
Abstract
Attenuated whole organism vaccines targeting the malaria liver stage reliably confer sterile immunity. These vaccines completely protect female mice from infection, but protection in male mice remains unproven. We discover that male mice vaccinated with prime-and-trap, a whole organism-based vaccine strategy, exhibit poorer protection against Plasmodium sporozoite challenge than females. We investigate this sex difference, and identify vaccinated males have fewer hepatic memory CD8+ T cells than females when scaling for liver biomass, and reduced inflammatory responses post-vaccination. Surgical hormone manipulation clarifies that the presence of testicular hormones hinders protection in male mice. The presence of androgens does not affect memory CD8+ T cell quantity nor quality, but reduces recruitment of CD8+ T cells in male liver tissues via a restricted inflammatory response. Here, we show both males and females form functional memory responses following prime-and-trap vaccination, but the presence of androgens during sporozoite challenge impair protection in male mice.
Similar content being viewed by others
Introduction
Vaccines that are durably effective are urgently needed to combat malaria, which resulted in 263,000 infections and 597,000 deaths in 20231. The complex lifecycle of Plasmodium parasites, the causative agent of malaria, begins with a bite from a mosquito carrying infectious sporozoites (spz). Spz migrate from the dermis to the bloodstream and are transported to the liver2. To initiate the liver stage, spz invade hepatocytes. After replication, hepatocytes release the infectious exoerythrocytic merozoites into the bloodstream initiating the disease-causing phase of Plasmodium infection and enabling onward transmission3. Since halting the liver stage could prevent disease and transmission, it is considered an attractive target for vaccine design.
Repeated vaccination with radiation-attenuated sporozoites (RAS) is the most protective experimental malaria vaccine to date that confers sterile immunity against the liver stage in humans4,5,6. The ability of RAS to protect against infection was first demonstrated in mouse models of Plasmodium7,8. RAS-based vaccines leverage the natural ability of spz to home to the liver while lacking the ability to advance to blood stage infection. Since their discovery, decades of research have sought to deconvolute the mechanisms of immune protection conferred by RAS and other similar attenuated spz-based liver stage vaccines. The collective data indicate that memory CD8+ T cells are a defining requirement for a protective immune response in the liver9,10,11. In particular, liver tissue-resident memory CD8+ T (Trm) cells and effector memory CD8+ (Tem) cells are important for durable protection6,12,13,14,15. Knowledge of these immune correlates of protection inspired the design of a heterologous liver stage vaccine called ‘prime-and-trap’ (P&T). The priming component of the P&T vaccine is designed to induce peripheral antigen-specific CD8+ T cells, and the trapping component is intended to recruit or ‘trap’ circulating antigen-specific memory CD8+ T cells in the liver to become liver Trms13,16. Different iterations of this P&T strategy have protected female mice16,17, but no data is so far available in male mice.
Sex-specific differences to vaccines are a well-documented phenomenon, with a general trend for females to mount more robust immune responses than males18,19,20. Three main sex-based factors drive differences in immune responses: sex hormones, sex-chromosome linked genes, and physiological differences18,19,21. These three factors intrinsically modify the tissue microenvironment22, leaving T cell-based vaccines especially susceptible to sex-divergent outcomes. The liver is also known to be one of the most sexually dimorphic organs in mice23 and humans24. Yet, pre-clinical studies of spz-based liver stage vaccines have almost exclusively relied on female mice. For example, in a review by Nganou-Makamdop and Sauerwein, of the 17 reviewed papers on spz immunization in mice, none reported using male mice, 12/17 reported using exclusively female mice, and 5/17 reported no sex information25 (Table ST1). This underscores the need to consider sex as a biological variable and delineate protection requirements in male mice.
The limited data we do have on the impact of sex on immune responses to liver stage malaria vaccines come from a single recent pre-clinical study26. Following repeated RAS vaccination, female mice were found to have more hepatic IFN-γ-producing CD8+ T cells and greater protection against P. berghei spz challenge than male mice. This reduced protection in males could be overcome by the removal of testicular hormones prior to vaccination26. Male mice are also known to experience more severe blood stage malarial disease than female mice across several rodent Plasmodium spp27,28. Thus, there is at least some evidence that biological sex modifies immune responses to both attenuated and virulent Plasmodium parasites.
Given the limited information on liver stage malaria vaccination in male mice, here we provide a comprehensive evaluation of P&T vaccine-induced responses on the basis of sex. Through a series of P&T immunization and challenge studies in male and female mice, we evaluate sex-specific differences in protection outcomes and memory CD8+ T cell populations. We further assess the contribution of sex hormones in modifying protection and the quality, quantity, and spatial recruitment of P&T-induced memory CD8+ T cells. Together, this study provides an in-depth characterization of the impact of biological sex and sex hormones on a liverstage malaria vaccine.
Results
Male mice exhibit less protection after prime-and-trap vaccination
To assess the influence of sex on the efficacy on liver stage malaria vaccines, we utilized a two-dose heterologous P&T regimen16 in male and female BALB/cJ mice. This P&T regimen involves epidermal DNA plasmid vaccination by gene gun on days 0 and 2. The DNA vaccine encodes a P. yoelii circumsporozoite protein (CSP) minigene designed to maximize CD8+ T cell activation and minimize antibody responses16. Twenty-eight days after DNA vaccination, a single dose of 2 × 104 P. yoelii RAS (Py-RAS) was injected IV to trap circulating CSP-specific CD8+ T cells in the liver. Mice were challenged 28 days later with IV administered 0.5–1.0 × 103 infectious wild-type P. yoelii spz (Py-WT) (Fig. 1a) and monitored for blood stage patency to assess protection. All female mice were protected (19/19), whereas only 15% of male mice (3/20) were protected (Fig. 1b). Regardless of sex, all naïve mice were susceptible to infection.

a Scheme of male (M) and female (F) BALB/cJ mice prime-and-trap (P&T) vaccination experiments. b Percentages of mice protected or not protected against challenge with 0.5 − 1.0 × 103 Py-WT sporozoites, as measured by blood parasitemia up to day 14 post-challenge. Numbers above bars indicate numbers of protected mice out of the total mice challenged, derived from 4 independent experiments. c Representative rainbow images of luminescence in livers 44 h post infection (hpi) with 1 × 104 Py-Luc sporozoites. Rainbow scales are expressed in radiance (p/s/cm2/sr). d Quantification of bioluminescent signal in log-transformed total flux (p/s) from mice at 44 hpi (2 independent replicates, n = 8–10/group). Groups in b were compared using a two-sided Fisher’s exact test, and groups in d were compared by a two-sided Wilcoxon test; only relevant comparisons are depicted. Error bars represent mean ± s.e.m; ns = p > 0.05, ***p < 0.001. Source data are provided as a Source Data file.
To determine if vaccinated male mice could achieve partial liver stage protection, we next monitored liver burden by in vivo bioluminescent imaging in P&T vaccinated mice of both sexes after virulent spz challenge. Mice were challenged 28 days after P&T using 1 x 104 P. yoelii spz that constitutively express a GFP-luciferase fusion protein (Py-Luc)29 and evaluated at 44 h post-injection (hpi). Naïve male and female mice were likewise challenged. Vaccinated female mice showed a 45-fold reduction in luminescent signal compared to naïve female mice, whereas vaccinated male mice showed only an 11-fold reduction compared to naïve males (Fig. 1c, d). Notably, there was no difference in luminescence signal at 44 hpi between naïve female and male mice (Fig. 1c, d), suggesting that liver stage replication of the parasite was similar for both sexes. Additionally, augmenting P&T with a more potent prime or with a co-administered Py-RAS with the glycolipid adjuvant 7DW8-530 did not overcome the low rate of protection observed in male mice (Fig. S1).
To confirm that sex-bias in protection was not unique to this mouse and parasite strain, we conducted similar P&T studies using C57BL/6 mice and P. berghei parasites for trapping (Pb-RAS) and challenge (Pb-WT). DNA encoding PbTRAP and PbRPL6 was used for priming, since these two antigens can be recognized in C57BL/617,31 (Fig. S2a). As expected, P&T vaccinated female C57BL/6 mice showed increased protection over vaccinated male mice (Fig. S2b, c), confirming that P&T vaccination induced a sex-bias in protection across two mouse malaria models.
Female mice generate higher density of memory CD8+ T cells in the liver
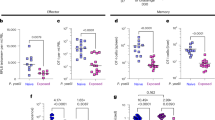
Given the importance of T cell responses to spz vaccines13,32, we first sought to phenotype memory CD8+ T cells in male and female mice following P&T vaccination (Fig. 2a). Twenty-eight days after vaccination, we isolated liver and spleen lymphocytes and ex vivo stimulated with PyCSP280-288 (SYVPSAEQI) peptide prior to intracellular cytokine staining (ICS) to evaluate CD8+ T cells at this memory timepoint. PyCSP280-288 is a known immunodominant epitope in the Py-RAS/BALB/cJ mouse model, and responsive cells represent a major population of antigen-specific CD8+ T cells after P&T vaccination16. A higher total CD4/CD8 ratio was observed in vaccinated male livers compared to female livers, but no significant difference in the CD4/CD8 ratio was found in the spleen (Fig. 2b, c, S5a). Male livers weighed significantly more than female livers in both naïve and vaccinated mice (Fig. 2e); thus, liver weights were incorporated in the analysis (results expressed per gram of liver) to account for the potential impact of sex differences in physical size on T cell numbers.

a Male (M) and female (F) BALB/cJ mice were vaccinated with the prime-and-trap (P&T) regimen. 28 days later, liver cells were analyzed by flow cytometry. b Representative flow plots of CD4 and CD8 surface markers on CD3+ cells isolated from the liver of mice. c Ratio of CD4-to-CD8 T cells in the liver of P&T vaccinated (+) and unvaccinated mice (-) female (F) and male (M) BALB/cJ mice. d Representative flow plots of IFN-γ (top), CD107a (middle), and TNF (bottom) on CD8+ T cells isolated from the liver and ex vivo stimulated with PyCSP280-288 peptide. e Liver weights of unvaccinated (open circle) or P&T vaccinated (closed circle) mice. f, g Quantification of CD8+ T cells responding to PyCSP280-288 (SYVPSAEQI) peptide defined by total number of IFN-γ+ and/or TNF+ and/or CD107a+ producing CD8+ T cells per gram of liver (f) and as proportion of CD3+ T cells (g). h, i Quantification of proportion of IFN-γ+ (left), TNF+ (middle), CD107a+ (right) (h), and IFN-γ+ TNF+ CD107a+ (i) producing CD8+ T cells in the liver. Detailed gating information is included in Supplementary Fig. S3. Data for c–i are shown from 3 independent experiments (n = 14 P&T groups, n = 5–7 unvaccinated group per sex). j Representative flow cytometry plot depicting CSP-tetramer+CD44hi CD8+ T cells Trm, Tem, and Trm phenotypes by markers CD69 (left) and CXCR3 (right). See supplementary Fig. S4 for the gating tree. k Frequency of CSP-tetramer+CD44hi Tem (CD62L-CD69-), Tcm (CD62L+CD69-), Trm (CD62L-CD69+) subsets of CD8+ T cells in the liver. Individual p-values for significant differences are shown, colored to correspond to memory T cell subsets where relevant. l. Frequency of CSP-tetramer+CXCR3+CD62L-CD69+CD44hi cells of CD8+ T cells in the liver. Data for j–l are shown from 2 independent experiments (n = 8–9). Statistical significance for data in c, f–i, k–I, was determined by Kruskal-Wallis test with Dunn’s multiple comparison and data in e was evaluated with a two-sided Wilcoxon Test; error bars represent mean ± s.e.m; ***p < 0.001, **p < 0.01, *p < 0.05, ns p > 0.05. Source data are provided as a Source Data file.
With differences in liver weight by sex acknowledged, we then evaluated the vaccine-induced memory CD8+ T cell populations. Upon peptide stimulation, liver Trm cells generated by P&T vaccination are known to express IFN-γ, TNF, and the degranulation marker CD107a13. We thus evaluated the total cell number, frequency, and effector function of CSP-specific CD8+ T cells isolated from the liver and spleen of male and female P&T-immunized mice. First, we examined the number of total cytokine-producing CSP-specific CD8+ T cells defined as any cell expressing IFN-γ, TNF, CD107a, or any combination of the three (Fig. 2d). Females showed higher cell counts per gram of liver compared to males (Fig. 2f). However, when evaluating total CSP-specific liver CD8+ T cells as a percent of CD3+ T cells, cytokine-producing CD8+ T cell frequencies did not significantly differ between male and female mice (Fig. 2g). Second, we evaluated the relative production of IFN-γ, TNF, and CD107a within the respective CD8+ T cell population. Vaccinated male and female mice had similar frequencies of IFN-γ, TNF, and CD107a producing CD8+ T cells in the liver (Fig. 2h). Third, we evaluated a putative cytotoxic ‘functional’ memory population defined as CD8+IFN-γ+TNF+CD107a+ 33,34. Within the liver CD8+ T cells repertoire, cells expressing all three markers were detected at similar frequencies in both male and female mice (Fig. 2i). In the spleen, there were also similar frequencies of IFN-γ+TNF+CD107a+ producing CD8+ T cells in male and female mice (Fig. S5e).
We also evaluated antigen-specific memory CD8+ T cells induced by P&T vaccination in another experiment using the H-2kd tetramer specific for MHC class I peptide Kd-PyCSP280-288. We defined putative Trm cells as CSP-tetramer+CD62L-CD69+CD44hiCD8+or CSP-tetramer+CXCR3+CD62L-CD69+CD44hiCD8+ (Fig. 2j). P&T vaccinated females and males showed an increased frequency of CSP-specific Trm cells in the liver at 28 days post-immunization, though males did not reach significance. Using either definition of CSP-specific Trm cells in the liver, there was no significant difference in Trm cell frequency between vaccinated male and female mice as reported as a proportion of CD8+ T cells (Fig. 2k, l, S6d, e). In the spleen, most cells had an effector memory phenotype (Tem: CSP-tetramer+CD62L-CD69-CD44hi), and there were no significant differences in the proportion of CSP-specific Tem cells between vaccinated male and female mice (Fig. S6d, e).
Taken together, P&T vaccination induces more antigen-specific memory CD8+ T cells per gram of liver by ICS in female mice compared to male mice. However, the frequency of these cells as a proportion of CD3+ T cells did not differ between male and female mice. Also, there were no detectable difference in effector function between male and female mice based on expression of IFN-γ, TNF, and/or CD107a. Finally, there was no detectable difference in the frequency of putative Trm cells. Thus, sex divergent outcomes in P&T vaccinated mice are potentially associated with differences in density, but not the quality, of vaccine-induced memory CD8+ T cells.
Sex-bias in hepatic inflammatory response to RAS immunization
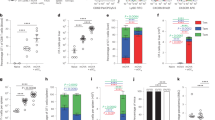
We examined gene expression profiles following RAS immunization to contextualize the sex-specific microenvironment that traps and guides memory CD8+ T cells in the liver. The acute inflammatory response of the liver following attenuated spz immunization mediates the formation of functional hepatic spz-specific CD8+ T cell responses35. Thus, to capture the transcriptional profile during initial innate phase and peak CD8+ T cell production phase, bulk mRNA sequencing was performed on whole livers isolated 44 h or 6 days post-RAS immunization. Mice were either primed with DNA encoding PyCSP or were naïve, then immunized with Py-RAS (Fig. 3a). Principal component analysis (PCA) identified sex as a major driver of variation in the transcriptome, mapping to the first principal component (PC1) that explained 35.6% of variation (Fig. 3b), even when omitting sex chromosomal-linked contributions (Fig. S7a) and stratifying by vaccination status (Fig. 3c). Differential expression analysis of mock-infected male and female mice confirmed differentially expressed genes (DEGs) in the Cytochrome P450 (Cyp) enzyme superfamily (Fig. S7b), aligning with known sex differences in the liver36. DEGs were examined relative to mock-infected controls within sex to account for this baseline variability between sexes (Fig. S7d).

a Experimental schematic. Livers from mock vaccinated, prime-and-trap vaccinated (P&T), or RAS-only (RAS) vaccinated male (M) and female (F) BALB/cJ mice were collected 44 h or 6 days after the indicated vaccine for RNAseq (n = 4 per group). b Principal Component (PC) analysis of male and female transcriptomes following vaccination (containing all vaccinated and mock-vaccinated mice). c Hierarchical clustering on differentially expressed genes (FDR < 0.05 and |LogFC | > 1.5, Benjamini-Hochberg adjusted) and unique to response to vaccination (not shared with baseline differences between the mock-vaccinated male and female mice) with column annotations for sex, vaccine status, and timepoint. Euclidean distance metric was used for sample clustering. d GSEA of selected Hallmark (top) and Gene Ontology Biological Processes pathways (bottom) with genes ranked by fold-change values relative to mock injected mice by sex. NES = normalized enrichment score. Column annotations depict sex and vaccine status as designated in panel (c). e Heatmap and hierarchal clustering of genes that contain at least one differentially expressed gene in either timepoint (44 h or 6 days) (FDR < 0.01, |LogFC | > 2, Benjamini-Hochberg adjusted) and appear in the Gene Ontology Biological Processes pathway ‘T cell activation’. Column annotations depict sex, timepoint, and vaccine status. f Calculated pathway score of aggregate samples by T cell activation pathways for the 44 h and 6-day timepoint (n = 8/group) relative to mock samples (n = 4). Detailed explanation of calculation in Methods section. g Heatmap and hierarchal clustering of genes that contain at least one differentially expressed gene at the 6 days timepoint (FDR < 0.05, |LogFC | > 1, Benjamini-Hochberg adjusted) and appear in the Hallmark ‘Interferon alpha response’, or Gene Ontology Biological Processes pathways ‘Phagocytosis’ and ‘Response to chemokine’. Column annotations depict sex and vaccine status (c). h Calculated pathway score of aggregate samples by ‘Interferon alpha response’, ‘Phagocytosis’, and ‘Response to chemokine’ pathways at day 6 post-immunization (n = 8/group) relative to mock samples (n = 4). Data produced from one independent replicate. The figure f and h represent the mean value with 95% confidence intervals; **p < 0.01, *p < 0.05, ns p > 0.05.
Prior studies comparing early and late-arresting whole spz vaccines in female mice have determined that type I IFN gene signature at the time of immunization impairs hepatic CD8+ T cell memory via a CD8+ T cell-extrinsic mechanism35. IFN signaling pathways have also been a documented response to wild-type37,38 and attenuated spz39 in the liver. Yet, despite a well-established female bias toward heightened interferon signaling responses upon inflammation21, a direct comparison between sexes following attenuated spz delivery remains to be reported. Thus, we performed gene sequence enrichment analysis (GSEA) on previously defined transcriptional patterns40,41. During the initial innate immune response phase at 44 h post-RAS immunization, male and female mice responded similarly for several pathways, including IFN-α response(Fig. S7e, Fig. 3d). Further evaluation of differentially expressed genes identified a gene signature shared between male and female mice related to liver inflammation (Saa1, Saa3) (Fig. S7c).
The day 6 timepoint captures the transcriptome during peak T cell recruitment, proliferation, and ongoing innate immune cell recruitment in the liver following clearance of RAS from infected hepatocytes4,32. Consistent with this, the gene ontology pathway for T cell activation was significantly induced in both vaccinated male and female mice at this timepoint (Fig. 3d), though females experienced a significantly higher response than male mice (Fig. 3e, f). The T cell activation pathway includes genes indicative of a CD8+ T cell response, including key genes encoding CD8, CD3, and CD28 surface markers. It is notable that there was a lack of detectable Plasmodium 18S rRNA at 6 days post-immunization, indicating that transcriptomes were driven by inflammation resulting from RAS clearance and the onset of adaptive cellular responses (Fig. S7f). Indeed, unsupervised clustering revealed that liver gene expression signatures clustered first by sex, then by timepoint and vaccine type (Fig. 3c).
Finally, further evaluation of the inflammatory environment during this T cell response at day 6 was conducted. Networks associated with responses to IFN-α were upregulated in female mice, but not in male mice at this timepoint (Fig. 3d). Further investigation found little to no induction of genes related to proteins associated with a type I IFN response (Gbp2, Gbp3, ifi44, ifit2) and CXCR3 ligands (Cxcl9, Cxcl10) in male mice (Fig. 3g). Female mice also experienced an increased IFN-γ response compared to male mice (Fig. 3d, S7g, h). Several other pathways were evaluated due to their potential to influence hepatic CD8+ T cell memory responses38: phagocytosis was upregulated in females but not males, whereas both male and female mice induced responses to chemokines at this timepoint (Fig. 3g, h). Overall, this suggests that while male mice can form chemokine cues important for CD8+ T cells, there is a diminished response to inflammatory signals, like interferons, when compared to female mice.
Androgens, not estrogens, alter protection from malaria challenge
Given the known effects of sex hormones on altering adaptive immune responses, we sought to evaluate the impact of sex hormones on protection from P&T vaccination. To evaluate whether estrogens in females or androgens in males contributed to the discordance in protection by sex following vaccination, we removed the gonads in adult mice at least 14 days prior to P&T vaccination and then later quantified liver burden 44 h post-spz challenge by Plasmodium 18S rRNA RT-PCR to assess protection (Fig. 4a). For females, ovariectomy (OVX, removing estrogens and progestins) had no effect on protection outcome, with OVX females maintaining protection (Fig. 4b, S8a). For males, orchiectomy (ORX, removing androgens) led to high levels of protection, a reversal of the outcome in intact males (Fig. 4c, S8b). IFN-γ ELISPOT on splenocytes obtained at post-challenge timepoints did not demonstrate significant differences in IFN-γ producing cells between intact and OVX females or intact and ORX males (Fig. S8d). Taken together, this data illustrates that among female mice, OVX to ablate circulating estrogens does not affect protection outcomes, whereas ORX to ablate androgens prior to vaccination significantly improves protection in male mice.

a Experimental regimen with orchiectomy (ORX) in males (M), ovariectomy (OVX) in females (F), or sham (SHAM) surgery followed by prime-and-trap (P&T) vaccination. 28 days later, mice were either challenged or lymphocytes were analyzed by flow cytometry. b, c Protection following 1 × 103 Py-WT sporozoite challenge defined as no detectable pan-Plasmodium 18S rRNA copies 44 h post-challenge. Data are shown from 2 independent experiments (n = 8–10/group). Only relevant comparisons are shown. d Liver weight of ORX and SHAM operated male BALB/cJ mice vaccinated with P&T regimen and harvest 28 days later. e Quantification of total responsive CD8+ T cells to PyCSP280-288 peptide defined by total number of IFN-γ+, TNF+, and/or CD107a+ producing CD8+ T cells corrected by weight of liver. f Ratio of CD4-to-CD8 T cells in the liver. g Total number of IFN-γ+, TNF+, and/or CD107a+ producing CD8+ T cells as proportion of CD3+ T cells in the liver. h, i Quantification of proportion of IFN-γ+ TNF+ CD107a+ (h) and IFN-γ+ (left), TNF+ (middle), and CD107a+ (right) producing CD8+ T cells in the liver. Data for d–i are shown from 2 independent experiments (n = 7–9/group). j Frequency of CSP-tetramer+CD44hi Tem (CD62L-CD69-), Tcm (CD62L+CD69-), Trm (CD62L-CD69+) subsets of total CD8+ T cell. k Quantitative evaluation of TCF1 MFI on CSP-tetramer+ Trm cells in the liver (CSP-tetramer+CD62L-CD69+CD44hiCD8+ T cells). l, m Proportion of PD-1+ (l) and LAG-3+ (m) on liver Trm cells. Data for j–m are shown from 2 independent experiments (n = 7–9/group). Statistical significance for data in b and c was determine by two-sided Fisher exact test; data in e–j was determined by Kruskal–Wallis test with Dunn’s multiple comparison and data in d and k–m was evaluated with the two-sided Wilcoxon Test. In j, individual adjusted p-values for significant differences are shown, colored to correspond to memory T cell subsets where relevant. Error bars represent mean ± s.e.m; box plot depicts median, interquartile range, and whiskers extending to maximum and minimum values within 1.5x of the interquartile range; ***p < 0.001, **p < 0.01, *p < 0.05, ns p > 0.05. Source data are provided as a Source Data file.
Androgens do not alter hepatic CD8+ T cell memory populations
Next, we characterized the impact of androgens on the memory CD8+ T cell repertoire in male mice after P&T immunization by ICS. Male mice underwent ORX or equivalent sham surgeries (SHAM), were P&T vaccinated, and then the vaccine response at 28 days post-immunization was evaluated as above. As previously reported42, we observed that livers from vaccinated ORX mice were significantly smaller compared to the vaccinated SHAM mice (Fig. 4d). We analyzed the number of CSP-specific CD8+ T cells per gram of tissue, and did not find a significant difference between ORX and SHAM mice (Fig. 4e). There was no difference in the CD4/CD8 T cell ratio in the livers of ORX and SHAM mice (Fig. 4f). Additionally, there was no significant difference in the frequency of CSP-specific CD8+ T cells as a percent of CD3+ T cells in the liver (Fig. 4g) and the spleen (Fig. S8e, f). Next, we assessed differences in cytokine-producing CD8+ T cells in ORX P&T immunized mice upon re-stimulation compared to SHAM mice. Both P&T vaccinated groups had increased levels of cytokine producing CD8+ T cells as compared to the naïve controls, but there was no significant difference in the frequency of IFN-γ+, TNF+, or CD107a+ T cells, or T cells expressing all three markers in the liver between the ORX and SHAM groups (Fig. 4h, i). A similar pattern was seen when comparing the frequency of IFN-γ+TNF+CD107a+ T cells in the spleen (Fig. S8g). Thus, androgens do not appear to alter the total number, frequency, or function of CSP-specific CD8+ T cells memory cells in the liver as measured by ICS.
In a separate experiment, we characterized the impact of androgens on memory CD8+ T cell subsets in ORX versus SHAM male mice after P&T vaccination by tetramer staining. ORX and SHAM male mice showed similar frequencies of total memory CSP-tetramer+CD44hiCD8+ T cells in the liver and spleen (S9a,d). Although Trm and Tem phenotypes were present in both ORX and SHAM vaccinated mice livers and spleen, there were no differences in Trm or Tem composition (Fig. 4j, S9b, e). Taken together, androgens do not appear to alter the frequency of CSP-specific CD8+ T cells memory subsets in the liver.
Since there was no difference in the number or proportion of CSP-specific CD8+ T cells in the livers of immunized ORX and SHAM male mice 28 days post-immunization, other factors must mediate the improved protection that occurs after withdrawal of androgens. We next evaluated the quality of CSP-tetramer+ Trm cells for markers of dysfunction. Androgen receptor signaling has been linked to a more dysfunctional T cell state via the transcription factor TCF143,44,45. Thus, we hypothesized that the presence of androgens may alter TCF1 expression and increase expression of co-inhibitory molecules like programmed cell death protein (PD-1) and lymphocyte activation gene 3 (LAG-3) that have also been connected to dysfunctional states46. As expected, higher expression of TCF1 was observed on central memory CD8+ T cells (Tcm) in the liver than on Tem and Trm cells (Fig. S9g). However, importantly, there was no difference in TCF1, PD-1, and LAG-3 expression on tissue resident CD8+ T cells (defined as CSP-tetramer+CD62L-CD69+CD44hi CD8+ T cells) in ORX compared to SHAM vaccinated mice (Fig. 4k–m).
In all, there was no detectable difference between the frequency, density, functionality profiles, or exhaustion/dysfunction status, of antigen-specific CD8+ T cells in male mouse livers at 28 days post-immunization following ORX or SHAM surgeries.
Hormone environment at time of challenge impacts protection outcomes
The question remained: why are intact male mice not protected by vaccination? Theoretically, androgens could influence the efficacy of P&T vaccination during two potential phases: 1) the production phase of immune memory at the time of vaccination, or 2) the recall phase of immune memory at the time of challenge. To identify at what stage androgens alter protection outcomes, ORX was performed before or after P&T vaccination and then mice were challenged three weeks after the post-P&T ORX procedures to allow the hormone environment in the liver to re-equilibrate (Fig. 5a). SHAM mice likewise received surgery at the post-P&T timepoint. As in previous experiments, male mice were protected by P&T when ORX was performed prior to vaccination. Remarkably, when ORX was performed after P&T vaccination, the mice were also protected (Fig. 5b, S10). However, since ORX is irreversible, this method of hormone manipulation did not allow for evaluation of the effect of androgens uniquely at the time of vaccination.

a Schedule of orchiectomy (ORX) in male (M) mice before or after prime-and-trap (P&T) vaccination as shown. After the completion of surgery at day 56, mice were rested 21 days to establish a new hormone equilibrium and then challenged with 1 × 103 Py-WT sporozoites. b Protection after Py-WT sporozoite challenge by pan-Plasmodium 18S rRNA RT-PCR from livers collected 44 h post-challenge. Data are shown from 2 independent experiments (n = 8–10/group). c Schedule of administration of acyline (Acy) in male mice at day 5 and 4 prior to either P&T vaccination (prior to gene gun and RAS) or prior to challenge, or both. Testosterone (T) add-back group received 3 injections of testosterone on day 3 and 1 prior to challenge and 1 day following challenge. Mice were challenged 28 - 42 days post P&T vaccination. d Protection after Py-WT sporozoite challenge by pan-Plasmodium 18S rRNA RT-PCR from livers collected 44 h post-challenge. Data are shown from 2 - 3 independent experiments (n = 10–14/group). Statistical significance for data in b and d was determined by a two-sided Fisher exact test. Only relevant comparisons are shown. Error bars represent mean ± s.e.m; ***p < 0.001, **p < 0.01, *p < 0.05, ns p > 0.05. Source data are provided as a Source Data file.
To further interrogate at which phase of vaccination and challenge androgens were exerting immune effects, we developed an acute and reversible model to suppress testosterone production in male mice. Acyline is a gonadotropin-releasing hormone (GnRH) antagonist that suppresses downstream production of testosterone in a dose-dependent manner and has previously been applied in mice47, and humans48,49. We optimized the dose of acyline for the BALB/cJ mouse model by selecting the dose that most consistently retained suppressed testosterone levels for over 7 days but under 14 days to ensure that hormone levels were maintained during peak immune responses of the P&T regimen (Fig. S11a, b). Next, we depleted testosterone prior to both steps of P&T vaccination, prior to spz challenge, or both (Fig. 5c). We found reducing testosterone at vaccination or challenge was sufficient to increase protection outcomes in male mice (Fig. 5d). To further confirm that testosterone at time of challenge altered protection outcomes, we added back testosterone and once again saw a reduction in protection (Fig. 5d). We further confirmed that administration of acyline, testosterone, and ORX surgery did not significantly alter liver burden in unvaccinated mice after a challenge dose of 1000 Py-WT sporozoites (Fig. S11f, g). Together with the above liver CD8+ T cell characterization data, these findings lead us to conclude that the hormone environment at the time of challenge is a driving factor behind limited vaccine-induced protection in intact male mice.
Androgens inhibit CD8+ T cell recruitment response
The mechanisms by which vaccine-induced CD8+ T cells enact their effector mechanisms at the time of challenge involve three mechanisms: cytolytic pathways, cytokine pathways33, and “sensing and alarm” to facilitate recruitment of other immune cells50. To study the direct impact of androgens on CD8+ T cells cytolytic enzyme and cytokine production, we used an ex vivo stimulation assay where we exogenously supplied dihydrotestosterone (DHT), the metabolically active form of testosterone, and evaluated the impact on CD8+ T cells in an otherwise hormone-free media environment. Intact male and female mice, and ORX male mice were P&T vaccinated (Fig. 6a), and lymphocytes were isolated from the liver and stimulated with PyCSP280-288 peptide in the presence or in the absence of DHT (Fig. 6b). CD8+ T cells from ORX male mice showed a reduction in IFN-γ and Granzyme B, but not TNF and CD107a, in the presence of DHT (Fig. 6c, d). In the presence of DHT, there was no reduction in proportion of a putative functional CD8+ T cells expressing IFN-γ, TNF and CD107a (Fig. 6e). However, in female mice Granzyme B+ functional CD8+ T cells were reduced (Fig. 6f). Thus, overall, the presence of DHT reduced the activity of CD8+ T cells via inhibition of IFN-γ+ and Granzyme B+, however, this fold reduction was quite slight.

a Female (F), male (M), and orchiectomized male (ORX) BALB/cJ mice were vaccinated with the prime-and-trap (P&T) regimen. 35 days later, liver cells were analyzed by flow cytometry. b Representative flow plots of IFN-γ (top), TNF (middle top), CD107a (middle bottom), and Granzyme B (bottom) on CD8+ T cells isolated from the liver and ex vivo stimulated with PyCSP280-288 peptide in hormone free condition with dihydrotestosterone added to media ( + DHT) or with ethanol control (- DHT). c, d Quantification of proportion of cytokines IFN-γ (c - left) and TNF (c -right) and cytolytic markers Granzyme B (d - left) and CD107a (d - right) producing CD8+ T cells in the liver. e Quantification of proportion of IFN-γ+TNF+CD107a+ producing CD8+ T cells in the liver. f Quantification of proportion of Granzyme B+IFN-γ+TNF+CD107a+ producing CD8+ T cells in the liver. Data are shown from 2 independent experiments (n = 10-12/group, naïve composed of n = 6 per sex). Statistical significance for data in (c–f) was determined by a paired two-sided Wilcoxon Test. Box plot depicts median, interquartile range, and whiskers extending to maximum and minimum values within 1.5x of the interquartile range; ***p < 0.001, **p < 0.01, *p < 0.05, ns p > 0.05. Source data are provided as a Source Data file.
Next, we evaluated the impact of androgens on the “sensing and alarm” function of Trm cells, which involves antigen-specific Trm cells serving as a local sensor and initiating an innate-like alarm signal to recruit more CD8+ T cells and other immune cells. Previous work identified a positive association between the number of CD8+ T cells per infected hepatocyte and P. yoelii parasite clearance51. Thus, we used immunohistochemistry to quantify inflammatory foci and CD8+ cells52 in P&T vaccinated and unvaccinated female, male, and ORX male mice challenged with Py-WT spz. For this experiment, we intentionally selected a high challenge dose of 1 x 105 spz to ensure sufficient immune foci for spatial quantification (Fig. 7a, b). As expected, we observed a higher number of inflammatory foci and CD8+ cells at 38 - 44hpi in P&T vaccinated mice compared to unvaccinated mice challenged with Py-WT spz and mock-injected control mice (Fig. 7c). Within the vaccinated mice, female and ORX male mice had more and larger inflammatory foci compared to male mice (Fig. 7d, e). Furthermore, within each inflammatory focus, there was a higher total number and density of CD8+ cells in female and ORX male mice compared to intact male mice (Fig. 7f–h). Thus, the presence of androgens reduced the number and size of inflammatory foci and impaired the recruitment of CD8+ cells to these foci in vaccinated male mice compared to female or ORX male mice.

a Female (F), male (M), and orchiectomized (ORX) male BALB/cJ mice were vaccinated with the prime-and-trap regimen (P&T) or left unvaccinated (-). 28 days later, mice were challenged with 1×105 Py-WT spz and liver was split for fixation for immunohistochemistry and RT-PCR of host genes between 38 and 44 h. b Representative inflammatory foci as determined with immunohistochemistry (IHC) in liver tissue captured under light microscope in vaccinated (P&T + Py-WT), unvaccinated (Py-WT), and mock-challenge mice where red demarks CD8+ cells and blue marks nuclear stain. One slice of left lateral lobe was analyzed per mouse and all inflammatory foci were counted within each lobe. c Average number of inflammatory foci per mouse corrected to tissue size evaluated (mm2) in vaccinated, unvaccinated, and unvaccinated mock-challenged mice. d, e Average number of inflammatory foci (d) and size of foci (μm2) (e) in female (F), male (M), and orchiectomized male (M ORX) mice that were vaccinated (+) and challenged with spz. f The average number of CD8+ cells within foci per each vaccinated, unvaccinated, and unvaccinated mock-challenged mouse. g, h. Within vaccinated and challenged F, M, and M ORX mice, the number of CD8+ cells in each foci and the density of CD8+ cells scaled to size of respective foci (CD8+ cells per foci size (mm2)). i Gene expression of Cxcl9, Cxcl10, Ifng in vaccinated and challenged female, male, and ORX male mice calculated as log2 fold change relative to pooled mock-challenged mice. Data are shown from 2 independent experiments (n = 5-8/group), except for the male P&T + Py-WT ORX group that was included in one experiment (n = 5 mice). Average values were calculated per mouse (c, d, f). Statistical significance for data was determined with Kruskal-Wallis test with Dunn’s multiple comparison. Error bars represent mean ± s.e.m; box plot depicts median, interquartile range, and whiskers extending to maximum and minimum values within 1.5x of the interquartile range; ***p < 0.001, **p < 0.01, *p < 0.05, ns p > 0.05. Source data are provided as a Source Data file.
Next, having determined that androgens impact CD8+ cell recruitment to immune foci, we sought to confirm that there was reduced inflammation in male mice. From the same set of mice, we evaluated the gene expression of key pro-inflammatory markers Cxcl9, Cxcl10, and Ifng in the liver at the time of challenge by gene-specific RT-PCR. Previous studies have shown that production of chemokines, CXCL9 and CXCL10, and the cytokine IFN-γ correlate with attracting and enhancing T cell responses to inflammatory foci33,53,54,55,56,57. We found significantly higher expression of Cxcl9, but not Cxcl10 and Ifng, in female and ORX male mice compared to male mice at the time of challenge (Fig. 7i). Taken together, this data demonstrates that the presence of androgens at time of challenge slightly reduces memory CD8+ T cell cytolytic capacity and cytokine production and reduces inflammation connected with Cxcl9 expression, linking to subsequent impaired recruitment of additional immune cells in male mice.
Discussion
Liver stage malaria vaccine research has predominantly been conducted in female mice. Through a series of P&T immunization and challenge studies in male and female mice, we identified sex-specific differences in protection outcomes, CD8+ T cell density, and inflammation in the liver. We further show that although androgens modulate protection outcomes, the quantity and quality of vaccine-induced CD8+ T cells did not correlate with sex hormone-mediated protection. Instead, through a step-wise assessment of modified vaccination regimens to overcome reduced protection, we determined that androgens at the time of challenge impede an otherwise protective adaptive immune response. Ultimately, we attribute reduced protection in vaccinated males to androgen-impaired recruitment of CD8+ T cells in male liver tissues via a restricted inflammatory response.
Sex hormones, physiology, and sex chromosomes are factors potentially contributing to sex-specific malaria liver stage vaccine outcomes. These three factors have important implications for the production, function, and recall responses of memory CD8+ T cells in the liver. First, with respect to sex hormones, it is well established that sex hormones influence many immune system functions18,19,21. Both androgens and estrogens have been linked to altering inflammation, interferon signaling, and CD8+ T cell frequency and function58,59,60,61,62,63,64. One prior report also found that sex hormones alter protection outcomes in adult C3H/HeNCr MTV mice vaccinated with two doses of radiation-attenuated P. berghei expressing P. falciparum CSP26. They further identified testosterone as a key modulator of protection through a series of orchiectomies, ovariectomies, and add-back hormone studies26. Here, we found a similar trend in response to P&T vaccination – that androgens and not estrogens are the drivers behind differential protection to P&T vaccination in the P.yoelii/BALB/cJ model.
Theoretically, androgens could influence the efficacy of P&T vaccination during two potential phases: 1) the production phase of immune memory at the time of vaccination, or 2) the recall phase of immune memory at the time of challenge. Some of our data suggests that androgens may influence the production phase of vaccination, since by acutely and reversibly altering the hormone environment at the time of vaccination, we were able to increase protection in male mice. However, it is also possible that administering acyline at the time of vaccination may have altered protection outcomes by influencing long-lasting CD8+ T cell-extrinsic factors within the liver65. Indeed, our other evidence indicates that poor protection in intact male mice is not driven by the production phase of vaccination. First, increasing CD8+ T cell responses with more immunogenic priming or adjuvanted trapping methods did not significantly increase protection in males. Second, the quality and quantity of the vaccine-induced CD8+ T cell repertoire were not significantly different between ORX and SHAM male mice. This was surprising, as there is literature describing how the androgen receptor accelerates terminally exhausted CD8+ T cells via direct transcriptional activation of TCF1/Tcf743. We thus hypothesized that intact male mice would have more dysfunctional CD8+ T cells as shown by increased LAG-3+, PD-1+, and TCF1lo expression and/or reduced production of IFN-γ, TNF, and CD107a. Yet, neither direct ex vivo phenotyping nor in vitro stimulation under normal stimulation conditions could establish a connection between androgens and T cell dysfunction. We therefore concluded that androgens do not alter the production of vaccine-specific memory CD8+ T cells nor appear to alter their permanent program within a one-month time period, although we acknowledge from in vitro assays that we cannot exclude a potential impact of androgens on CD8+ T cell effector function in vivo.
The alternative explanation was that the immune effects of androgens at the recall phase were driving sex-bias in protection outcomes from P&T vaccination. Remarkably, when we reduced androgens levels after P&T vaccination but prior to spz challenge, high levels of protection in males were achieved, confirming this was the key driver of the sex-specific outcomes in our model. To effectively protect mice during this recall phase, Trm cells must first detect and migrate toward infected hepatocytes13. This involves inflammatory signaling, and under inflammatory conditions, the speed and motility of local CD8+ T cells has been shown to increase53,66. Thereafter, Trm cells employ cytolytic and cytokine-mediated effector mechanisms to eliminate infection4,33,67,68. Importantly, cytokines produced by Trm cells also activate and recruit more immune cells toward infected hepatocytes, an innate-like process often referred to as ‘sensing and alarm,38,50,51. Thus, having shown that effects of androgens at the recall phase were driving sex-bias in protection outcomes, we next sought to separately and more discretely probe cytolytic and cytokine effector function versus cell recruitment and propagated inflammation.
To mimic the direct impact of androgens on CD8+ T cell cytolytic and cytokine function, we repeated ex vivo stimulation in hormone-free conditions and exogenously added DHT into the media. There was a small but significant reduction in Granzyme B expression across all groups and IFN-γ in ORX male mice. This supports prior literature that had identified androgen receptor (AR) interaction with the open chromatin regions associated with Ifng and Gzmb in effector memory CD8+ T cells and regulates rapid production of IFN-γ and Granzyme B upon TCR stimulation44. However, in the same assay, we were unable to detect a difference in CD107a expression, a marker of degranulation, indicating that we were unable to connect Granzyme B expression to a functional readout of cytolytic capacity. Whether this is a technical issue or a true biological phenomenon remains to be determined. Additionally, previous work had identified a non-essential role of Granzyme B and other cytolytic mechanisms in mediating protection in female mice67. Thus, while were able to find evidence for direct effects of androgens suppressing CD8+ T cell effector functions, these effects were minor under conditions of acute antigen stimulation, differing from the increased function of CD8+ T cells observed during androgen deprivation with chronic antigen stimulation models43,44,45.
To assess how androgens influence the sense and alarm function of Trm cells, and their ability to recruit immune cells to the site of infection, we evaluated the spatial distribution of hepatic CD8+ cells during Plasmodium spz challenge. Previous work has identified that multiple CD8+ T cells per infected hepatocyte are important to clear infection in the P. yoelii/BALB/cJ model51. Here we confirmed that male mice have fewer inflammatory foci with CD8+ cells, and within these foci, there are fewer CD8+ cells compared to female and ORX male mice. This parallels previously observed testosterone-mediated suppression of T cell recruitment observed in the hepatic infection model of cholangitis69. As this assay marks CD8+ cells, it is important to note it captures both antigen-specific CD8+ T cells, but also nonspecific activated CD8+ cells attracted to foci. Given we were unable to determine a definitive direct effect of androgens on CD8+ T cell cytokine production in our other assays, this difference in foci formation is likely also a byproduct of other androgen-mediated immune cell functions. One possible explanation could be reduced MHC Class I presentation on infected hepatocytes in male mice. This is supported by a recent study that identified AR signaling represses MHC Class I expression in a tumor microenvironment as a mechanism in which hormones modulate escape from CD8+ T cell killing70.
A second explanation for reduced recruitment of CD8+ cell to immune foci in male mice could be CD8+ T cell extrinsic and related to general inflammation and other immune cells. Androgens are also known to have an immunosuppressive effect on inflammation and innate immune cells59,65,71. We show restricted expression of CXCL9, an IFN-γ inducible chemokine, in male mice following attenuated and virulent spz infection. During viral infections, CXCL9-CXCR3 interactions are important for clustering of antigen-specific effector CD8+ T cells around infected cells72. As androgens did not affect CXCR3 expression on antigen-specific and nonspecific CD8 T cells (Fig. S9h), future work should examine their impact on CXCL9-producing cells. Two potential culprits are hepatic neutrophil and monocyte populations, which are known to be affected by testosterone in the context of other hepatotropic infections73,74,75. Further investigations into how the sex-specific inflammatory milieu might alter innate and adaptive immune cell recruitment toward infected hepatocytes at the time of challenge will thus be a key focus of future work.
Biological sex also alters physiology. While physiological differences by sex were not a focus here, we also observed that the male liver was about 35% larger than that of female mice. Previous work found a threshold of number of effector CD8+ T cells and infected hepatocytes is needed to confer sterile protection32. While we did not identify a difference in parasite biomass between naïve male and female mice 44 h after spz challenge, vaccine-induced CD8+ T cell number after accounting for differences in liver mass revealed a higher density in females. Future research could thus also seek to explore whether physical size influences a CD8+ T cell threshold requirement in malaria liver stage vaccine efficacy in pre-clinical and clinical studies.
Equally, while sex chromosomes were not directly evaluated here, the X and Y chromosomes encode a wide array of immune response-linked proteins58. The presence of two X chromosomes (XX) and absence of the Y chromosome is clearly not a requirement for protection since we were able to protect male mice by altering the hormone environment. However, genes mapped to the X-chromosome exhibited sex-specific expression in response to P&T, such as Cxcr3 and Il2rg. CXCR3 is an important homing marker and is constitutively expressed on Trm cells during homeostasis76. IL-2Rγ transduces common γ-chain cytokine signals such IL-2, IL-7, and IL-15, which are important signals for the survival of memory CD8+ T cells77. These genes therefore also warrant further evaluation for sex biased regulation. However, since male mice were capable of being protected, we can conclude these sex-linked genes were not the major driver of differential protection to P&T vaccination.
Finally, a critical question is how these findings may relate to human vaccination and infection. A recent meta-analysis of the attenuated spz vaccine PfSPZ in humans identified higher levels of antigen-specific antibodies in females compared to males. However, no difference in PfSPZ vaccine efficacy based on sex was identified78. Although sex-specific immunogenicity is observed in humans and mice, the absence of sex specific protection outcomes in humans may be linked to inherent differences, such as the longer liver stage in humans79 and distinct endocrine systems80,81. Given the paucity of information about how biological sex interacts with the immune-parasite axis during Plasmodium liver stage infection in mice and humans, further evaluation of the basic biology of how males and females respond to attenuated and virulent Plasmodium parasites is warranted.
Taken together, our results indicate that there are sex-specific differences in protection outcomes and hepatic memory CD8+ T cell number relative to liver size. However, restricted CD8+ T cell recruitment toward infected hepatocytes via a reduced inflammatory response is the driving factor behind the inability to protect male mice. Although ovariectomies did not decrease protection, we could not assess if there was an increase in efficacy because intact female mice are already completely protected, leaving the potential role of ovarian hormones uncertain. Additionally, the gene expression data is limited by measuring the RNA levels and no protein level analysis was conducted. This work highlights why male mice serve as a more stringent model for malaria liver stage vaccine evaluation. While we were unable to overcome the natural androgen-mediated barrier of the male liver with alternative P&T strategies, a malaria liver stage vaccine that can elicit complete sterile immunity by overcoming the immunosuppressive liver microenvironment in males may be an indicator of a promising malaria vaccine. Beyond malaria liver stage vaccines, capturing sex differences in pre-clinical and clinical studies is a National Institute of Health mandate and an important step towards understanding intrinsic biases in vaccine development.
Methods
Ethics statement
Animal studies were performed according to the regulations of the institutional animal care and use committee. Approval was obtained from the University of Washington (UW) Institutional Animal Care and Use Committee (IACUC) under protocol 4317-01. The UW IACUC adheres to the NIH Office of Laboratory Animal Welfare standards (OLAW welfare assurance # D16-00292).
Mice
Male and female BALB/cJ and C57BL/6 J mice (4-6 weeks old) were obtained from Jackson Laboratories and housed at UW in an IACUC-approved animal facility. All mice were used under an approved IACUC protocol (4317-01 to SCM). Mice experience a 14h light and 10h dark cycle in a 72°F and 30–70% humidity room. Primary euthanasia was conducted with CO2 inhalation, followed by a secondary dissection method.
DNA vaccine preparation
The P. yoelii circumsporozoite protein (PyCSP) DNA vaccine plasmids were constructed in the pUb.3 vector with an N-terminal ubiquitin tag and purified as described16,82,83. For all vaccinations, Escherichia coli heat-labile toxin (LT)-encoding plasmid was used as an adjuvant in a 1:10 ratio with the PyCSP minigene or full-length vaccines84. The PyCSP-minigene plasmid encodes the codon-optimized SYVPSAEQI epitope and the PyCSP plasmid encodes the full-length codon-optimized CSP protein without the major repeat regions, (GPGAPQ)5 and (PPQQ)4. All plasmids were Sanger sequenced at Azenta Life Sciences before use. Gene gun DNA vaccine cartridges were constructed by loading DNA onto gold powder (Technic Inc., 12-507) and then coating loaded gold onto Tefzel tubing (McMaster-Carr, 5583K44) as previously described16,83. Abdominal fur was trimmed and mice were vaccinated on the abdomen using a PowderJect-style gene gun using two cartridges per day on days 0 and 2 (0.5 µg DNA per cartridge).
Plasmodium sporozoites for challenge and vaccination
Female Anopheles stephensi mosquitoes infected with wild-type (WT) P. yoelii (strain 17XNL), GFP/luciferase-expressing P. yoelii 17XNL (Py-Luc)29, or P. berghei (strain ANKA) were reared at Seattle Children’s Research Institute (Seattle, WA) or UW (Seattle, WA). Fresh spz were obtained by salivary gland dissection 14–18 days post-infection followed by Accudenz (Fisher Scientific, NC9951010) gradient purification85. All spz were diluted in Schneider’s insect media for administration (Thermo Fisher Scientific, 21720024). Figure legends specify the dose for each experiment. For Py-WT spz challenge, 1x103 freshly dissected Py-WT spz were injected unless stated otherwise. For Pb-WT spz challenge, 200 freshly dissected Pb-WT spz were injected. Blood stage protection after spz challenge was assessed by Giemsa (Millipore Sigma, 48900-500ML-F) stained thin blood smear microcopy on 3–14 days post-challenge. Mice were deemed protected if blood smears remained negative for parasites up to 14 days.
For spz vaccinations, radiation attenuation of freshly dissected P. yoelii or P. berghei spz was done by exposing wild-type spz (Py-WT or Pb-WT) to 10,000 rads via an X-ray source (Rad Source). Cryopreserved radiation-attenuated P. yoelii17XNL spz (10,000 rads by C0-60) were produced, purified, and vialed by Sanaria Inc. (Rockville, MD) as described9,29. All spz used for immunization or challenge studies were administered intravenously (retro-orbital) in a volume of 100 μL per mouse. For Py- and Pb-RAS immunizations, the standard dose was 2x104 spz.
Parasite burden quantification
Luciferase-based in vivo parasite imaging of the liver was conducted with an in vivo bioluminescent imager (Xenogen IVIS Spectrum, Caliper Life Sciences)86. Briefly, mice previously infected with 104 Py-Luc spz by intraperitoneal injection with 5 mg of firefly D-Luciferin (Goldbio, LUCK-1G) prior to undergoing isoflurane anesthesia. Bioluminescence was acquired and measured as previously reported using a one-minute acquisition time. IVIS images were quantitatively evaluated using Living Image 3.0 software (Perkin Elmer) with regions of interest (ROI) placed around the abdominal area overlying the liver. ROI measurements were recorded in total flux (p/sec/cm2/sr).
To quantify P. yoelii liver burden in BALB/cJ mice by reverse transcription polymerase chain reaction (RT-PCR), mice were sacrificed, half of the liver was excised and pulverized by bead beating in NucliSENS lysis buffer (bioMérieux, 280134). For P. berghei liver burden in C57BL/6 mice, whole liver was excised, gently mashed through a 100-mm cell strainer and washed with RPMI mixture, and a fraction was collected into NucliSENS lysis buffer. In both methods, nucleic acids were extracted as previously described using a bioMérieux EasyMAG (bioMérieux)87. RNA was subjected to RT-PCR using the SensiFAST Probe Lo-ROX Kit (Bioline; Thomas Scientific, C755H82) using a predesigned HEX-labelled mouse Gapdh RT-PCR assay (IDT Inc) multiplexed with a Pan-Plasmodium 18S rRNA assay (forward: AAAGTTA[ + A]GGGA[ + G][ + T]GAAGA; reverse: AA[ + G]ACTTTGATTTCTC[ + A]TAAGG; [+X] denotes a locked nucleic acid) as described32. Cycling conditions consisted of one cycle of 45 °C for 10 min, one cycle of 95 °C for 2 min, and 45 cycles of 95 °C for 5 s and 50 °C for 35 s on a QuantStudio5 real-time PCR machine (Thermo Fisher Scientific). Plasmodium 18S rRNA copy numbers per reaction were determined against a calibration curve of quantified Armored RNA encoding full-length Plasmodium 18S rRNA (Asuragen); data were also normalized to mouse GAPDH. Results were reported to a cycle threshold (CT) of 35 cycles. Mice were deemed completely protected if RT-PCR results were below the limit of detection (no cycle threshold detected or detected at >35 cycles).
Quantification of inflammatory genes
To quantify host inflammatory genes from mouse liver, nucleic acids were isolated using the previously outlined method above. Inflammatory genes were evaluated with commercially available PrimeTimeTM qPCR primers: Cxcl9 (forward: 5’-CAAATCCCTCAAAGACCTCAAAC-3’, reverse: 5’-GATCTCCGTTCTTCAGTGTAGC-3’), Cxcl10 (forward: 5’-ATTTTCTGCCTCATCCTGCT-3’, reverse: 5’-TGATTTCAAGCTTCCCTATGGC-3’), and Ifng (forward: 5’-CTGAGACAATGAACGCTACACA-3’, reverse: 5’-TCCACATCTATGCCACTTGAG-3’). Data was normalized to mouse Gapdh. First, we evaluated if the baseline gene expression pattern of Cxcl9, Cxcl10, and Ifng were differentially expressed between a mock-infected female, male, and ORX male mice for each gene. No difference was detected in gene expression across all three genes (Kruskal Wallis test: Cxcl9 (p = 0.175), Cxcl10 (p = 0.754), Ifng (p = 0.465)), so we pooled all the mock samples into a single control group for relative comparison. Values were then transformed relative to log2 values to compare challenged mice relative to the pooled control group.
Tissue processing for flow cytometry
Unperfused livers were collected into RPMI media (Fisher Scientific, 11-875-119) containing 5% fetal bovine serum (FBS; Millipore Sigma, F4135-500ML), and supplemented with glutamine (ThermoFisher Fisher Scientific, 35050061) (RPMI mixture). Each liver was gently mashed through a 100-mm cell strainer (BioPioneer, DGN258367) and washed with RPMI mixture. Cell suspensions were centrifuged at 80 x g for 1 min at 22 °C with no brake. Isolated supernatants were then centrifuged at 450 x g for 10 min to pellet cells. Cells were resuspended in 10 mL of 35% Percoll (GE Healthcare, Fisher Scientific, 45-001-753) in HBSS (ThermoFisher Scientific, 24020117) supplemented with 100 U heparin (Millipore Sigma, H3393) before centrifugation at 830 x g at 22 °C with no brake. The cell pellet was incubated in 3 mL RBC lysis solution (150 mM NH4Cl, 10 mM KHCO3; 0.1 mM EDTA Na2·2H2O, pH 7.4) for 3 min before being washed with 10 mL of RPMI mixture and spun at 500 x g at 4 °C for 8 min. Final pellets were resuspended in 200 µL of RPMI mixture and moved to 96-well plates for antibody staining. For hormone-free conditions observed in Fig. 6, RPMI mixture was made without phenol red (Fisher Scientific, 11-835-030) and used charcoal-stripped FBS (ThermoFisher Scientific, A3382101).
Splenic lymphocytes were isolated by teasing tissue through a 100-mm cell strainer, washed with RPMI mixture and centrifuged at 460 x g at 4 °C for 5 min. The cell pellet was incubated in 3 mL RBC lysis solution for 3 min before being washed with 10 mL of RPMI mixture and spun at 460 x g at 4 °C for 5 min. Cells were resuspended in 1 mL of RPMI mixture and counted. Cells were diluted, and one million splenocytes per sample were used for antibody staining. For ELISPOTS, splenocytes were cryopreserved in freezing media (FBS/10% DMSO) prior to thawing for ELISPOT procedures, except in the case of the C57BL/6 J mice where splenocytes were not previously frozen.
Antibody staining and flow cytometry
Antibody panels used for staining were optimized to the BALB/cJ mouse model and are provided in Supplementary Table 2. For experiments corresponding to intracellular cytokine staining (ICS) in Figs. 2 and 4, lymphocytes were incubated at 37 °C with PyCSP peptide SYVPSAEQI (10 μM final) (Genemed Synthesis) and CD107a antibody (BD Biosciences, 558661) for 18 hours. Brefeldin A was added (Millipore Sigma, B7651-5MG), and cells were incubated for an additional 4 hours. Cells were washed, stained for live/dead designation, and resuspended in Fc-block (Fisher Scientific, BDB553141) for 15 min. Cells were washed and stained for cell surface markers and the intracellular markers IFN-γ (Biolegend, 505830), TNF (BD Biosciences, 554418), and Granzyme B (BD Biosciences, 560212) using the BD Cytofix/Cytoperm reagent (Fisher Scientific, BDB554722) according to manufacturer’s instructions. For experiments corresponding to Figs. 2 and 4, following liver lymphocyte isolation, cells were immediately stained for live/dead designation and resuspended in Fc-block. Lymphocytes were incubated with H-2Kd-PyCSP280-288-specific tetramer for 30 min at 4 °C prior to staining with surface antibodies and then intranuclear staining. Samples underwent intranuclear staining for marker TCF1 using the Foxp3/Transcription Factor buffer set (Thermo Fisher, 00-5523-00) for 30 min at 4 °C. For experiments corresponding to Fig. 6, lymphocytes were incubated at 37 °C with either 500 nM of dihydrotestosterone (DHT solution, Millipore Sigma, D-073) (or methanol vehicle control) for 18 hours. Cells were then stimulated with PyCSP peptide (or DMSO vehicle control). Brefeldin A was added after 2 hours and cells were incubated for an additional 4 hours. ICS was then performed after stimulation as outlined above. Flow cytometry data were acquired on a BD LSR Fortessa (BD Biosciences) equipped with a high-throughput sampler and configured with blue (488 nm), green (532 nm), red (628 nm), violet (405 nm), and ultraviolet (355 nm) lasers using standardized good clinical laboratory practice procedures to minimize variability.
Flow cytometry data analysis
Initial compensation, gating, and quality assessment of flow cytometry data were performed using FlowJo v.10 (TreeStar). Representative gating trees for ICS panel and intranuclear panel are shown in Supplementary Figs. 3 and 4. The surface marker and ICS flow cytometry data were then processed using the OpenCyto framework in the R programming environment88.
Ex vivo IFN-γ ELISPOT
PyCSP peptide (SYVPSAEQI) (Genemed Synthesis) was reconstituted in DMSO. Mouse IFN-γ ELISPOT was conducted by stimulating 5×105 splenocytes with PyCSP peptide (or DMSO vehicle control) at 1 µg/ml for 18 hr at 37 °C followed by development following manufacturer guidelines as reported previously89. The number of spot-forming units (SFU) in each well was calculated using an ImmunoSpot 5.1 Analyzer (Cellular Technology Limited). SFU were normalized to DMSO control wells and reported as SFU per million splenocytes.
Immunohistochemistry of inflammatory foci and CD8+ cells
Following infection with either P. yoelii or mock injection with loading media (Schneiders), livers from mice were harvested at 38–44 h post infection (hpi) and left lateral lobe was fixed 24 h in 10% neutral-buffered formalin (MilliporeSigma, HT501128-4L), paraffin-embedded and serially sectioned at 4 μm. Paraffin sections were stained using a BOND-RX automatic IHC system (Leica, Buffalo Grove, IL, USA) using heat-mediated EDTA antigen retrieval pH 8 (100 °C for 10 and 20 min). Sections were double stained with anti-CD8a (Rat monoclonal IgG2a eBioscience,14-0808-02; 1:1000) and anti-CD11c (Rabbit monoclonal, Cell Signaling, 97585; 1:500). Leica Bond Mixed Refine reagents (HRP and 3,3’-diaminobenzidene-based) and Leica Bond Red Refine reagents (AP and Fast Red-based) were used for detection and slides were counterstained with hematoxylin. Isotype-matched IgG antibodies were included as negative controls.
The CD8+ positive stained-cells are illustrated by red staining. One liver lobe section was analyzed per mouse and all inflammatory foci within a lobe were enumerated on scanned whole slide digital images at 20X on a NanoZoomer Digital Pathology (Hamamatsu Photonics). Inflammatory foci were defined as regions of aggregated cells and nuclei higher than the background density of nuclei and containing at least one CD8+ positive-stained cell. Foci in question on IHC slides were confirmed as inflammatory foci by comparing scanned whole slide images of IHC slides to corresponding inflammatory foci on hematoxylin and eosin-stained glass slides of the same liver lobe section for a subset of the samples. Once identified, a perimeter was drawn around the foci that was either applied with the circle tool or for more irregular shapes, was drawn freehand using with the area being calculated by the NDP.view2 software (Hamamatsu Photonics). Once the perimeter was established, a reviewer counted the number of CD8+ positive stained-cells within each focus, only considering markers with clear CD8+ positive membranous/intracellular staining. Tissue spots that were missing, contained staining artifacts, damaged or had unclear staining were excluded from analysis. Of note, slides were also stained with CD11c, however, due to unclear staining, was not used for quantitative evaluation. Analysis was performed by one investigator in a blinded manner with help and guidance from a board-certified veterinary pathologist, Dr. Klug.
Orchiectomy and ovariectomy surgeries
For orchiectomy (ORX), male mice were anesthetized by isoflurane inhalation (Isoflurane, USP, Dechra Veterinary Products), with doses of 4-5% for induction and 2-3% for maintenance. A 5 mg/kg dose of meloxicam (Pivetal Alloxate, Aspen Veterinary Resources, LTD; Patterson Veterinary, 07-893-7565) was administered subcutaneously (SC) to each mouse for systemic analgesia. Lubricant ophthalmic ointment (Pivetal Artificial Tears, Aspen Veterinary Resources, LTD; Patterson Veterinary, 07-894-6156) was applied to both eyes to prevent corneal desiccation. Thermal support was provided using a warm water circulating heating pad. The intended incision site was surgically prepped with three alternating scrubs of 10% povidone-iodine (Medline, MDS093902) and 70% ethanol. A small, midline, scrotal incision was made with tissue scissors. Each testicle was identified singly, and the associated spermatic cord was clamped with hemostats for at least 10 seconds, and the testicle removed. The incision was closed using surgical tissue adhesive (3 M Vetbond Tissue Adhesive 1469, 3 M; Patterson Veterinary, 07-805-5031). Sham ORX surgeries were performed using the same surgical protocol without surgical removal of the testicles. All ORX and SHAM surgeries were performed at least 14 days prior to experimental treatment.
For ovariectomies (OVX), female mice were anesthetized by isoflurane inhalation with doses of 4-5% for induction and 2-3% for maintenance. A 5 mg/kg dose of meloxicam and 0.1 mg/kg dose of buprenorphine (generic Buprenorphine Hydrochloride for Injection, Par Sterile Products; Patterson Veterinary, 07-892-5235) were administered subcutaneously to each mouse for systemic analgesia. A 0.02 ml dose of 0.5% lidocaine/0.25% bupivacaine mixture (VEDCO Lidocaine Hydrochloride 2%, Sparhawk Laboratories, Patterson Veterinary 07-892-4324; 0.25% Bupivacaine Hydrochloride Injection, USP, HF Acquisition Co., LLC, Medline 0409-1160-01) was also administered SC at each of the intended bilateral flank incision sites prior to incision for local anesthesia. The intended incision sites were surgically prepped with three alternating scrubs of 10% povidone-iodine and 70% ethanol. The first flank incision was made on one side of the mouse using iris tissue scissors, through the skin and abdominal muscle layers. The ovary was identified within the ovarian fat pad, exteriorized out of the skin incision, and a suture ligature was tied immediately proximal to the ovary using 4-0 or 5-0 absorbable suture. The ovary was removed by incising through the oviduct immediately distal to the suture ligature. The abdominal wall was then closed using 4-0 or 5-0 absorbable suture. The skin incision was closed using surgical tissue adhesive (Vetbond Tissue Adhesive 1469, 3 M). This process was repeated on the contralateral flank to remove the second ovary. Sham OVX surgeries were performed using the same surgical protocol without surgical removal of the ovaries. All OVX and SHAM surgeries were performed at least 14 days prior to experimental treatment.
Administration of acyline and testosterone
Acyline (lot # RDZ001) was provided by John K. Amory, MD, MPH, at the University of Washington. The lyophilized vials were stored at -20 °C until resuspension. At the time of use, acyline was resuspend to a concentration of 2 mg/mL with sterile Molecular Biology Grade Water (Fisher Scientific, SH31191LS). Resuspended vials were stored for a maximum of 1 week at 4 °C. Unless stated otherwise, mice were subcutaneously injected 5 and 4 days prior to treatment (gene gun prime, RAS trap, or WT spz challenge) to give time for the hormone environment to equilibrate. For testosterone treatment, Testosterone propionate (Millipore Sigma, T1875-5G) was dissolved in sesame oil (Millipore Sigma, S3547-250ML) at a concentration of 20 µg/µL. Mice were weighed to determine average weight per group, and then subcutaneously injected with 100 µg testosterone/g body weight at 3 timepoints (3- and 1-day pre-challenge and 1-day post-challenge), unless stated otherwise.
Testosterone quantification in serum
Serum was collected via chin bleed and stored in -20 °C until testing. To measure testosterone levels in serum, Testosterone ELISA kits (Crystal Chem) were used per manufacturer guidelines. Single absorbance values of each sample were measured with CLARIOstar Plus Microplate Reader (BMG Labtech) and analyzed using MARS data analysis interface. A calibration curve was generated using manufacturer-provided standards (0,0.1, 0.4, 1.5, 6.0, and 25 ng/mL), and a four parametric logistic (4-PL) curve fit was used to calculate sample values.
Immunization with CSP peptide-pulsed DCs
Male BALB/cJ bone marrow-derived DCs were cultured and prepared for immunization as described89. On day 9 of culture, media was supplemented with lipopolysaccharide (0.1 µg/mL; Millipore Sigma, L2654-1MG) and PyCSP peptide (1 µg/mL)89. The next day, cells were washed in PBS and counted, and then 1 x 106 cells were injected intradermally (ID) into mice. ID immunizations were administered as 100 µL per mouse split across two ID injections in the lower back near the base of the tail.
Glycolipid adjuvant preparation
Good manufacturing practice (GMP) grade glycolipid adjuvant 7DW8-5 powder was provided by Moriya Tsuji (Columbia U) and been previously applied to increase protection of spz-based liver stage vaccines90. 7DW8-5 was dissolved in DMSO (Millipore Sigma, D2650-100ML), aliquoted, and stored at -20 °C. Single use aliquots were thawed at 56 °C for 10 min and then sonicated in an ultrasonic water bath for five min to break micelle formation. The 7DW8-5 was mixed with diluted Py-RAS immediately before administration to mice. All mice received 2 µg of 7DW8-5 adjuvant.
Gene expression analysis
For RNA sequencing, the liver was perfused with perfusion buffer (PBS with 2 mM EDTA). Following perfusion, the left lateral lobe was sectioned and preserved in RNAlater (Millipore Sigma, R0901-500ML). Total RNA was extracted from the tissue and mRNA library was prepared with TruSeq stranded mRNA (Illumina). A sequencing depth of 40 M total reads was performed on a NovaSeq6000 S4 150PE flowcell. RNA extraction and Illumina sequencing was performed by Psomagen. RNA sequencing libraries were aligned with STAR aligner and quality filtered by median coefficient of variation coverage <0.75, duplicate mapped reads >0.75, total aligned sequences ≤106. No samples were omitted after this process. Counts were normalized for RNA composition using the trimmed mean of M-values normalization method, filtered to protein-coding genes with ≥0.1 count per million in at least three samples, and converted to log2 count per million with quality weights using voom. This resulted in 15,120 genes for analysis. Differential expression was determined separately for each time point using the pairwise contrast model of: ~0 + group. Differentially expressed genes (DEGs) were defined as genes with a false-discovery rate (FDR) < 0.05 and |logFC|> 1.5. Hypergeometric enrichment against Broad MSigDBHallmark (H) and canonical pathway (C2) gene sets40,91 was completed for significant genes, and here significant genes were defined at FDR less than 0.05. Gene set enrichment analysis (GSEA) was performed again H and C5 gene sets with contrast fold changes using fast GSEA.
Pathway score calculation
To identify the effect of the vaccine on the pathway response in female and male mice, we conducted a linear regression analysis separately for each time point (acute and adaptive). We took the normalized data and subsetted the data to the genes found in the MSigDB 2022.1.Mm mouse gene sets, which are named in the figures. In order to use the lm function in R to perform the linear regression analysis, we first normalized the data to ensure that the geometric mean for each gene was 1. Next, we computed the geometric mean weight for each sample across gene sets and used these values as weights in the linear regression analysis. For each mouse, we then calculated the pathway gene score by taking a sum of gene expression across respective response gene sets. Next, we evaluated the significance of the interaction term between sex and vaccination status. Confidence intervals for sex-specific differential expression were computed from the same linear regression. R 4.2.2 was used for all calculations.
Statistics
Details of the statistical tests applied to datasets shown in the figures can be found in the above methods and in the corresponding figure legends. All data points and n values reflect biological replicates. Data analyzed included outliers unless these could be explained by technical error. Data collection and analysis were not performed blind to the conditions of the experiments. Data were assumed to be not normal, thus non-parametric test were employed. Where relevant, statistical tests were two-sided and unpaired, unless stated otherwise. Individual p-values are provided in the figures. Statistical analyses were performed in R (version 3.5.1) or GraphPad Prism 9.1.2 Software. Plots used in this manuscript were generated with GraphPad Prism and the following R packages: ggplot2, ggpubr, and ComplexHeatmap.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The accession number for the bulk RNA-seq data reported in this paper is found at GEO accession number GSE253755. All other data are available in the article and its Supplementary files. Any additional requests should be directed to the corresponding author Source data are provided with this paper.
Code availability
R codes are available at [https://github.com/carduncombe/Duncombe-et-al.-Nature-Communications.-2025..git].
References
World Malaria Report 2024. (World Health Organization, Geneva, 2024).
Sinnis, P. & Zavala, F. The skin: where malaria infection and the host immune response begin. Semin Immunopathol. 34, 787–792 (2012).
Cowman, A. F., Healer, J., Marapana, D. & Marsh, K. Malaria: biology and disease. Cell 167, 610–624 (2016).
Schmidt, N. W., Butler, N. S., Badovinac, V. P. & Harty, J. T. Extreme CD8 T cell requirements for anti-malarial liver-stage immunity following immunization with radiation attenuated sporozoites. PLoS Pathog. 6, e1000998 (2010).
Scheller, L. F. & Azad, A. F. Maintenance of protective immunity against malaria by persistent hepatic parasites derived from irradiated sporozoites. Proc. Natl Acad. Sci. USA 92, 4066–4068 (1995).
Ishizuka, A. S. et al. Protection against malaria at 1 year and immune correlates following PfSPZ vaccination. Nat. Med. 22, 614–623 (2016).
Nussenzweig, R. S., Vanderberg, J., Most, H. & Orton, C. Protective immunity produced by the injection of x-irradiated sporozoites of Plasmodium berghei. Nature 216, 160–162 (1967).
El-Moamly, A. A. & El-Sweify, M. A. Malaria vaccines: the 60-year journey of hope and final success-lessons learned and future prospects. Trop. Med. Health 51, 29 (2023).
Schofield, L. et al. Gamma interferon, CD8+ T cells and antibodies required for immunity to malaria sporozoites. Nature 330, 664–666 (1987).
Weiss, W. R., Sedegah, M., Beaudoin, R. L., Miller, L. H. & Good, M. F. CD8+ T cells (cytotoxic/suppressors) are required for protection in mice immunized with malaria sporozoites. Proc. Natl Acad. Sci. USA 85, 573–576 (1988).
Weiss, W. R. & Jiang, C. G. Protective CD8+ T lymphocytes in primates immunized with malaria sporozoites. PLoS One 7, e31247 (2012).
Seder, R. A. et al. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 341, 1359–1365 (2013).
Fernandez-Ruiz, D. et al. Liver-resident memory CD8(+) T cells form a front-line defense against malaria liver-stage infection. Immunity 45, 889–902 (2016).
Tse, S. W., Radtke, A. J. & Zavala, F. Induction and maintenance of protective CD8+ T cells against malaria liver stages: implications for vaccine development. Mem. Inst. Oswaldo Cruz 106, 172–178 (2011).
Holz, L. E. et al. CD8(+) T cell activation leads to constitutive formation of liver tissue-resident memory T cells that seed a large and flexible niche in the liver. Cell Rep. 25, 68–79 e64 (2018).
Olsen, T. M., Stone, B. C., Chuenchob, V. & Murphy, S. C. Prime-and-trap malaria vaccination to generate protective CD8(+) liver-resident memory T cells. J. Immunol. 201, 1984–1993 (2018).
Valencia-Hernandez, A. M. et al. A Natural peptide antigen within the Plasmodiumribosomal protein RPL6 confers liver T(RM) cell-mediated immunity against malaria in mice. Cell Host Microbe 27, 950–962.e957 (2020).
Flanagan, K. L., Fink, A. L., Plebanski, M. & Klein, S. L. Sex and gender differences in the outcomes of vaccination over the life course. Annu Rev. Cell Dev. Biol. 33, 577–599 (2017).
Aaby, P. et al. The non-specific and sex-differential effects of vaccines. Nat. Rev. Immunol. 20, 464–470 (2020).
Khramtsova, E. A., Davis, L. K. & Stranger, B. E. The role of sex in the genomics of human complex traits. Nat. Rev. Genet. 20, 173–190 (2019).
Klein, S. L. & Flanagan, K. L. Sex differences in immune responses. Nat. Rev. Immunol. 16, 626–638 (2016).
Oliva, M. et al. The impact of sex on gene expression across human tissues. Science 369, eaba3066 (2020).
Yang, X. et al. Tissue-specific expression and regulation of sexually dimorphic genes in mice. Genome Res. 16, 995–1004 (2006).
Kur, P., Kolasa-Wolosiuk, A., Misiakiewicz-Has, K. & Wiszniewska, B. Sex hormone-dependent physiology and diseases of liver. Int. J. Environ. Res. Public Health 17, 2620 (2020)
Nganou-Makamdop, K. & Sauerwein, R. W. Liver or blood-stage arrest during malaria sporozoite immunization: the later the better?. Trends Parasitol. 29, 304–310 (2013).
Vom Steeg, L. G., Flores-Garcia, Y., Zavala, F. & Klein, S. L. Irradiated sporozoite vaccination induces sex-specific immune responses and protection against malaria in mice. Vaccine 37, 4468–4476 (2019).
Cernetich, A. et al. Involvement of gonadal steroids and gamma interferon in sex differences in response to blood-stage malaria infection. Infect. Immun. 74, 3190–3203 (2006).
Legorreta-Herrera, M. et al. Sex hormones modulate the immune response to Plasmodium berghei ANKA in CBA/Ca mice. Parasitol. Res. 114, 2659–2669 (2015).
Miller, J. L. et al. Quantitative bioluminescent imaging of pre-erythrocytic malaria parasite infection using luciferase-expressing Plasmodium yoelii. PLoS One 8, e60820 (2013).
Coelho-Dos-Reis, J. G., Li, X. & Tsuji, M. Development of a novel mechanism-based glycolipid adjuvant for vaccination. F1000Res 7, 676 (2018).
Hafalla, J. C. et al. Identification of targets of CD8(+) T cell responses to malaria liver stages by genome-wide epitope profiling. PLoS Pathog. 9, e1003303 (2013).
Spencer, A. J. et al. The threshold of protection from liver-stage malaria relies on a fine balance between the number of infected hepatocytes and effector CD8(+) T cells present in the liver. J. Immunol. 198, 2006–2016 (2017).
Butler, N. S., Schmidt, N. W. & Harty, J. T. Differential effector pathways regulate memory CD8 T cell immunity against Plasmodium berghei versus P. yoelii sporozoites. J. Immunol. 184, 2528–2538 (2010).
Cooney, L. A. et al. Short-lived effector CD8 T cells induced by genetically attenuated malaria parasite vaccination express CD11c. Infect. Immun. 81, 4171–4181 (2013).
Minkah, N. K. et al. Innate immunity limits protective adaptive immune responses against pre-erythrocytic malaria parasites. Nat. Commun. 10, 3950 (2019).
Waxman, D. J. & Holloway, M. G. Sex differences in the expression of hepatic drug metabolizing enzymes. Mol. Pharm. 76, 215–228 (2009).
Liehl, P. et al. Host-cell sensors for Plasmodium activate innate immunity against liver-stage infection. Nat. Med 20, 47–53 (2014).
Lefebvre, M. N. et al. Expeditious recruitment of circulating memory CD8 T cells to the liver facilitates control of malaria. Cell Rep. 37, 109956 (2021).
Miller, J. L., Sack, B. K., Baldwin, M., Vaughan, A. M. & Kappe, S. H. I. Interferon-mediated innate immune responses against malaria parasite liver stages. Cell Rep. 7, 436–447 (2014).
Liberzon, A. et al. The molecular signatures database (MSigDB) hallmark gene set collection. Cell Syst. 1, 417–425 (2015).
Ashburner, M. et al. Gene ontology: tool for the unification of biology. The gene ontology consortium. Nat. Genet. 25, 25–29 (2000).
Hall, K. & Korenchevsky, V. Liver Changes in male rats after castration and injection of sex hormones. Br. Med. J. 1, 438–441 (1938).
Yang, C. et al. Androgen receptor-mediated CD8(+) T cell stemness programs drive sex differences in antitumor immunity. Immunity 55, 1268–1283.e1269 (2022).
Guan, X. et al. Androgen receptor activity in T cells limits checkpoint blockade efficacy. Nature 606, 791–796 (2022).
Kwon, H. et al. Androgen conspires with the CD8(+) T cell exhaustion program and contributes to sex bias in cancer. Sci. Immunol. 7, eabq2630 (2022).
Wherry, E. J. & Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 15, 486–499 (2015).
Sanz, E. et al. RiboTag analysis of actively translated mRNAs in Sertoli and Leydig cells in vivo. PLoS One 8, e66179 (2013).
Herbst, K. L. et al. A single dose of the potent gonadotropin-releasing hormone antagonist acyline suppresses gonadotropins and testosterone for 2 weeks in healthy young men. J. Clin. Endocrinol. Metab. 89, 5959–5965 (2004).
Herbst, K. L., Anawalt, B. D., Amory, J. K. & Bremner, W. J. Acyline: the first study in humans of a potent, new gonadotropin-releasing hormone antagonist. J. Clin. Endocrinol. Metab. 87, 3215–3220 (2002).
Schenkel, J. M., Fraser, K. A., Vezys, V. & Masopust, D. Sensing and alarm function of resident memory CD8(+) T cells. Nat. Immunol. 14, 509–513 (2013).
Cockburn, I. A. et al. In vivo imaging of CD8+ T cell-mediated elimination of malaria liver stages. Proc. Natl Acad. Sci. USA 110, 9090–9095 (2013).
Hoffman, S. L. et al. Sporozoite vaccine induces genetically restricted T cell elimination of malaria from hepatocytes. Science 244, 1078–1081 (1989).
Harris, T. H. et al. Generalized Levy walks and the role of chemokines in migration of effector CD8+ T cells. Nature 486, 545–548 (2012).
Kastenmuller, W. et al. Peripheral prepositioning and local CXCL9 chemokine-mediated guidance orchestrate rapid memory CD8+ T cell responses in the lymph node. Immunity 38, 502–513 (2013).
Nakanishi, Y., Lu, B., Gerard, C. & Iwasaki, A. CD8(+) T lymphocyte mobilization to virus-infected tissue requires CD4(+) T-cell help. Nature 462, 510–513 (2009).
Rahimi, R. A. & Luster, A. D. Chemokines: critical regulators of memory T cell development, maintenance, and function. Adv. Immunol. 138, 71–98 (2018).
Griffith, J. W., Sokol, C. L. & Luster, A. D. Chemokines and chemokine receptors: positioning cells for host defense and immunity. Annu Rev. Immunol. 32, 659–702 (2014).
Fish, E. N. The X-files in immunity: sex-based differences predispose immune responses. Nat. Rev. Immunol. 8, 737–744 (2008).
Shepherd, R., Cheung, A. S., Pang, K., Saffery, R. & Novakovic, B. Sexual dimorphism in innate immunity: the role of sex hormones and epigenetics. Front Immunol. 11, 604000 (2020).
Jacobsen, H. & Klein, S. L. Sex differences in immunity to viral infections. Front Immunol. 12, 720952 (2021).
Brown, M. A. & Su, M. A. An inconvenient variable: Sex hormones and their impact on T cell responses. J. Immunol. 202, 1927–1933 (2019).
Henze, L., Schwinge, D. & Schramm, C. The effects of androgens on T cells: Clues to female predominance in autoimmune liver diseases?. Front Immunol. 11, 1567 (2020).
Pujantell, M. & Altfeld, M. Consequences of sex differences in type I IFN responses for the regulation of antiviral immunity. Front Immunol. 13, 986840 (2022).
Cervantes-Candelas, L. A. et al. 17beta-estradiol is involved in the sexual dimorphism of the immune response to malaria. Front Endocrinol. (Lausanne) 12, 643851 (2021).
Gubbels Bupp, M. R. & Jorgensen, T. N. Androgen-induced immunosuppression. Front Immunol. 9, 794 (2018).
Overstreet, M. G. et al. Inflammation-induced interstitial migration of effector CD4(+) T cells is dependent on integrin alphaV. Nat. Immunol. 14, 949–958 (2013).
Doolan, D. L. & Hoffman, S. L. The complexity of protective immunity against liver-stage malaria. J. Immunol. 165, 1453–1462 (2000).
Schenkel, J. M. & Masopust, D. Tissue-resident memory T cells. Immunity 41, 886–897 (2014).
Schwinge, D. et al. Testosterone suppresses hepatic inflammation by the downregulation of IL-17, CXCL-9, and CXCL-10 in a mouse model of experimental acute cholangitis. J. Immunol. 194, 2522–2530 (2015).
Chesner, L. N. et al. Androgen receptor inhibition increases MHC Class I expression and improves immune response in prostate cancer. Cancer Discov. 15, 481–494 (2024).
Kasarinaite, A., Sinton, M., Saunders, P. T. K. & Hay, D. C. The influence of sex hormones in liver function and disease. Cells 12, 1604 (2023).
Sung, J. H. et al. Chemokine guidance of central memory T cells is critical for antiviral recall responses in lymph nodes. Cell 150, 1249–1263 (2012).
Er-Lukowiak, M. et al. Testosterone affects type I/type II interferon response of neutrophils during hepatic amebiasis. Front Immunol. 14, 1279245 (2023).
Bizzaro, D. et al. Sex-dependent differences in inflammatory responses during liver regeneration in a murine model of acute liver injury. Clin. Sci. (Lond.) 132, 255–272 (2018).
Hoffmann, J. P., Liu, J. A., Seddu, K. & Klein, S. L. Sex hormone signaling and regulation of immune function. Immunity 56, 2472–2491 (2023).
Masopust, D. & Soerens, A. G. Tissue-resident T cells and other resident leukocytes. Annu Rev. Immunol. 37, 521–546 (2019).
Chin, S. S. et al. T cell receptor and IL-2 signaling strength control memory CD8(+) T cell functional fitness via chromatin remodeling. Nat. Commun. 13, 2240 (2022).
Kc, N. et al. Increased levels of anti-PfCSP antibodies in post-pubertal females versus males immunized with PfSPZ Vaccine does not translate into increased protective efficacy. Front Immunol. 13, 1006716 (2022).
De Niz, M. & Heussler, V. T. Rodent malaria models: insights into human disease and parasite biology. Curr. Opin. Microbiol. 46, 93–101 (2018).
Staley, K. & Scharfman, H. A woman’s prerogative. Nat. Neurosci. 8, 697–699 (2005).
Janne, M., Deol, H. K., Power, S. G., Yee, S. P. & Hammond, G. L. Human sex hormone-binding globulin gene expression in transgenic mice. Mol. Endocrinol. 12, 123–136 (1998).
Imai, J., Otani, M., Sakai, T. & Hatta, S. Purification of the subcellular compartment in which exogenous antigens undergo endoplasmic reticulum-associated degradation from dendritic cells. Heliyon 2, e00151 (2016).
Stone, B. C. et al. Complex minigene library vaccination for discovery of pre-erythrocytic Plasmodium T cell antigens. PLoS One 11, e0153449 (2016).
Arrington, J. et al. Plasmid vectors encoding cholera toxin or the heat-labile enterotoxin from Escherichia coli are strong adjuvants for DNA vaccines. J. Virol. 76, 4536–4546 (2002).
Kennedy, M. et al. A rapid and scalable density gradient purification method for Plasmodium sporozoites. Malar. J. 11, 421 (2012).
Billman, Z. P., Seilie, A. M. & Murphy, S. C. Purification of Plasmodium sporozoites enhances parasite-specific CD8+ T cell responses. Infect. Immun. 84, 2233–2242 (2016).
Murphy, S. C. et al. Real-time quantitative reverse transcription PCR for monitoring of blood-stage Plasmodium falciparum infections in malaria human challenge trials. Am. J. Trop. Med Hyg. 86, 383–394 (2012).
Finak, G. et al. OpenCyto: an open source infrastructure for scalable, robust, reproducible, and automated, end-to-end flow cytometry data analysis. PLoS Comput Biol. 10, e1003806 (2014).
Murphy, S. C., Kas, A., Stone, B. C. & Bevan, M. J. A T-cell response to a liver-stage Plasmodium antigen is not boosted by repeated sporozoite immunizations. Proc. Natl Acad. Sci. USA 110, 6055–6060 (2013).
Watson, F. N. et al. Ultra-low volume intradermal administration of radiation-attenuated sporozoites with the glycolipid adjuvant 7DW8-5 completely protects mice against malaria. Sci. Rep. 14, 2881 (2024).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Acknowledgements
We thank Tess Seltzer, Veronica Primavera, Cecilia Kalthoff, and Alexis Kaushansky (Seattle Children’s Research Institute) and staff at the University of Washington insectary for assistance and support of P. yoelii–infected mosquito production and the NIH Tetramer Core Facility (contract number 75N93020D00005) for providing PyCSP monomers. We thank the veterinary and husbandry staff of the UW Department of Comparative Medicine. We thank Paul Edlefsen, Ewelina Kosmider, and Danica Shao (Fred Hutch) for statistical support and Moriya Tsuji for the 7DW8-5 adjuvant. We thank Dr. Jenna Klug (University of Washington), Nishi Ivanov, and Brendy Fountaine for pathology and immunohistochemistry support. This research was supported by NIH grant 1R01AI141857 to S.C.M. and National Science Foundation fellowship 1000341692 to C.J.D.
Author information
Authors and Affiliations
Contributions
Designed research: C.J.D., N.S., F.N.W., with guidance from E.D.L., C.S., M.J.S., S.C.M. Performed the experiments: C.J.D., N.S., A.S.P., F.N.W., K.B., E.N.C., M.J.S., A.C.K., A.M.S. Produced DNA gene gun vaccines: A.C.K. Analyzed data: C.J.D., N.S., K.A.D., E.D.L. Wrote article: C.J.D., S.C.M. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
C.J.D., N.S., F.N.W., A.S.P., E.D.L., K.B., E.N.C, A.C.K., A.M.S, C.S. and M.J.S. declare no competing interests. S.C.M. filed a patent application on selected aspects of the prime-and-trap concept through the University of Washington. S.C.M. has equity in a startup company (Sound Vaccines, Inc.) that is negotiating with the University of Washington for rights to this intellectual property. The relationship between the authors and Sound Vaccines, Inc., has been reviewed by the University of Washington and complies with all University and State of Washington policies on such activities. K.A.D. is a paid contractor for Seattle Biosoftware, which had no input or association with this study.
Peer review
Peer review information
Nature Communications thanks Joseph Hoffmann and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Duncombe, C.J., Sen, N., Watson, F.N. et al. Androgens inhibit protective CD8+ T cell responses against pre-erythrocytic malaria parasites in mice. Nat Commun 16, 5172 (2025). https://doi.org/10.1038/s41467-025-60193-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-60193-7
This article is cited by
-
Palmitoylation of TBK1 enhances the type I interferon signaling and strengthens anti-malarial immunity in mice
Nature Communications (2025)
