
Abstract
The resistance to radiotherapy of prostate cancer is driven by interactions within the tumor microenvironment, particularly between prostate cancer cells and tumor-associated macrophages, however the underlying mechanisms remain poorly understood. In this study, we found that STAT1 enhanced the transcription of critical glycolytic enzymes, leading to an increase in lactate secretion from prostate cancer cells. Then, the lactate was transported to macrophages via the MCT1 transporter, activating the NFκB1 pathway, which subsequently promoted macrophage polarization to the M2 phenotype and activated the transcription of MCP-1. MCP-1 was secreted from macrophages interacted with the CCR2 receptor on prostate cancer cells, thereby activating the JAK/STAT1 pathway, ultimately contributing to the progression of prostate cancer and its resistance to radiotherapy. Taken together, our findings identified a STAT1/lactate/NFκB1/MCP-1 positive feedback mechanism as a driver of prostate cancer progression and resistance to radiotherapy that functioned by interaction to macrophages, which could be potential therapeutic targets for the advanced prostate cancer.
Similar content being viewed by others


Introduction
Prostate cancer (PCa) has become the most frequent malignancy in males worldwide, with a continuously increasing incidence in recent years [1]. Radical radiotherapy (RT) was still regarded as one of the standard therapeutic strategies for localized PCa, however, the proportion of initially diagnosed PCa patients was recognized as advanced PCa [2]. In terms of pathology, clinical biology and genetics, high heterogeneity was presented in advanced PCa, which contributed to an unsatisfied response to radiotherapy and induced a worse prognosis furtherly [3]. Previous studies have proposed that DNA abnormal repair and oxidative stress might be involved in resistance to radiotherapy [4], but the exact mechanism remains poorly understood. It is of vital importance to investigate the mechanism underlying resistance to radiotherapy of PCa.
Tumor microenvironment (TME), collectively constituted of cancer cells, immune cells, fibroblasts, signal molecules and extracellular matrix, was considered as the major role in tumor progression [3, 5]. Tumor-associated macrophages (TAMs) are pivotal elements of tumor immune microenvironment (TIME). It was well known that macrophages polarized to M1 or M2 by induction of different cytokines. M1 macrophages exerted anti-tumor effects in the early stage of tumor development, while M2 macrophages exerted pro-tumorigenic effects by remodeling immunosuppressive microenvironments [6, 7]. It was found that PCa patients with poor response to immunotherapy were characterized by high infiltration of M2 macrophages [8]. It was reported that proportion of M2 macrophage infiltration was elevated in process of PCa progression to castration-resistant prostate cancer (CRPC) [9]. In addition, emerging research evidence on the effects of macrophages in TME on resistance to radiotherapy has been increasing. Different radiation doses and modalities induced different macrophage phenotypes that established sensitivity to radiotherapy [10, 11]. It was discovered previously that macrophages regulated immune environment to modulate sensitivity to radiotherapy by secreting chemokines or altering surface antigens [12, 13]. However, there is still a substantial gap in our knowledge of the regulatory mechanism of macrophages in resistance to radiotherapy of PCa.
Aerobic glycolysis is imperative for the progression of PCa. Compared to normal prostate epithelial cells, PCa cells presented a lower oxygen consumption, but a higher rate of glucose uptake and lactate production. Under aerobic conditions, tumor cells quickly generated a large amount of adenosine triphosphate by aerobic glycolysis instead of oxidative phosphorylation. This adaptive metabolic change of tumor cells was known as the Warburg effect [14]. Lactate, a primary products of aerobic glycolysis, was capable of activating endothelial cells for angiogenesis through hypoxia inducible factor (HIF) dependent and HIF independent pathways as well as remodeling suppressive TIME for immune escape [15, 16]. Researches has demonstrated that M2 polarization was induced via lactate-Gp132 signal in breast cancer and high concentration of lactate was accompanied by increased infiltration of M2 macrophages in TME of head and neck squamous cancer [17, 18]. Nevertheless, the mechanism of communication between lactate derived from prostate cancer cells and macrophages remains incomplete.
Signal transducers and activators of transcription 1 (STAT1) acted as a transcription factor, was involved in various biological processes including cell proliferation, differentiation and metabolism by means of enhancing transcription for target genes [19, 20]. A range of research has reported that STAT1 participates in several diverse processes of tumorigenesis, progression and resistance to therapy in multiple cancer types, including breast cancer, prostate cancer and lung cancer [21,22,23]. In early stage, of PCa, STAT1 was identified as a tumor suppressor with evidence suggesting that high STAT1 expression was correlated to longer cancer-specific survival of patients with localized PCa, conversely, increasing amounts of studies indicated activation of STAT1 was accountable for resistance to endocrine therapy and chemotherapy in advanced PCa [24,25,26,27,28,29]. Overall, the role of STAT1 in malignant progression and resistance to radiotherapy of PCa and its exact mechanism remain poorly understood.
This study concentrated on the interaction between PCa cells and macrophages in TME, intending to explore the effect of STAT1 on PCa progression and resistance to radiotherapy and its molecular mechanism through in vitro and in vivo experiments. Meanwhile, protein microarray coupled with in vitro validation revealed that monocyte chemotactic protein 1 (MCP-1) was the key molecule for maintaining signal transmission between PCa cells and macrophages. Furthermore, we aimed to provide mechanistic insight into the positive feedback regulation on the janus kinase (JAK)/STAT1 pathway in PCa cells by MCP-1.
Material and methods
Cell culture
Normal human epithelial cells (RWPE-1, CL-0200), human prostate cancer cell lines including DU145 (CL-0075), PC-3 (CL-0185), 22RV1 (CL-0004), LNCaP (CL-0143), human embryonic kidney cells (293T, CL-0005) and mouse prostate cancer cells (RM-1, CL-0198) were purchased from Procell Life Science & Technology Co., Ltd (Wuhan, China). Human prostate cancer cell lines, C4-2 (HTX1927) and C4-2B (HTX1928), were purchased from Otwo Biotech Inc. (Shenzhen, China). Human leukemia monocytic cell line (THP-1, TCH-C361) was purchased from Haixing Biosciences Co., Ltd (Suzhou, China). RWPE-1, 22RV1, C4-2, C4-2B, LNCaP, THP-1 and RM-1 were cultured with RPMI-1640 medium added with 10% fetal bovine serum (FBS). DU145 was cultured with MEM medium added with 10% FBS. PC-3 was cultured with F-12K medium with 10% FBS. 293 T was cultured with DMEM medium supplemented with 10% FBS. All cells were incubated in 5% CO2 at 37 °C. For macrophage differentiation, THP-1 cells were stimulated with 200 nM phorbol 12-myristate 13-acetate (PMA) for 48 h and differentiated into M0 macrophages, which was followed by the subsequent addition of lactate and conditioned medium [30,31,32].
Clinical specimens
PCa tissue and benign prostatic hyperplasia (BPH) tissue were obtained from the hospital. A total of 20 pairs of fresh postoperative prostate cancer tissues and adjacent normal tissues from patients with PCa in the hospital were collected for protein extraction and western blot. All patients signed written informed consent before sample collection.
Generation of radioresistant cells
The radioresistant PC-3 cell line (designated PC-3R) was established from parental human prostate cancer PC-3 cells (designated PC-3S) via repeated cycles of fractionated irradiation. Briefly, when parental PC-3 cells reached 70–80% confluence, they were irradiated with 6 Gy per fraction using an RS2000-Pro X-ray irradiator (Radsource Technologies, Inc., USA). The irradiation procedure was repeated for 6 fractions (cumulative dose 36 Gy), with 48–72 h of recovery between fractions to allow cell repair and repopulation. 30% of the irradiated cells were re-seeded, and subsequent irradiation was performed when the cells reached 80% confluence again.
Radiation clonogenic survival assay
The clonogenic assay was performed to compare the radiosensitivity of PC-3R and PC-3S cell lines. Briefly, 100 cells were seeded per well in 12-well plates and allowed to adhere for 6–8 h. The cells were then irradiated with single doses ranging from 0 to 6 Gy (0, 2, 4, 6 Gy). After 10–14 days, cells were fixed with 4% paraformaldehyde and stained with a crystal violet staining solution. The number of colonies was counted (defined as >50 cells), and the survival fractions were calculated based on the plating efficiency of the non-irradiated controls.
RNA-sequencing (RNA-seq)
RNA seq was conducted by Seqhealth Technology Co., Ltd (Wuhan, China). Briefly, total RNA was isolated from radiosensitive PC-3 cells and radioresistant PC-3 cells. Following quality control, the RNA samples were used for library preparation and subsequent high-throughput sequencing. The resulting data were analyzed to identify differentially expressed genes.
RNA inference and lentivirus transduction
Small interfering RNA (siRNA) targeting STAT1 and nuclear factor kappa-B 1 (NFκB1) were constructed by Shanghai GenePharma Co., Ltd (Shanghai, China). The STAT1 overexpression lentivirus and NFκB1 knockdown lentivirus were purchased from Shanghai GENECHEM Co., Ltd (Shanghai, China). siRNA sequences and lentiviral sequences were described in Table S1. Cells were transfected using Lipofectamine™ 2000 reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions, when grown to approximately 70–80% confluences. RNA and protein were extracted 48 h later. Lentivirus transfection was conducted according to the manufacturer’s instructions; cells and patient-derived organoids (PDOs) were selected by puromycin treatment.
Extraction and culture of primary BMDMs
Bone marrow-derived macrophages (BMDMs) were isolated from male c57BL/6J mice, aged between 6 and 8 weeks. After euthanasia, the tibias and femurs were immediately collected and dissected free of surrounding muscle tissue. The bones were sterilized in 70% ethanol, rinsed with PBS, and both ends were cut off. The bone marrow was then flushed out by repeated rinsing with PBS. The bone marrow suspension was centrifuged at 1200 rpm for 5 min. The pellet was resuspended in red blood cell lysis buffer and incubated at room temperature for 3–5 min. The cells were seeded in culture dishes and incubated overnight in complete medium (high-glucose DMEM supplemented with 10% heat-inactivated FBS and 1% penicillin/streptomycin) to remove rapidly adhering cells. The non-adherent cells were collected and re-seeded in new culture dishes with the addition of 50 ng/mL recombinant M-CSF to induce BMDM differentiation.
Quantitative real-time polymerase chain reaction (qRT-PCR)
The total RNA was extracted using FastPure Cell/Tissue Total RNA Isolation Kit (Vazyme). The cDNA was synthesized by HiScript III All-in-one RT SuperMix Perfect for qPCR (Vazyme). Then, the relevant products were amplified by Taq Pro Universal SYBR qPCR Master Mix (Vazyme) using QuantStudio™ 5 real-time PCR system (Applied Biosystems). The relative expression was normalized based on the expression of β-actin. The sequences of primers were described in Table S2.
Western blot
The total proteins from cells and tissues were extracted by RIPA lysis buffer (P0013B, Beyotime). Protein concentration was measured using the bicinchoninic acid (BCA) assay kit (KGB2101-100, KeyGen BioTECH, China). Proteins were separated in 10% SDS-PAGE gels and transferred to polyvinylidene fluoride membranes. Then, the membranes were incubated with primary antibodies overnight at 4 °C, which was followed by incubation with HRP-conjugated goat anti-rabbit or anti-mouse IgG for 1 h. Lastly, the protein bands were developed and exposed using an enhanced chemiluminescence reagent. The primary antibodies were used as indicated in Table S3.
Enzyme-linked immunosorbent assay (ELISA)
ELISA assay was applied to detect the MCP-1 concentration in the supernatant from macrophages. The specific steps were carried out according to the instructions of the MCP-1 ELISA Kit (SEA087Hu, Cloud-Clone Corp).
Glucose uptake assay
Cells were planted in six-well plates at 4 × 105 cells/well and were cultured with glucose-free medium for 2 h. After starvation, fresh medium was replaced. The glucose concentration of cell supernatant was measured using a glucose fluorometric assay kit (E-BC-F037, Elabscience).
Lactate production assay
Cells were planted in six-well plates at 4 × 105 cells/well. After the cells became adherent, the medium was replaced for a further 24h incubation. D-lactate colorimetric assay kit (E-BC-K002-M, Elabscience) was used to analyze lactate content of cell supernatant.
Cellular extracellular acidification rate (ECAR) and oxygen consumption rate (OCR) assays
ECAR and OCR were measured using the Seahorse XF24 Extracellular Flux Analyzer (Seahorse, USA). DU145 and PC-3 cells pretreated with various conditions were planted into Seahorse XF24 cell plates at 2 × 104/well at 37 °C, 5% CO2 overnight. Then, based on the manufacturer’s protocols, glycolytic flux and mitochondrial metabolic flux were evaluated by a glycolytic stress test kit (Seahorse) and mitochondrial stress test kit (Seahorse), separately.
Immunohistochemistry (IHC)
The PCa tissue and BPH tissue were embedded with paraffin. Paraffin sections were placed oven for 1 h at 60 °C. Then, paraffin sections were deparaffinized with xylene and hydrated with ethanol at different concentration gradients. Antigen repair for sections was performed using citric acid antigen repair buffer. After blocking endogenous peroxidase activity and non-specific staining, sections were incubated with STAT1 antibody (10144-2-AP, Proteintech) or CD163 antibody (83285-4-RR, Proteintech) overnight at 4 °C. The next day, the sections were incubated with biotin-labeled goat anti-mouse/rabbit IgG and Streptomyces anti-biotin protein-peroxidase (KIT-9720, MaiXin.BIO). Subsequently, diaminobenzidine staining and hematoxylin counterstaining were performed. Finally, sections were dehydrated with gradient of ethanol and then made transparent with xylene, followed by sealing with neutral gum.
Dual-luciferase reporter assay
The hexokinase 2(HK2), phosphofructokinase, muscle (PFKM), pyruvate kinase M2 (PKM2), MCP-1 promoter wild type and mutant firefly luciferase reporter plasmids and the renilla luciferase vector plasmids were constructed by Shanghai GenePharma Co., Ltd (Shanghai, China). 293 T cells were co-transfected with plasmids using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA) following the manufacturer’s instructions. After 48 h of co-transfection, firefly luciferase value and renilla luciferase value were determined by Dual-Lumi™ Dual Luciferase Reporter Assay kit (Beyotime Biotechnology, Shanghai, China). The relative luciferase activities were obtained through calculation as firefly fluorescence value/renilla fluorescence value.
CUT&Tag assay
CUT&Tag assay was performed using NovoNGS® CUT&Tag® 4.0 High-Sensitivity Kit (NovoProtein, N259-YH01) based on the manufacturer’s instructions. To put it briefly, cells were treated with formaldehyde to strengthen the binding between transcription factors and DNA. Following the tight binding between cells and ConA beads, p-STAT1 antibody (Cell Signaling Technology, CST#7649) or NFκB1 antibody (Cell Signaling Technology, CST#8242) was added to samples for 2 h. Subsequently, secondary antibodies were added for incubation for 1 h. DNA fragmentation and DNA amplification were performed sequentially. Finally, qRT-PCR was used to analyze enrichment of the target gene promoter by transcription factors.
Immunofluorescence
Cells were seeded in six-well confocal plates at 1 × 105 cells/well. Cells were fixed with 4% paraformaldehyde for 30 min and were incubated with 0.3% Triton X-100 for 20 min. For blocking, 1% bovine serum albumin (BSA) was added for 1 h. Cells were incubated with primary antibody overnight at 4 °C and then incubated with fluorescent dye-conjugated anti-rabbit IgG H&L, protected from light for 1 hour at room temperature. DAPI was utilized for nuclear staining for 15 min. At last, fluorescence images were visualized by fluorescence confocal microscopy.
GSH-CAA-440 protein microarray screening of differentially expressed secreted proteins
GSH-CAA-440 protein microarray and data analysis were performed by H-Wayen biotechnologies (Shanghai, China). The specific operations were described as follows. After completely air drying the glass slide, 100 μl sample diluent was added into each well and incubated at room temperature for 30min to block the slides. Then, 100 μl samples were added to each well and incubated with arrays overnight at 4 °C. After incubation with biotinylated antibody cocktail, 80 μl Cy3 equivalent dye-conjugated streptavidin was added to each well and the slide was incubated protected from light for 1 h. Finally, fluorescent signals were detected by a laser scanner.
Cell counting assay
Cells were seeded in a 96-well plate at 5 × 103 cells/well, and then the plate was placed into an incubator to culture in 5% CO2 at 37 °C. 100 μl cell counting kit-8 (CCK-8) solution formulated by 10 μl CCK-8 regent and 90 μl complete medium was added at every predetermined time point. After incubation for 1 h, the optical density values at 450 nm were read using a microplate reader.
Colony formation assay
Cells were seeded into a 6-well plate at 1 × 103 cells/well. For each group, three repeated wells were set up, and different treatments were conducted. After approximately 10 days of cultivation, the colony formation of cells was observed under a light microscope. Sequentially, the cells were fixed using 4% paraformaldehyde for 30 min, and were stained using 1% crystal violet. Colony formation of cells for each well was captured and counted with the CTL Immunospot analyzer (Cellular Technology Ltd., USA).
Transwell assay
Cells were starved for 48 h with serum-free medium. For the Transwell chambers, 500 μl medium containing 30% fetal bovine serum was added into the lower well, and cells were planted into the upper well at 4 × 104 cells. For the transwell invasive assay, the 20 μg Matrigel Matrix (354234, Corning) was coated in the upper well before planting of cells. The Transwell chambers were placed into 37 °C incubator for 48 h. In order to evaluate the migrative and invasive abilities, the Transwell inserts were taken out, and non-migrated cells in the upper chamber were removed. We use 4% paraformaldehyde and 1% crystal violet for cell fixation and staining separately. At least 3 visual fields were randomly photographed under an inverted microscope.
Wound-healing assay
Cells in logarithmic growth phase were seeded into a 6-well plate at 3 × 105 cells per well. The cells were scratched vertically with a 1 ml gun head when they achieved 90% confluence. The unattached cells were washed off with phosphate buffer saline (PBS). The migrative images of cells were captured at 0 h using an inverted microscope. After that, cells were replaced with fresh serum-free medium and continued to be cultured for 24 h. The cell migration at the same location was recoded at 24 h with an inverted microscope.
Comet assay
Comet assay was performed using the comet assay kit (STA-351, Cell Biolabs, CA, USA). Lysis buffer, alkaline solution and electrophoresis solution were prepared and pre-chilled at 4 °C in advance. Agarose gel was heated in a boiling water bath for 10 min and then cooled in a 37 °C water bath for at least 20 min. The cells (1 × 105/ml) were combined with 75 μl of agarose gel at 37 °C in a 1:10 (v/v) ratio and were spread on the comet slide. The slide was placed in a dark environment for 30 min to solidify. Subsequently, the slides were immersed in lysis buffer at 4 °C for 1 hour, followed by alkaline solution at 4 °C for 30 min. The slides were placed in the electrophoresis chamber and electrophoresed for 30 min at a voltage of 20 V and a current of 300 mA. After electrophoresis, the slides were washed in pre-cooled deionized water and then stained with DAPI for 10 min after drying. Images were captured with a fluorescence microscope (Olympus), and tail moments were analyzed using Comet Assay Software Project (CASP).
In vivo tumorigenesis assay
Five-week-old male athymic BALB/c nude mice and c57BL6/J mice were purchased from Shanghai SLAC Laboratory Animal Co., Ltd (Shanghai, China). Mice were randomly grouped, and each group contained 5 mice. For in vivo assays to evaluate STAT1 functions, DU145 and PC-3 cells with stable STAT1 overexpression or a negative control vector were prepared. For each nude mouse, a total of 5 × 106 cells suspended with 200 μl medium and a hydrogel mixture were inoculated subcutaneously in the subaxillary region of nude mice. Mice were intraperitoneally given different drugs every 3 days, starting on Day 9 after cell injection. For in vivo assays to evaluate MCP-1 functions, DU145 and PC-3 cells were inoculated subcutaneously in the subaxillary region of each nude mouse at 5 × 106 cells, separately. Mice were randomly divided into four groups and were intratumorally given different drug treatments every 3 days, starting on Day 9 after cell injection. Detailed program of drug administration during the experiment period was shown in Table S4 and Fig. S1A. After injection for 9 days, the tumor size was assessed every 3 days simultaneously in a random and blinded fashion to draw tumor growth curve. The tumor volume was calculated as the following formula: tumor volume (V)=π/6×long diameter× short diameter2. After injection for 30 days, nude mice were sacrificed, and the weight of tumors was determined. In the RM-1 xenograft c57BL6/J mouse model, cells were injected subcutaneously into each mice at 5 × 105 cells. Drug treatment or radiation treatment was started on Day 9 following cell injection. Different treatment schemes for each group were described in Table S5 and displayed in Fig. S1B. Mice were sacrificed, and the weight of tumors was detected after injection for 21 days.
Patient-derived organoid (PDO) models
Patient-derived prostate cancer tissue specimens were enzymatically digested and cultured in Matrigel-based 3D matrices to establish PDO models. Upon reaching approximately 100 μm in diameter, organoids in all groups underwent radiation (5 Gy) exposure. Size and morphology were monitored microscopically. PDOs from different groups were seeded into 96-well plates and subjected to the radiation treatments. After 72h, the survival of PDOs was assessed by CelITiter-Glo (CTG) assay.
Statistical analysis
The statistical analyses for data were conducted by GraphPad Prism 9.0 (GraphPad Prism Software, Inc., USA) and SPSS version 26.0 software (IBM Corp, Armonk, NY, USA). The sample sizes were determined with reference to previous publications that detected significant effects using comparable methodologies and group sizes. Besides, sample size power analysis was performed using PASS and G*Power software. All data represent at least three independent experiments. No samples or animals were excluded from the analyses in this study. All experimental subjects met the pre-established health and viability criteria throughout the experimental period. Quantitative data following normal distribution were indicated as mean ± standard deviation (SD), and comparisons between groups were analyze using one-way ANOVA or Students t test. Repeated measures ANOVA was performed to assess repeated measurement data. P < 0.05 was regarded as a statistically significant difference.
Results
STAT1 was highly expressed in prostate cancer and associated with poor outcomes
To identify the potential mechanisms involved in radiotherapy resistance of PCa, we constructed the radioresistant PC-3 cells (PC-3R). PC-3R cells exhibited significantly greater radiation resistance than PC-3S cells, as determined by clonogenic survival assays (Fig. S2A). Furtherly, PC-3R and PC-3S cell samples were used to conduct RNA-seq. Compared to PC-3S cells, STAT1 was identified as the most significantly upregulated gene in PC-3R cells (Fig. S2B). These results preliminarily suggested that STAT1 was closely correlated with radiotherapy resistance of PCa.
To further investigate the potential role of STAT1 in PCa, we firstly performed qRT-PCR and western blot to compare the expression of STAT1 between normal prostate epithelial cells and different PCa cell lines. It was shown in Figs. 1A and S2C that the expression of STAT1 in PCa cell lines including DU145, PC-3, 22RV1, C4-2, C4-2B, was higher than that in normal prostate epithelial cells RWPE-1. Notably, DU145 and PC-3 presented a relatively higher expression of STAT1 than other PCa cell lines, thus were used to perform further experiments. The result was verified by an immunohistochemistry assay that compared to BPH tissue, PCa tissue showed a higher expression of STAT1 (Fig. 1B). Additionally, we observed by western blot that in 20 matched pairs of prostate cancer and adjacent normal tissues, the expression and the activity of STAT1 were higher in prostate cancer tissues (Fig. 1C, D). Furtherly, we performed survival analysis for PCa patients from our institution based on expression of STAT1 in PCa tissues assessed by IHC. Kaplan-Meier survival analysis revealed that patients with higher expression of STAT1 presented a worse survival outcome (Fig. 1E). These observations identified STAT1 might act as an oncogene in PCa.

A Relative mRNA expression of STAT1 in prostate cancer cell lines detected by qRT-PCR. B Expression of STAT1 in prostate cancer tissues (n = 80) and benign prostatic hyperplasia tissues (n = 80) detected by immunohistochemistry. C, D Expression of p-STAT1 and STAT1 in 20 pairs of matched prostate cancer tissue and adjacent normal tissues detected by western blot. E Survival analysis for PCa patients with high STAT1 expression and low STAT1 expression. F, G qRT-PCR and H, I Western blot used to detect knockdown and overexpression efficiency of STAT1. J, K The glucose uptake of DU145 cells and PC-3 cells after STAT1 knockdown or STAT1 overexpression. L, M The lactate production of DU145 cells and PC-3 cells after STAT1 knockdown or STAT1 overexpression. N, O ECAR and P, Q OCR of DU145 cells and PC-3 cells after STAT1 knockdown or STAT1 overexpression. Data were presented as mean ± SD of at least three independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 by paired two-tailed Student’s t test or unpaired two-tailed Student’s t test or one-way ANOVA test.
STAT1 promoted the transcription of glycolytic key enzymes and enhanced aerobic glycolysis in prostate cancer cells
STAT1 was well known as a significant transcription factor extensively involved in aerobic glycolysis of pancreatic ductal adenocarcinoma and multiple myeloma [33, 34]. However, whether STAT1 regulated aerobic glycolysis of prostate cancer is still unclear. To investigate the role of STAT1 in the aerobic glycolysis of PCa cells, the glucose uptake, lactate production and energy metabolism of DU145 and PC-3 cells were detected after knockdown and overexpression of STAT1. As shown in Fig. 1F–I, the efficiency of STAT1 knockdown and overexpression in DU145 and PC-3 cells was detected by qRT-PCR and western blot. Glucose uptake assay discovered that glucose uptake was significantly reduced after knockdown of STAT1, while glucose uptake was significantly increased after overexpression of STAT1 (Fig. 1J, K). Likewise, lactate production assay confirmed that lactate level in cell supernatant was significantly decreased after knockdown of STAT1, while increased after overexpression of STAT1 (Fig. 1L, M). Subsequently, in order to visually observe the alterations of glycolysis and mitochondrial respiration of PCa cells, seahorse XF analyzer was used to measure ECAR and OCR of DU145 and PC-3 cells. It was shown that STAT1-knockdown cells exhibited a lower glycolytic capacity and STAT1-overexpression cells had a higher glycolytic capacity (Fig. 1N, O). We also found that the respiratory capacity was higher after knockdown of STAT1, while it was lower after overexpression of STAT1 (Fig. 1P, Q). These results preliminarily indicated that STAT1 enhanced the aerobic glycolysis of PCa cells.
To determine whether STAT1 affected glycolysis through regulating expression of glycolytic key enzymes, we performed qRT-PCR and western blot to detect the expression level of HK2, PFKM, PKM2 following successful knockdown of STAT1 in DU145 and PC-3 cells, and then it was found that HK2, PFKM and PKM2 all showed a lower expression level after knockdown of STAT1 (Fig. 2A–D). Moreover, we constructed stable DU145 and PC-3 cell lines of STAT1 overexpression to further verify the effect of STAT1 on the expression of glycolytic key enzymes. As we anticipated, HK2, PFKM and PKM2 were more highly expressed in stable DU145 and PC-3 cells of STAT1 overexpression compared to vector control (Fig. 2E–H). These results demonstrated that not only did STAT1 affect mRNA expression levels of HK2, PFKM and PKM2, but it also had an impact on their protein expression levels, which suggested that STAT1 probably influenced aerobic glycolysis via transcriptional regulation of HK2, PFKM and PKM2.

A, B Following STAT1 knockdown, relative mRNA expression of HK2, PFKM, PKM2 in DU145 cells and PC-3 cells detected using qRT-PCR. C, D Following STAT1 knockdown, relative protein expression of HK2, PFKM, PKM2 in DU145 cells and PC-3 cells detected using western blot. E, F Following STAT1 overexpression, relative mRNA expression of HK2, PFKM, PKM2 in DU145 cells and PC-3 cells detected using qRT-PCR. G, H Following STAT1 overexpression, relative protein expression of HK2, PFKM, PKM2 in DU145 cells and PC-3 cells detected using western blot. I–K Wild type and mutate type of HK2, PFKM, PKM2 promoters binding site designed for dual-luciferase reporter assay. L–N Relative luciferase activities in wild-type and mutate type promoters of HK2, PFKM, PKM2 in control and STAT1 overexpression 293T cells detected by dual-luciferase reporter assay. O, P The combination of STAT1 with HK2, PFKM, PKM2 in DU145 and PC-3 cells shown using CUT & Tag-qPCR assay with anti-IgG or anti-p-STAT1 antibody. Q–S Correlation analysis between expression of STAT1 and expression of HK2, PFKM and PKM2 of PCa tissues. Data were presented as mean ± SD of at least three independent experiments. **P < 0.01, ***P < 0.001, ****P < 0.0001 by unpaired two-tailed Student’s t test or one-way ANOVA test.
STAT1 formed homodimeric or heterodimeric dimers and then translocated into the cell nucleus to exert the function of a transcriptional activator [35]. By use of JASPAR database (https://jaspar.elixir.no/), it was predicted that there were binding sites for the STAT1 transcriptional factor in the promoters of HK2, PFKM and PKM2. To verify whether STAT1 transcriptional factor directly bound to promoters of HK2, PFKM and PKM2, dual-luciferase reporter assay and CUT & Tag-qPCR assay were carried out. As revealed by dual-luciferase reporter assay, when compared to vector control, STAT1 overexpression markedly enhanced the luciferase activity of wild-type promoters of HK2, PFKM and PKM2, but not mutated type promoters of HK2, PFKM and PKM2 in 293T cells (Fig. 2I–N). For the CUT & Tag-qPCR assay, p-STAT1 antibody was used to bind the p-STAT1 transcriptional factor and then DNA fragments binding to p-STAT1 were amplified by PCR. CUT & Tag-qPCR assay confirmed that promoters of HK2, PFKM and PKM2 could be significantly enriched by p-STAT1, but not IgG control (Fig. 2O, P). Collectively, the above results suggested that the STAT1 transcriptional factor could bind to promoters of HK2, PFKM and PKM2 and subsequently activated expression of HK2, PFKM and PKM2.
To further enhance the association between expression of STAT1 and expression of glycolytic key enzymes (HK2, PFKM and PKM2) in PCa, we performed correlation analysis according to the results of immunohistochemical staining for STAT1, HK2, PFKM and PKM2 in 80 samples of PCa.
The result showed that expression of STAT1 was positively correlated with expression of HK2, PFKM and PKM2 (Fig. 2Q–S), which further elucidated a strong positive correlation between expression of STAT1 and aerobic glycolysis in PCa.
STAT1 promoted progression and resistance to radiotherapy of prostate cancer via activating the glycolysis pathway
Given the pronounced enhancing effects of STAT1 on glycolysis in prostate cancer cells, we hypothesized that STAT1 might aggravate the malignant phenotype of prostate cancer through the glycolysis pathway. Experiment groups were set as follows: vector group, OE-STAT1 group, OE-STAT1 + DMSO group, OE-STAT1 + 2-DG group. Cell counting assay demonstrated that STAT1 overexpression prominently elevated the ability to proliferate of PCa cells, which were rescued by further adding 2-DG (Fig.S3A, B). The promoting effect of STAT1 on the proliferation of PCa cells was reconfirmed using a colony formation assay, and this effect could be inhibited by 2-DG (Fig. S3C). Coherently with this result, the Transwell assay revealed that capacities of migration and invasion were prominently elevated by the overexpression of STAT1, however, this impact was inhibited by addition of 2-DG (Fig. S3D). It was identified again by wound-healing assay that accompanied by treatment of 2-DG, the promoting function of STAT1 on migration of PCa cells were denervated obviously (Fig. S3E). Subsequently, we established subcutaneous xenograft tumor model to examine whether the same effect could be exerted in vivo. Compared with vector group, a dramatical elevation of tumor growth rate, tumor volume and tumor weight were all observed in OE-STAT1 group, while 2-DG also rescued promoting function of STAT1 on prostate cancer growth in vivo (Fig. 3A–C). Furthermore, we also detected the antitumor effect of STAT1-knockdown and treatment with 2-DG in PCa cells. CCK-8 assay and colony formation assay demonstrated that both STAT1-knockdown and treatment with 2-DG markedly inhibited the proliferative ability of PCa cells (Fig. S4A–C). Migrative and invasive capacities of PCa cells were also prominently attenuated by STAT1-knockdown as well as by treatment with 2-DG, as revealed by Transwell assays (Fig. S4D, E). The above results provided essential evidence for pro-oncogenic role of STAT1 via glycolysis in PCa both in vitro and in vivo.

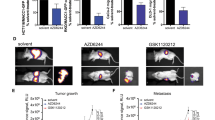
A, B The tumor growth curves and C weight of BALB/c mice injected with DU145 and PC-3 cells, accompanied by drugs administration in vivo xenograft tumorigenicity model. D, E Comet assays were used to detect DNA damage repair of DU145 and PC-3 cells at the indicated time following irradiation. F, G Immunofluorescence was used to evaluate γ-H2AX foci formation of DU145 cells at 8 h and 24 h after irradiation. H–J Representative images of PDOs underwent irradiation, which was employed to perform CTG assay. Data were presented as mean ± SD of at least three independent experiments. *P < 0.05 by repeated-measures ANOVA test in (B). *P < 0.05, **P < 0.01, ****P < 0.0001 by one-way ANOVA test in (C, E, G, I, J).
Furtherly, we investigated whether STAT1 affected resistance to radiotherapy of PCa and whether glycolysis was involved in enhancement of resistance to radiotherapy. Comet assays and immunofluorescence assays were performed to evaluate DNA damage. As shown in Fig. 3D, E, at 8 h after irradiation, tail moments of PCa cells were significantly shorter following STAT1 overexpression and this effect was disrupted by further treatment of 2-DG. Furthermore, this phenomenon was reconfirmed by examining γ-H2AX foci levels after 8 h and 24 h irradiation (Figs. 3F, G, and S4F, G). In PDO models, we initially found that the PDOs in OE-STAT1 group exhibited significantly larger size and more preferable survival compared to the PDOs in vector control group, under irradiation. Strikingly, further addition of 2-DG could markedly reverse effect caused by STAT1 overexpression on growth of PDOs (Fig. 3H–J). These results suggested STAT1 accelerated DNA damage repair through glycolysis pathway, which demonstrated STAT1 possessed potential to induce radiotherapy resistance via glycolysis.
Lactate derived from prostate cancer cells promoted M2 macrophages polarization
Previous studies have found that metabolite was crucial interactive signal between tumor cells and macrophages in TME [36]. We firstly explore whether glycolytic metabolite derived from PCa cells played a regulatory role on macrophages polarization. THP-1 cells were co-cultured with 200 nM phorbol 12-myristate 13-acetate (PMA) for 48 h and then M0 macrophages were identified by morphology observation and CD68 expression detection. It was observed that THP-1 cells changed from a suspended state to an attached state and extended filopodia (Fig. S4H). Besides, protein expression of M0 macrophage marker CD68 of THP-1 cells was also increased after induction by PMA (Fig. S4I). These results universally indicated M0 macrophages had been successfully induced. We treated DU145 and PC-3 cells with glycolysis inhibitor 2-DG, conditioned medium collected after 24 h, were used to culture M0 macrophages for 48 h. The result was showed by qRT-PCR that compared with control group, 2-DG group had a higher M1 macrophage markers (CD80, iNOS) expression and a lower M2 macrophage markers (IL-10, VEGF) expression (Fig. 4A, B). Western blot was also performed to detect protein expression of M1 macrophage markers and M2 macrophage markers. The result of western blot was in agreement with qRT-PCR (Fig. 4C–F).

A, B qRT-PCR and C–F western blot detecting mRNA and protein expression of M1 macrophage markers (CD80, iNOS) and M2 macrophage markers (IL-10, VEGF) in M0 macrophages co-cultured with conditioned medium from DU145 and PC-3 cells treated by complete medium or glycolysis inhibitor 2-DG for 48 h, respectively. G, H qRT-PCR and I–L western blot detecting mRNA and protein expression of M1 macrophage markers (CD80, iNOS) and M2 macrophage markers (IL-10, VEGF) in M0 macrophages co-cultured with conditioned medium from DU145 and PC-3 cells treated by complete medium or LDHA inhibitor GSK2837808A or LDHA inhibitor GSK2837808A adding with lactate for 48 h, respectively. M, N qRT-PCR and O–R western blot detecting mRNA and protein expression of M1 macrophage markers (CD80, iNOS) and M2 macrophage markers (IL-10, VEGF) in M0 macrophages co-cultured with conditioned medium from control or stable STAT1-overexpressed DU145 and PC-3 cells by treatment with glycolysis inhibitor 2-DG or 2-DG adding with lactate for 48 h, respectively. Data were presented as mean ± SD of three independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 by unpaired two-tailed Student’s t test or one-way ANOVA test.
Subsequently, we determined which metabolite from the glycolysis of prostate cancer cells contributed to M2 macrophage polarization. Considering that the exacerbation of glycolysis was accountable for the accumulation of lactate, we treated DU145 and PC-3 cells with glycolysis inhibitor 2-DG and glycolysis activator fructose-2,6-biphosphatase 3 (PFKFB3) at a concentration of 0.2 mM and 100 μΜ and the lactate levels of the cell supernatant were detected after 24h. We found that compared with the control group, lactate in cell supernatant was obviously decreased after treatment with 2-DG, while it was obviously increased after treatment with PFKFB3 (Fig.S4J). Hence, we speculated that it was lactate in the metabolite from glycolysis that drove M2 macrophages polarization. We treated DU145 and PC-3 cells with complete medium (Control) and lactate dehydrogenase (LDHA) inhibitor GSK2837808A (20 μM, LDHA inhibitor), respectively, and the conditioned medium collected after 24 h was co-cultured with M0 macrophages for 48 h. For rescue experiments, exogenous lactate was directly added to conditioned medium collected after treatment with the LDHA inhibitor to induce M0 macrophages. It was suggested by qRT-PCR that compared to the control group, M1 macrophage markers were increased and M2 macrophage markers were decreased in the LDHA inhibitor group remarkably, but these effects were abolished by further addition of 10 mM lactate (Fig. 4G, H). The protein expression trend detected by western blot was consistent with the mRNA expression trend (Fig. 4I–L). These results indicated that lactate derived from PCa cells actually promoted M2 macrophage polarization.
Next, we verified whether this effect was regulated by STAT1. We collected conditioned medium from stable STAT1 overexpression (OE-STAT1) and vector control (Vector) DU145 and PC-3 cells, respectively. In addition, glycolysis inhibitor 2-DG (OE-STAT1+2-DG) and an equal volume of DMSO (OE-STAT1+DMSO) were used to treat STAT1 overexpression DU145 and PC-3 cells for 24h, and then the conditioned medium was collected, respectively. The above conditioned medium from different groups was used to co-culture with M0 macrophages for 48 h. Moreover, lactate was added into conditioned medium from STAT1 overexpression DU145 and PC-3 cells treated by 2-DG, and collectively treated M0 macrophages for 48 h. It was revealed by qRT-PCR and western blot that, both in mRNA expression level and protein expression level, the OE-STAT1 group exhibited a lower expression of M1 macrophage markers and a higher expression of M2 macrophage markers than the vector group. Compared to the OE-STAT1 group, expression of M1 macrophage markers was increased and expression of M2 macrophage markers was decreased in the OE-STAT1 + 2-DG group; however, this effect was significantly reversed by supplementation of 10 mM lactate (Fig. 4M–R). As demonstrated above, lactate from glycolysis, regulated by STAT1 of PCa cells, promoted M2 macrophage polarization.
Moreover, to explore the effect of metabolites under irradiation conditions on macrophage polarization, we obtained conditioned medium from DU145 and RM-1 treated with 2-DG and irradiation. The conditioned media were cultured with the M0 macrophages and BMDMs, respectively. Both qRT-PCR and western blot analyses demonstrated that under radiation conditions, the 2-DG group exhibited a significant increase in M1 macrophage markers expression and a significant decrease in M2 macrophage markers expression compared to the control group (Fig. S5A–F). Subsequently, to detect the effect of lactate derived from irradiated PCa cells on macrophages polarization, we collected conditioned medium from irradiated DU145 and RM-1 cells treated with complete medium or LDHA inhibitor. The conditioned media were then incubated with M0 macrophages and BMDMs, respectively. It was found that M1 macrophage markers expressions were significantly elevated and M2 macrophage markers expressions were significantly decreased after treatment with LDHA inhibitor, however, further addition of lactate significantly rescued this effect (Fig. S5G-L). Additionally, we also detected whether this effect was regulated by STAT1 under irradiation condition. We irradiated DU145 and RM-1 cells with stable STAT1 overexpression, along with their corresponding controls. We further treated STAT1 overexpression cells with 2-DG and an equal volume of DMSO. Conditioned media were then collected from all groups and used to culture M0 macrophages and BMDMs. We discovered that compared to the vector control group, the OE-STAT1 group exhibited lower levels of M1 macrophage markers and higher levels of M2 macrophage markers. Compared to the OE-STAT1 group, the further addition of 2-DG led to a decrease in M2 macrophage markers and an increase in M1 macrophage markers, however, addition of lactate could reverse this effect (Fig. S5M–R). Collectively, under radiation conditions, lactate from glycolysis regulated by STAT1 of PCa cells also drove M2 macrophages polarization.
Monocarboxylate transporter 1 (MCT1)-dependent lactate uptake in macrophages promoted M2 macrophage polarization via activation of NFκB1 pathway
NFκB transcriptional factor family, was regarded as key regulatory factor for immune response and inflammatory response in TME [37]. It was reported previously that a lack of NFκB1 could induce M1 macrophage polarization to increase the expression of pro-inflammatory factors [38]. Therefore, we speculated that M2 macrophage polarization promoted by lactate was possibly dependent on activation of the NFκB1 pathway. MCT1, as an important transporter of lactate, was widely distributed in macrophages, lymphocytes, neutrophils and endothelial cells [39, 40]. Therefore, we first assessed the effect of lactate on NFκB1 pathway in macrophages. It was shown by western blot that expression of NFκB1 p50 and NFκB1 p65 in macrophage nucleus was prominently elevated, following addition of lactate, which could be rescued by further addition of MCT1 inhibitor AZD3965 (Fig. 5A, B). The result of immunofluorescence was exhibited as Fig. 5C, D, that stimulation with 10 mM lactate significantly enhanced nuclear translocation of NFκB1 p50 and NFκB1 p65, but this enhancement was diminished when the macrophages were co-stimulated with lactate and 10 μM MCT1 inhibitor AZD3965. Besides, the phosphorylation level of IKBα was significantly elevated after treatment with lactate, but this effect was attenuated after co-treatment with lactate and MCT1 inhibitor AZD3965 (Fig. 5E). These results together proved that lactate activated NFκB1 pathway in macrophages and this activation strictly depended on MCT1 transportation.

A, B Western blot detecting nucleus and cytoplasmic distribution of NFκB1 p65 and NFκB1 p50 in M0 macrophages treated with complete medium (Control) or lactate (LA) or lactate added with MCT1 inhibitor AZD3965 (LA + AZD3965). C, D The immunofluorescence staining detecting nucleus translocation of NFκB1 p65 (green) and NFκB1 p50 (green) in M0 macrophages treated with complete medium (Control) or lactate (LA) or lactate added with MCT1 inhibitor AZD3965 (LA + AZD3965). E Western blot detecting activation of IκBα in M0 macrophages treated with complete medium (Control) or lactate (LA) or lactate adding with MCT1 inhibitor AZD3965 (LA + AZD3965). F qRT-PCR and G Western blot detecting mRNA and protein expression of M1 macrophage markers (CD80, iNOS) and M2 macrophage markers (IL-10, VEGF) in M0 macrophages treated with complete medium (Control) or lactate (LA) or lactate adding with MCT1 inhibitor (LA + AZD3965) or lactate adding with NFκB1 pathway inhibitor (LA + JSH-23), respectively. Data were presented as mean ± SD of three independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 by one-way ANOVA test.
In order to define whether MCT1-dependent lactate uptake affected M2 macrophages polarization through activation of the NFκB1 pathway, we grouped as described below: M0 macrophages treated with complete medium (Control), M0 macrophages treated with lactate (LA), M0 macrophages co-treated with lactate and MCT1 inhibitor (LA + AZD3965), M0 macrophages co-treated with lactate and 30 μM NFκB1 inhibitor JSH-23 (LA + JSH-23). The results detected by qRT-PCR and western blot illustrated that compared to the control group, M1 macrophage markers were remarkably down-regulated and M2 macrophage markers were remarkably up-regulated in the LA group. However, following co-treatment of lactate with MCT1 inhibitor or NFκB1 inhibitor, the M1 macrophage markers were remarkably up-regulated, and M2 macrophage markers were remarkably down-regulated (Fig. 5F-G). These results clarified that MCT1-dependent lactate uptake in macrophages promoted M2 macrophage polarization via activation of NFκB1 pathway.
MCT1-dependent lactate uptake in macrophages facilitated MCP-1 transcription via activation of NFκB1 pathway
It has been found that activation of NFκB1 pathway by lactate in endothelial cells and cancer-associated fibroblasts could drive gene transcription of cytokines, including IL-8 and CXCL9 [41, 42]. It was conjectured that activation of NFκB1 pathway by lactate possibly modulated downstream cytokine gene transcription in macrophages. To search for possible key cytokines regulated by NFκB1, we performed a protein microarray to screen differentially expressed secreted proteins from the cell supernatant of NFκB1 knockdown and negative control macrophages. It was found that secretion of MCP-1 was decreased after NFκB1 knockdown. The screening and clustering diagrams of differentially secreted proteins were displayed in Fig. 6A–C. Furthermore, gene ontology (GO) analysis and Kyoto Encyclopedia of genes and genomes (KEGG) analysis were performed for the differentially secreted proteins. GO analysis revealed that differential secreted proteins were mainly enriched in regulation of leukocyte migration and mononuclear cell migration in biological process; vesicle lumen, vacuolar lumen and membrane in cellular component; tumor necrosis factor (TNF) signaling pathway, CXCR chemokine receptor binding and CCR chemokine receptor binding in molecular function (Fig. S6A–C). The result of KEGG analysis showed that differential secreted proteins were primarily enriched TNF signaling pathway, vital protein interaction with cytokine and cytokine receptor, NFκB signaling pathway and chemokine signaling pathway (Fig. S6D).

A Volcano plot of differentially expressed proteins between siNFκB1 and siNC groups screened by protein microarray. B Scatterplot of differentially expressed proteins between siNFκB1 and siNC groups screened by protein microarray. C Heatmap of differentially expressed proteins between siNFκB1 and siNC groups screened by protein microarray. D qRT-PCR and E western blot showing the shRNA lentiviral knockdown efficiency of NFκB1 and the effect of NFκB1 knockdown on MCP-1 mRNA and protein expression in macrophages, respectively. F ELISA showing the effect of NFκB1 knockdown on secretive level of MCP-1 in macrophages. G Wild type and mutate type of MCP-1 promoter binding site designed for dual-luciferase reporter assay. H Relative luciferase activities in wild-type and mutate type promoters of MCP-1 in control and NFκB1 overexpression 293 T cells detected by dual-luciferase reporter assay. I, J The combination of NFκB1 p65 or NFκB1 p50 with MCP-1 in macrophages shown using CUT & Tag-qPCR assay with anti-IgG or anti-NFκB1 p65 antibody or anti-NFκB1 p50 antibody. K qRT-PCR and L western blot detecting mRNA and protein expression of MCP-1 in macrophages treated with complete medium (Control) or lactate (LA) or lactate adding with MCT1 inhibitor (LA + AZD3965) or lactate adding with NFκB1 pathway inhibitor (LA + JSH-23), respectively. M ELISA detecting secretive level of MCP-1 in macrophages treated with complete medium (Control) or lactate (LA) or lactate adding with MCT1 inhibitor (LA + AZD3965) or lactate adding with NFκB1 pathway inhibitor (LA + JSH-23). Data were presented as mean ± SD of three independent experiments. **P < 0.01, ****P < 0.0001 by unpaired two-tailed Student’s t test or one-way ANOVA test.
For validation of the results from the protein microarray, we performed a series of experiments following the knockdown of NFκB1 in macrophages. qRT-PCR and western blot found that the mRNA and protein expression of MCP-1 were decreased significantly along with successful knockdown of NFκB1 in macrophages (Fig. 6D, E). Analysis of ELISA exhibited a lower secretion level of MCP-1 after knockdown of NFκB1 in macrophages (Fig. 6F). Aimed to confirm transcriptionally regulatory effect of NFκB1 on MCP-1, dual-luciferase reporter assay and CUT & Tag-qPCR assay were implemented successively. Using dual-luciferase reporter assay in 293 T cells, it was discovered that compared to vector group, overexpression NFκB1 strengthened luciferase activity of MCP-1 wild type promoter, while luciferase activity of MCP-1 mutated type promoter was still restricted after overexpression NFκB1 (Fig. 6G, H). For CUT & Tag-qPCR assay, we use NFκB1-p50 antibody and NFκB1-p65 antibody to bind corresponding proteins and amplified enriched DNA fragment using PCR. Interestingly, we discovered that NFκB1-p65 significantly enriched the MCP-1 promoter compared to IgG, however, there was no significant difference between anti-NFκB1-p50 group and IgG group (Fig. 6I, J). These results supported the conclusion that NFκB1-p65, instead of NFκB1-p50, identified and bound to MCP-1 promoter to activate MCP-1 transcription in macrophages.
Subsequently, to clarify the effect of MCT1-dependent lactate uptake on secretion of MCP-1 in macrophages and whether this effect occurred via activation of NFκB1 pathway, we established four treatment groups as following: M0 macrophages treated with complete medium (Control), M0 macrophages treated with lactate (LA), M0 macrophages co-treated with lactate and MCT1 inhibitor (LA + AZD3965), M0 macrophages co-treated with lactate and NFκB1 inhibitor JSH-23 (LA + JSH-23). As revealed by qRT-PCR and western blot, compared to the control group, the expression level of MCP-1 in macrophages was significantly increased by treatment with lactate. Nevertheless, combined treatment of lactate and MCT1 inhibitor or lactate and NFκB1 inhibitor, the inductive impact of lactate on MCP-1 was diminished obviously (Fig. 6K, L). Likely, the secretion of MCP-1 detected by ELISA presented the trend agreeing with that revealed by qRT-PCR and western blot (Fig. 6M). Based on the results described above, current evidence illustrated that MCT1-dependent lactate uptake in macrophages facilitated MCP-1 transcription via activation of NFκB1 pathway.
MCP-1 promoted glycolysis, progression and resistance to radiotherapy of prostate cancer via activation of JAK/STAT1 pathway
JAK/STAT1 was proven as a downstream signaling pathway of multiple chemokines [43]. MCP-1 was combined with chemokine C-C-motif receptor 2 (CCR2) to activate a downstream signaling pathway [44]. We evaluated the expression of MCP-1 and CCR2 in normal prostate epithelial cells and different prostate cancer cell lines. We found that the expression of MCP-1 and CCR2 in prostate cancer cell lines was higher than that in RWPE-1. In addition, the aggressive cell lines PC-3, DU145, C4-2B, and C4-2 presented higher expression levels of MCP-1 and CCR2, compared with cell lines of 22RV1 and LNCaP (Fig. S6E–H). It was also revealed that compared to adjacent normal tissues, PCa tissues exhibited a higher activation of JAK signaling, as indicated by a marked increase in phosphorylation levels (Fig. S6I). These results indicated that MCP-1/CCR2 and JAK/STAT1 signaling pathway were more active in PCa.
To explore whether MCP-1 signal from macrophages activated JAK/STAT1 pathway of prostate cancer cells through identifying and binding to CCR2, complete medium (Control) and 200 ng/ml recombinant MCP-1 (MCP-1) were used to culture DU145 and PC-3 cells for 24 h. Rescue experiments in DU145 and PC-3 cells were conducted by co-treatment with recombinant MCP-1 and 30 μΜ CCR2 antagonist INCB8761 (MCP-1 + INCB8761), or recombinant MCP-1 and 5 μΜ JAK inhibitor ruxolitinib. By using western blot to evaluate the activation of JAK/STAT1 pathway, we found that DU145 and PC-3 cells exhibited an increased phosphorylation JAK and phosphorylation STAT1 with treatment of MCP-1, hinting at JAK/STAT1 pathway was activated by MCP-1. However, this activation of JAK/STAT1 caused by MCP-1 was suppressed both in DU145 and in PC-3 cells furtherly treated with CCR2 antagonist or JAK inhibitor (Fig. 7A), confirming that JAK/STAT1 was the downstream signal of MCP-1/CCR2 in PCa cells.

A Western blot detecting the phosphorylation level of JAK and STAT1 in DU145 and PC-3 cells treated with complete medium (Control) or recombinant MCP-1 (MCP-1) or recombinant MCP-1 adding with CCR2 antagonist (MCP-1 + INCB8761) or recombinant MCP-1 adding with JAK inhibitor (MCP-1+Ruxolitinib). B The glucose uptake and C the lactate production of DU145 cells and PC-3 cells treated with complete medium (Control) or recombinant MCP-1 (MCP-1) or recombinant MCP-1 adding with CCR2 antagonist (MCP-1 + INCB8761) or recombinant MCP-1 adding with JAK inhibitor (MCP-1+Ruxolitinib). D, E ECAR and F, G OCR of DU145 cells and PC-3 cells treated with complete medium (Control) or recombinant MCP-1 (MCP-1) or recombinant MCP-1 adding with CCR2 antagonist (MCP-1 + INCB8761) or recombinant MCP-1 adding with JAK inhibitor (MCP-1+Ruxolitinib). Data were presented as mean ± SD of at least three independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 by one-way ANOVA test.
Considering the proven promoting function of STAT1 on glycolysis, we attempted to detect glucose uptake and lactate production of DU145 and PC-3 cells treated with the four different interventions as mentioned above. It was discovered that capacity of glucose uptake and lactate production was increased by MCP-1, and addition of CCR2 antagonist or JAK inhibitor was responsible for inhibition of this enhanced glycolysis caused by MCP-1(Fig. 7B, C). It was also verified by Seahorse XF for detecting cellular energy metabolism that glycolytic activity indicated by ECAR was significantly elevated after addition of MCP-1, however, CCR2 antagonist or JAK inhibitor reversed the enhanced ECAR caused by MCP-1 (Fig. 7D, E). As shown in Fig. 7F, G, we found that the variation tendency of OCR was opposite to ECAR in DU145 and PC-3 cells. These results indicated that activation of JAK/STAT1 pathway was necessary for MCP1-induced glycolysis in PCa cells.
A series of functional experiments was performed to assess the functions of MCP-1 on PCa cells, and these functions were exerted with or without involvement of JAK/STAT1 pathway. Proliferation of DU145 and PC-3 cells was obviously improved after culture with recombinant MCP-1, but proliferation was attenuated by the addition of CCR2 antagonist or JAK inhibitor, as uncovered by cell counting assay and colony formation assay (Fig. S6J–L). It was found by Transwell assay that migrative and invasive abilities of DU145 and PC-3 cells treated with recombinant MCP-1 were aggravated, while this aggravation was blocked by CCR2 antagonist or JAK inhibitor (Fig. S7A, B). Wound-healing assay revealed results consistent with Transwell assay that migrative ability elevated by MCP-1 was attenuated remarkably when DU145 and PC-3 cells were treated with CCR2 antagonist or JAK inhibitor (Fig. S7C). We also determined that MCP-1 favored growth of tumor in subcutaneous xenograft tumor in nude mice. With the simulation of MCP-1, the tumor showed a larger size, a faster growth and a heavier weight. However, this effect disappeared upon the addition of either a CCR2 antagonist or a JAK inhibitor (Fig. S7D–I).
Furtherly, we explored whether MCP-1 aggravated resistance to radiotherapy of PCa using comet assays and immunofluorescence assays. Treatment with MCP-1 decreased the length of tail moments at 8 h after irradiation in PCa cells. Interestingly, these reduced tail moments were rescued by further treatment with CCR2 antagonist or JAK inhibitor following irradiation at 8 h (Fig. 8A, B). In addition, the result consistent with comet assays was revealed by immunofluorescence assays that formation γ-H2AX foci significantly decreased after addition of MCP-1, however, γ-H2AX foci levels were elevated by further supplementation with CCR2 antagonist or JAK inhibitor at 8 h and 24 h after irradiation (Figs. 8C, D and S7J, K).

A, B DNA damage repairs of DU145 and PC-3 cells at the indicated time following irradiation were detected using comet assays. C, D γ-H2AX foci formations of DU145 cells at 8 h and 24 h after irradiation were measured by immunofluorescence. E, F The tumor growth curves and G weight of c57BL6/J mice implanted with RM-1 cells, accompanied by further intratumoral injection with different drugs in vivo xenograft tumorigenicity model. H, I The tumor growth curves and J weight of c57BL6/J mice implanted with RM-1 cells, accompanied by further radiation and intratumoral injection with different drugs in vivo xenograft tumorigenicity model. K A working model illustrating a positive feedback loop mechanism between prostate cancer cells and macrophages. Data were presented as mean ± SD of at least three independent experiments. *P < 0.05 by repeated-measures ANOVA test in (F, I). *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 by one-way ANOVA test in (B, D, G, J).
In light of the above results, we further explored whether JAK inhibitors and CCR2 antagonists could improve the sensitivity to radiotherapy of PCa in vivo. In the mouse RM-1 subcutaneous tumor model, we initially found that both the CCR2 antagonist and the JAK inhibitor significantly suppressed tumor growth, and the combination of them even exhibited a more effective inhibition for tumor growth (Fig.8E–G). Subsequently, the effects of CCR2 antagonist and JAK inhibitor on tumor growth under radiation exposure were investigated. It was found that under radiation, CCR2 antagonist or JAK inhibitor could markedly decreased tumor growth, compared the radiation alone group. Of greater concern was that combination of CCR2 antagonist with JAK inhibitor showed the most significant enhancement for sensitivity to radiation of PCa (Fig. 8H–J). Furthermore, we found that compared to the radiation alone group, the activation of STAT1 and the expression of glycolytic key enzymes (HK2, PFKM, PKM2) were significantly decreased by treatment with CCR2 antagonist or JAK inhibitor, and combination of them produced a maximal inhibitory effect (Fig. S8A–C). Compared to the radiation alone group, individual administration of either CCR2 antagonist or JAK inhibitor presented a lower infiltration of M2 macrophages, furtherly, combination of them exhibited a synergistic effect, achieving a more profound decrease (Fig. S8D, E). Then, we also explored the impact of STAT1 on tumor growth under radiation exposure. We discovered that compared with the radiation alone group, STAT1 overexpression combined with radiation promoted the tumor growth and exhibited a higher STAT1 activation and a higher expression of HK2, PFKM and PKM2, but the addition of CCR2 antagonist markedly suppressed the facilitation effect of STAT1 overexpression. Additionally, compared with the radiation alone group, CCR2 antagonist combined with radiation significantly inhibited tumor growth, and showed a lower STAT1 activation and a lower expression of HK2, PFKM and PKM2; however, overexpression of STAT1 could reverse these effects (Fig. S8F–K).
Consequently, these data elucidated that MCP-1 secreted from macrophages promoted glycolysis, progression and resistance to radiation of PCa via activation of JAK/STAT1 pathway and preliminarily indicated that JAK inhibitors and CCR2 antagonists may have the potential to improve the sensitivity to radiotherapy. The Fig. 8K exhibited a STAT1/ lactate /NFκB1/MCP-1 positive feedback loop between PCa cells and macrophages.
Discussion
TIME consisted of cancer cells and infiltrating immune cells. In prostate cancer TIME, immune cells functioned as an anti-tumor immune response relying on CD8+T cells. It was equally important that the accumulation of immunosuppressive cells covering TAMs, suppressive myeloid cells and regulatory T cells facilitated immune escape for PCa cells [45]. TAMs were an essential component of prostate cancer TIME. Cytokines and metabolic production were considered as a significant transmissive molecular regulating interaction between PCa cells and macrophages. Acted as the glycolic end product, lactate was conducive for cancer progression through establishing a suppressive TIME [16]. Previous reports drew a conclusion that lactate exacerbated polarization toward M2 macrophages, resulting in progression of breast cancer and cervical cancer [17, 18]. In this study, the result was concordant with findings for other solid tumors, that lactate secreted from PCa cells drove alteration of macrophage subtype from M0 to M2. However, the regulatory molecular network on the secretion of lactate by PCa cells is still unrevealed.
Previous researches have reported the potential molecular mechanism of lactate-induced M2 macrophages in cancers. Jiang et al. revealed that ectopic Zeb1 increased the transcriptional expression of glycolytic key enzymes to produce lactate, which promoted M2 macrophage polarization via the PKA/CREB signaling pathway, thus driving the breast cancer proliferation, migration, and chemoresistance [32]. Cai et al. found that in hepatocellular carcinoma, SRSF10 stabilized MYB RNA, which enhanced the transcription of glycolytic enzymes GLUT1, HK1, and LDHA, leading to lactate production, which drive M2 macrophage polarization by inducing histone H3K18 lactylation to activate pro-tumoral genes [46]. Zhang et al. discovered that lactate from tumor cells promoted M2 macrophage polarization via mTORC2-Akt/ERK signaling to trigger CCL17 secretion, which subsequently activated the CCR4/mTORC1 axis in tumor cells and promoted pituitary adenoma invasion [47]. Similar to study, by Cai et al. and Jiang et al., our study found that STAT1 enhanced the transcription of glycolytic key enzymes, inducing production of lactate in PCa cells, which also identified aerobic glycolysis as the upstream driver of lactate production in tumor cells. In addition, our study revealed bidirectional communication between PCa cells and macrophages mediated by lactate and MCP-1 as signaling molecules. This bidirectional interaction, rather than unidirectional regulation, formed a positive feedback loop that promoted tumor progression, highlighting the dynamic nature of metabolic-immune crosstalk in the TIME, which was consistent with the findings of Zhang et al. Our study identified a unique positive feedback loop in which STAT1 initiated glycolytic gene (HK2, PFKM, PKM2) upregulation, leading to lactate production. The lactate subsequently activated the NFκB pathway in macrophages, inducing MCP-1 secretion that activated JAK/STAT signaling in PCa cells, thereby completing the loop through STAT1 reactivation. This STAT1/lactate/NFκB1/MCP-1 positive feedback mechanism was identified as a driver of PCa progression and resistance to radiotherapy that functioned by interaction to macrophages, revealing potential therapeutic targets for advanced PCa.
Aerobic glycolysis metabolic reprogramming enabled the rapid provision of sufficient energy for tumor cells, which fostered an environment conducive to the growth of tumor cells. In regard to the development of novel tumor therapy, there is a rising demand for further identification of oncogenic signals mediated by aerobic glycolysis metabolic reprogramming within tumor cells. Modulation of tumor energy metabolism depended on signal transducers and activators of transcription (STAT) protein family to some extent [48]. Smith et al. revealed that activation of PRL-3 enhanced glycolysis, which, in a positive feedback loop, activated STAT1, thereby promoting the progression of multiple myeloma [49]. Findings were uncovered by Huangfu et al. that RBM4 modulated polarization to M1 macrophages through targeting glycolysis regulated by STAT1 [50]. Pitroda et al. also discovered that STAT1 promoted the progression of squamous cell carcinoma and resistance to radiotherapy by regulating glycolysis-related genes [51]. In our study, it was demonstrated that STAT1 enhanced transcriptional activities of HK2, PFKM and PKM2, the key glycolytic enzymes, thereby inducing secretion of lactate in PCa cells. This revealed a novel mechanism by which lactate derived from STAT1-mediated aerobic glycolysis regulated the interactive function between PCa cells and macrophages.
In addition, STAT1 played a dual role by exerting pro-oncogenic and anti-oncogenic effects simultaneously, and this dual function depended on related molecular mechanism. Mechanistically, a high level of ROS promoted anti-apoptotic effect by activation of JAK/STAT1 pathway, while STAT1 induced the generation of NQO1 confirmed as ROS scavenger, leading to a reduction of ROS, which was more effective in suppressing progression of tumor [52, 53]. Likewise, acted as tumor suppressor, STAT1 prolonged the cancer-specific survival in the early stage of PCa, its mechanism has not been clearly elucidated yet [24]. As a key transcription factor mediating interferon signaling, STAT1 typically exerts tumor-suppressive effects by activating genes involved in immune surveillance, which may underlie its protective role in early PCa. On the contrary, STAT1 facilitated resistance to enzalutamide and cancer stemness, dependent on upstream activation of JAK in the advanced stage of PCa [25]. Additionally, it has been proposed by Bachmann et al. that high expression of STAT1 in PCa exhibited a correlation with resistance to chemotherapy and radiotherapy [28]. The functional switch of STAT1 from a tumor suppressor to a promoter in advanced PCa likely arises from chronic activation within an altered tumor microenvironment, which reprograms its downstream transcriptional programs towards supporting survival, stemness, and therapy resistance. As discovered by our study, STAT1 overexpression obviously optimized the proliferative, migrative, invasive ability and induced resistance to radiotherapy of prostate cancer cells, but the pro-oncogenic and pro-resistant effect were significantly inhibited by STAT1 along with inhibition of aerobic glycolysis, indicating fulfillment of pro-oncogenic and pro-resistant effect relied on aerobic glycolysis of prostate cancer cells, thus clarifying a key mechanism whereby STAT1 promoted oncogenesis and conferred radiotherapy resistance in advanced PCa.
Transmembrane transfer activities of lactate were mediated by the monocarboxylate transporter (MCT) family. Végran et al. found that lactate transferred to endothelial cells by MCT1, activated NFκB1/IL-8 signal to enhance angiogenesis of breast cancer and colorectal cancer [42]. The result showed by Wang et al. that lactate transferred to macrophages by MCT1 strengthened histone lysine lactylation to induce expression of anti-inflammation cytokines [39]. Furthermore, several studies suggested that there was a close correlation between activation of NFκB1 pathway and polarization to M2 macrophages in both tumor microenvironment and inflammation microenvironment [54,55,56]. In this study, the regulatory function of macrophage polarization by lactate was determined with the treatment of MCT1 inhibitor and NFκB1 pathway inhibitor. This study indicated that lactate transferred to macrophages by MCT1 had utility as activator of NFκB1 pathway to promote polarization toward M2 macrophages.
NFκB1, which proved to be a transcription factor, was highly expressed in macrophages and regulated the expression of miscellaneous cytokines [57, 58]. Conditioned medium from THP-1 cells with a knockdown of NFκB1 was collected and probed on a protein microarray. Combined protein microarray with verified experiments, it was discovered that MCP-1 displayed a dramatic reduction of expression and secretion after NFκB1 knockdown. MCP-1 was reported as an indispensable chemokine for regulating migration and infiltration of macrophages. In TME, MCP-1 was regarded as a key signal driving the crosstalk among tumor cells, macrophages and CD8+T cells, shaping immunosuppressive microenvironment [59, 60]. Su et al. reported that MCP-1/CCR2 was an effective signal by recruitment for M2 macrophages and Treg, beneficial for immune escape and angiogenesis of PCa cells [61]. Klusa et al. and Qian et al. unveiled that MCP-1/CCR2 signal was associated with resistance to radiotherapy and chemotherapy in PCa, respectively [62, 63]. However, there is still a lack of a complete explanation for the effect and mechanism of MCP-1/CCR2 signal in the TME of PCa. It was demonstrated by Biswas et al. that MCP-1/CCR2 signal stimulated phosphorylation of JAK and triggered further activation of STAT1 in peritoneal macrophages from mice [64]. The phenomenon that JAK/STAT1 pathway was activated by MCP-1/CCR2 signaling to increase infiltration of macrophages and endothelial cells was observed by Shahrara et al. in a rat model of rheumatoid arthritis [65]. Our results illustrated that MCP-1 secreted by macrophages bound to CCR2 receptor of PCa cells, activating JAK/STAT pathway, further elevating glycolysis level, accordingly promoting progression of PCa. Moreover, mouse models were utilized to explore the effect of inhibitors targeting the pathways on tumor progression under irradiation. It was noteworthy that the use of CCR2 antagonist and JAK inhibitor in combination with radiation significantly inhibited tumor growth in vivo, which provided preliminary evidence to support clinical application of CCR2 antagonist and JAK inhibitor in assistance of radiotherapy.
As demonstrated above, our study revealed a positive feedback loop between macrophages and PCa cells in TME. The interactive mechanism was described detailly as following: a transcriptional enhancement of the glycolic key enzymes by STAT1 increased secretion of lactate. Lactate from PCa cells was transferred to macrophages by MCT1, accountable for activating NFκB1 and inducing MCP-1 secretion from macrophages further. It was worth being concerned that MCP-1 bound to CCR2 receptor of PCa cells, resulting in activation of JAK/STAT1 pathway. It was not ignored that in this positive feedback loop, STAT1 simultaneously promoted proliferation, invasion and migration of PCa by targeting glycolysis pathway and polarization to M2 macrophages by activation of NFκB1 signal. As the core molecular in this positive feedback loop, STAT1 and MCP-1 could be exploited as a potential therapeutic target for PCa.
However, there are limitations of this study to note. The specific molecular mechanism concerning the activation of NFκB1 by lactate has not been sufficiently clarified. Moreover, the M0 macrophages derived from THP-1 cells were used to simulate the macrophages in TME, but they were still incompletely substituted for primary macrophages extracted from PCa tissue. Finally, there is a requirement for exploring clinical applications for CCR2 receptor antagonist and JAK inhibitor.
Conclusion
This study revealed a positive feedback loop between PCa cells and macrophages. STAT1 activated the transcription of HK2, PFKM and PKM2, inducing secretion of lactate in PCa cells; lactate activated NFκB1 pathway via MCT1 transporter, on one hand, promoting macrophages polarization toward M2 phenotype, on the other hand, enhancing expression and secretion of MCP-1 from macrophages; Binding of MCP-1 to CCR2 receptor activated JAK/STAT1 pathway, accelerating the progression of PCa. This molecular network amplified the interactive signal between PCa cells and macrophages, driving progression and resistance to radiotherapy of PCa. In addition, accompanied by the preliminary exploration of the CCR2 antagonist and JAK inhibitor in vivo, the potential application value of improvement in radiotherapy sensitivity of PCa was initially found.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73:17–48.
Valle LF, Lehrer EJ, Markovic D, Elashoff D, Levin-Epstein R, Karnes RJ, et al. A systematic review and meta-analysis of local salvage therapies after radiotherapy for prostate cancer (MASTER). Eur Urol. 2021;80:280–92.
Haffner MC, Zwart W, Roudier MP, True LD, Nelson WG, Epstein JI, et al. Genomic and phenotypic heterogeneity in prostate cancer. Nat Rev Urol. 2021;18:79–92.
Chaiswing L, Weiss HL, Jayswal RD, Clair DKS, Kyprianou N. Profiles of radioresistance mechanisms in prostate cancer. Crit Rev Oncog. 2018;23:39–67.
Xiao Y, Yu D. Tumor microenvironment as a therapeutic target in cancer. Pharmacol Ther. 2021;221:107753.
Gao J, Liang Y, Wang L. Shaping polarization of tumor-associated macrophages in cancer immunotherapy. Front Immunol. 2022;13:888713.
Luo S, Yang G, Ye P, Cao N, Chi X, Yang WH, et al. Macrophages are a double-edged sword: molecular crosstalk between tumor-associated macrophages and cancer stem cells. Biomolecules. 2022;12:850.
JiaWei Z, ChunXia D, CunDong L, Yang L, JianKun Y, HaiFeng D, et al. M2 subtype tumor associated macrophages (M2-TAMs) infiltration predicts poor response rate of immune checkpoint inhibitors treatment for prostate cancer. Ann Med. 2021;53:730–40.
Zarif JC, Baena-Del Valle JA, Hicks JL, Heaphy CM, Vidal I, Luo J, et al. Mannose receptor-positive macrophage infiltration correlates with prostate cancer onset and metastatic castration-resistant disease. Eur Urol Oncol. 2019;2:429–36.
Prakash H, Klug F, Nadella V, Mazumdar V, Schmitz-Winnenthal H, Umansky L. Low doses of gamma irradiation potentially modifies immunosuppressive tumor microenvironment by retuning tumor-associated macrophages: lesson from insulinoma. Carcinogenesis. 2016;37:301–13.
Jones KI, Tiersma J, Yuzhalin AE, Gordon-Weeks AN, Buzzelli J, Im JH, et al. Radiation combined with macrophage depletion promotes adaptive immunity and potentiates checkpoint blockade. EMBO Mol Med. 2018;10:e9342.
Lee HL, Tsai YC, Pikatan NW, Yeh CT, Yadav VK, Chen MY, et al. Tumor-associated macrophages affect the tumor microenvironment and radioresistance via the upregulation of CXCL6/CXCR2 in hepatocellular carcinoma. Biomedicines. 2023;11:2081.
Bian Z, Shi L, Kidder K, Zen K, Garnett-Benson C, Liu Y. Intratumoral SIRPα-deficient macrophages activate tumor antigen-specific cytotoxic T cells under radiotherapy. Nat Commun. 2021;12:3229.
Johar D, Elmehrath AO, Khalil RM, Elberry MH, Zaky S, Shalabi SA, et al. Protein networks linking Warburg and reverse Warburg effects to cancer cell metabolism. Biofactors. 2021;47:713–28.
Fitzgerald G, Soro-Arnaiz I, De Bock K. The warburg effect in endothelial cells and its potential as an anti-angiogenic target in cancer. Front Cell Dev Biol. 2018;6:100.
Hayes C, Donohoe CL, Davern M, Donlon NE. The oncogenic and clinical implications of lactate induced immunosuppression in the tumour microenvironment. Cancer Lett. 2021;500:75–86.
Chen P, Zuo H, Xiong H, Kolar MJ, Chu Q, Saghatelian A, et al. Gpr132 sensing of lactate mediates tumor-macrophage interplay to promote breast cancer metastasis. Proc Natl Acad Sci USA. 2017;114:580–5.
Ohashi T, Aoki M, Tomita H, Akazawa T, Sato K, Kuze B, et al. M2-like macrophage polarization in high lactic acid-producing head and neck cancer. Cancer Sci. 2017;108:1128–34.
Fu XY, Schindler C, Improta T, Aebersold R, Darnell JE. The proteins of ISGF-3, the interferon alpha-induced transcriptional activator, define a gene family involved in signal transduction. Proc Natl Acad Sci USA. 1992;89:7840–3.
Lim CP, Cao X. Structure, function, and regulation of STAT proteins. Mol Biosyst. 2006;2:536–50.
Palakurthi B, Fross SR, Guldner IH, Aleksandrovic E, Liu X, Martino AK, et al. Targeting CXCL16 and STAT1 augments immune checkpoint blockade therapy in triple-negative breast cancer. Nat Commun. 2023;14:2109.
Jiang Q, Li J, Wang J, Zhang W. Inhibition of CDKL3 downregulates STAT1, thus suppressing prostate cancer development. Cell Death Dis. 2023;14:189.
Taniguchi H, Caeser R, Chavan SS, Zhan YA, Chow A, Manoj P, et al. WEE1 inhibition enhances the antitumor immune response to PD-L1 blockade by the concomitant activation of STING and STAT1 pathways in SCLC. Cell Rep. 2022;39:110814.
Hatziieremia S, Mohammed Z, McCall P, Willder JM, Roseweir AK, Underwood MA, et al. Loss of signal transducer and activator of transcription 1 is associated with prostate cancer recurrence. Mol Carcinog. 2016;55:1667–77.
Luo Y, Yang X, Basourakos SP, Zuo X, Wei D, Zhao J, et al. Enzalutamide-resistant progression of castration-resistant prostate cancer is driven via the JAK2/STAT1-dependent Pathway. Front Mol Biosci. 2021;8:652443.
Patterson SG, Wei S, Chen X, Sallman DA, Gilvary DL, Zhong B, et al. Novel role of Stat1 in the development of docetaxel resistance in prostate tumor cells. Oncogene. 2006;25:6113–22.
Wang S, Han L, Li J, Liu Y, Wang S. Inflammatory molecules facilitate the development of docetaxel-resistant prostate cancer cells in vitro and in vivo. Fundam Clin Pharmacol. 2022;36:837–49.
Bachmann SB, Frommel SC, Camicia R, Winkler HC, Santoro R, Hassa PO. DTX3L and ARTD9 inhibit IRF1 expression and mediate in cooperation with ARTD8 survival and proliferation of metastatic prostate cancer cells. Mol Cancer. 2014;13:125.
Grass GD, Alfonso JCL, Welsh E, Ahmed KA, Teer JK, Pilon-Thomas S, et al. The radiosensitivity index gene signature identifies distinct tumor immune microenvironment characteristics associated with susceptibility to radiation therapy. Int J Radiat Oncol Biol Phys. 2022;113:635–47.
Zhao S, Mi Y, Guan B, Zheng B, Wei P, Gu Y, et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J Hematol Oncol. 2020;13:156.
Wang YC, Wu YS, Hung CY, Wang SA, Young MJ, Hsu TI, et al. USP24 induces IL-6 in tumor-associated microenvironment by stabilizing p300 and β-TrCP and promotes cancer malignancy. Nat Commun. 2018;9:3996.
Jiang H, Wei H, Wang H, Wang Z, Li J, Ou Y, et al. Zeb1-induced metabolic reprogramming of glycolysis is essential for macrophage polarization in breast cancer. Cell Death Dis. 2022;13:206.
Xia Q, Jia J, Hu C, Lu J, Li J, Xu H, et al. Tumor-associated macrophages promote PD-L1 expression in tumor cells by regulating PKM2 nuclear translocation in pancreatic ductal adenocarcinoma. Oncogene. 2022;41:865–77.
Vandsemb EN, Rye MB, Steiro IJ, Elsaadi S, Rø TB, Slørdahl TS, et al. PRL-3 induces a positive signaling circuit between glycolysis and activation of STAT1/2. FEBS J. 2021;288:6700–15.
Majoros A, Platanitis E, Kernbauer-Hölzl E, Rosebrock F, Müller M, Decker T. Canonical and non-canonical aspects of JAK-STAT signaling: lessons from interferons for cytokine responses. Front Immunol. 2017;8:29.
Lyssiotis CA, Kimmelman AC. Metabolic interactions in the tumor microenvironment. Trends Cell Biol. 2017;27:863–75.
Mitchell JP, Carmody RJ. NF-κB and the transcriptional control of inflammation. Int Rev Cell Mol Biol. 2018;335:41–84.
Somma D, Kok FO, Kerrigan D, Wells CA, Carmody RJ. Defining the role of nuclear factor (NF)-κB p105 subunit in human macrophage by transcriptomic analysis of NFKB1 knockout THP1 cells. Front Immunol. 2021;12:669906.
Wang N, Wang W, Wang X, Mang G, Chen J, Yan X, et al. Histone lactylation boosts reparative gene activation post-myocardial infarction. Circ Res. 2022;131:893–908.
Manosalva C, Quiroga J, Hidalgo AI, Alarcón P, Anseoleaga N, Hidalgo MA, et al. Role of lactate in inflammatory processes: friend or foe. Front Immunol. 2021;12:808799.
Martinez-Outschoorn UE, Curry JM, Ko YH, Lin Z, Tuluc M, Cognetti D, et al. Oncogenes and inflammation rewire host energy metabolism in the tumor microenvironment: RAS and NFκB target stromal MCT4. Cell Cycle. 2013;12:2580–97.
Végran F, Boidot R, Michiels C, Sonveaux P, Feron O. Lactate influx through the endothelial cell monocarboxylate transporter MCT1 supports an NF-κB/IL-8 pathway that drives tumor angiogenesis. Cancer research. 2011;71:2550–60.
Li HS, Watowich SS. Innate immune regulation by STAT-mediated transcriptional mechanisms. Immunol Rev. 2014;261:84–101.
Xu M, Wang Y, Xia R, Wei Y, Wei X. Role of the CCL2-CCR2 signalling axis in cancer: mechanisms and therapeutic targeting. Cell Prolif. 2021;54:e13115.
Dong L, Myers KV, Pienta KJ. Understanding the tumor-immune microenvironment in prostate cancer. Curr Opin Oncol. 2021;33:231–7.
Cai J, Song L, Zhang F, Wu S, Zhu G, Zhang P, et al. Targeting SRSF10 might inhibit M2 macrophage polarization and potentiate anti-PD-1 therapy in hepatocellular carcinoma. Cancer Commun. 2024;44:1231–60.
Zhang A, Xu Y, Xu H, Ren J, Meng T, Ni Y, et al. Lactate-induced M2 polarization of tumor-associated macrophages promotes the invasion of pituitary adenoma by secreting CCL17. Theranostics. 2021;11:3839–52.
Li YJ, Zhang C, Martincuks A, Herrmann A, Yu H. STAT proteins in cancer: orchestration of metabolism. Nat Rev Cancer. 2023;23:115–34.
Smith CN, Blackburn JS. PRL-3 promotes a positive feedback loop between STAT1/2-induced gene expression and glycolysis in multiple myeloma. FEBS J. 2021;288:6674–6.
Huangfu N, Zheng W, Xu Z, Wang S, Wang Y, Cheng J, et al. RBM4 regulates M1 macrophages polarization through targeting STAT1-mediated glycolysis. Int Immunopharmacol. 2020;83:106432.
Pitroda SP, Wakim BT, Sood RF, Beveridge MG, Beckett MA, MacDermed DM, et al. STAT1-dependent expression of energy metabolic pathways links tumour growth and radioresistance to the Warburg effect. BMC Med. 2009;7:68.
Lee JK, Edderkaoui M, Truong P, Ohno I, Jang KT, Berti A, et al. NADPH oxidase promotes pancreatic cancer cell survival via inhibiting JAK2 dephosphorylation by tyrosine phosphatases. Gastroenterology. 2007;133:1637–48.
Totten SP, Im YK, Cepeda Cañedo E, Najyb O, Nguyen A, Hébert S, et al. STAT1 potentiates oxidative stress revealing a targetable vulnerability that increases phenformin efficacy in breast cancer. Nat Commun. 2021;12:3299.
Tian J, Chen W, Xiong Y, Li Q, Kong S, Li M, et al. Small extracellular vesicles derived from hypoxic preconditioned dental pulp stem cells ameliorate inflammatory osteolysis by modulating macrophage polarization and osteoclastogenesis. Bioact Mater. 2023;22:326–42.
Li Y, Chen Z, Han J, Ma X, Zheng X, Chen J. Functional and therapeutic significance of tumor-associated macrophages in colorectal cancer. Front Oncol. 2022;12:781233.
Xu F, Wei Y, Tang Z, Liu B, Dong J. Tumor‑associated macrophages in lung cancer: friend or foe? Mol Med Rep. 2020;22:4107–15.
Korbecki J, Simińska D, Gąssowska-Dobrowolska M, Listos J, Gutowska I, Chlubek D, et al. Chronic and cycling hypoxia: drivers of cancer chronic inflammation through HIF-1 and NF-κB activation: a review of the molecular mechanisms. International journal of molecular sciences. 2021;22:10701.
Zhang M, Liu ZZ, Aoshima K, Cai WL, Sun H, Xu T, et al. CECR2 drives breast cancer metastasis by promoting NF-κB signaling and macrophage-mediated immune suppression. Sci Transl Med. 2022;14:eabf5473.
Xie M, Lin Z, Ji X, Luo X, Zhang Z, Sun M, et al. FGF19/FGFR4-mediated elevation of ETV4 facilitates hepatocellular carcinoma metastasis by upregulating PD-L1 and CCL2. J Hepatol. 2023;79:109–25.
Miyamoto T, Murakami R, Hamanishi J, Tanigaki K, Hosoe Y, Mise N, et al. B7-H3 suppresses antitumor immunity via the CCL2-CCR2-M2 macrophage axis and contributes to ovarian cancer progression. Cancer Immunol Res. 2022;10:56–69.
Su W, Han HH, Wang Y, Zhang B, Zhou B, Cheng Y, et al. The polycomb repressor complex 1 drives double-negative prostate cancer metastasis by coordinating stemness and immune suppression. Cancer Cell. 2019;36:139–55.e10.
Klusa D, Lohaus F, Franken A, Baumbach M, Cojoc M, Dowling P, et al. Dynamics of CXCR4-positive circulating tumor cells in prostate cancer patients during radiotherapy. Int J Cancer. 2023;152:2639–54.
Qian DZ, Rademacher BL, Pittsenbarger J, Huang CY, Myrthue A, Higano CS, et al. CCL2 is induced by chemotherapy and protects prostate cancer cells from docetaxel-induced cytotoxicity. Prostate. 2010;70:433–42.
Biswas SK, Sodhi A. Tyrosine phosphorylation-mediated signal transduction in MCP-1-induced macrophage activation: role for receptor dimerization, focal adhesion protein complex and JAK/STAT pathway. Int Immunopharmacol. 2002;2:1095–107.
Shahrara S, Amin MA, Woods JM, Haines GK, Koch AE. Chemokine receptor expression and in vivo signaling pathways in the joints of rats with adjuvant-induced arthritis. Arthritis Rheum. 2003;48:3568–83.
Funding
This study was supported by Joint Funds for the Innovation of Science and Technology, Fujian Province (Grant No. 2025Y9143), the Major Scientific Research Program for Young and Middle-aged Health Professionals of Fujian Province, China (Grant No. 2022ZQNZD006) and the “Eyas Plan” Youth Top-notch Talent Project of Fujian Province (Grant No. SCYJHBJRC-XN2021).
Author information
Authors and Affiliations
Contributions
Writing—original draft: Jia-Yin Chen, Yu-Ting Xue, Bin Lin, Xu-Yun Huang, Fei Lin. Writing— review and editing: Zhi-Bin Ke, Ning Xu. Methodology: Jia-Yin Chen, Wan-Jin Zhang. Investigation: Yu-Ting Xue, Xu-Yun Huang. Formal analysis: Bin Lin, Dong-Ning Chen, Data curation: Fei Lin, Zhi-Bin Ke, Conceptualization: Jia-Yin Chen, Supervision: Yong Wei, Qing-Shui Zheng, Project administration: Xue-Yi Xue, Ning Xu.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval and consent to participate
All experiments were performed in accordance with relevant institutional guidelines and regulations of Fujian Medical University and national guidelines and regulations of China. All animal experiments were approved by the Experimental Animal Ethics Committee of Fujian Medical University (approval number: IACUC FJMU 2024-Y-1037). All methods were performed in accordance with the relevant guidelines and regulations of the ethics committee.
Consent for publication
Informed consent for publication of the manuscript was obtained from all authors.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by Boris Zhivotovsky
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, JY., Xue, YT., Lin, B. et al. A feedback mechanism from prostate cancer cells to macrophages, reinforced by STAT1, regulates tumor progression and resistance to radiotherapy. Cell Death Dis 17, 282 (2026). https://doi.org/10.1038/s41419-026-08577-5
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41419-026-08577-5
