
Abstract
Dietary polysaccharides have long been recognized for their capacity to modulate gut microbiota composition and metabolic activities, making them promising resources for functional food development. In this study, we investigated the effects of Poria cocos (P. cocos) polysaccharides on the structure and metabolism of human gut microbiota using an in vitro fecal fermentation model. Our results revealed that P. cocos polysaccharides were readily utilized by the gut microbiota, as evidenced by a significant decrease in pH and an increase in SCFAs concentrations. Notably, the relative abundance of beneficial bacteria (e.g., Lactobacillus and Bifidobacterium) increased, whereas that of potentially pathogenic taxa (e.g., Escherichia-Shigella and Bilophila) decreased. Furthermore, P. cocos polysaccharides enhanced the production of key microbial metabolites, significantly upregulating compounds such as L-cystine and etelcalcetide. Collectively, these findings underscore the beneficial role of P. cocos polysaccharides in promoting intestinal health and highlight their potential as prebiotics in the functional food industry.
Similar content being viewed by others


Introduction
P. cocos is a functional, edible, and medicinal fungus widely cultivated and utilized in East Asia1,2. It has been extensively used in the production of medicinal, nutritional, and health product formulations. Prolonged consumption of P. cocos reportedly exerts several beneficial effects, including regulating chest qi, promoting diuresis, and invigorating the body3. Additionally, P. cocos is commonly incorporated into a variety of culinary preparations, such as soups, porridges, snacks, and desserts, or consumed directly3,4. In addition, its application in healthcare products has been linked to improvements in sleep quality5. To date, more than 198 bioactive compounds have been identified in P. cocos6. Among these, P. cocos polysaccharide (FL) constitutes approximately 70–90% of the dry weight of its sclerotia, making it the most abundant component of the fungus6.
P. cocos polysaccharides consist of ribose, arabinose, xylose, mannose, glucose, and galactose2. They exhibit a broad spectrum of pharmacological activities, including anti-inflammatory, antioxidant, antitumor, anti-aging, and anti-hyperglycemic properties7,8. Following oral administration, polysaccharides are minimally digested in the small intestine but can act as potential prebiotics for gut microorganisms, thereby shaping the composition and metabolic function of the gut microbiota1. The gut microbiota, the most abundant microbial community in the human body, serves as an important link between dietary factors and diseases. Emerging evidence suggests that gut microbiota dysbiosis is closely associated with the onset and progression of numerous diseases, including obesity9, constipation10, and colitis11. Hence, the structure of the gut microbiota profoundly influences host health, and its imbalance can trigger various pathological conditions12. Consequently, the gut microbiota and its metabolites represent promising targets for disease prevention and treatment13.
Recent studies have highlighted the ability of plant-derived polysaccharides, particularly those obtained from traditional Chinese medicinal plants, to selectively stimulate the proliferation of beneficial gut bacteria, such as Bifidobacterium and Lactobacillus. These polysaccharides promote the growth of beneficial microorganisms and facilitate the production of beneficial metabolites, thereby contributing to normal physiological functions, immune regulation, and disease prevention. For instance, polysaccharides isolated from Gastrodia elata stimulate the growth of probiotics, including Bifidobacterium, Collinsella, Prevotella, and Faecalibacterium, while inhibiting the abundance of pathogenic bacteria, such as Shigella, Dorea, Desulfovibrio, and Blautia. These polysaccharides exhibit significant prebiotic activity and support human health14. Similarly, polysaccharides extracted from Ganoderma lucidum have been shown to alleviate obesity, reduce chronic inflammation, reduce chronic inflammation, and ameliorate insulin resistance in mice models, primarily attributed to gut microbiota modulation. These findings suggest that Ganoderma lucidum polysaccharides are potential prebiotics for managing obesity and its associated complications15. Despite these promising results, the mechanisms by which FL regulates the gut microbiota and its metabolites remain unclear. Elucidating these regulatory pathways is of great significance for promoting host health and preventing disease.
In this study, we utilized 16S rRNA sequencing and non-targeted metabolomics to elucidate the regulatory effects of FL on the gut microbiota and its key metabolites and evaluate its prebiotic potential. These findings are anticipated to provide critical data and a robust theoretical foundation for the development and application of FL in functional foods.
Results and discussion
Changes in pH and SCFAs
The pH changes observed during in vitro fermentation generally reflect the degree of microbial degradation and utilization of dietary polysaccharides. The pH changes during the fermentation of FL are shown in Fig. 1A. The initial pH value of the FL was not significantly different from that of the control. There was no significant change in the pH during the fermentation process in the control. As shown in Fig. 1A, the initial pH of the FL was not significantly different from that of the control, and no significant change was observed throughout fermentation. In contrast, the pH of FL significantly decreased as fermentation progressed, stabilizing after 24 h without further significant variation. The reduction in pH observed in the FL group is presumably related to microbial carbohydrate metabolism and the subsequent accumulation of organic acids, including short-chain fatty acids (SCFAs)16,17. In parallel with the decline in pH, the total SCFA concentration significantly increased in the FL group (Fig. 1B). This indicates that gut microorganisms can metabolize FL to produce SCFAs, particularly with the significant enrichment of acetic and propionic acids (Figure C and D).

A pH changes. B Total SCFAs. C Acetic acid. D Propionic acid. E Butyric acid. Total SCFAs represent the sum of acetic, propionic, and butyric acids. Different lowercase letters indicate significant differences within the same group at different time points (P < 0.05), whereas different uppercase letters indicate significant differences between the FL and control groups at the same time points (P < 0.05).
SCFAs are the major products of the colon fermentation of indigestible carbohydrates18. A previous study demonstrated that the proportions of acetate, propionate, and butyrate in the colon are approximately 57: 22: 2119. It is important to note that SCFAs concentrations and profiles are influenced by multiple factors, including microbial community composition, substrate type and concentration, and fermentation duration20,21. In vitro fermentation experiments have indicated that polysaccharides vary in their decomposition rates and yield distinct SCFAs profiles. For instance, starch degrades more rapidly than xylan, with starch fermentation producing predominantly butyric acid, whereas xylan fermentation mainly yields acetic acid20. Acetic acid is a critical energy source for the brain, heart, and peripheral tissues and is primarily produced by Bifidobacterium and Lactobacillus22,23. The significant increase in acetic acid levels in FL may be related to the elevated relative abundance of these microorganisms (Fig. 4I). As the most abundant SCFA in peripheral circulation, acetic acid can reduce appetite by crossing the blood-brain barrier via central homeostasis mechanisms22 and modulate the expression of regulatory neuropeptides that inhibit appetite and activating acetyl-CoA carboxylase24. Propionic acid has also been shown to reduce food intake, ameliorate fatty acid metabolism25, and enhance insulin sensitivity26.
In summary, in vitro fecal fermentation of FL stimulated the production of beneficial compounds, reducing the pH of the surrounding environment. These SCFAs have beneficial effects on the colon, gut microbiota, and other organs of the body. These findings suggest that FL may confer prebiotic benefits and possess potential health-promoting properties.
Fourier transform infrared spectroscopy
Fourier transform infrared spectroscopy (FT-IR) was used to examine the conformational characteristics of glycosidic bonds and functional groups of FL polysaccharides at different fermentation times (0, 6, 12, 24, and 48 h). As shown in Fig. 2, the absorption peaks near 2928 cm−1 are indicative of C-H stretching vibrations in alkyl groups, a signature commonly associated with polysaccharides27. The peak at approximately 1637 cm−1 corresponds to the bending vibration of the C = O groups28. In addition, the absorption range between 1000 and 1200 cm−1 is attributable to the tensile vibration of the pyran ring29. The presence of both α- and β-glycosidic bonds in FL is suggested by the characteristic absorption peaks observed at approximately 930 cm−1 and 851 cm−130. Notably, an absorption peak, known to be characteristic of glucose, appeared at 774 cm−1 at 12, 24, and 48 h of fermentation31, indicating that FL contains glucose or has been degraded into glucose by intestinal microorganisms. Overall, the FT-IR spectra remained largely consistent throughout the fermentation process, suggesting that although intestinal microorganisms degrade and utilize FL, they exert no significant influence on its structural characteristics.

0 h, 6 h, 12 h, 24 h, and 48 h indicate the FT-IR spectra at different time points of fermentation.
Diversity analysis of gut microbiota
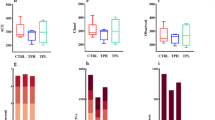
Polysaccharides are vital dietary components that modulate the gut microbiota and improve host health22,32. Sparse curves were generated as an indirect measure of species abundance to determine whether the sequencing amount was sufficient to reflect the species diversity within each sample33. The sparse curves for the Amplicon Sequence Variant (ASV) levels Sobs and Shannon indices (Fig. 3A, B) indicated that the sequencing data were adequate for assessing gut microbial diversity of the samples. The coverage index reflects the coverage of the community, Chao1 and Sobs indices indicate the richness of the community, and Shannon and Simpson indices represent the diversity of the community34. As illustrated in Fig. 3C, the coverage indices for the FL and Control groups were not significantly different and were close to 1, implying that the sequencing results accurately reflected the true microbial composition of the samples. Although no significant differences in richness (Chao1 and Sobs) were observed between the two groups, significant differences in diversity (Shannon and Simpson) were observed (Fig. 3D–G). This suggests that FL influences gut microbiota diversity without markedly affecting species richness. These results are consistent with those of previous studies showing that pectic polysaccharides from okra35 and citrus pectin36 have been shown to significantly reduce intestinal flora diversity. Similarity assessment was performed based on the bray_curtis algorithm at the ASV level (Fig. 3H). All samples were clustered into two distinct groups: FL and control. Principal Component Analysis (PCA) corroborated these findings, demonstrating clear separations in community structure and composition between the two groups, with PC1 and PC2 accounting for 33.59% and 23.71% of the total variance, respectively (Fig. 3I). Collectively, these results indicate that FL significantly affects the structural composition of the gut microbiota.

A Sobs rarefaction curve at the ASV level. B Shannon rarefaction curve at the ASVs level. C Coverage index of ASV levels. D Chao1 index at ASV level. E Sobs index at the ASV level. F Shannon index at the ASV level. G Simpson index at the ASV level. H Hierarchical clustering at the ASV level. I PCA based on ASV levels.
Analysis of gut microbiota composition
The gut microbiota plays diverse and vital roles in energy metabolism, nutrient absorption, and immune regulation32. Elucidating the relationship between FL and gut microbiota can help prevent diseases and promote health by regulating the composition of the gut microbiota. At the phylum level (Fig. 4A–F), the main constituents were Firmicutes, Proteobacteria, Desulfobacterota, Bacteroidota, Actinobacteriota, and a small proportion of Synergistota. Notably, Firmicutes was significantly more abundant in the FL group than in the control group, whereas Proteobacteria and Desulfobacterota was significantly lower than that in the control. A similar increase in Firmicutes abundance has been reported when Symphytum officinale L. polysaccharides were fermented in vitro37. Firmicutes play a vital role in fermenting indigestible carbohydrates to generate SCFAs, which provide energy to the host and modulate metabolic processes, thus promoting host health38. In contrast, Proteobacteria, the predominant phylum of gram-negative bacteria, includes many pathogenic microorganisms that can disrupt the gut microbiota, induce low-grade inflammation, and potentially contribute to colon cancer22,39.

A Community composition between the two groups at the phylum level. B Comparison of relative abundance of Firmicute. C Comparison of the relative abundance of Proteobacteria. D Comparison of relative abundance of Desulfobacterota. E Comparison of relative abundance of Bacteroidota. F Comparison of relative abundance of Actinobacteriota. G Relative abundance between two groups at the genus level. H Heatmap of relative abundance between the two groups at the genus level. I Significant differences in microbial groups between the two groups. J LEfSe was used to compare the gut microbiota between the control and FL groups. K LDA score for taxa differing between the control and FL groups.
At the genus level, stacked histograms and heatmap plots were used to represent the relative abundance and composition of species between the two groups (Fig. 4G, H). It can be visualized from the graphs that the composition of the gut microbiota in the FL was significantly different from that in the control. Megamonas, Parabacteroides, Streptococcus, Faecalibacterium, Lactobacillus, Prevotella, Enterococcus, and Bifidobacterium, etc., were predominant in the FL group, whereas. FL significantly reduced the abundance of potentially harmful bacteria, such as Escherichia-Shigella, Bilophila, Fusobacterium, Dorea, Klebsiella, Peptostreptococcus, Sellimonas. Consistent with these observations, Fig. 4I shows significant alterations in the relative abundances of Megamonas, Escherichia-Shigella, Bilophila, Lachnoclostridium, Lactobacillus, Dialister, Enterococcus, Bifidobacterium, and Prevotella. Fusobacterium, a pathogen that proliferates under nutrient-deficient conditions, was inhibited by FL treatment. This finding aligns with that of Wu et al., who observed that the fermentation of snow chrysanthemum polysaccharides significantly reduced the abundance of Fusobacterium, Klebsiella, and Escherichia-Shigella40. In addition, Megamonas, which belongs to the phylum Firmicutes, can ferment carbohydrates into acetic and propionic acids, which corroborates the SCFA results (shown in Fig. 1) and has been implicated in inflammatory bowel disease and neurological disorders41. Bifidobacteria and Lactobacillus, commonly found in the gastrointestinal tracts of humans and animals, are essential for maintaining host health. For example, Lactobacillus supports gut health by producing a range of SCFAs38 and in vitro fermentation of tamarind seed polysaccharides has been shown to increase both SCFAs and Lactobacillus abundance42. Bifidobacteria are a group of probiotics in the phylum Actinobacteria that are highly beneficial to human intestinal health43. To identify statistically significant taxonomic groups that differed between the FL and control groups, LEfSe combined with linear discriminant analysis (LDA = 4) was used (Fig. 4J, K). In total, 46 characteristic microbial groups at each taxonomic level, ranging from phyla to genera, were detected between the two groups. Among them, 16 microbial groups including Actinobacteria, Bifidobacteriales, Lactobacillales, Megamonas, and Prevotellaceae, were enriched in the FL group. 30 microbial groups exhibiting significant differences were enriched in the control group, including Clostridia, Lachnospiraceae, Lachnospirales, Bilophila, Desulfovibrionia, Enterobacteriaceae, Gammaproteobacteria, etc.
Overall, these findings suggest that FL promotes the proliferation of beneficial gut microbes while inhibiting the growth of potentially pathogenic and harmful bacteria, thereby enhancing the production of beneficial metabolites such as SCFAs. These characteristics are consistent with the concept of prebiotic activity.
Comparative analysis of metabolites of gut microbiota
Microbial metabolites are key mediators of dietary nutritional factors that modulate the gut microbiota and play an essential role in the prevention and amelioration of metabolic disorders, thereby contributing to human health. In the present study, we examined metabolites in the FL fermentation broth. The PCA results (Fig. 5A) demonstrated that the QC samples were well aggregated, with a close distance within the group and significant distances between the groups, with an overall explanation of 84.70%. The PLS-DA analysis results further showed that the metabolites of FL and control were completely distinguished (Fig. 5B), with significant variability between the control and experimental groups. The PLS-DA model validation results indicated an intercept of −0.5571 for the Q2 regression line on the Y-axis, indicating a robust predictive capability and suitability for subsequent analyses (Fig. 5C).

A PCA analysis. B PLS-DA analysis. C Permutation test. D Upset. E HMDB compound classification. F Metabolite clustering analysis was performed.
A comparative analysis of gut microbiota metabolites was conducted in both positive and negative ion modes using FL fermentation broth and a blank group. The results indicated the presence of 3030 and 2965 metabolites in the two groups (n = 3), with 2733 metabolites common between the two groups. Furthermore, 297 metabolites were unique to the FL group and 232 to the blank control group (Fig. 5D). Based on the HMDB 4.0 database, the differential metabolites were categorized into 19 categories (Fig. 5E). Organic acids and their derivatives constituted the largest proportion (27.70%), followed by lipids and lipid-like molecules (23.88%), organic chemical compounds (14.22%), and organic oxygen compounds (9.76%). Cluster analysis revealed distinct metabolite profiles across the groups (Fig. 5F), indicating that FL substantially altered the metabolite profile of the gut microbiota.
Analysis of differential metabolites
Volcano map analysis distinctly illustrated both upregulated and downregulated metabolites, revealing a total of 90 significantly different metabolites between the two groups: 50 upregulated and 40 downregulated (Fig. 6A). The contributions of the metabolites were assessed using the weighting coefficient of the OPLS-DA model, which ranked their contributions based on the VIP score. Accordingly, the top 20 upregulated metabolites and their contribution orders are shown in Fig. 6B, including L-3-cyanoalanine, tetragastrin, etelcalcetide, adenosine 5’-monophosphate, and L-cystine. L-cystine, an amino acid derivative, possesses antioxidant properties that protect against oxidative stress and tissue damage44. Adenosine 5’-monophosphate, a promising anti-obesity agent, has been shown to ameliorate obesity45. Dietary supplementation with adenosine 5’-monophosphate has been shown to modulate lipid metabolism in fattening pigs, enhance meat quality, and promote the formation of flavor compounds, such as glutamine, arginine, glycine, and alanine, in muscle tissue46. Differential metabolite analysis was performed to examine the effects of FL on serum metabolic profiles. The top 20 downregulated metabolites were identified (Fig. 6C), including Phe Leu, P-octopamine, xanthine, and pantothenic acid. Xanthine, a typical purine, undergoes direct metabolism to uric acid via xanthine oxidase. Elevated uric acid levels can promote the formation of sodium urate crystals, which accumulate in the joints, causing inflammation and potentially leading to gout47. Furthermore, FL affected 25 KEGG secondary pathways (Fig. 6D), including six main categories: Metabolism, Human Diseases, and Cellular Processes, etc. Among them, metabolism had the most annotated secondary pathways, particularly amino acid, carbohydrate, and lipid metabolism. Further enrichment analyses of the differentially expressed metabolites (Fig. 6E) indicated the highest enrichment in the pancreatic cancer pathway, followed by cAMP signaling and choline metabolism in cancer pathways. Additional pathways with significant enrichment included renin secretion, antifolate resistance, fat digestion and absorption, Cushing syndrome, phospholipase D signaling, and other metabolic pathways.

A Differential volcano map. B Upregulated metabolite VIP score chart. C Downregulated metabolite VIP score chart. D KEGG functional pathway analysis. E KEGG enrichment analysis. PS PS (20:4(5Z,8Z,11Z,14Z)/20:5(5Z,8Z,10E,14Z,17Z)-OH (12)), PGP PGP (22:6(4Z,7Z,11E,13Z,15E,19Z)-2OH (10S,17)/i-15:0). 6-(4-carboxy-3-hydroxyphenoxy)-: 6-(4-carboxy-3-hydroxyphenoxy)-3,4,5-trihydroxyoxane-2-carboxylic acid, 2,4-Pyrimidinediamine 2,4-Pyrimidinediamine, 5-(5-(1-piperazinylmethyl)-1,3,4-oxadiazol-2-yl)-N4-propyl-N2-(2-(4-pyridinyl) ethyl)-.
Collectively, these findings suggest that the biological activity of FL may be partially attributed to its fermentation by the gut microbiota, which promotes the generation of beneficial metabolites while mitigating harmful metabolites. Accordingly, FL has potential as a prebiotic agent in the intestinal environment.
Correlation analysis
Microbial metabolites are an important link between gut microorganisms and host health. Investigating the associations between intestinal microorganisms and their metabolites provides critical insights into the influence of dietary polysaccharides on gut microbiota and host health. As shown in Fig. 7, the beneficial bacterium Lactobacillus exhibited a significant positive correlation with metabolites such as raffinose, chondroitin 6-sulfate, panose, core oligosaccharide, maltotriose, L-cystine, and etelcalcetide. Similarly, Bifidobacterium abundance was positively correlated with etelcalcetide, adenosine 5’-monophosphate, tetragastrin, L-cystine, and chondroitin 6-sulfate. Raffinose, a trisaccharide composed of D-fructose, D-galactose, and D-glucose, promotes the proliferation of Lactobacillus and Bifidobacterium, highlighting its potential as a prebiotic48. Etelcalcetide is a novel calcium mimetic used to treat secondary hyperparathyroidism by directly binding to calcium-sensitive receptors, thereby reducing circulating parathyroid hormone and calcium levels49.

Red represents positive correlation and green represents negative correlation.*represents significant positive/negative correlation. Note: (Z)-7-: (Z)-7-[(1R,3S,4S,5R)-3-[(E,3R)-3-Hydroxyoct-1-enyl]-6-thiabicyclo[3.1.1]heptan-4-yl]hept-5-enoic acid. Quillaic acid 3-: Quillaic acid 3-[galactosyl-(1-2)-[rhamnosyl-(1-3)]-glucuronide] 28-[apiosyl-(1-3)-xylosyl-(1-4)-rhamnosyl-(1-2)-[rhamnosyl-(1-3)]-4-acetyl-fucosyl] ester.
Research indicates that panose and chondroitin 6-sulfate both displayed negative correlations with harmful bacteria, such as Escherichia-Shigella, Bilophila, Peptostreptococcus, and Klebsiella, while showing positive correlations with beneficial bacteria, including Megamonas, Lactobacillus, Parabacteroides, and Faecalibacterium. Panose has antioxidant and antidiscoloration effects and is recognized as a functional sugar with diverse biological properties50. Additionally, it serves as a prebiotic that can modulate the balance of the gut microbiota51. In contrast, the metabolite Xanthine demonstrated significant positive correlations with harmful bacteria such as Bilophila, Lachnoclostridium, Escherichia-Shigella, Dialister, Klebsiella, and Peptostreptococcus, while displaying negative correlation with beneficial bacteria such as Megamonas, Lactobacillus, Parabacteroides, and Faecalibacterium. Xanthine is generated through the deamination of guanine by guanine deaminase and the oxidation of hypoxanthine by xanthine oxidase. Subsequently, xanthine is further oxidized by xanthine oxidase to form uric acid52, whose excessive accumulation leads to hyperuricemia, a condition that has become the second most prevalent metabolic disorder after diabetes mellitus53, posing a substantial threat to human health and quality of life. In summary, FL is closely related to the intestinal flora and its associated metabolites. By modulating intestinal flora, FL stimulates the production of microbial metabolites, thereby exerting a profound influence on host health.
P. cocos polysaccharide is a polysaccharide with both therapeutic and nutritional potential, capable of modulating the composition of intestinal microbiota and associated metabolites during human fecal fermentation. At the phylum level, FL significantly increased the relative abundance of Firmicutes and decreased the relative abundance of Proteobacteria and Desulfobacterota. At the genus level, FL promoted the proliferation of beneficial bacteria and suppressed harmful taxa. Moreover, it enhances the production of SCFAs, such as acetic and propionic acids, thereby lowering environmental pH and maintaining intestinal homeostasis. In addition, FL significantly enriched beneficial metabolites, including L-cystine, etelcalcetide, and adenosine 5’-monophosphate, while diminishing harmful metabolites such as xanthine. Correlation analyses further revealed that these changes in intestinal flora were closely aligned with alterations in metabolite profiles.
Overall, these findings underscore the potential of FL to remodel the intestinal microbiota and regulate key metabolites, offering significant promise for improving host health. Consequently, FL exhibits prebiotic properties that may prove valuable for developing functional foods designed to enhance human well-being.
Materials and methods
Materials and reagents
P. cocos polysaccharides (90% purity) were obtained from Xi’an Kepin Biotechnology Co., Ltd. Short-chain fatty acid (SCFA) standards were purchased from Macklin Biochemical Technology Co., Ltd. (Shanghai, China).
In vitro fermentation
The culture medium was prepared as previously described protocols54,55. The pH was adjusted to 7.0 ± 0.2, and the medium was sterilized at 121 °C for 15 min. Fresh fecal samples were collected from four healthy volunteers (n = 4, male: female =1:1, aged 20−30) who maintained a healthy diet, were in good health, and had not used antibiotics, probiotics, or prebiotics in the previous 6 months. Informed consent was obtained from all participants, and each participant completed a questionnaire survey. The study was supervised and approved by the Institutional Review Board of the College of Food Science, Southwest University. Fecal inoculum was prepared as previously described55,56.
In vitro fermentation of FL was performed as described previously56,57. Briefly, 1% (w/v) FL was dissolved in the fermentation medium, and the medium without FL was used as a control. After pre-warming at 37 °C, the medium was inoculated with a 10% (v/v) human fecal homogenate, mixed thoroughly, and incubated at 37 °C under anaerobic conditions. Each group was prepared in triplicates. Samples were collected at different time points, and the pH was determined. All samples were stored at −80 °C until further analysis.
FT-IR analysis
FT-IR was used to examine the changes in infrared spectra during fermentation. The supernatant of the fermentation broth was subjected to freeze-drying, and 2 mg samples were weighed for KBr tableting, with a scanning range of 4000-400 cm-158.
Determination of SCFAs
SCFAs were extracted using anhydrous ether following a previously described method59. The determination of SCFAs was based on the previously described method60 with slight modifications. The detector temperature was set to 250 °C, the sample injection volume was 0.5 µL, and the split ratio was 12:1. The heating program was as follows: 100 °C for 0.5 min, increased to 160 °C at 4 °C/min, and then raised to 220 °C at 10 °C/min. All analyses were performed using an Agilent 8890-5977B gas chromatograph.
Analysis of gut microbiota
After 48 h of fermentation, the precipitate was collected, and total DNA was extracted using the YH-feces stool DNA extraction kit (Shanghai Meiji Suhua Biomedical Technology Co., Ltd.). The V3-V4 region of the 16S rRNA was amplified using the primers 338 F (ACTCCTACGGGAGGCAGCAG) and 806 R (GGACTACHVGGGTWTCTAAT)61. Sequencing was performed on the Illumina PE300 platform, and the resulting data were processed using the Majorbio Cloud platform (https://cloud.majorbio.com).
Gut microbiota metabolites analysis
Fermentation supernatant (200 µL) was precisely pipetted, and 200 µL of the extraction solution was added to extract the metabolites. The sample injection volume was 3 μL, the flow rate was 0.40 mL/min, and the column temperature was maintained at 40 °C. Following completion of the computer operation, the LC-MS raw data were imported into the metabolomics processing software Progenesis QI (Waters Corporation, Milford, USA) for handing. Simultaneously, MS and MS/MS information was cross-referenced with the metabolic public databases HMDB (http://www.hmdb.ca/), Metlin (https://metlin.scripps.edu/), and Majorbio self-built databases, and metabolite information was obtained from these databases. All data analyses were performed using the Majorbio Biomedical Cloud Platform (https://cloud.majorbio.com).
Statistical analysis
GraphPad Prism 9.0.2 was used for drawing and data representation. The mean ± SD was calculated for all experiments, with three replicates, and statistical significance was set at P < 0.05. Statistical analyses were performed using SPSS 20.0. The correlation between microorganisms and differential metabolites was analyzed using Spearman’s rank correlation test62. Additionally, the Chiplot (https://www.chiplot.online/) tool was used to draw a correlation heatmap.
Data availability
Data will be made available on request.
References
Duan, Y. et al. Poria cocos polysaccharide improves intestinal barrier function and maintains intestinal homeostasis in mice. Int. J. Biol. Macromol. 249, 125953 (2023).
Cheng, Y. et al. Structural characterization and hepatoprotective activity of a galactoglucan from poria cocos. Carbohydr. Polym. 263, 117979 (2021).
Wang, Q., Yang, K., Wei, X., Qiao, W. & Chen, L. Untargeted metabolomics analysis reveals dynamic changes in co-fermentation with human milk-derived probiotics and poria cocos. Front. Microbiol. 13, 1032870 (2022).
Jin, J. et al. Insights into triterpene acids in fermented mycelia of edible fungus poria cocos by a comparative study. Molecules 24, 1331 (2019).
Kim, H., Choi, H., Park, B., Ju, H. & Kim, Y. Efficacy of poria cocos extract on sleep quality enhancement: A clinical perspective with implications for functional foods. Nutrients 15, 4242 (2023).
Lu, J. et al. Phytochemistry and biological activities of poria. J. Chem. 2021, 1–20 (2021).
Li, X. et al. Molecular basis for poria cocos mushroom polysaccharide used as an antitumour drug in China. J. Cell. Mol. Med. 23, 4–20 (2019).
Nie, A. et al. Phytochemistry and pharmacological activities of wolfiporia cocos (f.a. Wolf) ryvarden & gilb. Front. Pharmacol. 11, 505249 (2020).
Zhang, Z. et al. FOS/GOS attenuates high-fat diet induced bone loss via reversing microbiota dysbiosis, high intestinal permeability and systemic inflammation in mice. Metab. -Clin. Exp. 119, 154767 (2021).
Zhou, X. et al. Efficacy of Bacillus coagulans BC01 on loperamide hydrochloride-induced constipation model in kunming mice. Front. Nutr. 9, 964257 (2022).
Wang, M. et al. Traditional chinese medicine: a promising strategy to regulate the imbalance of bacterial flora, impaired intestinal barrier and immune function attributed to ulcerative colitis through intestinal microecology. J. Ethnopharmacol. 318, 116879 (2024).
Wilmes, P. et al. The gut microbiome molecular complex in human health and disease. Cell Host Microbe 30, 1201–1206 (2022).
Arifuzzaman, M. et al. Inulin fibre promotes microbiota-derived bile acids and type 2 inflammation. Nature 611, 578–584 (2022).
Gan, Q. et al. Digestive characteristics of gastrodia elata blume polysaccharide and related impacts on human gut microbiota in vitro. J. Ethnopharmacol. 328, 118064 (2024).
Chang, C. et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 6, 7489 (2015).
Fu, X. et al. Structural characterization and in vitro fermentation of a novel polysaccharide from sargassum thunbergii and its impact on gut microbiota. Carbohydr. Polym. 183, 230–239 (2018).
Guo, C. et al. Crataegus pinnatifida polysaccharide alleviates colitis via modulation of gut microbiota and scfas metabolism. Int. J. Biol. Macromol. 181, 357–368 (2021).
Koh, A., De Vadder, F., Kovatcheva-Datchary, P. & Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 165, 1332–1345 (2016).
Cummings, J. H., Pomare, E. W., Branch, W. J., Naylor, C. P. & Macfarlane, G. T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 28, 1221–1227 (1987).
Macfarlane, S. & Macfarlane, G. T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 62, 67–72 (2003).
Dalile, B., Van Oudenhove, L., Vervliet, B. & Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 16, 461–478 (2019).
Zhang, X. et al. In vitro fermentation of gracilaria lemaneiformis sulfated polysaccharides and its agaro-oligosaccharides by human fecal inocula and its impact on microbiota. Carbohydr. Polym. 234, 115894 (2020).
Xu, S. et al. Microbial catabolism of porphyra haitanensis polysaccharides by human gut microbiota. Food Chem. 289, 177–186 (2019).
Sun, P., Sun, D. & Wang, X. Effects of scutellaria barbata polysaccharide on the proliferation, apoptosis and emt of human colon cancer ht29 cells. Carbohydr. Polym. 167, 90–96 (2017).
Wang, M. et al. Purified fraction of polysaccharides from fuzhuan brick tea modulates the composition and metabolism of gut microbiota in anaerobic fermentation in vitro. Int. J. Biol. Macromol. 140, 858–870 (2019).
Al-Lahham, S. H., Peppelenbosch, M. P., Roelofsen, H., Vonk, R. J. & Venema, K. Biological effects of propionic acid in humans; Metabolism, potential applications and underlying mechanisms. BBA-Mol. Cell Biol. L 1801, 1175–1183 (2010).
Su, Y. & Li, L. Structural characterization and antioxidant activity of polysaccharide from four auriculariales. Carbohydr. Polym. 229, 115407 (2020).
Jia, X. et al. Purification, structural characterization and anticancer activity of the novel polysaccharides from rhynchosia minima root. Carbohydr. Polym. 132, 67–71 (2015).
Dong, X., Zhu, C., Huang, G. & Xiao, J. Fractionation and structural characterization of polysaccharides derived from red grape pomace. Process Biochem 109, 37–45 (2021).
Jiang, P. et al. Structural characteristics, antioxidant properties and antiaging activities of galactan produced by mentha haplocalyx briq. Carbohydr. Polym. 234, 115936 (2020).
Zhang, L. et al. Characterization and antioxidant activities of polysaccharides from thirteen boletus mushrooms. Int. J. Biol. Macromol. 113, 1–7 (2018).
Chen, L. et al. Simulated digestion and fermentation in vitro by human gut microbiota of polysaccharides from helicteres angustifolia l. Int. J. Biol. Macromol. 141, 1065–1071 (2019).
Seidling, W. et al. Comparing observer performance in vegetation records by efficiency graphs derived from rarefaction curves. Ecol. Indic. 109, 105790 (2020).
Ding, Y. et al. In vitro digestion under simulated saliva, gastric and small intestinal conditions and fermentation by human gut microbiota of polysaccharides from the fruits of lycium barbarum. Int. J. Biol. Macromol. 125, 751–760 (2019).
Wu, D. et al. In vitro digestion and fecal fermentation behaviors of a pectic polysaccharide from okra (abelmoschus esculentus) and its impacts on human gut microbiota. Food Hydrocoll. 114, 106577 (2021).
Mao, G. et al. Depolymerized rg-i-enriched pectin from citrus segment membranes modulates gut microbiota, increases scfa production, and promotes the growth of Bifidobacterium spp., Lactobacillus spp. and Faecalibaculum spp. Food Funct. 10, 7828–7843 (2019).
Cao, Z. et al. Ultrasonic enzyme-assisted extraction of comfrey (symphytum officinale l.) polysaccharides and their digestion and fermentation behaviors in vitro. Process Biochem 112, 98–111 (2022).
Li, Q. et al. In vitro fecal fermentation characteristics of bamboo shoot (phyllostachys edulis) polysaccharide. Food Chem. X 11, 100129 (2021).
Huang, F. et al. In vitro digestion and human gut microbiota fermentation of longan pulp polysaccharides as affected by Lactobacillus fermentum fermentation. Int. J. Biol. Macromol. 147, 363–368 (2020).
Wu, D. et al. Dynamic changes of structural characteristics of snow chrysanthemum polysaccharides during in vitro digestion and fecal fermentation and related impacts on gut microbiota. Food Res. Int. 141, 109888 (2021).
Zou, R. et al. Changes in the gut microbiota of children with autism spectrum disorder. Autism Res. 13, 1614–1625 (2020).
Li, X. et al. Dynamic digestion of tamarind seed polysaccharide: Indigestibility in gastrointestinal simulations and gut microbiota changes in vitro. Carbohydr. Polym. 239, 116194 (2020).
Han, R. et al. In vitro digestibility and prebiotic activities of a sulfated polysaccharide from gracilaria lemaneiformis. J. Funct. Food 64, 103652 (2020).
Dai, W. & Chen, Q. M. Fresh medium or l-cystine as an effective nrf2 inducer for cytoprotection in cell culture. Cells 12, 291 (2023).
Pardo, F. et al. Molecular implications of adenosine in obesity. Mol. Asp. Med. 55, 90–101 (2017).
Rao, S. et al. Effects of dietary adenosine and adenosine 5’-monophosphate supplementation on carcass characteristics, meat quality, and lipid metabolism in adipose tissues of finishing pigs. Meat Sci. 201, 109174 (2023).
Liu, F., Shang, T., Han, L., He, N. & Liu, X. Electrochemical investigation on kinetics of xanthine metabolism and inhibition effect of febuxostat on xanthine oxidase activity. Electroanalysis 35 (2023).
Mao, B. et al. In vitro fermentation of raffinose by the human gut bacteria. Food Funct. 9, 5824–5831 (2018).
Blair, H. A. Etelcalcetide: First global approval. Drugs 76, 1787–1792 (2016).
Wang, M. et al. Discovery of a new microbial origin cold-active neopullulanase capable for effective conversion of pullulan to panose. Int. J. Mol. Sci. 23, 6928 (2022).
Makelainen, H., Hasselwander, O., Rautonen, N. & Ouwehand, A. C. Panose, a new prebiotic candidate. Lett. Appl. Microbiol. 49, 666–672 (2009).
Maiuolo, J., Oppedisano, F., Gratteri, S., Muscoli, C. & Mollace, V. Regulation of uric acid metabolism and excretion. Int. J. Cardiol. 213, 8–14 (2016).
Zhao, R. et al. Engineered Escherichia coli nissle 1917 with urate oxidase and an oxygen-recycling system for hyperuricemia treatment. Gut Microbes 14, 2070391 (2022).
Mao, B. et al. In vitro fermentation of lactulose by human gut bacteria. J. Agric. Food Chem. 62, 10970–10977 (2014).
Zhou, X. et al. In vitro fermentation characteristics of fucoidan and its regulatory effects on human gut microbiota and metabolites. Food Chem. 465, 141998 (2025).
Bai, J. et al. Systematic assessment of oat β-glucan catabolism during in vitro digestion and fermentation. Food Chem. 348, 129116 (2021).
Sayar, S., Jannink, J. & White, P. J. Digestion residues of typical and high-β-glucan oat flours provide substrates for in vitro fermentation. J. Agric. Food Chem. 55, 5306–5311 (2007).
Ma, Y., Jiang, S. & Zeng, M. In vitro simulated digestion and fermentation characteristics of polysaccharide from oyster (crassostrea gigas), and its effects on the gut microbiota. Food Res. Int. 149, 110646 (2021).
Wang, L. et al. Bifidobacteria exert species-specific effects on constipation in BALB/C mice. Food Funct. 8, 3587–3600 (2017).
Hu, J., Nie, S., Li, C. & Xie, M. In vitro fermentation of polysaccharide from the seeds of plantago asiatica l. by human fecal microbiota. Food Hydrocoll. 33, 384–392 (2013).
Liu, C. et al. Denitrifying sulfide removal process on high-salinity wastewaters in the presence of halomonas sp. Appl. Microbiol. Biotechnol. 100, 1421–1426 (2016).
Zhang, X. et al. Effects of in vitro fermentation of atractylodes chinensis (DC.) Koidz. polysaccharide on fecal microbiota and metabolites in patients with type 2 diabetes mellitus. Int. J. Biol. Macromol. 253, 126860 (2023).
Acknowledgements
This work was supported by Major Science and Technology Special Projects in Tibet Autonomous Region (No. XZ202201ZD0001N), the Technology Innovation and Application Development Special Project of Chongqing, China (CSTB2024TIAD-LUX0002), Fundamental Research Funds for the Central Universities (SWU-KQ22050 and SWU-KQ22076), University Innovation Research Group in Chongqing (No. CXQT21007), Key Construction Disciplines of Traditional Chinese Medicine in Chongqing (2021-4322190044). All authors thank Dr. Yan Li from Analytical & Testing Center of Southwest University for making the GC-MS 8890-5977B (Agilent Technologies, Santa Clara, CA, USA) available and for assistance in the determination of SCFAs.
Author information
Authors and Affiliations
Contributions
X. Zh and Y.Y. L. Designed, Methodology, Software, Writing-original draft, Writing-review & editing. Y.H. Y., L.W., J.H. X. Methodology. Ch. W. Conceptualization, Writing-original draft. J.J. S. Conceptualization, Investigation. Sh.J. L. Investigation. J. Y. B. Conceptualization, Supervision, Validation. H.Y. S. Funding acquisition, Supervision, Writing-original draft, Writing-review & editing. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhou, X., Li, Y., Yang, Y. et al. Regulatory effects of Poria cocos polysaccharides on gut microbiota and metabolites: evaluation of prebiotic potential. npj Sci Food 9, 53 (2025). https://doi.org/10.1038/s41538-025-00416-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41538-025-00416-9
