
Abstract
Monoallelic deletion of the short arm of chromosome 17 (del17p) is a recurrent abnormality in cancers with poor outcomes. Best studied in relation to haematological malignancies, associated functional outcomes are attributed mainly to loss and/or dysfunction of TP53, which is located at 17p13.1, but the wider impact of deletion of other genes located on 17p is poorly understood. 17p is one of the most gene-dense regions of the genome and includes tumour suppressor genes additional to TP53, genes essential for cell survival and proliferation, as well as small and long non-coding RNAs. In this review we utilise a data-driven approach to demarcate the extent of 17p deletion in multiple cancers and identify a common loss-of-function gene signature. We discuss how the resultant loss of heterozygosity (LOH) and haploinsufficiency may influence cell behaviour but also identify vulnerabilities that can potentially be exploited therapeutically. Finally, we highlight how emerging animal and isogenic cell line models of del17p can provide critical biological insights for cancer cell behaviour.
Similar content being viewed by others


Introduction
Deletion of the whole or part(s) of the short arm of chromosome 17 (del17p) is a recurrent chromosomal aberration observed in many cancers [1,2,3,4]. del17p is particularly common in solid cancers such as pancreatic ductal adenocarcinomas and colorectal carcinomas where more than 60% of tumours have been reported to contain malignant cells bearing this copy number alteration (CNA) [1, 5,6,7]. Other cancers such as non-small cell lung cancer [8], prostate cancer [9], breast cancer [4] and squamous cell carcinoma of the cervix [10] also harbour this abnormality. Importantly, del17p is also observed in haemic cancers such as chronic lymphocytic leukaemia (CLL) [11], multiple myeloma (MM) [12], diffuse large B cell lymphoma (DLBCL) [13], mantle cell lymphoma (MCL) [14] and acute myeloid leukaemia (AML) [2], where it is associated with rapid progression, short survival and resistance to therapies [15, 16]. From a mechanistic point of view our understanding of the impacts of the loss of multiple genes is limited. The vast majority of studies investigating del17p in cancer have focussed on dysfunction of TP53 which is located at 17p13.1. Whilst the p53 protein encoded by TP53 has an important function in protecting cells from genomic and environmental insults, and in many cases is accompanied by nonsense and missense TP53 mutations that lead to functional loss of this protein [17], studies of CLL [16, 18], and other cancers [1, 8, 9, 12, 19], have shown that del17p is associated with adverse prognosis even in the absence of p53 dysfunction. It is only recently that we have begun to appreciate the wider impact of del17p on cancer cells. The purpose of this review is to assess the impact of del17p on cell behaviour and suggest new horizons for future research and therapies.
Why is a better understanding of such impact important? Firstly, it is clear that the emergence of del17p is more common after DNA-damaging chemotherapy and/or radiotherapy and likely signifies clonal selection and outgrowth [20]. Such clones are invariably and inherently more resistant to further chemotherapy/radiotherapy. Secondly, even in diseases such as CLL, where targeted agents such as ibrutinib have largely replaced chemotherapeutic strategies, outcomes are significantly worse among patients with del17p, mutated TP53 [21, 22] or complex karyotypes [23] associated with del17p [24]. Finally, a better understanding of cancer relevant haploinsufficiencies resulting from loss of heterozygosity (LOH) at 17p may identify as yet uncharacterised targets to inform novel therapies that exploit synthetic lethality.
Anatomy of del17p
Chromosome 17 is highly susceptible to non-allelic homologous recombination due to its unusually complex rearrangement and duplication structure [25]. The presence of low copy number repeats, particularly in the short arm, foster this form of recombination to generate microdeletions of varying sizes. The best characterised are Miller-Dieker syndrome (MDS) and isolated lissencephaly sequence (ILS) which result from variable deletion of a critical 1.3 Mbp region within band 17p13.3 spanning between the essential genes PAFAH1B1 and YWHAE [26], and Smith–Magenis syndrome which in approximately 80% of cases is associated with microdeletion of a 3.7 Mbp region in band 17p11.2 [27]. The genes affected by these deletions lead to defects in craniofacial and brain development, as well as neurological sequelae such as epilepsy. Other microdeletion syndromes include hereditary neuropathy with pressure palsies which results from deletion of a chromosomal section in 17p12 containing PMP22 [28], as well as 17p13.1 and 17p13.2 microdeletion syndromes [29, 30]. These genetic conditions usually arise spontaneously in an autosomal dominant fashion, and range in severity with respect to gene expression and clinical impact. Importantly, microdeletions within 17p13.3 and 17p13.2 tend to be more severe such that affected individuals rarely reach adulthood [26, 29]. In contrast, the impact of microdeletions within 17p13.1, 17p12 and 17p11.2 in affected individuals are milder and do not curtail life expectancy [27, 30]. Importantly, these autosomal microdeletions of 17p do not appear to increase the risk of cancers in affected individuals [30].
In contrast to microdeletions, macro-deletions of 17p within malignant cells span a wider genomic region that varies widely between individual cancers. To understand the nature of this variability we used data available on the DepMap (https://depmap.org/portal/) [31, 32] and cBioPortal (https://www.cbioportal.org/) [17, 33] platforms to investigate the linkage of CNA among genes located on 17p. Figure 1A shows the relationship between copy numbers of TP53 (often used clinically as an indicator of 17p deletion) and of four other genes situated telomeric (POLR2A, PAFAH1B1) or centromeric (MAP2K4, LLGL1) to TP53. The high degree of concomitant loss, particularly of genes in the direct vicinity of TP53, strongly suggests co-deletion with TP53 and that such CNA occurs in a continuous fashion. This notion is illustrated in Fig. 1B where further analysis of this relationship across all cell lines available on the DepMap portal shows that CNAs of genes located telomeric to TP53 are more likely to show such concomitant loss than those that are centromeric. Furthermore, Fig. 1B also shows that the size of deletion is most likely to extend from GEMIN4 at the telomere to MAP2K4 close to the centromere. Application of this algorithm to cell lines derived from specific cancer types showed largely similar sizes of 17p loss with the exception of prostate adenocarcinoma where there was evidence for the occurrence of discreet microdeletions as opposed to the large-scale deletion observed associated with the other cancer cell lines (Fig. 1C). Large-scale deletion of 17p was also evident in AML and DLBCL using data obtained from primary patient samples within the cBioPortal platform (Fig. 1D), an observation made more evident by clustering the samples from individual patients. This showed that del17p is likely to encompass nearly the entire short arm in DLBCL, AML, breast, pancreatic and colorectal cancers (Fig. 1E). Importantly, this analysis also revealed the respective prevalence of del17p in patient samples from each of these tumours, revealing that this aneuploidy is more likely to be observed in solid than in haemic malignancies. With respect to prostate cancers, our findings were in keeping with those using cancer cell lines and del17p was manifested in about half of cases as microdeletions within 17p13.1 (Fig. 1E). Our findings are supported by studies reporting large deletions encompassing most of the entire arm of 17p in CLL [34], DLBCL [35], MM [36], AML [2] and some solid tumours [4,5,6, 9], typically using either SNP arrays or fluorescence in-situ hybridisation techniques such as chromosome painting. Key to the insight provided by the above analysis is that despite the much higher prevalence of del17p within solid tumours, our understanding of del17p and its impact on patient outcomes is mainly derived from studies of haemic malignancies.

A Dot plots showing copy number alteration (CNA) data of the four indicated genes (upper panel shows where they are located on chromosome 17p) plotted relative to TP53 CNA. The CNA data is derived from the cancer cell lines available on DepMap. B Graph showing the frequency of genes located on 17p that are co-deleted with TP53 in a continuous fashion, giving insight into the size of deletion that is most commonly observed in cells bearing del17p. The location of TP53 is indicated by the dotted red line, as are the positions of the genes illustrated in part A. The data used to construct this graph is taken from all cell lines available on DepMap. C Graphs showing the frequency of genes located on 17p that are co-deleted with TP53 in a continuous fashion using data (from DepMap) associated with cell lines derived from the indicated cancers. The location of TP53 is indicated by the dotted red line, as are the positions of the genes illustrated in part A. D Graphs showing the frequency of genes located on 17p that are co-deleted with TP53 in a continuous fashion using data (from cBioPortal) associated with primary cells derived from the indicated cancers. The location of TP53 is indicated by the dotted red line, as are the positions of the genes illustrated in part A. E Hierarchical cluster plots indicating 17p deletion size (x-axis) within cells from individual patients (y-axis) calculated from data recorded in cBioPortal for primary cells from the indicated cancers.
Is del17p the cause or result of genetic instability and/or p53 inactivation?
A large question that looms over the study of del17p is whether it is the cause or consequence of genetic instability and/or p53 inactivation. This is because del17p is often associated with tumours with high chromosomal instability (CIN) and/or with p53 which has become dysfunctional, and because del17p is not always associated with aggressive tumour behaviour; a study of CLL patient outcomes where del17p is observed at diagnosis reported the presence of cases where disease progression was slow [37]. However, slow disease progression in this situation could be the result of association with good prognostic indicators such as mutated IGHV genes and/or to the presence of a low percentage of cells with del17p and/or whether TP53 was mutated as was reported by Yuan et al. [16] in a subsequent study of a larger cohort. This same study reported that presence of mutated TP53 together with del17p correlated with the poorest clinical outcomes, and this brings the question of whether dysfunction of p53 is a driver of 17p loss. Zenz et al. [38] published a detailed mutational profile of TP53 in CLL patient samples with and without del17p, noting that such mutation was present in 76% of cases. Such prevalence is consistent with other tumours such as AML and colorectal and pancreatic cancer (Fig. 2), but other malignancies, such as breast cancer and multiple myeloma, show considerably less, suggesting that association between mutated TP53 and del17p may be cancer dependent. In this respect, TP53 LOH may contribute to p53 inactivation due to haploinsufficiency. This effect has been shown in isogenic cell lines where monoallelic disruption of TP53 expression leads to lower p53 mRNA and protein levels to impact p53-mediated gene expression [39]. Furthermore, one study investigating the relevance of del17p in cell lines derived from MM showed that TP53 haploinsufficiency is associated with reduced p53 activation in response to etoposide or nutlin [40]. Regarding the effect of del17p in primary cancer cells, gene expression profiling revealed under-expression of p53 in CLL cells with a 17p deletion [41], but here it is important to note that p53 mRNA expression may be reduced by mechanisms other than TP53 LOH [17, 33]. Nevertheless, a recent study demonstrated that the introduction of gain-of-function missense mutations into p53, notably R175H and R273H, resulted in increased levels of aneuploidy in recipient cell lines compared to those which received nonsense mutations [42]. Importantly, del17p was not the most common deletion observed within this experimental system. However, studies of fibroblasts derived from patients with Li-Fraumeni syndrome (LFS), an inherited disorder where germ-line mutations in TP53 are of a similar nature to those reported by Redman-Rivera et al. do show LOH associated with the chromosome arm containing the wildtype allele of TP53 as part of observed increased genomic instability [43, 44]. Thus, dysfunctional p53 can drive 17p deletion, but it seems missense mutations are favoured, a notion supported by the cancers we studied for this review where TP53 with missense mutations are significantly represented in tumours with del17p (Fig. 2).

Data from cBioPortal on mutation within TP53 was analysed against the presence or absence of del17p for the indicated cancers (brca breast invasive carcinoma, coad colorectal adenocarcinoma, dlbc diffuse large B cell lymphoma, laml acute myeloid leukaemia, paad pancreatic adenocarcinoma, prad prostate adenocarcinoma) and illustrated according to A count within the total data set available, or as B a percentage within each variable (loss vs. intact 17p). TP53 mutation classification is indicated by the colour scheme.
Is del17p a cause or consequence of genetic instability? Certainly dysfunction of p53 is a driver of such instability and there is reported association with del17p, complex karyotypes, and chromothripsis in CLL [24] and other cancers [45]. A recent study comparing patterns of chromosomal instability in colorectal cancers showed that 17p deletion occurs earlier in cases with high levels of chromosomal complexity compared to cases with lower levels [46]. These findings are in keeping with an earlier report investigating TP53 inactivation and complex karyotype in CLL cells [47]. To this end, several genes on 17p are involved in maintaining genomic integrity. These include RPA1, WRAP53 and CTC1 which also play a direct role in maintaining telomere length, implying that del17p may also influence replicative senescence. In keeping with this, senescence was induced in cells by depletion of MIS12, a key protein encoded at 17p13.2 that is also involved in kinetochore formation [48]. TOP3A, encoded at 17p11.2 and part of the BTR (Bloom syndrome protein (BLM)–TOP3A–RecQ-mediated genome instability proteins (RMI1/2)) dissolvasome complex, is another gene affected by del17p that has a dual role in maintaining genomic integrity and preserving telomere length [49]. In CLL cells there is a reported increase in telomere sister chromatid exchange rates that is correlated with strong down-regulation of TOP3A expression [50]. Telomere dysfunction is a feature of progressive CLL [51] and this can lead to the generation of inter-chromosomal fusions, including with 17p, as reported by Escudero et al. [52]. Here, it is important to note that 17p may be particularly sensitive to the effects of telomere shortening because its telomere is the shortest within the genome [53]. This notion is supported by one report linking further shortening of the 17p telomere to chromosome instability and heteroallelic loss of 17p in Barrett’s oesophagus and development of cancer [54], and by others where short telomeres in general are linked to del17p and increased genetic complexity in CLL and Barrett’s oesophagus [54,55,56,57,58]. Thus, it would seem that telomere shortening is a driver of 17p deletion because of its known role as a driver of genetic instability in cells (reviewed by Maciejowski and de Lange [59]). However, within this context there is also requirement for loss of p53 function in order for cells to tolerate such instability [59], explaining why TP53 mutation is so strongly associated with del17p and high CIN in cancers. Nevertheless, del17p can occur in cancers where p53 is wildtype (Fig. 2), and does feature as a singular aneuploidy in CLL and other tumour types. For this latter reason, this review will focus on the impact of del17p in cells.
Potential cellular and biological consequences of 17p deletion – protein coding genes
Data obtained from OMIM and UCSC Genome Browser indicate that 17p contains 328 protein coding genes as well as 25 miRNAs, 292 lncRNAs, and 153 pseudogenes. Chromosome 17 is second only to chromosome 19 in terms of gene density [25], with bands 17p13.3 and 17p13.2 being rich in gene numbers. Importantly, human chromosome 17 shares high synteny with the distal approximate 60 Mb region of mouse chromosome 11, with cytobands 11B2 – 11B4 on mouse chromosome 11 being representative of human chromosome 17p (Fig. 3). This means that mouse models of del17p can be created if reported embryonic lethality can be overcome by induction of tissue-specific deletion as has been demonstrated in key papers [2, 60].

A chromosome map comparing gene arrangement between mouse chromosome 11B and human chromosome 17. The cytobands 11B2, 11B3 and 11B4 are marked for reference to human 17p.
An important question here is whether haploinsufficiency of genes other than TP53 that are located on 17p is relevant for cancer biology. We previously investigated the gene expression profile of primary CLL cells obtained from patients with or without del17p and identified several genes in addition to TP53 that were under-expressed in cases with del17p including YWHAE, PRPF8, PAFAH1B1, and NUP88 [41]. Using a bioinformatics approach similar to the one reported by Li et al. [4] we examined TGCA data [17, 33] for genes located on 17p that are differentially expressed between samples with or without del17p obtained from patients with different types of cancer. An UpSet plot of this analysis revealed a panel of 76 genes, including the four we identified in CLL, that were consistently under-expressed, suggesting haploinsufficiency, in del17p samples from AML, colorectal, pancreatic, prostate and breast cancers (Fig. 4, Table 1). A significant feature of this panel is the inclusion of eighteen genes designated “common essential” as determined by CRISPR/Cas9 and siRNA screens of cancer cell lines [31, 32]. This designation relates to an absolute requirement of the protein produced by these genes for the survival of affected cells and, together with our observation of reduced expression of these genes in cells with del17p, suggests potential vulnerability. Indeed, haploinsufficiency with respect to expression of these genes has the potential of making cells bearing del17p less fit. This concept is largely unexplored with the exception of a study reporting the development of a tool, known as BISCUT, which was used to show correlation between cellular fitness and aneuploidy [61]. Although not relevant to del17p, this study identified WRN, a gene located on chromosome 8, as a haploinsufficient TSG able to drive tumour growth. This is important in the context of this review because 27 of 1217 recognised tumour suppressor genes (including TP53) are located on 17p at a significantly higher density than on other chromosome arms [62], and our analysis shows that 11 TSGs have lower expression in cells with del17p (Table 1). An additional notion to consider is that cells, including malignant cells, take cues from their environment that regulate their growth, a phenomenon known as cell competition (reviewed in Morata [63]). Here, cells with del17p may be outcompeted by other cancer cells that may be more fit in tumours that are heterogeneous, but this homoeostasis can rapidly change following interventions, such as when therapy is applied, leading to expansion and dominance of cell clones bearing del17p. The concept of cell competition could, therefore, be used to explain why del17p is not always associated with aggressive disease in CLL [37]. The proportion of malignant cells carrying del17p clone also plays a role where reports indicate that when this proportion is high, disease outcome in CLL [16] and MM [64] is poor and vice versa. The notion of increased vulnerability is supported by reports showing that reduced expression of POLR2A in del17p prostate and breast cancer cells is associated with enhanced sensitivity to the fungal toxin α-amanitin [4, 9]. Similarly, LOH and haploinsufficiency have been observed for the splicing factor PRPF8 in AML where reports indicate that reduced expression of this gene lead to increased proliferation of cells bearing del17p as well as increased sensitivity to the drug meayamycin which is an inhibitor of pre-mRNA splicing [65]. Mutation of RPA1 is reported to contribute to the development of lymphoid tumours in mice [66], while reduced expression of this gene is linked to enhanced cell sensitivity to PARP inhibitors such as olaparib [67]. Although not represented within our panel of downregulated genes, LOH and haploinsufficiency of MYBBP1A in pancreatic cancer cells is reported to increase the activity of PARP inhibitors through a mechanism involving the eviction of a minimal pool of chromatin-bound MYBB1A [68].
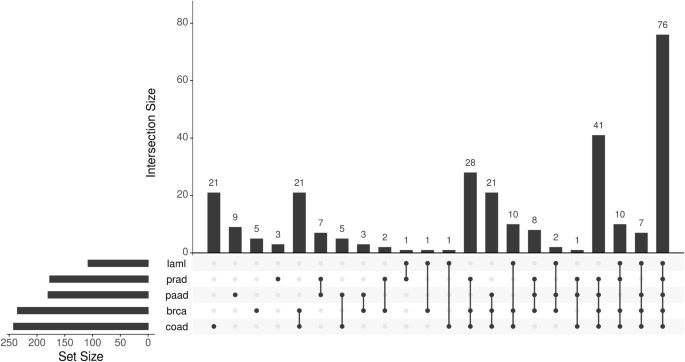
Gene expression in primary malignant cells with or without del17p was compared (with DESeq2), focusing on genes located on 17p that were downregulated in del17p cells. Set size refers to the number of genes showing change in cells from the indicated cancers. Intersection size refers to the shared genes between the different cancers analysed. Cells from 5 cancers were compared; laml acute myeloid leukaemia, prad prostate adenocarcinoma, paad pancreatic adenocarcinoma, brca breast invasive carcinoma, coad colorectal adenocarcinoma. The data for analysis was taken from cBioPortal.
As indicated in the paragraph above, LOH and haploinsufficiency will likely impact Tumour Suppressor Genes (TSGs), potentially helping to explain why cells with del17p can be easily selected and are more resistant to therapy. Of the 11 TSGs identified from the UpSet analysis (Fig. 4, Table 1), several are involved in epigenetic and transcriptional control of gene expression: PHF23 functions as a H3K4me3 reader and deficiency in its expression, such as that associated with del17p, limits the ability of cells to differentiate and promotes their outgrowth as immature malignant cells [69]. SMYD4 is another TSG involved in epigenetic regulatory control, heterozygous loss and downregulation of its expression is reported to facilitate breast cancer development as well as contribute to poor outcomes in patients with other tumour types [70, 71]. ZBTB4 functions as a transcriptional repressor that is reported to regulate expression of the CDK inhibitor P21CIP1 during cellular stress, providing cells with an essential survival signal with which to overcome common front-line therapeutics such as vinscristine [72]. Other TSGs are involved in vesicular trafficking in cells; reduced expression of GABARAP in multiple myeloma cells bearing del17p is associated with inability to express eat-me signal calreticulin protein interactors leading to increased disease resistance to agents which induce immunogenic cell death in this disease [73], and reduced expression of INPP5K and MYO1C have been shown to contribute to tumour development in a rat model of endometrial carcinoma [74]. Finally, although not present within the list of genes affected by LOH and haploinsufficiency (Table 1), one study found that LOH and haploinsufficiency of the TSG ALOX15B contributed to the development of DLBCL in the Eμ-Myc mouse model of this disease [2].
To more fully understand the impact of del17p on cells we performed an enrichment analysis of the affected genes listed in Table 1 using Metascape (https://metascape.org) [75]. Figure 5 shows that the enrichment term “Smith Magenis and Potocki Lupski syndrome copy number variation” is the most significantly represented. Such association can be expected given the majority of genes listed in Table 1 are located within the 17p13.1, 17p12 and 17p11.2 cytobands. However, this result is also surprising because gene density within these cytobands is less than what is observed in 17p13.3 and 17p13.2 [25]. One conclusion from this finding is that genes located in cytobands 17p13.1, 17p12 and 17p11.2 are functionally more likely to be affected by haploinsufficiency in cancers where del17p is a feature and therefore play a significant role in cell behaviour and disease outcome. Of particular interest is the enrichment of genes involved in mitochondrion organisation and in negative regulation of carbohydrate metabolic process. Mitochondrial biogenesis is reported to be increased in CLL cells with del17p [76], potentially in response to increased expression of p21 and Myc [77], and is consistent with the increased cell growth that is required to sustain the shorter lymphocyte doubling times associated with del17p CLL [78]. The rapid cell proliferation associated with del17p can also potentially be explained by haploinsufficiency of MNT which antagonises Myc [79]; downregulation of MNT protein is reported to facilitate MYC-driven transformation in Burkitt’s lymphoma [80] and is associated with adverse prognosis in renal clear cell carcinoma [81]. Further support for del17p resulting in increased cell proliferation is provided by enrichment of genes represented within the cell cycle. For example, reported reduction of PAFAH1B1 and resulting increased malignant cell growth in hepatocellular carcinoma [82] is likely a result of del17p that is commonly observed in this disease [83]. Similarly, reduced expression of the phosphatase CTDNEP1 is reported to facilitate aggressive tumour behaviour in medulloblastoma through amplification of Myc [84]. Finally, increased cell proliferation would be supported by changes to metabolic processes; del17p in CLL cells is associated with increased basal metabolism [85] resulting in increased lactate production and respiration, metabolic alterations observed to closely resemble the switch to aerobic glycolysis that promotes proliferation and survival of CLL cells exposed to B cell receptor (BCR) crosslinking.

The panel of 76 genes identified from the UpSet plot analysis was subjected to enrichment pathway analysis using the Metascape tool (https://metascape.org). Reactome and GO terms are listed according to their probability score.
Cellular and biological consequences of 17p deletion – non-protein coding genes
Chromosome 17p also contains sequences encoding 25 miRNA, 292 long non-coding RNA (lncRNA), and 16 snoRNAs. In keeping with the high gene density observed in this region [25], the majority of these sequences are clustered within 17p13.3 – 17p13.1. 16 of the 25 miRNA encoding sequences are situated within introns of protein-coding genes, six miRNAs fall within enhancer or promoter-like sequences of genes, and three (miR-22, -497, and -195) are coded for within host long non-coding RNAs (Fig. 6A) and likely signify co-regulation and overlapping functions. The predicted targets of the 25 miRNA-encoding sequences on 17p were explored using miRDB (https://mirdb.org/). Targets with a prediction score >90 were visualised using Cytoscape [86] and EnrichmentMap [87], revealing significant enrichment of REACTOME ‘signal transduction’ as well as gene transcription pathways (Fig. 6B).

A Pie chart showing the frequency of microRNA genes located within their own host gene, or within promoter or enhancer regions / intronic gene sequences of other genes. B Cytoscape EnrichmentMap visualisation of DAVID REACTOME pathway enrichment analysis of predicted targets with a target score of >90 for all 17p-derived miRNAs. The sizes of each node represent the number of genes within that particular pathway, while the node colour indicates the False Discovery Rate q significance value. The thickness of each connection indicates the number of genes shared between adjacent nodes (p < 0.01, q < 0.1, overlap threshold = 0.5).
Importantly, miR-22, miR-195, miR-497, and miR-132 are recognised by the Cancer Dependency Map (DepMap, https://depmap.org/portal/) as tumour suppressors. miR-132, together with the other member of the same cluster (miR-212), has been implicated in B cell development and signalling. Specifically, loss of this cluster in mice exhibited reduced B cell apoptosis leading to increased numbers of total B cells in peripheral blood, bone marrow, and spleen [88], whereas enforced expression of miR-132 within the Eμ-Myc mouse-derived haematopoietic stem and progenitor cells limited the development of B-cell cancers [88], a finding that is consistent with other reports demonstrating tumour-suppressive properties of this miRNA [89, 90]. An important target of miR-212 and miR-132 is FOXO3 [91]. Overexpression of this transcription factor is associated with short survival in cancers such as PDAC [92, 93] where del17p is present in a high proportion of cases [68]. FOXO3 is also implicated in the pathogenesis of AML. Specifically, reduction in FOXO3 expression induces differentiation and apoptosis in AML cell lines and limits the development of AML in murine models [94]. Curiously, the gene coding for Activator of RhoGEF and GTPase, ABR, is located very close to the miR-132/212 cluster on 17p13.3 and is relevant because low expression of this gene in cells with del17p is reported to correlate with repressed malignant-cell differentiation and poor outcome in AML [95]. miR-22 is a recognised tumour suppressor in malignancies of the liver, lung and myeloid lineage [96,97,98], affecting processes such as epithelial-mesenchymal transition, cell differentiation and cell cycle (reviewed in [99]). The miR-497/195 cluster was first found reported to have tumour suppressive properties as early as 2011, when Li et al. found that the pair were significantly downregulated in breast cancer samples [100]. More recent studies have found that miR-497/195 is also under-expressed in other cancers [101, 102], low levels being associated with short patient survival. The prevalence of miR-497/195 under-expression is in keeping with the close proximity of the host gene to TP53, meaning that deletion of TP53 would almost always be accompanied by co-deletion of the miR-497/195 cluster. Interestingly, the molecular targets of miR-497/195 (reviewed in [103, 104]) are different to those of miR-22, but the affected cellular processes, regulation of cell differentiation and of cell cycle, cell migration and tumour metastasis, are broadly similar. This suggests potential for a combined effect on these processes resulting from low expression of these miRs, as what might be expected if their host genes are affected by haploinsufficiency in cells with del17p.
Less is known about the lncRNA genes located on 17p. At least one, PRAL (p53 regulation-associated lncRNA), is reported to function as a tumour suppressor by regulating the ability of HSP90 to stabilise p53 protein levels [105,106,107]. Low expression of PRAL is reportedly associated with poor prognosis in non-small cell lung cancer [108], a finding similar to that in a more recent study showing that low PRAL expression in the malignant cells of multiple myeloma, especially those with 17p deletion, correlated with significantly shorter disease-free and overall survival [105]. This same study further showed that PRAL potentiates the anti-multiple myeloma effects of bortezomib by sponging miR-210 and inhibiting its ability to repress expression of bone morphogenetic protein 2 (BMP2). These findings suggest that low levels of PRAL confer bortezomib resistance in multiple myeloma cells bearing a 17p deletion. Consistent with its ability to regulate p53 protein levels, low PRAL expression resulting from del17p has also been reported to confer resistance to DNA-damaging agents in cell lines derived from hepatocellular carcinoma and lung cancer [106]. A second lncRNA linked to p53 stabilisation is SNHG29 (small nucleolar RNA host gene 29) [109]. The role of this lncRNA as a tumour suppressor was recently described by Liu et al. [110] who demonstrated its influence on the genomic binding profile of the transcriptional activator EP300 and correlated low expression with aggressive tumour behaviour in AML. Other potential tumour suppressive effects include stabilising the expression of YAP and facilitating downregulation of programmed death ligand 1 (PD-L1) on tumour cells [111], and in maintaining cell stemness in AML by regulating expression of ETV6 target genes [112]. Importantly, SNHG29 is a host gene for the small nucleolar RNA (snoRNA) genes SNORA49A, SNORA49B and SNORD65, all of which have been linked to the pathogenesis of ALL and AML [113,114,115], likely through their described contribution to tumorigenesis and cellular function by catalysing post-transcriptional modifications of ribosomal, transfer and mRNAs to influence protein production [116].
Role of del17p in tumorigenesis
Does del17p have a role in tumour development? The potential for such a role is great considering the density of TSGs on this chromosome arm [62]. Moreover, 8 proto-oncogenes are also present on 17p [117], but there are no reports of driver mutations within these genes that contribute to tumorigenesis in any cancer. On balance, however, del17p is unlikely to be a driver of tumorigenesis but more of a facilitator of other drivers, either by helping to make these drivers happen or by releasing restriction of their effects. Within this model, deletion of 17p is a discrete process within a single clone that occurs subsequent to its malignant transformation and relatively early in the evolution of the tumour. Evidence for this hypothesis is provided by studies showing the presence of del17p clones in monoclonal B lymphocytosis [118, 119], the precursor condition leading to CLL, in monoclonal gammopathy of undetermined significance (MGUS) and smoldering multiple myeloma (SMM) [120, 121], and of genomic instability in colorectal cancer which place del17p as an early event preceding CIN [46, 122]. Here, as discussed in this review, consideration must also be given to cell-essential gene haploinsufficiency which potentially weakens the ability of cells bearing del17p to effectively compete with other tumour clones. This means that expansion of such cells is accompanied by some sort of selective pressure resulting either from cell intrinsic factors, from therapeutic intervention (expansion of del17p clones is often observed at disease relapse in CLL [123] and MM [124]) and/or from the tumour microenvironment. Cell-intrinsic pressures may be caused by presence of driver mutations such as KRas(G12D) described for colorectal adenoma to carcinoma progression [122], whereas with tumour microenvironment CLL clones that are less able to take advantage of its ability to provide engagement of BCR, which is a key driver of CLL clonal expansion [11], are more likely to manifest as indolent disease [37]. Thus, where cell-intrinsic factors and/or tumour microenvironment is favourable, del17p clones will be selected and make up a greater proportion of the malignant clone in de-novo disease, ultimately contributing to shortened survival and increased disease aggressiveness as is observed associated with CLL and MM [16, 64]. This may be particularly relevant with respect to metastasis; dysregulation of miR-324-5p is reported to enhance the migratory and invasive abilities of MM cells by regulating the function of the SCFβ-TrCP E3 ligase [125], downregulated expression of GAS7 is reported to promote metastasis in neuroblastoma [126], and increased representation of malignant cells with del17p is associated with lymphnode metastases in intrahepatic cholangiocarcinoma [127] and with metastases in other cancers [128]. Figure 7A presents a proposed model of the factors contributing to the expansion of del17p clones, including the influence of therapy resistance as is discussed in the next section.

A Summary of the role del17p plays in tumorigenesis. del17p is an early event where selection processes dictate expansion of the clone which, in turn, adds to therapy resistance and increased aggressive behaviour / metastatic potential. The inset list indicates the cancer types where del17p is associated with aggressive tumour behaviour and poor clinical outcome. CLL chronic lymphocytic leukaemia, MM multiple myeloma, AML acute myeloid leukaemia, DLBCL diffuse large B cell lymphoma, MCL mantle cell lymphoma, ALL acute lymphoblastic leukaemia, HCC hepatocellular carcinoma, CRC colorectal carcinoma. B Summary of the impact of del17p on malignant cell vulnerability and resistance to therapy. Vulnerability / Therapy resistance likely results from LOH and haploinsufficiency of the listed cell-essential genes or TSGs, respectively.
Can del17p confer therapy resistance independently of p53?
In addition to the role of del17p in cancer biology, it is also important to consider how this aneuploidy may influence therapy response. Therapy resistance has been best studied in haemic malignancies, particularly in CLL, where studies employing serial sampling of patients show expansion of clonal populations of malignant cells carrying loss of 17p at disease relapse [20, 129]. These findings have since been expanded to include AML [130], MM [131, 132] and MCL [14]. Importantly, the size of the clone carrying del17p is important for outcome in MM [133] and in CLL [16], where patients who have more cells with this aneuploidy have increased resistance to therapy and a worse clinical outcome (Fig. 7A). TP53 inactivation with loss of cellular processes mediated by wild-type p53 such as apoptosis, cell-cycle arrest and DNA repair, clearly plays an important role in mediating therapy resistance associated with del17p, and this has been reviewed elsewhere [134]. Additionally, as discussed earlier in this review del17p is associated with chromosomal instability in cancers and it is well known that chromosomal instability is a feature of therapy-resistant tumours as has been recently reviewed by Lukow and Sheltzer [135].
This section, therefore, only considers evidence linking del17p with other mechanisms of therapy resistance and is summarised in Fig. 7B. Interestingly, del17p is linked to higher rates of rituximab elimination in CLL, although the underlying mechanism is unclear [136]. In breast cancer, LOH at 17p13.3 was found to correlate with loss of oestrogen and progesterone receptor expression [137], with implications for hormonal therapy. Loss of MKK4, encoded by the tumour suppressor gene MAP2K4 at 17p12, may also contribute to therapy resistance. In addition to potentially driving the selection of malignant clones bearing del17p due to the unusually linear gene-dosage phenotype of MAP2K4 [138], inactivating mutations in these genes have been shown to impair JNK signalling and co-operate with mutant K-RAS to accelerate pancreatic ductal adenocarcinoma [139]. Through its ability to regulate JNK activity, MKK4 promotes Mcl-1 degradation [140], and loss of this effect might therefore explain why del17p is associated with shorter progression-free survival in CLL patients treated with the Bcl-2 inhibitor venetoclax [141]. In colorectal cancer, resistance to chemotherapy and adverse clinical outcome have been linked to low expression of CLDN7, a gene located at 17p13.1 that functions as a TSG in cells where p53 is functionally intact [142]. Finally, reduced expression of ELP5, which is also located at 17p13.1, is reported to confer gemcitabine resistance in gallbladder cancer [143]. ELP5 regulates p53 IRES-dependent translation as part of the Elongator complex – a hexameric RecA-like ATPase that provides tRNA-specific binding sites.
Some TSGs located on chromosome 17p work in tandem with p53, and haploinsufficiency of these genes could therefore potentially allow cancer cells greater latitude to evade the effects of DNA-damaging chemotherapy. The high levels of cellular stress induced by chemotherapy activate p53, initiating downstream functions including cell death to maintain genomic integrity. One mechanism of p53-mediated cell death involves the downregulation of the cystine transporter SLC7A11, resulting in reduced cystine import and an iron-dependent form of cell death known as ferroptosis [144]. Key to the latter process is the formation of lipid peroxides. Notably, five of the six genes responsible for lipid peroxide production (ALOX12, ALOX12B, ALOX15, ALOX15B and ALOX3E) are located on chromosome 17p, four of them (ALOX12, ALOX12B, ALOX15B and ALOX3E) within 1Mbp of TP53. Reduced expression of any of the five ALOX genes on 17p has been shown to increase resistance to ferroptosis in various cell models [145,146,147,148,149], and selective downregulation of ALOX15 has been linked with chemo-resistance in gastric and other cancers [150, 151]. Furthermore, deletions of ALOX12 and ALOX15B have been shown to cooperate with p53 to facilitate tumorigenesis [2, 148, 152]. The observation that phenethyl isothiocyanate, a natural compound that lowers intracellular glutathione, induced the accumulation of reactive oxygen intermediates and preferentially killed CLL cells bearing del17p [153] suggests that it may be possible to activate the ferroptosis death pathway selectively in cancer cells with del17p.
Therapy resistance associated with del17p may also result from the impact of the deletion on tumour-microenvironmental interactions. Notably, loss of 17p in triple negative breast cancer has recently been reported to correlate with reduced tumour infiltration with cytotoxic T-cells and adverse clinical outcome [4]. A similar observation has been made in hepatocellular carcinoma, where del17p was associated with resistance to immune-checkpoint inhibitors [154].
Can LOH of genes on chromosome 17p be exploited therapeutically?
Loss of 17p could create vulnerabilities in cancer cells that are potentially exploitable. Such synthetic lethality could be particularly effective given the survival/growth advantage associated with del17p subclones and their enrichment following chemotherapy. Notably, thirteen genes deemed essential for cell replication/survival located in 17p (Fig. 2B) [31, 32] show lower expression when there is LOH and are potentially targetable with drugs. This review has already discussed how POLR2A LOH leads to lower expression of RNA polymerase II (RNAP2) in del17p prostate and breast cancer cells, and how this renders such cells more sensitive to the cytotoxic effects of the fungal toxin α-amanatin [4, 9]. This vulnerability could be therapeutically exploited through antibody-drug conjugates, which are ideal delivery conduits for compounds such as α-amanitin which are highly toxic. This approach is now being tested in recent reports on the development of an antibody-drug conjugate that combines an anti-B cell maturation antigen (BCMA) antibody with α-amanitin (HDP-101) for the treatment of MM with del17p [155, 156]. HDP-101 is currently undergoing phase I clinical evaluation where early reports indicate that it is well tolerated in the dosage cohorts tested and may have clinical benefit [157]. Another example of synthetic lethality associated with del17p is the enhanced cell sensitivity to PARP inhibitors associated with LOH and haploinsufficiency of PRPF8, RPA1 and MYBBP1A [67, 68]. Although not investigated in the specific context of 17p deletion, a recent report has described the targeting by proteasomal degradation of proline-, glutamic acid- and leucine-rich protein 1 (PELP1), an oestrogen receptor co-activator encoded at 17p13.2, in the treatment of ER+ breast cancer [158]. According to DepMap and cBioPortal, PELP1 LOH is associated with haploinsufficiency in many tumours, raising the possibility that del17p might confer increased sensitivity to the PELP1 degrader, SMIP34. PELP1 also functions as a glucocorticoid receptor co-repressor, potentially explaining the unexpected effectiveness of therapy containing high-dose methylprednisolone in del17p CLL [159, 160]. LOH of the proteasome 20S Subunit Beta 6 (PSMB6), located at 17p13.2, also results in haploinsufficiency, and in myeloma the STAT3 inhibitor, stattic, is reported to inhibit PSMB6 protein function to restore sensitivity to the proteasome inhibitor, bortezomib [161]. Finally, there is evidence to suggest that the metastatic behaviour of tumour cells might be influenced by arsenic trioxide which displaces zinc from CLIP170 resulting in disruption of the PAFAH1B1/NDEL1/dynein microtubule complex [162]. Cancer cells with del17p may be more sensitive to this compound due to low expression of PAFAH1B1 and NDEL1 resulting from LOH/haploinsufficiency of the corresponding genes at 17p13.3 and 17p13.1, respectively.
Models of 17p deletion
Despite the widespread occurrence of 17p deletion across multiple cancer types, study of its impact on tumour cell behaviour is restricted by the limited availability of isogenic cell lines that can be used for direct comparison. The first description of 17p13 deletion modelling was performed using a MICER (Mutagenic Insertion and Chromosome Engineering Resource) strategy to introduce loxP sites on to mouse chromosome 11B3 corresponding to a 4 Mb region that is syntenic to human 17p13.1 [2]. The heterozygous deletion produced through this approach was not lethal to embryonic stem cells but blocked embryonic development likely because of the essential role played by genes within this region during this process. Nevertheless, selective targeting of the 11B3 deletion into the mouse hematopoietic compartment accelerated cancer development in mouse models of B cell lymphoma and AML. These models were further used to elucidate ALOX12 as a gene responsible for facilitating ferroptosis during p53-mediated tumour suppression [148] as well as characterise a role for the H3K4me3 reader PHF23 as a tumour suppressor gene [69]. Hitherto, only two isogenic del17p cell lines have been reported, both of which were generated through Cas9 and sgRNAs targeting of WDR81 (telomere) and MAP2K3 (centromere) to remove the majority of 17p [4, 9]. These cell lines, derived from prostate (DU145) and breast (HS578T) cancer, respectively, have been used in xenograft models to demonstrate the efficacy of targeting haploinsufficient expression of POLR2A with antibody-drug conjugates containing α-amanitin. Interestingly, the DU145 del17p isogenic cell line was used in a CRISPR-based genetic screen which identified an essential role for RING box protein-1 (RBX1) and General transcription factor IIH subunit 1 (GTF2H1) specifically in cells with del17p [9]. This same study showed that RBX1, an E3 ubiquitin ligase, functions to ubiquitinylate RNAP2 to activate its RNA transcription activity and that blocking its expression in cells bearing 17p deletion makes them highly sensitive to the effects of α-amanitin. Together these studies demonstrate the power of an isogenic cell line approach to uncover new therapeutic targets, and call for the creation of further cell lines, representing other cancers, that can be used to investigate the mechanisms responsible for the rapidly progressive, drug resistant phenotype associated with this aneuploidy, as well as to explore novel therapeutic approaches based on synthetic lethality. These studies will likely involve generation of cell lines able to model specific patterns of 17p deletions representative of those found in diseases such as prostate cancer (Fig. 1E). Here, several technologies have been developed to study aneuploidy in cells, with most involving CRISPR/Cas9 to introduce DNA double strand breaks close to the centromere in order to generate chromosome arm deletions (reviewed in Truong, Cané and Lens [163]). With respect to deletion of smaller regions along 17p, caution will be required due to the presence of low copy repeats within chromosome 17 that make re-insertion of removed parts of the chromosome into the unaffected allele via non-allelic homologous recombination a possibility [25].
Conclusions and future directions
There is clearly much to learn from focused research on the wider biological impact of del17p in cancer, over and above p53 dysfunction alone now that the required investigative tools are becoming available. A chief focus so far has been to investigate how del17p contributes to cancer development and whether this can be therapeutically exploited due to synthetic lethality created by haploinsufficiency of genes critical for tumour cell survival. An important caveat here is that the contribution of del17p to disease pathogenesis has only been modelled in DLBCL and AML, and that essential genes are likely to vary with tumour type [31, 32]. Consequently, there is need to develop additional models representing other cancers where this aneuploidy is prevalent, such as pancreatic ductal adenocarcinoma and colorectal carcinoma. This might extend to further interrogation of existing or the development of new mouse models to study the impact of del17p on how this aneuploidy affects the pathogenesis of cancer. Such models have the potential to yield significant insight because of the high degree of synteny between mouse chromosome 11B and human chromosome 17, particularly with respect to 17p (Fig. 3) [25]. As already discussed within this review, seminal work by the Lowe group has used this model to show how del17p accelerates disease progression in DLBCL and AML [2, 69]. Importantly, the deletion created by this model, targeting mouse 11B3 (human 17p13.1) encompasses 31 of the 76 genes that are downregulated in cells with del17p (Table 1), this suggests that further interrogation of this model will yield interesting insight. Study of del17p within a mouse model now needs to be extended to other cancers, an approach that will need to use tissue-specific activation of Cre as has been previously used to create cardiac tissue-specific deletions of mouse chromosome 11 [60] because of the lethality associated with deleting mouse embryonic stem bearing loss of particular regions of chromosome 11. Additional del17p isogenic cell lines need also to be created for comparative studies of how different cancers adapt to selective pressures arising from the tumour microenvironment and exposure to different therapies. The data on deletion size for the cancers presented in Fig. 1 indicates that deletion of the entire short arm is common but not universal, and that tumour-specific models might be required. An interesting hypothesis that could be explored in mouse and isogenic models is that malignant cells harbouring del17p shape their microenvironment in a specific way, a notion made all the more intriguing by a recent review of how driver mutations impact the tumour microenvironment in lung cancer [164]. Finally, if del17p can be detected at single-cell resolution (e.g. using gene signatures associated with del17p) to differentiate between 17p-intact and -deleted cells, it should be possible to investigate how these different cell populations interact with each other within tumours. In summary, del17p is notable for its occurrence across a range of tumour types, its association with aggressive disease behaviour, and its propensity for clonal selection by therapy. Although del17p is strongly associated with TP53 mutation, the association is only partial and it is clear that its deleterious effects result from p53-dependent and/or independent mechanisms. Understanding these mechanisms promises to identify new drug targets, while therapeutic strategies based on synthetic lethality due to LOH/haploinsufficiency of essential genes on 17p set the scene for a Shakespearian fate whereby del17p cancer cells are hoist by their own petard.
References
Murphy SJ, Hart SN, Halling GC, Johnson SH, Smadbeck JB, Drucker T, et al. Integrated genomic analysis of pancreatic ductal adenocarcinomas reveals genomic rearrangement events as significant drivers of disease. Cancer Res. 2016;76:749–61.
Liu Y, Chen C, Xu Z, Scuoppo C, Rillahan CD, Gao J, et al. Deletions linked to TP53 loss drive cancer through p53-independent mechanisms. Nature. 2016;531:471–5.
Ben-David U, Amon A. Context is everything: aneuploidy in cancer. Nat Rev Genet. 2020;21:44–62.
Li Y, Sun Y, Kulke M, Hechler T, Van der Jeught K, Dong T, et al. Targeted immunotherapy for HER2-low breast cancer with 17p loss. Sci Transl Med. 2021;13:eabc6894.
Harada T, Chelala C, Bhakta V, Chaplin T, Caulee K, Baril P, et al. Genome-wide DNA copy number analysis in pancreatic cancer using high-density single nucleotide polymorphism arrays. Oncogene. 2008;27:1951–60.
Bardi G, Sukhikh T, Pandis N, Fenger C, Kronborg O, Heim S. Karyotypic characterization of colorectal adenocarcinomas. Genes Chromosomes Cancer. 1995;12:97–109.
Sayagués JM, Abad Mdel M, Melchor HB, Gutiérrez ML, González-González M, Jensen E, et al. Intratumoural cytogenetic heterogeneity of sporadic colorectal carcinomas suggests several pathways to liver metastasis. J Pathol. 2010;221:308–19.
Chmara M, Wozniak A, Ochman K, Kobierska G, Dziadziuszko R, Sosinska-Mielcarek K, et al. Loss of heterozygosity at chromosomes 3p and 17p in primary non-small cell lung cancer. Anticancer Res. 2004;24:4259–63.
Li Y, Liu Y, Xu H, Jiang G, Van der Jeught K, Fang Y, et al. Heterozygous deletion of chromosome 17p renders prostate cancer vulnerable to inhibition of RNA polymerase II. Nat Commun 2018;9:4394.
Wilting SM, Smeets SJ, Snijders PJ, van Wieringen WN, van de Wiel MA, Meijer GA, et al. Genomic profiling identifies common HPV-associated chromosomal alterations in squamous cell carcinomas of cervix and head and neck. BMC Med Genomics. 2009;2:32.
Kipps TJ, Stevenson FK, Wu CJ, Croce CM, Packham G, Wierda WG, et al. Chronic lymphocytic leukaemia. Nat Rev Dis Primers. 2017;3:17008.
Avet-Loiseau H, Hulin C, Campion L, Rodon P, Marit G, Attal M, et al. Chromosomal abnormalities are major prognostic factors in elderly patients with multiple myeloma: the intergroupe francophone du myélome experience. J Clin Oncol. 2013;31:2806–9.
Chapuy B, Stewart C, Dunford AJ, Kim J, Kamburov A, Redd RA, et al. Molecular subtypes of diffuse large B cell lymphoma are associated with distinct pathogenic mechanisms and outcomes. Nat Med. 2018;24:679–90.
Yi S, Yan Y, Jin M, Bhattacharya S, Wang Y, Wu Y, et al. Genomic and transcriptomic profiling reveals distinct molecular subsets associated with outcomes in mantle cell lymphoma. J Clin Invest. 2022;132:e153283.
Hallek M, Fischer K, Fingerle-Rowson G, Fink AM, Busch R, Mayer J, et al. Addition of rituximab to fludarabine and cyclophosphamide in patients with chronic lymphocytic leukaemia: a randomised, open-label, phase 3 trial. Lancet. 2010;376:1164–74.
Yuan YY, Zhu HY, Wu JZ, Xia Y, Liang JH, Wu W, et al. The percentage of cells with 17p deletion and the size of 17p deletion subclones show prognostic significance in chronic lymphocytic leukemia. Genes Chromosomes Cancer. 2019;58:43–51.
Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, Sumer SO, et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal 2013;6:pl1.
Yu L, Kim HT, Kasar S, Benien P, Du W, Hoang K, et al. Survival of del17p CLL depends on genomic complexity and somatic mutation. Clin Cancer Res. 2017;23:735–45.
Poeta ML, Manola J, Goldwasser MA, Forastiere A, Benoit N, Califano JA, et al. TP53 mutations and survival in squamous-cell carcinoma of the head and neck. N Engl J Med. 2007;357:2552–61.
Zenz T, Kröber A, Scherer K, Häbe S, Bühler A, Benner A, et al. Monoallelic TP53 inactivation is associated with poor prognosis in chronic lymphocytic leukemia: results from a detailed genetic characterization with long-term follow-up. Blood. 2008;112:3322–9.
Brieghel C, Aarup K, Torp MH, Andersen MA, Yde CW, Tian X, et al. Clinical outcomes in patients with multi-Hit TP53 chronic lymphocytic leukemia treated with ibrutinib. Clin Cancer Res. 2021;27:4531–8.
Bomben R, Rossi FM, Vit F, Bittolo T, Zucchetto A, Papotti R, et al. Clinical impact of TP53 disruption in chronic lymphocytic leukemia patients treated with ibrutinib: a campus CLL study. Leukemia. 2023;37:914–8.
Kittai AS, Miller C, Goldstein D, Huang Y, Abruzzo LV, Beckwith K, et al. The impact of increasing karyotypic complexity and evolution on survival in patients with CLL treated with ibrutinib. Blood. 2021;138:2372–82.
Salaverria I, Martín-Garcia D, López C, Clot G, García-Aragonés M, Navarro A, et al. Detection of chromothripsis-like patterns with a custom array platform for chronic lymphocytic leukemia. Genes Chromosomes Cancer. 2015;54:668–80.
Zody MC, Garber M, Adams DJ, Sharpe T, Harrow J, Lupski JR, et al. DNA sequence of human chromosome 17 and analysis of rearrangement in the human lineage. Nature. 2006;440:1045–9.
Chen CP, Chang TY, Guo WY, Wu PC, Wang LK, Chern SR, et al. Chromosome 17p13.3 deletion syndrome: aCGH characterization, prenatal findings and diagnosis, and literature review. Gene. 2013;532:152–9.
Rinaldi B, Villa R, Sironi A, Garavelli L, Finelli P, Bedeschi MF. Smith-Magenis Syndrome-clinical review, biological background and related disorders. Genes. 2022;13.
van Paassen BW, van der Kooi AJ, van Spaendonck-Zwarts KY, Verhamme C, Baas F, de Visser M. PMP22 related neuropathies: Charcot-Marie-Tooth disease type 1A and Hereditary Neuropathy with liability to Pressure Palsies. Orphanet J Rare Dis. 2014;9:38.
Yamamoto N, Okazaki S, Kuki I, Yamada N, Nagase S, Nukui M, et al. Possible critical region associated with late-onset spasms in 17p13.1-p13.2 microdeletion syndrome: a report of two new cases and review of the literature. Epileptic Disord. 2022;24:567–71.
Schluth-Bolard C, Sanlaville D, Labalme A, Till M, Morin I, Touraine R, et al. 17p13.1 microdeletion involving the TP53 gene in a boy presenting with mental retardation but no tumor. Am J Med Genet A. 2010;152a:1278–82.
Hart T, Chandrashekhar M, Aregger M, Steinhart Z, Brown KR, MacLeod G, et al. High-Resolution CRISPR screens reveal fitness genes and genotype-specific cancer liabilities. Cell. 2015;163:1515–26.
McDonald ER 3rd, de Weck A, Schlabach MR, Billy E, Mavrakis KJ, Hoffman GR, et al. Project DRIVE: A compendium of cancer dependencies and synthetic lethal relationships uncovered by large-scale, deep RNAi screening. Cell. 2017;170:577–92.e10.
Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, Aksoy BA, et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012;2:401–4.
Edelmann J, Holzmann K, Miller F, Winkler D, Bühler A, Zenz T, et al. High-resolution genomic profiling of chronic lymphocytic leukemia reveals new recurrent genomic alterations. Blood. 2012;120:4783–94.
Bea S, Zettl A, Wright G, Salaverria I, Jehn P, Moreno V, et al. Diffuse large B-cell lymphoma subgroups have distinct genetic profiles that influence tumor biology and improve gene-expression-based survival prediction. Blood. 2005;106:3183–90.
Boyd KD, Ross FM, Tapper WJ, Chiecchio L, Dagrada G, Konn ZJ, et al. The clinical impact and molecular biology of del(17p) in multiple myeloma treated with conventional or thalidomide-based therapy. Genes Chromosomes Cancer. 2011;50:765–74.
Tam CS, Shanafelt TD, Wierda WG, Abruzzo LV, Van Dyke DL, O’Brien S, et al. De novo deletion 17p13.1 chronic lymphocytic leukemia shows significant clinical heterogeneity: the M. D. Anderson and Mayo Clinic experience. Blood. 2009;114:957–64.
Zenz T, Vollmer D, Trbusek M, Smardova J, Benner A, Soussi T, et al. TP53 mutation profile in chronic lymphocytic leukemia: evidence for a disease specific profile from a comprehensive analysis of 268 mutations. Leukemia. 2010;24:2072–9.
Lynch CJ, Milner J. Loss of one p53 allele results in four-fold reduction of p53 mRNA and protein: a basis for p53 haplo-insufficiency. Oncogene. 2006;25:3463–70.
Teoh PJ, Chung TH, Sebastian S, Choo SN, Yan J, Ng SB, et al. p53 haploinsufficiency and functional abnormalities in multiple myeloma. Leukemia. 2014;28:2066–74.
Lin K, Lane B, Carter A, Johnson GG, Onwuazor O, Oates M, et al. The gene expression signature associated with TP53 mutation/deletion in chronic lymphocytic leukaemia is dominated by the under-expression of TP53 and other genes on chromosome 17p. Br J Haematol. 2013;160:53–62.
Redman-Rivera LN, Shaver TM, Jin H, Marshall CB, Schafer JM, Sheng Q, et al. Acquisition of aneuploidy drives mutant p53-associated gain-of-function phenotypes. Nat Commun. 2021;12:5184.
Burt EC, James LA, Greaves MJ, Birch JM, Boyle JM, Varley JM. Genomic alterations associated with loss of heterozygosity for TP53 in Li-Fraumeni syndrome fibroblasts. Br J Cancer. 2000;83:467–72.
Boyle JM, Mitchell EL, Greaves MJ, Roberts SA, Tricker K, Burt E, et al. Chromosome instability is a predominant trait of fibroblasts from Li-Fraumeni families. Br J Cancer 1998;77:2181–92.
Bernard E, Nannya Y, Hasserjian RP, Devlin SM, Tuechler H, Medina-Martinez JS, et al. Implications of TP53 allelic state for genome stability, clinical presentation and outcomes in myelodysplastic syndromes. Nat Med. 2020;26:1549–56.
Golas MM, Gunawan B, Cakir M, Cameron S, Enders C, Liersch T, et al. Evolutionary patterns of chromosomal instability and mismatch repair deficiency in proximal and distal colorectal cancer. Colorectal Dis. 2022;24:157–76.
Lazarian G, Tausch E, Eclache V, Sebaa A, Bianchi V, Letestu R, et al. TP53 mutations are early events in chronic lymphocytic leukemia disease progression and precede evolution to complex karyotypes. Int J Cancer. 2016;139:1759–63.
Zhang S, Wu Z, Shi Y, Wang S, Ren J, Yu Z, et al. FTO stabilizes MIS12 and counteracts senescence. Protein Cell. 2022;13:954–60.
Temime-Smaali N, Guittat L, Wenner T, Bayart E, Douarre C, Gomez D, et al. Topoisomerase IIIalpha is required for normal proliferation and telomere stability in alternative lengthening of telomeres. EMBO J. 2008;27:1513–24.
Medves S, Auchter M, Chambeau L, Gazzo S, Poncet D, Grangier B, et al. A high rate of telomeric sister chromatid exchange occurs in chronic lymphocytic leukaemia B-cells. Br J Haematol. 2016;174:57–70.
Lin TT, Letsolo BT, Jones RE, Rowson J, Pratt G, Hewamana S, et al. Telomere dysfunction and fusion during the progression of chronic lymphocytic leukemia: evidence for a telomere crisis. Blood. 2010;116:1899–907.
Escudero L, Cleal K, Ashelford K, Fegan C, Pepper C, Liddiard K, et al. Telomere fusions associate with coding sequence and copy number alterations in CLL. Leukemia. 2019;33:2093–7.
Martens UM, Zijlmans JM, Poon SS, Dragowska W, Yui J, Chavez EA, et al. Short telomeres on human chromosome 17p. Nat Genet. 1998;18:76–80.
Finley JC, Reid BJ, Odze RD, Sanchez CA, Galipeau P, Li X, et al. Chromosomal instability in Barrett’s esophagus is related to telomere shortening. Cancer Epidemiol Biomarkers Prev. 2006;15:1451–7.
Dos Santos P, Panero J, Palau Nagore V, Stanganelli C, Bezares RF, Slavutsky I. Telomere shortening associated with increased genomic complexity in chronic lymphocytic leukemia. Tumour Biol. 2015;36:8317–24.
Steinbrecher D, Jebaraj BMC, Schneider C, Edelmann J, Cymbalista F, Leblond V, et al. Telomere length in poor-risk chronic lymphocytic leukemia: associations with disease characteristics and outcome. Leuk Lymphoma. 2018;59:1614–23.
Jebaraj BMC, Tausch E, Landau DA, Bahlo J, Robrecht S, Taylor-Weiner AN, et al. Short telomeres are associated with inferior outcome, genomic complexity, and clonal evolution in chronic lymphocytic leukemia. Leukemia. 2019;33:2183–94.
Hoxha M, Fabris S, Agnelli L, Bollati V, Cutrona G, Matis S, et al. Relevance of telomere/telomerase system impairment in early stage chronic lymphocytic leukemia. Genes Chromosomes Cancer. 2014;53:612–21.
Maciejowski J, de Lange T. Telomeres in cancer: tumour suppression and genome instability. Nat Rev Mol Cell Biol. 2017;18:175–86.
Zheng B, Sage M, Sheppeard EA, Jurecic V, Bradley A. Engineering mouse chromosomes with Cre-loxP: range, efficiency, and somatic applications. Mol Cell Biol. 2000;20:648–55.
Shih J, Sarmashghi S, Zhakula-Kostadinova N, Zhang S, Georgis Y, Hoyt SH, et al. Cancer aneuploidies are shaped primarily by effects on tumour fitness. Nature. 2023;619:793–800.
Zhao M, Sun J, Zhao Z. TSGene: a web resource for tumor suppressor genes. Nucleic Acids Res. 2012;41:D970–D6.
Morata G. Cell competition: a historical perspective. Dev Biol. 2021;476:33–40.
Laurence L, Marion E, Valérie T, Thierry S, Soraya W, Steven R, et al. Mutations in TP53 are exclusively associated with del(17p) in multiple myeloma. Haematologica. 2010;95:1973–6.
Kurtovic-Kozaric A, Przychodzen B, Singh J, Konarska MM, Clemente MJ, Otrock ZK, et al. PRPF8 defects cause missplicing in myeloid malignancies. Leukemia. 2015;29:126–36.
Wang Y, Putnam CD, Kane MF, Zhang W, Edelmann L, Russell R, et al. Mutation in Rpa1 results in defective DNA double-strand break repair, chromosomal instability and cancer in mice. Nat Genet. 2005;37:750–5.
Murai J, Pommier Y. BRCAness, Homologous recombination deficiencies, and synthetic lethality. Cancer Res. 2023;83:1173–4.
Hsieh A, Pitarresi JR, Lerner J, Donahue G, Hsiehchen D, Rustgi AK, et al. Growth of pancreatic cancers with hemizygous chromosomal 17p loss of MYBBP1A can be preferentially targeted by PARP inhibitors. Sci Adv. 2020;6:eabc4517.
Chen M, Chen X, Li S, Pan X, Gong Y, Zheng J, et al. An epigenetic mechanism underlying chromosome 17p deletion-driven tumorigenesis. Cancer Discov. 2021;11:194–207.
Hu L, Zhu YT, Qi C, Zhu YJ. Identification of Smyd4 as a potential tumor suppressor gene involved in breast cancer development. Cancer Res. 2009;69:4067–72.
Olivera Santana BL, de Loyola MB, Gualberto ACM, Pittella-Silva F. Genetic alterations of SMYD4 in solid tumors using integrative multi-platform analysis. Int J Mol Sci. 2024;25:6097.
Weber A, Marquardt J, Elzi D, Forster N, Starke S, Glaum A, et al. Zbtb4 represses transcription of P21CIP1 and controls the cellular response to p53 activation. EMBO J. 2008;27:1563–74.
Gulla A, Morelli E, Johnstone M, Turi M, Samur MK, Botta C, et al. Loss of GABARAP mediates resistance to immunogenic chemotherapy in multiple myeloma. Blood. 2024;143:2612–26.
Hedberg Oldfors C, Dios DG, Linder A, Visuttijai K, Samuelson E, Karlsson S, et al. Analysis of an independent tumor suppressor locus telomeric to Tp53 suggested Inpp5k and Myo1c as novel tumor suppressor gene candidates in this region. BMC Genet. 2015;16:80.
Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. 2019;10:1523.
Ogasawara MA, Liu J, Pelicano H, Hammoudi N, Croce CM, Keating MJ, et al. Alterations of mitochondrial biogenesis in chronic lymphocytic leukemia cells with loss of p53. Mitochondrion. 2016;31:33–9.
Kienle DL, Korz C, Hosch B, Benner A, Mertens D, Habermann A, et al. Evidence for distinct pathomechanisms in genetic subgroups of chronic lymphocytic leukemia revealed by quantitative expression analysis of cell cycle, activation, and apoptosis-associated genes. J Clin Oncol. 2005;23:3780–92.
Baumann T, Moia R, Gaidano G, Delgado J, Condoluci A, Villamor N, et al. Lymphocyte doubling time in chronic lymphocytic leukemia modern era: a real-life study in 848 unselected patients. Leukemia. 2021;35:2325–31.
Liaño-Pons J, Arsenian-Henriksson M, León J. The multiple faces of MNT and its role as a MYC modulator. Cancers. 2021;13:4682.
Niu F, Dzikiewicz-Krawczyk A, Koerts J, de Jong D, Wijenberg L, Fernandez Hernandez M, et al. MiR-378a-3p is critical for Burkitt lymphoma cell growth. Cancers. 2020;12:3546.
Prochownik EV, Wang H. Normal and neoplastic growth suppression by the extended Myc network. Cells. 2022;11:4087.
Xing Z, Tang X, Gao Y, Da L, Song H, Wang S, et al. The human LIS1 is downregulated in hepatocellular carcinoma and plays a tumor suppressor function. Biochem Biophys Res Commun. 2011;409:193–9.
Jin Q, Cheng M, Xia X, Han Y, Zhang J, Cao P, et al. Down-regulation of MYH10 driven by chromosome 17p13.1 deletion promotes hepatocellular carcinoma metastasis through activation of the EGFR pathway. J Cell Mol Med. 2021;25:11142–56.
Luo Z, Xin D, Liao Y, Berry K, Ogurek S, Zhang F, et al. Loss of phosphatase CTDNEP1 potentiates aggressive medulloblastoma by triggering MYC amplification and genomic instability. Nat Commun. 2023;14:762.
Kluckova K, Clear AJ, D’Avola A, Rassenti LZ, Kipps TJ, Gribben JG, et al. B-cell receptor signaling induced metabolic alterations in chronic lymphocytic leukemia can be partially bypassed by TP53 abnormalities. Hemasphere. 2022;6:e722.
Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13:2498–504.
Merico D, Isserlin R, Stueker O, Emili A, Bader GD. Enrichment Map: A Network-Based Method for Gene-Set Enrichment Visualization and Interpretation. PLoS ONE. 2010;5:e13984.
Mehta A, Mann M, Zhao JL, Marinov GK, Majumdar D, Garcia-Flores Y, et al. The microRNA-212/132 cluster regulates B cell development by targeting Sox4. J Exp Med. 2015;212:1679–92.
Zhang S, Hao J, Xie F, Hu X, Liu C, Tong J, et al. Downregulation of miR-132 by promoter methylation contributes to pancreatic cancer development. Carcinogenesis. 2011;32:1183–9.
You J, Li Y, Fang N, Liu B, Zu L, Chang R, et al. MiR-132 suppresses the migration and invasion of lung cancer cells via targeting the EMT regulator ZEB2. PLoS ONE. 2014;9:e91827.
Mehta A, Zhao JL, Sinha N, Marinov GK, Mann M, Kowalczyk MS, et al. The microRNA-132 and microRNA-212 cluster regulates hematopoietic stem cell maintenance and survival with age by buffering FOXO3 expression. Immunity. 2015;42:1021–32.
Tenbaum SP, Ordóñez-Morán P, Puig I, Chicote I, Arqués O, Landolfi S, et al. β-catenin confers resistance to PI3K and AKT inhibitors and subverts FOXO3a to promote metastasis in colon cancer. Nat Med. 2012;18:892–901.
Lees J, Hay J, Moles MW, Michie AM. The discrete roles of individual FOXO transcription factor family members in B-cell malignancies. Front Immunol. 2023;14:1179101.
Sykes SM, Lane SW, Bullinger L, Kalaitzidis D, Yusuf R, Saez B, et al. AKT/FOXO signaling enforces reversible differentiation blockade in myeloid leukemias. Cell. 2011;146:697–708.
Namasu CY, Katzerke C, Bräuer-Hartmann D, Wurm AA, Gerloff D, Hartmann JU, et al. ABR, a novel inducer of transcription factor C/EBPα, contributes to myeloid differentiation and is a favorable prognostic factor in acute myeloid leukemia. Oncotarget. 2017;8:103626–39.
Jiang X, Hu C, Arnovitz S, Bugno J, Yu M, Zuo Z, et al. miR-22 has a potent anti-tumour role with therapeutic potential in acute myeloid leukaemia. Nat Commun. 2016;7:11452.
Cui S, Chen Y, Guo Y, Wang X, Chen D. Hsa-miR-22-3p inhibits liver cancer cell EMT and cell migration/ invasion by indirectly regulating SPRY2. PLoS ONE. 2023;18:e0281536.
Yang X, Su W, Li Y, Zhou Z, Zhou Y, Shan H, et al. MiR-22-3p suppresses cell growth via MET/STAT3 signaling in lung cancer. Am J Transl Res. 2021;13:1221–32.
Wang J, Li Y, Ding M, Zhang H, Xu X, Tang J. Molecular mechanisms and clinical applications of miR-22 in regulating malignant progression in human cancer (Review). Int J Oncol. 2017;50:345–55.
Li D, Zhao Y, Liu C, Chen X, Qi Y, Jiang Y, et al. Analysis of MiR-195 and MiR-497 expression, regulation and role in breast cancer. Clin Cancer Res. 2011;17:1722–30.
Bai J, Xu J, Zhao J, Zhang R. lncRNA SNHG1 cooperated with miR-497/miR-195-5p to modify epithelial–mesenchymal transition underlying colorectal cancer exacerbation. J Cell Physiol. 2020;235:1453–68.
Chae D-K, Park J, Cho M, Ban E, Jang M, Yoo YS, et al. MiR-195 and miR-497 suppress tumorigenesis in lung cancer by inhibiting SMURF2-induced TGF-β receptor I ubiquitination. Mol Oncol. 2019;13:2663–78.
Yu W, Liang X, Li X, Zhang Y, Sun Z, Liu Y, et al. MicroRNA-195: a review of its role in cancers. Onco Targets Ther. 2018;11:7109–23.
Yang G, Xiong G, Cao Z, Zheng S, You L, Zhang T, et al. miR-497 expression, function and clinical application in cancer. Oncotarget. 2016;7:55900–11.
Xiao G, Li Y, Wang Y, Zhao B, Zou Z, Hou S, et al. LncRNA PRAL is closely related to clinical prognosis of multiple myeloma and the bortezomib sensitivity. Exp Cell Res. 2018;370:254–63.
Su P, Wang F, Qi B, Wang T, Zhang S. P53 regulation-association long non-coding RNA (lncRNA PRAL) inhibits cell proliferation by regulation of P53 in human lung cancer. Med Sci Monit. 2017;23:1751–8.
Zhou CC, Yang F, Yuan SX, Ma JZ, Liu F, Yuan JH, et al. Systemic genome screening identifies the outcome associated focal loss of long noncoding RNA PRAL in hepatocellular carcinoma. Hepatology. 2016;63:850–63.
Wang H, Wang J, Liang CF, Zhou T. Expression of long non-coding RNA PRAL as a potential biomarker for diagnosis in non-small-cell lung cancer patients is associated with the inhibition of cell proliferation and metastasis. Clin Lab. 2018;64:1341–8.
Jiang J, Hu H, Chen Q, Zhang Y, Chen W, Huang Q, et al. Long non-coding RNA SNHG29 regulates cell senescence via p53/p21 signaling in spontaneous preterm birth. Placenta. 2021;103:64–71.
Liu S, Zhou J, Ye X, Chen D, Chen W, Lin Y, et al. A novel lncRNA SNHG29 regulates EP300- related histone acetylation modification and inhibits FLT3-ITD AML development. Leukemia. 2023;37:1421–34.
Ni W, Mo H, Liu Y, Xu Y, Qin C, Zhou Y, et al. Targeting cholesterol biosynthesis promotes anti-tumor immunity by inhibiting long noncoding RNA SNHG29-mediated YAP activation. Mol Ther. 2021;29:2995–3010.
Wang F, Tian X, Zhou J, Wang G, Yu W, Li Z, et al. A three‑lncRNA signature for prognosis prediction of acute myeloid leukemia in patients. Mol Med Rep. 2018;18:1473–84.
Huang W, Sun YM, Pan Q, Fang K, Chen XT, Zeng ZC, et al. The snoRNA-like lncRNA LNC-SNO49AB drives leukemia by activating the RNA-editing enzyme ADAR1. Cell Discov. 2022;8:117.
Ussowicz M, Marcel V, Long FNV, Kazanowska B, Diaz JJ, Wołowiec D. Analysis of the rRNA methylation complex components in pediatric B-cell precursor acute lymphoblastic leukemia: A pilot study. Adv Clin Exp Med. 2020;29:107–13.
Huang R, Liao X, Li Q. Integrative genomic analysis of a novel small nucleolar RNAs prognostic signature in patients with acute myelocytic leukemia. Math Biosci Eng. 2022;19:2424–52.
Huang Z-H, Du Y-P, Wen J-T, Lu B-F, Zhao Y. snoRNAs: functions and mechanisms in biological processes, and roles in tumor pathophysiology. Cell Death Discovery. 2022;8:259.
Liu Y, Sun J, Zhao M. ONGene: A literature-based database for human oncogenes. J Genet Genomics. 2017;44:119–21.
Sekar A, Griffin R, Parikh SA, Genovese G, Robinson DP, Norman AD, et al. Mosaic chromosomal alterations (mCAs) in individuals with monoclonal B-cell lymphocytosis (MBL). Blood Cancer J. 2024;14:193.
Griffin R, Wiedmeier-Nutor JE, Parikh SA, McCabe CE, O’Brien DR, Boddicker NJ, et al. Differential prognosis of single and multiple TP53 abnormalities in high-count MBL and untreated CLL. Blood Adv. 2023;7:3169–79.
Tang G, Wu Y, Lin P, Toruner GA, Hu S, Li S, et al. Cytogenetic profile in monoclonal gammopathy of undetermined significance, smoldering and symptomatic multiple myeloma: A Study of 1087 patients with highly purified plasma cells. Cancers. 2023;15:5690.
Bustoros M, Sklavenitis-Pistofidis R, Park J, Redd R, Zhitomirsky B, Dunford AJ, et al. Genomic profiling of smoldering multiple myeloma identifies patients at a high risk of disease progression. J Clin Oncol. 2020;38:2380–9.
Hermsen M, Postma C, Baak J, Weiss M, Rapallo A, Sciutto A, et al. Colorectal adenoma to carcinoma progression follows multiple pathways of chromosomal instability. Gastroenterology. 2002;123:1109–19.
Malcikova J, Stano-Kozubik K, Tichy B, Kantorova B, Pavlova S, Tom N, et al. Detailed analysis of therapy-driven clonal evolution of TP53 mutations in chronic lymphocytic leukemia. Leukemia. 2015;29:877–85.
Lakshman A, Painuly U, Rajkumar SV, Ketterling RP, Kapoor P, Greipp PT, et al. Natural history of multiple myeloma with de novo del(17p). Blood Cancer J. 2019;9:32.
Zhang L, Liu D, Tang B, Xu A, Huang H, Su Y, et al. MicroRNA-324-5p suppresses the migration and invasion of MM cells by inhibiting the SCF(β-TrCP) E3 ligase. Oncol Lett. 2018;16:5331–8.
Dong Z, Yeo KS, Lopez G, Zhang C, Dankert Eggum EN, Rokita JL, et al. GAS7 deficiency promotes metastasis in MYCN-driven neuroblastoma. Cancer Res. 2021;81:2995–3007.
Jansen R, Moehlendick B, Bartenhagen C, Tóth C, Lehwald N, Stoecklein NH, et al. ACGH detects distinct genomic alterations of primary intrahepatic cholangiocarcinomas and matched lymph node metastases and identifies a poor prognosis subclass. Sci Rep. 2018;8:10637.
Qin LX. Chromosomal aberrations related to metastasis of human solid tumors. World J Gastroenterol. 2002;8:769–76.
Zenz T, Häbe S, Denzel T, Mohr J, Winkler D, Bühler A, et al. Detailed analysis of p53 pathway defects in fludarabine-refractory chronic lymphocytic leukemia (CLL): dissecting the contribution of 17p deletion, TP53 mutation, p53-p21 dysfunction, and miR34a in a prospective clinical trial. Blood. 2009;114:2589–97.
Saygin C, Hu E, Zhang P, Sher S, Lozanski A, Doong TJ, et al. Genomic analysis of cellular hierarchy in acute myeloid leukemia using ultrasensitive LC-FACSeq. Leukemia. 2021;35:3406–20.
Merz M, Jauch A, Hielscher T, Mai EK, Seckinger A, Hose D, et al. Longitudinal fluorescence in situ hybridization reveals cytogenetic evolution in myeloma relapsing after autologous transplantation. Haematologica. 2017;102:1432–8.
Croft J, Ellis S, Sherborne AL, Sharp K, Price A, Jenner MW, et al. Copy number evolution and its relationship with patient outcome-an analysis of 178 matched presentation-relapse tumor pairs from the Myeloma XI trial. Leukemia. 2021;35:2043–53.
Thanendrarajan S, Tian E, Qu P, Mathur P, Schinke C, van Rhee F, et al. The level of deletion 17p and bi-allelic inactivation of TP53 has a significant impact on clinical outcome in multiple myeloma. Haematologica. 2017;102:e364–e7.
Guo Y, Wu H, Wiesmüller L, Chen M. Canonical and non-canonical functions of p53 isoforms: potentiating the complexity of tumor development and therapy resistance. Cell Death Dis. 2024;15:412.
Lukow DA, Sheltzer JM. Chromosomal instability and aneuploidy as causes of cancer drug resistance. Trends Cancer. 2022;8:43–53.
Bagacean C, Tempescul A, Ternant D, Banet A, Douet-Guilbert N, Bordron A, et al. 17p deletion strongly influences rituximab elimination in chronic lymphocytic leukemia. J Immunother Cancer. 2019;7:22.
Roncuzzi L, Brognara I, Baiocchi D, Amadori D, Gasperi-Campani A. Loss of heterozygosity at 17p13.3-ter, distal to TP53, correlates with negative hormonal phenotype in sporadic breast cancer. Oncol Rep. 2005;14:471–4.
Cunningham SC, Gallmeier E, Hucl T, Dezentje DA, Abdelmohsen K, Gorospe M, et al. Theoretical proposal: Allele dosage of MAP2K4/MKK4 could rationalize frequent 17p loss in diverse human cancers. Cell Cycle. 2006;5:1090–3.
Davies CC, Harvey E, McMahon RF, Finegan KG, Connor F, Davis RJ, et al. Impaired JNK signaling cooperates with KrasG12D expression to accelerate pancreatic ductal adenocarcinoma. Cancer Res. 2014;74:3344–56.
Wang W, Wang YQ, Meng T, Yi JM, Huan XJ, Ma LP, et al. MCL-1 degradation mediated by JNK activation via MEKK1/TAK1-MKK4 contributes to anticancer activity of new tubulin inhibitor MT189. Mol Cancer Ther. 2014;13:1480–91.
Mato AR, Thompson M, Allan JN, Brander DM, Pagel JM, Ujjani CS, et al. Real-world outcomes and management strategies for venetoclax-treated chronic lymphocytic leukemia patients in the United States. Haematologica. 2018;103:1511–7.
Hou Y, Hou L, Liang Y, Zhang Q, Hong X, Wang Y, et al. The p53-inducible CLDN7 regulates colorectal tumorigenesis and has prognostic significance. Neoplasia. 2020;22:590–603.
Xu S, Zhan M, Jiang C, He M, Yang L, Shen H, et al. Genome-wide CRISPR screen identifies ELP5 as a determinant of gemcitabine sensitivity in gallbladder cancer. Nat Commun. 2019;10:5492.
Jiang L, Kon N, Li T, Wang SJ, Su T, Hibshoosh H, et al. Ferroptosis as a p53-mediated activity during tumour suppression. Nature. 2015;520:57–62.
Li X, Xiong W, Wang Y, Li Y, Cheng X, Liu W. p53 activates the lipoxygenase activity of ALOX15B via inhibiting SLC7A11 to induce ferroptosis in bladder cancer cells. Lab Invest. 2023;103:100058.
Li J, Tang LL, Ma J. Survival-related indicators ALOX12B and SPRR1A are associated with DNA damage repair and tumor microenvironment status in HPV 16-negative head and neck squamous cell carcinoma patients. BMC Cancer. 2022;22:714.
Yang X, Liu J, Wang C, Cheng KK, Xu H, Li Q, et al. miR-18a promotes glioblastoma development by down-regulating ALOXE3-mediated ferroptotic and anti-migration activities. Oncogenesis. 2021;10:15.
Chu B, Kon N, Chen D, Li T, Liu T, Jiang L, et al. ALOX12 is required for p53-mediated tumour suppression through a distinct ferroptosis pathway. Nat Cell Biol. 2019;21:579–91.
Shintoku R, Takigawa Y, Yamada K, Kubota C, Yoshimoto Y, Takeuchi T, et al. Lipoxygenase-mediated generation of lipid peroxides enhances ferroptosis induced by erastin and RSL3. Cancer Sci. 2017;108:2187–94.
Zhang H, Deng T, Liu R, Ning T, Yang H, Liu D, et al. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol Cancer. 2020;19:43.
Kazan HH, Urfali-Mamatoglu C, Yalcin GD, Bulut O, Sezer A, Banerjee S, et al. 15-LOX-1 has diverse roles in the resensitization of resistant cancer cell lines to doxorubicin. J Cell Physiol. 2020;235:4965–78.
Qi L, Pan X, Chen X, Liu P, Chen M, Zhang Q, et al. COX-2/PGE2 upregulation contributes to the chromosome 17p-deleted lymphoma. Oncogenesis. 2023;12:5.
Liu J, Chen G, Pelicano H, Liao J, Huang J, Feng L, et al. Targeting p53-deficient chronic lymphocytic leukemia cells in vitro and in vivo by ROS-mediated mechanism. Oncotarget. 2016;7:71378–89.
Cui X, Han L, Cui L, Fu G, Liu E, Wang D, et al. Immune index: A gene and cell prognostic signature for immunotherapy response prediction in hepatocellular carcinoma. Pharmacol Res. 2023;187:106583.
Singh RK, Jones RJ, Shirazi F, Qin L, Zou J, Hong S, et al. Novel anti-B-cell maturation antigen alpha-amanitin antibody-drug conjugate HDP-101 shows superior activity to belantamab mafodotin and enhanced efficacy in deletion 17p myeloma models. Res Sq [Preprint]. 2024 Jan:rs.3.rs-3843028.
Figueroa-Vazquez V, Ko J, Breunig C, Baumann A, Giesen N, Pálfi A, et al. HDP-101, an anti-BCMA antibody-drug conjugate, safely delivers amanitin to induce cell death in proliferating and resting multiple myeloma cells. Mol Cancer Ther. 2021;20:367–78.
Raab MS, Kaufman JL, Richard S, Grosicki S, Takacs I, Strassz A, et al. Hdp-101, an anti-BCMA antibody-drug conjugate with a novel payload amanitin in patients with relapsed multiple myeloma, initial findings of the first in human study. Blood. 2023;142:3334.
Altwegg KA, Viswanadhapalli S, Mann M, Chakravarty D, Krishnan S, Liu Z, et al. A first-in-class inhibitor of ER coregulator PELP1 targets ER+ Breast Cancer. Cancer Res. 2022;82:3830–44.
Choi YB, Ko JK, Shin J. The transcriptional corepressor, PELP1, recruits HDAC2 and masks histones using two separate domains. J Biol Chem. 2004;279:50930–41.
Pettitt AR, Jackson R, Carruthers S, Dodd J, Dodd S, Oates M, et al. Alemtuzumab in combination with methylprednisolone Is a highly effective induction regimen for patients with chronic lymphocytic leukemia and deletion of TP53: Final results of the National Cancer Research Institute CLL206 trial. J Clin Oncol. 2012;30:1647–55.
Yuan C, Yuan M, Li W, Cheng H, Luo J, Zhang Q, et al. The STAT3 inhibitor stattic overcome bortezomib-resistance in multiple myeloma via decreasing PSMB6. Exp Cell Res. 2023;429:113634.
Gao L, Xue B, Xiang B, Liu KJ. Arsenic trioxide disturbs the LIS1/NDEL1/dynein microtubule dynamic complex by disrupting the CLIP170 zinc finger in head and neck cancer. Toxicol Appl Pharmacol. 2020;403:115158.
Truong MA, Cané-Gasull P, Lens SMA. Modeling specific aneuploidies: from karyotype manipulations to biological insights. Chromosome Res. 2023;31:25.
Mansouri S, Heylmann D, Stiewe T, Kracht M, Savai R. Cancer genome and tumor microenvironment: Reciprocal crosstalk shapes lung cancer plasticity. eLife. 2022;11:e79895.
Acknowledgements
Preparation of this article was supported by Northwest Cancer Research UK and Bloom Appeal (NWCRTBA2021.02) grant funding to AC, MG, ARP, NK and JRS. The authors would like to thank Dr Marco Sciacovelli (University of Liverpool) for critical reading of this manuscript.
Author information
Authors and Affiliations
Contributions
FvK, AC, MAG and JRS analysed data and designed, edited and wrote the manuscript. AK analysed data and helped write the manuscript. JS analysed data. ARP and NK helped write and edit the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests financial or otherwise Preparation of this article was supported by a Northwest Cancer Research UK and Bloom Appeal (NWCRTBA2021.02) grant funding to AC, MG, ARP, NK and JRS.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
van Kampen, F., Clark, A., Soul, J. et al. Deletion of 17p in cancers: Guilt by (p53) association. Oncogene 44, 637–651 (2025). https://doi.org/10.1038/s41388-025-03300-8
Received:
Revised:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41388-025-03300-8
This article is cited by
-
Low expression of ALOX15B modulates immunosuppressive tumor microenvironment in diffuse large B-cell lymphoma via the TAP1/MHC-I axis
Journal of Experimental & Clinical Cancer Research (2026)
