
Abstract
Hypertrophic scars are a stubborn form of dermal fibrosis that impairs quality of life. Although 5-ALA-mediated photodynamic therapy holds promise, its efficacy is undermined by poor transdermal delivery and rapid metabolism into non-photosensitive heme. Here, we introduce a “zero-waste” strategy that repurposes 5-ALA-derived heme to synergistically amplify ferroptosis. This is achieved by co-encapsulating 5-ALA and baicalin within human H-ferritin, subsequently incorporated into polyvinylpyrrolidone microneedles. The resulting system enables targeted delivery to hypertrophic scar fibroblasts with pH-responsive, programmable drug release. Upon administration, 5-ALA generates protoporphyrin IX to initiate photodynamic therapy. Baicalin is then released to induce ferroptosis and synergize with the reactive oxygen species and heme accumulated during photodynamic therapy, thereby overstimulating the HO-1–heme metabolic axis. This cascade promotes the release of Fe²⁺ and CO, further amplifying ferroptotic responses. Moreover, the ferroptotic stress triggers mitophagy and mitochondrial Fe²⁺ efflux. By harnessing 5-ALA metabolic byproducts, this strategy achieved markedly prolonged anti-scar efficacy in the female rabbit ear HS tissues, surpassing that of conventional therapies.
Similar content being viewed by others
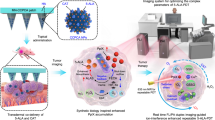


Introduction
Hypertrophic scar (HS), characterized by excessive proliferation of fibroblasts and over-deposition of extracellular matrix, has been regarded as a complex skin disorder, imposing substantial social and financial burdens1,2,3. Studies have reported that the incidence of HS following linear surgical wounds is around 35%, with the rate rising to as high as 72% in burn patients4,5. Local corticosteroid injections, particularly triamcinolone acetonide (TA) and compound betamethasone (CB), remain the first-line treatments for HS6,7,8. However, their variable efficacy and potential side effects, such as skin atrophy and pigment deposition, limit their use. As an innovative alternative, photodynamic therapy (PDT) is gaining attention for its high efficacy and precise controllability. The Food and Drug Administration (FDA) has approved 5-aminolevulinic acid (5-ALA) as a photosensitizer precursor, which is metabolized to protoporphyrin IX (PpIX), generating abundant reactive oxygen species (ROS) for hypertrophic scar fibroblasts (HSFs) killing upon light exposure9. However, the further metabolism of PpIX into heme not only results in the loss of the photosensitizer but also poses potential toxicity to normal tissues10,11. For decades, 5-ALA-derived heme has been regarded as a metabolic waste, with numerous studies focused on mitigating its accumulation12,13,14. From the perspective of natural material recycling, metabolic byproducts may be repurposed into functional resources under suitable conditions. Inspired by this, we are exploring the possibility of turning these useless heme into a potential “auxiliary energy source” to amplify the therapeutic efficacy of PDT.
Numerous studies have demonstrated that heme can be catabolized into Fe2+ and CO through the action of heme oxygenase-1 (HO-1), thereby inducing iron overload and ferroptosis15,16,17,18. Coincidentally, it has been observed that HSFs are naturally addicted to iron, suggesting ferroptosis as a potential alternative treatment19. Inspired by this, 5-ALA-derived heme may become a potential source for ferroptosis, amplifying the therapeutic effect for HS. However, due to the limited abundance of HO-1 within cells, catalyzing the excessive accumulation of heme to facilitate the process of ferroptosis presents a significant challenge. Therefore, efforts to excessively activate the HO-1-heme metabolic axis and optimize the utilization of 5-ALA metabolic waste are critical for establishing an efficient ferroptosis bioreactor post-PDT.
Baicalin is a flavonoid extracted from the root of Scutellaria baicalensis. As one of the oldest medicinal plants recorded in Chinese ancient pharmacopeia, baicalin has been widely used to treat inflammation, infection and cancer for centuries20,21,22. Recent studies have highlighted baicalin’s ability to enhance HO-1 expression, suggesting that co-delivering baicalin with 5-ALA may initiate a complex cascade reaction23,24. Furthermore, baicalin has been shown to independently trigger ferroptosis in cancer treatment25,26,27,28. When combined with the enhanced HO-1-heme metabolic axis, this may create a continuous ferroptosis bioreactor within HSFs. Compared to other ferroptosis inducers such as erastin, baicalin is more affordable and exhibits lower toxicity to normal tissues. However, its poor solubility and lack of specific targeting severely restrict its clinical application29,30. Recently, various nanocarriers have been employed to improve the poor bioavailability of hydrophobic drugs and enhance their therapeutic efficiency31,32. Among these, human H-ferritin (HFn) has garnered significant interest due to its excellent water solubility, high drug loading capacity and intrinsic targeting ability towards transferrin receptor 1 (TFR1), which is often overexpressed in tumor cells33,34,35. Interestingly, HSFs also overexpress TFR1, suggesting that the HFn-based delivery strategy may hold promise for the targeted therapy against HS19.
Herein, a zero-waste “ferroptosis amplifier” capable of targeting HSFs was engineered by co-encapsulating 5-ALA and baicalin into an HFn nanocage (FAB NPs). To enhance transdermal delivery, these FAB NPs were integrated with polyvinylpyrrolidone (PVP) microneedles (FAB@MN), providing a painless and convenient mode of administration (Fig. 1A). Once penetrating the epidermal barrier, FAB@MNs target HSFs via the TFR1-HFn interaction, thereby enhancing intracellular drug delivery and retention (Fig. 1B). Then, a synergistic therapy for HS can be implemented by PDT and ferroptosis based on multistage cascade metabolites (Fig. 1C). Specifically, 5-ALA is converted into PpIX to initiate PDT, followed by the release of baicalin to induce ferroptosis. Meanwhile, baicalin synergizes with the ROS and heme accumulated during PDT, activating the HO-1-heme metabolic axis to release Fe²⁺ and CO, further amplifying ferroptosis. This was not merely an amplification of a single process but a multi-layered feedback loop. As iron-catalyzed Fenton reactions continue, the generated ROS further upregulate HO-1, thus driving heme degradation and propelling ferroptosis. Unlike traditional approaches that focus solely on the immediate effects of PDT, this strategy repurposes 5-ALA metabolic waste as an “auxiliary energy source” to trigger a sustained ferroptosis cascade. Therefore, each brief PDT session sparks a sustained ferroptosis effect, enabling FAB@MN to significantly inhibit HSFs after a single laser exposure (Fig. 1D). Despite the potential for further optimization, this approach offers a compelling strategy for harnessing 5-ALA-derived byproducts to enhance the synergistic efficacy of PDT and ferroptosis, with promising implications for the treatment of HS and other related disorders.

A Fabrication process of the FAB@MN patch. B The targeting capacity of FAB NPs toward HSFs. C Formation of a ferroptosis amplifier via reutilization of 5-ALA photodynamic waste. D Therapeutic process of FAB@MN for HS.
Results and discussion
Fabrication and characteristics of FAB@MN
Herein, we reported a one-pot method to co-encapsulate 5-ALA and baicalin within the hollow core of the HFn nanocage, which were then incorporated into PVP MN patches (Fig. 2A). Specifically, 5-ALA and baicalin were mixed with HFn solutions and heated to 60 °C. It has been demonstrated that hyperthermia can expand the channels of HFn, enabling the encapsulation of small molecules without compromising the protein structure33,36. Transmission electron microscopy (TEM), dynamic light scattering (DLS), and Zeta potential analysis were performed to characterize HFn and the newly synthesized FAB NPs. As shown in Fig. 2B, HFn exhibited a typical ring-like structure with a hollow core, while FAB NPs appeared as solid spheres, likely due to the filling of 5-ALA and baicalin. Notably, dual drug loading did not alter the high monodispersity and good colloidal stability of HFn, while markedly enhancing the water solubility of baicalin, thus amplifying its potential for biomedical applications (Fig. 2B, C and Fig. S1). According to the DLS data, the average diameters of HFn and FAB NPs were 10.11 nm and 11.68 nm, respectively (Fig. 2D). The slight increase in the size of FAB NPs is likely due to the encapsulated drugs. And the more negative Zeta potential of FAB NPs may be attributed to the inherent negative charge of baicalin molecules (Fig. 2D)37.

A Schematic illustration of steps for the preparation of FAB@MN. B TEM and C digital images of HFn and FAB NPs solutions (Scale bar: 20 nm). D DLS and Zeta potential analysis (n = 3 biologically independent samples). E UV-Vis spectra of 5-ALA, baicalin, HFn and FAB NPs. F Native-PAGE characterization. G (i) The SEC and (ii) CD analysis of HFn, FA NPs, FB NPs and FAB NPs. H The schematic diagram of MN fabrication and transdermal drug delivery. I Digital photos and J mechanical compression test of PVP MN and FAB@MN patches. K SEM image of FAB@MN. L Typical images of rabbit HS skin after applying FAB@MN (Scale bar: 2 mm). M (i) Fluorescent photograph of FAB@MN (FAB NPs were labeled by red RhB) (Scale bar: 50 μm) and (ii) the distribution of RhB-labeled FAB NPs in the depths of rabbit HS after FAB@MN insertion for 5 min (Scale bar: 100 μm). Data represent the mean ± SD and a two-sided unpaired Student’s t-test was used for the comparison in (D). Experiments in (B), (F), (K), and (M) were each performed thrice independently, yielding similar results each time. Source data are provided as a Source Data file.
Ultraviolet-visible (UV-Vis) spectroscopy further confirmed the successful encapsulation of 5-ALA and baicalin within HFn, as indicated by their distinct characteristic absorption peaks (Fig. 2E). The high-performance liquid chromatography (HPLC) analysis determined that the encapsulation efficiency of 5-ALA was 57.42 ± 2.46%, with a drug loading capacity of 19.43 ± 0.67%. For baicalin, the encapsulation efficiency was 37.99 ± 0.60%, and the drug loading capacity was 12.86 ± 0.19% (Fig. S2). To confirm that the drug loading process did not affect the structure of HFn, comprehensive analyses including native polyacrylamide gel electrophoresis (Native-PAGE), circular dichroism (CD) spectra and size-exclusion chromatography (SEC) were conducted. As shown in Fig. 2F, FAB NPs appeared as a ≈ 440 kDa protein in Native-PAGE, similar to that of HFn. The CD spectra and SEC elution profiles of FA NPs (HFn encapsulating 5-ALA), FB NPs (HFn encapsulating baicalin) and FAB NPs also closely matched those of HFn, confirming the secondary structure and oligomerization states of HFn remained unchanged after drug encapsulation (Fig. 2G(i, ii)). Additionally, the stability of FAB NPs in cell culture media was assessed, revealing no significant change in size over 48 h under physiological conditions (Fig. S3).
To facilitate the in vivo biomedical applications, we developed a percutaneous delivery system for FAB NPs utilizing PVP MNs (Fig. 2H). Specifically, a 20% PVP solution containing FAB NPs was cast into a poly-dimethylsiloxane (PDMS) mold to dry overnight. The optical images demonstrated that the fabricated MN patch consisted of 225 needles spread over a 14 × 14 mm² area, with the patch turning slightly yellow after loading with FAB NPs (Fig. 2I). Scanning electron microscopy (SEM) further revealed that each MN was pyramid-shaped, with 340 μm in diameter at the base and 800 μm in height (Fig. 2K). Additionally, the mechanical hardness of the fabricated FAB@MN was evaluated, showing a force of 0.11 N/needle, which was sufficient for skin penetration (Fig. 2J). Intriguingly, the mechanical strength of FAB@MN was found to be higher than that of the blank PVP MN, possibly due to the hydrogen bonding and electrostatic interactions between the proteins and polymers. The penetration ability of FAB@MN was further demonstrated on rabbit ear HS models, where regular perforations were observed in the skin following insertion (Fig. 2L). And the fluorescence staining images revealed these MNs successfully penetrated the epidermal layer, capable of delivering Rhodamine B (RhB)-labeled FAB NPs to the deeper dermal layer (Fig. 2M(i, ii)).
Targeting ability and programmable release behavior of FAB NPs
Although various nanodrugs have been explored for eliminating HSFs, the enhancement of their targeting capabilities has often been neglected. Recently, the HFn protein-based drug delivery system has gained widespread use in cancer treatments, emerging as a hotspot in the field of nanomedicine33,38. Studies have demonstrated that HFn not only possesses excellent drug-loading capacity but also exhibits pH-responsive disassembly and high targeting specificity for TFR139,40,41. In this study, HFn was employed to encapsulate 5-ALA and baicalin (FAB NPs), aiming to optimize the therapeutic effects of free drugs while minimizing damage to normal tissues. To further explore the therapeutic potential of these FAB NPs for HS, their targeting ability and release behavior were thoroughly characterized.
Initially, double-blind molecular docking was conducted using HFn (PDB ID: 7CK9) as the receptor to evaluate the structural stability of 5-ALA and baicalin encapsulation. As shown in Fig. 3A, both molecules exhibited stable binding on the HFn surface, forming fourteen intermolecular contacts comprising five hydrogen bonds, seven van der Waals forces, and two π–alkyl interactions. The binding energies for 5-ALA and baicalin with HFn were calculated as −4.4 and −9.4 kcal mol−1 respectively, indicating that the resulting complex structures were stable, thereby laying a solid foundation for FAB NPs to retain the TFR1-targeting capability of native HFn. Previous studies have successfully employed HFn to target the overexpressed TFR1 in tumor cells, yielding promising therapeutic outcomes33,41. According to our results from western blotting and quantitative reverse transcription polymerase chain reaction (qRT-PCR), the TFR1 level in HSFs was significantly higher than that in normal fibroblasts (Fig. 3B(i–iii) and Fig. S4). The immunohistochemical staining also demonstrated the increased TFR1 levels in scarring tissues, which potentially offered more binding sites for FAB NPs (Fig. S5). To verify the targeting ability of FAB NPs towards HSFs, we constructed RhB-labeled FAB NPs and co-incubated them with normal fibroblasts and HSFs, respectively. As shown in Fig. 3B(iv) and Fig. S6, the fluorescence intensity in HSFs was 7.43-fold higher than in normal fibroblasts, demonstrating the superior targeting efficacy of FAB NPs toward HSFs. Flow cytometry analysis further confirmed these findings, with results consistent with the observed fluorescence patterns (Fig. S7).

A Molecular docking simulation of 5-ALA and baicalin with HFn. B Targeting ability of FAB NPs towards HSFs. (i) The schematic diagram illustrating the difference in uptake of FAB NPs by normal fibroblasts and HSFs. (ii) Protein and (iii) relative mRNA expression of TFR1 in normal fibroblasts and HSFs (n = 3 biologically independent samples). (iv) Representative fluorescence images of RhB-labeled FAB NPs internalized by normal fibroblasts and HSFs (green, Actin-Tracker Green-stained cytoskeleton; blue, DAPI-stained nucleus; red, RhB-labeled FAB NPs; Scale bar: 10 μm). C The deep penetration of FAB NPs in 3D scarring spheroid model. (i) The fabrication and treatment process of scarring spheroids. (ii) CLSM images of scarring spheroids after different treatments (Green, Fluorescein Isothiocyanate (FITC)-labeled 5-ALA; Red, RhB-labeled baicalin. Scale bar: 50 μm). (iii) 3D surface plot of the 75 μm sections from different groups. D The subcellular location and release profile of FAB NPs. (i) Representative CLSM images of FAB NPs colocalized with lysosomes after endocytosis (green, Lyso-Tracker Green-stained lysosome; blue, Hoechst-stained nucleus; red, RhB-labeled FAB NPs. Scale bar: 5 μm). (ii) Cumulative release profiles of 5-ALA and baicalin from FAB NPs in solutions at different pH values in vitro (n = 3 biologically independent samples). (iii) The FEL of 5-ALA-HFn (left) and baicalin-HFn (right). E Scheme illustrating the therapeutic strategies and characteristics of FAB NPs for HSFs. Data represent the mean ± SD. Two-sided unpaired Student’s t-test was used for the comparison in B (iii). Experiments in (C) were performed thrice independently, yielding similar results each time. Source data are provided as a Source Data file. Schematic in (C) was created using BioRender. Yuan, C. (2025) https://BioRender.com/r1hiu7x.
As many drugs proven effective in 2D cell cultures lose their potency in dense scar tissues in vivo, we further investigated the deep penetration abilities of FAB NPs in 3D scarring spheroid models using confocal laser scanning microscopy (CLSM) (Fig. 3C(i)). As shown in Fig. 3C(ii), the spheroids treated with free 5-ALA and baicalin displayed weak fluorescence, with drugs predominantly confined to the periphery. In contrast, spheroids incubated with FAB NPs exhibited widespread fluorescence throughout the entire structure. The fluorescence intensity of the treated spheroids was further quantified by 3D surface plots, revealing distinct signals penetrating deep into the spheroid core following incubation with FAB NPs (Fig. 3C(iii)). Considering the excellent targeting and penetration abilities of FAB NPs in both 2D and 3D scar models, we proceeded to assess their specific cytotoxicity towards HSFs. Normal fibroblasts and HSFs were exposed to varying concentrations of FAB NPs, with the Cell Counting Kit-8 (CCK8) assay revealing significantly greater cytotoxicity towards HSFs, highlighting their marked selectivity for HS (Fig. S8). Based on this, we further developed a microneedle system for the localized delivery of FAB NPs, enhancing treatment precision while minimizing potential harm to surrounding tissues.
The subcellular location and release behavior of FAB NPs were also explored. As shown in Fig. 3D(i) and Fig. S9, a large portion of FAB NPs colocalized with Lyso-Tracker (a typical marker of lysosome) after internalization. Recent studies have indicated that HFn remains stable in neutral environments, but its subunit interactions weaken and disassemble under acidic conditions33,42. Leveraging this property, FAB NPs were expected to selectively release 5-ALA and baicalin in acidic lysosomes, thus minimizing premature leakage during administration. As depicted, a complete release of 5-ALA and baicalin was observed after 24 h of incubation at pH 5.0, significantly faster than that at pH 7.4 (Fig. 3D(ii)). Moreover, notable discrepancies were observed in the release rates of 5-ALA and baicalin, particularly within the initial 4 h, with approximately 81.41% of 5-ALA and 53.87% of baicalin released. To explore the mechanisms behind these differential release behaviors, we conducted molecular dynamics simulations and computed the free energy landscape (FEL) and free binding energy for each drug-protein interaction. In cases of weak or unstable protein-ligand interactions, the FEL typically exhibits multiple minima clusters. As expected, the 5-ALA-HFn complex had four small energy clusters, whereas the baicalin-HFn complex exhibited only a single concentrated energy cluster, indicating relatively stronger binding stability (Fig. 3D(iii)). Consistently, the binding free energy of 5-ALA-HFn was calculated as −10.68 kcal mol−1, higher than that of baicalin-HFn at −42.05 kcal mol−1. A lower binding free energy often indicates a stronger binding affinity, which may explain the slower release of baicalin compared to 5-ALA from the FAB NPs (Fig. S10). The diagram in Fig. 3E summarizes the precise targeting and unique drug release properties of FAB NPs in treating HS. Initially, FAB NPs selectively bind to the significantly overexpressed TFR1 on the surface of HSFs, achieving targeted drug localization and minimizing side effects on surrounding healthy tissues. Once internalized, FAB NPs degraded within the lysosomal acidic environment, initiating a programmable sequential release of the encapsulated drugs. Specifically, the rapid release of 5-ALA quickly initiates PDT, providing early therapeutic effects, while the slow release of baicalin offers sustained support. This sequential release pattern allows for a programmable approach to treat HS, significantly improving the precision and efficiency of the therapy.
Enhanced anti-scarring effects through FAB@MN-mediated 5-ALA metabolic waste reutilization
5-ALA serves as a precursor of photosensitizer PpIX, demonstrating potent photodynamic activity against HSFs9,43. However, a significant challenge arises as PpIX is quickly metabolized into non-photosensitive heme13,14. To address this challenge, we developed a waste-to-energy strategy to fully reutilize 5-ALA metabolic waste heme to enhance its anti-scar efficacy. In this strategy, FAB@MN releases 5-ALA to biosynthesize PpIX for PDT, during which substantial heme accumulates. Subsequently, baicalin significantly boosts the expression of HO-1 through its synergistic interaction with the ROS and heme accumulated from PDT. This upregulation facilitates the decomposition of heme into therapeutically beneficial Fe2+ and CO, thereby inducing oxidative damage to HSFs. Moreover, this cascade establishes a multi-layered feedback loop, where iron-catalyzed Fenton reactions enhance ROS production, thus further promoting HO-1 overexpression. Through this strategy, we aim to transform the “end” metabolic waste heme into a bioavailable intracellular fuel to suppress HSF activity (Fig. 3I).
As the rate-limiting enzyme in heme catabolism, HO-1 efficiently converts heme into Fe²⁺, CO, and biliverdin44,45. Studies have shown that excessive HO-1-mediated heme metabolism can exacerbate cellular oxidative damage, offering potential for cancer treatment46,47,48. In this study, we observed differential HO-1 expression levels across the FA@MN, FB@MN and FAB@MN groups, which correlate closely with their cellular effects. As shown, HO-1 expression was elevated in both the FA@MN and FB@MN groups compared to the control, but significantly lower than in the FAB@MN group (Fig. 4A, B). The slight upregulation of HO-1 expression observed in the FA@MN group may be attributed to a negative feedback response triggered by elevated levels of ROS and heme following PDT49,50,51. And the increase in the FB@MN group suggested that baicalin was a potent enhancer of HO-1. To better understand the interactions between baicalin and HO-1 protein at the atomic level, molecular docking studies were subsequently conducted. As shown, baicalin formed hydrogen bond interactions with multiple residues of HO-1, showing a strong binding affinity of −12.5 kcal mol−1 (Fig. 4C). These findings suggest that baicalin may directly bind to the HO-1 protein, thereby modulating its activity or expression to activate the HO-1–heme metabolic axis. Particularly following 5-ALA-mediated PDT, the increased ROS and heme further amplified the baicalin’s effects, initiating a powerful synergy to upregulate HO-1 in the FAB@MN group.

A Western blot and B the corresponding quantitative analysis of HO-1 proteins in different groups (n = 3 biologically independent samples). C Molecular docking visualization of baicalin with HO-1 protein. D Quantitative analysis of intracellular heme in different groups by ELISA kits (n = 3 biologically independent samples). E Fluorescence images of CO labeled by COP-1 and Fe²⁺ labeled by FerroOrange in HSFs under different treatment conditions (Scale bar: 50 μm). F DCFH-DA staining images of different groups (green, DCFH-DA; blue, Hoechst-stained nucleus. Scale bar: 50 μm). G Apoptosis analysis of HSFs by flow cytometry after different treatments. H Statistical analysis of the cell viability of HSFs (n = 3 biologically independent samples). I Scheme illustrating FAB@MN enhanced their cytotoxic effect by recycling 5-ALA photodynamic waste heme. Data represent the mean ± SD. One-way ANOVA followed by Tukey’s post hoc test was used for comparisons in (B), (D) and (H), and all tests were two-sided. The exact p-values have been indicated in the figures. Experiments in (E, F) were each performed thrice independently, yielding similar results each time. Source data are provided as a Source Data file. Schematic in (I) was created using BioRender. Yuan, C. (2025) https://BioRender.com/r1hiu7x.
We then analyzed the key metabolites within the 5-ALA-PpIX-heme-Fe²⁺/CO/biliverdin axis across different groups. Utilizing enzyme-linked immunosorbent assay (ELISA) and fluorescence imaging techniques, we quantified the PpIX and heme levels. As shown in Fig. S11, both the FA@MN and FAB@MN groups exhibited a similar increase in PpIX. However, significant differences were observed in heme, with the FA@MN group showing a notable increase compared to the control, while the FAB@MN group exhibited minimal changes, indicating enhanced heme degradation by FAB@MN (Fig. 4D). Tracking the heme decomposition products, we observed no significant biliverdin fluorescence across the groups, likely due to its rapid degradation by abundant biliverdin reductase enzymes (Fig. S12)18,52. However, the FA@MN group exhibited enhanced fluorescence for Fe2+ and CO relative to the control, confirming the excess heme and ROS generated during PDT upregulated HO-1, thus promoting heme degradation (Fig. 4E). During 5-ALA treatment, the conversion of PpIX to heme typically increased iron uptake, explaining the higher free Fe2+ levels in the FA@MN group, despite substantial Fe2+ binding by heme18,53,54. Notably, the FAB@MN group displayed the strongest fluorescence for Fe²⁺ and CO, which was largely reversed by the HO-1 inhibitor, emphasizing the critical role of overexpressed HO-1 in heme reutilization. Interestingly, no detectable CO was observed in the FB@MN group, likely due to the limited heme available in cells. And the elevated Fe²⁺ levels in this group could be attributed to baicalin’s independent ferroptosis mechanism as reported in previous researches25,26.
Recently, Fang et al. made a groundbreaking discovery by leveraging downstream metabolites of 5-ALA-derived heme, such as Fe²⁺ and CO, to amplify oxidative stress in cancers18. Inspired by this, we explored whether the FAB@MN could effectively promote oxidative cytotoxicity against HSFs through the enhanced 5-ALA-PpIX-heme-HO-1 metabolic axis. Using DCFH-DA staining, we evaluated the oxidative stress levels of HSFs across different groups. The results revealed that FA@MN, FB@MN and FAB@MN all elevated intracellular ROS levels, with FAB@MN inducing the most significant increase (Fig. 4F and S13). Although baicalin can be used as an antioxidant, numerous studies have highlighted its dual role in inducing ROS and ferroptosis, with this activity varying based on different cellular environments55,56,57. Notably, FAB@MN elicited a synergistic effect that exceeded the additive impact of FA@MN and FB@MN, resulting in a pronounced elevation of intracellular ROS. And this increase was significantly mitigated by the HO-1 inhibitor, further supporting the pivotal role of the HO-1-heme metabolic axis in driving ROS storm in HSFs. As the iron-catalyzed Fenton reaction progresses, the accumulated ROS act as an activator to further upregulate HO-1 expression, thus establishing a self-perpetuating feedback loop (Fig. 4I(i))51. By repurposing 5-ALA–derived heme as a therapeutic energy source, FAB@MN offers a promising strategy to enhance anti-scarring effects following PDT.
As indicated by the CCK-8 assay, the relative cell viability in the FA@MN and FB@MN groups decreased to 72.2% and 61.9% respectively, while only 13.8% of HSFs survived in the FAB@MN group after a single irradiation, significantly outperforming the combined effects of FA@MN and FB@MN (Fig. 4H). However, inhibition or knockdown of HO-1 notably reduced FAB@MN’s therapeutic efficacy, increasing cell survival to 36.6% and 42.9% respectively (Fig. 4H and Fig. S14). Flow cytometry analysis also revealed that the HO-1 inhibitor significantly lowered the apoptosis rate in the FAB@MN group, emphasizing FAB@MN’s role in enhancing HSFs killing by upregulating the HO-1-heme metabolic axis (Fig. 4G and Fig. S15). Building on these findings, it can be concluded that FAB@MN not only generates PpIX to initiate PDT but also upregulates HO-1 to decompose heme for oxidative damage (Fig. 4I). This chemical reaction storm has been demonstrated to enhance the anti-scarring effects of PDT, effectively converting waste metabolites into treasure.
FAB@MN triggers continuous ferroptosis in HSFs
Ferroptosis is a regulated form of cell death characterized by the accumulation of Fe2+ and lipid peroxidation (LPO)58,59. Recent studies have shown that enhanced heme degradation and the subsequent release of Fe²⁺ serve as potent inducers of ferroptosis16,60. Concurrently, the role of CO in exacerbating mitochondrial damage and driving ferroptosis has become increasingly evident18,61,62. Building on these findings, we propose that FAB@MN-mediated reutilization of 5-ALA-derived heme can effectively trigger ferroptosis in HSFs (Fig. 5A). Additionally, studies have indicated that baicalin not only promoted HO-1 expression but may also independently induce ferroptosis, with this dual action further reinforcing the continuous ferroptosis effects of FAB@MN25,26,27,28.

A Schematic illustration to reveal the mechanism of 5-ALA photodynamic waste augmented ferroptosis in HSFs. B Relative levels of LIP, C GSH and D MDA in HSFs with different treatments (n = 3 biologically independent samples). E Representative fluorescence images of intracellular LPO in different groups by staining with C11-BODIPY581/591 probe (Scale bar: 50 μm). F Western blot analysis and G, H corresponding grayscale quantification of GPX4 and xCT proteins in HSFs treated with various interventions (n = 3 biologically independent samples). I Fluorescent assessment of MMP level in HSFs using JC-1 fluorescent probe (Scale bar: 50 μm). J Representative fluorescent images of mitochondrial morphology in HSFs (Red, Mito-Tracker Deep Red-stained mitochondria; blue, Hoechst-stained nucleus. Scale bar: 10 μm). K TEM images of mitochondrial ultrastructure. Structures colored red indicate mitochondria. L Flow cytometry analysis of MMP in HSFs stained by TMRM probe. M Statistical data of mitochondrial individuals in HSFs based on fluorescent images with Mito-Tracker Deep Red probe (n = 3 biologically independent samples). Data represent the mean ± SD. One-way ANOVA followed by Tukey’s post hoc test was used for comparisons in (B–D), (G, H), (M), and all tests were two-sided. The exact p-values have been indicated in the figures. Experiments in (E), (I–K) were each performed thrice independently, yielding similar results each time. Source data are provided as a Source Data file. Schematic above (I–L) was created using BioRender. Wang, S. (2025) https://BioRender.com/7i8m28b.
To elucidate the role of ferroptosis in the effects of FAB@MN, we introduced the necroptosis inhibitor necrostatin-1 (Nec-1), apoptosis inhibitor Z-VAD-FMK, and ferroptosis inhibitor deferoxamine (DFO) for cell viability assays (Fig. S16). The results showed that Nec-1 and Z-VAD-FMK provided minimal rescue effects, likely by suppressing PDT-induced damage. In contrast, DFO significantly increased the cell viability from 14.9% to 76.2%, suggesting ferroptosis played a dominant role in FAB@MN-induced cell death independent of PDT. According to our strategy, FAB@MN transformed 5-ALA metabolic waste heme into a potent storm of Fe2+ and CO. This cascade, along with baicalin’s inherent ferroptosis potential, initiated robust and sustained ferroptosis in HSFs. To demonstrate it, we investigated several ferroptosis biomarkers across different groups. As shown in Fig. 5B, FAB@MN-treated HSFs exhibited the highest labile iron pool (LIP), which was 6.07-fold higher than that of the control group. While the level of antioxidants such as GSH dropped to 35.70% of the control, significantly lower than those in other groups (Fig. 5C). Furthermore, using the C11-BODIPY581/591 probe to assess intracellular LPO, we observed heightened green fluorescence in FAB@MN-treated HSFs compared to the FB@MN group, indicating increased LPO production via heme biotransformation following PDT (Fig. 5E and Fig. S17). Intracellular malondialdehyde (MDA), the end product of lethal LPO, also increased by 1075.53% in FAB@MN-treated HSFs, far exceeding other groups (Fig. 5D). Notably, DFO treatment significantly reversed these ferroptosis biomarkers, reinforcing the role of ferroptosis in FAB@MN-induced cell death. Furthermore, FAB@MN-treated HSFs showed the most pronounced downregulation of GPX4 and xCT, with expression levels significantly lower than those observed in the FB@MN group (Fig. 5F, H). Previous research has shown that baicalin can downregulate GPX4 and xCT, triggering LPO accumulation and ferroptosis25,63. Additionally, excessive activation of the HO-1-heme metabolic axis may further exacerbate the downregulation of GPX4 and xCT64,65,66. To validate this, we performed HO-1 knockdown, resulting in a significant restoration of GPX4 and xCT, thus confirming the pivotal role of the 5-ALA-PpIX-HO-1-heme-Fe²⁺ metabolic axis in FAB@MN-mediated ferroptosis (Fig. S18).
As previously reported, mitochondrial dysfunction is closely involved in ferroptosis. Therefore, we investigated the mitochondrial structure and mitochondrial membrane potential (MMP) of HSFs treated with different materials. Initially, the JC-1 and TMRM probe was utilized as the fluorescent indicator for MMP detection. As shown, FAB@MN-treated HSFs exhibited the brightest JC-1 green fluorescence and the weakest red fluorescence, alongside a significant decrease in TMRM intensity, suggesting the most pronounced MMP depolarization (Fig. 5I, L and Fig. S19). Subsequently, mitochondrial morphology analysis revealed that FAB@MN induced abnormal mitochondrial structures in HSFs, including rod-like, spherical and fragmented shapes, in stark contrast to the elongated and interconnected mitochondria observed in the control group (Fig. 5J). TEM images confirmed severe mitochondrial shrinkage and reduced cristae in FAB@MN-treated HSFs, consistent with mitochondrial changes typically associated with ferroptosis19. And the quantitative analysis via ImageJ software showed an increase in mitochondrial number but a reduction in branching length after FAB@MN treatment (Fig. 5M and Fig. S20). Notably, all observed mitochondrial damage observed in the FAB@MN group could be rescued by DFO, further confirming that the mitochondrial damage in HSFs was associated with ferroptosis.
FAB@MN-induced crosstalk between ferroptosis and mitophagy in HSFs
FAB@MN acted as a ferroptosis bioreactor by leveraging 5-ALA metabolic waste heme and the inherent ferroptosis property of baicalin. To further explore its underlying mechanisms, transcriptome sequencing was conducted. As shown in Fig. 6A, the volcano plot revealed a total of 2413 differentially expressed genes (DEGs) following the treatment with FAB@MN, with 1364 up-regulated and 1049 down-regulated. Gene ontology (GO) enrichment analysis revealed significant enrichment of DEGs in processes such as mitochondrial membrane permeability, stress response to metal ions, positive regulation of fatty acid oxidation, and cellular response to oxidative stress (Fig. 6B). Gene set enrichment analysis (GSEA) based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) database highlighted the activation of pathways related to HIF1, mitophagy and ferroptosis, with key genes like HO-1, xCT, p62, and LC3B implicated (Fig. 6C, D). Additionally, protein-protein interaction (PPI) network analysis of DEGs enriched in both ferroptosis and mitophagy revealed extensive interactions, indicating a possible association between FAB@MN-induced ferroptosis and mitophagy (Fig. 6E). Previous research have revealed the intricate crosstalk between ferroptosis and mitophagy67,68. In the early stages of ferroptosis, mitophagy is typically activated to remove damaged mitochondria and alleviate oxidative stress. However, excessive mitophagy can release iron from mitochondria, further amplifying ferroptosis and perpetuating a vicious cycle69,70. Based on this, we hypothesized that FAB@MN not only induced ferroptosis but also triggered mitophagy, with potential direct or indirect interactions between these two processes.

A Volcano plot of DEGs. DEGs were identified using the R package DESeq2 with multiple testing correction. Genes with P.adjust <0.05 and |log2FoldChange | > 1 were considered significant. B Results of GO enrichment analysis for DEGs based on hypergeometric analysis. C KEGG pathway analysis of DEGs. ES calculated by permutation test, normalized to NES. Nominal p-value estimated via 1000 phenotype permutations, comparing observed ES to null distribution based on sign. D Circular visualization of the results of functional enrichment analysis. E PPI network of DEGs involved in ferroptosis and mitophagy. F (i) TEM images of mitophagosome in control and FAB@MN group. (ii) Representative fluorescence images showing mitophagy (red, Lyso-Tracker Red-stained lysosome; green, Mito-Tracker Green-stained mitochondria; blue, Hoechst-stained nucleus). (iii) Schematic illustration of mitophagy formation in FAB@MN group. G Western blot analysis of the expression of p62, LC3B, PINK1 and Parkin proteins in HSFs treated with different interventions. H Statistical analysis of the cell viability of HSFs (n = 3 biologically independent samples). NC, negative control siRNA. I Flow cytometry analysis of intracellular ROS in HSFs stained by DCFH-DA probe. J CLSM images of intracellular Fe2+ upon different treatments (Scale bar: 50 μm). K Fluorescence images of LPO stained with C11-BODIPY581/591 probe (Scale bar: 50 μm). L Schematic of the ferroptosis mechanism induced by mitophagy. Data represent the mean ± SD. One-way ANOVA followed by Tukey’s post hoc test was used for comparisons in (H). The exact p-values have been indicated in the figures. Experiments in (F), (J, K) were each performed thrice independently, yielding similar results each time. Source data are provided as a Source Data file. Schematic in (F) and (L) created using BioRender. Wang, S. (2025) https://BioRender.com/7i8m28b.
As shown in Fig. 6F(i), TEM analysis revealed FAB@MN significantly enhanced the formation of mitophagosomes in HSFs. And the fluorescence imaging also confirmed a marked increase in colocalization between mitochondria (stained with Mito-Tracker) and lysosomes (stained with Lyso-Tracker), further supporting the enhancement of mitophagy activity (Fig. 6F(ii, iii)). To elucidate the molecular mechanism of FAB@MN-induced mitophagy, the expression levels of several mitophagy-related proteins including LC3B, p62, PINK1 and Parkin were subsequently measured by immunoblotting assays (Fig. 6G). As expected, FAB@MN markedly upregulated the protein ratio of LC3B-II and LC3B-I, while decreasing p62 levels, indicating the formation of autophagosomes and the occurrence of autophagy. Furthermore, the abundance of PINK1 and Parkin proteins also significantly increased, suggesting that FAB@MN activated mitophagy via the classical PINK1/ Parkin pathway. Under normal conditions, PINK1 is rapidly degraded. However, when mitochondria are damaged, it accumulates and acts as a beacon to recruit Parkin to the surface of mitochondria. Parkin, endowed with E3 ubiquitin ligase activity, subsequently tags these damaged mitochondria, recruiting substantial p62 to the mitochondrial outer membrane. This promotes LC3B-I lipidation, ultimately leading to autophagosomal engulfment of the damaged mitochondria71,72,73.
To investigate the potential interplay between mitophagy and ferroptosis mediated by FAB@MN, we assessed the influence of the iron chelator DFO on mitophagy regulation. As expected, DFO effectively reduced the accumulation of Parkin and PINK1 while promoting the upregulation of p62, demonstrating that FAB@MN-induced iron overload played a pivotal role in abnormal mitophagy activation within HSFs (Fig. S21). We also explored the involvement of mitophagy in FAB@MN-mediated ferroptosis by silencing PINK1 (Fig. S22). As shown, PINK1 knockdown partially restored cell viability in the FAB@MN-treated group, boosting survival from 16.7% to 44.81% (Fig. 6H). Further analysis revealed that PINK1 knockdown not only reduced ROS levels induced by FAB@MN but also significantly decreased the fluorescence intensities of Fe²⁺ and LPO (Fig. 6I–K and Fig. S23). The role of mitophagy in ferroptosis remains controversial, often viewed as a double-edged sword. In some cases, mitophagy can remove damaged mitochondria and maintain cellular function68. However, under specific conditions, excessive mitophagy may release free Fe2+ to promote ferroptosis74. In this study, FAB@MN triggered iron overload, which subsequently mediated aberrant mitophagy activation via the PINK1/Parkin pathway, leading to elevated Fe2+ and LPO levels in HSFs and amplifying ferroptosis (Fig. 6L).
In vivo anti-scarring effects of FAB@MN
Despite the availability of various treatment options, managing HS remains a significant challenge, underscoring the urgent need for a universally effective and minimally invasive therapeutic approach7,75,76. Motivated by the promising in vitro results of FAB@MN, we further investigated its anti-scarring effects in vivo. Specifically, we established rabbit HS models and applied treatments once a week for a month (Fig. 7A). The results revealed that a single light exposure induced the highest levels of ROS in FAB@MN group, supporting the synergy between PDT and ferroptosis (Fig. 7B, C). After four-week treatments, scars in the FAB@MN group became smooth and indistinguishable from the surrounding healthy skin, with therapeutic effect significantly surpassing those of FA@MN, FB@MN, and even commercial corticosteroids like TA and CB (Fig. 7D and Fig. S24). Consistent with previous studies, although TA and CB injections led to some lightening of the scars, residual pigmentation persisted following treatments6,8. To assess efficacy more comprehensively, we measured scar color, thickness and hardness in each group (Fig. S25A). As shown in Fig. S25B, the redness value of FAB@MN-treated scars (2.00 ± 0.10) was closest to that of normal skin (1.97 ± 0.31). Ultrasound imaging further confirmed FAB@MN’s superior ability to reduce scar thickness, with a significant reduction from 3.70 ± 0.54 mm to 0.46 ± 0.09 mm (Fig. 7E and Fig. S24, S27A). Moreover, FAB@MN treatment resulted in a marked decrease in scar hardness, with a 53.9% reduction (17.53 ± 2.11 HOO) compared to untreated scars (38.03 ± 2.97 HOO) (Fig. S25C). Notably, both FA@MN and FB@MN exhibited limited therapeutic effects in vivo, suggesting short-term PDT or baicalin-induced ferroptosis alone was ineffective. While FAB@MN leveraged a series of 5-ALA metabolic waste to establish a sustained bioreactor integrating PDT and ferroptosis, thereby providing an effective strategy for scar treatment and recurrence prevention.

A Experimental timeline of animal studies. B Fluorescence imaging of ROS levels in rabbit ear HS models after different treatments, acquired using an In Vivo Imaging System (IVIS). C ROS staining of HS from different groups (Scale bar: 200 μm). D Representative photographs of the scars with four rounds of treatments (Scale bar: 5 mm). E Ultrasound images of scars after predetermined treatments (Scale bar: 2 mm). F H&E, G Masson, Sirius Red and GPX4 immunohistochemical staining of scar tissues after various treatments. H Scheme diagram of efficient anti-scarring efficacy of FAB@MN treatment. Experiments in (C), (F, G) were each performed thrice independently, yielding similar results each time. Source data are provided as a Source Data file. Some components of the schematic in (A) were created using BioRender. Wang, S. (2025) https://BioRender.com/7i8m28b.
Histological analyses were further conducted to assess the therapeutic outcomes of various treatments. Hematoxylin-eosin (H&E) staining showed the HS treated with FAB@MN returned to the smooth appearance of normal skin, while the other groups exhibited varying degrees of protuberance, with densely packed and disorganized fibers (Fig. 7F and Fig. S26A). Numerous studies have shown that abnormally proliferating HSFs secrete excessive collagen, particularly resulting in the accumulation of type I collagen and disrupting the collagen balance9,77. Therefore, direct targeting and destroying HSFs could potentially restore the collagen structure and alleviate fibrosis9,19,78. As expected, Masson staining demonstrated that FAB@MN significantly reduced collagen deposition and promoted more organized collagen fiber alignment at the HS sites (Fig. 7G and Figs. S26B, S27B). Sirius Red staining revealed a significant reduction in yellow-stained type I collagen in the FAB@MN group, suggesting substantial remodeling of the abnormal collagen matrix (Fig. 7G). Besides, the GPX4 level in HS tissues was also evaluated to assess the ferroptosis effects via immunohistochemical staining. As shown, FAB@MN notably reduced the area of GPX4-positive cells and exhibited a more effective therapeutic outcome than FB@MN, likely due to the ferroptosis cascade initiated by 5-ALA photodynamic waste (Fig. 7G, and Fig. S27C).
The biosafety of FAB@MN in vivo was verified by the H&E staining of vital organs (heart, liver, spleen, lung, and kidney), as well as hematological and biochemical analyses. No significant organ damage was observed in FAB@MN-treated group, and all blood parameters remained within normal limits after one month of treatment, confirming its negligible systemic toxicity in vivo (Figs. S28, S29). As a targeted ferroptosis nanomedicine for HS, FAB@MN not only enhances the therapeutic outcomes but also minimizes damage to healthy tissues (Fig. 7H). Furthermore, by utilizing byproducts from PDT, we have optimized the use of 5-ALA, ensuring efficient waste conversion and enhancing the synergistic effects between PDT and ferroptosis. Even compared to first-line corticosteroids, FAB@MN exhibits superior therapeutic efficacy, offering a promising alternative for HS management and presenting a potential strategy applicable to other pathological scars, such as keloids.
Methods
Ethical statement
This study complies with all applicable ethical regulations. Both human tissue collection and animal experiments were approved by the Ethics Committee of the Affiliated Stomatological Hospital of Chongqing Medical University (Approval No. CQHS-REC-2024-058). Human tissues were obtained with written informed consent from all participants and processed in accordance with the Declaration of Helsinki. Animal procedures were conducted in accordance with the Guidelines for the Care and Use of Laboratory Animals issued by the National Institutes of Health (NIH Publication No. 86–23, revised 1985) and the ARRIVE guidelines.
Cell culture and animals
Nine HS tissues and the matched normal skin tissues were provided by patients undergoing surgical treatment, and each patient has voluntarily signed an informed consent form. All participants in this study did not have any known systemic diseases and were not receiving any treatments that could potentially affect the study results. In brief, tissues were first cut into 3 mm-thick slices, and then digested overnight with 0.25% neutral protease II. Subsequently, the epidermal layer was peeled off, and further digestion was carried out for 4–6 h at 37 °C with 0.2% Collagenase II. After digestion, cells were filtered and collected using a cell strainer with a 75 μm pore size, then resuspended in Dulbecco’s Modified Eagle Medium (DMEM) containing 10% FBS and cultured at 37 °C in an atmosphere of 5% CO2. The method of isolating fibroblasts from normal skin was similar to the aforementioned process for obtaining HSFs with minor modifications.
Eighteen female Oryctolagus cuniculus (New Zealand White rabbits, 8–10 weeks old, ~2.5–3.0 kg) were obtained from the Experimental Animal Center of Chongqing Medical University (Chongqing, China). Animals were fed standard laboratory chow (Catalog No. 1016708478371979264, Beijing Keao Xieli Feed Co., Ltd., China) containing 17% protein, 3% fat, 10–15% fiber, and 11% moisture. They were housed individually under controlled conditions (25 ± 2 °C, 55 ± 2% relative humidity, 12-h light/dark cycle). Group allocation was performed in a blinded manner by a technician using a random number table to ensure unbiased distribution and intergroup comparability. Animals were monitored daily for general health, including activity, posture, intake of food and water, body weight, and wound healing. Signs of pain or distress were closely monitored. Humane euthanasia was performed under veterinary supervision upon evidence of sustained weight loss, anorexia, or pain-related behaviors. At the end of the study, all animals were humanely euthanized via CO2 inhalation using a gradual-fill method (20–30% of chamber volume per minute) in a sealed chamber. Death was confirmed by the cessation of respiration and loss of corneal reflex.
Preparation of FAB@MN
The synthesis of FAB@MN followed a detailed protocol. Initially, 5 mg of 5-ALA was dissolved in PBS, while 5 mg of baicalin was dissolved in ethanol. Then, 10 mg of HFn was dissolved in Tris-HCl buffer (20 mM, pH 7.5). The solutions of 5-ALA and baicalin were gradually added to the HFn solution under gentle stirring to prevent ethanol-induced denaturation of the protein. After mixing, the blend was subsequently incubated in a 60 °C water bath for 4 h, followed by centrifugation at 16,099 g for 15 min. The supernatant was collected and dialyzed four times in PBS using a dialysis bag (10 kDa), with the dialysis fluid replaced every 4 h to remove unencapsulated baicalin and 5-ALA. Subsequently, the obtained FAB NPs were mixed with a 20% PVP solution and poured into a PDMS mold, then placed under a vacuum of 600 mmHg for 1 min to ensure the solution completely filled the MN cavities. Finally, it was dried overnight in the dark before being demolded.
Characterizations of FAB NPs and FAB@MN
The protein structure changes of HFn were characterized with size-exclusion chromatography (SEC, Agilent 1260 Infinity II, USA) and circular dichroism (CD, JASCO J-1500, Japan). Particle size and Zeta potential of the materials were measured using a Zetasizer Nano ZS90 (Malvern, UK). The ultraviolet absorption spectra of the materials were determined using a multimode microplate reader (PerkinElmer EnSpire, USA). Nanoparticle morphology was visualized using a JEM-1400 Plus transmission electron microscope (TEM, Tokyo, Japan), with grid staining performed using 1% (w/v) uranyl acetate for several minutes before observation. Molecular docking simulation was performed with Auto Dock Vina software (Version 1.2.0), and molecular dynamics simulation was performed by the GROMACS (2022) program. The fabricated MN patches were then examined by scanning electron microscopy (SEM, FEI Hillsboro, USA), and their mechanical properties were assessed using a universal material testing machine (INSTRON, USA). To evaluate the skin penetration ability of FAB@MN (with RhB-labeled FAB NPs), we vertically inserted it into rabbit ear HS for 5 min. The treated skin sample was then embedded in optimal cutting temperature (OCT), and sliced into 10 μm sections using a cryotome (CM1860, Leica). Subsequently, these skin sections were placed on adhesive microscope slides and observed using a multi-function microscope.
Targeting ability and cellular uptake of FAB NPs
To investigate the targeting ability of FAB NPs towards HSFs, RhB-labeled FAB NPs were added into HSFs for 4 h. Then, cells were fixed in 4% cold formaldehyde, and the nuclei and cytoskeleton were stained with DAPI and Actin-Tracker-Red respectively. The fluorescence emitted from the treated cells was evaluated using a CLSM (Leica TCS SP8, Germany) and flow cytometry (BD Biosciences, USA). To further assess the deep penetration capability of FAB NPs in a 3D cell model, we initially planted 5 × 104 cells/well in ultra-low attachment round-bottom 48-well plates (Corning, USA) to cultivate 3D cell spheroids. After the formation of 3D scarring spheroids, the fluorescent-labeled NPs were co-incubated for 4 h, and the fluorescence images were captured and analyzed using a CLSM.
In vitro drug release and subcellular colocalization of FAB NPs
We employed High-Performance Liquid Chromatography (HPLC, Agilent, USA) for the precise quantification of 5-ALA and baicalin concentrations. The encapsulation efficiency (%) and loading capacity (%) of 5-ALA and baicalin were calculated using the following formulas: encapsulation efficiency (%) = amount of encapsulated drugs/amount of drugs added; loading capacity (%) = amount of encapsulated drugs /amount of FAB NPs. To examine the release profiles of FAB NPs, 5 mL of FAB NPs solution were put into a dialysis bag (10 kDa) that was directly immersed into 20 mL PBS at 37 °C with pH values of 7.4 and 5.0. At predetermined time intervals, the concentrations of the released drugs were tested. Stability evaluation was conducted by incubating FAB NPs with cell culture medium containing 10% FBS for 48 h, and assessed their stability using a Zetasizer Nano ZS90 (Malvern, UK). For the subcellular colocalization imaging experiments, HSFs were seeded into 48-well plates at a density of 1 × 104 cells/well and incubated with RhB-labeled FAB NPs for 4 h. After removing the culture medium, the cells were washed twice with cold PBS. Lysosomes and nuclei were stained with Lyso-Tracker and Hoechst 33342 for 30 and 10 min respectively. After staining, the intracellular fluorescence signals were captured with CLSM.
In vitro analysis of 5-ALA metabolic products in HSFs
To evaluate the metabolic products of 5-ALA generated in HSFs, the cells were co-cultured with different materials in the dark for 4 h, then irradiated with a 633 nm laser for 10 min, and subsequently incubated at 37 °C for 24 h. For heme assessment, the cells were gently scraped and subjected to ultrasonication. The cell suspension was then centrifuged at 11,180 g for 10 min to collect the supernatant, and the heme content was analyzed using an ELISA kit. Simultaneously, the cells were stained with COP-1 (Ruixi Biotechnology, China) for CO detection and FerroOrange (Beiren Chemical Technology, Beijing) for Fe2+ detection. These metabolic products were then examined using a fluorescence microscope, allowing for the measurement of fluorescence signals for CO, Fe2+, and biliverdin (autofluorescence). Molecular docking of baicalin with HO-1 was performed using Auto Dock Vina software (Version 1.2.0).
In vitro cytotoxicity assay
In vitro cytotoxicity was assessed using the Cell Counting Kit-8 (CCK-8). In brief, HSFs were seeded at a density of 5 × 103 cells/well in a 96-well plate and cultured for 24 h. Subsequently, these cells were exposed to various experimental materials for 4 h, followed by laser irradiation at 633 nm (40 mW/cm², LWRPD630, China) for 10 min. After an additional 24-h incubation period, a diluted CCK-8 solution was administered to each well and incubated at 37 °C for 4 h. The absorbance at 450 nm was then measured using a microplate reader. Similarly, the treated HSFs underwent Annexin V-FITC/PI staining following the supplied protocol and were then analyzed with flow cytometry.
Ferroptosis evaluation of FAB@MN
HSFs were seeded at a density of 5 × 104 cells/well in 24-well plates and cultured overnight. Subsequently, these cells were exposed to various experimental materials for 4 h, followed by laser irradiation for 10 min. After an additional 24-h incubation, the HSFs were stained with C11-BODIPY 581/591 for LPO analysis for 30 min, and analyzed using a fluorescence microscope and flow cytometry. In addition, the LIP, LPO, MDA and GSH amounts were monitored using an LIP, LPO, MDA and GSH assay kit according to the instructions. To assess the mitochondrial structure and MMP of HSFs, JC-1, TMRM and Mito-Tracker probes were incubated with cells for 30 min respectively, and the intracellular fluorescence was evaluated with CLSM or flow cytometry. For a detailed examination of the mitochondrial ultrastructure, the treated cells were fixed with 2.5% glutaraldehyde and observed using biological transmission electron microscopy (Bio-TEM, Tokyo, Japan). Mitophagy was evaluated using a dual staining method with Lyso-Tracker Green and Mito-Tracker Deep Red. Specifically, the treated HSFs were first stained with Mito-Tracker Deep Red at 37 °C for 30 min, then incubated with Lyso-Tracker Green for another 30 min, and finally stained with Hoechst 33342 for 10 min to label the cell nuclei.
Western blot analysis
Cells were collected using a cell scraper and transferred into lysis buffer containing RIPA: PMSF: phosphatase inhibitors: protease inhibitors at a ratio of 100:1:2:2. The mixture was then sonicated and centrifuged at 14,000 g for 15 min at 4 °C, and the protein concentration was measured using a BCA assay kit. Depending on the molecular weight of the proteins, appropriate Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE) gels were selected for protein separation, and the proteins were transferred to a polyvinylidene fluoride (PVDF) membrane. The membrane was incubated overnight with specific antibodies, followed by incubation with horseradish peroxidase (HRP)-conjugated secondary antibodies (1:2000, Zhongshan Golden Bridge, China). At last, the membranes were analyzed using an image reader (LAS-4000; Fujifilm, Japan). All uncropped scans of blot presented in the main text have been included in the Source Data file, and those related to the Supplementary Figs. have been provided in Supplementary Information.
In vivo anti-scarring efficacy of FAB@MN
The HS model was established using a previously detailed method. Under anesthesia, a 1 cm diameter full-thickness wound was created on the ventral side near the proximal end of the rabbit’s ear, with careful removal of the perichondrium to avoid damaging the cartilage beneath. Four weeks post-surgery, a firm and dense HS had formed at the wound sites, and these scars were randomly divided into six groups: Control group, FA@MN group, FB@MN group, FAB@MN group, TA-TRANSTON® group and CB-DIPROSPAN® group. Four hours post MN insertion, the treated regions were exposed to laser irradiation for 10 min, and this process was repeated weekly for a total of four weeks. The scar morphology within each group was captured and recorded using a digital camera. Upon completion of the treatment, ultrasound imaging of the scars was conducted using the Aixplorer ultrasound system (Super-Sonic Imagine, France), enabling direct measurement of scar thickness. Specifically, ultrasound transmission gel was uniformly applied over the scarred region, and the scars were analyzed with an ultrasonic probe. Scar hardness and color were measured using a Shore durometer (HT-65100, Lantai Instruments Co., Ltd, China) and a 3nh spectrophotometer (three Enshi technology Co., Ltd, Guangdong, China). Following these procedures, the rabbits were sacrificed and HS tissues were collected for histological analysis through H&E, Masson and Sirius Red staining. Additionally, immunohistochemical staining with GPX4 antibody was performed. For ROS assays, 10 µL of a 10 mM DCFH-DA solution was precisely injected into the rabbit ear scar tissue before treatment. Upon completing the treatment, detailed imaging of the scar areas was performed using the animal in vivo imaging system (IVIS). Meanwhile, the scar tissue samples were harvested for fluorescence analysis.
Statistical analysis
All analyses and comparisons were performed using SPSS 20.0 software (IBM, USA). Results were presented as mean ± SD. The two-sided t-test was employed for comparisons between two groups, while one-way ANOVA was used for analyzing the variance among multiple groups. p-value < 0.05 indicated a significant difference between groups.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data generated this study are provided in the main text and Supplementary Information. The transcriptomic data generated in this study have been deposited in the NCBI Gene Expression Omnibus (GEO) under accession code GSE300586: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE300586. Source data are provided with this paper.
References
Yang, Z. R. et al. Endogenous stimuli-responsive separating microneedles to inhibit hypertrophic scar through remodeling the pathological microenvironment. Nat. Commun. 15, 2038 (2024).
Grabowski, G., Pacana, M. J. & Chen, E. Keloid and hypertrophic scar formation, prevention, and management: standard review of abnormal scarring in orthopaedic surgery. J. Am. Acad. Orthop. Surg. 28, e408–e414 (2020).
Oliveira, G. V. et al. Treatment of keloids and hypertrophic scars. Position statement of the Brazilian expert group GREMCIQ. J. Eur. Acad. Dermatol Venereol. 35, 2128–2142 (2021).
Li, Y. et al. Liposomes loaded with 5-fluorouracil can improve the efficacy in pathological scars. Int J. Nanomed. 19, 7353–7365 (2024).
Jeschke, M. G. et al. Scars. Nat. Rev. Dis. Prim. 9, 64 (2023).
Li, C. et al. ADSCC-CM-induced keratin hydrogel-based bioactive microneedle patch containing triamcinolone acetonide for the treatment of pathological scar. Adv. Funct. Mater. 34, 2400457 (2024).
Frech, F. S. et al. Hypertrophic scars and keloids: advances in treatment and review of established therapies. Am. J. Clin. Dermatol 24, 225–245 (2023).
Chen, Z. et al. Layered GelMA/PEGDA hydrogel microneedle patch as an intradermal delivery system for hypertrophic scar treatment. ACS Appl Mater. Interfaces 15, 43309–43320 (2023).
Huang, Y. et al. Fully armed photodynamic therapy with spear and shear for topical deep hypertrophic scar treatment. J. Control Release 343, 408–419 (2022).
Campbell, N. K., Fitzgerald, H. K. & Dunne, A. Regulation of inflammation by the antioxidant haem oxygenase 1. Nat. Rev. Immunol. 21, 411–425 (2021).
Kumar, S. & Bandyopadhyay, U. Free heme toxicity and its detoxification systems in human. Toxicol. Lett. 157, 175–188 (2005).
Li, A. et al. Boosting 5-ALA-based photodynamic therapy by a liposomal nanomedicine through intracellular iron ion regulation. Acta Pharm. Sin. B. 11, 1329–1340 (2021).
Chen, Y. et al. Desferrioxamine enhances 5-aminolaevulinic acid-induced protoporphyrin IX accumulation and therapeutic efficacy for hypertrophic scar. J. Pharm. Sci. 112, 1635–1643 (2023).
Howley, R. et al. Analysis of renal cell carcinoma cell response to the enhancement of 5-aminolevulinic acid-mediated protoporphyrin IX fluorescence by iron chelator deferoxamine. Photochem Photobio. 99, 787–792 (2023).
Luo, P. et al. Celastrol induces ferroptosis in activated HSCs to ameliorate hepatic fibrosis via targeting peroxiredoxins and HO-1. Acta Pharm. Sin. B 12, 2300–2314 (2022).
Menon, A. V. et al. Excess heme upregulates heme oxygenase 1 and promotes cardiac ferroptosis in mice with sickle cell disease. Blood 139, 936–941 (2022).
Jiang, X., Stockwell, B. R. & Conrad, M. Ferroptosis: mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 22, 266–282 (2021).
Fang, F. et al. Continuous spatiotemporal therapy of a full-API nanodrug via multi-step tandem endogenous biosynthesis. Nat. Commun. 14, 1660 (2023).
Zhao, B. et al. Ferroptosis-mediated synergistic therapy of hypertrophic scarring based on metal–organic framework microneedle patch. Adv. Funct. Mater. 33, 2300575 (2023).
Guo, L. T. et al. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J. Neuroinflammation 16, 95 (2019).
Singh, S., Meena, A. & Luqman, S. Baicalin mediated regulation of key signaling pathways in cancer. Pharm. Res 164, 105387 (2021).
Zhao, Q. Y. et al. Baicalin inhibits Escherichia coli isolates in bovine mastitic milk and reduces antimicrobial resistance. J. Dairy Sci. 101, 2415–2422 (2018).
Hu, Q. et al. Baicalin and the liver-gut system: pharmacological bases explaining its therapeutic effects. Pharm. Res. 165, 105444 (2021).
Xu, L. et al. pH-responsive hydrogel as a potential oral delivery system of baicalin for prolonging gastroprotective activity. Pharmaceutics 15, 257 (2023).
Wen, R. J. et al. Baicalin induces ferroptosis in osteosarcomas through a novel Nrf2/xCT/GPX4 regulatory axis. Phytomedicine 116, 154881 (2023).
Kong, N. et al. Baicalin induces ferroptosis in bladder cancer cells by downregulating FTH1. Acta Pharm. Sin. B 11, 4045–4054 (2021).
Yuan, J. et al. Baicalin enhances the efficacy of 5-Fluorouracil in gastric cancer by promoting ROS-mediated ferroptosis. Biomed. Pharmacother. 164, 114986 (2023).
Wen, Z. et al. Baicalin induces ferroptosis in oral squamous cell carcinoma by suppressing the activity of FTH1. J. Gene Med 26, e3669 (2024).
Zhang, L. et al. Preparation and characterisation of baicalin magnesium and its protective effect in ulcerative colitis via gut microbiota-bile acid axis modulation. Phytomedicine 126, 155416 (2024).
Yan, Y. et al. Endogenous crude Scutellaria baicalensis polysaccharide robustly enhances one-pot extraction and deglycosylation of baicalin. Int J. Biol. Macromol. 263, 130349 (2024).
Qi, D. et al. Baicalin/ambroxol hydrochloride combined dry powder inhalation formulation targeting lung delivery for treatment of idiopathic pulmonary fibrosis: Fabrication, characterization, pharmacokinetics, and pharmacodynamics. Eur. J. Pharm. Biopharm. 188, 243–253 (2023).
Li, C. et al. Baicalin nanocomplexes with an in situ-forming biomimetic gel implant for repair of calvarial bone defects via localized sclerostin inhibition. ACS Appl Mater. Interfaces 15, 9044–9057 (2023).
Wang, Z. R. et al. Bioengineered dual-targeting protein nanocage for stereoscopically loading of synergistic hydrophilic/hydrophobic drugs to enhance anticancer efficacy. Adv. Funct. Mater. 31, 2102004 (2021).
Wang, C. et al. Ferritin-based targeted delivery of arsenic to diverse leukaemia types confers strong anti-leukaemia therapeutic effects. Nat. Nanotechnol. 16, 1413–1423 (2021).
Wang, X. et al. A protein-based cGAS-STING nanoagonist enhances T cell-mediated anti-tumor immune responses. Nat. Commun. 13, 5685 (2022).
Pang, J. et al. Ferritin-nanocaged ATP traverses the blood-testis barrier and enhances sperm motility in an asthenozoospermia model. ACS nano 16, 4175–4185 (2022).
Mi, X. et al. Folic acid decorated zeolitic imidazolate framework (ZIF-8) loaded with baicalin as a nano-drug delivery system for breast cancer therapy. Int J. Nanomed. 16, 8337–8352 (2021).
Dong, Y. et al. ERK-peptide-inhibitor-modified ferritin enhanced the therapeutic effects of paclitaxel in cancer cells and spheroids. Mol. Pharm. 18, 3365–3377 (2021).
Gu, C. et al. His-mediated reversible self-assembly of ferritin nanocages through two different switches for encapsulation of cargo molecules. ACS nano 14, 17080–17090 (2020).
Sevieri, M. et al. Deciphering the role of H-ferritin nanocages in improving tumor-targeted delivery of indocyanine green: combined analysis of murine tissue homogenates with UHPLC-MS/MS and fluorescence. ACS omega 8, 48735–48741 (2023).
Zhang, J. et al. H-ferritin-nanocaged gadolinium nanoparticles for ultra-sensitive MR molecular imaging. Theranostics 14, 1956–1965 (2024).
Jia, W. et al. Brain-targeted HFn-Cu-REGO nanoplatform for site-specific delivery and manipulation of autophagy and cuproptosis in glioblastoma. Small 19, e2205354 (2023).
Chen, Y. et al. Functional transdermal nanoethosomes enhance photodynamic therapy of hypertrophic scars via self-generating oxygen. ACS Appl Mater. Interfaces 13, 7955–7965 (2021).
Horii, S. et al. 5-aminolevrinic acid exhibits dual effects on stemness in human sarcoma cell lines under dark conditions. Int J. Mol. Sci. 24, 6189 (2023).
Hou, J. et al. 5-Aminolevulinic acid with ferrous iron induces permanent cardiac allograft acceptance in mice via induction of regulatory cells. J. Heart Lung Transpl. 34, 254–263 (2015).
Zhu, L. et al. Self-catalyzed nitric oxide nanocomplexes induce ferroptosis for cancer immunotherapy. J. Control Release 377, 524–539 (2024).
Li, R. et al. Heme oxygenase 1-mediated ferroptosis in Kupffer cells initiates liver injury during heat stroke. Acta Pharm. Sin. B 14, 3983–4000 (2024).
Zeng, Y. L. et al. Iridium(III) photosensitizers induce simultaneous pyroptosis and ferroptosis for multi-network synergistic tumor immunotherapy. Angew. Chem. Int Ed. Engl. 63, e202410803 (2024).
Sansaloni-Pastor, S., Varesio, E. & Lange, N. Modulation and proteomic changes on the heme pathway following treatment with 5-aminolevulinic acid. J. Photochem Photobio. B 233, 112484 (2022).
Nakayama, T. et al. Photoirradiation after aminolevulinic acid treatment suppresses cancer cell proliferation through the HO-1/p21 pathway. Photodiagnosis Photodyn. Ther. 28, 10–17 (2019).
Chiang, S. K., Chen, S. E. & Chang, L. C. A dual role of heme oxygenase-1 in cancer cells. Int J. Mol. Sci. 20, 39 (2018).
Mei, J. et al. Production of bilirubin by biotransformation of biliverdin using recombinant Escherichia coli cells. Bioprocess Biosyst. Eng. 45, 563–571 (2022).
Cho, H. R. et al. Malignant glioma: MR imaging by using 5-aminolevulinic acid in an animal model. Radiology 272, 720–730 (2014).
Li, J. et al. Effects of feeding 5-aminolevulinic acid on iron status in weaned rats from the female rats during gestation and lactation. Anim. (Basel) 12, 2869 (2022).
Ueda, S. et al. Baicalin induces apoptosis via mitochondrial pathway as prooxidant. Mol. Immunol. 8, 781–791 (2002).
Lin, M. Y. et al. Baicalin enhances chemosensitivity to doxorubicin in breast cancer cells via upregulation of oxidative stress-mediated mitochondria-dependent apoptosis. Antioxid. (Basel) 10, 1506 (2021).
Lan, M. et al. Activating caspase-8/Bid/ROS signaling to promote apoptosis of breast cancer cells by folate-modified albumin baicalin-loaded nanoparticles. Nanotechnology. 33 https://doi.org/10.1088/1361-6528/ac197b (2022).
Liang, C. et al. Recent progress in ferroptosis inducers for cancer therapy. Adv. Mater. 31, e1904197 (2019).
Sun, S. et al. Targeting ferroptosis opens new avenues for the development of novel therapeutics. Signal Transduct. Target Ther. 8, 372 (2023).
Miyamoto, H. D. et al. Iron overload via heme degradation in the endoplasmic reticulum triggers ferroptosis in myocardial ischemia-reperfusion injury. JACC Basic Transl. Sci. 7, 800–819 (2022).
Cao, W. et al. Exogenous carbon monoxide promotes GPX4-dependent ferroptosis through ROS/GSK3β axis in non-small cell lung cancer. Cell Death Discov. 10, 42 (2024).
Yao, X. et al. Multifunctional nanoplatform for photoacoustic imaging-guided combined therapy enhanced by CO induced ferroptosis. Biomaterials 197, 268–283 (2019).
Shao, L. et al. Baicalin enhances the chemotherapy sensitivity of oxaliplatin-resistant gastric cancer cells by activating p53-mediated ferroptosis. Sci. Rep. 14, 10745 (2024).
Han, S. et al. HO-1 contributes to luteolin-triggered ferroptosis in clear cell renal cell carcinoma via increasing the labile iron pool and promoting lipid peroxidation. Oxid. Med Cell Longev. 2022, 3846217 (2022).
Guo, Z. et al. High expression levels of haem oxygenase-1 promote ferroptosis in macrophage-derived foam cells and exacerbate plaque instability. Redox Biol. 76, 103345 (2024).
Li, X. et al. HO-1 upregulation promotes mitophagy-dependent ferroptosis in PM2.5-exposed hippocampal neurons. Ecotoxicol. Environ. Saf. 277, 116314 (2024).
Li, T. PM2.5-induced iron homeostasis imbalance triggers cardiac hypertrophy through ferroptosis in a selective autophagy crosstalk manner. Redox Biol. 72, 103158 (2024).
Yang, P. et al. Ionizing radiation-induced mitophagy promotes ferroptosis by increasing intracellular free fatty acids. Cell Death Differ. 30, 2432–2445 (2023).
Li, J. et al. β-amyloid protein induces mitophagy-dependent ferroptosis through the CD36/PINK/PARKIN pathway leading to blood-brain barrier destruction in Alzheimer’s disease. Cell Biosci. 12, 69 (2022).
Yu, F. et al. Dynamic O-GlcNAcylation coordinates ferritinophagy and mitophagy to activate ferroptosis. Cell Discov. 8, 40 (2022).
Li, J. et al. PINK1/Parkin-mediated mitophagy in neurodegenerative diseases. Ageing Res Rev. 84, 101817 (2023).
Zheng, Y. et al. STOML2 potentiates metastasis of hepatocellular carcinoma by promoting PINK1-mediated mitophagy and regulates sensitivity to lenvatinib. J. Hematol. Oncol. 14, 16 (2021).
Kumar, A. V., Mills, J. & Lapierre, L. R. Selective autophagy receptor p62/SQSTM1, a pivotal player in stress and aging. Front Cell Dev. Biol. 14, 793328 (2022).
Doric, Z. & Nakamura, K. Mice with disrupted mitochondria used to model Parkinson’s disease. Nature 599, 558–560 (2021).
Atiyeh, B. S. Nonsurgical management of hypertrophic scars: evidence-based therapies, standard practices, and emerging methods. Aesthetic Plast. Surg. 44, 1320–1344 (2020).
Mokos, Z. B. et al. Current therapeutic approach to hypertrophic scars. Front Med (Lausanne) 4, 83 (2017).
Xu, Y. et al. Single-dose of integrated bilayer microneedles for enhanced hypertrophic scar therapy with rapid anti-inflammatory and sustained inhibition of myofibroblasts. Biomaterials 312, 122742 (2025).
Zhao, B. et al. Supramolecular assembly-enabled transdermal therapy of hypertrophic scarring through concurrent ferroptosis-apoptosis. Adv. Funct. Mater. 2416011 (2024).
Acknowledgments
This work was supported by the National Science Fund for Excellent Young Scholars (32322044, C.T.), National Natural Science Foundation of China (32071362, C.T.), Key International (Regional) Joint Research Program funded by National Natural Science Foundation of China (NSFC) (82220108019, J.P.), the Science Fund for Distinguished Young Scholars of Chongqing (CSTB2022NSCQ-JQX0012, C.T.), the CQMU Program for Youth Innovation in FutureMedicine (W0077, C.T.).
Author information
Authors and Affiliations
Contributions
Y.C. conceived the idea and designed the experiments; Y.C., S.W., C.X.M., Q.Y.L., and D.Q.F. prepared and characterized the materials; Y.C., S.W., Y.P.L., X.C., J.L.Z., and Z.X.Y performed in vitro and in vivo experiments; Y.C., S.W., and X.Y.Z. contributed to analysis and discussion of the results; Y.C. and S.W. wrote the paper; Q.Q.H., P.J., and T.C. revised the manuscript; T.C. supervised the overall study.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Salvador Gonzalez, Cheng Zhan, who co-reviewed with Guoshu Bi, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Chen, Y., Wang, S., Mao, C. et al. 5-ALA photodynamic metabolite-powered zero-waste ferroptosis amplifier for enhanced hypertrophic scar therapy. Nat Commun 16, 8321 (2025). https://doi.org/10.1038/s41467-025-63438-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-63438-7
