
Abstract
In this study, the effect of cumulative ACEs experienced on human maternal DNA methylation (DNAm) was estimated while accounting for interaction with domains of ACEs in prenatal peripheral blood mononuclear cell samples from the Maternal and Developmental Risks from Environmental Stressors (MADRES) pregnancy cohort. The intergenerational transmission of ACE-associated DNAm was also explored used paired maternal (N = 120) and neonatal cord blood (N = 69) samples. Replication in buccal samples was explored in the Children’s Health Study (CHS) among adult parental (N = 31) and pediatric (N = 114) samples. We used a four-level categorical indicator variable for ACEs exposure: none (0 ACEs), low (1–3 ACEs), moderate (4–6 ACEs), and high (>6 ACEs). Effects of ACEs on maternal DNAm (N = 240) were estimated using linear models. To evaluate evidence for intergenerational transmission, mediation analysis (N = 60 mother-child pairs) was used. Analysis of maternal samples displayed some shared but mostly distinct effects of ACEs on DNAm across low, moderate, and high ACEs categories. CLCN7 and PTPRN2 was associated with maternal DNAm in the low ACE group and this association replicated in the CHS. CLCN7 was also nominally significant in the gene expression correlation analysis among maternal profiles (N = 35), along with 11 other genes. ACE-associated methylation was observed in maternal and neonatal profiles in the COMT promoter region, with some evidence of mediation by maternal COMT methylation. Specific genomic loci exhibited mutually exclusive maternal ACE effects on DNAm in either maternal or neonatal population. There is some evidence for an intergenerational effect of ACEs, supported by shared DNAm signatures in the COMT gene across maternal-neonatal paired samples.
Similar content being viewed by others
Introduction
Adverse Childhood Experiences (ACEs) refers to a collection of adverse events that occurs before a child turns 18 years old [1]. Accumulating ACEs are associated with a heightened risk of several unfavorable outcomes – including all-cause mortality and other chronic disease like cardiovascular disease [2]. ACEs are common, with 57% of adults reporting more than one ACE [2]. Exposure to ACEs can be quantified as an increasing number of events overall, or assessed within domains of abuse (sexual, physical, or emotional), neglect (emotional or physical), and dysfunction (depression, incarceration, domestic violence, substance abuse, and mental illness) [1]. Risk patterns of ACEs can manifest multi-generationally: children of individuals who experiences ACEs are at increased risk for experiencing ACEs themselves [3, 4]. Strikingly, this multigenerational effect of ACEs is present from birth: infants born to mothers with high ACEs exhibit aberrant stress responses compared to infants of mothers with no ACEs [5]. Such evidence may speak to larger biological and adaptive processes activated in response to chronic early life stress [6].
Improvement in trauma informed care (TIC) can potentially reduce risk of physical and mental health outcomes for individuals exposed to ACEs [7]. Investigating the biological underpinnings of ACEs can aid in evaluation of implemented TIC in healthcare settings. One biological consequence of ACEs is reflected in DNA methylation (DNAm) changes. DNAm is the molecular state captured when a methyl group is covalently bonded to the cytosine base of DNA, potentially influencing gene expression and increasing later life disease risk. Traumatic events can also induce stress which can trigger a physiological response by cells. The effects of early life adversity, maltreatment, and/or ACEs have been studied extensively within human and animal models in relation to DNAm [8,9,10]. Literature based on animal models is experimental and usually examines the effect of an isolated adverse event. Most human studies have focused on either specific domains or overall number of ACEs and few studies have explored the combinatorial effect of such factors. It is important to consider these two measures in tandem because the magnitude and domain of ACEs may contribute to variability in DNAm in human populations [11].
This study first aimed to examine the effect of the total number of ACEs experienced on maternal peripheral blood mononuclear cell (PBMC) DNAm during pregnancy while accounting for interaction with specific domains of ACEs. The possible intergenerational transmission of ACE-associated DNAm was also explored using paired maternal and child PBMC DNAm profiles in a mediation analysis. Relationships between maternal PBMC DNAm and maternal PBMC gene expression was evaluated. The analysis was conducted in a discovery and replication study from two population-based cohorts in Los Angeles, CA, using two distinct tissue types. Data from the Maternal and Developmental Risks from Environmental and Social Stressors (MADRES) pregnancy cohort [12] using maternal prenatal and cord blood collected at birth PBMC DNAm and gene expression was used as the discovery population and the Children’s Health Study (CHS) using family-based parental adult and pediatric child buccal cell data served as the replication population [13, 14]. In humans, there is extremely limited evidence for intergenerational transmission of ACEs and is currently an ongoing area of research. Conclusions regarding intergenerational transmission should be limited without coupled genotyping data.
Materials and methods
Discovery study sample and recruitment
Recruitment of pregnant participants in the MADRES prospective pregnancy cohort began in 2015. Participants were recruited through four prenatal providers in Los Angeles, California if they were less than 30 weeks gestational age at cohort entry and at least 18 years old. The Institutional Review Board (IRB) at the University of Southern California authorized MADRES study protocol, and informed consent was provided by study participants prior to cohort entry (IRB #HS-15-00498). Additional exclusion criteria include incarceration during recruitment, multiple pregnancy, and HIV-positive status. Pregnant participants were interviewed at each trimester, birth, and postnatally for up to a year. Details on all variables collected and additional exclusion criteria in the MADRES study is available here [12].
MADRES maternal DNAm
Blood samples were collected in EDTA tubes (BD# 366643) and transported upright on ice to the lab at Norris Cancer Center, where they were centrifuged within 1 h of collection and overlying plasma was removed. PBMCs were isolated from 10 mL peripheral blood samples collected during the early and late pregnancy visit (Fig. S1). DNA was extracted using the AllPrep DNA kit (Qiagen). Plasma aliquots were stored within 1 h of collection at −20 °C. Bisulfite conversion of DNA was conducted using the EZ DNAm Kit (Zymo Research) and was performed in batches, with families plated together. Then, DNAm was quantified using the Illumina Infinium HumanMethylationEPIC (850 K) assay using the manufacturer’s recommended protocol with no other modifications.
MADRES cord blood DNAm
10 mL of umbilical cord blood was collected at birth and stored within 24 h of delivery. Samples were collected by hospital staff and placed in a cooler on ice, then transported by couriers from the delivery hospital to the laboratory. Blood samples were not frozen until after centrifugation to separate the plasma, PBMCs, and red blood cells. PBMCs were frozen and thawed at −20 °C before DNA extraction. Bisulfite converted using the EZ DNAm Kit (Zymo Research) and was performed in batches, with familial samples plated together. The Illumina Infinium HumanMethylationEPIC was used to quantify DNAm under the same protocols as the maternal arrays.
MADRES DNAm quality control
All data analysis was performed in R (version v4.1.0, R Core Team 2021). Quality control and normalization of data were performed separately for maternal and cord blood samples. Sample and probe level quality control were performed using standard protocols outlined by the minfi Bioconductor package [15]. Briefly, poor detection p-values were computed across probes, representing those probes with no significant difference in detection between background and control probes, and were removed from the analysis. If a sample had more than 10% of poor (p > 0.01) detection p-value probes, it was removed from the analysis. Cross-reactive and polymorphic probes were also removed [16, 17]. Outlier individuals, or those displaying a median probe intensity below the minfi default value of 10.5, were also removed from the analysis. Sex predicted from intensity of X and Y chromosomes was used as a quality control check. Noob background correction for dye-bias [18] followed by quantile normalization [19] was used for normalization. SNP-associated probes were removed from the analysis. Log-transformed beta-values were used in downstream regression analysis [15]. Figure S2 is a consort diagram of our quality control and sample normalization process. In the maternal samples, 5% of samples failed quality control. In cord blood samples, 15% were dropped due to low median intensities (N = 16) and sex discrepancy (N = 3).
MADRES maternal mRNA
Maternal PBMC gene expression levels for this analysis were profiled for the same mothers with DNAm profiles in pregnancy. PBMCs were isolated from 10 mL of whole blood collected in early pregnancy (N = 35). Total mRNA was extracted using the Qiagen Allprep DNA/RNA Isolation Kit and sequenced by the Dartmouth Genomics Core Laboratory. Quality of mRNA was assigned using a fragment analyzer instrument, while mRNA quantity was determined using qubit. The Kapa RNA HyperPrep and RiboErase plus globin kit was used with 500 ng of high-quality mRNA as input. For final library amplification, 12 PCR cycles were used. NextSeq500 was used to sequence all samples, resulting in the generation of 25 million, 75 bp single end reads per sample. RTA v2.4.11 was used for base calling and changed into fastqs using bcl2fastq v2.20.0.0422. Samples displayed good quality with a depth between 20 and 70 million reads for each sample with a median alignment rate of 85%. MultiQC plots were generated to examine sample quality across maternal profiles – no samples were flagged for removal from the analysis based on phred score, per base N content, or per sequence GC content [20]. The HISAT2 aligner was used to map reads to the reference; GRCh genome annotation number 97 from Ensembl [21, 22]. FeatureCounts was used to quantify reads to exons [23]. Transcripts per million (TPM) log2-transformed counts were generated from raw counts in downstream correlative analysis [24]. Our sample was limited to participants with available ACE questionnaire and DNAm data (N = 35). These data are publicly available in Gene Omnibus Expression (Accession Number: GSE18175).
Children’s health study DNAm
The CHS is a prospective cohort study that recruited schoolchildren from the Southern California region from the 1990s to 2000s [13, 14]. The study’s goal was to determine the effects of air pollution and other environmental exposures on respiratory health. Buccal cell samples were collected from a subset of 235 recruited participants and their families. Approximately two decades after baseline, a convenience sample of adult-aged index participants were invited to participate in a follow-up mail-based study. Buccal samples were collected from the adult participant, their partner, and their child. Informed consent was provided (IRB #HS-17-00778). Pediatric buccal cells were collected with swab using the Oragene OC-175 kit or toothbrush collection methods for each family trio in approximately 5 mL of buffer. DNA was extracted with the Oragene prepIT L2P kit. The extracted DNA was stored locally at −80 °C. The Zymo EZ DNAm kit was used to perform bisulfite conversion, and the Illumina EPIC DNAm protocol was used to generate the data. For the purposes of replication, the CHS samples was limited to the recently collected adult (N = 31) and offspring pediatric (N = 114) samples. All sample and probe level filters applied to the discovery population were applied to the replication population in the CHS. Figure S3 is a consort diagram for the pediatric and parental populations. 12% of our samples and 90% of probes were retained from the parental population, while 67% of samples and 90% of probes were retained in the pediatric participants.
Cell Type estimation
Cell type estimation was conducted separately on discovery and replication datasets using generated, normalized DNA methylation profiles using DNA extraction from samples that had not experienced a previous freeze-thaw cycle. The Houseman method using the EPIC reference platform was executed in minfi [15] using the FlowSorted.Blood.EPIC data for cord blood and whole blood, respectively [25]. Cord blood and blood composite cell type was used for neonatal and maternal data respectively in MADRES. For buccal cell data in the CHS, immune cell type proportions were estimated using HEpiDISH [26].
Genetic factors
Given the confounding impact of ancestral based SNP-variation on DNAm in this analysis, we used EPISTRUCTURE principal components generated from the quality-controlled DNAm dataset. The top two EPISTRUCTURE principal components were used in maternal and neonatal analyses. EPISTRUCTURE methods are described elsewhere. Briefly, the program uses sparse principal component analysis and was validated in multi-ethnic populations [27].
Measurement of adverse childhood experiences
The Adverse Childhood Experiences (ACEs) CDC-Kaiser questionnaire is shown in Fig. S4 [1]. The ACEs questionnaire was administered during the second pregnancy trimester in MADRES. In CHS, the questionnaires were distributed during the mail-based follow-up and were administered to both parents and children. The same questionnaire was administered in both the discovery and replication populations. ACE scores were treated as a categorical indicator variable for all analyses: none (0 ACEs), low (1–3 ACEs), moderate (4–6 ACEs), and high (>6 ACEs).
Covariate selection
In addition to cell type, batch, gestational age at sample collection [28, 29], and genetic covariates identified above, confounders were identified a priori from the literature using a directed acyclic graph (DAG) (Fig. C1, Supplementary Text). For estimating maternal effects, the following covariates were included: maternal age at birth, pre-pregnancy body mass index (BMI), a composite variable for National Institute of Health (NIH) race categories, ethnicity, and nativity status (Black and Non-Hispanic, Foreign-born Hispanic, Multiracial, US Born Hispanic, and White Non-Hispanic), any evidence of glucose dysregulation, any evidence of a hypertensive disorder, socioeconomic status defined as highest maternal education attained (less than high school, high school or some college, and any college degree), nulliparity status (binary defined as first pregnancy), prenatal smoking use (binary defined as mothers who reported smoking in both their early and late pregnancy visits), and maternal depressive symptoms using the Center for Epidemiology Studies for Depression (CES-D) Scale (cut-off of 16 for probable clinical depression) [30]. Depressive symptoms were also measured in binary questionnaire (have you ever been diagnosed with depression by a doctor?) in CHS compared to MADRES. In addition, the top two surrogate variables produced from surrogate variable analysis (SVA) was used to control for technical variation in our maternal and neonatal datasets. SVA uses probe level data to estimate unwanted sources of heterogeneity and produces variables that capture the variability in said heterogeneity [31]. The replication population adjusted for the same covariates, including sex in the parental population. The pediatric CHS model also included a covariate for ACE score of the child. Technical covariates were not adjusted for in mediation analysis. Additional information on covariate selection can be found in the supplemental text.
Statistical analysis
The objective was to estimate effects of ACEs on DNAm and then determine if these effects show evidence of intergenerational transmission using a mediation analysis.
Effects of each ACE category on maternal DNAm was estimated by adjusting for confounders detailed above. Linear regression models were implemented in limma [32] accounting for correlation between the early and late pregnancy visit (n = 120, N = 240) in our prenatal samples (Fig. S1) to estimate differentially methylation positions (DMPs) and differentially methylated regions (DMRs) [33, 34]. Interactions with domains of abuse and neglect were analyzed within the low and moderate ACE categories because individuals in the high ACE group all had experiences across the three domains. A similar approach for the main effect was used in the neonatal cord blood dataset. Then, using the average beta value among the probes mapped to maternal DMRs annotated to mRNA-sequencing genes, Pearson correlation were used to measure the direction and strength of the associations between maternal DNAm and TPM-normalization maternal expression of the associated gene during pregnancy.
Second, overlapping significant (FDR < 0.05) DMRs between maternal and neonatal datasets were used as candidates for the intergenerational mediation analysis. Significance of the indirect pathway was tested using a quasi-Bayesian method for p-value and confidence interval estimation from the mediation package [35]. Mothers reporting persistent prenatal smoking (n = 3) were removed from the mediation analysis due to potential for exposure-induced mediator-outcome confounding, resulting in a total of 60 mother-child pairs (Fig. S2). Additional statistical models for mediation sensitivity analysis are described in the supplemental text.
Pathway analysis
We conducted a pathway analysis by inputting all FDR significant DMR annotated genes in DMRcate [33, 34] into shinyGO for Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment [36].
Results
Characteristics of the study populations
Characteristics of MADRES participants (N = 240, n = 120) are presented in Table 1. Individuals with higher ACE scores were more likely to have lower educational attainment (p = 7.88 × 10−4). Prevalence of ACEs also varied significantly by race, ethnicity, and foreign-born status (p = = 7.23 × 10−8). Individuals also differed in their diabetes prevalence (p = 0.0112), with a lower prevalence in the low ACE group. Characteristics of CHS index adult subjects (n = 31) and pediatric participants (n = 173) are listed in Tables S1 and S2.
Associations between ACEs and maternal DMPs/DMRs
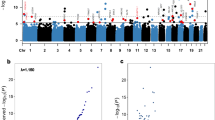
In a DMP analysis using individuals with no ACEs as the reference, we identified 784 DMPs associated with a low ACE score, 710 DMPs associated with a moderate ACE score, and 313 DMPs associated with a high ACE score (Fig. 1). 67%, 61% and 62% of DMPs had lower methylation levels in low, moderate, and high ACE scores compared to the group with no ACEs. In a DMR analysis using individuals with no ACEs as the reference, we identified 138 DMRs associated with low ACE score, 133 with a moderate ACE score, and 56 with a high ACE. A table of the top 10 DMRs is listed in Table 2. Two genes and 10 DMPs (Fig. 2) were consistently identified as having DMRs across the ACE categories: COMT/TXNRD2 and KCNQ1.

Top five p-value significant (FDR < 0.05) annotated genes for each ACEs category: none (0 ACEs), low (1-3 ACEs), moderate (4-6 ACEs), and high (>6 ACEs) are labelled. All red and blue dots are significant probes (FDR < 0.05), all black dots are those that did not meet FDR significance (FDR > 0.05). Blue indicates a log-fold change with methylation higher in ACE groups versus the unexposed individuals, and red indicates a log-fold change with methylation lower in exposed versus unexposed individuals.

Boxplot of beta values for Maternal CpG cg20656154 significant (FDR < 0.05) across higher ACE scores. There was a trend for lower methylation in higher ACE scores compared to those with no ACEs.
DMR interactions by ACE domain of abuse and neglect
49 DMRs were associated with abuse in the low ACE category, and 166 in the moderate group. Across the low and moderate group, the following DMRs were consistently identified for the following genes: LINC01044, CFDP1, CYP1B1, CERS3, PRKXP1, PPAPDC3, EIF2AK4, CCDC9, C6orf25.
Exposure to neglect was associated with 68 DMRs among the low ACE group and 141 DMRs in the moderately exposed group. Among individuals with a low or moderate ACE score with experiences of neglect, the following DMRs were shared: ABAT, MBP, C2CD2L, HLA-DPB2, FRMD4A, MSL3P1, DLEU7. The top five DMRs for each ACE group and domain are listed in Table 3.
Effect of ACEs on neonatal DMPs/DMRs
Among 69 neonates with available cord blood data paired to maternal ACEs, 8 significant DMPs were associated with maternal ACEs in the low category, 2 in the moderate category, and none in the high category. 1 DMR med FDR < 0.05 criteria for significance in the low ACE group (COMT) and 2 DMRs were statistically significant in the moderate ACE group (COMT and ZFP57).
Intergenerational mediation analysis
We tested for mediation of the indirect effect of ACEs on neonatal DNAm by maternal DNAm for COMT/TXNRD2, a gene with a statistically significant (FDR < 0.05) DMRs identified independently in both maternal and neonatal samples. COMT was the only statistically significant (FDR < 0.05) DMR identified across all three ACE categories in prenatal samples and both the low and moderate ACE group among neonatal samples. COMT regional DNAm was partially (63%) mediated by maternal DNAm comparing individuals who were exposed to ACEs versus those who were not. The indirect effect was 0.14 with a 95% CI of (0.0025, 0.32) (p-value = 0.044) (Fig. 3).

Effect of Maternal ACEs on neonatal COMT methylation is partially (64%) mediated by maternal COMT methylation. This indirect effect was significant (p-value = 0.044) with a 95% CI of (0.0025, 0.32).
Correlation between maternal methylation in DMRs and gene expression
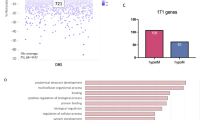
Among 144 statistically significant low ACE DMRs mapped to HCNC gene names, 12 displayed a nominally significant Pearson correlation (p < 0.05) with gene expression in the first trimester (Table S3). These included: NMNAT2, HDAC4, GAK, MFAP3, VARS2, MAFK, PON1, LYNX1, TUBGCP5, CLCN7, GPR108, AND TXNRD2.
Replication cohort: ACE-associated adult DMRs
In our evaluation of ACEs and adult DNAm in the adult replication cohort, a total of 82 DMRs were found in the low ACE group, 60 in the moderate ACE group, and six in the high ACE group. We replicated 2 DMRs from the low ACEs group and 1 DMRs from the moderate ACEs group in the CHS at an FDR < 0.05. Among the low ACE group DMRs, the CLCN7 and PTPRN2 gene were replicated (Fig. S5). DMRs in both genes were statistically significant in the moderate ACE group in CHS. Among the moderate ACE groups, the FOXK1 replicated across CHS and MADRES. There were no shared DMRs across the high ACE groups. Effect sizes across overlapping statistically significant DMRs were consistent in direction for both cohorts. When comparing the direction of effect among probes identified (FDR < 0.05) in MADRES prenatal samples, approximately 50% of probes were consistent in the same direction in our replication population (Fig. S6).
Replication cohort: parental ACE intergenerational pediatric DMRs
A total of 53 DMRs were found in the low ACE group, 36 in the moderate ACE group, and five in the high group. We replicated no DMRs from the neonatal DMRs in MADRES evaluated at an FDR < 0.05. Among the maternal MADRES genes, the GAK was replicated in the low ACE group. The replicated GAK DMR overlapped and were consistent in direction for the two cohorts.
Pathway analysis
To capture shared pathways influenced by exposure to ACEs, we input all unique DMP-associated genes to ShinyGo [36]. This was done independently for the maternal discovery and adult replication cohorts to determine potential overlap. Gene ontology analysis of the discovery population indicated the top pathways to be enriched for KEGG processes in nicotine addiction, GABAergic synapse, and oxytocin signaling pathway (Fig. 4). For the replication population, there were no enriched KEGG pathways.

Significant (FDR < 0.05) maternal DMP genes in MADRES input to ShinyGO to generate KEGG pathways.
Discussion
Few studies have explored shared signatures of childhood adversity on an epigenome-wide scale in multiple generations. Prior studies have generally been limited to candidate probe or regional studies in a single generation. Specifically, there has been focus on candidate genes including BDNF, NR3C1, AVP, and FKBP5 [9]. No DMRs in the main effect or interaction model demonstrated a statistically significant (FDR < 0.05) overlap with these candidate genes. However, there was overlap in our main effect and interaction models with other studies using genome-wide scans, including CACNA2D4 [37], ALS2 [38], OPRL1 [39], PRDM16, and C8orf31 [10]. ZFP57 was statistically significant in the interaction of abuse in the maternal population (Table 3) and the main effect in our neonatal population and has been associated with a shared multigenerational signature of neglect in human studies [40]. Moreover, CLCN7 and PTPRN2 were two overlapping statistically significant DMRs in our low ACE group in the discovery and replication population. PTPRN2 has been implicated with exposure to stress, violence and associated with mood state [41, 42]. CLCN7 is associated with osteoporosis [43]. We also found evidence of nominally significant Pearson correlations with 13 CpGs that were significant in the DNAm analysis, including CLCN7. These 13 CpGs, including PON1, have been previously implicated in DNAm signatures of maternal ACEs. [40] However, future studies should incorporate gene expression analyses with larger sample sizes, compared to our N = 35, and use models adjusted for relevant confounders.
We identified a DMR in the COMT/TXNRD2 gene for both mothers and neonates. We therefore tested whether maternal methylation levels in this gene mediated ACE impacts on neonatal methylation levels. COMT/TXNRD2 met the criteria for intergenerational mediation of ACEs on neonatal methylation levels. The DMR was identified in the north shore and island region of the COMT gene, overlapping with the end of the TXNRD2 gene and located in the 22q11.2 region [44]. This gene is involved in dopamine metabolism and methylation of this gene is associated with stress, pain sensitivity, and diagnosis of several neurological outcomes in adults [45]. Increased COMT methylation has been associated with malnutrition and impaired cognition, consistent in the direction of our adult discovery cohort [46]. Increasing promoter methylation of COMT has been associated with an increased risk of experiencing stress, consistent with the direction in our adult discovery cohort [47] (Fig. 5). Although COMT promoter methylation has been associated with Val158 polymorphism [48] twin studies indicate that there is residual variation not explained by genotype [49, 50]. The COMT Val158 polymorphism has a frequency of approximately 40% in Latin American populations [51], and therefore has the potential to bias our results away from the null. Therefore, even with control for EPISTRUCTURE genetic ancestry principal components, we cannot rule out the possibility of residual confounding by genotype. This intergenerational effect in the discovery population did not replication, and the COMT/TXNRD2 gene was not statistically significant in either adult or pediatric participants.

Beta Values for COMT regional methylation in maternal PBMC and neonatal cord blood samples in MADRES.
The lack of replication for some of our findings between MADRES and the CHS may reflect differences in tissue types, sex distributions, race, ethnicity, foreign born status, or age differences between the two cohorts. Our discovery population also had a higher prevalence of moderate and high ACE score exposure. However, there was also a higher prevalence of college educated individuals in our replication population, which may indicate that the replication population had access to resources to mitigate effects of ACEs, effectively biasing results towards the null. Moreover, the replication population was a convenience sample from the original cohort. It is therefore possible that selection bias may be distorting our results. It is also important to note the low prevalence of depressive and stress symptoms across the discovery and replication cohort, given that this is not consistent with the literature [1]. Therefore, the generalizability of these results may be limited.
This study used a DAG to guide covariate selection. We adjusted for confounders to promote exchangeability in our analysis. However, assumptions of consistency of the effect of ACEs on DNAm may be violated given that effects of time and circumstance may render shared experience among individuals with a similar ACE score. With regards to mediation analysis, assumptions of exchangeability are met. Exposure-induced mediator outcome confounding may be present due to maternal adulthood trauma, adulthood socioeconomic status, and maternal BMI. Maternal BMI was tested for evidence of exposure-induced mediator outcome confounding using a multiple sequential mediators’ product method [52] and adult socioeconomic status was restricted to the most prevalence group in our sample. Effects of maternal adulthood trauma were unmeasured in our cohort, so these may be conflating estimates. Additional discussion on DAG pathways and mediation sensitivity analysis can be found in the supplemental text.
This study provides empirical supportive evidence for the expectation of an intergenerational effect of ACEs on an epigenome-wide scale. One strength of the study was the rich discovery dataset, which enabled us to test for effects of ACEs on DNAm. However, the sample size in our discovery population was modest and may have been subject to Type I error. We were also limited in the overlap of findings for our replication cohort.
In this study, we observed that higher ACE scores were associated with DNAm both in our discovery prenatal sample and adult replication population. While some effects were conserved across low, medium, and high ACE scores, most effects were distinct across ACE categories and domains. More DMRs were shared between low and moderately exposed ACE categories than low and high exposed ACE categories, in part possibly due to higher statistical power.
Overall, two FDR-significant DMRs in our prenatal discovery population, PTPRN2 and CLCN7, replicated in our adult buccal cell population. CLCN7 was also statistically significant in the gene expression correlation analysis in the discovery population. Among the neonatal cord blood discovery and pediatric buccal replication populations, the GAK gene was replicated. A majority of identified DMRs and DMPs in the discovery and replication population were mapped to Open Sea regions.
Gene expression was correlated (p < 0.05) with DNAm across 12 DMRs. Six had an inverse correlation between DNAm and gene expression. Future studies should consider the impact of ACE DNAm signatures on gene expression in larger samples and analysis using adjusted models adjusted for cell composition.
The COMT/TXNRD2 DMR demonstrated an intergenerational association in our discovery population, but this effect did not persist in the replication analysis. Moreover, in our adult discovery population, methylation of this DMR was positively associated with gene expression.
Data availability
MADRES methylation data is available in the database of Genotypes and Phenotypes (Accession Number: phs003194.v1.p1). MADRES Gene expression data are publicly available in Gene Omnibus Expression (Accession Number: GSE18175). CHS methylation data is available upon reasonable request to Dr. Carrie Breton (breton@usc.edu).
Code availability
Code is available upon request to authors.
References
Felitti VJ, Anda RF, Nordenberg D, Williamson DF, Spitz AM, Edwards V, et al. Relationship of childhood abuse and household dysfunction to many of the leading causes of death in adults. The Adverse Childhood Experiences (ACE) Study. Am J Prev Med. 1998;14:245–58. https://doi.org/10.1016/s0749-3797(98)00017-8.
Webster EM. The impact of adverse childhood experiences on health and development in young children. Glob Pediatr Health. 2022;9:2333794X221078708. https://doi.org/10.1177/2333794X221078708.
Schickedanz A, Escarce JJ, Halfon N, Sastry N, Chung PJ. Intergenerational associations between parents’ and children’s adverse childhood experience scores. Child (Basel). 2021;8:747. https://doi.org/10.3390/children8090747.
Schofield TJ, Donnellan MB, Merrick MT, Ports KA, Klevens J, Leeb R. Intergenerational continuity in adverse childhood experiences and rural community environments. Am J Public Health. 2018;108:1148–52. https://doi.org/10.2105/AJPH.2018.304598.
Gray SAO, Jones CW, Theall KP, Glackin E, Drury SS. Thinking across generations: unique contributions of maternal early life and prenatal stress to neonatal physiology. J Am Acad Child Adolesc Psychiatry. 2017;56:922–9. https://doi.org/10.1016/j.jaac.2017.09.001.
Smith KE, Pollak SD. Early life stress and development: potential mechanisms for adverse outcomes. J Neurodev Disord. 2020;12:34 https://doi.org/10.1186/s11689-020-09337-y.
Oral R, Ramirez M, Coohey C, Nakada S, Walz A, Kuntz A, et al. Adverse childhood experiences and trauma informed care: the future of health care. Pediatr Res. 2016;79:227–33. https://doi.org/10.1038/pr.2015.197.
Folger AT, Nidey N, Ding L, Ji H, Yolton K, Ammerman RT, et al. Association between maternal adverse childhood experiences and neonatal SCG5 DNAm-effect modification by prenatal home visiting. Am J Epidemiol. 2022;191:636–45. https://doi.org/10.1093/aje/kwab270.
Parade SH, Huffhines L, Daniels TE, Stroud LR, Nugent NR, Tyrka AR. A systematic review of childhood maltreatment and DNAm: candidate gene and epigenome-wide approaches. Transl Psychiatry. 2021;11:134 https://doi.org/10.1038/s41398-021-01207-y.
Houtepen LC, Hardy R, Maddock J, Kuh D, Anderson EL, Relton CL, et al. Childhood adversity and DNAm in two population-based cohorts. Transl Psychiatry. 2018;8:266. https://doi.org/10.1038/s41398-018-0307-3
Weston A, Harris CC Assessment of Causation by the Bradford-Hill Criteria. In: Kufe DW, Pollock RE, Weichselbaum RR, Bast RC, Gansler TS, et al., editors. Holland-Frei Cancer Medicine. 6th edition. Hamilton (ON): BC Decker; 2003. Available from: https://www.ncbi.nlm.nih.gov/books/NBK13302/.
Bastain TM, Chavez T, Habre R, Girguis MS, Grubbs B, Toledo-Corral C, et al. Study design, protocol and profile of the maternal and developmental risks from environmental and social stressors (MADRES) pregnancy cohort: a prospective cohort study in predominantly low-income Hispanic women in Urban Los Angeles. BMC Pregnancy Childbirth. 2019;19:189. https://doi.org/10.1186/s12884-019-2330-7.
Peters JM, Avol E, Navidi W, London SJ, Gauderman WJ, Lurmann F, et al. A study of twelve Southern California communities with differing levels and types of air pollution. I. Prevalence of respiratory morbidity. Am J Respir Crit Care Med. 1999;159:760–7. https://doi.org/10.1164/ajrccm.159.3.9804143.
Gauderman WJ, Gilliland GF, Vora H, Avol E, Stram D, McConnell R, et al. Association between air pollution and lung function growth in southern California children: results from a second cohort. Am J Respir Crit Care Med. 2002;166:76–84. https://doi.org/10.1164/rccm.2111021.
Aryee MJ, Jaffe AE, Corrada-Bravo H, Ladd-Acosta C, Feinberg AP, Hansen KD, et al. “Minfi: A flexible and comprehensive Bioconductor package for the analysis of Infinium DNAm microarrays.”. Bioinformatics. 2014;30:1363–9. https://doi.org/10.1093/bioinformatics/btu049
Pidsley R, Zotenko E, Peters TJ, et al. Critical evaluation of the Illumina MethylationEPIC BeadChip microarray for whole-genome DNAm profiling. Genome Biol. 2016;17:208. https://doi.org/10.1186/s13059-016-1066-1
McCartney DL, Walker RM, Morris SW, McIntosh AM, Porteous DJ, Evans KL. Identification of polymorphic and off-target probe binding sites on the Illumina Infinium MethylationEPIC BeadChip. Genom Data. 2016;9:22–4. https://doi.org/10.1016/j.gdata.2016.05.012.
Triche TJ Jr, Weisenberger DJ, Van Den Berg D, Laird PW, Siegmund KD. Low-level processing of Illumina Infinium DNA Methylation BeadArrays. Nucleic Acids Res. 2013;41:e90. https://doi.org/10.1093/nar/gkt090.
Touleimat N, Tost J. Complete pipeline for Infinium(®) Human Methylation 450K BeadChip data processing using subset quantile normalization for accurate DNA methylation estimation. Epigenomics. 2012;4:325–41. https://doi.org/10.2217/epi.12.21.
Ewels Philip, Magnusson Måns, Lundin Sverker, Käller Max. MultiQC: summarize analysis results for multiple tools and samples in a single report. Bioinformatics. 2016;32:3047–8. https://doi.org/10.1093/bioinformatics/btw354
Kim D, Paggi JM, Park C, Bennett C, Salzberg SL. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat Biotechnol. 2019;37:907–15. https://doi.org/10.1038/s41587-019-0201-4
Cunningham F, Allen JE, Allen J, Alvarez-Jarreta J, Amode MR, Armean IM, et al. Ensembl 2022. Nucleic Acids Res. 2022;50:D988–D995. https://doi.org/10.1093/nar/gkab1049
Liao Y, Smyth GK, Shi W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics. 2014;30:923–30. https://doi.org/10.1093/bioinformatics/btt656.
Wagner GP, Kin K, Lynch VJ. Measurement of mRNA abundance using mRNA-seq data: RPKM measure is inconsistent among samples. Theory Biosci. 2012;131:281–5. https://doi.org/10.1007/s12064-012-0162-3.
Salas LA, Zhang Z, Koestler DCRondiAB, Hansen HM, Molinaro AM, et al. Enhanced cell deconvolution of peripheral blood using DNAm for high-resolution immune profiling. Nat Commun. 2022;13:761. https://doi.org/10.1038/s41467-021-27864-7
Zheng SC, Breeze CE, Beck S, Teschendorff AE. Identification of differentially methylated cell types in epigenome-wide association studies. Nat Methods. 2018;15:1059–66. https://doi.org/10.1038/s41592-018-0213-x.
Rahmani E, Shenhav L, Schweiger R, Yousefi P, Huen K, Eskenazi B, et al. Genome-wide methylation data mirror ancestry information. Epigenetics Chromatin. 2017;10:1. https://doi.org/10.1186/s13072-016-0108-y.
Adkins RM, Thomas F, Tylavsky FA, Krushkal J. Parental ages and levels of DNAm in the Thomas newborn are correlated. BMC Med Genet. 2011;12:47 https://doi.org/10.1186/1471-2350-12-47.
Non AL, Binder AM, Kubzansky LD, Michels KB. Genome-wide DNAm in neonates exposed to maternal depression, anxiety, or SSRI medication during pregnancy. Epigenetics. 2014;9:964–72. https://doi.org/10.4161/epi.28853.
Zarei K, Kahle L, Buckman DW, Choi K, Williams F. Parent-child nativity, race, ethnicity, and adverse childhood experiences among United States children. J Pediatr. 2022 Aug:S0022-347600672-2. https://doi.org/10.1016/j.jpeds.2022.07.050. Epub ahead of print. PMID: 35944710.
Leek JT, Johnson WE, Parker HS, Jaffe AE, Storey JD. The sva package for removing batch effects and other unwanted variation in high-throughput experiments. Bioinformatics. 2012;28:882–3. https://doi.org/10.1093/bioinformatics/bts034.
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, et al. “limma powers differential expression analyses formRNA-sequencing and microarray studies.”. Nucleic Acids Res. 2015;43:e47. https://doi.org/10.1093/nar/gkv007.
Peters TJ, Buckley MJ, Statham AL, Pidsley R, Samaras K, Lord RV, et al. “De novo identification of differentially methylated regions in the human genome.”. Epigenetics Chromatin. 2015;8:6 http://www.epigeneticsandchromatin.com/content/8/1/6.
Peters TJ, Buckley MJ, Chen Y, Smyth GK, Goodnow CC, Clark SJ. “Calling differentially methylated regions from whole genome bisulphite sequencing with DMRcate.” Nucleic Acids Res. 2021;49. https://academic.oup.com/nar/article/49/19/e109/6329576.
Tingley D, Yamamoto T, Hirose K, Keele L, Imai K. “mediation: R package for causal mediation analysis.”. J Stat Softw. 2014;59:1–38. http://www.jstatsoft.org/v59/i05/
Ge SX, Jung D, Yao R. ShinyGO: a graphical gene-set enrichment tool for animals and plants. Bioinformatics. 2020;36:2628–9. https://doi.org/10.1093/bioinformatics/btz931.
Cecil CAM, Zhang Y, Nolte T. Childhood maltreatment and DNAm: a systematic review. Neurosci Biobehav Rev. 2020;112:392–409. https://doi.org/10.1016/j.neubiorev.2020.02.019.
Labonte B, Suderman M, Maussion G, Navaro L, Yerko V, Mahar I, et al. Genome-wide epigenetic regulation by early-life trauma. Arch Gen Psychiatry. 2012;69:722–31.
Zhang H, Wang F, Kranzler HR, Zhao H, Gelernter J. Profiling of childhood adversity-associated DNA methylation changes in alcoholic patients and healthy controls. PLoS One. 2013;8:e65648. https://doi.org/10.1371/journal.pone.0065648.
León I, Herrero Roldán S, Rodrigo MJ, López Rodríguez M, Fisher J, Mitchell C, et al. The shared mother-child epigenetic signature of neglect is related to maternal adverse events. Front Physiol. 2022;13:966740. https://doi.org/10.3389/fphys.2022.966740.
Yan Q, Forno E, Cardenas A, Qi C, Han YY, Acosta-Pérez E, et al. Exposure to violence, chronic stress, nasal DNAm, and atopic asthma in children. Pediatr Pulmonol. 2021;56:1896–905. https://doi.org/10.1002/ppul.25372.
Rasmi Y, Shokati A, Hassan A, Aziz SG, Bastani S, Jalali L, et al. The role of DNAm in progression of neurological disorders and neurodegenerative diseases as well as the prospect of using DNAm inhibitors as therapeutic agents for such disorders. IBRO Neurosci Rep. 2022;14:28–37. https://doi.org/10.1016/j.ibneur.2022.12.002.
Sobacchi C, Villa A, Schulz A, Kornak U. CLCN7-Related Osteopetrosis. 2007 Feb 12 [updated 2022 Jan 20]. In: Adam MP, Mirzaa GM, Pagon RA, et al. editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2023. PMID: 20301306.
Schneider M, Debbané M, Bassett AS, Chow EW, Fung WL, van den Bree M, et al. International consortium on brain and behavior in 22q11.2 deletion syndrome. Psychiatric disorders from childhood to adulthood in 22q11.2 deletion syndrome: results from the international consortium on brain and behavior in 22q11.2 deletion syndrome. Am J Psychiatry. 2014;171:627–39. https://doi.org/10.1176/appi.ajp.2013.13070864.
Walton E, Liu J, Hass J, White T, Scholz M, Roessner V, et al. MB-COMT promoter DNAm is associated with working-memory processing in schizophrenia patients and healthy controls. Epigenetics. 2014;9:1101–7. https://doi.org/10.4161/epi.29223.
Peter CJ, Fischer LK, Kundakovic M, Garg P, Jakovcevski M, Dincer A, et al. DNAm signatures of early childhood malnutrition associated with impairments in attention and cognition. Biol Psychiatry. 2016;80:765–74. https://doi.org/10.1016/j.biopsych.2016.03.2100.
Wiegand A, Blickle A, Brückmann C, Weller S, Nieratschker V, Plewnia C. Dynamic DNAm changes in the COMT gene promoter region in response to mental stress and its modulation by transcranial direct current stimulation. Biomolecules. 2021;11:1726. https://doi.org/10.3390/biom11111726.
Thomas M, Banet N, Wallisch A, Glowacz K, Becker-Sadzio J, Gundel F, et al. Differential COMT DNAm in patients with borderline personality disorder: genotype matters. Eur Neuropsychopharmacol. 2019;29:1295–1300. https://doi.org/10.1016/j.euroneuro.2019.09.011.
Mill J, Dempster E, Caspi A, Williams B, Moffitt T, Craig I. Evidence for monozygotic twin (MZ) discordance in methylation level at two CpG sites in the promoter region of the catechol-O-methyltransferase (COMT) gene. Am J Med Genet B Neuropsychiatr Genet. 2006;141B:421–5. https://doi.org/10.1002/ajmg.b.30316.
Smederevac S, Delgado-Cruzata L, Mitrović D, Dinić BM, Bravo TT, Delgado M, et al. Differences in MB-COMT DNAm in monozygotic twins on phenotypic indicators of impulsivity. Front Genet. 2023;13:1067276. https://doi.org/10.3389/fgene.2022.1067276.
L Phan, Y Jin, H Zhang, W Qiang, E Shekhtman, D Shao, et al. “ALFA: Allele Frequency Aggregator.” National Center for Biotechnology Information, U.S. National Library of Medicine, 10 Mar. 2020, www.ncbi.nlm.nih.gov/snp/docs/gsr/alfa/.
VanderWeele TJ, Vansteelandt S. Mediation analysis with multiple mediators. Epidemiol Methods. 2014;2:95–115. https://doi.org/10.1515/em-2012-0010.
Acknowledgements
We would like to acknowledge Dr. Nicholas Mancuso for his advice on methylation-based software tools for ancestry-based SNP adjustment. Funding was provided by National Institute of Health grants (P50ES026086, P50MD015705, R00 ES030400) and the Environmental Protection Agency Grant (83615801). Additionally, funding was provided by the Southern California Environmental Health Sciences Center (5P30ES007048), and the Dartmouth Cancer Center Support Grant (P30 CA023108).
Author information
Authors and Affiliations
Contributions
Sahra Mohazzab-Hosseinian led the project, conducted analysis, interpreted results, and wrote up the manuscript. Dr. Erika Garcia, Dr. Joseph Wiemels, Dr. Crystal Marconett, Dr. Karina Corona, Dr. Shohreh Farzan, and Dr. Theresa M. Bastain consulted in analysis and assisted in editing and reviewing the manuscript. Dr. Caitlin G. Howe obtained funding for gene expression analyses, and edited and revised the manuscript. Dr. Helen Foley managed lab protocol and edited and revised the manuscript. Dr. Carrie V. Breton obtained funding for the study, reviewed the manuscript, consulted on analysis, and edited the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mohazzab-Hosseinian, S., Garcia, E., Wiemels, J. et al. Effect of parental adverse childhood experiences on intergenerational DNA methylation signatures from peripheral blood mononuclear cells and buccal mucosa. Transl Psychiatry 14, 89 (2024). https://doi.org/10.1038/s41398-024-02747-9
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41398-024-02747-9
