
Abstract
Background
Exercise training often produces less weight loss than expected, a phenomenon termed exercise-induced energy compensation, but the underlying mechanisms remain unclear. This study aimed to quantify metabolic and behavioral compensation to aerobic exercise training.
Methods
Sixteen sedentary adults with overweight completed a 12-week supervised aerobic walking intervention targeting an energy expenditure of 20 kcal/kg/week. Total daily energy expenditure was measured using doubly labeled water, and whole-room calorimetry was used to assess changes in resting and sleeping metabolic rate (RMR, SMR) and diet-induced thermogenesis (DIT). Volumes of highly metabolic organs were quantified by magnetic resonance imaging. Physical activity was monitored objectively, walking economy was assessed during standardized treadmill walking, and dietary intake was evaluated using self-report and intake-balance methods. A parallel mouse exercise model was used to explore tissue-level adaptations.
Results
Exercise training induces substantial energy compensation, resulting in minimal body weight loss despite improved body composition. Total daily energy expenditure increases, while RMR and SMR decrease, accounting for most of the compensatory response. Liver and kidney volumes decrease by 5%, while brain volume remains unchanged. Exercise improves walking economy and leads to smaller-than-expected increases in daily moderate-to-vigorous physical activity. Dietary intake and DIT remain unchanged. In mice, exercise is associated with increased cellular density and mitochondrial content in the liver, indicating structural and metabolic remodeling.
Conclusions
Aerobic exercise training engages compensatory physiological and behavioral mechanisms that constrain energy expenditure. Reductions in basal metabolism, improved movement efficiency, and selective remodeling of metabolically active organs may collectively limit exercise-induced weight loss.
Plain language summary
Many people exercise to lose weight, but results are often smaller than expected. This study explored why this happens. We studied adults with overweight who completed a 12-week supervised walking program and mice who were also regularly exercised. We measured how much energy they used each day, how their bodies used energy at rest, how active they were, and the size of key internal organs using advanced imaging. We found that body weight changed very little despite regular exercise. The body adapted by using less energy at rest, walking more efficiently, and reducing the size of highly metabolic organs such as the liver and kidneys. These changes helped conserve energy and may explain why exercise alone does not lead to major weight loss
Similar content being viewed by others
Introduction
Exercise is commonly recommended as a strategy for weight loss and weight maintenance1. However, exercise intervention studies have demonstrated that exercise alone, absent dietary modifications, results in less weight loss than expected. The prevailing hypothesis is that individuals behaviorally compensate for the increased exercise-induced energy expenditure (ExEE) through elevations in energy intake2. However, it is possible that exercise-induced decreases in non-exercise energy expenditure3,4,5 also contribute to the less-than-expected weight loss.
The effect of exercise on total daily energy expenditure (TDEE) is assumed to be a linear relationship: that is, increased physical activity (PA) is assumed to result in elevated TDEE in a dose-response manner, known as the additive model. However, recent free-living and population-based studies have challenged this assumption. In a study performed from diverse adult populations across Africa and North America, the observed relationship between physical activity and TDEE was linear at low activity levels but plateaued at high activity levels6. Based on these findings, a constrained TDEE model was suggested, in which the body metabolically adapts to offset the increase in ExEE by reducing energy expenditure in one or more components of TDEE7,8. This metabolic adaptation (MA) is also referred to as “adaptive thermogenesis”, “metabolic compensation”, or exercise-induced energy compensation (ExEC). The constrained model may explain why exercise studies show modest (2–3 kg), less than predicted, weight loss despite strenuous exercise (> 2000 kcal/week)1,8,9,10,11,12,13,14,15,16,17,18. Therefore, although exercise is associated with various health benefits, the effectiveness of exercise to induce weight loss is debatable.
From an evolutionary perspective, the constrained energy model suggests that sustained increases in PA may trigger compensatory downregulation of non-essential metabolic processes in order to preserve resting metabolic rate (RMR) and vital organ function, which are prioritized for survival8. However, two recent systematic reviews have reported mixed results on the effect of aerobic exercise on RMR, with published studies demonstrating a high risk of bias due to heterogeneous methodologies and a limited number of interventions19,20. ExEC may be mediated by changes in the sizes of vital organs, leading to a decreased metabolic rate. Although the brain (240 kcal/kg/d), liver (200 kcal/kg/d), and kidneys (400 kcal/kg/d) represent only about 5% of the body weight21,22,23, they account for 51% of the total RMR. In contrast, adipose tissue and skeletal muscle have relatively low metabolic rate (4 and 13 kcal/kg/day, respectively) at rest21,22,24. Studies in humans have shown that weight loss induced by caloric restriction is associated with reduced organ size25,26,27,28,29,30,31, except for the brain, where the mass increased or remained unchanged26,30,32,33,34. If this reduction in organ size leads to a reduction in the metabolic rate of these highly metabolically active organs, this would lower the body's energetic demands to balance overall energy needs. In contrast, the current literature concerning the impact of exercise-induced elevated EE on organ size and the relationship to MA is scarce.
The effects of exercise on other components of TDEE, such as sleeping metabolic rate (SMR) and diet-induced thermogenesis (DIT), have received markedly less scrutiny. Very few interventional trials have tested the effects of chronic aerobic exercise on SMR in humans 35,36,37,38. Evidence from studies that measured the effect of calorie restriction on TDEE suggests that changes in DIT may contribute to the decreases in TDEE29,39,40,41,42. Furthermore, with a substantial portion of the modern-day 24-h cycle spent in a postprandial state, changes in DIT could considerably impact TDEE43. There have been contradictory reports on how exercise affects DIT, with negative, positive, or no significant associations between DIT and exercise44,45,46,47,48.
Behavioral adaptations such as reducing spontaneous and occupational PA9,16,35,49,50,51,52,53,54,55,56,57 and increasing sedentary time may reduce non-exercise PA during structured exercise intervention9,16,36,50,51,58,59,60,61,62,63. Therefore, individuals who practice exercise training may increase their sedentary time and decrease their non-exercise PA56,64,65. It is also possible that improvements in exercise economy result in reduced EE costs associated with non-exercise PA. In caloric restriction-induced weight loss studies, physiological adaptations result in an approximately 20% increase in muscle work economy66. However, whether such an effect occurs in exercise-induced weight loss remains unclear. According to a recent systematic review, only 23% of exercise intervention studies noted a reduction in physical activity energy expenditure (PAEE)67 because it can only be indirectly approximated by subtracting RMR from TDEE (after accounting for DIT), it would be more methodologically sound to directly assess metabolic economy within skeletal musculature.
The constrained model predicts that a decrease in one or more components of TDEE (i.e., SMR, RMR, DIT, and/or non-exercise PA) contributes to less-than-expected increases in TDEE during chronic aerobic exercise training. We conducted this study to quantitatively assess physiological and behavioral compensation to exercise and to determine whether ExEC mechanisms act, at least partly, by diminishing the volume of metabolically active organs and enhancing cellular energy economy.
Methods
Study participants
Sixteen sedentary (n = 8 women), overweight (BMI 25–30 kg/m2) adults between the ages of 21–45 years were recruited to participate in this 12-week supervised exercise intervention study (Fig. 2). Exclusion criteria included participation in another exercise or weight loss program in the past 6 months, non-stable weight (>±5%) over the past 6 months, current regular exercise >1 h per week, smoking within the past 6 months, being postmenopausal, breastfeeding, pregnant, or pregnant within the past 6 months, contraindications for MRI scans (tattoos, intrauterine devices, permanent makeup, etc.), previous bariatric surgery, cardiopulmonary disease (e.g., recent myocardial infarction or unstable angina), musculoskeletal or neuromuscular impairments that preclude exercise training, other health issues include cancer, diabetes, thyroid disease, hypertension, chronic renal failure, cognitive impairments, or use of drugs that potentially impact body metabolism. A standard medical questionnaire prepared by the Israeli Ministry of Sport was used to evaluate potential study recruits, and a physician approved the results.
Ethics
The study was approved by the Helsinki committee of Sheba Medical Center (7214-20-SMC) and the ethics committee of Tel Aviv University (approval no. 0001932-4 for human study and TAU-MD-IL-2504-121-5 for animal experiment). All participants provided informed consent before participating in the study, either in person or via Zoom (approved during the pandemic). The human study was registered on 7 July 2020 at ClinicalTrials.gov (NCT04460040) and MyTrial.gov (7214-20SMC) sites.
Screening phase and eligibility
The study included a screening stage before participant enrollment. The study was advertised on social media (primarily on Facebook) with a brief description of the study, and potential participants were asked to complete an electronic questionnaire designed to assess eligibility. Eligible responders were then invited to an orientation session where basic anthropometric measurements (weight, height) were obtained, and detailed information about the study was provided. Following this visit, potential participants underwent a 1-week run-in phase designed to assess adherence rate by having them wear an accelerometer for 7 days and perform two 30-minute exercise sessions outdoors or in the gym and a 30-min session in the whole-room indirect calorimetry at 5 km/h. Screening information from all participants was documented and maintained in a screening log. Eligible consenting participants were scheduled to start the control phase.
Study design
To determine the effect of exercise on MA, the first phase of the study comprised two weeks of pre-intervention control phase (day -14 to day 0), when participants did not engage in any exercise training but were monitored for TDEE and its components (RMR, SMR, and DIT). The volume of highly metabolic organs was evaluated, and the walking economy, body composition, and daily PA intensity were assessed (Fig. 2). Subsequently, the participants commenced a 12-week active walking phase, targeting 65–75% of HRmax68 for 12 weeks. During this period, they were restricted from engaging in any other forms of exercise.
Exercise was individually tailored to achieve a target exercise energy expenditure of 20 kcal/kg/week. This target was explicitly chosen to be (a) consistent with current recommendations for the amount of exercise required to elicit health benefits and weight loss1,69,70,71, (b) appropriate and achievable for a sedentary overweight population. Given the evidence of MA, we expected that within 1–3 months, participants would adapt to the exercise regime, and their TDEE would return to near the level before the change in exercise72. Individual weekly energy expenditure was calculated pre-intervention from the VO2-HR ratio, determined during the VO2max test73,74,75, and a weekly exercise plan was created. In the first two weeks of the active phase, participants performed 70% of the weekly plan to adapt to the new regime and avoid injury. After 6 weeks, the prescribed ExEE was recalibrated for each participant to maintain the target of 20 kcal/kg/week. This adjustment was based on the participant’s current body weight at week 6 and their updated Oxygen consumption (VO₂)–HR ratio, which was derived from a near-maximal VO₂ test. The revised heart rate–based energy expenditure prescription ensured that the ExEE remained aligned with both the participant’s physiological capacity and the relative dosing goal throughout the 12-week intervention.
Once a week, participants exercised on a treadmill in the whole-room indirect calorimetry laboratory under supervised conditions; the other exercise sessions were conducted at home or in a gym on a treadmill or outdoors. All exercise training was monitored using an optical HR sensor (Polar, OH1) paired to a smartphone to track exercise adherence to HR range goals objectively. Real-time HR monitoring for participants was enabled76,77. Participants utilized the online platform https://flow.polar.com/diary to promptly upload their exercise data via the ‘Polar Beat’ application. From the data obtained from their devices, we were able to calculate several key parameters, including exercise duration (minutes/week), mean intensity during exercise (percentage of maximal HR), and the frequency of exercise (sessions/week). To promote adherence to the exercise regimen, participants were given the autonomy to choose their preferred frequency of exercise sessions per week, with a minimum requirement of 3 sessions, as well as in determining the intensity during each session. Additionally, we conducted weekly telephone or text message consultations with participants. During these consultations, we reviewed exercise data collected from the HR sensor and assessed any potential obstacles hindering their participation in the exercise program. We aimed to identify and address these obstacles to optimize their commitment to the program.
Female participants informed the investigators about the onset of their menstrual cycle. Day 1 of the menstrual cycle was considered the starting point. Unless a participant was using continuous hormonal oral contraceptives, women were only tested during the luteal phase (days 15–28) to assess walking economy and any component of TDEE. This approach aimed to minimize any confounding effects of hormones78,79,80. Also, participants were asked to report any exercise-related adverse events or injuries during the study.
All post-intervention assessments were conducted during the final 14 days of the intervention period, while participants were still actively engaged in the prescribed training sessions. This study is reported in accordance with the CONSORT 2025 guidelines for the reporting of clinical trials (Table S1).
Body composition assessment
Body weight was measured to the nearest 10 g on a digital scale (Seca, 'mBCA') after 12 h of overnight fasting while wearing light clothing. Height was measured barefoot with a wireless digital stadiometer (Seca 274) to the nearest 0.1 cm at screening. Waist and hip circumference were measured with a tape measure81. To determine the individual dosage of DLW (doubly labeled water), FFM was evaluated using multi-frequency bioelectrical impedance analysis (BIA) with the Seca “mBCA” body composition analyzer after a 12-h fasting period. Changes in body composition, including FM and FFM, were assessed using the DLW method82,83. Body weight, waist and hip circumference, and body composition measurements were made in the control phase and the post-intervention period, with post-intervention assessments completed within the final 7 days of the 12-week exercise intervention to detect the MA changes due to exercise84.
Energy metabolism assessment
Doubly labeled water
TDEE was measured using DLW, the gold standard methodology for assessing free-living energy expenditure 85. TDEE was evaluated over 10 days at baseline (control phase) and during the last 14 days of the intervention. This time frame was considered a representative assessment considering the 2–4 month time span of the study86,87. Before DLW dosing, a baseline urine sample was obtained following a 12-h overnight fast to determine the background abundances of 2H and 18O. Participants then consumed an oral dose containing 1.8 g/kg total body water (TBW), estimated as 73% of FFM of 10 atom percent excess (APE) 18O and 0.12 g/kg TBW of 99.9 APE 2H. Participants provided urine samples 4 and 5 h after dosing and were sedentary for the entire 5-h period, with only very limited PA, such as working on a computer or reading a book. On day 10 of the DLW dosing, participants were instructed to discard their first urine void of the day. They then visited the laboratory, where their second and third voided urines of the day were collected at a 45-min inter-collection interval. Participants abstained from eating or drinking for 1 h before the second void.
Sample aliquots (4 mL) were frozen at −80 ⁰C until analysis. They were shipped on dry ice to the University of Colorado for analysis. Frozen urine samples were thawed and centrifuged and then analyzed for 18O and 2H enrichment using Off-Axis Integrated Cavity Output Spectroscopy (OA-ICOS), as previously described88. TBW was calculated as the average of dilution spaces of 2H and 18O, and CO2 production rate was calculated using a modification89 of the original two-point equation of Schoeller et al. 90. TDEE was calculated using the equation of Weir, assuming a respiratory quotient of 0.86, and averaged over 10 days91.
Whole human room calorimetry
The components of TDEE (SMR, RMR, DIT, and ExEE) were measured before and after the intervention using indirect calorimetry in a small whole-room indirect calorimetry (~11,500 liters). The room is furnished with a bed, chair, and desk with internet access and telephone. An airlock was used for the exchange of food. Air in the calorimeter was removed at a rate of 240 L/min. O2 uptake and CO2 production were measured using gas analyzers (Promethion Systems), and gas concentration was analyzed using the ExpeData software (Sable Systems International, models GA-3 m2 and FG-250 flow generator). The O2 and CO2 gas analyzers were calibrated before each experiment and once a week using standardized gas mixtures92. To calculate energy expenditure, Weir's equation was used91. Repeated quality control tests of the whole-room indirect calorimetry using a 4-h N2 injection test showed a mean recovery of 97.8 ± 4.3% for O2 consumption93. All post-intervention assessments in whole-room indirect calorimetry were conducted during the final 7 days of the 12-week exercise intervention
Sleeping metabolic rate
SMR was measured in the whole-room indirect calorimetry, which allowed normal sleep without needing a facemask, hood, or probes17,49,94. Participants arrived at 7 p.m. after having a typical light Israeli dinner at home. The dinner comprised an independently selectable assortment of white cheese, eggs, tuna fish, bread, fresh vegetables, and fruits. Participants were instructed to maintain a sitting posture until 11 p.m. Subsequently, they were directed to turn off the lights and sleep for 7 h, waking up at 6 a.m. Several factors have been shown to modulate SMR, including daily PA, sleep stages, menstrual cycle, daily DIT, and environmental temperature95,96,97. To mitigate the influence of such confounders, participants were tested at a thermoneutral room temperature (22 °C–27 °C)98, had their last light meal at 7:00 p.m. at home, refrained from beverages containing caffeine and alcohol, and performed their last exercise session at least 24 h before the test. A radar sensor was used to continuously monitor participants' body movements during their stay in the room to ensure that they maintained a minimum level of PA99. Upon awakening, participants were escorted out of the room, voided their bladders, allowed them to brush their teeth, and entered to consume standardized/test meal.
Resting metabolic rate
RMR was measured pre- and post-intervention on a separate morning visit, distinct from the SMR measurement night, in the whole-room indirect calorimetry over 60 min. Measurements were made between 7 a.m. and 9 a.m. after a 12-h overnight fast and at least 24 h after the last bout of any structured exercise. Participants were lightly clothed, and measurements were made in a thermoneutral (22 °C–25 °C), dimly lit, quiet room. Participants were instructed to remain motionless and awake supine.
Diet-induced thermogenesis and energy intake
Following an overnight sleep in the whole-room indirect calorimetry, participants consumed a standardized breakfast meal (Fig. 6A) within 30 min of waking at 6:15 a.m., after ~11 h of fasting. Energy expenditure was continuously measured for 4 h while the participant remained stationary in a supine position. Only very low-intensity PA, such as watching television or a smartphone, reading a book, etc., was allowed44,99,100. The standardized/test meal was analyzed for energy and macronutrients in a formal laboratory setting (AminoLab LTD). DIT was determined as a cumulative increase in energy expenditure.
Study participants were instructed to maintain their habitual diet and to maintain the same caloric intake. During the control phase and in the final week of the exercise intervention, participants meticulously logged all food and drink consumption, including photographs of all meals, using the MealLogger© smartphone application (food record)101,102. To ensure the accuracy and completeness of the food records, phone interviews (24-h diet recall) were conducted the morning after each recorded day to recall any items that may have been overlooked. The total self-reported energy intake was calculated using “Tzameret”, the electronic food and nutrient database administered by the Israeli Ministry of Health103 Energy intake was estimated using the “intake-balance method”.
Walking economy
Walking economy was assessed pre- and post-intervention during treadmill walking at low-intensity (2 km/h) and moderate-intensity (4 km/h) speeds, with post-intervention measurements conducted within the final 7 days of the exercise program. For LIPA assessment, participants commenced with a 4-min seated rest before engaging in 4 min of walking. Although 4 min may be considered a relatively short duration, previous studies indicate that steady-state oxygen uptake is typically achieved within 2–3 min at low intensities in healthy individuals104,105. Oxygen consumption and carbon dioxide production were measured breath-by-breath using a face mask by an indirect calorimetry metabolic cart (Quark CPE, COSMED). The metabolic system was calibrated using both a 3-L calibration syringe and standard gas calibration (16% O2 and 5% CO2) before each test. The tests were conducted at the Sylvan Adams Sports Institute at Tel Aviv University. The MIPA was assessed at a fixed walking speed of 4 km/h for 45 min in the whole-room indirect calorimetry chamber and was considered an additional session conducted separately from the routine exercise schedule to enhance compliance. Although the absolute intensity was fixed across participants, relative intensity may have varied individually; this standardization enabled direct pre–post comparisons of energy cost. Importantly, the energy expenditure from this session was included within the total prescribed weekly dose of 20 kcal/kg/week. All measurements were performed following an overnight fast (12 h), 2 h from any beverage consumption, and 24 h from consumption of caffeine, alcohol, or exercise training. Additionally, all tests were concluded at 10:00 a.m.
Measurements of organ volume, visceral fat distribution, and hepatic fat content
The volume of highly metabolically active organs and adipose tissue was measured by MRI (a gold standard for assessment of human organ size106) in the control phase and after 12 weeks of exercise intervention. All MRI scans were performed using the Magnetom Prisma 3-Tesla 64-channel head coil scanner of Siemens (Medical Solutions) at the Alfredo Federico Strauss Center for Computational Neuroimaging located at Tel Aviv University. High-resolution, T1-weighted images of the brain were collected using a rapid gradient echo (MPRAGE) pulse sequence with a voxel size of 0.9*0.9*0.9 mm parallel imaging factor of 2, TR/TI/TE = 2400/1000/2.78 ms, and flip angle = 8. T2-weighted anatomical images were acquired using a sampling perfection with application-optimized contrast using the flip angle evolution SPACE sequence, with the following parameters: TR/TE = 3200/554 ms, resolution of 0.9 mm3. A field map was acquired using spin-echo echo planar imaging with opposite phase-encoding directions.
The volume of human liver, kidneys, and abdominal fat was determined based on T1 Dixon scans of 3-mm slice thickness with no interslice gap, with voxel size of 1.2*1.2*5.0 mm, parallel imaging factor of 2, TR/TE1/TE2 = 4/1.24/2.47 ms, flip angle = 9 as previously described107. Images were acquired in axial, coronal, and sagittal planes. Each subject underwent scanning for in-phase, out-of-phase, fat-phase, and water-phase images, utilizing a 17-s breath-hold technique to minimize motion-related artifacts.
To precisely determine human visceral adipose tissue distribution, MRI scans were conducted at three axial levels (L5-L4, L4-L3, and L3-L2). A continuous line was drawn over the superficial fascia to differentiate between deep subcutaneous adipose tissue and superficial subcutaneous adipose tissue. Mean values for visceral adipose tissue, deep subcutaneous adipose tissue, and superficial subcutaneous adipose tissue were calculated based on the three axial slices.
Following the MRI scans, manual raw image analysis of the liver and kidneys was performed using a custom MATLAB (MathWorks) code developed in-house specifically for this study. The code was designed to leverage segmentation algorithms to accurately identify and isolate the different organ regions from the surrounding anatomical structures and the various adipose tissues present in the acquired images. Once the regions of interest were segmented, the custom MATLAB code employed volumetric analysis techniques to calculate the volume. All MRI scans of the liver and kidneys were divided equally between two investigators for analysis, with each investigator assigned 8 human participants and 3 mice. Investigators performing manual segmentation and volumetric analysis were blinded to both the intervention timepoint and participant identity. To evaluate the consistency and reliability of these analyses, the interclass correlation coefficient (ICC) was employed to compute the external and internal validity of the liver and kidney MRI scans. In sub-sample analysis, the ICC for internal validity was 0.995% [95% confidence interval (CI): 0.981, 0.999] and 0.961% (95% CI: 0.955, 0.967) corresponding to each investigator, respectively, and the ICC for external validity, representing the agreement between the two investigators, was 0.991% (95% CI: 0.977, 0.996). Hepatic human fat content was determined using MRS, a well-validated108 analytical technique that detects radiofrequency electromagnetic signals emitted by atomic nuclei within molecules. A voxel measuring 20 × 20 × 20 mm was carefully positioned within the right hepatic lobe, avoiding major vessels based on a preliminary survey scan. The acquired data were analyzed using the INSPECTOR software (Columbia Technology Venture license 17130, 2016) by an experienced researcher blinded to the intervention groups. The total hepatic fat fraction within the image was determined as the ratio of the sum of the area under all fat peaks to that of the area under all fat and water peaks.
Monitoring physical activity
Alterations in daily free-living non-exercise PA and sedentary behavior were evaluated for 7 days before and during the final 7 days using a tri-axial accelerometer (ActiGraph GT9X Link)109. The accelerometer was securely fastened to the participant's non-dominant wrist using disposable straps to ensure the device could not be intentionally removed during the monitoring period. The data from the accelerometer were utilized to estimate metabolic equivalents (METs) and to quantify the duration of different PA intensities.
Cardiorespiratory assessment
Cardiorespiratory status was evaluated pre- and post-exercise intervention, with post-assessment conducted during the final 7 days. Sleeping BP, diastolic, systolic, and HR were measured using photoplethysmography technology110. The photoplethysmography device was placed on the wrist and transmitted light in specific red (~650 nm) and infrared (~880 nm) wavelengths through the tested tissue. The measurements were taken during the assessment of SMR using a BB-613W device (BioBeat). Maximal aerobic capacity (VO₂max) was assessed using a graded treadmill exercise test. The test began with a 2-min standing rest on the treadmill to collect baseline cardiorespiratory data. The exercise phase started at a speed of 4 km/h for men and 3 km/h for women, with speed increased every 2 min until reaching a maximum of 7 km/h for men and 6 km/h for women, all performed at a 1% incline. Once the target speed was reached, the treadmill gradient was increased by 2% every minute, while maintaining the final speed, until volitional exhaustion.VO₂ and HR were continuously monitored to confirm maximal effort. VO₂ was measured using an indirect calorimetry metabolic cart, as detailed in Section 5.769,111. This test was also used to determine each participant’s HRmax for exercise prescription and VO₂–HR calibration purposes.
Liver volume and phenotype of hepatocytes in mice
Mice
Animal experiments were performed in accordance with the guidelines of the Tel Aviv University Institutional Animal Care and Use Committee with institutional policies and approved protocols (IACUC permit: 01-21-047). Mice were housed in individually ventilated cages in reverse light with 22 ± 1 °C temperature and 32–35% humidity with access to ad libitum water and food intake (mouse chow diet pellets). 22-week-old C57BL/6JRccHsd female mice (Envigo) were habituated for 12 days upon their arrival to the animal facility before experiment initiation. Female mice were chosen based on their increased metabolic response to exercise compared with males112.
Training protocol
All mice were age- and sex-matched, housed under identical conditions, and experiments were performed in parallel to minimize batch effects. Mice were arbitrarily assigned to either the active group (forced treadmill running) or to the control group (sedentary) in alternating order at the time of assignment, prior to treatment initiation, to ensure balanced group distribution, as previously described112, n = 10 mice in each group. Briefly, every day before the start of the exercise, mice were habituated in the experimental room for 20 min. On the first day, following the lighting habituation, mice were habituated on the treadmill (Panlab Harvard Apparatus) for 10 min and at a speed of 5 cm/s. From day 2 and onward, the mice were habituated in the experimental room for 20 min, followed by treadmill exercise starting at 18 cm/s for 5 min. The speed was then increased by 2 cm/s increments until reaching 24 cm/s, which was sustained for 8 min. The speed gradually decreased by 2 cm/s every minute until 18 cm/s. The total duration of the exercise session was 20 min per mouse; performed every other day for 8 weeks. The treadmill protocol included a mild electrical stimulus (0.2 mA), used only as a motivational cue during adaptation (see Supplementary Movie 1). Under our conditions, mice rarely experienced shocks during running, and cumulative exposure was minimal (<2 s per session). Control mice were habituated to the experimental room in the same way as the active group and were then placed on to the treadmill apparatus for 20 min (total experimental time) without turning on the machine (no speed, no electric shock). Following the exercise, the mice were returned to their home cages. This study is reported in accordance with the ARRIVE guidelines for the reporting of animal experiments (Table S2). All quantitative mouse outcome analyses including organ weights, histological scoring, image quantification and MRI were performed in a blinded manner, with investigators unaware of group allocation at the time of data acquisition and analysis.
Organ volume quantification in mice
At the end of the exercise protocol, organ volumes were quantified using a Prisma 7-Tesla 64-channel scanner (Siemens Medical Solutions) at the Alfredo Federico Strauss Center for Computational Neuroimaging, Tel Aviv University. The volumes of the kidneys were determined using T1-weighted Dixon sequences with a slice thickness of 1 mm, no interslice gap, and a voxel size of 0.2 × 0.2 × 1.0 mm, parallel imaging factor of 2, TR/TE1/TE2 = 4/1.1/2.2 ms, flip angle = 10°. Images were acquired in axial, coronal, and sagittal planes. Scans were performed for in-phase, out-of-phase, fat-phase, and water-phase images, with measures taken to minimize motion artifacts.
Tissue processing
At the conclusion of the training protocol, mice were anesthetized, livers were harvested, and either flash frozen in liquid nitrogen (for proteomic analysis) or fixed with 4% paraformaldehyde at 4 °C dehydrated in a graded ethanol series, and embedded in paraffin wax. The PFA-fixed and paraffin embedded liver tissues were sliced into 10-mm sections and dried overnight at 37 °C. Sections were stained with hematoxylin (Sigma-Aldrich) and eosin (Sigma-Aldrich) and mounted with DPX Mountant (Sigma-Aldrich), according to the manufacturer’s instructions, and imaged at 20x magnification with a Nikon brightfield microscope. Blind quantification of the nuclei was done by using a defined ROI (kept consistent through all images) in 10 random areas of each image (1 slide scan per mouse, n = 10 in both the control and active groups). Stained nuclei were counted using the counting tool on the ImageJ program (n ≥ 2 mice per group).
RT-qPCR analysis of mouse liver tissue
RT-qPCR analyses of mitochondrial genes ND1 and S16 were performed by de-paraffinizing 4–10 slices liver tissue. First, 500 µl of xylene was added, tissue was incubated at 50 °C for 3 min, and the supernatant was removed (18,000 × g, 1 min, room temperature). Next, 500 µl EtOH was added, samples were vortexed and centrifuged as above, and the supernatant was removed. To the supernatant was added 500 µl Proteinase K buffer (20 µM Tris∙HCl, pH 8, 1 µM CaCl2, 0.5% SDS), and vortexing and centrifugation were carried out as above. Finally, the samples were incubated with 0.5 mg/mL Proteinase K (150–250 µl depending on tissue size) overnight at 37 °C. Next, DNA was extracted using the GenElute Mammalian Genomic DNA Miniprep Kit (Sigma Aldrich, G1N350) as per the manufacturer's instructions with the following adjustments: The lysis solution was added at an equal volume of the Proteinase K incubation buffer, and EtOH volume was adjusted in the same manner. Samples were eluted with 100 µl of UPW.
Proteomic analysis of mouse liver tissue
At the conclusion of the training protocol, organs were harvested and flash frozen for proteomic analysis. Samples were then digested by trypsin, analyzed by LC-MS/MS on Exploris 480 (Thermo). Data analysis includes Maxquant 2.4 analysis and identification with the andromeda search algorithms against the mouse proteome from the Uniprot database, and a decoy database (in order to determine the false discovery rate). All the identified peptides were filtered with high confidence −1% FDR threshold. (*FDR = false discovery rate, is the estimated fraction of false positives in a list of peptides). Quantitation was done by calculating the peak area of each peptide. The abundance of the protein is the sum of all associated peptide group abundances. In addition to the original intensities, LFQ normalized intensities were added after transformation to log2. Missing values were replaced by 17 (in log 2). T-tests were done using the Perseus software and were added to the report.
Proteomics data were then filtered for proteins whose fold change was ≥ 1.2 (ratio active:control) and were uploaded to the QIAGEN IPA (QIAGEN Inc.) system. The upstream regulator analysis was generated through the use of QIAGEN IPA (QIAGEN Inc)113. Similarly, the same list of enriched proteins was uploaded to the Proteomap system (https://bionic-vis.biologie.uni-greifswald.de/)114,115.
Sample size calculation
The sample size calculation for the primary outcome, the within-subject change in TDEE following PA, was informed both the theoretical framework proposed by Pontzer et al. 6, demonstrating constrained energy expenditure, and empirical data from long-term aerobic exercise interventions. While Pontzer et al. describe the expected metabolic compensation with increased activity, they do not provide an intervention-based change in kcal/day. Therefore, we used results from studies employing doubly labeled water, such as Willis et al. 16, which reported increases of approximately 250–300 kcal/day in TDEE in response to structured exercise. To remain conservative, we selected a meaningful within-subject effect size of 120 kcal/day. Assuming a correlation of 0.5 between baseline and post-intervention TDEE, α = 0.05, and β = 0.2 (i.e., 80% power), the required sample size to detect a meaningful change in TDEE was calculated to be 15 participants. We therefore enrolled 16 participants to ensure sufficient power.
Statistical analysis
Quality control was performed by investigators blinded to intervention time points. Before the statistical analysis, the following assessments were completed for each variable: (1) minimum and maximum values; (2) mean ± SD and range; (3) number of participants included with no missing data. Changes in continuous variables from pre- to post-intervention were presented as means ± SD. During the data analysis process, no outlier manipulation or variable transformation was performed, ensuring that all the data was analyzed in its original form without any alterations, allowing for an accurate and unbiased analysis of the research findings. Due to the small sample size (n = 16), all variables were tested for normal distribution by the Shapiro–Wilk test. Variables not meeting normality assumptions were analyzed using non-parametric tests, including the Signed Rank Wilcoxon test or Mann–Whitney U tests as appropriate based on study design. Otherwise, analyses were performed using Student’s T-tests. Linear correlations were analyzed using Spearman's two-tailed correlation rank test. The null hypothesis was tested against two-directional alternatives at the nominal 0.05 significance level. All statistical analyses were performed using SPSS software (SPSS Statistics for Windows, version 23; IBM Corp.). Figures were created using Graph Pad Prism (GraphPad Software, v. 8.4.1).
Data processing
The cumulative weekly adherence of individuals to the exercise protocol was determined by calculating the total duration, in minutes, of exercising at 65% or 75% of their HRmax during each session. As already described, weight compensation is the difference between the amount of weight loss predicted from ExEE and observed weight loss from baseline (day 1) to follow-up (Eq. 1):
Additionally, it can be displayed as a percentage (Eq. 2):
Predicted weight loss was calculated using two methods: the conventional model assumed changes in 1 kg of body weight is equal to 7700 kcal116,117, and a validated dynamic energy balance NIH model118 (link to the web model), which accounts for factors including age, sex, body weight, body fat, height, RMR, EI, PA level, ExEE change, and length of intervention. The dynamic energy model does not account for the impact of ExEC. Positive values in weight change and percentages indicate a lower weight loss than expected. Hence, a weight change value of zero or 0% signifies a complete absence of any deviation from the expected weight change119.
To calculate ExEC changes in body energy (stored or lost), were quantified from the pre- and post-differences in FM and FFM. These changes in body composition reflect long-term shifts in energy balance120. Using the established energy densities for FM and FFM121 and incorporating the variables of time (t) and change (∆), the following equation was applied (Eq. 3):
Furthermore, the individual ExEC was calculated by subtracting the changes in body energy changes from the additional ExEE (Eq. 4). The percentage of ExEC was then determined by dividing this value by the ExEE 117,121,122 (Eq. 5):
Negative values of ExEC indicate that not all structured ExEE contribute to changes in FM and FFM. Additionally, 0% ExEC indicates perfect alignment with ExEE and the absence of any energy compensation, whereas 100% indicates that all ExEE was compensated for. It is important to note that ExEC encompasses both components of the energy balance equation, including changes in energy expenditure and energy intake, thereby reflecting both physiological and behavioral adaptation.
To evaluate the presence of post-intervention MA in terms of energy expenditure at the levels of TDEE, SMR, and RMR, we compared energy expenditure at baseline to the values at the end of the 12 weeks of exercise intervention by two different approaches. First, we calculated EE relative to FFM. Second, a linear regression analysis following the procedures of Galgani and Santos123, with baseline (pre-intervention) TDEE/SMR/RMR parameters as the dependent variable and baseline FFM as the independent variable (the only significant covariate). We then quantified MA at the group level by the difference (residual) between the measured post-intervention energy expenditure and that which was predicted (Eq. 6) from the current FFM using the baseline linear relationship.
Furthermore, the percentage MA was determined for TDEE, SMR, and RMR by dividing MA by the predicted energy expenditure (Eq. 7). A negative value indicates the presence of MA.
However, to identify MA at the individual level (Eq 8), we compared the difference between residual values from pre- to post-intervention, as suggested by Galgani and Santos123 (EE component: TDEE, SMR, or RMR). Negative residuals (▲Residuals<0) indicate metabolic adaptation—i.e., measured energy expenditure is lower than predicted based on FFM—while positive residuals indicate higher-than-predicted EE. While no standardized cutoff for MA exists in the literature, we applied a 5% or greater reduction as a practical indicator of meaningful physiological change. This approach reflects the understanding that even small declines in energy expenditure can meaningfully influence energy balance and weight outcomes.
The SMR was calculated using published procedures with pre-determined intervals: the sum energy expenditure between 02:00 a.m. and 5:00 a.m.124,125 and extrapolated to 24 h (multiplying by 8). Movement was continuously monitored using a radar-based detection system embedded in the whole-room indirect calorimetry. Only data from periods with spontaneous PA below 1.5% were used to SMR estimation126. While this approach minimizes the influence of wakefulness or PA, we cannot confirm with certainty that participants were continuously asleep during this period. RMR was determined as the lowest consecutive 20-min stable (<10% of average SD) interval during the measurement127. RMR measurements that did not meet these criteria were considered invalid and excluded from the analysis. TDEE, SMR, and RMR changes were adjusted for changes in FFM and were reported relative to FFM, as FFM is the primary predictor of energy expenditure 19. DIT was calculated as the area under the curve of a 4-h consecutive interval after breakfast128,129. To ensure maximal precision in testing the energy cost of arousal, only measurements of SMR, RMR, and DIT in which spontaneous PA was less than 1.5%, as determined by radar, were considered valid. Energy expenditure values were obtained from minute-to-minute intervals and used Weir's equation91 as follows (Eq. 9):
HR and BP were averaged during SMR. PAEE was calculated as the difference between TDEE and RMR, adjusted for the estimated DIT, 10% from TDEE130,131 or using individual-level DIT data as follows (Eq. 10):
Although DIT was measured following a standardized meal in whole-room indirect calorimetry, we did not extrapolate these data to obtain individualized 24-h DIT estimates. Instead, for the purpose of calculating PAEE using Eq. 9, a fixed value of 10% of TDEE was assumed, consistent with conventional practice in metabolic studies.
PA level (Eq. 11) was calculated as the ratio of TDEE to RMR132:
Data generated from the low-intensity walking economy test was downloaded from the metabolic cart using a Microsoft Excel spreadsheet. To approximate steady-state VO₂ and VCO₂ levels, energy expenditure was averaged over the final 2 min of the 4-min walking bout. While this duration is generally sufficient for reaching steady-state at low intensities104,105 we acknowledge that individual variability may limit its confirmation in all participants133. Gases during the moderate-intensity walking muscle economy test in whole-room indirect calorimetry were defined within a 20-min steady-state phase.
Accelerometer data was converted to raw 30 Hz and 1-min epoch CSV files to establish non-exercise PA. The raw CSV files were used to estimate METs and time spent in various PA intensities (i.e., sedentary behaviors (<1.5 METs), LIPA (1.5–2.99 METs), MIPA (3.0-5.99 METs), vigorous-intensity PA (VIPA; ≥ 6.0 METs), and MVPA (≥3.0 METs)) using the Hibbing two-regression adult non-dominant wrist algorithm retrieved from Hibbing et al. 134. Only data collections capturing full 24-hour periods of accelerometer monitoring were considered eligible for inclusion in the analytical sample. The accelerometer data has the potential to categorize certain activities recorded during the structured walking exercise as VIPA rather than MIPA135 because the accelerometer translates movements into PA intensities rather than HR levels, and because, during outdoor exercise, there may naturally be higher spikes of physical activity that can be categorized as VIPA. Therefore, we combined both MIPA and VIPA intensities into the category of MVPA.
TDEE was assumed to be equal to EI (EI = TDEE) when participants were in a weight-stable state and energy balance (pre-intervention), making it an unbiased biomarker of the “true” pre-energy intake. To measure long-term changes in free-living energy intake, we employed the following validated ”intake-balance method”136 (Eq. 12):
Minor energy losses through feces and urine, typically representing approximately 5% and 0.5% of total energy intake, respectively137, were not explicitly modeled. However, because total energy intake was compared within the same individuals before and after the intervention, these losses are expected to remain constant and therefore do not affect the interpretation of within-subject change.
DIT was calculated using the incremental area under the curve (iAUC) for postprandial energy expenditure over a 4-h period. To account for baseline metabolic rate, SMR—measured during the night immediately prior to the DIT assessment—was subtracted from each postprandial energy expenditure time point. This approach was used instead of RMR, which was assessed on a separate day, to ensure temporal consistency.
Results
Recruitment and characteristics of the study population
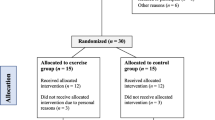
We assessed 859 people for eligibility and willingness to participate in this exercise interventional study. Of these, 60 were found eligible and consented to receiving further information regarding the study and were invited to a 30-min face-to-face or Zoom interview. Eighteen participants commenced the run-in phase; two withdrew before being allocated to the exercise intervention (Fig. 1). All 16 participants who started the exercise intervention completed the study. Of these participants, 50% were female, and all were middle-aged and overweight (BMI = 28.1 ± 2.1 kg/m2, 24.5–31.2 and age = 39 ± 4 years, 32.8–44.3 respectively). Based on medical record evaluation using NCEP ATP III criteria138, 5 out of 16 participants met the diagnostic threshold for metabolic syndrome, indicating a heterogeneous metabolic profile within the study cohort. The trial was conducted between May 2021 and July 2022, and the initial characteristics and changes stratified by gender of the participants are summarized in Table S3. The study design is outlined in Fig. 2.

CONSORT flow diagram Consolidated flow of participant progression through sequential stages of enrollment, allocation, and study completion.

Measurements were conducted prior to intervention (control phase) and within the final two weeks of a 12-week supervised aerobic exercise intervention (intervention phase). A Total daily energy expenditure and body composition were measured using the isotope dilution method (doubly labeled water). B Volumes of highly metabolic organs (brain, liver, and kidney) were assessed using magnetic resonance imaging. C Whole room indirect calorimetry was used to assess: D resting metabolic rate, E diet-induced thermogenesis, F sleeping metabolic rate, and G exercise economy. H Exercise energy expenditure and cardiovascular fitness were monitored using ametabolic cart, I daily physical activity was monitored using a tri-axial accelerometer, and J exercise training was evaluated using an optical heart rate sensor. Written informed consent for publication of identifiable images was obtained from all participants shown. BMI body mass index, kcal kilocalories, wk week.
Physical activity adherence over 12 weeks of the monitored intervention
During the 12-week intervention, participants performed supervised exercise (treadmill walking) prescribed to induce an ExEE of 20 kcal/kg/week (1636 ± 195 kcal/week). This quantitative objective was disseminated to the participants as aggregate weekly temporal commitments, apportioned across two specified PA intensities: 65% and 75% of maximum heart rate (HRmax). The overall adherence to the exercise protocol was 95.4% ± 8.2%. Specifically, participants allocated approximately 37% and 58% of their individual session durations, respectively, to intensity levels pegged at 65% and 75% of HRmax. Participants completed 11.8 ± 0.1 weeks of exercise, consisting of 43.6 ± 8.9 total exercise sessions, corresponding to 3.9 ± 0.7 sessions per week. The prescribed walking activities resulted in 200 ± 30 min per week, equivalent to 27.9 ± 4.9 min daily (Table S4). No exercise-related adverse events or injuries were reported during the study.
Changes in weight, body composition, and cardiovascular fitness
The mean weight loss was −0.2 ± 1.9 kg [not significant (NS), range: −3.1 to +4.3, Fig. 3A]. However, fat mass (FM) significantly decreased (−1.2 ± 2.1 kg, p = 0.033), and fat-free mass (FFM) significantly increased (1.0 ± 1.0 kg, p = 0.001; Fig. 3A). No changes were observed in hip or waist circumference or waist-to-hip ratio (Table S3). An abdominal magnetic resonance imaging (MRI) scan was conducted to further explore changes in FM. These scans revealed no notable changes in total abdominal, deep, or superficial subcutaneous fat. However, visceral fat decreased by ~10% (−12.4 ± 19.4 cm3, p = 0.027) (Fig. 3B). Additionally, maximum oxygen consumption (VO2max) increased by 1.8 ± 2.9 ml/kg/min (p = 0.034) and 0.2 ± 0.3 L/min (p = 0.055), indicating improved cardiovascular fitness. No changes were detected in nocturnal heart rate (HR) or blood pressure (BP) measurements (Table S3). Changes in body composition differed by gender, with reductions in FM observed primarily in women, while changes in FFM were generally comparable between men and women. Other anthropometric measures and cardiovascular fitness did not differ significantly when stratified by sex. (Table S3).

A Individual changes in body weight and body composition determined using DLW method. B Individual changes in abdominal regional fat measured by MRI. C Individual exercise-induced energy compensation. D The association between ExEE and exercise-induced energy compensation. E The association between FM percentage and energy compensation. F Observed versus predicted weight loss using two different models. G Individual observed versus predicted weight loss using two different models. Data are means ± SD. *p < 0.05 and **p < 0.01. Statistical analyses were performed using the Signed Rank Wilcoxon test and Spearman correlation coefficients (r). n = 16, except for B (n = 15). DLW doubly labeled water, Ex EE exercise-induced energy expenditure, FFM fat-free mass, FM fat mass, kcal kilocalories, kg kilograms, MRI magnetic resonance imaging.
Exercise-induced energy compensation
The cumulative estimated ExEE was 18,465 ± 3177 kcal, equivalent to 222 ± 40 kcal per day or 1556 ± 278 kcal per week. However, based on changes in body composition and assuming an energetic cost of 9500 kcal/kg for FM and 1200 kcal/kg for FFM121, the changes in body energy reserves were −10,260 ± 19,206 kcal (−125 ± 223 kcal per day). Thus, there was a notable energy discrepancy (ExEC) of 8205 ± 20,282 kcal, equivalent to 97 ± 235 kcal per day. This compensation accounted for 39 ± 116% of the ExEE in our cohort. Of the 16 participants, 11 exhibited ExEC, while 5 did not, with substantial and expected inter-individual variability observed (Fig. 3C).
Weekly ExEE demonstrated a near-significant correlation with ExEC (r = 0.38, p = 0.14; Fig. 3D). However, after adjusting for pre-intervention body weight, this association was no longer close to statistical significance, suggesting that this relationship may be largely driven by body size rather than a dose-dependent compensatory mechanism. Exercise dose, quantified by the frequency (number of sessions per week) and duration of exercise (time spent exercising per week or session), was not associated with ExEC. However, there was a robust positive correlation between changes in %FM and ExEC (r = 0.91, p < 0.001; Fig. 3E), indicating that greater compensation is associated with less decrease in fat mass. Importantly, no significant correlation was detected between the weekly ExEE and changes in FM (r = 0.41, p = 0.116).
Predicted and observed weight change
We investigated the difference between the measured and predicted weight change using both the conventional static model (7700 kcal/kg) and the dynamic energy balance model (NIH)116,117. These models predicted weight reductions of 2.4 ± 0.4 kg and 4.8 ± 1.7 kg, respectively. However, there was no observable change in mean body weight (−0.2 ± 1.9 kg). Our findings indicate a lower-than-expected weight loss of 89 ± 86% (2.2 ± 2.1 kg) compared to the static model and 96 ± 54% (4.6 ± 2.7 kg) compared to the dynamic (Fig. 3F). Although only eight participants experienced weight loss (50% did not lose or gain weight), all participants exhibited discrepancies in weight change compared to the predicted values, with significant variability (Fig. 3G). Notably, a trend (p = 0.070) toward an association between ExEC and the gap between modeled and measured weight indicates that greater adaptation is associated with less body weight change.
Total daily energy expenditure
At baseline, the absolute TDEE was 2508 ± 428 kcal/day, ranging from 1932 to 3616 kcal/day (Fig. 4A). Following 12 weeks of exercise, TDEE significantly increased by 194 ± 304 kcal/day (p = 0.022). The estimated post-intervention TDEE, calculated from the sum of daily ExEE and the predicted TDEE (Table S5), was 2724 ± 427 kcal, not statistically different from the average measured TDEE of 2702 ± 442 kcal. Thus, there was no MA when the entire cohort was considered (21 ± 274 kcal/day). However, individual residual analyses revealed MA in TDEE in 3 participants (Fig. 4B, Table S6), who showed a decrease of −169 ± 10 kcal/day in TDEE, in contrast to an increase of 227 ± 77 kcal/day in those without adaptation. Additionally, there was no significant correlation between MA in TDEE and weight change or body energy stores. Baseline TDEE was associated with the amount of ExEC (r = 0.61, p = 0.013, Fig. 1S)

A TDEE pre- and post-exercise. B Individual metabolic adaptation in TDEE. C SMR pre- and post-exercise. D Individual metabolic adaptation in SMR. E RMR pre- and post-exercise. F Individual metabolic adaptation in RMR Data are as means ± SD. *p < 0.05. Statistical analyses were performed using the Signed Rank Wilcoxon test. TDEE, SMR (n = 16). RMR measured pre- and post-intervention (n = 15 and n = 14, respectively); F n = 13. Panels B, D, and F display individual-level metabolic adaptation, as proposed by Galgani and Santos (Obesity, 2016), defined by a negative shift in residuals (measured minus predicted energy expenditure) from pre- to post-intervention. Participants exhibiting metabolic adaptation greater than 5% of their baseline predicted energy expenditure are indicated by red lines. RMR resting metabolic rate, SMR sleeping metabolic rate, TDEE total daily energy expenditure.
Sleeping metabolic rate and resting metabolic rate
The average SMR at baseline was 1498 ± 322 kcal per day. However, the absolute post-intervention SMR (1395 ± 214) changed (Fig. 4c). Nevertheless, a difference of −105 ± 174 kcal/day (−6.7 ± 11.7%, p = 0.030) was observed when the average measured SMR was compared with the predicted value of 1499 ± 177 kcal (Table S5), suggesting an effect of MA on SMR. Approximately 60% of participants (10 out of 16) exhibited MA in SMR (Fig. 4D and Table S7), demonstrated by a significant decrease in SMR (–361 ± 65 kcal/day) compared to an increase in those without adaptation (+278 ± 80 kcal/day; p = 0.001).
The analysis of RMR included 15 pre-intervention measurements and 14 measurements post-intervention; due to technical issues, not all subjects were included. The average RMR was 1645 ± 246 kcal/day at baseline (Fig. 4E). The average RMR post-intervention decreased to 1561 ± 227 kcal—a reduction of 74 ± 108 kcal/day (p = 0.040). A comparative analysis of the measured and predicted RMR (Table S5) revealed a notable discrepancy, with an average decrease of 87 ± 118 kcal/day (p = 0.019) following the intervention. This finding indicates a significant MA in RMR of 5.2 ± 7.7%. Of interest, MA in RMR was observed in approximately 70% of participants (9 out of 13; Fig. 4F and Table S6), characterized by a significant difference in RMR change between groups: participants without MA showed an average increase of 43 ± 46 kcal/day, whereas those with MA exhibited a decrease of –150 ± 18 kcal/day. Notably, the daily ExEC of 97 kcal indicates that the MA occurred entirely at the levels of SMR and RMR, which decreased by −105 kcal and −87 kcal, respectively. No significant correlations were observed between ExEC and the magnitude of MA at either the SMR or RMR level. However, baseline SMR was significantly associated with the degree of ExEC (r = 0.51, p = 0.044; Figure S1) and showed a marginal association with baseline RMR (r = 0.44, p = 0.098; Fig. S1)
Volumes of highly metabolically active organs
To explore the physiological mechanism contributing to ExEC, participants underwent a 3-Tesla MRI scan to evaluate changes in volumes of highly metabolically active organs (Fig. 5A). Sixteen participants underwent pre-intervention MRI scans. One participant declined to undergo post-intervention scanning, and another had permanent face makeup that prevented brain imaging. Liver volumes decreased from the pre-intervention measurement by 3.8% (56 ± 94 cm³, p = 0.047; Fig. 5B). There was an average decline in total kidney volume of 5.1% (16 ± 21 cm³, p = 0.017, Fig. 5C). No significant changes were observed in the total brain volume (Fig. 5D).

A Example of kidneys and liver volume quantification using a 3-Tesla MRI scans and home-based MATLAB post-processing image analysis. Volumes of B liver, C total kidney, and D brain pre- and post-intervention. E Liver FM, FFM and liver water content pre- and post-intervention. Association between volumes of F liver, G total kidney, and H brain and TDEE pre-intervention. I Association between changes in liver and kidney volume. Data are means ± SD. *p < 0.01. Statistical analyses were performed using the Signed Rank Wilcoxon test for (B–E) and Spearman correlation coefficients (r) for (G–I). Brain n = 16 and n = 14, liver fat n = 15 and n = 12 pre- and post-intervention, respectively. Liver and kidney volumes n = 15. cm centimetres, FFM fat-free mass, FM fat mass, MRI magnetic resonance imaging, TDEE total daily energy expenditure.
To calculate changes in mass, the specific density of each organ139 was multiplied by the volume. This calculation revealed a loss of 59 ± 99 grams in the liver and 16 ± 22 grams in the kidneys. Assuming metabolic rates of these organs based on published values139, this corresponds to an energy equivalent loss of 19 kcal/day. This energy deficit is approximately 20% of the approximately 100 kcal/day MA observed at the level of SMR and RMR. There were no significant changes from pre- to post intervention in the average liver fat percentage (5.2 ± 5.6% vs. 5.9 ± 8.7%) or in absolute liver fat mass (83 ± 91 vs. 101 ± 153 g). However, there was a significant reduction in the FFM component of the liver (−72 ± 104 g), without corresponding changes in liver water content, resulting in an overall decrease in liver mass of −59 g (Fig. 5E). Comparing individuals with and without MA in TDEE, SMR, and RMR, we found no significant differences in the changes of liver or kidney volume between groups.
A positive correlation was observed between the volume of various organs and TDEE, SMR, and RMR. Specifically, at baseline, there was a strong association between TDEE and brain volume (r = 0.78, p < 0.001), liver volume (r = 0.67, p = 0.007), and total kidney volume (r = 0.66, p = 0.008) (Fig. 5F–H). Similarly, we found associations between pre-intervention RMR and brain volume (r = 0.71, p = 0.003), liver volume (r = 0.55, p = 0.043), and total kidney volume (r = 0.88, p < 0.001). Post-intervention, the correlations persisted with brain volume (r = 0.59, p = 0.026 for TDEE; r = 0.62, p = 0.025 for RMR) and liver volume (r = 0.50, p = 0.058 for TDEE; r = 0.81, p < 0.001 for RMR). However, the correlations with total kidney volume were no longer significant. For SMR, there was considerable heterogeneity. Pre-intervention, there was a significant positive correlation between SMR and brain volume (r = 0.52, p = 0.039), but no correlations were observed between SMR and liver or kidney volumes. Intriguingly, post-intervention analysis revealed substantial positive correlations between SMR and the volumes of the liver (r = 0.63, p = 0.013) and total kidneys (r = 0.51, p = 0.052), although the correlation with brain volume diminished. No significant correlations were observed between changes in organ shrinkage and the magnitude of MA at either the SMR or RMR level. Finally, there was a negative association between the volumetric changes in the liver and kidney (r = −0.63, p = 0.012) (Fig. 5I). Notably, this relationship was not observed with the brain volumetric changes.
Diet-induced thermogenesis and energy intake
We evaluated changes in DIT using a standardized meal of 559 kcal with macronutrient proportions of 54% carbohydrates, 19% protein, and 27% fat (Fig. 6A).

A Energy intake, nutrient intake, and components of standardized breakfast test meal. B DIT pre- and post-intervention. C Energy intake pre- and post-intervention. D PAEE pre- and post-intervention. E PAL pre- and post-intervention. F Time on average spent sleeping, sedentary, LIPA, and MVPA. G Walking economy pre- and post-intervention. Data are means ± SD. *p < 0.05. All statistical analyses were performed using the Signed Rank Wilcoxon test. All panels: n = 16, except DIT (n = 13). Energy-intake data were derived using the intake-balance method. DIT Diet-induced thermogenesis, FFM fat-free mass, g grams, h hour, EI energy intake, kcal kilocalories, LIPA low-intensity physical activity, ml milliliter, MIPA moderate intensity physical activity, MVPA moderate-to-vigorous physical activity, PA physical activity, PAEE physical activity energy expenditure, PAL physical activity level, VO2 volume of oxygen.
DIT, calculated as the incremental area under the curve (iAUC) for energy expenditure over the 4-h postprandial period, was 46 ± 28 kcal pre-intervention and 53 ± 31 kcal post-intervention (Fig. 6B), with no significant difference observed. Notably, three pre-intervention DIT values were excluded due to physiologically implausible negative values (i.e., SMR exceeded postprandial energy expenditure). There was also substantial inter-individual variability in DIT responses, with changes ranging from –106 kcal to +63 kcal, despite all participants consuming the same standardized meal. There was no significant association between the magnitude of MA and changes in DIT.
The participants' energy intake was evaluated pre- and post-intervention over 6.9 ± 0.3 and 6.6 ± 0.7 days, respectively, with a minimum stipulation of 5 days. Self-reported daily energy intake remained similar, averaging 2076 ± 475 kcal/day before the intervention and 1994 ± 408 kcal/day afterward. In contrast, energy intake estimated using the intake-balance method was substantially higher—2508 ± 428 kcal/day pre-intervention and 2577 ± 561 kcal/day post-intervention (Fig. 6C)—with no significant change over time. The difference between self-reported and objectively measured intake was 432 ± 234 kcal/day pre-intervention and 583 ± 514 kcal/day post-intervention (p = 0.001), indicating a consistent underreporting bias. Notably, no significant changes in energy intake were observed from pre- to post-intervention, regardless of the method used.
Physical activity monitoring
Further analysis was conducted to assess the effect of ExEE on changes in objectively measured PA. The average daily pre-intervention PAEE was 631 ± 252 kcal/day, with values ranging from 225 to 1304 kcal/day. When calculated using the individual DIT approach, the mean was 657 ± 300 kcal/day, with a broader range of 32 to 1378 kcal/day. Daily PAEE increased significantly by an average of 225 ± 258 kcal (201 ± 379 kcal/day using the individual DIT approach) following the exercise intervention (Fig. 6D). There was no significant difference between the PAEE values calculated using the standardized 10% DIT assumption and those derived from the individual DIT method. Notably, daily ExEE increased by 215 ± 75 kcal. Thus, there was a significant (p = 0.004) increase in the PA level ratio from 1.55 ± 0.16 to 1.74 ± 0.23 (Fig. 6E). Moreover, there was a negative correlation (r = −0.48, p = 0.085) that did not reach significance between changes in RMR and changes in PAEE (i.e., a decrease in either RMR or PAEE but not both).
During the run-in and pre-intervention phases, participants spent ~8 h per day sedentary (475 ± 64 and 495 ± 71 min/day, respectively); thus, their PA behavior was consistent in the lead-up to the intervention (481 ± 67 min/day), as detailed in Fig. 6F. Baseline accelerometry data indicated participants spent an average of 67% of the day in sleep or sedentary behaviors before the intervention. Low-intensity PA (LIPA) occupied about 27% of each day, and only 6% involved moderate to vigorous PA (MVPA). In other words, the participants in our study spent about 94% of each day with very low PA levels before beginning the study protocol.
In the post-interventional phase, no statistically significant alterations were detected in sleep, sedentary, or LIPA time (Fig. 6F). Moderate-intensity PA (MIPA) and vigorous-intensity PA (VIPA) were combined to yield MVPA. There was an overall increase of 15 ± 23 min daily of MVPA during the intervention period (p = 0.020, Fig. 6F). However, it should be noted that this increase fell short of the expected 28 min of MVPA.
Walking economy assessment
We measured energy expenditure during low- and moderate-intensity paced walking to evaluate changes in walking economy, reflecting typical daily walking in a Western lifestyle. We observed a reduction in relative VO2 during low-intensity walking (from 12.26 ± 2.44 to 10.83 ± 1.40 ml/min/FFM, p = 0.057). A more substantial decrease in relative VO2 was observed during moderate-intensity walking (from 15.84 ± 2.29 to 14.08 ± 1.82 ml/min/FFM, p < 0.001) (Fig. 6G).
Mechanistic hypothesis: structural changes in highly metabolic organs post exercise is associated with alterations in metabolic efficiency via the AMPK pathway
To elucidate the underlying mechanisms by which exercise-induced organ volume reductions result in organ energetics alterations, we established an in vivo exercise model140 similar to the human cohort protocol (Fig. 7A). An example of kidney volume quantification using 7-Tesla MRI scans is shown in Fig. 7B. We found that total kidney volume was significantly reduced by MRI analysis (Fig. 7C), which demonstrates that our animal model recapitulates the organ shrinkage post exercise phenomena we saw in human (Fig. 5C). In parallel with the human data, post-intervention brain volume did not differ between groups (control: 188.7 ± 6.6 mm³ vs. exercise: 192.8 ± 6.5 mm³; p = 0.347). Notably, we were unable to present the liver data since the assessment of liver volume in mice via MRI proved challenging due to the complex geometry of the liver, particularly its lobulated shape, which complicates 3D segmentation.

A Schematic representation of mouse exercise training model and downstream analyses performed. B Example of kidneys volume quantification using a 7-Tesla MRI scans and home-based MATLAB postprocessing image analysis. C Volume of total kidney in control and exercise mice. D Left: Representative hematoxylin and eosin staining of control (top) and exercised (bottom) livers, with a ROI showing the boundaries used to count. Right: Blind quantification of nuclei shown as the average number of nuclei across 10 random fields from each organ, n = 12 Control, n = 8 Exercise mice. E Relative mitochondrial DNA expression of S16 and ND1 (mtDNA) normalized to nuclear DNA (mtDNA/nDNA). F Overlap of cytokines known to be released after exercise (termed exerkines) and predicted upstream regulators generated from IPA cytokine analysis of liver proteomics in control and exercise mice. G Volcano plot of predicted upstream regulators generated from IPA transcription factor analysis of liver proteomic in control and exercise mice, with MTOR (downregulated, blue) and YAP1 (upregulated, red) highlighted. Data are means ± SD. *p < 0.05, **P < 0.01, ****p < 0.0001. Statistical analyses were performed using the Signed Rank Wilcoxon test in C and using unpaired Student’s t-test in (D, E). n = 3 per group in (C, E); n = 4 per group in (F, G). Blind quantification of nuclei per region of interest across 4 random fields from each organ sample (n ≥ 2 per group) in (E). cm centimetres, MRI magnetic resonance imaging, mtDNA mitochondrial DNA, nDNA nuclear DNA.
Next, liver samples taken from exercise and control groups were subjected to proteomics analysis via mass spectrometry141 (Supplementary Data 2), followed by functional relevance by Proteomap analysis. Exercise induces changes in the cytoskeleton, adhesion and metabolic characterizations of the liver (Figs. S2–S4). To further reveal these phenotypic changes, we subjected livers from exercised and control mice to pathology analysis. We found a significant increase in hepatocyte cellular density, by counting nuclei inside defined regions of interest revealing the number of cells per ROI significantly increased after exercise (Fig. 7D). This increase in cellular density supports the proteomic evidence of structural reorganization following exercise (Figs. S2–S4), particularly pathways related to cytoskeletal remodeling and cell-cell interactions.
To understand the metabolic changes, we quantified the mitochondria content in each cell by utilizing two genes (S16 and ND1) that are part of the stable part of mitochondrial DNA (mtDNA) and normalized this to nuclear DNA (nDNA)142. This revealed a significant increase in mtDNA (Fig. 7E), indicating an increase in total mitochondria per cell. Because cells that are densely packed have lower mitochondrial efficiency143, hypothesize that the observed mitochondria are less efficient due to the high density of the tissue.
To further understand the molecular mechanism leading to decrease in mitochondrial efficiency due to higher cell density, we performed predicted upstream regulators analysis using the proteomics data of control and exercise liver141 (Supplementary Data 2). We found that approximately 30% of the upstream cytokines are exerkines (Fig. 7F), known muscle and exercise released cytokines144,145. Further, among the key upstream regulators we found AMP-activated protein kinase (AMPK) (Supplementary Data 3). AMPK is an evolutionarily conserved serine/threonine kinase that plays a pivotal role in regulating cellular energy and structure homeostasis. It is known that under conditions of low cellular energy, such as an elevated AMP/ATP ratio, AMPK is activated to restore energy balance, by promoting catabolic processes that generate ATP while inhibiting energy-consuming anabolic processes145,146.
An additional key target of AMPK activity is the actin cytoskeleton, by either phosphorylation of actin-associated proteins such as cofilin and filamin, which are involved in actin filament remodeling147,148,149,150 or by modulation of Rho family GTPases activity (RhoA, Rac1, Cdc42)151,152,153, which are central regulators of actin filament organization, membrane protrusions, and cell contractility154,155,156. Further, It is known that exercise-induced AMPK activation occurs in various tissues, including the liver157. This response is mediated by the cytokine IL-6, which directly activates AMPK158. Further, exercise-induced activation of AMPK is significantly impaired in IL-6 knockout mice158.
Taking together what is known in the literature and our own observations, we represent a mechanistic hypothesis that exercise induced IL-6 increases AMPK activity leading to changes in both the structure of the cell and metabolic state of the mitochondria in the liver. In support of this hypothesis, it is known that AMPK downregulates mTOR159 which is a core activator of mitochondrial function160,161 and that AMPK regulates YAP162 a known regulator of the cytoskeleton163,164. Interestingly, we found a decrease in mTOR expression and an increase in YAP in the upstream regulator analysis (Fig. 7G). Favoring our hypothesis by which the AMPK activity may be a player in how decreased metabolic works parallel to structural changes in the liver after exercise.
Discussion
Metabolic adaptation and weight loss compensation
In this study, we quantitatively assessed physiological and behavioral adaptation in sedentary, overweight participants who underwent a 3-month exercise intervention. Our results revealed that overweight individuals experienced a 40% ExEC attributed to structured aerobic exercise. Recent studies reported comparable results, demonstrating exercise-induced ExEC averaging approximately 45% with a range from 28% to 63%52,120,165,166. From a practical point of view, it is imperative for current exercise guidelines aimed at weight loss to incorporate and adjust for the ExEC effect by appropriately augmenting the recommended volume of ExEE by 40%. Alternatively, personalized exercise volumes may be prescribed to accommodate the variability in ExEC among individuals.
Notably, within our cohort of sixteen participants, all achieved less weight loss than anticipated3. Moreover, a nearly significant positive correlation was observed between ExEC and the discrepancy between expected and actual weight change (limited sample sizes may explain the lack of significance) and a positive correlation with changes in FM, indicating that higher levels of ExEC were associated with smaller changes in weight and FM. These findings are consistent with existing literature that suggests that exercise training without dietary intervention leads to significantly lower weight loss than projected based on the ExEE3,4,5.
Individuals with higher levels of ExEE initially appeared to exhibit greater ExEC. However, this association did not remain statistically significant after adjusting for baseline body weight. Additionally, dose-dependent relationships were observed between ExEC and baseline TDEE, SMR, and to a lesser extent RMR. These findings suggest that individuals with greater body mass, and consequently higher baseline energy expenditure, may exhibit stronger compensatory responses. This likely reflects a homeostatic regulation mechanism, whereby larger individuals experience greater energy conservation not only as a function of exercise dose but also due to their underlying metabolic demands. Notably, a similar correlation between baseline TDEE and ExEC was recently reported38, lending further support to the presence of dose-related compensation effects.
Furthermore, our prescribed exercise dose was 20 kcal per kilogram, equivalent to 1500 kcal per week (range 1000–2100 kcal), but there is substantial evidence for ExEC even at lower ExEE, such as 700 kcal per week18. Thus, the dose-response relationship between a higher level of ExEE and the enhancement of ExEC remains controversial. There may be a threshold at which increased ExEE no longer results in further exacerbation of ExEC. Exploring recent scientific literature reveals mixed outcomes. Some studies support the idea that higher ExEE correlates with increased ExEC (e.g., comparing 1800 kcal vs. 3600 kcal167 and 700 kcal vs. 1760 kcal18 weekly). In contrast, other research suggests that lower ExEE can lead to equivalent120 (e.g., comparing 1500 kcal vs. 2700 kcal weekly) or even greater ExEC165 (e.g., comparing 1500 kcal vs. 3000 kcal). These conflicting findings highlight the complex and non-linear nature of the ExEE–ExEC relationship and suggest that body size and baseline energy expenditure may confound interpretations unless carefully accounted for. Further research is needed to determine whether a physiological threshold exists beyond which increased ExEE no longer elicits greater ExEC.
Notably, average energy intake, whether self-reported or objectively estimated, did not significantly change from pre- to post-intervention. This supports the interpretation that the less-than-expected weight loss was not primarily driven by increased caloric intake but rather by compensatory adaptations in energy expenditure. However, the absence of measurable exercise-induced energy compensation (ExEC) in five participants suggests that additional factors on the intake side—such as inter-individual variability in energy excretion or retention—may also have contributed in some cases. Taken together, these observations reinforce an expenditure-centered explanation for ExEC while acknowledging that intake-related processes could play a secondary role in certain individuals.
Body composition, body fat pools, and cardiovascular fitness
Despite the absence of weight reduction, participants lost approximately 1 kg of FM and gained the equivalent in FFM. This observation indicates a beneficial change in body composition, favoring lean muscle gain over FM, which is crucial for metabolic health and physical fitness168. Specifically, the exercise intervention led to a reduction of about 10% in visceral fat, which is a more relevant and stronger predictor for obesity-related morbidity and mortality than weight changes169. In this context, it is important to emphasize that anthropometric surrogate markers, including waist circumference, hip circumference, or a combination of both, did not accurately reflect the favorable effects of chronic exercise on abdominal fat in our study. We observed an increase in VO2max, which is commonly used as a marker of improved cardiorespiratory fitness and is strongly linked to a reduced risk of chronic diseases and premature mortality170,171. This enhancement of VO2max highlights the significance of promoting regular exercise in public health guidelines, even in cases where substantial weight loss is not observed. This underscores the importance of enhanced aerobic fitness and oxygen consumption that results from MVPA, even in the absence of weight loss.
Components of TDEE
During the intervention, TDEE increased an average of 220 kcal per day. The absence of a difference between the measured post-intervention TDEE and the predicted TDEE suggests that MA did not occur. These findings cast doubt on the constrained model, which posits that the body undergoes MA in response to increased PA, leading to a less significant increase in TDEE than initially projected. Thus, while the exercise dose of 20 kcal/kg/week is considered moderate-to-high in intervention studies, it is uncertain whether this load was sufficient to reach the threshold at which TDEE becomes constrained—particularly in sedentary individuals. According to the constrained energy model, individuals with low initial activity levels may still operate within the linear range of the PA–TDEE relationship. Therefore, the degree to which our findings reflect constrained energy expenditure remains speculative and may vary across individuals. Nevertheless, it was evident that approximately 50% (~100 kcal) of the daily ExEE (~220 kcal) was compensated for, providing strong evidence for the presence of physiological and/or behavioral adaptations.
Contrary to the commonly held belief that routine exercise will "boost" your metabolism while at rest, we did not observe significant changes in SMR in alignment with other studies that reported no changes in SMR due to exercise35,36,37,38. These studies utilized whole-room indirect calorimetry, the gold standard measurement. Further investigation is warranted to gain deeper insights into these discrepant findings.
The observed downregulation in basal energy expenditure, both at the level of RMR and SMR, accounts for the estimated MA of approximately 100 kcal per day. These results indicate that MA primarily occurs through reducing the body's basal energy demands as a protective mechanism to preserve weight. Recently, Careau et al. published a comprehensive cross-sectional study utilizing data from the International Atomic Energy Agency DLW database166. Their findings align with those of our interventional study, indicating that a decrease in RMR accounts for the majority of the observed MA. MA at the level of both SMR and RMR affected approximately 63% and 85% respectively of the study participants. Still, there was substantial inter-individual variation, underscoring the potential variability in response to exercise interventions. Furthermore, this MA magnitude of around 100 kcal aligns with the results obtained during caloric restriction (i.e., diet-only induced MA)172, hinting at similar underlying mechanisms of energy balance regardless of whether it is achieved through ExEE or caloric-induced weight loss.
Our findings further refine the constrained TDEE model originally proposed by Pontzer6, which posits that energy compensation occurs predominantly within the PAEE component, while RMR remains stable. In contrast, our results suggest that compensation occurred primarily within the resting components, namely, SMR and RMR, despite an increase in TDEE. This indicates that MA in response to structured exercise may emerge through reductions in intrinsic metabolic cost, not solely through behavioral or energetic changes in PAEE. These findings provide an important extension of the constrained energy framework and may help reconcile variability observed across populations and interventions.
Highly metabolic organ volume changes
To our knowledge, this study is the first to report a reduction in the volume of metabolically active organs following exercise training in humans, which may represent an adaptive response to increased energy demands or other unrelated physiological processes. Considering the substantial metabolic demands associated with exercise, we hypothesize that organs adapt themselves to meet the body's metabolic requirements via their size, similar to the effects of calorie restriction-induced weight loss interventions28,31,34,172. Both during caloric restriction28,31,34,172 and exercise, the body selectively preserves the volume of the brain. Thus, the impact of loss of volume on internal metabolic organs on RMR is unlikely to be the only mechanism underlying compensatory reductions in energy expenditure that led to MA in RMR. Our data supports the notion that organ mass loss is comparable during a negative energy balance, providing valuable insights into the challenges faced during weight loss journeys.
Together the MRI, proteomics, staining results, and RT-qCPR indicates contradicting results in that the total organ volume significantly decreased after chronic exercise, however, the total number of cells and the number of mitochondria per cell increased. Cells that are densely packed show lower mitochondrial efficiency143. Given that we observed reduced organ volume accompanied by higher cell density and increased mitochondrial number, we hypothesize that although there are more mitochondria present per cell, their efficiency may be reduced due to increased tissue density. This could represent one mechanism linking organ shrinkage to metabolic state, alongside other non-mitochondrial processes. This raises the question as to which phenomenon comes first. It may be that first, the tissue increases its mitochondria per cell upon exercise to better compete for its metabolic needs109,141, then to maintain homeostasis at resting, perhaps the organ then shrinks the volume of the tissue (via fat or water loss). However, it could also be two independent processes, one being a response to metabolic pressure (increasing number of mitochondria), and the other is a response to reduced fat and water in the tissue (decreased tissue volume). Both of these should be investigated via a time course analysis of the tissues during the exercise intervention; however, this would be outside the scope of this paper.
While our findings demonstrate that reductions in liver and kidney volume occur alongside exercise-induced decreases in RMR, we acknowledge that the causal direction of this relationship cannot be inferred from our study design. It remains unclear whether organ shrinkage actively contributes to MA by lowering basal energy demands, or whether it is a secondary consequence of broader adaptive processes, such as reduced energetic availability or systemic endocrine changes. Organ remodeling could also represent an energy-conserving byproduct of constrained TDEE, without playing a mechanistic role in the regulation of metabolic rate. Given this uncertainty, our interpretation of organ volume loss as a potential driver of energy compensation should be viewed as hypothesis-generating. Longitudinal or interventional studies that manipulate organ size or assess tissue-specific energetics over time will be necessary to establish causality.
Intriguingly, we observed a notable inverse correlation between changes in volume within the liver and kidneys. This suggests that the body undergoes distinctive adaptations in response to ExEE, leading to a reduction in the size of either the liver or the kidneys. This underscores the highly individualized nature of the body's response to exercise.
Although we initially hypothesized that reductions in organ volume contribute directly to MA, the absence of significant differences in liver or kidney volume changes and MA (based on TDEE, RMR, and SMR) suggests alternative explanations. Organ shrinkage may reflect physiological remodeling unrelated to energy expenditure. For instance, prolonged aerobic training alters hemodynamics and may reduce resting organ perfusion needs, leading to volumetric adaptations without direct energetic consequences173. Chronic exercise also modulates systemic inflammation and immune activity, potentially decreasing tissue-resident immune cells or interstitial edema174. Intra-organ shifts in glycogen or associated water content could reduce organ volume independently of changes in metabolically active cell mass. While liver fat percentage remained unchanged on average, regional reductions in ectopic lipid content could still contribute subtly to reduced organ size175. Additionally, exercise-induced rebalancing of autonomic tone and alterations in circulating anabolic and catabolic hormones (e.g., IGF-1, cortisol, catecholamines) may influence tissue growth dynamics176. Finally, under energetic constraint, a strategic redistribution of resources may favor skeletal muscle and cardiovascular adaptations at the expense of visceral organ mass, a prioritization that may serve functional rather than metabolic efficiency goals8. These possibilities underscore the complexity of exercise-induced organ remodeling and highlight the need for mechanistic studies examining tissue-specific energy regulation beyond gross volume changes.
While reductions in the mass of metabolically active organs were estimated to explain approximately 20% of the resting and sleeping metabolic adaptation, the remaining component likely reflect systemic functional changes rather than structural ones. These may include improved skeletal muscle mitochondrial efficiency, reduced sympathetic activity, and a decline in inflammation, oxidative stress, and immune activation. In parallel, lower circulating levels of thyroid hormone (T3), leptin, and adiponectin—each known to potentially influence resting energy expenditure—may further contribute to the reduction in basal metabolic rate following aerobic training.
Several studies have accurately predicted RMR (r of approximately 0.85) using the combined volume of organs with high metabolic rates (such as the brain, liver, kidneys, and heart) and those of tissues like skeletal muscle and adipose tissue139,177. Our study corroborates these findings, demonstrating a robust positive correlation between brain, liver, and kidney volumes and basal metabolism measured by SMR and RMR. A similar pattern was observed for TDEE, likely due to the strong association between basal metabolism and overall daily metabolism.
Utilizing a two-compartment body composition approach, where body weight is divided into FM and fat-free mass FFM, we treated FFM as a unified component without distinguishing among tissue types. However, our findings reveal a meaningful shift in its internal composition: while total FFM increased by approximately 1 kg, we simultaneously observed reductions in the volume of metabolically active organs such as the liver and kidneys, and possibly others not directly measured. This suggests that the gain in FFM was likely driven by increases in skeletal muscle mass, whereas the reduction in organ volume reflects a selective remodeling within the FFM compartment. Importantly, this shift—toward a lower proportion of high-metabolic-rate tissue—may contribute to the observed reductions in SMR and RMR, offering an additional mechanistic perspective on MA to exercise training.
Free-living physical activity
From an energetics standpoint, it is important to note that PAEE and PA level were not suppressed by exercise, indicating no compensatory changes in PA. By directly quantifying changes in walking economy at low intensities representative of non-exercise PA and moderate levels reflective of planned exercise, a reduction in energy expenditure associated with activity was observed. These observations, previously established in caloric restriction-induced weight loss studies in both monkeys178 and humans133, are supported by our study. Our findings enhance our understanding of the mechanisms underlying ExEC in alignment with the constrained energy expenditure model.
Our results demonstrate that the additional time allocated to exercise corresponds to merely 50% of the anticipated changes in overall daily MVPA. This suggests a compensatory behavioral adjustment characterized by a non-intentional reduction in MVPA duration outside of structured exercise sessions. In other words, the integration of structured exercise is offset by a decreased engagement in MVPA during other daily activities, such as opting to drive instead of walking. From an evolutionary standpoint, this is a logical approach to "trim down" nonessential aspects of daily PA. This behavioral compensation could partially underline the lower-than-expected weight loss observed during exercise interventions.
Accelerometer data showed no significant shifts in either sleep duration, sedentary time, or LIPA upon exercise intervention. Consistent with modifications in voluntarily structured MVPA, there might have been a reduction in spontaneous PA or non-exercise PA throughout the entire 24-h period. This is difficult to detect for several reasons. Firstly, there was substantial variability among participants. Secondly, accelerometers have a limited ability to detect changes in lower levels of PA accurately135. Thirdly, we only assessed non-exercise PA pre- to post-intervention. It is possible that alterations in non-exercise PA might temporarily have been observed during the initial weeks of the intervention, as it is known that feelings of fatigue can lead to a decrease in PA, specifically in the early stage. Lastly, the prescribed activity level of 20 kcal/kg or approximately 30 min per day may not be enough to induce a noticeable behavioral compensatory reaction, a conclusion supported by previous studies16,18,55.
Diet-induced thermogenesis
Conflicting findings have emerged regarding the impact of exercise on DIT44. In our study, we did not observe any evidence of exercise-induced MA at the level of DIT. There was a notable divergence in DIT among participants, although identical meals were consumed. This suggests a significant variance in the capacity to convert surplus caloric intake into energy expenditure, implying differing propensities for weight fluctuation. It would be ideal to assess changes in response to meals varying in composition, timing, and quantity. This rigorous approach would help minimize the possibility of drawing erroneous conclusions.
Strengths and limitations of the study
This investigation had methodological strengths given the application of established criterion measures, whole-room indirect calorimetry and DLW, for determining energy expenditure. Rigorous oversight of the customized exercise programs conferred high adherence to individualized regimens. Additional strengths were the 3-Tesla MRI, considered to be the gold standard for organ sizing and measurement of discrete adipose depots. As previous works suggest different relationships between ExEE levels and the extent of compensatory responses179, we specifically investigated the association between ExEC at doses similar to those recommended for weight loss and weight loss maintenance (around 20 kcal/kg).
The study did have limitations. A priori power analysis targeted recruitment of 15 participants based on a 150 kcal/day SD in TDEE. Although absolute changes in TDEE revealed no MA, this may reflect adaptations remaining below the detection threshold reported in DLW reliability studies. Another limitation worth noting is the lack of a parallel control group, which may limit causal interpretation; although the intervention was conducted under highly controlled conditions, we cannot fully exclude the influence of time-related or behavioral confounders common in free-living settings.
Another potential source of bias relates to participants’ increasing familiarity with the metabolic chamber across repeated measurements, which could reduce psychological stress during post-intervention assessments. However, this effect was likely minimal, as all participants underwent a 30-minute exercise run-in followed by 30 min of quiet sitting before data collection (≈1 h total acclimation). The RMR measurement lasted 60 min, with only the most stable 20 min analyzed, and SMR was assessed after participants had spent about four hours in the chamber before sleep. These standardized conditions likely minimized, though did not completely eliminate, any habituation-related effects; thus, the observed reductions in RMR and SMR are more plausibly attributed to true metabolic adaptations rather than chamber familiarity.
We also acknowledge that the quantification of hepatic lipid content, which was derived from a 20 × 20 × 20 mm MR spectroscopy voxel representing a small fraction of total liver volume. Although this voxel size is standard for MR spectroscopy protocols and provides an optimal signal-to-noise ratio, it captures less than 1% of total liver tissue. Therefore, some degree of spatial heterogeneity in lipid distribution cannot be excluded, and part of the observed liver volume reduction may theoretically reflect lipid loss outside the quantified region. We also acknowledge that, given participants had overweight and metabolic syndrome in some cases, the observed reduction in kidney volume could partly reflect the loss of metabolically inert intra-renal lipid depots rather than true tissue remodeling. Although kidney lipid accumulation is relatively small compared with other organs, its reduction may have contributed to the measured decrease in kidney volume.
An additional methodological limitation concerns is that we inferred organ-level metabolic activity by quantifying mass rather than directly assaying energetics or energy costs. Analyses of our mouse model suggested reduced cellular energetic efficiency coinciding with exercise-induced alterations to hepatocellular packing geometry and mitochondrial numbers. Taking together, these multi-scale regulatory modifications offer biologically plausible mechanisms.
The estimation of ExEE may be considered a limitation. It was not feasible to measure ExEE for every session using indirect calorimetry due to logistical constraints and the labor-intensive nature of the method. However, it is worth mentioning that one weekly session was conducted in a controlled laboratory setting (whole-room indirect calorimetry), and in the remaining sessions, HR was monitored. This approach provides a reasonably reliable assessment of ExEE for the majority of the sessions. Furthermore, this study did not consider ExEE increases in RMR due to the excess post-exercise oxygen consumption (EPOC) effect. Nevertheless, it is worth noting that the EPOC effect typically ranges between 6-15%180, resulting in an additional daily EE of 13–33 kcal per day in our study. Given these values, it is plausible to expect a higher weight loss adaptation due to the cumulative effect of ExEE and the EPOC effect.
Two aspects of PAEE and PA level estimates could potentially impact our conclusions. Firstly, PAEE and PA level are estimation variables obtained by subtracting RMR from TDEE or by using their ratio. This approach introduces uncertainty compared to direct measurements of energy expenditure minute by minute throughout the day, which is impractical in free-living conditions181. The reliance on assumptions and the magnification of variance introduced by each measure contributes to heightened uncertainty. Secondly, in the assessment of PAEE, we assumed a fixed daily value of 10% for DIT, disregarding the inter-individual variability that may exist. Although DIT was measured following a standardized test meal in the whole-room indirect calorimetry, these values were not extrapolated to estimate 24-h DIT, as the controlled test conditions may not accurately reflect real-life meal timing, composition, and thermogenic responses. Therefore, we adopted the conventional approximation of 10% of TDEE for DIT to maintain consistency with prior literature. Nonetheless, we acknowledge that using individualized DIT values could enhance the precision of PAEE estimation. The lack of accurate PAEE underscores the limited evidence in the literature regarding reductions in PAEE during exercise interventions67,182.
Another limitation is the limited reproducibility of DIT, particularly in studies with small sample sizes such as ours. Previous research has shown substantial day-to-day and inter-individual variability in DIT, with coefficients of variation often exceeding 25–30% even under tightly controlled laboratory conditions44,183. This inherent variability reduces the statistical power to detect meaningful changes or draw firm conclusions about DIT-related adaptations in our cohort.
It is also important to acknowledge that this study did not include assessments of endocrine biomarkers such as leptin and triiodothyronine (T3), which are known regulators of energy homeostasis and may contribute to metabolic adaptation. While our primary focus was on physiological and behavioral outcomes, the absence of hormonal data limits the mechanistic interpretation of the observed changes in energy expenditure and organ volume. Future research incorporating endocrine profiling will be valuable in further elucidating the hormonal pathways underlying exercise-induced metabolic adaptations.
In the preclinical study, kidney volume was assessed both in vivo and ex vivo, whereas liver volume was evaluated only in vivo. This limitation, stemming from logistical constraints and tissue prioritization that necessitated a kidney-focused approach, may have limited the translational strength of the findings. Also, a limitation of the mouse experiment is that, despite careful habituation and minimal use of mild electrical stimulation, contextual differences inherent to treadmill exposure between exercised and sedentary mice cannot be fully excluded. However, stimulation was brief, infrequent, and continuously monitored, and thus unlikely to have materially influenced the observed outcomes.
Lastly, our study population was remarkably homogeneous with individuals aged between 33 and 44 years, all overweight (BMI ranging from 25 to 31). This particular characteristic of our sample restricts the generalizability of our findings to broader populations, including both younger and older subjects and individuals belonging to different BMI groups.
Taken together, our data indicated significant compensation in ExEE, culminating in the absence of observable weight reduction. This ExEC implies a complete MA at the level of basal metabolism. Furthermore, the study elucidated physiological mechanisms characterized by a reduction in the volume of highly metabolically active organs. In the context of PA, the participants demonstrated behavioral compensation with a less-than-expected elevation in MVPA throughout the day resulting from structured exercise and physiological compensatory mechanism, evidenced by increased walking economy during MVPA. However, DIT and LIPA energy expenditure did not change following the chronic PA.
In summary, our findings provide new insight into how aerobic exercise training can induce both behavioral and physiological compensations that constrain total daily energy expenditure. These adaptations, manifested as increased locomotor economy and reduced non-exercise energy expenditure, may contribute to the wide inter-individual variability in weight change observed following exercise interventions. While energy intake undoubtedly interacts with these processes, the complex interplay between energy expenditure, intake, and retention remains only partially understood. A deeper understanding of these physiological mechanisms—rather than focusing solely on caloric intake—will be essential for refining strategies aimed at preventing and managing obesity.
Data availability
All source data underlying the graphs presented in Figs. 3–7 are provided as Supplementary Data in Excel format (Supplementary Data 1). For the mouse study, the data are available in Sheinboim D. et al. 141, and the mass spectrometry–based proteomics data reported in this paper have been deposited in the PRIDE repository under accession numbers PXD035630 and PXD035648. In addition, requests for further data can be submitted directly via our laboratory website (https://www.gepnerlab.com/data-research-requests).
References
Oppert, J. M. et al. Exercise training in the management of overweight and obesity in adults: synthesis of the evidence and recommendations from the European Association for the Study of Obesity Physical Activity Working Group. Obes. Rev. 22, 1–12 (2021).
Caruso, L., Zauli, E. & Vaccarezza, M. Physical exercise and appetite regulation: new insights. Biomolecules 13, 1170 (2023).
Dhurandhar, E. J. et al. Predicting adult weight change in the real world: a systematic review and meta-analysis accounting for compensatory changes in energy intake or expenditure. Int. J. Obes. 39, 1181–1187 (2015).
Jansen, S. C. P. et al. A systematic review and meta-analysis of the effects of supervised exercise therapy on modifiable cardiovascular risk factors in intermittent claudication. J. Vasc. Surg. 69, 1293–1308.e2 (2019).
Bellicha, A. et al. Effect of exercise training on weight loss, body composition changes, and weight maintenance in adults with overweight or obesity: an overview of 12 systematic reviews and 149 studies. Obes. Rev. 22, 1–13 (2021).
Pontzer, H. et al. Constrained total energy expenditure and metabolic adaptation to physical activity in adult humans. Curr. Biol. 26, 410–417 (2016).
Pontzer, H. Constrained total energy expenditure and the evolutionary biology of energy balance. Exerc. Sport Sci. Rev. 43, 110–116 (2015).
Pontzer, H. Energy constraint as a novel mechanism linking exercise and health. Physiology 33, 384–393 (2018).
Washburn, R. A. et al. Does increased prescribed exercise alter non-exercise physical activity/energy expenditure in healthy adults? A systematic review. Clin. Obes. 4, 1–20 (2014).
Donnelly, J. E. et al. Effects of a 16-month randomized controlled exercise trial on body weight and composition in young, overweight men and women: the Midwest Exercise Trial. Arch. Intern. Med. 163, 1343–1350 (2003).
King, N. A., Hopkins, M., Caudwell, P., Stubbs, R. J. & Blundell, J. E. Individual variability following 12 weeks of supervised exercise: identification and characterization of compensation for exercise-induced weight loss. Int. J. Obes. 32, 177–184 (2008).
Hopkins, M. et al. The adaptive metabolic response to exercise-induced weight loss influences both energy expenditure and energy intake. Eur. J. Clin. Nutr. 68, 581–586 (2014).
Donnelly, J. E. et al. Aerobic exercise alone results in clinically significant weight loss for men and women: Midwest Exercise Trial 2. Obesity 21, E219–E228 (2013).
Bouchard, C. et al. The response to exercise with constant energy intake in identical twins. Obes. Res. 2, 400–410 (1994).
Riou, M. È. et al. Predictors of energy compensation during exercise interventions: a systematic review. Nutrients 7, 3677–3704.
WILLIS, E. A. et al. Nonexercise energy expenditure and physical activity in the Midwest Exercise Trial 2. Med. Sci. Sports Exerc. 46, 2286–2294 (2014).
Wang, X., Bowyer, K. P., Porter, R. R., Breneman, C. B. & Custer, S. S. Energy expenditure responses to exercise training in older women. Physiol. Rep. 5, e13360 (2017).
Martin, C. K. et al. Effect of different doses of supervised exercise on food intake, metabolism, and non-exercise physical activity: the E-MECHANIC randomized controlled trial. Am. J. Clin. Nutr. 110, 583–592 (2019).
Halsey, L. G. et al. Does eating less or exercising more to reduce energy availability produce distinct metabolic responses? Philos. Trans. R. Soc. Lond. B Biol. Sci. 11, 378 (2023).
MacKenzie-Shalders, K., Kelly, J. T., So, D., Coffey, V. G. & Byrne, N. M. The effect of exercise interventions on resting metabolic rate: a systematic review and meta-analysis. J. Sports Sci. 38, 1635–1649 (2020).
Aragon, A. A. et al. International society of sports nutrition position stand: Diets and body composition. J. Int. Soc. Sports Nutr. 14 (2017).
Dulloo, A. G., Jacquet, J., Solinas, G., Montani, J. P. & Schutz, Y. Body composition phenotypes in pathways to obesity and the metabolic syndrome. Int. J. Obes. 34, S4–S17 (2010).
Nelson, K. M., Weinsier, R. L., Long, C. L. & Schutz, Y. Prediction of resting energy expenditure from fat-free mass and fat mass. Am. J. Clin. Nutr. 56, 848–856 (1992).
Kinney, J. M. & Clintec International Inc. Energy Metabolism: Tissue Determinants and Cellular Corollaries (Raven Press, 1992).
Martin, A., Fox, D., Murphy, C. A., Hofmann, H. & Koehler, K. Tissue losses and metabolic adaptations both contribute to the reduction in resting metabolic rate following weight loss. Int. J. Obes. 46, 1168 (2022).
Bosy-Westphal, A., Kahlhöfer, J., Lagerpusch, M., Skurk, T. & Müller, M. J. Deep body composition phenotyping during weight cycling: relevance to metabolic efficiency and metabolic risk. Obes. Rev. 16, 36–44 (2015).
Gallagher, D. et al. Changes in skeletal muscle and organ size after a weight-loss intervention in overweight and obese type 2 diabetic patients. Am. J. Clin. Nutr. 105, 78–84 (2017).
Bosy-Westphal, A. et al. Contribution of individual organ mass loss to weight loss-associated decline in resting energy expenditure 1-3. Am. J. Clin. Nutr. 90, 993–1001 (2009).
Müller, M. J. & Bosy-Westphal, A. Effect of over- and underfeeding on body composition and related metabolic functions in humans. Curr. Diab Rep. 4, 1221–1227 (2019).
Heshka, S. et al. Resting energy expenditure and organ-tissue body composition 5 years after bariatric surgery. Obes. Surg. 30, 587–594 (2020).
Shen, W. et al. Effect of 2-year caloric restriction on organ and tissue size in nonobese 21- to 50-year-old adults in a randomized clinical trial: the CALERIE study. Am. J. Clin. Nutr. 114, 1295–1303 (2021).
Bosy-Westphal, A. et al. Contribution of individual organ mass loss to weight loss-associated decline in resting energy expenditure. Am. J. Clin. Nutr. 90, 993–1001 (2009).
Pourhassan, M. et al. Impact of body composition during weight change on resting energy expenditure and homeostasis model assessment index in overweight nonsmoking adults. Am. J. Clin. Nutr. 99, 779–791 (2014).
Müller, M. J. et al. Metabolic adaptation to caloric restriction and subsequent refeeding: the Minnesota Starvation Experiment revisited. Am. J. Clin. Nutr. 102, 807–819 (2015).
Broskey, N. T. et al. Effect of aerobic exercise-induced weight loss on the components of daily energy expenditure. Med. Sci. Sports Exerc. 53, 2164–2172 (2021).
Meijer, G. A. et al. The effect of a 5-month endurance-training programme on physical activity: evidence for a sex-difference in the metabolic response to exercise. Eur. J. Appl. Physiol. Occup. Physiol. 62, 11–17 (1991).
Bingham, S. A., Goldberg, G. R., Coward, W. A., Prentice, A. M. & Cummings, J. H. The effect of exercise and improved physical fitness on basal metabolic rate. Br. J. Nutr. 61, 155–173 (1989).
Flanagan, E. W. et al. No evidence for metabolic adaptation during exercise-related energy compensation. iScience 27, 109842 (2024).
Reinhardt, M. et al. A human thrifty phenotype associated with less weight loss during caloric restriction. Diabetes 64, 2859–2867 (2015).
Park, M. Y. et al. Dietary factors and eating behaviors affecting diet-induced thermogenesis in obese individuals: a systematic review. J. Nutr. Sci. Vitaminol. 66, 1–9 (2020).
Nelson, K. M. et al. Effect of weight reduction on resting energy expenditure, substrate utilization, and the thermic effect of food in moderately obese women. Am. J. Clin. Nutr. 55, 924–933 (1992).
Dulloo, A. G., Miles-Chan, J., Schutz, Y. & Montani, J.-P. Targeting lifestyle energy expenditure in the management of obesity and health: from biology to built environment. Obes. Rev. 19, 3–7 (2018).
Isacco, L. & Miles-Chan, J. L. Gender-specific considerations in physical activity, thermogenesis and fat oxidation: implications for obesity management. Obes. Rev. 19, 73–83 (2018).
Granata, G. P. & Brandon, L. J. The thermic effect of food and obesity: discrepant results and methodological variations. Nutr. Rev. 60, 223–233 (2002).
Jones, P. P., Van Pelt, R. E., Johnson, D. G. & Seals, D. R. Role of sympathetic neural activation in age- and habitual exercise-related differences in the thermic effect of food. J. Clin. Endocrinol. Metab. 89, 5138–5144 (2004).
Poehlman, E. T., Melby, C. L. & Badylak, S. F. Relation of age and physical exercise status on metabolic rate in younger and older healthy men. J. Gerontol. 46, B54–B58 (1991).
De Jonge, L. The thermic effect of food and obesity: a critical review. Obes. Res. 5, 622–631 (1997).
Morio, B. et al. Effects of 14 weeks of progressive endurance training on energy expenditure in elderly people. Br. J. Nutr. 80, 511–519 (1998).
Melanson, E. L., Keadle, S. K., Donnelly, J. E., Braun, B. & King, N. A. Resistance to exercise-induced weight loss: compensatory behavioral adaptations. Med. Sci. Sports Exerc. 45, 1600–1609 (2013).
Goran, M. I. & Poehlman, E. T. Endurance training does not enhance total energy expenditure in healthy elderly persons. Am. J. Physiol. Endocrinol. Metab. 263, E950–E957 (1992).
Herrmann, S. D. et al. Energy intake, nonexercise physical activity, and weight loss in responders and nonresponders: the Midwest Exercise Trial 2. Obesity 23, 1539–1549 (2015).
Riou, M. -È. et al. Energy compensation following a supervised exercise intervention in women living with overweight/obesity is accompanied by an early and sustained decrease in non-structured physical activity. Front. Physiol. 10, 1–12 (2019).
Myers, C. A. et al. Examination of mechanisms (E-MECHANIC) of exercise-induced weight compensation: study protocol for a randomized controlled trial. Trials 15, 212 (2014).
Fedewa, M. V., Hathaway, E. D., Williams, T. D. & Schmidt, M. D. Effect of exercise training on non-exercise physical activity: a systematic review and meta-analysis of randomized controlled trials. Sports Med. 47, 1171–1182 (2017).
Schutz, Y., Nguyen, D. M. T., Byrne, N. M. & Hills, A. P. Effectiveness of three different walking prescription durations on total physical activity in normal- and overweight women. Obes. Facts 7, 264–273 (2014).
Melanson, E. L. The effect of exercise on non-exercise physical activity and sedentary behavior in adults. Obes. Rev. 18, 40–49 (2017).
Mansfeldt, J. M. & Magkos, F. Compensatory responses to exercise training as barriers to weight loss: changes in energy intake and non-exercise physical activity. Curr. Nutr. Rep. 12, 327–337 (2023).
Colley, R. C., Hills, A. P., King, N. A. & Byrne, N. M. Exercise-induced energy expenditure: implications for exercise prescription and obesity. Patient Educ. Couns. 79, 327–332 (2010).
Keytel, L. R., Lambert, M. I., Johnson, J., Noakes, T. D. & Lambert, E. V. Free living energy expenditure in post menopausal women before and after exercise training. Int. J. Sport Nutr. Exerc. Metab. 11, 226–237 (2001).
McLaughlin, R., Malkova, D. & Nimmo, M. A. Spontaneous activity responses to exercise in males and females. Eur. J. Clin. Nutr. 60, 1055–1061 (2006).
Van Etten, L. M. L. A., Westerterp, K. R., Verstappen, F. T. J., Boon, B. J. B. & Saris, W. H. M. Effect of an 18-wk weight-training program on energy expenditure and physical activity. J. Appl, Physiol. 82, 298–304 (1997).
Paravidino, V. B., Mediano, M. F. F., Hoffman, D. J. & Sichieri, R. Effect of exercise intensity on spontaneous physical activity energy expenditure in overweight boys: a crossover study. PLoS ONE 11, 1–12 (2016).
Wang, X. & Nicklas, B. J. Acute impact of moderate-intensity and vigorous-intensity exercise bouts on daily physical activity energy expenditure in postmenopausal women. J. Obes. 2011, 342431 (2011).
Kozey-Keadle, S. et al. Changes in sedentary time and physical activity in response to an exercise training and/or lifestyle intervention. J. Phys. Act. Health 11, 1324–1333 (2014).
Hand, G. A. et al. The effect of exercise training on total daily energy expenditure and body composition in weight-stable adults: a randomized, controlled trial. J. Phys. Act. Health 17, 456–463 (2020).
Rosenbaum, M. et al. Effects of experimental weight perturbation on skeletal muscle work efficiency in human subjects. Am. J. Physiol. Regul. Integr. Comp. Physiol. 285, R183–R192 (2003).
Silva, A. M. et al. What is the effect of diet and/or exercise interventions on behavioural compensation in non-exercise physical activity and related energy expenditure of free-living adults? A systematic review. Br. J. Nutr. 119, 1327–1345 (2018).
Garber, C. E. et al. Quantity and quality of exercise for developing and maintaining cardiorespiratory, musculoskeletal, and neuromotor fitness in apparently healthy adults: guidance for prescribing exercise. Med. Sci. Sports Exerc. 43, 1334–1359 (2011).
PAGAC Physical Activity Guidelines Advisory Committee Report. Wash. DC US 67, 683 (2008).
DONNELLY, J. E. et al. Appropriate physical activity intervention strategies for weight loss and prevention of weight regain for adults. Med. Sci. Sports Exerc. 41, 459–471 (2009).
LS Pescatello, R Arena, D Riebe, P. T. ACSM’s Guidelines for Exercise Testing and Prescription (Wolters Kluwer/Lippincott Williams & Wilkins Health, 2014).
Pontzer, H. The crown joules: energetics, ecology, and evolution in humans and other primates. Evol. Anthropol. 26, 12–24 (2017).
Hiilloskorpi, H. K., Pasanen, M. E., Fogelholm, M. G., Laukkanen, R. M. & Mänttäri, A. T. Use of heart rate to predict energy expenditure from low to high activity levels. Int. J. Sports Med. 24, 332–336 (2003).
Lam, Y. Y. & Ravussin, E. Analysis of energy metabolism in humans: a review of methodologies. Mol. Metabol. 5, 1057–1071 (2016).
Warren, J. M. et al. Assessment of physical activity - a review of methodologies with reference to epidemiological research. Eur. J. Cardiovasc. Prev. Rehabil. 17, 127–139 (2010).
Hermand, E., Cassirame, J., Ennequin, G. & Hue, O. Validation of a photoplethysmographic heart rate monitor: Polar OH1. Int. J. Sports Med. 40, 462–467 (2019).
Hettiarachchi, I. T., Hanoun, S., Nahavandi, D. & Nahavandi, S. Validation of Polar OH1 optical heart rate sensor for moderate and high intensity physical activities. PLoS ONE 14, e0217288 (2019).
Howe, J. C., Rumpler, W. V. & Seale, J. L. Energy expenditure by indirect calorimetry in premenopausal women: variation within one menstrual cycle. J. Nutr. Biochem. 4, 268–273 (1993).
Solomon, S. J., Kurzer, M. S. & Calloway, D. H. Menstrual cycle and basal metabolic rate in women. Am. J. Clin. Nutr. 36, 611–616 (1982).
Benton Id, M. J., Hutchins, A. M. & Dawes, J. J. Effect of menstrual cycle on resting metabolism: a systematic review and meta-analysis. https://doi.org/10.1371/journal.pone.0236025 (2020).
World Health Organization. Waist Circumference and Waist-Hip Ratio: Report of a WHO Expert Consultation (World Health Organization, 2011).
Seon Yeong Leea & Assessment, D. G. methods in human body composition. Curr. Opin. Clin. Nutr. Metab. Care 11, 566–572 (2008).
Buchowski, M. S. Doubly labeled water is a validated and verified reference standard in nutrition research. J. Nutr. 144, 573–574 (2014).
Donnelly, J. E. & Smith, B. K. Is exercise effective for weight loss with ad libitum diet? Energy balance, compensation, and gender differences. Exerc Sport Sci. Rev. 33, 169–174 (2005).
Westerterp, K. R. Doubly labelled water assessment of energy expenditure: principle, practice, and promise. Eur. J. Appl. Physiol. 117, 1277–1285 (2017).
Levine, J. A. Non-exercise activity thermogenesis (NEAT). Best Pract Res Clin Endocrinol Metab. 16, 679–702 (2002).
Westerterp, K. R. Physical activity and physical activity induced energy expenditure in humans: Measurement, determinants, and effects. Front. Physiol. 4, 90 (2013).
Berman, E. S. F. et al. Direct analysis of δ2H and δ18O in natural and enriched human urine using laser-based, off-axis integrated cavity output spectroscopy. Anal. Chem. 84, 9768–9773 (2012).
Speakman, J. R., Nair, K. S. & Goran, M. I. Revised equations for calculating CO2 production from doubly labeled water in humans. Am. J. Physiol. Endocrinol. Metab. 264, E912–7 (1993).
Schoeller, D. A. et al. Energy expenditure by doubly labeled water: validation in humans and proposed calculation. Am. J. Physiol. 250, R823–R830 (1986).
Weir, J. B. & de, V. New methods for calculating metabolic rate with special reference to protein metabolism. J. Physiol. 109, 1–9 (1949).
Brychta, R. J., Rothney, M. P., Skarulis, M. C. & Chen, K. Y. Optimizing energy expenditure detection in human metabolic chambers. In Proc. 31st Annual International Conference of the IEEE Engineering in Medicine and Biology Society: Engineering the Future of Biomedicine, EMBC 2009 vol. 2009 6864–6868 (IEEE Computer Society, 2009).
Rising, R., Foerster, T., Arad, A. D., Albu, J. & Pi-Sunyer, X. Validation of whole room indirect calorimeters: refinement of current methodologies. Physiol. Rep. 5, 1–13 (2017).
Kayaba, M. et al. Energy metabolism differs between sleep stages and begins to increase prior to awakening. Metabolism 69, 14–23 (2017).
Schoffelen, P. F. M. & Westerterp, K. R. Intra-individual variability and adaptation of overnight- and sleeping metabolic rate. Physiol. Behav. 94, 158–163 (2008).
Goldberg, G. R., Prentice, A. M., Davies, H. L. & Murgatroyd, P. R. Overnight and basal metabolic rates in men and women. Eur. J. Clin. Nutr. 42, 137–144 (1988).
Meijer, G. A., Westerterp, K. R., Saris, W. H. & ten Hoor, F. Sleeping metabolic rate in relation to body composition and the menstrual cycle. Am. J. Clin. Nutr. 55, 637–640 (1992).
Soares, M. J. & Müller, M. J. Resting energy expenditure and body composition: critical aspects for clinical nutrition. Eur. J. Clin. Nutr. 72, 1208–1214 (2018).
Schutz, Y., Ravussin, E., Diethelm, R. & Jequier, E. Spontaneous physical activity measured by radar in obese and control subjects studied in a respiration chamber. Int. J. Obes. 6, 23–28 (1982).
Ho, K. K. Y. Diet-induced thermogenesis: fake friend or foe? J. Endocrinol. 238, R185–R191 (2018).
Fernández-Verdejo, R., Aguirre, C. & Galgani, J. E. Issues in measuring and interpreting energy balance and its contribution to obesity. Curr. Obes. Rep. 8, 88–97 (2019).
Alshurafa, N. et al. Counting bites with bits: expert workshop addressing calorie and macronutrient intake monitoring. J. Med. Internet Res. 21, e14904 (2019).
Goshen, A. et al. Diet quality in relation to healthy ageing: the Israeli Longitudinal Study on Aging (ILSA)-a study protocol. BMJ Open 9, e024673 (2019).
Luciano, F., Ruggiero, L., Minetti, A. E. & Pavei, G. Move less, spend more: the metabolic demands of short walking bouts. Proc. Biol. Sci. 291, 20241220 (2024).
Adeyeri, B., Thomas, S. A. & Arellano, C. J. A simple method reveals minimum time required to quantify steady-rate metabolism and net cost of transport for human walking. J. Exp. Biol. 225, jeb244471 (2022).
Zhou, W. & Xie, Y. Interactive medical image segmentation using snake and multiscale curve editing. Comput. Math Methods Med. 2013, 325903 (2013).
Gallagher, D. et al. Small organs with a high metabolic rate explain lower resting energy expenditure in African American than in white adults. Am. J. Clin. Nutr. 83, 1062–1067 (2006).
Pasanta, D. et al. Magnetic resonance spectroscopy of hepatic fat from fundamental to clinical applications. Diagnostics 11, 1–19 (2021).
Vähä-Ypyä, H., Vasankari, T., Husu, P., Suni, J. & Sievänen, H. A universal, accurate intensity-based classification of different physical activities using raw data of accelerometer. Clin. Physiol. Funct. Imaging 35, 64–70 (2015).
Nachman, D. et al. Twenty-four-hour ambulatory blood pressure measurement using a novel noninvasive, cuffless, wireless device. Am. J. Hypertens. 34, 1171–1180 (2021).
Howley, E. T., Bassett, D. R. & Welch, H. G. Criteria for maximal oxygen uptake: Review and commentary. Med. Sci. Sports Exerc. 27, 1292–1301 (1995).
Priego, T., Sánchez, J., Picó, C. & Palou, A. Sex-differential expression of metabolism-related genes in response to a high-fat diet. Obesity 16, 819–826 (2008).
Krämer, A., Green, J., Pollard, J. & Tugendreich, S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 30, 523–530 (2014).
Otto, A. et al. Systems-wide temporal proteomic profiling in glucose-starved Bacillus subtilis. Nat. Commun. 1, 137 (2010).
Liebermeister, W. et al. Visual account of protein investment in cellular functions. Proc. Natl. Acad. Sci. USA. 111, 8488–8493 (2014).
Thomas, D. M. et al. Can a weight loss of one pound a week be achieved with a 3500-kcal deficit? Commentary on a commonly accepted rule. Int. J. Obes. 37, 1611–1613 (2013).
Thomas, D. M. et al. Why do individuals not lose more weight from an exercise intervention at a defined dose? an energy balance analysis. Obes. Rev. 13, 835–847 (2012).
Kevin, D. Hall. Quantification of the effect of energy imbalance on bodyweight. Lancet 378, 826–837 (2011).
Wolfe, B. M. et al. Resting metabolic rate, total daily energy expenditure, and metabolic adaptation 6 months and 24 months after bariatric surgery. Obesity 26, 862–868 (2018).
Flack, K. D., Hays, H. M., Moreland, J. & Long, D. E. Exercise for weight loss: further evaluating energy compensation with exercise. Med. Sci. Sports Exerc. 52, 2466–2475 (2020).
Hall, K. D. What is the required energy deficit per unit weight loss? Int J. Obes. 32, 573–576 (2008).
Thomas, D. M. et al. Response to ‘Why is the 3500 kcal per pound weight loss rule wrong?’ Int. J. Obes. 37, 1614–1615 (2013).
Galgani, J. E. & Santos, J. L. Insights about weight loss-induced metabolic adaptation. Obesity 24, 277–278 (2016).
Van Etten, L. M., Westerterp, K. R. & Verstappen, F. T. Effect of weight-training on energy expenditure and substrate utilization during sleep. Med. Sci. Sports Exerc. 27, 188–193 (1995).
Chen, K. Y. et al. Room indirect calorimetry operating and reporting standards (RICORS 1.0): a guide to conducting and reporting human whole-room calorimeter studies. Obesity 28, 1613–1625 (2020).
Ravussin, E., Lillioja, S., Anderson, T. E., Christin, L. & Bogardus, C. Determinants of 24-hour energy expenditure in man. Methods and results using a respiratory chamber. J. Clin. Investig. 78, 1568–1578 (1986).
Irving, C. J., Eggett, D. L. & Fullmer, S. Comparing steady state to time interval and non-steady state measurements of resting metabolic rate. Nutr. Clin. Pract. 32, 77–83 (2017).
Usui, C. et al. Validity and reproducibility of a novel method for timecourse evaluation of diet-induced thermogenesis in a respiratory chamber. Physiol. Rep. 3, 1–13 (2015).
Matthews, J. N. S., Altman, D. G., Campbell, M. J. & Royston, P. Analysis of serial measurements in medical research. Br. Med. J. 300, 230–235 (1990).
Redman, L. M. et al. Metabolic and behavioral compensations in response to caloric restriction: Implications for the maintenance of weight loss. PLoS ONE 4, e4377 (2009).
Westerterp, K. R. Diet induced thermogenesis. Nutr. Metab. 1, 5 (2004).
Speakman, J. R. & Westerterp, K. R. Associations between energy demands, physical activity, and body composition in adult humans between 18 and 96 y of age. Am. J. Clin. Nutr. 92, 826–834 (2010).
Goldsmith, R. et al. Effects of experimental weight perturbation on skeletal muscle work efficiency, fuel utilization, and biochemistry in human subjects. Am. J. Physiol. Regul. Integr. Comp. Physiol. 298, R79–88 (2010).
Hibbing, P. R., Lamunion, S. R., Kaplan, A. S. & Crouter, S. E. Estimating energy expenditure with actiGraph GT9X inertial measurement unit. Med. Sci. Sports Exerc. 50, 1093–1102 (2018).
Liu, F., Wanigatunga, A. A. & Schrack, J. A. Assessment of physical activity in adults using wrist accelerometers. Epidemiol. Rev. 43, 65 (2021).
Sanghvi, A., Redman, L. M., Martin, C. K., Ravussin, E. & Hall, K. D. Validation of an inexpensive and accurate mathematical method to measure long-term changes in free-living energy intake. Am. J. Clin. Nutr. 102, 353–358 (2015).
Lund, J., Gerhart-Hines, Z. & Clemmensen, C. Role of energy excretion in human body weight regulation. Trends Endocrinol. Metab. 31, 705–708 (2020).
Grundy, S. M. et al. Diagnosis and management of the metabolic syndrome: an American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 112, 2735–2752 (2005).
Gallagher, D. et al. Organ-tissue mass measurement allows modeling of ree and metabolically active tissue mass. Am. J. Physiol. Endocrinol. Metab. 275, 37–39 (1998).
Fainstein, N. et al. Exercise intensity-dependent immunomodulatory effects on encephalomyelitis. Ann. Clin. Transl. Neurol. 6, 1647–1658 (2019).
Sheinboim, D. et al. An exercise-induced metabolic shield in distant organs blocks cancer progression and metastatic dissemination. Cancer Res. 82, 4164–4178 (2022).
Quiros, P. M., Goyal, A., Jha, P. & Auwerx, J. Analysis of mtDNA/nDNA ratio in mice. Curr. Protoc. Mouse Biol. 7, 47–54 (2017).
Kambach, D. M. et al. Disabled cell density sensing leads to dysregulated cholesterol synthesis in glioblastoma. Oncotarget 8, 14860–14875 (2017).
Chow, L. S. et al. Exerkines in health, resilience and disease. Nat. Rev. Endocrinol. 18, 273–289 (2022).
Docherty, S. et al. The effect of exercise on cytokines: implications for musculoskeletal health: a narrative review. BMC Sports Sci. Med. Rehabil. 14, 1–14 (2022).
Hardie, D. G., Ross, F. A. & Hawley, S. A. AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 13, 251–262 (2012).
Miranda, L. et al. AMP-activated protein kinase induces actin cytoskeleton reorganization in epithelial cells. Biochem. Biophys. Res. Commun. 396, 656–661 (2010).
Crosas-Molist, E. et al. AMPK is a mechano-metabolic sensor linking cell adhesion and mitochondrial dynamics to Myosin-dependent cell migration. Nat. Commun. 14, 2740 (2023).
Schubert, K. M. et al. The AMP-related kinase (AMPK) Induces Ca2 + -independent dilation of resistance arteries by interfering with actin filament formation. Circ. Res. 121, 149–161 (2017).
Blume, C. et al. AMP-activated protein kinase impairs endothelial actin cytoskeleton assembly by phosphorylating vasodilator-stimulated phosphoprotein. J. Biol. Chem. 282, 4601–4612 (2007).
Yan, Y. et al. Augmented AMPK activity inhibits cell migration by phosphorylating the novel substrate Pdlim5. Nat. Commun. 6, 6137 (2015).
Moon, S. et al. Interactome analysis of AMP-activated protein kinase (AMPK)-α1 and -β1 in INS-1 pancreatic beta-cells by affinity purification-mass spectrometry. Sci. Rep. 4, (2014).
You, G. Y. et al. Tiam-1, a GEF for Rac1, plays a critical role in metformin-mediated glucose uptake in C2C12 cells. Cell Signal. 25, 2558–2565 (2013).
Hall, A. Rho GTPases and the actin cytoskeleton. Science 279, 509–514 (1998).
Zaidel-Bar, R., Zhenhuan, G. & Luxenburg, C. The contractome–a systems view of actomyosin contractility in non-muscle cells. J. Cell Sci. 128, 2209–2217 (2015).
Sit, S. T. & Manser, E. Rho GTPases and their role in organizing the actin cytoskeleton. J. Cell Sci. 124, 679–683 (2011).
Park, H. et al. Coordinate regulation of malonyl-CoA decarboxylase, sn-glycerol-3-phosphate acyltransferase, and acetyl-CoA carboxylase by AMP-activated protein kinase in rat tissues in response to exercise. J. Biol. Chem. 277, 32571–32577 (2002).
Kelly, M. et al. AMPK activity is diminished in tissues of IL-6 knockout mice: The effect of exercise. Biochem. Biophys. Res. Commun. 320, 449–454 (2004).
Garza-Lombó, C., Schroder, A., Reyes-Reyes, E. M. & Franco, R. mTOR/AMPK signaling in the brain: cell metabolism, proteostasis and survival. Curr. Opin. Toxicol. 8, 102–110 (2018).
Ramanathan, A. & Schreiber, S. L. Direct control of mitochondrial function by mTOR. Proc. Natl. Acad. Sci. USA. 106, 22229–22232 (2009).
Morita, M. et al. mTOR coordinates protein synthesis, mitochondrial activity and proliferation. Cell Cycle 14, 473–480 (2015).
Mo, J. S. et al. Cellular energy stress induces AMPK-mediated regulation of YAP and the Hippo pathway. Nat. Cell Biol. 17, 500–510 (2015).
Pagliari, S. et al. YAP-TEAD1 control of cytoskeleton dynamics and intracellular tension guides human pluripotent stem cell mesoderm specification. Cell Death Differ. 28, 1193–1207 (2021).
Nardone, G. et al. YAP regulates cell mechanics by controlling focal adhesion assembly. Nat. Commun. 8, 1–13 (2017).
Flack, K. D., Ufholz, K., Johnson, L., Fitzgerald, J. S. & Roemmich, J. N. Energy compensation in response to aerobic exercise training in overweight adults. Am. J. Physiol. Regulat. Integr. Comp. Physiol. 315, R619–R626 (2018).
Careau, V. et al. Energy compensation and adiposity in humans. Curr. Biol. 31, 4659–4666.e2 (2021).
Rosenkilde, M. et al. Body fat loss and compensatory mechanisms in response to different doses of aerobic exercise–a randomized controlled trial in overweight sedentary males. Am. J. Physiol. Regul. Integr. Comp. Physiol. 303, R571–R579 (2012).
Kolnes, K. J., Petersen, M. H., Lien-Iversen, T., Højlund, K. & Jensen, J. Effect of exercise training on fat loss-energetic perspectives and the role of improved adipose tissue function and body fat distribution. Front Physiol. 12, 737709 (2021).
Neeland, I. J. et al. Visceral and ectopic fat, atherosclerosis, and cardiometabolic disease: a position statement. Lancet Diab. Endocrinol. 7, 715–725 (2019).
Wilson, M. G., Ellison, G. M. & Cable, N. T. Basic science behind the cardiovascular benefits of exercise. Br. J. Sports Med. 50, 93–99 (2016).
Aspenes, S. T. et al. Peak oxygen uptake and cardiovascular risk factors in 4631 healthy women and men. Med. Sci. Sports Exerc. 43, 1465–1473 (2011).
Most, J. & Redman, L. M. Impact of calorie restriction on energy metabolism in humans. Exp. Gerontol. 133, 110875 (2020).
Poole, D. C., Copp, S. W., Ferguson, S. K. & Musch, T. I. Skeletal muscle capillary function: contemporary observations and novel hypotheses. Exp. Physiol. 98, 1645–1658 (2013).
Gleeson, M. et al. The anti-inflammatory effects of exercise: mechanisms and implications for the prevention and treatment of disease. Nat. Rev. Immunol. 11, 607–610 (2011).
Hines, C. D. G. et al. T(1) independent, T(2) (*) corrected chemical shift based fat-water separation with multi-peak fat spectral modeling is an accurate and precise measure of hepatic steatosis. J. Magn. Reson Imaging 33, 873–881 (2011).
Hackney, A. C. Stress and the neuroendocrine system: the role of exercise as a stressor and modifier of stress. Expert Rev. Endocrinol. Metab. 1, 783–792 (2006).
Wang, Z. et al. Evaluation of specific metabolic rates of major organs and tissues: Comparison between men and women. Am. J. Hum. Biol. 23, 333–338 (2011).
Yamada, Y. et al. Long-term calorie restriction decreases metabolic cost of movement and prevents decrease of physical activity during aging in rhesus monkeys. Exp. Gerontol. 48, 1226–1235 (2013).
Flack, K. D. et al. Altered motivation states for physical activity and ‘appetite’ for movement as compensatory mechanisms limiting the efficacy of exercise training for weight loss. Front. Psychol. 14, 1098394 (2023).
Laforgia, J., Withers, R. T. & Gore, C. J. Effects of exercise intensity and duration on the excess post-exercise oxygen consumption. J. Sports Sci. 24, 1247–1264 (2006).
Speakman, J. R. & Pontzer, H. Quantifying physical activity energy expenditure based on doubly labelled water and basal metabolism calorimetry: what are we actually measuring? Curr. Opin. Clin. Nutr. Metab. Care 26, 401–408 (2023).
Halsey, L. G. The mystery of energy compensation. Physiol. Biochem Zool. 94, 380–393 (2021).
Ruddick-Collins, L. C., King, N. A., Byrne, N. M. & Wood, R. E. Methodological considerations for meal-induced thermogenesis: measurement duration and reproducibility. Br. J. Nutr. 110, 1978–1986 (2013).
Acknowledgements
The authors thank Scott E. Crouter for analyzing the accelerometer PA data. This study was financially supported by a grant from the Binational Science Foundation (BSF_2019313) and the Israeli Cancer Association (grant number 20250161).
Author information
Authors and Affiliations
Contributions
T.K., Y.G., and E.L.M. were responsible for the conceptualization, study design, regulatory compliance, and overall supervision of the project. They also led the data analysis and were the primary authors of the manuscript. G.D.R. secured informed consent from the participants and provided ongoing medical oversight throughout the study. T.K. and E.Z. managed participant recruitment, data collection, and adherence monitoring. I.M. and D.P. contributed to the development of the measurement protocols and supported data analysis. P.M. and C.L. directed design, regulatory compliance, execution, and data analysis for the mouse experiments. M.R and G.C. conducted the mouse experiment. Chen.L contributed to the tissue structure proteomic analysis. D.A. analyzed the mouse brain imaging data.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Medicine thanks Guillermo Sanchez-Delgado who co-reviewed with Elisa Merchan-Ramirez; Jens Lund and Rodrigo Fernández-Verdejo for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Knaan, T., Ziv-Av, E., Dubnov-Raz, G. et al. Multilevel metabolic adaptation to exercise training. Commun Med 6, 244 (2026). https://doi.org/10.1038/s43856-026-01502-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43856-026-01502-z
