
Abstract
The deubiquitinating enzyme Ubiquitin specific peptidase 5 (USP5) has attracted substantial notice for its vital role in cancer progression. However, the USP5-mediated deubiquitination of corresponding protein substrates and its functional role in hepatocellular carcinoma (HCC) have not been fully investigated. Here, we demonstrated that USP5 expression was significantly elevated in HCC tissues. The overexpression of USP5 was closely associated with larger tumor sizes, more satellite nodules and tumor emboli, and predicted unfavorable clinical outcome in HCC patients as well. Functionally, USP5 facilitated cell proliferation, migration, and invasion, and induced lipid accumulation in vitro, along with enhanced tumor growth in vivo. Moreover, knockdown of USP5 expression showed a profound effect on lipidomic profiling, specially reduced the content of palmitic acid (PA). Treatment of PA could partially rescue the suppression of HCC mediated by USP5 knockdown. Further mechanistic investigation uncovered that Fatty acid synthase (FASN), the crucial enzyme catalyzing PA synthesis, was a downstream target of USP5. USP5 interacted with FASN, repressing the ubiquitination modification of FASN and preventing its degradation. Notably, the positive correlation between USP5 and FASN expression in HCC tissues was observed, and USP5 exerted oncogenic effects partly via FASN. Our findings revealed that USP5 promotes HCC progression through deubiquitinating FASN, and targeting the USP5-FASN-PA axis could potentially serve as a strategic approach for the therapy of HCC.
Similar content being viewed by others

Introduction
Hepatocellular carcinoma (HCC), which comprises approximately 90% of primary liver cancer cases, stands as one of the most formidable and widespread malignancies in humans [1]. The heterogeneity and complexity of HCC leads to late-stage diagnoses and unfavorable overall survival outcomes for patients [2]. Metabolic reprogramming serves as a well-established hallmark of cancer [3]. The liver, being the central hub of metabolism within the body, experiences profound alterations in metabolic processes in HCC. The dysregulation of lipids assumes pivotal roles in the development and progression of HCC, owing to the vast and intricate nature of lipids as complex biomolecules. Lipids exert a critical role in multiple biological processes, encompassing not only energy storage and metabolism, but also the transmission of signals, the regulation of immune responses, the modulation of inflammatory cascades, and the intricate interplay of cellular recognition [4,5,6]. For example, elevated palmitic acid (PA), a saturated fatty acid with a 16-carbon chain, is strongly linked to tumor progression through multiple oncogenic mechanisms. PA induces a pro-metastatic memory via Schwann cells and fosters an immunosuppressive tumor microenvironment [7, 8]. PA drives metastasis by reinforcing YAP signaling through the ZDHHC15-YAP positive feedback loop [9]. Moreover, PA promotes RAS palmitoylation, a critical step for its oncogenic activation [10]. Given these multifaceted roles, elucidating the regulatory mechanisms of PA synthesis is crucial for understanding and potentially targeting cancer progression.
Fatty acid synthase (FASN) is an essential enzyme responsible for the de novo synthesis of long-chain fatty acids, particularly PA, a process dependent on nicotinamide adenine dinucleotide phosphate (NADPH) as a reducing cofactor [11]. In normal cells, lipid demands are primarily met through exogenous uptake, keeping FASN expression at basal levels due to sufficient fatty acid supply from dietary sources and hepatic or adipose stores. In contrast, cancer cells exhibit marked FASN upregulation, as their accelerated proliferation and survival hinge on enhanced de novo lipogenesis to meet heightened metabolic demands [12,13,14]. This metabolic reprogramming underscores FASN’s critical role in sustaining tumor growth, making it a compelling therapeutic target for HCC [15]. FASN is highly expressed in HCC and correlates with sorafenib treatment response. Its transcription is regulated by upstream transcription factors, including liver X receptor (LXR) and sterol regulatory element-binding protein 1 (SREBP1) [16,17,18]. More importantly, several types of post-translational modification are also involved in FASN upregulation in HCC. Moreover, post-translational modifications contribute significantly to FASN upregulation in HCC. For example, S-palmitoylation of FASN mediated by the DHHC-type palmitoyltransferase ZDHHC20 enhances its protein stability, a process critical for hepatocarcinogenesis [19]. Additionally, heat shock protein 90α (Hsp90α) interacts with FASN and stabilizes it by suppressing ubiquitination [20]. Consequently, elucidating the mechanisms driving FASN overexpression in HCC is imperative for developing targeted interventions.
Ubiquitination represents a crucial post-translational modification that regulates protein activity, function, localization, and stability through a well-characterized enzymatic cascade. In this process, E1 (ubiquitin-activating enzyme), E2 (ubiquitin-conjugating enzyme), and E3 (ubiquitin ligase) sequentially catalyze the transfer of activated ubiquitin molecules to lysine residues on target proteins [21]. The addition of ubiquitin signals by E3 ubiquitin ligases is counteracted by around 100 deubiquitinating enzymes (DUBs) in humans. DUBs and E3 ligases work together to regulate ubiquitin signaling by conferring specificity for various substrates and/or ubiquitin signals. The balance between ubiquitination and deubiquitination is meticulously regulated to maintain proper proteostasis and enable appropriate responses to cellular stimuli and stressors. Consequently, DUBs have been implicated in all fundamental characteristics of cancer [22]. Based on sequence and domain conservation, DUBs are classified into nine superfamilies, including ubiquitin-specific proteases (USPs), ovarian tumor proteases (OTUs), ubiquitin C-terminal hydrolases (UCHs), and JAMM/MPN domain-associated metalloproteases (JAMMs) [22, 23]. Ubiquitin-specific protease 5 (USP5) is a member of the USP family, which governs a multitude of cellular processes, such as the repair of DNA double-strand breaks, modulation of inflammatory reactions, regulation of stress responses, glucose metabolism reprogramming, and epithelial-mesenchymal transition (EMT) [24,25,26,27,28]. USP5 is frequently overexpressed in HCC and correlates with aggressive tumor behavior. It promotes epithelial–mesenchymal transition (EMT) by deubiquitinating SLUG and inosine monophosphate dehydrogenase 2 (IMPDH2) [25, 29]. USP5 also interacts with the oncoprotein c-Myc, stabilizing it by inhibiting K48-linked polyubiquitination and thereby modulating glucose metabolism [28]. Moreover, USP5-mediated deubiquitination of lymphoid-specific helicase (LSH) enhances hepatocarcinogenesis through upregulation of solute carrier family 7 member 11 (SLC7A11), which suppresses ferroptosis in HCC cells [30]. Despite these established roles, the key protein substrates of USP5 in lipid metabolism during HCC progression remain poorly defined, highlighting an important area for investigation.
In this study, we demonstrated that USP5 silencing markedly suppresses HCC cell proliferation, migration, invasion, and PA accumulation. Moreover, we elucidated the molecular interaction between USP5 and fatty acid synthase (FASN), revealing that USP5 inhibits FASN ubiquitination, thereby stabilizing its expression in HCC. These findings highlight the oncogenic role of USP5 in driving HCC progression and propose USP5 as a promising therapeutic target for HCC treatment.
Materials and methods
Tissue collection
106 pairs of HCC and corresponding adjacent normal liver tissues were obtained from patients with HCC who underwent hepatectomy at Zhongshan Hospital of Xiamen University from 2013 to 2022. Prior to the surgery, these patients did not undergo interventional therapy, chemotherapy, or targeted therapy. Tissue samples were collected, immediately fixed in 10% neutral formalin solution. Written informed consent for this research was obtained from all patients. The informed consent document and experimental procedures were approved by the Ethics Committee of Zhongshan Hospital of Xiamen University in accordance with the World Medical Association Declaration of Helsinki.
Cell culture
HEK293T, Huh7, and SK-Hep-1 cells were purchased from the Cellcook Company (Guangzhou, China). The authentication of these cell lines was performed through comparison with the STR database, and all experiments were performed with mycoplasma-free cells. The cells were cultured in high-glucose Dulbecco’s modified Eagle medium (DMEM; Gibco, USA) supplemented with 10% fetal bovine serum (FBS, Vivacell, Shanghai, China) as well as Penicillin-Streptomycin (Gibco, USA) at 37 °C in a humidified incubator with 5% CO2 in the air.
Immunohistochemical (IHC) staining
Fixed tissues were processed using the Tissue-Tek TEC5 Embedding Centre (Sakura Finetek Co., Japan), following a standard protocol, and embedded in paraffin blocks as 4-μm thick sections. Briefly, IHC staining proceeded as follows: sections were deparaffinized, hydrated, and immersed in 3% H2O2 for 20 min at room temperature. Subsequently, they were incubated overnight at 4 °C with anti-USP5 (1:10000, ab10800, Abcam) and anti-FASN (1:1000, 10800-1-AP, Proteintech) antibodies. Slides were then treated with biotinylated goat anti-rabbit antibodies for 1 h and stained with diaminobenzidine (DAB; Maixin Biotechnology, China), followed by hematoxylin counterstaining (Maixin Biotechnology, China). To prevent bias, two clinical pathologists, blinded to experimental data, independently assessed IHC-stained sections. In summary, 100 cells were randomly counted in 200× microscopic fields and categorized into five groups based on the percentage of positive staining cells in HCC tissues: 0=negative; 1–3 = 1–25%; 4–6 = 26–50%; 7–9 = 51–75%; 10–12 = ≥ 76%. Scores from 0 to 6 indicated low-expression, while scores from 7 to 12 indicated high-expression.
Overexpression of USP5
Lentiviral particles expressing an empty vector and USP5-Flag were purchased from Public Protein/Plasmid Library (Jiangsu Province, China). The cells were infected with these lentiviruses in the presence of polybrene (8 μg/mL). After 48 h, the cells were used for the following experiments.
Knockdown of USP5 expression
Two different shRNAs targeting USP5 (shUSP5-1: CCACACGATTTGCCTCATT, shUSP5-2: TAGACATGAACCAGCGGAT) and one shRNA targeting FASN (shFASN: TACTGGATGCGTTCTTCAA) were inserted into pLV-shRNA-puro lentiviral, respectively. Lentiviral particles were produced in HEK293T cells according to the standard procedure. The lentiviral particles were collected 48 h after transfection. Cells were infected with viral supernatants using polybrene (8 μg/mL). After 48 h of infection, stable cells were selected using 5 μg/mL puromycin (Inovogen, China) for one week.
Cell proliferation detection
Cells were plated in triplicate at a density of 1 ~ 2 × 103 cells per well in 96-well culture plates. The cell proliferation was evaluated in three separate experiments through the use of the CCK-8 (Dojindo, China) assay at specified time intervals, following the manufacturer’s guidelines. The absorbance was quantified at 450 nm using a microplate reader (Infinite® F50, Tecan).
Transwell assay
To assess cell migration, the specified cells were resuspended with 100 μL serum-free DMEM and then applied to the upper chamber (8 μm pore size, Corning, USA). Cell invasion was tested using Matrigel Invasion Chamber (8 μm pore size, Corning, USA). The lower chamber was filled with 600 μL DMEM containing 10% FBS. Subsequent to 24 h of incubation, the cells that migrated or invaded were fixed with 4% paraformaldehyde at room temperature for 20 min, followed by staining with 1% crystal violet for 30 min. The cells were subsequently counted under a microscope.
Mass spectrometry
Huh7 cells were treated with 10 μM MG132 (MedChemExpress) for 6 h. Cell lysates were subjected to immunoprecipitation using anti-USP5 antibodies (Cell Signaling Technology, USA) overnight. Subsequently, these immunoprecipitated complexes were combined with Dynabeads Protein G (Invitrogen, USA) for an additional 2 h incubation. Gel electrophoresis was performed, followed by in-gel digestion to prepare samples for subsequent proteomic analysis. The LC-MS/MS analysis aimed at identifying proteins interacting with USP5 was carried out by Applied Protein Technology (Shanghai, China).
Protein extraction and western blotting
Cell lysis was performed using radioimmunoprecipitation assay (RIPA) lysis buffer (Beyotime, China). Following quantification with a BCA protein assay kit (Thermo), protein samples underwent separation through sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and subsequent transfer onto polyvinylidene difluoride (PVDF) membranes (Millipore, USA). After blocking with 5% non-fat milk (LABLEAD, China) for 1 h at room temperature, membranes were incubated with anti-USP5 (Cell Signaling Technology, USA), anti-β-actin (Axel Biotechnology, USA), or anti-FASN (Abcam, UK) antibodies. This was followed by incubation with anti-rabbit or anti-mouse secondary antibodies conjugated to horseradish peroxidase (HRP, Jackson). Immunoreactive proteins were then visualized using the enhanced chemiluminescence (ECL) detection system (Millipore, USA).
Co-immunoprecipitation
Total cellular proteins were extracted using co-immunoprecipitation (co-IP) lysis buffer (Beyotime, China). Cell lysates were incubated with anti-USP5 (Cell Signaling Technology, USA) or anti-FASN antibody (Abcam, UK) or a negative control IgG (Cell Signaling Technology, USA) and Dynabeads Protein G for immunoprecipitation (Invitrogen, USA) overnight at 4 °C. Subsequently, the beads were washed three times with lysis buffer, and the immunoprecipitated samples were collected for western blot analysis.
Animal study
A xenograft mouse model was established using 4-week-old male BALB/c athymic nude mice. A total of 2 × 106 Huh7 cells with shNC or shUSP5-1 or shUSP5-1 + FASN were subcutaneously injected into the nude mice, respectively. After approximately ten days, tumor size was assessed using a vernier caliper, and tumor volumes were calculated with the formula V = ½ (length × width2). 30 days post-injection, the mice were sacrificed under anesthesia. Then, the tumors were fixed in 10% neutral formalin, paraffin-embedded, and sectioned into 3 μm thick slices, which were used for further detection. All animal experiments were undertaken in accordance with relevant guidelines and granted approval by the Animal Care and Use Committee of Xiamen University.
Oil red O and Nile red staining
Cells were cultured in 6-well plates. At 70–80% confluence, a palmitic acid (PA; 0.25 mM; Kunchuang Biotechnology, Xi’an, China) and oleic acid (OA; 0.5 mM; Kunchuang Biotechnology, Xi’an, China) mixture in 0.5% BSA was added to the medium for 24 h. Cells were washed 3–4 times with PBS, fixed with 4% paraformaldehyde for 1 h, and incubated with Oil-Red O (Sigma, USA) for 1 h. Finally, the lipid droplets were visualized using light microscopy. For Nile red staining, cells were fixed with 4% paraformaldehyde solution, washed with PBS, and stained with Nile Red solution (7385-67-3, Solarbio, China) in the dark. In fluorescence microscopy, images were acquired after the samples were washed twice with PBS and stained with DAPI.
Targeted metabolomics
The lipidomic profiling was analyzed by Applied Protein Technology Company (Shanghai, China). In brief, tumor cells cultured to the logarithmic phase were treated with trypsin, centrifuged, and counted. The cells were then resuspended in the culture medium, adjusted to the desired concentration, plated in a 10 cm culture dish, and incubated overnight at 37 °C in a 5% CO2 atmosphere. When the cells reached 80% confluence, they were harvested and cryopreserved in liquid nitrogen. Samples were thawed on ice, and 50 µL of each sample was added into a 2 mL glass centrifuge tube, then mixed with 5 mL methylene dichloride-methanol (2:1 v/v). The mixture was incubated in an 80 °C water bath for 30 min for fatty-acid esterification. Then, 200 µL internal standard, 1 mL n-hexane, and 5 mL water were added and vortex mixed. The supernatant (500 µL) was subjected to GC-MS using an Agilent Model 7890–5977 system. To quantify medium- and long-chain fatty acids, a calibration curve for the concentration range of 0.1–2500 µg/mL was constructed. The internal standard was used to correct for injection variability and instrument response changes. Samples were separated with an Agilent DB-23 capillary GC column (60 m × 250 µm ID × 0.15 µm). The initial temperature was 50 °C for 3 min, increased to 220 °C at 10 °C/min, then to 250 °C at 15 °C/min, and held at 250 °C for 10 min. The carrier gas was helium (1.0 mL/min). Injection port and transmission line temperatures were 280 °C and 250 °C, respectively, under SIM mode.
Quantitative analysis of ubiquitination modifications
The ubiquitination proteomics was performed by Novogene (Beijing, China). Total Protein Extraction: Samples were ground in liquid nitrogen, lysed with 100 mM NH4HCO3 (pH 8), 8 M Urea, 0.2% SDS, and ultrasonicated on ice. Lysates were centrifuged (12000 g, 15 min, 4 °C) and the supernatant was collected. Proteins were reduced with 10 mM DTT (1 h, 56 °C) and alkylated with iodoacetamide (1 h, room temperature, dark). Samples were mixed with 4x volume of cold acetone and incubated at −20 °C for 2 h, then centrifuged. The pellet was washed with cold acetone and dissolved in 0.1 M TEAB (pH 8.5), 6 M urea. Protein Quality: Protein concentration was determined using the Bradford assay. 20 μg protein samples were analyzed on 12% SDS-PAGE, stained with Coomassie blue, and decolored until bands were clear. Peptide Preparation: 10 mg protein was digested with trypsin in 50 mM TEAB buffer (37°C, 4 h), followed by overnight digestion with CaCl2. pH was adjusted to <3 with formic acid, centrifuged, and the supernatant was desalted on a C18 column. Ubiquitin Peptide Enrichment: Lyophilized samples were dissolved in MOPS IAP buffer, mixed with anti-Ubiquitin Remnant Motif beads (2 h, 4 °C), washed, and eluted. Eluates were desalted and lyophilized. LC-MS/MS Analysis: Peptides were separated on a C18 column and analyzed with an EASY-nLCTM 1200 UHPLC system coupled to a Q Exactive HF-X mass spectrometer. MS and MS/MS settings included a full scan range of m/z 350–1500, resolution of 120,000 (MS) and 15,000 (MS/MS), AGC target values, and specific collision energies.
Statistical analysis
All statistical analyses were conducted utilizing SPSS software (Version 19.0). The data is presented as mean ± standard deviation (SD), and three independent experiments were carried out. Group-wise differences in the results were assessed using either Student’s t-test or a multi-way analysis of variance (ANOVA) test. The association between USP5 expression and clinical features of HCC patients was evaluated through the Chi-squared test. Spearman’s rank correlation was employed to scrutinize the relationship between USP5 and FASN expression. The Kaplan-Meier survival test and log-rank test were utilized to investigate the correlation between USP5 expression and the prognosis of HCC patients. Significance was assigned for p-values less than 0.05.
Results
Upregulation of USP5 is closely associated with poor prognosis in HCC
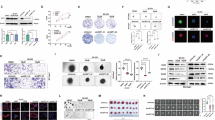
In The Cancer Genome Atlas (TCGA) database, we observed that USP5 is upregulated in most cancers (Supplementary Fig. 1A), including breast cancer (BRCA), cholangiocarcinoma (CHOL), and lung squamous cell carcinoma (LUSC). Notably, its expression is significantly higher in liver hepatocellular carcinoma (LIHC) compared to normal tissues, accompanied by a markedly lower promoter methylation level (Fig. 1A). To further evaluate the clinical significance of USP5 expression, we stratified the TCGA cohort into four quartiles based on risk scores: Q1 (below the 25th percentile, 93 patients), Q2 (between the 25th and 50th percentiles, 92 patients), Q3 (between the 50th and 75th percentiles, 92 patients), and Q4 (above the 75th percentile, 93 patients). Kaplan-Meier analysis revealed significant differences in overall survival among the four groups (p = 0.0083, Fig. 1B), with Q4 patients exhibiting the poorest prognosis. Notably, survival time progressively decreased from Q1 to Q4, and the Q4 group demonstrated the highest mortality rate and shortest survival duration (Fig. 1C). Furthermore, we assessed the association between USP5 expression and clinicopathological features using chi-square tests. In the TCGA cohort, the low-USP5 group not only showed a significant survival advantage but also correlated with lower TNM stage (p = 0.015) and HCC stage (p = 0.046) compared to the high-expression group (Table 1).

A Analysis of USP5 expression (top panel) and promoter methylation levels (bottom panel) in LIHC and adjacent normal tissues using UALCAN, an online platform for comprehensive cancer genomics analysis. B Kaplan–Meier survival curves of the TCGA-LIHC cohort showing that patients with high USP5 expression exhibited significantly poorer overall survival than those with low expression (log-rank test), analyzed using the Home for Researchers platform. C Distribution of USP5 expression scores (top panel) and corresponding survival status (middle panel) among HCC patients in the TCGA cohort. D Representative IHC staining of USP5 in clinical HCC tissues and matched adjacent non-tumor tissues (left panel), with quantitative analysis of pathologist-scored USP5 expression levels (right panel). Scale bar: 200 μm. E Kaplan–Meier analysis of tumor-free survival in the clinical cohort, showing significantly shorter survival in patients with high USP5 expression (log-rank test). F Kaplan–Meier analysis of overall survival in the clinical cohort, revealing significantly worse outcomes for patients with high USP5 expression (log-rank test).
To further validate USP5 expression in HCC, we performed immunohistochemical (IHC) staining on 106 paraffin-embedded tissue sections from our institutional cohort. Consistent with bioinformatics findings, USP5 expression was significantly upregulated in HCC tissues compared to adjacent non-tumor tissues (p < 0.05, Fig. 1D). Based on IHC scoring, patients were stratified into high- and low-expression groups. Kaplan-Meier analysis demonstrated that the high-USP5 group had significantly worse disease-free survival (DFS) and overall survival (OS) than the low-expression group (p < 0.05, Fig. 1E, F). Clinicopathological correlation analysis (Table 2) revealed that elevated USP5 expression was associated with aggressive tumor features, including larger tumor sizes (p = 0.003), higher incidence of satellite nodules (p = 0.001), and tumor emboli (p = 0.001). However, no significant associations were observed with age, sex, alpha-fetoprotein levels, HBV DNA load, tumor differentiation, or liver cirrhosis status. Notably, multivariate Cox regression identified USP5 overexpression as an independent prognostic factor for poor HCC outcomes (Supplementary Fig. 1B). Together, our integrated analysis of TCGA data and clinical specimens consistently establishes USP5 as both a biomarker of tumor aggressiveness and an independent predictor of unfavorable prognosis in HCC.
USP5 promotes the proliferation, migration and invasion of HCC cells in vitro
To investigate the biological function of USP5 in HCC, we first established USP5-knockdown models using two independent shRNAs in Huh7 and SK-Hep-1 cells, which achieved efficient USP5 silencing as confirmed by western blot analysis (Fig. 2A). Functional characterization revealed that USP5 depletion significantly impaired HCC cell proliferation, as evidenced by both CCK-8 assays (Fig. 2B) and colony formation assays (Fig. 2C), while transwell migration and Matrigel invasion assays demonstrated marked reductions in cell motility and invasive capacity (Fig. 2D). Conversely, USP5 overexpression in these cell lines promoted proliferative, migratory and invasive phenotypes (Fig. 3A–D). These experimental findings were further substantiated by TCGA database analysis, which revealed significant positive correlations between USP5 expression and key oncogenic processes, including DNA replication, tumor proliferation markers, and G2/M cell cycle checkpoint regulation (Supplementary Fig. 2A–C), collectively supporting an oncogenic role for USP5 in HCC progression.

A Western blot analysis demonstrating efficient USP5 knockdown in Huh7 and SK-Hep-1 cells transduced with two independent USP5-targeting shRNAs (shUSP5-1, shUSP5-2) compared to negative control shRNA (shNC). B Proliferation curves of control and USP5-knockdown cells measured by CCK-8 assay. C Colony formation ability following USP5 depletion. Left: Representative images of crystal violet-stained colonies. Right: Quantification of colony numbers. D Metastatic capacity following USP5 depletion was assessed by transwell assay. All data represent mean ± SD from three independent experiments. **p < 0.01, ***p < 0.001.

A Western blot validation of USP5 overexpression in Huh7 and SK-Hep-1 cells transduced with either empty vector (NC) or USP5-overexpressing lentivirus. β-Actin served as the loading control. B Proliferation curves of control and USP5-overexpressing cells measured by CCK-8 assay. C Representative images and quantitative analysis of cell clones formed by control and USP5-overexpressing cells. D Metastatic capacity following USP5 overexpression was assessed by transwell assay. Data are presented as mean ± SD from three independent experiments. **p < 0.01, ***p < 0.001.
USP5 regulates lipid accumulation in HCC cells
The role of USP5 in regulating lipid metabolism during HCC progression remains poorly understood. Intriguingly, our analysis of TCGA data revealed a significant negative correlation between USP5 expression and fatty acid degradation pathways (Fig. 4A), suggesting its potential involvement in lipid metabolic reprogramming. To experimentally validate this finding, we performed Oil Red O staining in USP5-knockdown Huh7 and SK-Hep-1 cells, which demonstrated significantly reduced lipid accumulation compared to controls (Fig. 4B). This observation was further corroborated by Nile Red staining, showing decreased intracellular lipid droplet formation following USP5 depletion (Fig. 4C).

A Negative correlation between USP5 expression and fatty acid degradation in the TCGA cohort. B Oil Red O staining showing significantly reduced lipid accumulation in Huh7 and SK-Hep-1 cells following USP5 knockdown compared to negative control. C Nile Red staining demonstrating decreased intracellular lipid droplet accumulation in HCC cells after USP5 downregulation. D The lipidomic profiling alteration occasioned by USP5 knockdown in Huh7 cells. E Cell proliferation was assessed in Huh7 cells transduced with shNC or shUSP5 and treated with either vehicle or 1 μM PA using CCK-8 assay. F Representative images (left) and quantification (right) of transwell assays performed in Huh7 cells transduced with shNC or shUSP5 and treated with either vehicle or 1 μM PA. All data represent mean ± SD from three independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001.
Comprehensive lipidomic profiling of control and USP5-silenced cells uncovered distinct alterations in fatty acid composition. Specifically, USP5 knockdown led to: (1) reduced levels of saturated fatty acids (SFAs), particularly palmitic acid (PA); and (2) increased abundance of polyunsaturated fatty acids (PUFAs), including linoleic acid (Fig. 4D). Given PA’s established role as an oncogenic lipid, we hypothesized it might mediate USP5’s pro-tumorigenic effects. Rescue experiments confirmed this hypothesis, as PA supplementation was capable of partially restoring the proliferative, migratory, and invasive capacities that were impaired by USP5 silencing (Fig. 4E, F). These findings establish USP5 as a novel regulator of lipid metabolism in HCC and identify PA as an important downstream effector mediating its oncogenic functions.
USP5 interacts with FASN and influences its stability via the ubiquitin-proteasome pathway
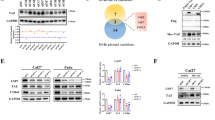
As a deubiquitinating enzyme, USP5 plays a pivotal role in modulating the ubiquitin-proteasome system through its cleavage of ubiquitin moieties from polyubiquitin chains. To elucidate the molecular mechanisms underlying USP5’s promotion of PA production and HCC progression, we performed comprehensive proteomic analyses. Co-immunoprecipitation (co-IP) coupled with mass spectrometry identified 896 potential USP5-interacting proteins (Fig. 5A). Parallel ubiquitination proteomics revealed 650 proteins exhibiting enhanced ubiquitination upon USP5 knockdown in Huh7 cells. Integration of these datasets yielded 126 high-confidence candidate targets likely regulated by USP5-mediated deubiquitination (Fig. 5B).

A Co-immunoprecipitation followed by mass spectrometry analysis of HEK-293T cells expressing exogenous USP5 identified 896 interacting proteins in total. The top five most abundant interacting proteins (excluding USP5 itself) are presented. B Comparative analysis revealed that among 650 proteins showing increased ubiquitination upon USP5 knockdown (by ubiquitin proteomics), 126 overlapped with the USP5-interacting proteins identified (Venn diagram representation). C Interaction between USP5 and FASN was confirmed by co-immunoprecipitation (co-IP) in both Huh7 and SK-Hep-1 cell lines. D Western blot analysis demonstrated that USP5 knockdown or overexpression significantly altered FASN protein levels.
Notably, FASN emerged as a top-ranking USP5-interacting protein, demonstrating markedly increased ubiquitination following USP5 depletion. This finding is particularly significant given FASN’s established role as the rate-limiting enzyme in lipid biosynthesis, where it catalyzes long-chain fatty acid production - especially palmitic acid (PA) - and serves as the central regulator of de novo lipogenesis [11]. These observations perfectly align with our above findings regarding USP5’s regulatory function in lipid metabolism. To further investigate FASN as a key downstream effector of USP5 in HCC progression, we conducted validation experiments. Co-IP assays confirmed a physical interaction between USP5 and FASN (Fig. 5C). To further substantiate this finding, binding between USP5 and FASN was also validated using purified recombinant proteins (Supplementary Fig. 3A) Intriguingly, while USP5 manipulation did not significantly affect FASN mRNA levels (Supplementary Fig. 3B). However, the protein expression of FASN showed a consistent decrease or increase trend following USP5 silencing or overexpression, respectively (Fig. 5D), indicating that USP5 was involved in the degradation of FASN.
To elucidate the mechanism underlying FASN degradation, we treated HCC cells with the proteasome inhibitor MG132. Notably, MG132 treatment effectively reversed the reduction in FASN protein levels caused by USP5 knockdown (Fig. 6A), suggesting proteasomal degradation as the primary pathway for FASN turnover. To further assess whether USP5 regulates FASN protein stability, we performed cycloheximide (CHX) chase experiments in Huh7 and SK-Hep-1 cells with control and USP5-depletion. The results demonstrated that USP5 deficiency significantly accelerated FASN protein degradation (Fig. 6B), supporting a critical role for USP5 in maintaining FASN stability. Consistent with these findings, ubiquitination assays revealed that USP5 knockdown substantially increased FASN ubiquitination, whereas USP5 overexpression reduced it (Fig. 6C, D), indicating that USP5 directly modulates FASN ubiquitination status. Finally, IHC analysis of clinical HCC specimens revealed a strong positive correlation between USP5 and FASN expression levels (Fig. 6E), further reinforcing their functional relationship. Collectively, these results demonstrate that USP5 stabilizes FASN protein by antagonizing its ubiquitination and subsequent proteasomal degradation in HCC cells.

A The control and USP5 knockdown Huh7 and SK-Hep-1 cells were treated with vehicle or the proteasome inhibitor MG132 (10 μM) for 10 h, and whole-cell lysates were prepared and subjected to immunoblotting. FASN was detected with β-actin serving as the loading control. B Cycloheximide (CHX, 20 μg/mL) chase experiments in control and USP5-knockdown cells showed time-dependent changes in FASN protein stability (upper: representative blots of FASN and β-actin; bottom: quantification of FASN/β-actin ratios). C, D Huh7 (C) and SK-Hep-1 (D) cells with USP5 knockdown or overexpression were treated with MG132 (10 μM, 10 h), followed by immunoprecipitation with an anti-FASN antibody and immunoblotting with the indicated antibodies. The levels of FASN ubiquitination were quantified. E Representative IHC staining of USP5 and FASN in sequential tissue sections (left). Scatter plot showing coordinated expression patterns across specimens, with statistical significance determined by Pearson correlation analysis (right).
The oncogenic function of USP5 is partially mediated by FASN in HCC
To investigate the functional consequences of USP5-mediated FASN regulation, we conducted rescue experiments by depleting FASN in USP5-overexpressing Huh7 cells (Fig. 7A). Functional characterization using CCK-8 proliferation assays, colony formation tests, and transwell migration/invasion analyses consistently demonstrated that FASN knockdown significantly attenuated, though did not completely abolish, the enhanced proliferative capacity, clonogenic potential, and metastatic behavior induced by USP5 overexpression (Fig. 7B-D). These results indicate that FASN serves as a crucial downstream effector mediating, at least in part, the oncogenic functions of USP5 in HCC progression.

A USP5-overexpressed cells were infected with lentiviral particles expressing shRNA targeting FASN. B CCK-8 assays demonstrated that FASN knockdown attenuated the pro-proliferative effects mediated by USP5 overexpression in Huh7 cells. C Colony formation assays confirmed that inhibition of FASN abrogated the enhanced proliferative capacity induced by USP5 overexpression. D Transwell migration and Matrigel invasion assays revealed that FASN downregulation markedly reduced the USP5-mediated enhancement of metastatic potential in Huh7 cells. Data are presented as mean ± SD from three independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001.
Downregulation of USP5 inhibits the growth of HCC in vivo
To validate the oncogenic role of USP5 in vivo, xenograft models were established by subcutaneous injection of control (shNC), USP5-knockdown (shUSP5), or USP5-knockdown with FASN-overexpressing (shUSP5+FASN) Huh7 cells into nude mice. Tumors derived from USP5-deficient cells exhibited markedly reduced volume and weight compared with control tumors, whereas ectopic expression of FASN partially rescued this growth inhibition (Fig. 8A, B). Immunohistochemical analysis further confirmed that USP5 depletion led to decreased FASN and Ki67 expression, while FASN overexpression restored both markers to intermediate levels (Fig. 8C). These findings provide in vivo evidence that USP5 promotes HCC progression by stabilizing FASN, highlighting the critical role of the USP5–FASN axis in tumor growth regulation.

A Representative photographs (left) and growth curves (right) of subcutaneous xenograft tumors derived from control (shNC), USP5-knockdown (shUSP5), and USP5-knockdown with FASN-overexpression (shUSP5+FASN) Huh7 cells. Tumor volumes were measured every 4 days using calipers (length × width²/2). B Quantitative analysis of excised tumor weights at study endpoint. C Representative immunohistochemical (IHC) staining of USP5, FASN, and the proliferation marker Ki67 in paraffin-embedded xenograft tumor sections derived from shNC, shUSP5, and shUSP5+FASN Huh7 cells. Quantification of staining density is shown on the right. Data are presented as mean ± SD. *p < 0.05, **p < 0.01.
Discussion
Emerging evidence has established USP5 as a critical oncoprotein across multiple malignancies, with its overexpression correlating with poor prognosis in glioma, ovarian cancer, osteosarcoma, pancreatic ductal adenocarcinoma, bladder cancer, HCC, and cholangiocarcinoma [25, 31,32,33,34,35,36]. Mechanistically, USP5 promotes tumorigenesis through diverse pathways, including enhancing proliferation, maintaining stemness, facilitating metastasis, modulating inflammatory responses, enabling immune evasion, and suppressing ferroptosis by deubiquitinating various protein substrates [30, 37,38,39]. Our current study further reinforces the clinical significance of USP5 in HCC, demonstrating its upregulation in patient specimens and strong association with aggressive clinicopathological features (larger tumor size, satellite nodules, and vascular invasion) as well as unfavorable outcomes. Notably, multivariate analysis identified USP5 as an independent prognostic factor for overall survival in HCC patients. Functionally, we elucidated that USP5 drives HCC progression by promoting malignant phenotypes and reprogramming lipid metabolism. These findings position USP5 as a promising prognostic biomarker and therapeutic target in cancer. Importantly, the recent development of specific USP5 inhibitors (degrasyn, WP1130, and EOAI) showing potent anti-tumor efficacy in preclinical models suggests that pharmacological targeting of USP5 could represent a novel therapeutic strategy for HCC treatment [30, 40,41,42]. The convergence of USP5’s multifaceted oncogenic roles and the availability of targeted inhibitors underscores its translational potential in cancer therapy.
Recent studies have identified several key functional substrates of USP5 in HCC, revealing its multifaceted role in tumor progression through distinct molecular mechanisms. First, USP5 stabilizes SLUG, a master regulator of EMT, through deubiquitination, thereby enhancing the metastatic potential of HCC cells [25]. Second, USP5 interacts with LSH, a crucial DNA methylation modulator, maintaining its protein stability via deubiquitination and subsequently upregulating SLC7A11 to protect HCC cells from ferroptosis [30]. Most notably, USP5 plays a pivotal role in the metabolic reprogramming of HCC cells by stabilizing the oncoprotein c-Myc. Through inhibition of K48-linked polyubiquitination, USP5 prevents c-Myc degradation and consequently activates multiple glycolytic enzymes, including lactate dehydrogenase A (LDHA), enolase 1 (ENO1), triosephosphate isomerase 1 (TPI1), glucose transporter 1 (SLC2A1/GLUT1), and pyruvate kinase M2 (PKM2) [28]. In addition to its roles in glycolysis, USP5 has been implicated in the regulation of lipid metabolism. A recent study revealed that LRP11 recruits USP5 to facilitate the deubiquitination and stabilization of RACK1, thereby promoting lipid accumulation in HCC cells [43]. However, the precise mechanism by which RACK1 influences this process remains unclear. Furthermore, it is still unknown whether USP5 directly targets key lipid metabolism regulators—such as FASN, acetyl-CoA carboxylase (ACC), ATP-citrate lyase (ACLY), and acyl-CoA synthetase long chain family member 4 (ACSL4). These findings suggest that USP5 coordinates a multifaceted oncogenic network involving EMT, epigenetic regulation, glycolysis, and—most notably—lipid metabolic reprogramming. This dimension of USP5 function remains largely unexplored and represents a major focus of the present study.
The dysregulation of lipid metabolism has emerged as a hallmark of cancer progression, with growing recognition of its critical role in tumorigenesis and metastasis. Central to this process are three key lipogenic enzymes - ACLY, ACC, and FASN - which are frequently overexpressed or hyperactivated across various malignancies to fuel cancer cell proliferation, invasion, and dissemination [44, 45]. In aggressive cancers, the intensification of FA synthesis has been widely depicted as a mechanism for generating cell proliferative membranes as well as regulating membrane composition and engendering fat stores. For instance, the induction of de novo lipogenesis via FASN in cancer cells is associated with lymph node metastasis and can induce migration and invasion in vitro. Cancer cells also promote the storage of newly synthesized lipids into lipid droplets as an energy reservoir to facilitate cancer cell dissemination. In order to proliferate in new niches, metastatic cells may require enhanced lipid biosynthesis [11, 46, 47]. Notably, FASN-mediated de novo lipogenesis has been specifically implicated in promoting lymph node metastasis and facilitating migratory capacity in vitro. Metastatic cells appear to possess an inherent epigenetic predisposition to upregulate lipogenic factors like FASN, explaining their consistent association with poor prognosis and multidrug resistance in clinical settings [48, 49]. These findings have spurred clinical interest in repurposing anti-obesity agents (e.g., orlistat) and developing novel FASN inhibitors as potential anticancer therapies, with several compounds currently under clinical evaluation [50,51,52]. To date, whether USP5 regulates lipid metabolism of HCC remains unclear. FASN protein levels can be regulated by the ubiquitination/deubiquitination pathway. For example, FBXW7β serves as an E3 ligase of FASN, leading to its degradation and suppressing colorectal cancer growth [53]. The deubiquitinating enzyme USP14 directly interacts with and increases FASN stability, which promotes liver triglyceride accumulation [14]. Our study provides novel insights into this regulatory network by identifying USP5 as a critical regulator of FASN in HCC. We demonstrate that USP5: (1) promotes lipid droplet accumulation and increases oncogenic PA levels; (2) physically interacts with FASN to reduce its ubiquitination and prolong its half-life; and (3) drives HCC progression through the FASN and PA. However, several mechanistic questions remain unresolved, including: (1) the specific ubiquitination sites on FASN targeted by USP5; (2) the particular polyubiquitin chain linkages (e.g., K48 vs K63) modulated by USP5; and (3) potential crosstalk between USP5 and other known FASN regulators like USP14. Addressing these questions will be crucial for developing targeted interventions against this pathway in HCC.
Conclusion
In conclusion, the results of our present study demonstrate that USP5 plays an important role in the progression of HCC via deubiquitination-mediated stabilization of FASN protein and subsequent enhancement of PA production. These findings not only reveal a novel USP5-FASN-PA regulatory axis driving HCC progression but also highlight USP5 as a promising molecular target for therapeutic intervention.
Ethics approval and consent to participate
The study received approval from the Ethics Committee of Zhongshan Hospital of Xiamen University, and informed consent was obtained from the participating patients. The animal study was approved by the Animal Care and Use Committee of Xiamen University, ensuring adherence to the fundamental principles of animal experimentation.
Data availability
The data generated in this study are available within the article and its Supplementary Data files. All other raw data are available upon a reasonable inquiry from the corresponding author.
References
Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73:17–48.
Villanueva A. Hepatocellular Carcinoma. N Engl J Med. 2019;380:1450–62.
Sun L, Zhang H, Gao P. Metabolic reprogramming and epigenetic modifications on the path to cancer. Protein Cell. 2022;13:877–919.
Hu B, Lin JZ, Yang XB, Sang XT. Aberrant lipid metabolism in hepatocellular carcinoma cells as well as immune microenvironment: A review. Cell Prolif. 2020;53:e12772.
Nakagawa H, Hayata Y, Kawamura S, Yamada T, Fujiwara N, Koike K. Lipid Metabolic Reprogramming in Hepatocellular Carcinoma. Cancers (Basel). 2018;10:447.
Sangineto M, Villani R, Cavallone F, Romano A, Loizzi D, Serviddio G. Lipid Metabolism in Development and Progression of Hepatocellular Carcinoma. Cancers (Basel). 2020;12:1419.
Bian Z, Xu C, Wang X, Zhang B, Xiao Y, Liu L, et al. TRIM65/NF2/YAP1 Signaling Coordinately Orchestrates Metabolic and Immune Advantages in Hepatocellular Carcinoma. Adv Sci (Weinh). 2024;11:e2402578.
Pascual G, Dominguez D, Elosua-Bayes M, Beckedorff F, Laudanna C, Bigas C, et al. Dietary palmitic acid promotes a prometastatic memory via Schwann cells. Nature. 2021;599:485–90.
Wang J, Shen D, Jiang J, Hu L, Fang K, Xie C, et al. Dietary Palmitic Acid Drives a Palmitoyltransferase ZDHHC15-YAP Feedback Loop Promoting Tumor Metastasis. Adv Sci (Weinh). 2025;12:e2409883.
Jin Q, Qi D, Zhang M, Qu H, Dong Y, Sun M, et al. CLDN6 inhibits breast cancer growth and metastasis through SREBP1-mediated RAS palmitoylation. Cell Mol Biol Lett. 2024;29:112.
Rohrig F, Schulze A. The multifaceted roles of fatty acid synthesis in cancer. Nat Rev Cancer. 2016;16:732–49.
Buckley D, Duke G, Heuer TS, O’Farrell M, Wagman AS, McCulloch W, et al. Fatty acid synthase - Modern tumor cell biology insights into a classical oncology target. Pharmacol Ther. 2017;177:23–31.
Chaturvedi S, Biswas M, Sadhukhan S, Sonawane A. Role of EGFR and FASN in breast cancer progression. J Cell Commun Signal. 2023;17:1249–82.
Liu B, Jiang S, Li M, Xiong X, Zhu M, Li D, et al. Proteome-wide analysis of USP14 substrates revealed its role in hepatosteatosis via stabilization of FASN. Nat Commun. 2018;9:4770.
Wang H, Zhou Y, Xu H, Wang X, Zhang Y, Shang R, et al. Therapeutic efficacy of FASN inhibition in preclinical models of HCC. Hepatology. 2022;76:951–66.
Li Y, Yang W, Zheng Y, Dai W, Ji J, Wu L. et al. Targeting fatty acid synthase modulates sensitivity of hepatocellular carcinoma to sorafenib via ferroptosis. J Exp Clin Cancer Res. 2023;42:6.
Ni M, Zhang B, Zhao J, Feng Q, Peng J, Hu Y, et al. Biological mechanisms and related natural modulators of liver X receptor in nonalcoholic fatty liver disease. Biomed Pharmacother. 2019;113:108778.
Yahagi N, Shimano H, Hasegawa K, Ohashi K, Matsuzaka T, Najima Y, et al. Co-ordinate activation of lipogenic enzymes in hepatocellular carcinoma. Eur J Cancer. 2005;41:1316–22.
Mo Y, Han Y, Chen Y, Fu C, Li Q, Liu Z, et al. ZDHHC20 mediated S-palmitoylation of fatty acid synthase (FASN) promotes hepatocarcinogenesis. Mol Cancer. 2024;23:274.
Ni W, Lin S, Bian S, Zheng W, Qu L, Fan Y, et al. USP7 mediates pathological hepatic de novo lipogenesis through promoting stabilization and transcription of ZNF638. Cell Death Dis. 2020;11:843.
Chen YJ, Wu H, Shen XZ. The ubiquitin-proteasome system and its potential application in hepatocellular carcinoma therapy. Cancer Lett. 2016;379:245–52.
Dewson G, Eichhorn PJA, Komander D. Deubiquitinases in cancer. Nat Rev Cancer. 2023;23:842–62.
Lange SM, Armstrong LA, Kulathu Y. Deubiquitinases: From mechanisms to their inhibition by small molecules. Mol Cell. 2022;82:15–29.
Liu Q, Wu Y, Qin Y, Hu J, Xie W, Qin FX, et al. Broad and diverse mechanisms used by deubiquitinase family members in regulating the type I interferon signaling pathway during antiviral responses. Sci Adv. 2018;4:eaar2824.
Meng J, Ai X, Lei Y, Zhong W, Qian B, Qiao K, et al. USP5 promotes epithelial-mesenchymal transition by stabilizing SLUG in hepatocellular carcinoma. Theranostics. 2019;9:573–87.
Nakajima S, Lan L, Wei L, Hsieh CL, Rapic-Otrin V, Yasui A, et al. Ubiquitin-specific protease 5 is required for the efficient repair of DNA double-strand breaks. PLoS One. 2014;9:e84899.
Nostramo R, Varia SN, Zhang B, Emerson MM, Herman PK. The Catalytic Activity of the Ubp3 Deubiquitinating Protease Is Required for Efficient Stress Granule Assembly in Saccharomyces cerevisiae. Mol Cell Biol. 2016;36:173–83.
Xia P, Zhang H, Lu H, Xu K, Jiang X, Jiang Y, et al. METTL5 stabilizes c-Myc by facilitating USP5 translation to reprogram glucose metabolism and promote hepatocellular carcinoma progression. Cancer Commun (Lond). 2023;43:338–64.
Jiang S, Jiang L, Xu Y, Ma Y, Deng Y, Jiao C, et al. USP5 deubiquitinates and stabilizes IMPDH2, to promote hepatocellular carcinoma progression. Oncogene. 2025;44:1936–48.
Yan B, Guo J, Wang Z, Ning J, Wang H, Shu L, et al. The ubiquitin-specific protease 5 mediated deubiquitination of LSH links metabolic regulation of ferroptosis to hepatocellular carcinoma progression. MedComm (2020). 2023;4:e337.
Cai H, Ke ZB, Chen JY, Li XD, Zhu JM, Xue YT, et al. Ubiquitin-specific protease 5 promotes bladder cancer progression through stabilizing Twist1. Oncogene. 2024;43:703–13.
Du Y, Lin J, Zhang R, Yang W, Quan H, Zang L, et al. Ubiquitin specific peptidase 5 promotes ovarian cancer cell proliferation through deubiquitinating HDAC2. Aging (Albany NY) 2019;11:9778–93.
Jiang X, You H, Niu Y, Ding Y, Chen Z, Wang H, et al. E2F1-regulated USP5 contributes to the tumorigenic capacity of glioma stem cells through the maintenance of OCT4 stability. Cancer Lett. 2024;593:216875.
Li J, Li H, Zhu W, Zhou B, Ying J, Wu J, et al. Deubiquitinase inhibitor degrasyn suppresses metastasis by targeting USP5-WT1-E-cadherin signalling pathway in pancreatic ductal adenocarcinoma. J Cell Mol Med. 2020;24:1370–82.
Ning F, Du L, Li J, Wu T, Zhou J, Chen Z, et al. The deubiquitinase USP5 promotes cholangiocarcinoma progression by stabilizing YBX1. Life Sci. 2024;348:122674.
Wu Q, Liu R, Yang Y, Peng J, Huang J, Li Z, et al. USP5 promotes tumorigenesis by activating Hedgehog/Gli1 signaling pathway in osteosarcoma. Am J Cancer Res. 2024;14:1204–16.
Pan J, Qiao Y, Chen C, Zang H, Zhang X, Qi F, et al. USP5 facilitates non-small cell lung cancer progression through stabilization of PD-L1. Cell Death Dis. 2021;12:1051.
Shi S, Pan X, Chen M, Zhang L, Zhang S, Wang X. et al. USP5 promotes lipopolysaccharide-induced apoptosis and inflammatory response by stabilizing the TXNIP protein. Hepatol Commun. 2023;7:e0193.
Xiao X, Shi J, He C, Bu X, Sun Y, Gao M, et al. ERK and USP5 govern PD-1 homeostasis via deubiquitination to modulate tumor immunotherapy. Nat Commun. 2023;14:2859.
Cheung BB, Kleynhans A, Mittra R, Kim PY, Holien JK, Nagy Z, et al. A novel combination therapy targeting ubiquitin-specific protease 5 in MYCN-driven neuroblastoma. Oncogene. 2021;40:2367–81.
Kapuria V, Peterson LF, Fang D, Bornmann WG, Talpaz M, Donato NJ. Deubiquitinase inhibition by small-molecule WP1130 triggers aggresome formation and tumor cell apoptosis. Cancer Res. 2010;70:9265–76.
Zheng Y, Wang L, Niu X, Guo Y, Zhao J, Li L, et al. EOAI, a ubiquitin-specific peptidase 5 inhibitor, prevents non-small cell lung cancer progression by inducing DNA damage. BMC Cancer. 2023;23:28.
Liang L, Jia W, Wang J, Feng Y, Zhu D, Zhao W, et al. LRP11 facilitates lipid metabolism and malignancy in hepatocellular carcinoma by stabilizing RACK1 through USP5 regulation. Mol Med. 2025;31:35.
Bian X, Liu R, Meng Y, Xing D, Xu D, Lu Z. Lipid metabolism and cancer. J Exp Med. 2021;218:e20201606.
Li C, Zhang L, Qiu Z, Deng W, Wang W. Key Molecules of Fatty Acid Metabolism in Gastric Cancer. Biomolecules. 2022;12:706.
Hoy AJ, Nagarajan SR, Butler LM. Tumour fatty acid metabolism in the context of therapy resistance and obesity. Nat Rev Cancer. 2021;21:753–66.
Koundouros N, Poulogiannis G. Reprogramming of fatty acid metabolism in cancer. Br J Cancer. 2020;122:4–22.
Menendez JA, Lupu R. Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat Rev Cancer. 2007;7:763–77.
Menendez JA, Lupu R. Fatty acid synthase: a druggable driver of breast cancer brain metastasis. Expert Opin Ther Targets. 2022;26:427–44.
Du Q, Liu P, Zhang C, Liu T, Wang W, Shang C, et al. FASN promotes lymph node metastasis in cervical cancer via cholesterol reprogramming and lymphangiogenesis. Cell Death Dis. 2022;13:488.
Lupu R, Menendez JA. Pharmacological inhibitors of Fatty Acid Synthase (FASN)—catalyzed endogenous fatty acid biogenesis: a new family of anti-cancer agents?. Curr Pharm Biotechnol. 2006;7:483–93.
Schcolnik-Cabrera A, Chavez-Blanco A, Dominguez-Gomez G, Taja-Chayeb L, Morales-Barcenas R, Trejo-Becerril C, et al. Orlistat as a FASN inhibitor and multitargeted agent for cancer therapy. Expert Opin Investig Drugs. 2018;27:475–89.
Wei W, Qin B, Wen W, Zhang B, Luo H, Wang Y, et al. FBXW7beta loss-of-function enhances FASN-mediated lipogenesis and promotes colorectal cancer growth. Signal Transduct Target Ther. 2023;8:187.
Funding
This work was supported by the Natural Science Foundation of Xiamen (No. 3502Z202373100 and No. 3502Z202373140), the Fujian Provincial Natural Science Foundation of China (No.2022J011350, No.2022J02056, No. 2023D001, and No. 2025J011413).
Author information
Authors and Affiliations
Contributions
Conceptualization: CX, ZY, and FW. Methodology: CX, ZY, FW, and HW. Investigation: QF, CL, XC, and PZ. Software, analysis pipeline: QY, and YL. Visualization: QF and YL. Resources: CX, ZY, and QF Supervision: CX, ZY, and HW. Funding acquisition: QF, CX, and ZY. Writing: CX and YL with inputs from all authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Fang, Q., Luo, C., Lu, Y. et al. Stabilization of FASN by USP5-mediated deubiquitination promotes hepatocellular carcinoma progression. Oncogenesis 14, 46 (2025). https://doi.org/10.1038/s41389-025-00589-8
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41389-025-00589-8
