
Abstract
The fall armyworm Spodoptera frugiperda (J.E. Smith) is a new serious destructive and widespread pest of corn which recently invaded subtropical regions worldwide. As this invasive species is spreading across the continent, it is vital to assess its susceptibility to currently used insecticides and establish the potential underlying resistance mechanism to better inform control programmes. In this study, we characterized the strains from eighteen fall armyworm populations from different countries in West Africa, established their susceptibility profiles to the main insecticides and genotyped the target site resistance alleles. The RFLP-PCR method showed that the majority of the population tested was a corn strain (72.5–100%) compared to the probable rice strain (0–27.5%). Tpi sequencing of the suspected rice strains revealed that almost all the samples analysed were from corn (> 97%). Additionally, the three insecticides tested, λ-cyhalothrin, chlorpyrifos and emamectin benzoate induced susceptible to moderate toxicity against this pest. Synergism tests performed to investigate the biochemical mechanism used by fall armyworm to breakdown λ-cyhalothrin indicated that metabolic enzymes (oxydases, esterase and the glutathion-S-transferase) play moderate roles in the resistance of λ-cyhalothrin observed in western Africa. Target mutation tests (qPCR) combined with previous synergetic tests showed that resistance to organophosphates and pyrethroids could be due to a biochemical mechanism + amino acid mutations (presence of the F290V mutation) or a biochemical mechanism (absence of the T929I mutation), respectively. The results of this study provide valuable information for supporting decisions related to sustainable fall armyworm control and applied resistance management.
Similar content being viewed by others

Introduction
Since 2016, the fall armyworm Spodoptera frugiperda(J.E. Smith) (Lepidoptera: Noctuidae) has become the world’s most important new invasive insect due to its rapid migration and the number of countries invaded in short time, raising socio-economic concerns about its impact on agricultural production1. Gorgen et al.2 and Otim et al.3were the first to report the migration of this species in Africa. Later, it spread throughout Africa and reached Asia, New Caledonia and Oceania4,5,6. This pest is also important because of its high reproductive capacity and large number of host plants, 353 species in 76 plant families, which include several economically important crops, such as corn, rice, sorghum, sugar cane, cotton and soybean1,7,8,9.
Based on the preferred host plants, two sympatric fall armyworm strains were reported: the corn strain (C-strain), which feeds on large grasses such as corn, sorghum (Sorghum bicolor) and cotton, and the rice strain (R-strain), which prefers small grasses such as rice, alfalfa, sugar cane and Johnson grass (Sorghum halepense)10,11. The taxonomic and genetic status of the two fall armyworm host strains remain uncertain and controversial. Genomic studies have been similarly inconclusive with reports for both the presence10and absence12of significant nuclear genomic differences between strains. However, strain-specific genetic markers, including segments of the mitochondrial cytochrome oxidase subunit I (COI) gene and the Z-chromosome Tpi gene, which codes for the domestic enzyme triosephosphate isomerase, have demonstrated strong correlations between genetic polymorphisms and host plants in fall armyworm in several locations in the Western Hemisphere13,14,15. According to the Lepidoptera ZW/ZZ sex determination system, fall armyworm males carry two copies of the Tpi gene, making it possible to obtain TpiC/TpiR heterozygotes, which are indicative of potential interstrain hybrids15,16. However, some laboratory studies on strain identification using the Tpi gene revealed partial intermating and consequently, significant barriers to interstrain hybridization17,18, proving that Tpi is an accurate marker of strain identity19,20. It should also be noted that physiology, sensitivity to insecticides and detoxification genes were other models (criteria) that differed between the two strains10,21,22.
According to Eschen et al.23, the potential economic losses caused by uncontrolled fall armyworm infection (insecticide resistance) are estimated to reach up to US$9.4 billion annually in Africa, corresponding to losses ranging from 11–58%23,24,25. Faced with the rapid invasion of the armyworm and the slowness of suggested management strategies, chemical control is the main method of pest management in Africa. However, treatment failures are observed by smallholders. To identify the best integrated pest management strategy for armyworms, chemical pesticides, which include more than 57 active ingredients with nine modes of action have been used. Among which, 34 active ingredients were found in fall armyworm-invading regions in Africa7,26. The genetic plasticity, high fecundity and in particular the intensive selection pressure of this pest leads to fall armyworm exhibiting resistance to main insecticide classes such as; organophosphates (chlorpyriphos)27,28,29, carbamates (carbaryl)28,30, pyrethroids (λ-cyhalothrin)27,31, spinosyns (spinosad)32, diamides (chlorantraniliprole)33,34 and genetically engineered crops expressing Bacillus thuringiensis(Bt) toxins27,35. In addition to the intrinsic variation in insecticide susceptibility associated with fall armyworm strains, insecticide resistance will depend on the preexistence of resistant alleles in the starting population and selection pressure on the newly invaded areas and spread19,36. Pyrethroids and organophosphates constitute the main insecticides used worldwide by smallholders because of their low application cost and low toxicity to humans and the environment. This is justified by the high number of compounds registered for decades and their frequent application37,38. However, farmers have reported treatment failures for pyrethroids and organophosphates in the field4,29, thereby leading to the overuse of pyrethroids by increasing concentrations, application frequencies or even the use of prohibited or unregistered compounds39,40. In contrast, the efficacy of emamectin benzoate, which acts on glutamate-gated chloride channel allosteric modulators has been proven in recent studies for lepidopteran pest control in West Africa and within different agronomic settings7,28,31.
Therefore, screening the resistance pattern of this pest is crucial for enhanced integrated pest management (IPM) strategies. Previous studies have highlighted target site mutations and biochemical mechanisms as the main resistance mechanisms against insecticides41. Observations by Carvalho et al.42 and Boaventura et al.27have underscored the target-site insensitivity to organophosphate (chlorpyrifos) and pyrethrinoid (λ-cyhalothrin). Three amino acid substitutions, A201S, G227 A, F290 V and T929I, L932 F, L1014 F, were detected in the genes encoding acetylcholinesterase (AChE) and voltage-gated sodium channel (VGSC) respectively43,44. In addition to the lack of information on the mapping of resistance mechanisms, no research has attempted to provide information on the genetic diversity and phenotypic profiles of resistance observed in several cropping systems across West Africa. In the present research, the susceptibility profiles of three insecticides were determined in fall armyworm populations after genotyping. Enzyme inhibitor synergy tests highlighting possible mechanisms of metabolic resistance and amino acid mutation frequencies of genes encoding AChE, VGSC, the target site of organophosphates (OPs), carbamates and pyrethroids were studied. The information generated will help to assess the risk to corn and rice strains in Africa and improve decision-making on the type of insecticides used for fall armyworm control across West Africa.
Results
Molecular characterization of the S. frugiperda strain
RFLP-PCR was performed with the mitochondrial primers 891 F-COI/C1303R-COI and JM76/JM77 digested with the EcoRV and MspI enzymes respectively, to identify sympatric strains of the C-strain and R-strain of the fall armyworm according to the protocol of Bonaventura et al.27. The PCR product obtained with the COI was 450 bp for both strains and contained the EcoRV restriction site for the R-strain (150 and 300 bp), while the C-strain fragment remained uncut. Similarly, when the JM76/JM77 primer was used to amplify the same DNA samples, the resulting PCR product was approximately 569 bp in length for both strains. The C-strain contained an MspI restriction site (approx. 487 and 72 bp) and the R-strain was uncut (Fig. 1). Using this method, 12.7%, 18.9%, 50.0%, 17.4% and 00.0% of the R-strains were obtained from fall armyworms collected from maize, sorghum, rice, onion and cabbage respectively in Benin. For the fall armyworms collected from maize in Niger, Togo and Mali 17.2%, 27.5% and 00.0% of the R-strains were obtained respectively. Altogether, 86.4%, 82.8%, 72.5% and 100% of the C-strains were obtained for all the samples tested in Benin, Togo, Niger and Mali respectively (Table S1).

Agarose gel electrophoresis showing PCR products (891 F-COI/C1303R-COI and JM76/JM77) digested with the restriction enzymes EcoRV and MspI, respectively, used for the identification of Spodoptera frugiperda host plant strains in field samples collected from Benin, Togo, Niger and Mali. M: molecular weight markers. [1–7] The PCR products amplified with the primers 891 F_COI and c1303R_COI (Table S3) did not contain an EcoRV-specific site, and the PCR products amplified with the JM76 and JM77 primers (Table S3) contained the FastDigest MspI-specific site. The C-strain was uncut with EcoRV and cut with MspI using the following primers (891 F_COI and c1303R_COI and JM76 and JM77). [8–18] The above PCR product after digestion contained a specific EcoRV site that was amplified using the 891 F_COI and c1303R_COI primers (Table S3) and was below the products obtained after digestion with FastDigest MspI (no specific site). The R-strain was cut with EcoRV and uncut with MspI using the following primers (891 F_COI and c1303R_COI and JM76 and JM77). 1 = FOuiMaD2; 2 = FAthMaD3; 3 = FGamMaD5; 4 = FMadMaA9; 5 = FAveMaA1; 6 = FAdkMaA6; 7 = FMalMaA2; 8 = FGpOiD7; 9 = FAgbMaD2; 10 = FAgaMaD17; 11 = FAthSoD15; 12 = FKouRiD1; 13 = FGpoOiD18; 14 = FAdkMaD9; 15 = FAmoMaD14; 16 = FDjiMaA11; 17 = FDjiMaD7; 18 = FMadMaA16.
Sequencing of Tpi for confirmation of molecular identification and genetic diversity analysis
Correlation between RFLP-PCR and sanger sequencing
In total, 75 samples found as R-strains after PCR RFLP were successfully sequenced and analysed. Of the 75 samples, only 01 (1.33%) was confirmed to belong to the R-strain, whereas the remaining were C-strain (97.33%) (Fig. 2e). These findings indicate that the PCR RFLP method is not suitable for differentiating between corn and rice strains.
Genetic diversity
A 303 bp fragment spanning the third and fourth exons (TpiE3; E4) of the coding region in the full-length nuclear and sex-linked Tpi gene was successfully amplified and sent for sequencing in 88 Fall armyworms from Benin (n=44), Togo (n=21) and Niger (n=23), 75 of which were successfully analysed (2n= 150). The genetic diversity parameters of the full fragments of the tpi sequences are given in Table 1 according to the different countries, the strains and host plants on which the pests were collected. Overall, 19 polymorphic sites defining 12 haplotypes were detected. A lower number of polymorphic sites (S=3) and a reduced haplotype diversity (H=3 haplotypes; hd= 0.3) were detected in the pests from Benin than in the other locations. Pests collected from maize were more polymorphic (H=12 haplotypes; hd= 0.5) than those collected from other host plants. Accordingly, the C-strain was more polymorphic than the R-strain, probably due to the very low number of R-strains. The overall haplotypes and nucleotides diversity were 0.542 and 0.006 respectively, with no significant differences between the sequences examined (p > 0.1) (Table 1). Assessment of the demographic expansion of populations based on neutrality tests gave a negative value for Tajima’s D (− 1.223), indicating that the populations under study were expanding (abundant rare alleles).

Genetic diversity parameters of Tpi in S. frugiperda collected in West Africa in relation to the host plant strain. (a) Phylogenetic tree and (b) haplotype network (using a maximum likelihood method with phase sequences) according to localities; (c) phylogenetic tree and (d) haplotype network (using a maximum likelihood method with phase sequences) according to host plants; (e) phylogenetic tree (using a maximum likelihood method with unphase sequences) according to strains; (f) haplotype network (using a maximum likelihood method with phase sequences) according to strains.
Distribution of haplotypes and phylogeny
The analysis involved 150 nucleotide sequences of 303 bp plus three reference nucleotide sequences, two for S. frugiperda(TpiCa1 and TpiCa2) from India45 and one for S. frugiperda(TpiRa1) from China46. A clear segregation of the two Spodoptera sub-species (bootstrap 1000) was observed. One cluster included S. frugiperda (TpiCa1 and TpiCa2) from Benin, Togo and Niger on different host plants (maize, sorghum, rice and onion), and the other cluster included S. frugiperda (TpiRa1) from only Togo (Fig. 2). Analysis of the haplotype network of the TPi gene based on locality, host plant and strain showed that there were 12 major haplotypes (Fig. 2) and mutations (21) between the haplotypes (Table 2) in these field populations. In general, the haplotypes of each C-strain (TpiCa1 and TpiCa2) were strongly related to a maximum of 1–7 mutational steps. The H10, H11 and H12 haplotypes were found only in Togo (TpiRa1) and were different from the C-strain reference sequence with 14 mutational steps (Table 2). When comparing the haplotypes of the two sub-species, 21 mutational steps were recorded, confirming the segregation of these species (Table 2; Fig. 2).
Among the five major haplotypes, the ancestor haplotype (H1) which had the highest frequency (41), belonged to the C-strain and was shared between all the countries and all the host plants. H7, the second major haplotype was specific to the C-strain and was specific to Niger and Togo, whereas the third (H5), which was also specific to corn, was present in Togo and Benin only (Fig. 2a, b). The phylogenetic tree constructed with TpiE3;E4 sequences clearly segregated the individuals of the two species, thus confirming their divergence; however, within each species, the main related branches of the tree of several different profiles were obtained and are indicative of intraspecific variation. The analysis of the maximum likelihood phylogenetic tree between the identified haplotypes did not reveal any haplotype clustering associated with localities or host plants but only between the rice and corn stains (Fig. 2).
Bioassays
The susceptibility profiles of fall armyworms in the study countries were performed with the main insecticides used by farmers (λ-cyhalothrin, chlorpyrifos and emamectin benzoate). The results of the susceptibility test performed with λ-cyhalothrin showed that the LD50values (44. 54–482.7 µg·ml−1) varied moderately with Benin population and slight variation was observed with Niger population (1394.61–1039.83 µg·ml−1) and Togo population (146.32–344.96 µg·ml−1). The LD50value obtained for the Mali population was 339.03 µg·ml−1 (Table 3). Low to moderate variation of RRs was observed with chlorpyrifos bioassay in the three tested populations. The most RRs variation was observed in Togo population ranging from 1.46 to 51.16 fold. However, in Benin and Niger, the susceptibility profile to chlorpyrifos varied respectively between 3.56 to 12.07 fold and 5.33 to 12.97 fold respectevely (Table 3). According to the results of the emamectin benzoate bioassay, the least DL50 variation (3.55–6.30 µg·ml−1) was observed between the three tested populations in Benin. Contrary to the benin population, a significante variation was observed in Togo (0.8–16.65) and Niger (0.52–12.68) for Togo and Niger populations respectively (Table 3).
Synergist assessment
The synergistic effects of PBO, DEF and DEM on the toxicity of λ-cyhalothrin were evaluated in several field populations in four countries (Table 4). In Benin, after combining the PBO + λ-cyhalothrin, the synergist ratio was around 1 fold in the four localities (Sagon, Agbangnizoun, Ouidah and Agatha) but the SRs were 5.47 and 6.07 in Zinvié and Athiémè respectively. Accoording to DEM + λ-cyhalothrin, the SRs were 2.38; 1.37 and 1.20 in Agbangnizoun, Agatha and Zinvié respectively. However, the DEF + λ-cyhalothrin was not taken account because of the samples size. In Togo, the combinaision of PBO and λ-cyhalothrin showed that the SRs varied slightly from 1.14 to 1.47 in the tested localities. Concerning DEF + λ-cyhalothrin, the SRs varied also slightly from 1.09 to 2.09. According to DEM + λ-cyhalothrin, highly variation (1.07 to 3.18) was observed in all prospeted localities (Table 4). In Niger, when we associate PBO and λ-cyhalothrin, we observed that the SRs varied between 1.17 and 5.56. About DEF + λ-cyhalothrin and DEM + λ-cyhalothrin combinaisons, the SRs varied from 1.16 to 4.84, and 1.08 to 3.77 respectively (Table 4). The combinaision of PBO, DEF and DEM with λ-cyhalothrin in Mali showed that the SRs were 4.59; 2.01 and 1.75 respectively. The observed synergistic activity suggested that metabolic enzymes (cytochrome P450 s, esterases and GSTs) are moderately involved in the resistance of armyworms to λ-cyhalothrin in West Africa.
Fluorescence-based allelic discrimination assays for target-site mutation detection
PCR-based allelic discrimination assays using fluorescent probes were used to detect the mutations associated with the observed resistance in the collected population. In total, 626 individuals from twenty-one populations across four countries (Benin, Togo, Niger and Mali) were tested for voltage-gated sodium channel (VGSC) and acetylcholinesterase (AChE) mutations in the invasive fall armyworm population in West Africa. Separate TaqMan assays were performed for each of the two mutations. As the target site, the F290 V mutation in AChE was the most frequent mutation found in all the populations tested. In Benin, twelve populations were tested on different host plants, and on average, 86.5%, 96.5%, 90%, 72% and 84.9% of the tested larvae were resistant (R/R and R/S) to maize, cabbage, rice, onion and sorghum, respectively (Figs. 3 and 4a). These resulted in allelic frequencies (F290 V) of 0.65 to 0.97 and 0.56 to 1 for susceptible (dead) and resistant (alive) populations, respectively (Table S2). Thus, we noted no significant difference in the frequency of the F290 V mutation between the susceptible (SS) and resistant (R/R and R/S) samples; consequently, the correlation was also not significant (P = 0.947). Similarly, for the Benin population, non-significant variations in resistance (R/R and R/S) were observed in the other countries (Togo, Niger and Mali) included in the study; 69.5%, 91.8% and 89.5% respectively (P = 0.537) (Figs. 3& 4bc). The allelic frequencies varied between 0.4 and 0.67; 0.67 and 0.94 for the susceptible population (dead); and between 0.31 and 1 and between 0.53 and 1 and 0.76 for the resistant population (alive) for the Togo, Niger and Mali population respectively. Interestingly, there was no significant difference in the frequency of the F290 V mutation between the susceptible and resistant samples; consequently, the correlation was also not significant (Table S2).
The T929I-kdr mutation was also screened in the four countries. The resistant allele was not present in any of the samples analyzed. In Benin, Togo, Niger and Mali, 100% of the tested larvae were susceptible to the different host plants (S/S) (Fig. 4 d).

Histogram showing the discrimination of different acetylcholinesterase alleles in Spodoptera frugiperda samples by an allele-specific real-time PCR fluorescent.

Real-time detection of the F290 V and T929I mutations in Spodoptera frugiperda by an allele-specific real-time PCR fluorescent probe in West Africa. Bivariate plot showing the discrimination of the acetylcholinesterase (F290 V) allele in Benin (a), Togo (b) and Niger and Mali (c). Bivariate plot showing the discrimination of the voltage-gated sodium channel (VGSC) (T929I) allele in Benin, Togo, Niger and Mali (d). Each dot represents a single larva. The blue circle represents susceptible SS homozygotes (F290; allele 2), the red lozenge mutant RR homozygotes (V290; allele 1) and the green square RS heterozygotes (F290/V290).
Discussion
Since the fall armyworm invasion in Africa in 2016, significant shifts in the production system have occurred for maize and some alternative host plants (onion, sorghum, cabbage, etc.). Smallholder farmers and national program transitioned to new production schemes to cope with the damage caused by this pest. Despite the many IPM programs funded and developed, synthetic insecticides are still the main option adopted by farmers because of their perception on its efficacity and rapid action in the native area26and the invasive area39. Nevertheless, farmers have claimed the ineffectiveness of certain classes of insecticides (pyrethroids, organophosphates). This could be due to the lack of information on the genetic characteristics (strain and pesticide resistance profile) developed by the pest or the distribution of sub-species. While two sympatric host plant strains of S. frugiperdahave been previously described the C-strain and the R-strain16,47, recent studies have reported that S. frugiperdapopulations present in Africa are interstrain hybrids, with the genetic background mostly from the corn strain46,48. In this study, the RFLP-PCR technique was used to examine the host-plant strain composition of each population. Among the 530 fall armyworm larvae analyzed, 84.8% and 15.2% were genotyped as C-strain and R-strain, respectively. This study is in contrast with the findings of Boaventura et al.49, in which more R strains (61% and 91.7%) were found in Kenya and Indonesia respectively. However, most related studies align with our results in Brazil and Puerto Rico27, China46, Togo and Ghana47and several african countries12,50. A portion of the Tpi gene was sequenced in this study to confirm the R-strain obtained via RFLP-PCR method. The Tpi haplotypes (H = 12) are still accurate markers of strain identity in Africa, and the relative frequencies of TpiC (H = 8), TpiR (H = 3), and TpiH (H = 1) should reveal evidence of differential mating similar to that observed in the Western Hemisphere47,48,50. The low number of haplotypes obtained in Benin (H = 3) suggested that after the first report of fall armyworm in Benin2, several mutations (haplotypes) could be developed during the species’ migration to other countries. Furthermore, this could also mean that IPM strategies do not allow other haplotypes to establish in Benin. In fact, the suspected R-strain of fall armyworm (RFLP analysis) was almost exclusively TpiC after sequencing, accounting for less than 3% (2/75) of the TpiR/TpiH obtained from the analyzed samples. The low percentages of TpiRs and TpiHs could be explained by the small number of males homozygous for TpiRs. Compared to the previous studies, another haplotype (TpiCa3) was established in Africa (Fig. 5). The presence of few R-strain fall armyworms in Africa could have significant consequences on the potential crops and constitute a risk for sustainable agriculture.
The high capacity of this species to evolve resistance to several chemical components requires a better understanding of the resistance mechanisms to chemical insecticides and the implementation of resistance management program. The susceptibility profile against λ-cyhalothrin, chlorpyrifos and emamectin benzoate showed a high variation of LD50 = 47.54 to 1039.82; LD50 = 206.7 to 39311.02 and LD50= 0.52 to 16.65 respectively in the survey countries. These results could explain the strong resistance variation across the countries and justified the several IPM strategies approach from a country to other. These phenotipic resistances observed were often supported by the genotypic resistance. Then, organophosphates target AChE and resistance is often associated with mutations in the ace-1 gene, leading to amino acid substitutions at the enzyme’s active site42. Our genotyping results shed more light on the presence of the amino acid substitution F290 V in 21 populations collected in Benin, Togo, Niger and Mali, as described by Carvalho et al.42 and Boaventura et al.27. Point mutations linked to organophosphate resistance have been described for Tuta absoluta (Meyrick) (Lepidoptera, Gelechiidae) and Plutella xylostella(L.) (Lepidoptera: Plutellidae), which have supported a reduction in AChE sensitivity toward carbamate and organophosphate insecticides51,52. A high frequency of the F290 V mutation in AChE (R/S and R/R) was found in all populations tested in the four countries (Fig. 3), suggesting that the AChE mutation was present in the pest from the native country. Moreover, the comparison between the dead and alive populations (P= 0.537) showed no significant difference and SRs values obtained suggesting that F290 V mutation are not only involved in the observed resistance. Otherwise, the L1014 F and T929I mutations are the most commonly reported kdr-type mutations in a range of arthropod species where resistance to pyrethroids is typically conferred42,49. In this study, only the T929I mutation was successfully screened for the involvement of the voltage-gated sodium channel (VGSC) in pyrethroid resistance. No mutations were found in the tested samples in any of the tested populations. This result suggested that the amino acid substitution T929I was not involved in the pyrethroid resistance observed after susceptibility testing with λ-cyhalothrin, as described by Zhang et al.46. The metabolic way is the main mechanism used by the fall armyworm populations tested. As mentioned in previous publications, our study results confirmed that pyrethroid resistance was involved mainly in metabolic processes53. Regarding fall armyworm susceptibility to emamectin benzoate in the different countries of our study, low concentrations of emamectin benzoate were needed compared to those of λ-cyhalothrin and chlorpyrifos. The resistance ratio considering the least susceptible population of fall armyworm to emamectin benzoate in Benin was constant (around 1.5) and varied between 1 and 24.38 in Niger and from 1 to 20.81 in Togo. These results are consistent with previous reports in Brazil54and China55 on resistance to emamectin benzoate. Nonetheless, the resistance level observed in West Africa was greater than the preceding reports. To investigate the role of metabolic resistance, synergist bioassays were performed with λ-cyhalothrin and a slight to high variation of SRs were observed. So that, the PBO, DEF and DEM synergist ration varied from 1.06 to 6.07; from 1.09 to 4.83; and from 1.07 to 6.82 respectively explaining the involvement of metabolic resistance. In a lot of cases, a low-grade insecticide was used to control 50% (LD50) of the fall armyworm population when λ-cyhalothrin is combined with synergists. This study correlates with previous studies demonstrating that metabolic patterns constitute the main route used by fall armyworms to thwart insecticide action in several countries53,55. Given the limited options for effective chemical classes for the control of S. frugiperda and the current reliance on the use of organophosphates, pyrethroids and avermectin in Africa, there is a need for ongoing monitoring of resistance development for more information on the rational choice of insecticides or biological strategies to be used to control pest damage. In summary, the informations provided by this study are a prerequisite for designing, implementation and monitoring of the resistance management strategies against this pest.
Conclusion
Based on our results, two (corn and rice) strains of fall armyworm are present in West Africa. The field ineffectiveness of organophosphates is likely compromised by the presence of the AChE F290 V allele in heterozygous and homozygous strains in West Africa, unlike pyrethroids, for which the kdr mutation does not appear at the genetic level. For the best IPM strategies to control fall armyworm, screening for insecticide resistance levels and diagnostic tools for detecting and monitoring the early presence of resistance alleles in the field are necessary to avoid multiple types of damage caused by this new invasive pest.
Material and Methods
Insect strains
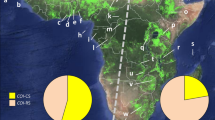
The fall armyworm larvae were collected from private property with the consent of the owners. Larvae were collected from different host plants and localities in Benin and from maize plants only in Niger, Togo and Mali (Fig. 6; Table 5). In total, four regions were prospected in Southern of Benin: Atlantic, Mono-Couffo, Ouèmé-Plateau and Zou; three regions in Niger: Niamey, Balleyarra and Maradi; two regions in Togo: Maritime and Plateau regions; and one region in Mali: Bamako. Approximately 200 larvae of all instars were collected from each location after reports of control failures of organophosphates or pyrethroids insecticides mainly used in maize crop production. Late-instar larvae and adults were examined to confirm that all individuals were fall armyworms based on diagnostic taxonomic characters56. After susceptibility testing with the F1 population, dead and alive larvae were stored at −20 °C and − 80 °C, respectively until DNA extraction and used for molecular analysis.

Diagrams of the relevant Tpi gene segments, with descriptions of the polymorphisms used to identify different haplotypes. Vertical lines in gene segments indicate sites of single-base polymorphism, with thicker bands indicating polymorphism diagnostic of strain identity. (a) Diagram of the Tpi sequence used for PCR amplification of the TpiE3; E4 exon segment. Site gTpi183 is diagnostic of Tpi-based strain identity at all locations. The gTpi165 and gTpi168 sites are also strain specific in the Western Hemisphere. The gTpi168 and gTpi180 sites are also strain specific in Africa. The gTpi192 and gTpi198 sites are polymorphic but not strain specific at all locations. (b) Tpi gene sequences.

Surveyed localities map of Spodoptera frugiperda in West Africa.
Insect rearing
Larvae were maintained on a natural diet (sprouted maize) at 25 ± 1 °C and 70–80% relative humidity (RH) with a photoperiod of 12:12 light: darkness (L: D) in the insectary. Briefly, larvae were reared on sprouted maize (corn soaked in water for 48 h, spread on paper towels and watered daily for 3 to 4 days until seedlings were obtained) in the insectary. The natural diet was changed every 3 days until the pupae stage. Pupae were collected and disinfected in 5% bleach. Emerging adult males and females from F0 larvae were pooled for mating. In separate mating cages (10 cm high and 10 cm diameter cylindrical plastic), 4–5 < 24 h female adults and 7–8 < 24 h male adults were introduced. The plastic was covered with a compress (Gauss) to allow for resting and eggs laying for females. Adults were fed with honey-soaked cotton (10%), which was subsequently renewed every 48 h. Eggs from this mating were incubated, and the emerging larvae (F1) from the hatched eggs were placed on a natural diet to obtain the L3 stage for susceptibility test.
Insecticides and chemicals
Insecticide characteristics (λ-cyhalothrin, chlorpyrifos and emamectin benzoate) used in this study were technical grade (purity > 98%) and were purchased as analytical standards from Sigma‒Aldrich (Taufkirchen, Germany) (Table 6). These insecticides were dissolved in acetone as stock solutions. Desired concentrations for bioassays were prepared from the stock solutions by serial dilution with acetone. The synergists, piperonyl butoxide (PBO), S,S, S-tributyl phosphorotrithioate (DEF) and diethyl maleate (DEM) were also obtained from Sigma‒Aldrich (Taufkirchen, Germany).
Molecular identification and analysis of the COI, JM76/77 and Tpi genes
DNA extraction
Genomic DNA was extracted from individual fall armyworm larvae (abdominal fragments of third/fourth or fifth instar larvae) using Livak protocols57 and stored at −20 °C for further analysis. Dead and alive populations from each insecticide susceptibility test (diagnostic dose) were selected for allelic discrimination assays and the same population was used for genetic speciation analysis, as shown in Tables S1 and 2. The quality and DNA concentration were determined with a microplate spectrophotometer (Multiskan SkyHigh, Thermo Scientific, Singapore).Characterization of S. frugiperda strains using PCR-RFLP. Two primers and two specific restriction enzymes were synthesized by Inqaba biotec (Pretoria, South Africa) for genotyping corn and rice strains of the fall armyworm population in West Africa following levy et al.58 and Boaventura et al.27 protocols. RFLP-PCR reactions were performed using the specific primers JM76 and JM77 (Table S3), and 10 µL of PCR product was digested in 1.0 µL of FastDigest MspI (BioLabs, New England) at 37 °C for 1 h. To validate the results, a second PCR spanning another restriction site was performed using designed forward (891 F_COI) and reverse (C1303R_COI) primers (Table S3). After amplification, the digestion step was performed by adding EcoRV (BioLabs, New England) according to the manufacturer’s instructions. Polymerase chain reaction (PCR) amplification was carried out in a 50 µL reaction mixture containing 9.2 µL of PCR Master Mix, 2 µL of primers (forward and reverse), 5 µL of DNA template and 31.8 µL of sterile H2O. The thermocycling program was an initial denaturation at 95 °C for 3 min, followed by 40 cycles of denaturation for 30 s at 95 °C, annealing at the appropriate temperature according to Table S3 for 30 s, and extension for 1 min at 72 °C, with a final elongation step of 5 min at 72 °C. The reaction was carried out in a Gene Pro Thermal Cycler (TC-E-48D; Hangzhou Bioer Technology Co., China). Negative controls without DNA were included in each experiment. At the end of the PCR, the amplification products were separated via electrophoresis in 1.5% agarose gels and stained with MIDORI Green Advance DNA stain (MG04, Nippon Genetics Europe GmbH, Germany). Molecular weights were estimated using 100 bp PCR markers from Promega.
Sequencing and species identification by characterization of Tpi Haplotypes
All individuals suspected to belong to the R-strain of fall armyworm after RFLP-PCR analysis were sequenced to confirm the molecular identification. According to Nagoshi et al.16, the portion of the Tpi gene between intron 2 and intron 4 was successfully used for genomic DNA extraction. After sequencing and cleaning, a 303 bp portion was maintained for genetic diversity. In addition, total genomic DNA was extracted and amplified using the forward (Tpi282 F) and reverse (Tpi850R) primers, as described in Table S3. The target region was amplified, purified and sequenced to screen for mutations associated with genetic differentiation. Briefly, a 12.5 µl PCR mixture containing OneTaq® Quick-Load®2X Master Mix with standard buffer (M0486, New England Biolabs, USA), 0.25 µl of each primer, 3.75 µl of nuclease water and 2 µl of the genomic DNA template was used. The thermocycling program consisted of an initial denaturation at 94 °C for 5 min, followed by 35 cycles of denaturation for 30 s at 94 °C; the respective annealing temperature according to Table S3 for 1 min, and extension for 90 s at 68 °C, with a final extension step of 10 min at 68 °C. The PCR products were visualized on an automated gel electrophoresis system according to the manufacturer’s instructions (E-BOX, Vilber Lourmat, Italy), purified using an enzymatic method (ExoSAP; Thermo Fisher Scientific, USA) and directly sequenced on both strands. The fragments were sequenced using the Nimagen Brilliant Dye™ Terminator Cycle Sequencing Kit V3.1 and BRD3-100/1000 according to the manufacturer’s instructions. The labeled products were then cleaned with the ZR-96 DNA Sequencing Clean-up Kit (D4053, Zymo Research Co., USA). The obtained sequences were aligned and corrected using BioEdit v7.2.5 software59. Alignment was performed using ClustalW multiple alignment integrated in BioEdit60and the genetic diversity parameters were assessed using DnaSP v6.12.0361and MEGA v7.062 softwares.
Bioassays
Bioassays were performed with third instar larvae (5–7 mg) of fall armyworm using a natural diet (sprouted maize) as described above. Topical application was performed for toxicity test with a hand microapplicator (Burkard Manufacturing, England), which was validated in several recent studies38. Dose‒response bioassays were carried out with seven concentrations, including the control (no insecticide) and each insecticide, and mortality was recorded at concentrations ranging from 0–100%. Approximately 100 to 250 larvae were exposed to each insecticide, and at least twenty-one larvae were treated with each concentration. One microliter of each dilution was applied topically on the pro-thoraxic dorsum of each insect. Larvae were tested individually in plastic cups (2.5 cm diameter; 3.5 cm height) containing 4 g of sprouted maize and punched at the top. The control mortality rate was less than 10%.
To evaluate the possible impact of metabolic resistance on the observed insecticide resistance, synergistic tests were performed with the three main synergists, PBO, DEF and DEM, to clarify the involvement of the oxydases, esterarses and the glutathion-S-transferase, respectively. Then, 20 µg·ml−1of PBO (piperonyl butoxide), DEF (S, S,S-tributyl phosphorotrithioate) or DEM (diethyl maleate) was added for 1 h prior to insecticide exposure. No mortality was observed in the fall armyworm after exposure to synergists alone. After treatment, the larvae were kept under the rearing conditions described above. Mortality was assessed every 24 h for 72 h for both test and control samples. Larvae were considered dead when unable to move if prodded with a blunt probe or brush63. Probit analysis was conducted using Polo-Plus v1.064 software to calculate lethal concentration values (LD50), 95% fiducial limits (F.L.),slopes and chi-square (X2). Prior to the Polo-Plus analysis, the Abbot65formula was used to correct the mortality. RRs were calculated using the most susceptible population as the reference baseline. The resistance levels for insecticides were classified as susceptible (RR ˂ 5.0), low level of resistance (5.0 ≤ RR ˂ 10.0), moderate level of resistance (10.0 ≤ RR ˂ 100.0), and high level of resistance (RR ≥ 100.0)66,67 (Lu et al., 2017; Zhang et al., 2017).
Fluorescence-Based Allelic Discrimination Assays.
Total larvae (dead and alive) resulting from first dilution greater than the DL50 from susceptibility test for each location was used for Allelic Discrimination Assays. The genomic DNA of individual fall armyworm was amplified using a primer pair designed with the Custom TaqMan Assay Design Tool (Inqaba biotec, Pretoria, South Africa). The primers Sf_taq_F290_F & Sf_taq_F290_R and TAT929I_F & TAT929I_R were used for the detection of the F290 V and T929I mutations in Ace1 and VGSC, respectively (Table S3). The probes Sf_F290_FAM and Sf_F290_mut_HEX and TAT929I-VIC and TAT929I-FAM were used in the F290 V and T929I assays, respectively (Table S3). The PCR reactions set up at a final volume of 10 µL and contained 5 µL of Universal Probes qPCR Master Mix (Luna, BioLabs, New England), 700 nM forward and reverse primers, 200 nM probes, 20–50 ng of gDNA and nuclease-free water. The PCR conditions were as follows: 1 cycle at 95 °C for 5 min; 40 cycles at 95 °C for 15 s, 60 °C for 30 s, and 72 °C for 1 min; and a final extension at 72 °C for 10 min. Real-time PCR was conducted with an Mx3000PQPCR System (Aligent Technologies, USA), and the end-point fluorescence values, taking cycle 35 as a threshold, were plotted as a Catter plot using MxPro–MX3000P analysis software (Roche Molecular Systems, Agilent Technologies, USA).
Data availability
All data generated or analyzed during this study are included in this published article. The sequences used for the phylogenetic analysis are deposited into GenBank and included in (PP457652 – PP457726).
References
Wang, J. et al. Migration risk of fall armyworm (Spodoptera frugiperda) from North Africa to Southern Europe. Front. Plant. Sci. 14, 1–12 (2023).
Goergen, G., Kumar, P. L., Sankung, S. B., Togola, A. & Tamò, M. First report of outbreaks of the fall armyworm spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS One. 11, 1–9 (2016).
Otim, M. H. et al. Detection of sister-species in invasive populations of the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) from Uganda. PLoS One. 13, e0194571 (2018).
Kumela, T. et al. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. Int. J. Pest Manag. 65, 1–9 (2019).
Sharanabasappa et al. First report of the fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag Hortic. Ecosyst. 24, 23–29 (2018).
Tay, W. T., Kuniata, L., James, W. & Walsh, T. Confirmation of Spodoptera frugiperda (Lepidoptera: Noctuidae) in Papua new Guinea by molecular diagnostics of mitochondrial DNA COI gene. BioInvasions Rec. 12, 103–116 (2023).
Tepa-Yotto, G. T., Chinwada, P., Rwomushana, I., Goergen, G. & Subramanian, S. Integrated management of Spodoptera frugiperda 6 years post detection in Africa: a review. Curr. Opin. Insect Sci. 52, 100928 (2022).
Montezano, D. G. et al. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 26, 286–300 (2018).
Barros, E. M., Torres, J. B., Ruberson, J. R. & Oliveira, M. D. Development of Spodoptera frugiperda on different hosts and damage to reproductive structures in cotton. Entomol. Exp. Appl. 137, 237–245 (2010).
Gouin, A. et al. Two genomes of highly polyphagous lepidopteran pests (Spodoptera Frugiperda, Noctuidae) with different host-plant ranges. Sci. Rep. 7, 1–12 (2017).
Silva-Brandão, K. L. et al. Loci under selection and markers associated with host plant and host-related strains shape the genetic structure of Brazilian populations of Spodoptera frugiperda (Lepidoptera, Noctuidae). PLoS One. 13, e0197378 (2018).
Schlum, K. A. et al. Whole genome comparisons reveal panmixia among fall armyworm (Spodoptera frugiperda) from diverse locations. BMC Genom. 22, 1–13 (2021).
Laura Juárez, M. et al. Host association of spodoptera frugiperda (Lepidoptera: Noctuidae) corn and rice strains in Argentina, Brazil, and Paraguay. J. Econ. Entomol. 105, 573–582 (2012).
Prowell, D. P., McMichael, M. & Silvain, J. F. Multilocus genetic analysis of host use, introgression, and speciation in host strains of fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 97, 1034–1044 (2004).
Nagoshi, R. N., Meagher, R. L. & Hay-Roe, M. Inferring the annual migration patterns of fall armyworm (Lepidoptera: Noctuidae) in the united States from mitochondrial haplotypes. Ecol. Evol. 2, 1458–1467 (2012).
Nagoshi, R. N. The fall armyworm triose phosphate isomerase (Tpi) gene as a marker of strain identity and interstrain mating. Ann. Entomol. Soc. Am. 103, 283–292 (2010).
Schöfl, G., Heckel, D. G. & Groot, A. T. Time-shifted reproductive behaviours among fall armyworm (Noctuidae: Spodoptera frugiperda) host strains: evidence for differing modes of inheritance. J. Evol. Biol. 22, 1447–1459 (2009).
Groot, A. T., Marr, M., Heckel, D. G. & SchÖfl, G. The roles and interactions of reproductive isolation mechanisms in fall armyworm (Lepidoptera: Noctuidae) host strains. Ecol. Entomol. 35, 105–118 (2010).
Nagoshi, R. N. et al. Genetic characterization of fall armyworm infesting South Africa and India indicate recent introduction from a common source population. PLoS One. 14, e0217755 (2019).
Jing, D. P. et al. Initial detections and spread of invasive Spodoptera frugiperda in China and comparisons with other noctuid larvae in cornfields using molecular techniques. Insect Sci. 27, 780–790 (2020).
Pashley, D. P. Current Status of Fall Armyworm Host Strains Author (s): Dorothy P. Pashley Source : The Florida Entomologist, Vol. 71, No. 3 (Sep., 1988), pp. 227–234 Published by : Florida Entomological Society Stable URL : (1988). https://www.jstor.org/stable/34954. Florida Entomol. Soc. Stable 71, 227–234.
Adamczyk Jr, J. J., Leonard, B. R., Graves, J. B., Adamczyk, J. Jr & Leonard, B. R. Toxicity of Selected Insecticides to Fall Armyworms (Lepidoptera: Noctuidae) in Laboratory Bioassay Studies Author (s): J and J. B. Graves Published by : Florida Entomological Society Florida Entomological Soc. Florida Entomol. 82, 230–236 (1999).
Eschen, R. et al. Towards estimating the economic cost of invasive alien species to African crop and livestock production. CABI Agric. Biosci. 2, 1–18 (2021).
Kassie, M. et al. Economic impacts of fall armyworm and its management strategies: evidence from Southern Ethiopia. Eur. Rev. Agric. Econ. 47, 1473–1501 (2020).
Chimweta, M., Nyakudya, I. W., Jimu, L. & Bray Mashingaidze, A. Fall armyworm [Spodoptera frugiperda (J.E. Smith)] damage in maize: management options for flood-recession cropping smallholder farmers. Int. J. Pest Manag. 66, 142–154 (2020).
Gutirrez-Moreno, R. et al. Field-Evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 112, 792–802 (2019).
Boaventura, D., Martin, M., Pozzebon, A., Mota-Sanchez, D. & Nauen, R. Monitoring of target-site mutations conferring insecticide resistance in spodoptera frugiperda. Insects 11, 1–15 (2020).
Ahissou, B. R. et al. Baseline toxicity data of different insecticides against the fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) and control failure likelihood Estimation in Burkina Faso. Afr. Entomol. 29, 435–444 (2021).
Garlet, C. G. et al. Field-evolved resistance to Chlorpyrifos by Spodoptera frugiperda (Lepidoptera: Noctuidae): Inheritance mode, cross-resistance patterns, and synergism. Pest Manag Sci. 77, 5367–5374 (2021).
Yu, S. J. Insecticide resistance in the fall armyworm, Spodoptera frugiperda (J. E. Smith). Pestic Biochem. Physiol. 39, 84–91 (1991).
Deshmukh, S. et al. Field efficacy of insecticides for management of invasive fall armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) on maize in India. Fla. Entomol. 103, 221–227 (2020).
Okuma, D. M. et al. Inheritance and fitness costs of Spodoptera frugiperda (Lepidoptera: Noctuidae) resistance to spinosad in Brazil. Pest Manag Sci. 74, 1441–1448 (2018).
Boaventura, D. et al. Detection of a Ryanodine receptor target-site mutation in diamide insecticide resistant fall armyworm, Spodoptera frugiperda. Pest Manag Sci. 76, 47–54 (2020).
Bolzan, A. et al. Selection and characterization of the inheritance of resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to Chlorantraniliprole and cross-resistance to other diamide insecticides. Pest Manag Sci. 75, 2682–2689 (2019).
Nonci, N., Pakki, S. & Muis, A. Field testing of synthetic inseticides on fall armyworm (Spodoptera Frugiferda J.E. Smith) in corn plant. IOP Conf. Ser. Earth Environ. Sci. 911, 012059 (2021).
Arias, O. et al. Population genetic structure and demographic history of Spodoptera frugiperda (Lepidoptera: Noctuidae): implications for insect resistance management programs. Pest Manag Sci. 75, 2948–2957 (2019).
Kansiime, M. K. et al. Farmer perception of fall armyworm (Spodoptera Frugiderda J.E. Smith) and farm-level management practices in Zambia. Pest Manag Sci. 75, 2840–2850 (2019).
Tossou, E. et al. Susceptibility profiles of helicoverpa armigera (Hübner) (lepidoptera: Noctuidae) to deltamethrin reveal a contrast between the Northern and the Southern Benin. Int. J. Environ. Res. Public. Health. 16, 1882–1897 (2019).
Sisay, B., Tefera, T., Wakgari, M., Ayalew, G. & Mendesil, E. The efficacy of selected synthetic insecticides and botanicals against fall armyworm, spodoptera Frugiperda, in maize. Insects 10, 10020045 (2019).
Tossou, E. et al. A novel approach for assessing technical grade and quality of Lambda-Cyhalothrin and Acetamiprid in insecticides used in agricultural systems by HPLC technique in Southern Benin. Agrochemicals 2, 551–560 (2023).
Mota-Sanchez, D. and J. C. W. The Arthropod Pesticide Resistance Database. IRAC, Michigan State University (2023). http://www.pesticideresistance.org
Carvalho, R. A., Omoto, C., Field, L. M., Williamson, M. S. & Bass, C. Investigating the molecular mechanisms of organophosphate and pyrethroid resistance in the fall armyworm Spodoptera frugiperda. PLoS One. 8, e62268 (2013).
Hilliou, F., Chertemps, T., Maïbèche, M. & Le Goff, G. Resistance in the genus spodoptera: key insect detoxification genes. Insects 12, 1–27 (2021).
Yainna, S. et al. Geographic monitoring of insecticide resistance mutations in native and invasive populations of the fall armyworm. 1–12 (2021).
Swathi, R. S. & Venkatesan, T. Identification of Spodoptera frugiperda by using Triose Phosphate Isomerase (TPI). Unpubl. Submitt. Genomic Resour. ICAR- NBAIR, Bellary Road, Banglore, Karnataka 560024, India (2021).
Zhang, L. et al. Genetic structure and insecticide resistance characteristics of fall armyworm populations invading China. Mol. Ecol. Resour. 20, 1682–1696 (2020).
Nagoshi, R. N. et al. The fall armyworm strain associated with most rice, Millet, and pasture infestations in the Western hemisphere is rare or absent in Ghana and Togo. PLoS One. 16, 1–16 (2021).
Nagoshi, R. N. et al. Genetic studies of fall armyworm indicate a new introduction into Africa and identify limits to its migratory behavior. Sci. Rep. 12, 1–12 (2022).
Boaventura, D. et al. Molecular characterization of Cry1F resistance in fall armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 116, 103280 (2020).
Nagoshi, R. N., Goergen, G., Plessis, H., van den Du, J. & Meagher, R. Genetic comparisons of fall armyworm populations from 11 countries spanning sub-Saharan Africa provide insights into strain composition and migratory behaviors. Sci. Rep. 9, 1–11 (2019).
Hyung, J. et al. Identification and characterization of ace1 -type acetylcholinesterase likely associated with organophosphate resistance in plutella Xylostella. 81, 164–175 (2005).
Haddi, K. Mutation in the ace-1 gene of the tomato leaf miner (Tuta absoluta) associated with organophosphates resistance. 1–8 (2015). https://doi.org/10.1111/jen.12386
Zhao, Y. X. et al. Susceptibility of fall armyworm, Spodoptera frugiperda (J.E.Smmith), to eight insecticides in China, with special reference to lambda-cyhalothrin. Pestic Biochem. Physiol. 168, 104623 (2020).
Muraro, D. S. et al. Inheritance patterns, cross-resistance and synergism in Spodoptera frugiperda (Lepidoptera: Noctuidae) resistant to Emamectin benzoate. Pest Manag Sci. 77, 5049–5057 (2021).
Chen, H. L. et al. Resistance monitoring and mechanism in the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) for Chlorantraniliprole from Sichuan Province, China. Front. Physiol. 14, 1–11 (2023).
Capinera, J. L. Identification fall armyworm, Spodoptera frugiperda. (1999).
Livak, K. J. Organization and mapping of a sequence on the drosophila melanogaster X and y chromosomes that is transcribed during spermatogenesis. Genetics 107, 611–634 (1984).
Levy, H. C., Garcia-Maruniak, A. & Maruniak, J. E. Strain identification of Spodoptera frugiperda (Lepidoptera: Noctuidae) insects and cell line: PCR-RFLP of cytochrome oxidase C subunit I gene. Fla. Entomol. 85, 186–190 (2003).
Hall, T. A. A. & User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows. 95–98 at (1999).
Thompson, J., Higgins, D. & Gibson, T. CLUSTAL W: (improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice). Encycl Genet. Genomics Proteom. Inf. 22, 376–377 (2008).
Rozas, J. et al. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 34, 3299–3302 (2017).
Kumar, S., Stecher, G. & Tamura, K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874 (2016).
Kranthi, K. Insecticide resistance monitoring, mechanisms and management manual. Central Institute for Cotton Research, Nagpur Pp (2005). https://doi.org/10.1016/0191-8141(80)90002-4
Robertson, J. L., Russell, R. M. & Savin, N. E. POlo: a user ’ s guide to probit or logit analysis. Pac. Southwest. Range Exp. Stn. (1980).
Abbot, W. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 265–267. https://doi.org/10.1093/jee/18.2.265a (1925).
Zhang, X. et al. Neonicotinoid insecticide resistance in the field populations of Sogatella furcifera (Horváth) in central China from 2011 to 2015. J. Asia Pac. Entomol. 20, 955–958 (2017).
Lu, Y. et al. Resistance monitoring of Chilo suppressalis (Walker) (Lepidoptera: Crambidae) to Chlorantraniliprole in eight field populations from East and central China. Crop Prot. 100, 196–202 (2017).
Acknowledgements
The authors thank numerous field researchers for collecting fall armyworm samples in Benin, Togo, Niger and Mali. The Agro-Eco-Health platform of the International Institute of Tropical Agriculture – Benin supported this research. I would like also to thank Abdoulaye Tahirou for financial support.
Funding
This research received funding from the Wellcome Trust International Intermediate Fellowship 214201/Z/18/Z awarded to Rousseau Djouaka. Support was also received from the Royal Norwegian Embassy in Mali for climate smart agricultural technologies for improved rural livelihoods and food security in Mali (Grant MLI-17-0008) and Niger (Grant NER-17-0005). Similarly, the authors thankfully acknowledge the financial support from the German Federal Ministry for Economic Cooperation and Development (BMZ) commissioned by the Deutsche Gesellschaft für Internationale Zusammenarbeit (GIZ) through the Fund International Agricultural Research (FIA) (grant number 81235252 GA).
Author information
Authors and Affiliations
Contributions
E.T., G.T.T.-Y., G.G. L.A., M.N., and R.D. designed the study; E.T., G.M.T., G.T.T.-Y., G.G., and R.D. conducted the experiments; E.T., M.T., D.N.N, C.W. analysed the data; E.T., R.D. wrote the manuscript; and M.T., C.W., M.T., G.T.T.-Y. and A.H.B.-G. crosschecked the manuscript prior to submission.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Tossou, E., Tepa-Yotto, G.T., Goergen, G. et al. Genetic variation associated with increased lambda-cyhalothrin resistance in Spodoptera frugiperda (Lepidoptera: Noctuidae) in West Africa. Sci Rep 15, 21812 (2025). https://doi.org/10.1038/s41598-025-99196-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-99196-1
