA). Cell viability was calculated as the ratio of the OD values from treated cells to those of control cells, representing the percentage of viable cells compared to the untreated controls.
Measurement of reactive oxygen species
Intracellular reactive oxygen species (ROS) levels were measured with a ROS detection kit (Beyotime, Shanghai, China). Leydig cells and TM3 cells were seeded into 6-well plates. When the cell density reached 70%, the cells were collected after 24 h of treatment, suspended in diluted DCFH-DA, incubated in a 37 °C cell culture incubator for 20 min, and washed thrice with serum-free DMEMF/12. Cells were resuspended with 200 μL PBS, and the fluorescence intensity of the samples was detected using a fluorescence microscope (Olympus, Tokyo, Japan) and a FACSCalibur cytometer (Becton-Dickinson, New Jersey, USA).
Intracellular mitochondrial membrane potential assays
The mitochondrial membrane potential (MMP) of Leydig cells was measured using a JC-1 (MCE, New Jersey, USA) dual-emission membrane potential-sensitive probe that exists as a green fluorescent monomer at low MMPs and forms aggregates with red/orange fluorescence at high MMPs. JC-1 was used as per the manufacturer’s instructions. The cells were digested with trypsin (Thermo Fisher, Massachusetts, USA), centrifuged at 400 × g for 5 min, and the supernatant was discarded. Subsequently, the cells were resuspended in 1 mL of JC-1 working solution and incubated at 37 °C in a 5% CO₂ incubator for 30 min. After incubation, the cells were washed twice with 2 mL of PBS by centrifugation at 400 × g for 5 min, followed by supernatant removal. The cells were resuspended in 1 mL of PBS and analyzed by flow cytometry on a FACSCalibur cytometer (Becton-Dickinson, New Jersey, USA). Healthy cells, characterized by red JC-1 aggregates in mitochondria, were detected via the FL2 channel. Apoptotic or unhealthy cells, containing green JC-1 monomers, were measured in the FL1 channel. The fluorescence intensity of the samples was observed under a fluorescence microscope (Olympus, Tokyo, Japan).
Malonaldehyde detection
The testes or cell samples were homogenized or lysed using RIPA Lysis Buffer (Beyotime, Shanghai, China); the supernatant was collected by centrifugation at 12,000 × g for 10 min. The malonaldehyde (MDA) concentration in testes and interstitial cells was measured using the MDA Assay Kit (Beyotime, Shanghai, China) according to the manufacturer’s instructions. A 0.1 mL of PBS (blank control), standards at different concentrations, or test samples to centrifuge tubes, followed by the addition of 0.2 mL MDA assay working solution. The mixtures were vortexed thoroughly and heated in a boiling water bath for 15 min. After cooling to room temperature, the mixtures were centrifuged at 1000 × g for 10 min. Subsequently, 200 μL of the supernatant was transferred to a 96-well microplate, and absorbance was measured at 532 nm using an enzyme labeller (BioTek, Vermont, USA). Protein concentration was measured with a BCA Protein Assay Kit (Beyotime, Shanghai, China), which was applied to calculate the MDA content of the sample per unit weight of protein.
Iron measurement
Testicular tissues or cells were collected, and the ferrous iron content was determined using a Ferrous Iron Colorimetric Assay Kit (Elabscience, Wuhan, China) or Cell Ferrous Iron Colorimetric Assay Kit (Elabscience, Wuhan, China), respectively, according to the manufacturers’ protocols. A 300 μL standard or sample was added to centrifuge tubes, followed by the addition of 150 μL of chromogenic solution to each tube. The mixture was vortexed thoroughly and incubated at 37 °C for 10 min, then centrifuged at 12,000 × g for 10 min. After centrifugation, 300 μL of the supernatant was transferred to the 96-well microplate. The optical density value at 593 nm was measured using an enzyme labeller (BioTek, Vermont, USA).
Transmission electron microscopy
Fresh testicular samples were collected and cut in 1 × 1 × 1 mm pieces. The samples were fixed in Gluta fixative (Biosharp, Shanghai, China) at 4 °C for 24 h. After dehydration with ethanol and acetone, the samples were embedded and sectioned into ultrathin sections. The sections were stained using lead citrate and uranyl acetate and observed with a transmission electron microscope (Philips, Amsterdam, Holland).
Statistical analysis
Results are expressed as mean values ± standard deviation (SD). Comparisons between two groups were made by a two-tailed Student’s t-test, and comparisons between multiple groups were made by one-way analysis of variance (ANOVA) followed by an LSD post hoc test. Levene’s test of equality of variances was used to test for homogeneity of variances. Three independent biological replicates were performed for each experiment. Significance was set at p < 0.05. Statistical analysis was conducted using SPSS 22.0 (IBM, New York, USA).
Data availability
scRNA-seq data have been published in our previous study15 (accession number: GSE254812). scATAC-seq data (accession number: PRJNA1403152) and miRNA sequencing data (accession number: PRJNA1377513) are provided in the NCBI repository. All numerical data are presented in Supplementary Data 1. Uncropped blot images are provided as Supplementary Figs. 8–11 in the Supplementary Material file. All other data are available from the corresponding author upon reasonable request.
References
Piche, M. E., Tchernof, A. & Despres, J. P. Obesity phenotypes, diabetes, and cardiovascular diseases. Circ. Res. 126, 1477–1500 (2020).
NCD-RisC, N. R. F. C. Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 387, 1377–1396 (2016).
Ameratunga, D., Gebeh, A. & Amoako, A. Obesity and male infertility. Best Pract. Res. Clin. Obstet. Gynaecol. 90, 102393 (2023).
Craig, J. R., Jenkins, T. G., Carrell, D. T. & Hotaling, J. M. Obesity, male infertility, and the sperm epigenome. Fertil. Steril. 107, 848–859 (2017).
Leisegang, K., Sengupta, P., Agarwal, A. & Henkel, R. Obesity and male infertility: mechanisms and management. Andrologia 53, e13617 (2021).
Bieniek, J. M. et al. Influence of increasing body mass index on semen and reproductive hormonal parameters in a multi-institutional cohort of subfertile men. Fertil. Steril. 106, 1070–1075 (2016).
He, X., Kuang, G., Wu, Y. & Ou, C. Emerging roles of exosomal miRNAs in diabetes mellitus. Clin. Transl. Med. 11, e468 (2021).
Kawata, R. et al. Exploration of small RNA biomarkers for testicular injury in the serum exosomes of rats. Toxicology 440, 152490 (2020).
Chen, T. et al. Gut-derived exosomes mediate the microbiota dysbiosis-induced spermatogenesis impairment by targeting meioc in mice. Adv. Sci. 11, e2310110 (2024).
Trefts, E., Gannon, M. & Wasserman, D. H. The liver. Curr. Biol. 27, R1147–R1151 (2017).
Lee, E. S. et al. Curcumin analog CUR5-8 ameliorates nonalcoholic fatty liver disease in mice with high-fat diet-induced obesity. Metabolism 103, 154015 (2020).
Grossmann, M., Wierman, M. E., Angus, P. & Handelsman, D. J. Reproductive endocrinology of nonalcoholic fatty liver disease. Endocr. Rev. 40, 417–446 (2019).
Liu, G. L. et al. Endoplasmic reticulum stress mediating downregulated StAR and 3-beta-HSD and low plasma testosterone caused by hypoxia is attenuated by CPU86017-RS and nifedipine. J. Biomed. Sci. 19, 4 (2012).
Yokoe, T. et al. Efficient identification of a novel cancer/testis antigen for immunotherapy using three-step microarray analysis. Cancer Res. 68, 1074–1082 (2008).
Yuan, C. et al. Increased oxidized low-density lipoprotein in mice exposed to a high-fat diet impaired spermatogenesis by inhibiting testosterone synthesis via the Klk1bs/Eid3 pathway. Clin. Transl. Med. 14, e1603 (2024).
Martin, L. J. Cell interactions and genetic regulation that contribute to testicular Leydig cell development and differentiation. Mol. Reprod. Dev. 83, 470–487 (2016).
Kowalczyk, A., Wrzecinska, M., Czerniawska-Piatkowska, E. & Kupczynski, R. Exosomes—spectacular role in reproduction. Biomed. Pharmacother. 148, 112752 (2022).
Schuldt, A. Great expectations of small RNAs. Nat. Rev. Mol. Cell Biol. 11, 676 (2010).
Derumeaux, G. A. & D’Humieres, T. MicroRNA, miR-122-5p, stiffens the diabetic heart. JACC Cardiovasc. Imaging 14, 1143–1145 (2021).
Zhao, Y. et al. Ferroptosis is critical for phthalates driving the blood-testis barrier dysfunction via targeting transferrin receptor. Redox Biol. 59, 102584 (2023).
O’Neill, L. M. et al. Stearoyl-CoA desaturase-2 in murine development, metabolism, and disease. Int. J. Mol. Sci. 21, 8619 (2020).
Lorito, N. et al. FADS1/2 control lipid metabolism and ferroptosis susceptibility in triple-negative breast cancer. EMBO Mol. Med. 16, 1533–1559 (2024).
Peleman, C. et al. Ferroptosis is a targetable detrimental factor in metabolic dysfunction-associated steatotic liver disease. Cell Death Differ. 31, 1113–1126 (2024).
Pope, L. E. & Dixon, S. J. Regulation of ferroptosis by lipid metabolism. Trends Cell Biol. 33, 1077–1087 (2023).
Magtanong, L. et al. Exogenous monounsaturated fatty acids promote a ferroptosis-resistant cell state. Cell Chem. Biol. 26, 420–432 (2019).
Kumar, A. et al. High-fat diet-induced upregulation of exosomal phosphatidylcholine contributes to insulin resistance. Nat. Commun. 12, 213 (2021).
Lagos-Quintana, M. et al. Identification of tissue-specific microRNAs from mouse. Curr. Biol. 12, 735–739 (2002).
Sato, K., Meng, F., Glaser, S. & Alpini, G. Exosomes in liver pathology. J. Hepatol. 65, 213–221 (2016).
Tobaruela-Resola, A. L. et al. Circulating miR-122-5p, miR-151a-3p, miR-126-5p and miR-21-5p as potential predictive biomarkers for Metabolic Dysfunction-Associated Steatotic Liver Disease assessment. J. Physiol. Biochem. https://doi.org/10.1007/s13105-024-01037-8 (2024).
Zhou, Z. et al. Circular RNA cVIM promotes hepatic stellate cell activation in liver fibrosis via miR-122-5p/miR-9-5p-mediated TGF-beta signaling cascade. Commun. Biol. 7, 113 (2024).
Fan, X., Qiao, W., Guo, X., Wang, J. & Zhao, L. METTL14-mediated miR-122-5p maturation stimulated tumor progression by targeting KAT2A in hepatocellular carcinoma. Sci. Rep. 15, 17884 (2025).
Hess, A. L. et al. Levels of circulating miR-122 are associated with weight loss and metabolic syndrome. Obesity 28, 493–501 (2020).
Raitoharju, E. et al. Blood hsa-miR-122-5p and hsa-miR-885-5p levels associate with fatty liver and related lipoprotein metabolism—the Young Finns Study. Sci. Rep. 6, 38262 (2016).
Trzybulska, D., Bobjer, J., Giwercman, A. & Tsatsanis, C. Serum microRNAs in male subfertility-biomarkers and a potential pathogenetic link to metabolic syndrome. J. Assist. Reprod. Genet. 34, 1277–1282 (2017).
Joshi, M. et al. Qualitative and quantitative assessment of sperm miRNAs identifies hsa-miR-9-3p, hsa-miR-30b-5p and hsa-miR-122-5p as potential biomarkers of male infertility and sperm quality. Reprod. Biol. Endocrinol. 20, 122 (2022).
Deng, H. et al. Serum exosomes from hepatitis B virus-infected patients inhibit glycolysis in Sertoli cells via miR-122-5p/ALDOA axis. Reprod. Biol. 24, 100845 (2024).
Cao, M. et al. Adipose mesenchymal stem cell-derived exosomal microRNAs ameliorate polycystic ovary syndrome by protecting against metabolic disturbances. Biomaterials 288, 121739 (2022).
Gao, S. et al. Brain-derived neurotrophic factor: A steroidogenic regulator of Leydig cells. J. Cell. Physiol. 234, 14058–14067 (2019).
Gao, L. et al. Cardio-renal exosomes in myocardial infarction serum regulate proangiogenic paracrine signaling in adipose mesenchymal stem cells. Theranostics 10, 1060–1073 (2020).
Wang, L. et al. Host liver-derived extracellular vesicles deliver miR-142a-3p induces neutrophil extracellular traps via targeting WASL to block the development of Schistosoma japonicum. Mol. Ther. 30, 2092–2107 (2022).
Acknowledgements
The research was supported by the National Natural Science Foundation of China (32272872, 32172726), Outstanding Young Talents and Technology Innovation Project of Jilin Province (20220508094RC), and the China Agriculture Research System of MOF and MARA (CARS-36).
Author information
Authors and Affiliations
Contributions
Xu Zhou, Chunjin Li, and Nan Wang conceived, designed, and supervised the study. Nan Wang, Xu Zhou, and Chunjin Li wrote and revised the manuscript. Nan Wang (1st), Boqi Zhang, and Jinxin Zong completed all experiments and visualizations. Tong Chen, Guitian He, and Maosheng Cao provided sequencing data analysis and technical support. Yueying Wang, Xue Chen, Yuxin Luo, Caomeihui Shen, and Yanan Sun performed data curation. All authors contributed to reviewing the manuscript and approved the final version.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Rajesh Sarkar and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Frank Avila and Kaliya Georgieva.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, N., Zhang, B., Chen, T. et al. Obesity impairs spermatogenesis via Leydig cell ferroptosis induced by liver-derived exosomal miR-122-5p. Commun Biol 9, 318 (2026). https://doi.org/10.1038/s42003-026-09612-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-026-09612-z
