
Abstract
Abdominal aortic aneurysm (AAA) and the attendant catastrophic event of rupture remain a leading cause of death. Germline mutation of RRAS2 cause Noonan Syndrome, yet little is known about its function in AAA pathogenesis. Integrated analysis of RNA-seq data revealed RRAS2 a potential regulator of AAA. Its mRNA and protein levels reduced in abdominal aorta of AAA patients and AAA mice. Mice with SMCs-specific knockout of Rras2 were more vulnerable to Ang II and porcine pancreatic elastase-induced AAA, while overexpression of Rras2 in SMCs inhibited AAA progression. Maternally expressed gene 3 (MEG3) prevented RRAS2 mRNA degradation through binding with ELAV-like protein 1 (ELAVL1/HuR). Mechanistically, loss of R-Ras2 reduces phosphorylation, nuclear translocation, and transcriptional activity of general transcription factor 2I (GTF2-I/TFII-I) to reduce contractile-related genes expression in a MET tyrosine kinase-dependent manner. Here, we show an essential role of R-Ras2 in preserving VSMCs homeostasis and provide potential therapeutic targets for AAA.
Similar content being viewed by others
Introduction
Abdominal aortic aneurysm (AAA) causes 1.3% of all deaths among men aged 65–85 years in developed countries and is always lethal when ruptured if leaving untreated1. Aortic aneurysm is characterized by a profound loss of integrity of the medial layer, including extracellular matrix degradation and SMC rarefaction due to apoptosis2,3. During AAA, vascular smooth muscle cells (VSMCs) lost contractile-related phenotype, with reduced levels of actin alpha 2, smooth muscle (ACTA2), transgelin (TAGLN), and calponin 1 (CNN1)4,5, and obtain ability to secrete matrix metalloproteinases to degrade the extracellular matrix and weaken the arterial wall, which potentiate the rupture of the aneurysm3.
Ras-related protein 2 (R-Ras2) is a small GTPase of the Ras subfamily. RRAS2 mutations cause Noonan Syndrome, a developmental disorder featured with craniofacial appearance, short stature, variable cognitive impairment, predisposition to malignancies, and a wide spectrum of cardiac defects6,7,8. Similar with other oncogenic Ras family member, constitutive active form of R-Ras2 activates MAP kinase and PI3K-Akt cascades9. Recent studies found important roles of R-Ras2 in platelet activation and thrombus stability10. Besides, R-Ras2 is a key regulator of autoimmunity, and enhanced negative selection during T cell development was observed in R-Ras2 knockout mice11. However, the role of R-Ras2 in VSMCs homeostasis and AAA remains unelusive. Previous study found that H-Ras, another member of Ras subfamily, could mediate VSMCs proliferation in carotid and coronary artery injuries12,13,14. In the current study, we sought to investigate whether R-Ras2 is also involved in regulating expressions of contractile-related proteins in VSMCs and, therefore, modulating the progression of AAA.
In this work, we report that loss of vascular smooth muscle cell (VSMC)-derived R-Ras2 reduced the phosphorylation and nuclear translocation of general transcription factor 2I (GTF2I/TFII-I), inhibited contractile-related protein expression, and promoted AAA formation. Maternally expressed gene 3 (MEG3) is identified as an upstream regulator of R-Ras2, it mediates the binding of ELAVL 1 with RRAS2 to prevent RRAS2 degradation. As expected, overexpression of MEG3 alleviates AAA.
Results
R-Ras2 reduces in VSMCs during AAA
To investigate genes potentially regulating VSCMs phenotypic transition and aortic aneurysm, we took advantage of the Gene Expression Omnibus database to gather common differentially expressed genes (DEGs) in Human aortic aneurysm compared with normal individuals (GSE140947), and ApoE−/− mice infused with angiotensin II (Ang II) compared with saline ones (GSE12591). Next, to identify the candidate genes potentially related to VSMCs phenotypic transition, we challenged human aortic smooth muscle cells (HASMCs) with Ang II and conducted RNA-seq to identify twenty DEGs (Fig. 1a). Among them, RAS-like family 11 member B (RAS11LB) and RAS-related 2 (RRAS2) were two most downregulated genes with related molecular functions, and emerged as main candidate genes linking VSMCs phenotypic switch to aortic aneurysm (Fig. 1b). Q-PCR demonstrated that the expression level of RRAS2 is much higher than RAS11LB in HASMCs, although both RASL11B and RRAS2 reduced with Ang II stimulation (Supplementary Fig. 1a, b). In addition, knockdown of RRAS2, rather than RAS11LB, could significantly reduce the expression of contractile-related markers, including Calponin 1, α-SMA, and SM22α (Fig. 1c and Supplementary Fig. 1c–e). Besides, the gene expression of OPN, MMP2, and MMP9 were increased in HASMCs lacking RRAS2 (Fig. 1d), confirming the critical role of RRAS2 in maintaining the contractile phenotype of VSMCs.

a Venn diagram showing the overlapping differentially expressed genes (DEGs) for abdominal aortic aneurysm (AAA) patients (GSE140947), AAA mouse (GSE12591), and HASMCs stimulated with Ang II. b Heatmap of the relative average expression of the 20 overlapped genes in (a). c Representative western blotting and quantification of α-SMA, Calponin 1, and SM22 in human aortic smooth muscle cells (HASMCs) transfected with siRNA targeting RRAS2. n = 6 distinct samples for each group, two-sided Unpaired t-test. d RRAS2, OPN, MMP2, and MMP9 expression in HASMCs transfected with siRNA targeting RRAS2. n = 6 distinct samples for each group, two-sided Unpaired t-test. e Representative western blotting and quantification of R-Ras2 expression in Sham and Ang II-induced AAA mice. n = 6 mice for each group, two-sided Unpaired t-test. f Representative immunofluorescent staining of aorta from control (Sham) and AAA mice for α-SMA (Green) and R-Ras2 (Red), IgG was used as the isotype control. Blue: DAPI. Scale bar: 40 μm. 3 independently experiments were repeated with similar results. g Representative western blotting and quantification of R-Ras2 expression in aortic specimens of controls (non-AAA) (n = 6) and AAA patients (n = 6), two-sided Unpaired t-test. h Immunofluorescent staining of AAA specimen for α-SMA (Green) and R-Ras2 (Red), IgG was used as the isotype control. Blue: DAPI. Scale bar: 20 μm. 3 independently experiments were repeated with similar results. i Western blotting analysis of α-SMA, Calponin 1, and R-Ras2 expression in HASMCs overexpressing R-Ras2 and incubated with Ang II. n = 6 distinct samples for each group, Two-way ANOVA test followed by Turkey’s multiple comparisons test. All data were represented as mean ± SEM. Source data are provided as a Source Data file.
We isolated mouse aortic smooth muscle cells (MASMCs), endothelial cells (HAECs), and bone marrow-derived macrophages (BMDMs) and compared the level of R-Ras2 in these cells, a higher level of R-Ras2 was observed in HASMCs compared with HAECs, and R-Ras2 expression was hardly detected in BMDMs (Supplementary Fig. 2a). Besides, Ang II treatment significantly inhibited R-Ras2 expression in HASMCs rather than in HAECs (Supplementary Fig. 2b, c). We then compared the mRNA level of ras homologous, including HRAS, KRAS, NRAS, RRAS1, RRAS2, and RRAS3 in HASMCs, and found higher levels of RRAS2 and RRAS1 in HASMCs (Supplementary Fig. 2d). The finding is in accordance with a previous study that R-Ras1 is mainly expressed in SMC15. We then detected whether Ang II treatment could regulate the expression of R-Ras1, and found that the protein level of R-Ras1 was unchanged with Ang II treatment (Supplementary Fig. 2e), indicating that R-Ras2 rather than R-Ras1 is involved in Ang II-induced VSMCs phenotypic transition. As expected, reduction of mRNA and protein levels of Rras2 were observed in aortic tissue of ApoE−/− mice infused with Ang II (Fig. 1e and Supplementary Fig. 1f). Co-immunofluorescent staining of α-SMA and R-Ras2 confirmed the reduction of R-Ras2 in the tunica media of abdominal aorta of AAA mice (Fig. 1f). More importantly, we found that R-Ras2 mRNA and protein abundance are lower in the human AAA lesions than in non-AAA aorta (Fig. 1g and Supplementary Fig. 1g). Co-immunofluorescent staining of R-Ras2 and α-SMA confirmed a reduction of R-Ras2 in the tunica media of abdominal aorta in AAA patients (Fig. 1h). Collectively, these results indicate the potential involvement of R-Ras2 in the progression of VSMCs phenotypic transition and AAA.
Key features of AAA includes increased migration, proliferation, and loss of VSMCs in the media layer along with AAA progress16. Knockdown of RRAS2 significantly increased the migration and proliferation ability of HASMCs, as determined by wound healing and transwell assays, and in situ zymography assay demonstrated that knockdown of RRAS2 increased MMPs activity (Supplementary Fig. 2f–i). We further determined whether knockdown of R-Ras2 increased HASMCs apoptosis upon TNF-α + CHX stimulation, and found that knockdown of RRAS2 made HASMCs more vulnerable to TNF-α + CHX stimulation, evidenced by increased level of cleaved-PARP1 (C-PARP1), Bax and reduced Bcl2 expression (Supplementary Fig. 2j). Overexpression of R-Ras2 restored the protein levels of α-SMA, Calponin 1 and SM22 in HASMCs stimulated with Ang II (Fig. 1i). Consistently, overexpression of R-Ras2 prevented excessive proliferation and migration of VSMCs induced by Ang II and inhibited MMPs activation (Supplementary Fig. 2k–n). Collectively, these results suggest that R-Ras2 is critical for maintaining VSMCs homeostasis.
SMCs-specific knockout of Rras2 aggravates AAA in mice
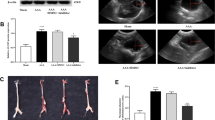
To investigate the role of R-Ras2 in the progression of AAA, we constructed SMCs-specific Rras2 knockout mice (Rras2 SMKO) by crossing Rras2 flox/flox (Rras2 f/f) mice with TaglnCre mice (without ApoE−/− background). The elimination of R-Ras2 in SMCs was determined by co-immunofluorescent staining of α-SMA and R-Ras2 (Supplementary Fig. 3a). Ang II infusion increased systolic blood pressure (SBP) in both genotypes (Supplementary Fig. 3b). Ang II treatment induced AAA in only 2 of the 20 Rras2 f/f mice, which is similar with a previous study17. However, Ang II treatment triggered AAA or lethal aneurysm dissection in 14 of the 20 Rras2 SMKO mice (Fig. 2a, b). The suprarenal maximal aortic lumen diameter is larger in Rras2 SMKO mice compared with Rras2f/f mice treated with Ang II (Fig. 2c). The aortic weight to body weight ratio was also increased in Rras2 SMKO mice with Ang II infusion (Fig. 2d). According to intravascular ultrasound imaging, diameters of abdominal aorta were significantly increased in Rras2 SMKO mice infused with Ang II (Fig. 2e). H&E staining revealed significant increase in aortic medial thickness, and Masson’s trichrome staining revealed increased deposition of collagen in Rras2 SMKO mice compared with those in Rras2 f/f mice infused with Ang II. In addition, EVG staining revealed more severe fragmentation of elastic fibers in Rras2 SMKO mice compared with Rras2 f/f littermates challenged with Ang II (Fig. 2f). We also found significantly reduced protein levels of α-SMA, Calponin 1 and SM22α in Rras2 SMKO mice compared with Rras2 f/f littermates upon Ang II treatment (Fig. 2g). Co-immunofluorescent staining of α-SMA and Calponin 1 accordingly revealed reduction of Calponin 1 in SMCs (Fig. 2h). Accordingly, Bax and C-PARP1 levels were increased in Rras2 SMKO mice treated with Ang II, while Bcl2 level in Rras2SMKO mice were reduced with Ang II infusion (Fig. 2i). This observation was further confirmed by TUNEL staining, where TUNEL-positive SMCs were increased with Ang II treatment (Fig. 2j). Finally, in situ zymography found increased activity of MMPs in the suprarenal aorta specimens of Rras2SMKO mice infused with Ang II (Supplementary Fig. 3c).

a Representative photograph of aortas isolated from Rras2f/f or Rras2SMKO mice infused with Ang II (1000 ng/kg/min) for 4 weeks. b Incidence of AAA, two-sided Fisher’s exact test. c Quantification of the suprarenal vascular outer maximal width, n = 20 mice for each group, two-sided Mann–Whitney test. d The ratio of the aorta weight to body weight. Rras2f/f + Ang II (n = 20) and Rras2SMKO + Ang II (n = 17). Two-sided Mann–Whitney test. e Ultrasound images and quantification of the maximal lumen abdominal aorta diameters of Rras2f/f + Ang II (n = 20) and Rras2SMKO + Ang II (n = 17) mice. Two-way RM ANOVA followed by Bonferroni’s multiple comparisons test. f H&E, Masson, and EVG staining of supra-renal abdominal aorta sections, and quantifications of collagen content and elastic fiber breaks. n = 6 mice for each group. Masson, two-sided Unpaired t-test with Welch’s correction. EVG, two-sided Mann–Whitney test. g Western blot of SM22, Calponin 1 and α-SMA. n = 6 mice for each group, two-sided Unpaired t-test. h Co-immunofluorescent staining of α-SMA (Green) and R-Ras2 (Red), IgG was used as the isotype control. Blue: DAPI. Scale bar: 40 μm. 3 independent experiments showed consistent results. i Western blot of Bax, Cleaved-PARP1 (C-PARP1), and Bcl-2 in aortic tissue of different treated mice, n = 6 mice for each group, two-sided Unpaired t-test. j TUNEL staining for apoptotic cells (Green). Red: α-SMA, Blue: DAPI. 3 independent experiments showed consistent results. Scale bar: 40 μm. Collagen deposition in (f), and data in (c, e, g, and i) were represented as mean ± SEM. Data in (d) and EVG in (f) were represented as the box plots indicating median (middle line), 25th, 75th percentile (box), as well as minimum and maximum values (whiskers). Source data are provided as a Source Data file.
The role of SMC-Rras2 in aortic aneurysm was further confirmed in the PPE-induced AAA model. Compared with Rras2f/f littermates challenged with PPE, Rras2SMKO mice exhibited increased infrarenal artery outer diameter and aortic weight to body weight (Supplementary Fig. 3d–f), as well as more progressive elastin degradation after 14 days of PPE incubation (Supplementary Fig. 3g). Consistently, the expression of R-Ras2 was reduced in tunica media of suprarenal abdominal aorta in Rras2SMKO mice compared with Rras2f/f littermates incubated with PPE (Supplementary Fig. 3h). We found that loss of Rras2 in SMCs of female mice also significantly aggravated aortic aneurysm formation (Fig. S4). These data collectively demonstrate that loss of Rras2 in SMCs could accelerate the progression of AAA.
Overexpression of Rras2 in SMCs alleviates AAA
We next overexpressed Rras2 in SMCs via cloning the expression cassette of Flag-Rras2 in an inverse, anti-sense orientation between two pairs of loxP sites into lentivirus vector, and infected ApoE−/−TaglnCre mice. Cre recombinase expression in SMCs mediated the inversion of Flag-Rras2 into a sense orientation and promoted R-Ras2 expression in SMCs (hereafter referred to as Lenti-Rras2) (Supplementary Fig. 5a). Co-immunofluorescent staining of Flag and α-SMA confirmed overexpression of R-Ras2 in the medial layer of aorta (Supplementary Fig. 5b). AAA was then induced in mice with subcutaneous infusion of Ang II for 4 weeks according to the experimental design shown in Supplementary Fig. 5c. Although Ang II infusion increased SBP both in Lenti-Rras2 and in Lenti-Vector mice (Supplementary Fig. 5d), overexpression of R-Ras2 in SMCs reduced the occurrence of aortic aneurysm or lethal aneurysm dissection (Fig. 3a, b). Besides, overexpression of R-Ras2 in SMCs inhibited abdominal aortic dilatation and the elevation of aortic weight to body weight in mice challenged with Ang II (Fig. 3c, d). The dilatation of ascending aorta, rather than descending aorta, thoracic aorta, and aortic arch was prevented with R-Ras2 overexpression (Supplementary Fig. 5e–h). The protein levels of α-SMA, Calponin 1 and SM22 were reduced significantly in aorta of Lenti-Vector mice infused with Ang II, and overexpression of R-Ras2 in SMCs restored their expression (Fig. 3e). Similarly, immunofluorescent staining revealed that overexpression of R-Ras2 in SMCs restored the level of Calponin 1 (Fig. 3f). Overexpression of R-Ras2 in SMCs could also reduce the expression of Bax and C-PARP1, and increase Bcl2 level (Fig. 3g). Histological staining revealed that overexpression of R-Ras2 in SMCs alleviated elastic fiber fragmentation and collagen deposition and reduced MMPs activity (Fig. 3h, i).

a Representative photograph of aortas. b Incidence of AAA. Lenti-Vector+ Ang II (n = 13) and Lenti-Rras2+ Ang II (n = 14), two-sided Fisher’s exact test. c Quantification of the maximal abdominal aorta lumen diameters. Lenti-Vector+ Saline (n = 12), Lenti-Vector + Ang II (n = 10), Lenti-Rras2+ Saline (n = 12) and Lenti-Rras2+ Ang II (n = 12). Two-way RM ANOVA followed by Bonferroni’s multiple comparisons test. d Ratio of aortic weight to body weight. Lenti-Vector+ Saline (n = 12), Lenti-Vector+ Ang II (n = 11), Lenti-Rras2+ Saline (n = 12) and Lenti-Rras2+ Ang II (n = 12). Two-way ANOVA test followed by Turkey’s multiple comparisons test. e Western blot and quantification of α-SMA, Calponin 1, and SM22 in aortic tissue of mice. n = 6 mice for each group. Two-way ANOVA with Turkey’s multiple comparisons test. f Immunofluorescent staining of α-SMA (Green) and Calponin1(Red) in suprarenal abdominal aortas, IgG was used as the isotype control. Blue: DAPI. Scale bar: 40 μm. 3 independent experiments showed consistent results. g Western blot of Bax, Cleaved-PARP1, and Bcl-2 in aortic tissue. n = 6 distinct samples for each group. Two-way ANOVA test followed by Turkey’s multiple comparisons test. h H&E staining, Masson, and EVG staining in supra-renal abdominal aorta. Scale bar = 50 μm. n = 6 distinct samples for each group. Collagen content, Two-way ANOVA test followed by Turkey’s multiple comparisons test; EVG elastin degradation score, two-sided Mann–Whitney test. i Activity of matrix metalloproteinases (MMPs) determined by in situ zymography. n = 6 distinct samples for each group, Two-way ANOVA test followed by Turkey’s multiple comparisons test. Elastin degradation score in (h) was represented as the box plots indicate median (middle line), 25th, 75th percentile (box), as well as minimum and maximum values (whiskers). The left data were represented as mean ± SEM. Source data are provided as a Source Data file.
Consistently, in PPE-induced AAA model, overexpression of Rras2 in SMCs reduced incidence of AAA (Supplementary Fig. 6a–c), inhibited the elevation of aortic weight to body weight ratio and ameliorated the dilatation of suprarenal abdominal aorta (Supplementary Fig. 6d, e). Importantly, overexpression of Rras2 in SMCs prevented elastin degradation induced by PPE (Supplementary Fig. 6f, g), and restored the expression of Calponin 1 and α-SMA (Supplementary Fig. 6h, i), indicating that R-Ras2 maintains the contractile phenotype of VSMCs and prevents PPE-induced AAA.
Given the effects of R-Ras2 in preventing VSMCs phenotypic transition and AAA, we sought to determine whether overexpression of Rras2 in SMCs could reverse the established AAA. We adopted a chronic advanced AAA model where AAA is induced by a combination of oral β-aminopropionitrile (BAPN) administration and perivascular PPE application (Supplementary Fig. 7a)18,19,20. Compared with the Ang II infusion model or the PPE model, this chronic advanced AAA model demonstrates persistent long-term aneurysm growth, with considerable thrombus formation and progression at the advanced stages of AAA development. 28 days after surgery, the aortic weight to body weight ratio and maximal external diameter of infrarenal abdominal aorta were significantly reduced in Lenti-Rras2 mice compared with Lenti-Vector (Supplementary Fig. 7b, c). Consistently, the Maximal external diameter of infrarenal abdominal aorta was reduced in Lenti-Rras2 mice compared with Lenti-Vector mice challenged with PPE + BAPN (Supplementary Fig. 7d). Importantly, overexpression of R-Ras2 reduced elastin degradation and restored the expression of Calponin 1 in VSMCs of infrarenal abdominal aorta (Supplementary Fig. 7e–h). Collectively, above findings demonstrated that overexpression of R-Ras2 in SMCs can both prevent AAA development and inhibit the growth of established aneurysms.
R-Ras2 maintains the contractile-related proteins expression in SMCs through preserving phosphorylation of TFII-I
Next, we tried to identify the downstream factors that mediate the protecting effect of R-Ras2. Previous study found that constitutive active form of R-Ras2 could increase the phosphorylation of Akt and ERK1/29. We knocked down R-Ras2 in HASMCs and detected the level of p-Akt (S473) and p-ERK1/2, unexpectedly, we did not observe any difference in phosphorylation of AKT or ERK1/2 (Supplementary Fig. 8a, b). We hypothesized that the relative contributions of R-Ras2 to basal Akt and ERK1/2 phosphorylation may be redundant in the presence of other ras family members9,21. Above results suggest that there are other effectors that specifically mediate the protective role of R-Ras2 in VSMCs homeostasis. Therefore, we investigated proteins in HASMCs that can interact with R-Ras2 through co-immunoprecipitation (CoIP) and liquid chromatography-tandem mass spectrometry (LC-MS/MS). A total of 740 proteins were identified, Gene Ontology (GO) analysis showed an enrichment of genes involved in blood vessel development (Fig. 4a). Among these genes, we found that transcription factor 2I (GTF2I, also known as TFII-I) is enriched on the promoters of genes related with SMCs proliferation, differentiation, and signaling pathway (Fig. 4b), according to in silico analysis of a TFII-I ChIP-seq data (GSE138234). Further analysis of this GEO dataset revealed that TFII-I binds with SMC contractile phenotype-related genes, including TAGLN, ACTA2, and CNN1 (Fig. 4c). CoIP assay confirmed the endogenous interaction of R-Ras2 with TFII-I (Fig. 4d). As expected, knock down of TFII-I reduced the expression of α-SMA, Calponin 1, and SM22 in HASMCs (Fig. 4e), and inhibited the protective effects of R-Ras2 upon Ang II stimulation (Fig. 4f).

a Gene Ontology (GO) categories of proteins that can interact with R-Ras2 and molecular functions of these proteins. b GO analysis of TFII-I target genes (ChIP-seq data: GSE138234). a, b: The p-value was calculated through the default hypergeometric method of enrichGO function with a one-tailed test. No correction was applied for multiple testing. c Recruitment of TFII-I to promoters of TAGLN, ACTA2, and CNN1 based on in silica analysis of the ChIP-seq data. d Interaction of R-Ras2 and TFII-I. e Western blotting of α-SMA, Calponin 1, SM22, and TFII-I in HASMCs transfected with siTFII-I. n = 6 distinct samples. α-SMA, TFII-I: two-sided Unpaired t-test with Welch’s correction. Calponin 1, SM22: two-sided Unpaired t-test. f R-Ras2-overexpressed HASMCs transfected with siTFII-I and treated with Ang II, the levels of SM22 and Calponin 1 were determined. n = 6 distinct samples. Two-way ANOVA test followed by Turkey’s multiple comparisons test. g Western blot analysis of TFII-I phosphorylation. n = 6 distinct samples. Two-sided Unpaired t-test. h Western blotting analysis of TFII-I level in nuclear and cytoplasm fractions. n = 6 distinct samples, two-sided Unpaired t-test. i Immunofluorescence staining of TFII-I in HASMCs stimulated with Ang II. Green: TFII-I; Blue: DAPI. Scale bar: 20 μm. 3 independent experiments showed consistent results. j Phosphorylation of TFII-I in aorta of non-AAA and AAA patients. n = 6 distinct samples, two-sided Unpaired t-test. k Diagram showing the locations of E-boxes, WT and MUT INR in the TAGLN promoter construct and deletion constructs used for promoter activity studies (left) and the relative luciferase activity (right). n = 6 distinct samples. Ordinary one-way ANOVA followed by Dunnett’s multiple comparisons test. l ChIP assay detected the enrichment of TFII-I to the promoter of ACTA2, CNN1, and TAGLN in HASMCs transfected with si-RRAS2. n = 6 distinct samples for each group. Two-sided Unpaired t-test. All data were represented as mean ± SEM. Source data are provided as a Source Data file.
Recent studies suggest that TFII-I plays an important role in signal transduction through its phosphorylation on tyrosine 248 (Y248). Phosphorylation of TFII-I mediates its nuclear translocation and transcriptional activity22,23,24. We observed that knockdown of RRAS2 significantly reduced the phosphorylation of Y248 in TFII-I (Fig. 4g). Further, we found that nuclear translocation of TFII-I was reduced in HASMCs transfected with si-RRAS2 (Fig. 4h). Immunofluorescent staining of TFII-I confirmed the reduction of nuclear TFII-I in HASMCs transfected with si-RRAS2 (Fig. 4i). In the aorta of AAA patients, we also observed a reduction in p-Y248 of TFII-I (Fig. 4j).
TFII-I can initiate gene transcription through binding to the E-box and INR elements. We analyzed the 2.0-kb human TAGLN promoter region and found several E-box motifs and an INR motif at the transcriptional starting site. We then constructed a series of luciferase reporter plasmids containing the truncated TAGLN promoters (−1656, −1288; −716), and TAGLN promoters with wild-type (WT) or mutated (MUT) INR binding motifs to investigate the binding site of TFII-I on TAGLN promoter. Luciferase assay found that overexpression of TFII-I increased the luciferase activity of WT rather than MUT or truncated promoters, confirming the binding of TFII-I to the INR motif in TAGLN promoter (Fig. 4k). Finally, ChIP assay confirmed that knockdown of R-Ras2 reduced the binding of TFII-I to the promoters of ACTA2, CNN1, and TAGLN (Fig. 4l). In fact, similar with our observations, it has been observed that association of TFII-I with GTPase activating protein might be downstream effector of the growth factor receptors, and this interaction would undergo signal-induced tyrosine phosphorylation of TFII-I, which eventually mediates dissociation of TFII-I from the complex and translocation to the nucleus for gene activation25.
MET tyrosine kinase is involved in the phosphorylation of TFII-I
To further determine the growth factor receptors involved in R-Ras2-mediated phosphorylation of TFII-I, we screened proteins approximal to R-Ras2 in HASMCs using live cell proximity-dependent biotin labeling of proteins (BioID) assay. BioID allows identification of transient or weak protein interactions in living cells that may be missed by conventional CoIP method, therefore, it is suitable for detecting the interaction between kinases and effectors26. BirA biotin ligase (termed BirA*), which conjugates biotin to proteins within the immediate vicinity (~10 nm), was fused in frame to the N-terminus of R-Ras2 (BirA*-RRAS2) and cloned into lentivirus for HASMC transfection. Cells were then treated with biotin, and the biotinylated proteins were affinity-captured with streptavidin-conjugated beads and analyzed by tandem MS spectrometry (Fig. 5a). A total of 567 proteins were identified, including Ras interactors, such as guanine nucleotide exchange factors (GEFs) and Ras-GTPase activating proteins (GAP). The approximation of TFII-I with R-Ras2 was also identified in the BioID assay. Functional annotation identified twelve proteins with kinases activity according to an expert-curated resource of pharmacological targets (IUPHAR/BPS Guide to PHARMACOLOGY). Among them, TTK and MET are tyrosine kinases (Fig. 5b). Knockdown of MET, rather than TTK, significantly reduced the phosphorylation of TFII-I (Fig. 5c and Supplementary Fig. 8c–e). BioID, PLA, and coimmunoprecipitation assays further confirmed the endogenous adjacency of MET with R-Ras2 (Fig. 5d–f). As expected, in the aorta of ApoE−/− mice, although infusion of Ang II reduced the interaction of R-Ras2 with MET, as demonstrated by reduced PLA signal, overexpression of R-Ras2 in VSMCs restored this interaction (Fig. 5g).

a BioID assay schema. b Western blotting analysis of biotinylated proteins with HRP-streptavidin in HASMCs transfected with BirA*-R-Ras2. c Phosphorylation of TFII-I in HASMCs transfected with si-MET. n = 6 distinct samples for each group. Two-sided Unpaired t-test. d Proximity of R-Ras2 with MET determined by BioID assay. e Proximity of R-Ras2 with MET determined by PLA assay. Scale bar: 40 μm, PLA positive signal: Red, DAPI: Blue. 3 independent experiments showed consistent results. f Coimmunoprecipitation of endogenous R-Ras2 with MET. g Proximity of R-Ras2 with MET determined by PLA assay in sections of abdominal aorta of AAA mice. Scale bar: 20 μm. PLA positive signal: Red, Elastin: Green, DAPI: Blue. 3 independent experiments showed consistent results. h HASMCs were stimulated with 0, 25, 50, and 100 ng/ml of hepatic growth factor (HGF), the level of TFII-I phosphorylation was determined. n = 6 distinct samples for each group. Ordinary one-way ANOVA followed by Dunnett’s multiple comparisons test. i HASMCs were transfected with lentivirus encoding HA-R-Ras2 F75A mutant (HA-R-Ras2 MUT1) and HA-R-Ras2 F75A/M78I/Y82A mutant (HA-R-Ras2 MUT2). The interaction of R-Ras2 with MET was determined. j HASMCs was transfected with si-RRAS2, and were transfected with lentivirus encoding R-Ras2 WT or HA-R-Ras2 MUT2, the phosphorylation of TFII-I, the protein levels of α-SMA, Calponin1, and SM22 were determined. n = 6 distinct samples: Ordinary one-way ANOVA followed by Turkey’s multiple comparisons test. All data were represented as mean ± SEM. Source data are provided as a Source Data file.
Next, we asked if MET is involved in phosphorylation of TFII-I. Hepatocyte growth factor (HGF) is the endogenous ligand of MET. We found that HGF dose-dependently increased the phosphorylation of TFII-I (Fig. 5h). Ras guanine-nucleotide exchange factor Son of sevenless (Sos) activates Ras in response to the activation of cell-surface receptors. It has been reported that at the heart of the interface between Ras, and Sos is a cluster of three hydrophobic side chains from the Switch 2 region of Ras (Tyr 64, Met 67, and Tyr 71)27. Among them, Tyr 64 is believed to be the most important residue mediating Ras-Sos binding28. According to the partial amino acid sequence alignment of human R-Ras2 with other Ras members, we mutated Phe75 (corresponding to Tyr64 in K-Ras, H-Ras, and N-Ras) of R-Ras2 to Ala (F75A)27 and determined the interaction between MET and R-Ras2. F75A mutation did not affect the affinity of R-Ras2 for MET. Further triple mutation of F75A/M78I/Y82A (corresponding to Y64/M67/Y71 in K-Ras, H-Ras and N-Ras) resulted in reduced binding affinity of R-Ras2 for MET (Fig. 5i). As expected, in HASMCs lacking endogenous R-Ras2, overexpression of wild-type R-Ras2, rather than F75A/M78I/Y82A mutant, rescued the phosphorylation of TFII-I (Fig. 5j). Smooth muscle cell contractile-related proteins, including α-SMA, Calponin 1 and SM22, were also restored with overexpression of R-RAS2 WT rather than R-RAS2 F75A/M78I/Y82A mutant (Fig. 5j). Collectively, these results indicate that MET is involved in R-Ras2-mediated phosphorylation and nuclear translocation of TFII-I, loss of R-Ras2 impairs this signaling pathway, and reduces TFII-I-dependent transcription of contractile-related genes in VSMCs.
MEG3 is the upstream regulator of R-Ras2
We next explored the upstream factors that regulate the expression of R-Ras2. We investigated the effects of Ang II on the transcriptional activity, mRNA stability, and protein stability of R-Ras2 in HASMCs. Unfortunately, we did not observe any difference in the transcriptional activity of RRAS2 promoter in HASMCs stimulated with saline or Ang II (Supplementary Fig. 9a). The stability of RRAS2 mRNA was further determined, and a reduction of half-life of RRAS2 mRNA upon Ang II treatment was observed (Supplementary Fig. 9b). We further detected protein stability of R-Ras2, and found no difference in the half-life of R-Ras2 protein in HASMCs treated with or without Ang II (Supplementary Fig. 9c). Moreover, neither chloroquine nor MG132 incubation restored the protein level of R-Ras2 upon Ang II (Supplementary Fig. 9d, e), excluding the involvement of proteasomal or lysosomal degradation. According to above observations, we further investigated factors that regulate the stability of RRAS2 mRNA.
Non-coding RNAs (ncRNAs), including microRNAs (miRNAs) and long non-coding RNAs (lncRNAs), circular RNA (circRNA) and et al., have been identified as critical regulators of post-transcriptional gene expression29. Of note, miRNAs and lncRNA primarily regulate gene expression by influencing stability of mRNA. We predicted miRNAs targeting RRAS2 using starbase and targetscan, and identified 49 overlapping candidates. We then collected the common different expressed miRNAs from human AAA biopsy samples (GSE62179, GSE63541), and tried to found miRNAs that both targeting RRAS2 and dysregulated during AAA. Unfortunately, we found no miRNAs targets that satisfy both criteria (Supplementary Excel 1).
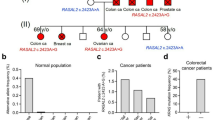
LncRNA is a type of RNA that is greater than 200 nucleotides in length without protein-coding function. LncRNAs regulate gene expression at the transcriptional and post-transcriptional levels, the cellular localization of lncRNAs are important features that are often linked to their functions30,31,32. We performed RNA-sequencing in HASMCs stimulated with Ang II (n = 2) or saline (n = 2) to identify different regulated lncRNAs (false discovery rate p-value < 0.05; fold change > 2 or < 0.5) (Fig. 6a). Cross-checking upregulated or downregulated transcripts in an experimental AAA mouse model (GSE153097) identified maternally expressed 3 (MEG3) as a species-independent lncRNA with potential implications for experimental AAA progression. In HASMCs stimulated with Ang II, Q-PCR found reduction of MEG3 and importantly, overexpression of MEG3 could inhibit Ang II-induced RRAS2 degradation (Supplementary Fig. 9f, g). The level of MEG3 reduced significantly both in AAA patients and AAA mice as determined by Q-PCR (Fig. 6b, c). Using fluorescence in situ hybridization (FISH) assay, we observed lower staining of MEG3 in patient specimens compared with healthy controls (Fig. 6d). Meanwhile, we found reduction of MEG3 staining in HASMCs stimulated with Ang II compared with Ctrl (Fig. 6e). Accordingly, knockdown of MEG3 reduced the protein level of R-Ras2, whereas overexpression of MEG3 significantly restored R-Ras2 level in HASMCs stimulated with Ang II (Fig. 6f, g). Besides, overexpression of MEG3 in HASMCs could reduce Bax and Cleaved-PARP1 levels upon TNF-α + CHX stimulation (Supplementary Fig. 9h–j), and prevent the migration of HASMCs as determined by scratch and transwell assay (Supplementary Fig. 9k–m). More importantly, overexpression of MEG3 restored the protein level of Calponin 1, SM22 and α-SMA in HASMCs stimulated with Ang II, and these beneficial effects were abolished with the loss of R-Ras2 (Fig. 6h). In situ zymography also demonstrated that overexpression of MEG3 could alleviate MMPs activation, and this protective effect was reversed with knockdown of RRAS2 (Fig. 6i). Collectively, above data demonstrate that MEG3 modulates the homeostasis of smooth muscle cell during AAA through R-Ras2.

a Volcano plot of deregulated transcripts in HASMCs stimulated with Ang II compared with PBS. The p-value was calculated by a Wilcoxon rank sum test. No correction was applied for multiple testing. b MEG3 expression level in aorta of AAA patients and healthy control individuals (non-AAA). n = 6 distinct samples for each group, two-sided Unpaired t-text. c Meg3 expression level in aorta of Sham and AAA mice. n = 5 mice for each group, two-sided Mann–Whitney test. d Fluorescence in situ hybridization (FISH) assay to determine the expression of MEG3 in aortic sections from AAA patients and non-AAA. Scale bar: 50 μm. MEG3: Red, DAPI: Blue. 3 independent experiments showed consistent results. e Fluorescence in situ hybridization (FISH) assay to determine the expression of MEG3 in HASMCs simulated with Ang II. Scale bar: 50 μm. MEG3: Red, DAPI: Blue. 3 independent experiments showed consistent results. f Representative western blotting and quantification of R-Ras2 in HASMCs transfected with si-MEG3. n = 6 distinct samples for each group, two-sided unpaired t-test. g Representative western blotting and quantification of R-Ras2 in HASMCs overexpressing MEG3 and stimulated with Ang II. n = 6 distinct samples for each group. Two-way ANOVA test followed by Turkey’s multiple comparisons test. h HASMCs were overexpressed with MEG3, followed by transfected with si-RRAS2 and stimulated with Ang II. Protein levels of Calponin 1, α-SMA, SM22, and R-Ras2 were determined and their relative expression levels were analyzed. n = 6 distinct samples for each group. Two-way ANOVA test followed by Turkey’s multiple comparisons test. i Representative image of in situ zymography for detection of MMPs activity (Green). DAPI: Blue. Scale bar = 50 μm. n = 6 distinct samples for each group. Two-way ANOVA test followed by Turkey’s multiple comparisons test. All data were represented as mean ± SEM. Source data are provided as a Source Data file.
MEG3 maintains mRNA stability of RRAS2 through binding with ELAVL1
To further investigate how MEG3 modulates the expression of R-Ras2, we first used the lncTar online tool (http://www.cuilab.cn/lnctar) to predict the interaction between MEG3 and RRAS2. Unfortunately, we did not find direct interaction between MEG3 and RRAS2 (data not shown). RNA-binding proteins (RBPs) are essential for lncRNA-mediated mRNA stability control. RNA pull-down assay and mass spectrometry (MS) analysis was then performed to identify potential MEG3 binding proteins (Supplementary Fig. 10a). We analyzed the following 4 datasets: MEG3 binding proteins identified from our RNA pulldown and MS analysis results, RRAS2 binding proteins predicted by CatRAPID algorithm, RBPs identified from CLIP-Seq dataset for MEG3, and RBPs identified from GO terms database. ELAV-like protein 1 (ELAVL1) was finally identified as the only potential MEG3 binding protein (Fig. 7a and Supplementary Fig. 10b). RNA stability assay found that knockdown of ELAVL 1 reduced the half-life of RRAS2 (Fig. 7b).

a Identification of ELAVL 1 as RRAS2 and MEG3 binding protein. b RRAS2 mRNA stability in HASMCs transfected with si-ELAVL 1. n = 3 distinct samples. c Interaction of MEG3 with ELAVL 1 determined by RNA pull-down assay. d Interactions of ELAVL1 with full-length or MEG3 truncations. e Interaction between MEG3 and ELAVL 1 determined by RIP assay. n = 5 distinct samples, two-sided Unpaired t-test with Welch’s correction. f The interaction between RRAS2 and ELAVL 1. n = 5 distinct samples, two-sided Unpaired t-test. g Protein level of R-Ras2 in HASMCs transfected with si-ELAVL 1. n = 6 distinct samples, two-sided Unpaired t-test with Welch’s correction. h Expression of R-Ras2 in HASMCs overexpressing with ELAVL1 and stimulated with Ang II. n = 6 distinct samples for each group. Two-way ANOVA test followed by Turkey’s multiple comparisons test. i HASMCs were overexpressed with MEG3, followed by transfected with si-ELAVL 1 and stimulated with Ang II, the level of RRAS2 were determined. n = 6 independent experiments. Two-way ANOVA test followed by Turkey’s multiple comparisons test. j Schematic of 3′UTR, CDS and 5′UTR of RRAS2 reporter plasmids. k, l Luciferase activity of RRAS2 reporter plasmids. n = 6 independent experiments for each group, two-sided Unpaired t-test. m Schematic prediction of ELAVL 1 binding AU-rich elements (AREs) within the 3′UTR of RRAS2. n Luciferase activity of RRAS2 3′UTR variants (Full, a–e) in HEK293 cells with empty or ELAVL1 overexpressing vector. n = 6 independent experiments for each group, two-sided Unpaired t-test. o RIP analysis of relative enrichment of RRAS2 transcripts in ELAVL 1-immunoprecipitates. n = 5 independent experiments for each group, two-sided Unpaired t-test with Welch’s correction. All data were represented as mean ± SEM. Source data are provided as a Source Data file.
We then confirmed the binding of ELAVL 1 to MEG3 via biotin-labelled RNA pulldown assay (Fig. 7c). To determine the precise binding regions of MEG3 for ELAVL 1, we constructed a series of biotinylated MEG3 fragments by RNA pulldown assay and found that ELAVL 1 was only detectable in the pull-down complex of 656–1070 nt MEG3 fragment (Fig. 7d). RNA immunoprecipitation (RIP) assay found that the enrichment of MEG3 to ELAVL 1 was reduced in HASMCs stimulated with Ang II (Fig. 7e). In addition, knockdown of MEG3 reduced the level of ELAVL 1-bound RRAS2 (Fig. 7f). Western blotting confirmed that knockdown of ELAVL 1 reduced the expression of R-Ras2 (Fig. 7g), and overexpression of ELAVL 1 restored the expression of R-Ras2 in HASMCs stimulated with Ang II (Fig. 7h). Although overexpression of MEG3 restored the level of RRAS2 in HASMCs stimulated with Ang II, silencing of ELAVL 1 inhibited the protective effect (Fig. 7i).
We then determined the binding regions of ELAVL 1 in RRAS2. Vectors containing 3′UTR, CDS, and 5′UTR of RRAS2 were constructed and co-transfected with si-ELAVL 1 or ELAVL 1 for luciferase assay. The result demonstrated that the 3′UTR region of RRAS2 bound ELAVL 1 (Fig. 7j–l). ELAVL 1 can regulate the mRNA stability of targeting genes by binding AU-rich elements (AREs), a major class of cis elements within 3′-UTR33. Close inspection of the 3′-UTR of RRAS2 transcript by web tools AREsite (http://rna.tbi.univie.ac.at/AREsite2/welcome) identified five AREs in tandem. We constructed luciferase reporter vectors containing full-length or series truncations of RRAS2 3′-UTR as demonstrated in Fig. 7m. Co-transfection of vectors overexpressing ELAVL 1 significantly increased the luciferase activity of full-length and 3′UTR-B, C rather than 3′-UTR-A, D or E, confirming that ELAVL 1 can bind the AREs in 559–693 nt of RRAS2 3′UTR (Fig. 7n). RIP analysis demonstrated an enrichment of RRAS2 transcripts in ELAVL 1-immunoprecipitated complex in HASMCs, which was reduced upon Ang II treatment (Fig. 7o). Taken together, these results suggest that ELAVL 1 mediates the binding of MEG3 with RRAS2, which maintains mRNA stability of RRAS2 and inhibits smooth muscle cell phenotypic switching.
Overexpression of MEG3 alleviates AAA
To evaluate the significance of MEG3 in AAA in vivo, recombinant adeno-associated virus serotype 9 (AAV9) vectors containing MEG3 driven by murine Sm22a promoter or control vector34 (AAVMEG3 and AAVvector, respectively, 1 × 1012 vp/mL, 50 μL/mouse) were intravenously injected into mice followed by subcutaneous implantation of Ang II minipump (Supplementary Fig. 11a). Four weeks later, the level of MEG3 was confirmed by FISH assay. MEG3 reduced in aorta of Ang II-infused mice, and overexpression of MEG3 inhibited Ang II-induced reduction (Supplementary Fig. 11c). As expected, overexpression of MEG3 significantly reduced the incidence of AAA in Ang II-treated mice (Fig. 8a, b), with no influence on SBP (Supplementary Fig. 11b). The increases in aorta weight and maximal lumen diameter of abdominal aorta were also prevented by overexpressing of MEG3 in Ang II-treated mice (Fig. 8c, d). Diameters of the ascending aorta, thoracic aorta, descending aorta, and aortic arch were comparable among the four groups (Supplementary Fig. 11d). The levels of SMCs contractile apparatus-related proteins, including α-SMA, Calponin 1 and SM22, were reduced significantly in AAVvector + Ang II mice, and overexpression of MEG3 restored their expression (Fig. 8e). Accordingly, co-immunofluorescent staining confirmed restoration of Calponin 1 and α-SMA expression in the aorta of AAVMEG3 + Ang II mice (Supplementary Fig. 11e), and Q-PCR revealed increased mRNA level of Acta2 and Cnn1 in the aorta of AAVMEG3 + Ang II compared with AAVvector + Ang II mice (Supplementary Fig. 11f). Moreover, H&E staining revealed reduced thickness of aortic medial, Masson’s trichrome staining showed alleviated collagen deposition, and EVG staining showed reduced elastin fiber fragmentation in AAVMEG3 + AngII compared with AAVvector + AngII mice (Fig. 8f). Finally, in situ zymography found that the increased activity of MMPs was abrogated in AAVMEG3 mice subjected to Ang II infusion (Fig. 8g).

a Representative photograph of aortas in each group. b Incidence of AAA. AAVvector+ Ang II (n = 12) and AAVMEG3+ Ang II (n = 11), two-sided Fisher’s exact test. N.D. means Not Detected. c Quantification of the maximal abdominal aorta lumen diameters in each group at different time points. Two-way RM ANOVA followed by Bonferroni’s multiple comparisons test. d Ratio of aortic weight to body weight. Two-way ANOVA test followed by Turkey’s multiple comparisons test. c, d: AAVvector + Saline (n = 5), AAVvector + Ang II (n = 8), AAVMEG3 + Saline (n = 5) and AAVMEG3 + Ang II (n = 9). e Western blot and quantification of α-SMA, Calponin 1, and SM22 in aortic tissue of mice subjected with different treatment. AAVvector + Saline (n = 5), AAVvector + AngII (n = 6), AAVMEG3 + Saline (n = 5) and AAVMEG3 + Ang II (n = 6). Two-way ANOVA test followed by Turkey’s multiple comparisons test. f Representative H&E staining, Masson, and EVG staining of supra-renal abdominal aorta. Scale bar = 50 μm. Collagen content, AAVvector + Saline (n = 5), AAVvector + AngII (n = 6), AAVMEG3 + Saline (n = 5) and AAVMEG3 + Ang II (n = 6). Two-way ANOVA test followed by Turkey’s multiple comparisons test, EVG elastin degradation score, AAVvector + Saline (n = 5), AAVvector + Ang II (n = 8), AAVMEG3 + Saline (n = 5), and AAVMEG3 + Ang II (n = 9), two-sided Mann–Whitney test. g Representative immunofluorescent staining of MMPs activity determined by in situ zymography. Scale bar: 20 μm. AAVvector + Saline (n = 5), AAVvector + AngII (n = 6), AAVMEG3 + Saline (n = 5) and AAVMEG3 + Ang II (n = 6). Two-way ANOVA test followed by Turkey’s multiple comparisons. Elastin degradation score in (f) was represented as the box plots indicating median (middle line), 25th, 75th percentile (box), as well as minimum and maximum values (whiskers). The left data were represented as mean ± SEM. Source data are provided as a Source Data file.
Next, we examined whether overexpression of MEG3 alleviates aneurysm development in PPE-induced AAA model (Supplementary Fig. 12a). Overexpression of MEG3 reduced incidence of AAA and aortic weight to body weight (Supplementary Fig. 12b–d), as well as reduced the maximal external diameter of infrarenal abdominal aorta (Supplementary Fig. 12e). EVG staining confirmed a lower grade of elastin degradation in AAVMEG3 mice compared with AAVVector mice challenged with PPE (Supplementary Fig. 12f, g). Moreover, the expression of Calponin1 and α-SMA in aortic wall were significantly restored in mice overexpressed with MEG3 (Supplementary Fig. 12h, i).
To explore the therapeutic potential of MEG3 for AAA in vivo, we intravenously injected AAVMEG3 or AAVVector into mice after the AAA model has been established (Supplementary Fig. 13a). MEG3 overexpression reduced the maximal external infrarenal aortic diameters and ratio of aorta weight to body weight upon PPE + BAPN stimulation (Supplementary Fig. 13b-d). Consistently, the expansion rate of abdominal aorta was reduced in AAVMEG3 mice compared with AAVVector mice challenged with PPE + BAPN (Supplementary Fig. 13e). In addition, overexpression of MEG3 alleviated elastin degradation (Supplementary Fig. 13f, g), and restored the expression of Calponin 1 in VSMCs of infrarenal abdominal aorta (Supplementary Fig. 13h, i). Collectively, these observations highlight a protective role of MEG3 for the prevention and treatment of AAA.
Discussion
In this study, we provided evidences to support an essential role of R-Ras2 in maintaining VSMCs homeostasis during AAA. We showed that R-Ras2 mediated phosphorylation of TFII-I, which induced nuclear translocation and transcriptional activation of TFII-I on genes including TAGLN, ACTA2, and CNN1. MET is the tyrosine kinase mediating the phosphorylation of TFII-I. Furthermore, our findings suggested that MEG3 prevented the decay of RRAS2 to maintain VSMCs homeostasis and alleviated the progression of AAA.
Accumulating evidence suggests Ras a critical regulator in VSMCs phenotypic switching and a putative target for pharmacological intervention in vascular proliferative disorders12,13,14,15. Signaling begins with the activation of tyrosine kinase receptors through recruiting GEF (Sos, for example) to induce Ras activation35. For example, H-Ras has been found to mediate VSMCs proliferation in carotid or coronary artery subjected to balloon injury and local delivery of negative mutants of H-Ras prevents acute vascular injury12,13,14. R-Ras1, on the contrary, is largely confined into fully differentiated smooth muscle cells, and R-Ras1-null mice showed exaggerated neointimal thickening in response to arterial injury15. The different roles of H-Ras and R-Ras1 in VSMCs proliferation and differentiation suggest unique roles of different Ras family members. It is well known that the Ras signaling pathway participates in many cellular biological processes, such as growth, differentiation, and apoptosis36. The excessive activation of the Ras signaling pathway is widely investigated in many malignant tumors, and inhibitor of K-Ras G12C mutant Sotorasib has been found to be effective in treating advanced solid tumors37. However, Sotorasib can induce hypertension37, which might be an off-target effect on wild-type K-Ras or other Ras family members. Besides, in tumor patients with N-Ras overactivation and treated with Ras pathway inhibitor MEK162, hypertension is a main adverse effect (https://clinicaltrials.gov/ct2/show/results/NCT01885195). These observations are in consistent with our study that the normal function of Ras is important for vascular homeostasis.
We noticed that RRAS2 mutations associated with Noonan Syndrome and cancer make R-Ras2 constitutively active. While in our study, we observed that loss of RRAS2 in the SMCs make the mice more vulnerable to AAA. It seems that both overactivated and dysfunction of R-Ras2 induce the vascular remodeling injury. This might because the diverge expression and downstream effectors of R-Ras2 in different vascular beds. Besides, constitutive active R-Ras2 in NIH3T3 cell line can increase the phosphorylation of ERK1/2 and Akt9, while knockdown of R-Ras2 in HASMCs had no influence on phosphorylation of ERK or Akt. We supposed that the basal activity of R-Ras2 is important for maintaining normal cellular function and different downstream effectors mediate the specific effects of R-Ras2 in different kinds of cells and tissues.
Rather than a general transcriptional factor, previous evidences show that the activity of TF-II-I is cell-type dependent and partner-interaction dependent. For example, TFII-I specifically repressed a subset of TGFβ-responsive genes in PANC1 and HepG2 cell lines through interacting with SPT5, a pausing complex component38. In human umbilical vein endothelial cells), p190RhoGAP stimulates VEGFR2 expression by restricting nuclear translocation of TFII-I, therefore controlling capillary network formation in retinal angiogenesis39. Here in SMCs, we found a new mechanism that mediates the phosphorylation and activation of TFII-I through MET and R-Ras2. Site directed mutation revealed detailed amino acids that mediate MET-dependent TFII-I activation by R-Ras2, which is first discovered in this study as far as we known. Hepatic growth factor (HGF), the endogenous ligand of MET, increased the phosphorylation of TFII-I in a dose-dependent manner. These observations uncovered the importance of MET-dependent signaling pathway in preserving TFII-I phosphorylation and VSMCs contractile phenotype.
Long noncoding RNAs (lncRNAs) can interfere with gene expression and post-transcriptional signaling at various levels. Accumulating evidence suggests that lncRNAs play a crucial role in vascular biology and cardiovascular diseases40,41,42,43. In this study, we observed reduced stability of RRAS2 and revealed that MEG3 is a critical regulator of RRAS2 stability through interacting with ELAVL1. Reduced level of MEG3 in VSMCs is observed during AAA, and overexpression of MEG3 can alleviate AAA in vivo. Interestingly, a recent study found that dysregulation of 3′UTR of RRAS2 reduces the R-Ras2 protein level during cellular senescence44. In addition, a single nucleotide polymorphism (rs8570) in the 3′UTR of RRAS2 links with higher expression of R-Ras2 in chronic lymphocytic leukemia patients45. Our study provides an alternative way for RRAS2 post-transcriptional regulation. More importantly, Evidences demonstrate that MEG3 is a lncRNA tumor suppressor, and its level reduced in many cancer cell lines46. Therefore, we believe that overexpression of MEG3 can prevent the progression of AAA with little tumorigenic risk.
In summary, our study provides the first evidence for an essential role of R-Ras2 in AAA through regulating VSMCs homeostasis. Our results showed that loss of R-Ras2 reduced SMCs contractile apparatus-related proteins expression through reducing TFII-I phosphorylation, nuclear translocation, and transcriptional activity in a tyrosine kinase MET-dependent manner. Our study also uncovered MEG3 an upstream regulator of R-Ras2 through preventing RRAS2 degradation, and provided evidence for the application of MEG3 in preventing AAA.
Methods
Human aorta samples
Human aortic tissue was collected in compliance with the Declaration of Helsinki. The use of AAA tissue was approved by the Ethics Committee of Nanjing Drum Tower Hospital (No. 2020-292-01). The use of the non-AAA samples was approved by the Ethics Committee of Nanjing First Hospital, Nanjing Medical University (KY20190404-03-KS-01). Informed consent was obtained from all participants. AAA was diagnosed with maximal abdominal aortic diameters ≥55 mm in men or ≥50 mm in women measured by computed tomographic angiography. Abdominal aortic tissues were collected during surgical repair for AAA at the Nanjing Drum Tower Hospital. The non-AAA samples used in this study were obtained from the Biospecimen Bank of the Nanjing First Hospital, Nanjing Medical University. The information of AAA patients and non-AAA was listed in Supplementary Table 1.
Animals
All animal experiments were conducted in accordance with the ARRIVE guidelines for the care and use of laboratory animals, and protocols were approved by the Nanjing Medical University Animal Care and Use Committee (No. IACUC-1911031). Rras2flox/flox (Rras2f/f, C57BL/6) mice were generated by the Animal Center of the Nanjing Medical University (Nanjing, China). Tagln-Cre mice with C57BL/6 background was a kind gift from Nanjing Drum Tower Hospital. Vascular smooth muscle cells (SMCs)-specific R-Ras2 knockout mice (Rras2SMKO) mice were generated by crossbreeding Rras2f/f mice with Tagln-Cre mice. Six-week-old male ApoE−/− mice were purchased from the Animal Center of the Nanjing University (Nanjing, China). Only male mice were used in this study, because male is an independent risk factor for AAA and the use of female mice would have resulted in much higher number of animals. In compliance with the principle of the Reduction, we conducted the study only on male mice. Mice were kept under a 12-h light/dark cycle at 23 °C with ad libitum access to food and water.
Abdominal aortic aneurysm model
Both male and female mice were used in this study. To induce murine Ang II-induced AAA model, 8- to 10-week-old ApoE−/− mice were infused with Ang II (1000 ng/kg per min) or saline via an osmotic minipump (Model 2004, Alzet) for 4 weeks. The mice were anesthetized with isoflurance (1.5%) and placed the pumps into subcutaneous space through a small incision in the back of the neck and skin were closed with sutures. The incision site was sutured and penicillin was injected intraperitoneally to prevent infection. To determine the AAA formation, measurements of aortic maximal diameter were recorded at the ascending aorta, aortic arch, descending aorta, thoracic aorta, and abdominal aorta with a Vevo 2100 High-Resolution Micro-Ultrasound System (FUJIFILM Visual Sonics Inc.) at 0, 14, and 28 days. According to the measurements of echocardiography, the AAA was defined as a suprarenal aortic diameter ≥1.5 times than the mean aortic diameter of the same segment in unchallenged mice with the same genetic background, aortic dissection was defined as the presence of hematoma within the aortic wall detected on gross examination, and aortic rupture and premature death were also included47. For blood pressure measurements, mice were placed into restrainers and SBP was measured with the noninvasive tail-cut blood pressure analysis system (BP2000, Visitech) at 0, 7, 14, 21, and 28 days after pumps transplantation. The mice died before the end of the study will be collected for gross image and incidence rate analysis, but excluded from diameter measurement, staining, and western blotting study.
The porcine pancreatic elastase (PPE)-induced AAA model was implemented as previously described48 with some improvements. In brief, mice were anesthetized with Isoflurane, the infrarenal abdominal aorta was isolated and then surrounded with a sterile gauze presoaked with 30 μL of elastase (66.7 units/mL, Sigma, E1250) or heat-inactivated PPE. After 5 min of incubation, the gauze was removed, the peritoneal cavity was flushed with sterile saline for 5 min before suturing. After 14 days, mice were anesthetized and euthanized by collection of blood and organs.
The chronic advanced AAA model was induced as previously reported18,19,20: briefly, 0.2% beta-aminopropionitrile (BAPN) drinking water was provided 2 days before aneurysm induction surgery to mice until the end of study. For aneurysm induction surgery, 30 μL of the active form of PPE (66.7 U/ml, Sigma, E1250) was applied on adventitia of the infrarenal abdominal aorta for 5 min. Then the gauze was removed and the peritoneal cavity was flushed with sterile saline for 5 min before suturing. After 28 days, mice were anesthetized and euthanized by collection of blood and organs.
Cell culture
HASMCs (6110, ScienCell) purchased from ScienCell Research Laboratories were cultured in Smooth Muscle Cell Medium (SMCM, 1101, ScienCell). Human aortic endothelial cells (HAECs) were a gift from Prof. Yong Xu (Nanjing Medical University), originally obtained from Lifeline Cell Technology (02357, Lifeline Cell Technology) and cultured in Endothelial Cell Medium (1001, ScienCell). HASMCs and HAECs were used between 6 and 8 passages. HEK293T cells were a gift from Prof. Qi Chen (Nanjing Medical University) and maintained in Dulbecco’s Modified Eagle’s Medium (Gibco) supplemented with 10% (v/v) FBS. All cells were cultured at 37 °C in an incubator with 5% CO2. HASMCs were starved in serum-free SMCM for 24 h followed by Angiotensin II (Ang II, 10−6M) treatments.
Luciferase reporter assays
Luciferase reporter assay was performed using the Dual-Luciferase Reporter Assay System (Promega). HEK293T or HASMCs were transient transfected with a corresponding firefly luciferase reporter vector and Renilla luciferase vector using Lipo3000 transfection reagent (Invitrogen) according to the manufacturer’s protocol. After incubation, cells were starved for 24 h. Cells were harvested 48 h after transfection and luciferase activity was measured by a luciferase assay kit (Promega, E1910). The luciferase activity is presented as the ratio of firefly luciferase to renilla luciferase activity.
Immunofluorescent staining
Cultured cells, frozen aortic sections from AAD mice, and the dewaxed paraffin sections of human aorta were incubated with 4% paraformaldehyde for 20 min, permeabilized with 0.3% Triton ×-100 for 15 min, and blocked with 10% Bovine serum albumin (BSA) for 45 min at room temperature. After incubation, samples were washed with PBS and incubated with primary antibodies at 4 °C overnight. The following day, samples were incubated with the corresponding secondary antibody for 2 h at room temperature. The nucleus was visualized by DAPI staining. Immunofluorescence confocal microscopy was performed with an LSM 800 confocal microscope (Carl Zeiss). Antibodies used in the study were listed in Supplementary.
MMPs activity
MMPs activity was detected by in situ zymography. Briefly, frozen aortic sections from AAD mice (5 μm) or cultured HAMSCs were incubated with fluorogenic gelatin substrate (EnzChek Proteinase Assay Kit, Invitrogen) dissolved to 25 mg/mL in zymography buffer according to the manufacturer’s protocol. The proteolytic activity of MMP was measured as green fluorescence by confocal microscopy (LSM 800) and the fluorescent intensity was measured by Image J.
Transwell assay
Transwell assay was performed using transwell chambers (8.0 μm, Corning). Briefly, HASMCs were stimulated and trypsinized, counted, and incubated in the upper chamber with 200 μl serum-free SMCM in a 12-well plate (5 × 105 cells/mL). The plate containing the transwell chambers was incubated overnight at 37 °C. Then, cells were fixed with methanol for 30 min and stained with 0.5% crystal violet for 20 min. To facilitate the counting of migrated cells, cells on the upper membrane that did not migrate through membrane were removed gently with a cotton bud. The migrated cells on the lower surface of the transwell were photographed in 3 random fields per well and calculated by Image-Pro Plus 6.0 software.
Chromatin immunoprecipitation (ChIP)
ChIP was performed as described before41. Briefly, HASMCs were cross-linked with formaldehyde (final concentration: 1%) for 10 min at 37 °C. A final concentration of 0.125 M glycine was added to terminate the cross-linking. Cells were washed, harvested, and lysed with 500 μL lysis. Next, the genomic DNA in cells was sonicated into 300–1000 bp fragments. 50 μl of lysate was separated and used as input. The rest of the lysate was rocked overnight with the protein A/G beads, which were pre-cleaned by BSA and salmon sperm. The following day, the mixture was further incubated (4 h, 4 °C) with anti-TFII I or normal IgG antibody. The beads were washed twice with Low Salt buffer (150 mM NaCl, 20 mM Tris-HCl, 0.1% SDS, 0.5% sodium deoxycholate, 1 mM EDTA, 1% NP-40), twice with High Salt buffer (500 mM NaCl, 20 mM Tris-HCl, 0.1% SDS, 0.5% sodium deoxycholate, 1 mM EDTA, 1% NP-40), twice with LiCl buffer (250 mM LiCl, 20 mM Tris-HCl, 0.1% SDS, 0.5% sodium deoxycholate, 1 mM EDTA, 1% NP-40) and twice with TE buffer (10 mM Tris-HCl, 1 mM EDTA). Protein-DNA complexes were eluted in elution buffer (1% SDS, 100 mM NaHCO3). The cross-linking DNA of eluted immunocomplexes was reversed by the incubation with proteinase K and NaCl (final concentration: 0.2 M) at 65 °C overnight. The next day, Phenol-chloroform extraction was performed to purify genomic DNA and the level of ACTA2, CNN1, and TAGLN promoters were determined by qPCR.
Isolation of nuclear and cytosolic fraction
HASMCs were washed with cold PBS and incubated with 300 μl lysis buffer A (10 mM HEPES, 0.1 mM EDTA, 1 mM KCl, 50 mM NaF, 0.1 mM EGTA, 1 mM Na3VO4, 1 mM DTT, and protease inhibitor cocktail) for 30 min on ice. Cells were scraped and collected in tubes, then 20 μl of NP-40 was added to each tube and vortex rapidly for 2 min followed by centrifugation at 12,000 × g for 5 min at 4 °C. The supernatant is the cytosolic protein, the precipitation is a nuclear fraction which was further incubated with 60 μl lysis buffer C (20 mM HEPES, 1 mM EDTA, 0.4 M NaCl, 50 mM NaF, 1 mM EGTA, 25% Glycerol, 1 mM DTT and protease inhibitor cocktail) and vortexed every 5 min for a total of 1 h. Finally, the lysate was centrifuged at 12,000 × g for 15 min at 4 °C and the supernatant is the cell nuclear protein.
Quantitative real-time PCR (qPCR)
Total RNA was extracted from cultured cells, mouse tissue, and human sample using RNAiso (Takara). Reverse transcription and cDNA synthesis were performed HiScript® III All-in-one RT SuperMix (Vazyme) and used as a template for qPCR. qPCR was performed by Hieff® qPCR SYBR Green Master Mix. The expression of target genes was normalized against GAPDH. All amplifications were done at least three technical and three biological replicates were performed. The sequence of primers used were shown in Supplementary Table 3.
Western blotting
Total protein from cultured cells, mouse tissue, and human samples were extracted using RIPA lysis buffer supplemented with a protease inhibitor cocktail. Lysates were subjected to sonication and centrifuged at 12,000 × g for 15 min at 4 °C. After centrifugation, the supernatants were collected and calculated protein concentration by BCA assay. Proteins were separated by SDS-PAGE and transferred onto the PVDF membrane (Millipore). Target proteins were incubated with primary antibodies overnight at 4 °C and 2 h at room temperature with secondary antibodies conjugated to HRP. Finally, enhanced chemiluminescence (Thermo) was used to measure protein expression levels. Gel bands were quantified with Image-Pro Plus 6.0 software. The antibodies used for Western blot were shown in Supplementary Table 2.
Scratch assay
HASMCs were plated on 12-well plates. After different treatments, the cells were starved in a serum-free medium for 24 h. After starvation, make a wound by scratching the cell monolayer in a straight line with a sterile P-200 pipette tip in each well. Pictures were taken immediately after scratch and 12 h later. For quantification, the migration area was measured using the Image-Pro Plus 6.0 software.
Fluorescence in situ hybridization (FISH)
In situ hybridization was performed as described in 2. Probes were designed and synthesized by GenePharma (Shanghai, China). The FISH analysis of MEG3 was performed on HASMCs or human aorta tissue and was examined using a FISH kit (GenePharma, China). Briefly, HASMCs were seeded in confocal dishes. After different treatments, cells were fixed with 4% paraformaldehyde for 15 min at room temperature, followed by 0.1% Triton ×-100 incubated for 15 min. The dishes were incubated with prehybridization buffer for 30 min at 37 °C. For the paraffin sections of human aorta tissue, dewaxing was performed first. Then paraffin sections were incubated with protease K at 37 °C for 20 min. Paraffin sections were denatured for 8 min at 78 °C, and dehydrated using graded ethanol for 2 min at room temperature. MEG3 FISH probes were pre-denatured for 5 min at 73 °C. Subsequently, a hybridization buffer with MEG3 FISH probes was added to the dishes or paraffin sections and incubated at 37 °C overnight. The following day, dishes were washed and stained with DAPI, the level of MEG3 was detected by confocal fluorescent microscopy. The sequence of probes used were shown in Supplementary Table 3.
siRNA transfection
Small interfering RNA (siRNA) against target genes and scramble siRNA were synthesized by GenePharma (China). HASMCs were transfected with siRNA (100 nM) via Lipofectamine 3000 Transfection Reagent (Thermo) according to the manufacturer’s instructions. All siRNA oligos used for target genes knockdown were shown in Supplementary Table 3.
Plasmid construction
The pcDNA 3.0 was used for overexpression of RRAS2 (NCBI Reference Sequence: NM_012250.6), ELAVL 1 (NCBI Reference Sequence: NM_001419.3), and MEG3 (NCBI Reference Sequence: NR_046466.1). All the gene synthesis and cloning steps were performed by Genewiz, Inc. (Suzhou, China).
For luciferase reporter assay vectors, a 2.0 kb RRAS2 (Homo sapiens) promoter was synthesized and subcloned into a pGL6 basic luciferase vector (Beyotime Biotechnology) and designated as RRAS2-Luc. Similarly, a total length of 2.0 kb promoter of TAGLN (Homo sapiens) and its truncated fragment for E-BOX element or mutated for INR element were subcloned into pGL6 basic luciferase vector and named TAGLN -Luc-(-2000), TAGLN -Luc-(-1656), TAGLN -Luc-(-1288), TAGLN -Luc-(-716), and TAGLN -Luc-(MUT) respectively. The 3′UTR parts corresponding to RRAS2 3′UTR-full, 3′UTR-A, 3′UTR-B, 3′UTR-C, 3′UTR-D, 3′UTR-E were subcloned into pGL3 basic luciferase vector (Promega).
Proximity-dependent biotin identification
For myc-BirA* proximity-labeling experiments, the myc-BirA* sequence is reported previously42. The RRAS2 sequence was created by gene synthesis and cloned in frame 3′ of myc-BirA* (BirA*-R-Ras2). HASMCs were transfected with BirA*-R-Ras2 fusions plasmid or control plasmid followed by the addition of 50 μmol/L biotin. After 48 h incubation, 4 × 107 cells were washed with PBS twice and lysed with 600 μL lysis buffer (50 mM Tris-HCl, pH 7.4, 500 mM NaCl, 0.2% SDS, 1 mM DTT, 2% Triton, and protease inhibitor cocktail). Cells were sonicated three times for 10 s followed by centrifugation at 16,500 × g for 10 min at 4 °C. The supernatant was collected and an equal volume of lysis buffer was added. To each tube, 50 ml of agarose beads was added and rotated overnight at 4 °C. The next day, tubes were centrifuged and the supernatants were removed. The agarose beads were washed sequentially with 2% SDS, wash buffer 1 (0.1% deoxycholic acid, 1% TritionX-100, 1 mM EDTA, 500 mM NaCl, 50 mM HEPES, pH 7.5) and wash buffer 2 (0.5% deoxycholic acid, 0.5% NP-40, 1 mM EDTA, 250 mM LiCl, 10 mM Tris-HCl, pH 7.4). Then, 50 mM Tris-HCl was added to resuspend the beads, and the beads were boiled at 99 °C for 5 min in an SDS loading buffer. Samples were further analyzed by mass spectrometry or by Western blot.
ChIP-sequencing datasets
The information of TFII I binding sites on the promoter of contractile genes (ACTA2, CNN1, and TAGLN) was based on the ChIP-sequencing datasets (GSE63057) and visualized by NCBI Genome Data Viewer (https://www.ncbi.nlm.nih.gov/genome/gdv/).
In vivo AAV9-mediated MEG3 overexpression
In vivo MEG3 overexpression was performed by AAV9 vectors34. Recombinant adeno-associated virus serotype 9 (AAV9) vectors containing MEG3 driven by murine SM22a promoter, or control vector (AAV9-pCMV-MEG3 or AAV9-pCMV-vector) were constructed by GeneChem Co., Ltd (Shanghai, China). AAV9-pCMV-MEG3/vector (5 × 1011 vector genomes (vg)/mice) were delivered by intravenous injection. Four weeks after AAV9 delivery, MEG3 overexpression was verified by FISH staining.
In vivo Lentivirus-mediated SMC-specific gene overexpression
Mice were infected with lentivirus expressing Rras2 or Vector via tail vein injection as described previously43. The construct was designed so that Cre recombinase induction could be used to mediate the inversion of Rras2 into a sense orientation44. We cloned the expression sequence of R-Ras2-Flag in an inverse, anti-sense orientation between two different pairs of loxP sites, which would be induced by Cre recombinase under the Tagln promoter. The Flag-tagged mouse Rras2 (NM_025846.2) was subcloned into the pLVX-FLEX-EF1a-ZsGreen vector (pLVX-FLEX-R-Ras2-Flag-EF1a-ZsGreen). All the plasmids were constructed by Viraltherapy Technologies Co. (Wuhan, China). Lentivirus was packaged in HEK293T cells and collected from the medium supernatant, and lentivirus particles were concentrated and purified. A total of 5×109 IFU/mL viruses were injected into the tail vein of each Tagln-Cre/ApoE-/−mouse, and an osmotic pump was implanted after 4 weeks.
RNA pull-down
MEG3 pulldown was performed using PierceTM Magnetic RNA-protein pull-down kit (20164, Thermo) according to the manufacturer’s instruction. Briefly, T7 High-Efficiency Transcription Kit (TranseGen, JT101-01) was used for MEG3 full-length (sense), antisense, and truncated fragments in vitro transcription, using templates with specific primers containing a T7 promoter: 5′TAATACGACTCACTATAGGG3′. Subsequently, all nucleotides produced by in vitro transcription were labeled with biotin by Pierce™ RNA 3′ End Desthiobiotinylation Kit (Thermo Fisher Scientific, 20164). A total of 5 × 107 HASMCs were added to 200 μl of IP lysis buffer (25 mM Tris-HCl PH = 7.4, 150 mM NaCl, 5 mM EDTA, 1% NP-40, 15% Glycerol, protease inhibitor, and RNase inhibitor), the lysis was frozen in liquid nitrogen and thawed repetitively for three times. Next, 50 pmol biotin-labeled nucleotides (MEG3 sense, antisense, or truncated fragments) and 50 μl streptavidin magnetic beads were mixed with a 1× RNA Capture Buffer and rotated at room temperature for 30 min. Then, unbound RNA was rinsed with 20 mM Tris (pH = 7.5). Finally, the RNA-protein binding reaction system (10 μl of 10× RNA-protein binding buffer, 30 μl of 50% glycine, and 30 μl of cell lysis buffer were added to streptavidin magnetic beads) was configured according to the kit instructions and rotated at 4 °C for 60 min. Next, beads were washed 3 times with 1 × wash buffer, followed by 50 μl Elution buffer eluted at 37 °C for 30 min and boiled with protein-loading buffer for 5 min. Samples were separated by SDS-PAGE and subsequently further analyzed by Western blotting and Mass spectrometry. Primers used for MEG3 in vitro transcription were listed in Supplementary Table 3.
Histomorphological analysis
H&E staining, Verhoeff’s Van Gieson (EVG) staining, and Masson’s trichrome staining were performed for histomorphological analysis. After sacrifice, mouse abdominal aortas were collected and fixed in 4% paraformaldehyde, followed by embedded in wax and sectioned at 5 µm. Samples were deparaffinized and rehydrated before subjected to H&E staining (Solarbio), EVG staining (Solarbio), and Masson’s trichrome (Solarbio) according to the manufacturer’s protocol. The quantitative analysis of aortic collagen deposition was performed by the Image-Pro Plus 6.0 software.
RNA immunoprecipitation (RIP)
The RIP protocol was performed as previously described with some modifications. Briefly, 5 × 106cells were cross-linked with 0.37% formaldehyde for 10 min and terminated with glycine. Then, cells were lysed in 500 μl lysis buffer (50 mM HEPES at pH 7.5, 0.4 M NaCl, 1 mM EDTA, 1 mM DTT, 0.5% Triton ×-100, 10% glycerol) containing 20 μL PMSF, 5 μl protease inhibitors cocktail (100×) and 5 μl RNase inhibitor (40 U/μL). After sonication and centrifugation, supernate and 5 μg of the antibody against ELAVL 1 (Abcam, #ab9110; control: normal rabbit IgG, Santa Cruz Biotechnology, #sc-2027) were incubated with protein G-agarose beads (GE Healthcare) overnight at 4 °C. The following day, beads were washed with IP lysis buffer five times. Then, 100 μL RIP buffer (50 mM HEPES at pH 7.5, 0.1 M NaCl, 5 mM EDTA, 10 mM DTT, 0.5% Triton ×-100, 10% glycerol, 1% SDS) containing 1 μL RNase inhibitor (40 U/μL) was added to the beads and incubated at 70 °C to reverse the crosslinks. Finally, RNA extraction was performed using RNAiso (Takara) and analyzed by qPCR using the primer sets listed in Supplementary Table 2. The RNA immunoprecipitated by ELAVL 1 was normalized to the input fraction.
Duolink assay (in situ proximity ligation assay) and immunofluorescence
Duolink assay was performed using Duolink In Situ Red Starter Kit (#DUO92101, Sigma-Aldrich) according to the manufacturer’s instructions as described previously49 (PMID: 32929201). Briefly, tissue or cells were washed three times with ice-cold PBS, fixed with 4% paraformaldehyde, permeabilized with 0.5% Triton ×-100, and blocked with 5% BSA in PBS before incubation with primary antibodies (against R-Ras2) at 4 °C overnight. Samples were washed and incubated with secondary antibodies and conjugated with oligonucleotides for 1 h at 37 °C. Ligation solution was added to each reaction after washing, and incubated at 37 °C for 30 min. Texas Red-labeled oligonucleotides and polymerase were then added to each reaction to amplify the previously ligated nucleotide circles. A distinct red spot was visualized as a positive signal, and each spot representing one cluster of protein-protein interaction. Nuclei were stained with DAPI. After mounting, cells were visualized using a Zeiss LSM900 laser microscope (ZEN2010B SP1, Zeiss).
Statistical analysis
Data are analyzed using GraphPad Prism version 9.0.1 (GraphPad Software, San Diego, CA). Continuous variables with normal distribution were expressed as mean ± SEM. Otherwise, data were expressed as median (interquartile range) Shapiro–Wilk test was used for determining the distribution of the data, and the homogeneity of variance was examined by Levene’s Test. Comparisons between two groups were performed using 2-tailed Student’s t-test, Mann–Whitney U or Fisher’s exact test, as appropriate. Differences among groups were evaluated using one-way or two-way ANOVA analysis, followed by Dunnett’s multiple comparisons test or Tukey’s test for post-hoc comparisons. The time-course of blood pressure and vascular lumen diameters change among groups were analyzed by Two-way RM ANOVA followed by Bonferroni’s multiple comparisons test. Differences with a P ≤ 0.05 was considered statistically significant.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The RNA-seq data generated in this study have been deposited in the GEO repository under accession code GSE291516. Mass spectrometry data are available via PRIDE with identifier PXD062059, PXD062094 and PXD062082. All data supporting the findings described in this manuscript are available in the article, in the Supplementary Information, and from the corresponding author upon request. Source data are provided with this paper.
References
Sakalihasan, N., Limet, R. & Defawe, O. D. Abdominal aortic aneurysm. Lancet 365, 1577–1589 (2005).
Golledge, J., Thanigaimani, S., Powell, J. T. & Tsao, P. S. Pathogenesis and management of abdominal aortic aneurysm. Eur. Heart J. 44, 2682–2697 (2023).
Sun, L. Y. et al. Nuclear receptor NR1D1 regulates abdominal aortic aneurysm development by targeting the mitochondrial tricarboxylic acid cycle enzyme aconitase-2. Circulation 146, 1591–1609 (2022).
Riedel, J. H., et al. Glucocorticoids target the CXCL9/CXCL10-CXCR3 axis and confer protection against immune-mediated kidney injury. JCI Insight 8, e160251 (2023).
Wang, J. et al. TCF7L1 accelerates smooth muscle cell phenotypic switching and aggravates abdominal aortic aneurysms. JACC Basic Transl. Sci. 8, 155–170 (2023).
Niihori, T. et al. Germline-activating RRAS2 mutations cause Noonan syndrome. Am. J. Hum. Genet. 104, 1233–1240 (2019).
Capri, Y. et al. Activating mutations of RRAS2 are a rare cause of Noonan syndrome. Am. J. Hum. Genet 104, 1223–1232 (2019).
Weinstock, N. I. & Sadler, L. The RRAS2 pathogenic variant p.Q72L produces severe Noonan syndrome with hydrocephalus: a case report. Am. J. Med. Genet. A 188, 364–368 (2022).
Clavain, L. et al. Characterization of mutant versions of the R-RAS2/TC21 GTPase found in tumors. Oncogene 42, 389–405 (2023).
Janapati, S. et al. TC21/RRas2 regulates glycoprotein VI-FcRgamma-mediated platelet activation and thrombus stability. J. Thromb. Haemost. 16, 1632–1645 (2018).
Martinez-Riano, A. et al. RRAS2 shapes the TCR repertoire by setting the threshold for negative selection. J. Exp. Med. 216, 2427–2447 (2019).
Indolfi, C. et al. Inhibition of cellular Ras prevents smooth muscle cell proliferation after vascular injury in vivo. Nat. Med. 1, 541–545 (1995).
Ueno, H., Yamamoto, H., Ito, S., Li, J. J. & Takeshita, A. Adenovirus-mediated transfer of a dominant-negative H-ras suppresses neointimal formation in balloon-injured arteries in vivo. Arterioscler Thromb. Vasc. Biol. 17, 898–904 (1997).
Work, L. M. et al. Short-term local delivery of an inhibitor of Ras farnesyltransferase prevents neointima formation in vivo after porcine coronary balloon angioplasty. Circulation 104, 1538–1543 (2001).
Komatsu, M. & Ruoslahti, E. R-Ras is a global regulator of vascular regeneration that suppresses intimal hyperplasia and tumor angiogenesis. Nat. Med. 11, 1346–1350 (2005).
Lu, H. et al. Cyclodextrin prevents abdominal aortic aneurysm via activation of vascular smooth muscle cell transcription factor EB. Circulation 142, 483–498 (2020).
Lu, H. et al. Hypercholesterolemia induced by a PCSK9 gain-of-function mutation augments angiotensin II-induced abdominal aortic aneurysms in C57BL/6 mice-brief report. Arterioscler Thromb. Vasc. Biol. 36, 1753–1757 (2016).
Lu, G. et al. A novel chronic advanced stage abdominal aortic aneurysm murine model. J. Vasc. Surg. 66, 232–242 e234 (2017).
Salarian, M. et al. Homeostatic, non-canonical role of macrophage elastase in vascular integrity. Circ. Res. 132, 432–448 (2023).
Yodsanit, N. et al. Targeted PERK inhibition with biomimetic nanoclusters confers preventative and interventional benefits to elastase-induced abdominal aortic aneurysms. Bioact. Mater. 26, 52–63 (2023).
Vega-Cuesta, P. et al. Ras2, the TC21/R-Ras2 Drosophila homologue, contributes to insulin signalling but is not required for organism viability. Dev. Biol. 461, 172–183 (2020).
Novina, C. D., Cheriyath, V. & Roy, A. L. Regulation of TFII-I activity by phosphorylation. J. Biol. Chem. 273, 33443–33448 (1998).
Hong, M. et al. Transcriptional regulation of the Grp78 promoter by endoplasmic reticulum stress: role of TFII-I and its tyrosine phosphorylation. J. Biol. Chem. 280, 16821–16828 (2005).
Egloff, A. M. & Desiderio, S. Identification of phosphorylation sites for Bruton’s tyrosine kinase within the transcriptional regulator BAP/TFII-I. J. Biol. Chem. 276, 27806–27815 (2001).
Roy, A. L. Signal-induced functions of the transcription factor TFII-I. Biochim. Biophys. Acta 1769, 613–621 (2007).
Roux, K. J., Kim, D. I., Raida, M. & Burke, B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 196, 801–810 (2012).
Boriack-Sjodin, P. A., Margarit, S. M., Bar-Sagi, D. & Kuriyan, J. The structural basis of the activation of Ras by Sos. Nature 394, 337–343 (1998).
Hall, B. E., Yang, S. S., Boriack-Sjodin, P. A., Kuriyan, J. & Bar-Sagi, D. Structure-based mutagenesis reveals distinct functions for Ras switch 1 and switch 2 in Sos-catalyzed guanine nucleotide exchange. J. Biol. Chem. 276, 27629–27637 (2001).
Poller, W. et al. Non-coding RNAs in cardiovascular diseases: diagnostic and therapeutic perspectives. Eur. Heart J. 39, 2704–2716 (2018).
Herman, A. B., Tsitsipatis, D. & Gorospe, M. Integrated lncRNA function upon genomic and epigenomic regulation. Mol. Cell 82, 2252–2266 (2022).
Grammatikakis, I. & Lal, A. Significance of lncRNA abundance to function. Mamm. Genome 33, 271–280 (2022).
Bridges, M. C., Daulagala, A. C. & Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 220, e202009045 (2021).
Ripin, N. et al. Molecular basis for AU-rich element recognition and dimerization by the HuR C-terminal RRM. Proc. Natl. Acad. Sci. USA 116, 2935–2944 (2019).
Zeng, Z. et al. Circular RNA CircMAP3K5 acts as a microRNA-22-3p sponge to promote resolution of intimal hyperplasia via TET2-mediated smooth muscle cell differentiation. Circulation 143, 354–371 (2021).
Simanshu, D. K., Nissley, D. V. & McCormick, F. RAS proteins and their regulators in human disease. Cell 170, 17–33 (2017).
Moore, A. R., Rosenberg, S. C., McCormick, F. & Malek, S. RAS-targeted therapies: is the undruggable drugged?. Nat. Rev. Drug Discov. 19, 533–552 (2020).
Hong, D. S. et al. KRAS(G12C) inhibition with sotorasib in advanced solid tumors. N. Engl. J. Med. 383, 1207–1217 (2020).
McCleary-Wheeler, A. L. et al. TFII-I-mediated polymerase pausing antagonizes GLI2 induction by TGFbeta. Nucleic Acids Res. 48, 7169–7181 (2020).
Mammoto, A. et al. A mechanosensitive transcriptional mechanism that controls angiogenesis. Nature 457, 1103–1108 (2009).
Michalik, K. M. et al. Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth. Circ. Res. 114, 1389–1397 (2014).
Cremer, S. et al. Hematopoietic deficiency of the long noncoding RNA MALAT1 promotes atherosclerosis and plaque inflammation. Circulation 139, 1320–1334 (2019).
Piccoli, M. T. et al. Inhibition of the cardiac fibroblast-enriched lncRNA Meg3 prevents cardiac fibrosis and diastolic dysfunction. Circ. Res. 121, 575–583 (2017).
Vacante, F. et al. CARMN loss regulates smooth muscle cells and accelerates atherosclerosis in mice. Circ. Res. 128, 1258–1275 (2021).
Chen, M. et al. 3’ UTR lengthening as a novel mechanism in regulating cellular senescence. Genome Res. 28, 285–294 (2018).
Hortal, A. M. et al. Overexpression of wild type RRAS2, without oncogenic mutations, drives chronic lymphocytic leukemia. Mol. Cancer 21, 35 (2022).
Xu, J., Wang, X., Zhu, C. & Wang, K. A review of current evidence about lncRNA MEG3: a tumor suppressor in multiple cancers. Front. Cell Dev. Biol. 10, 997633 (2022).
Luo, W. et al. Critical role of cytosolic DNA and its sensing adaptor STING in aortic degeneration, dissection, and rupture. Circulation 141, 42–66 (2020).
Zhao, G., et al. BAF60c prevents abdominal aortic aneurysm formation through epigenetic control of vascular smooth muscle cell homeostasis. J. Clin. Invest. 132, e158309 (2022).
Hou, J. et al. PD-L1-mediated gasdermin C expression switches apoptosis to pyroptosis in cancer cells and facilitates tumour necrosis. Nat. Cell Biol. 22, 1264–1275 (2020).
Acknowledgements
Lentivirus expressing Rras2 was designed with the help of Professor Jingjing Ben from Department of Pathophysiology, Key Laboratory of Targeted Intervention of Cardiovascular Disease, Nanjing Medical University. This work is supported by the grants from National Health Commission (NHC) Major Special Program on the Four Major Chronic Diseases received by Y.J. (2023ZD0503100), National Natural Science Foundation of China (8224121, 82121001 received by Y.J., 82270504 and 82422071 received by S.S.L., and 82222009 received by L.P.X.). Open Project Fund of the State Key Laboratory of Frigid Zone Cardiovascular Disease, Ministry of Science and Technology (HDHY2024001 received by S.S.L.).
Author information
Authors and Affiliations
Contributions
Y.J., L.P.X., Y.H., and S.S.L. designed and supervised the study. J.Y.C. and S.S.L. performed data analysis and drafted the manuscript. J.Y.C., K.Z., and R.L. performed the in vivo experiments. X.Q.W., S.X.S., and Y.Z. performed the in vitro experiments. T.Y.S., Z.X.S., and X.L.T. helped to collect the clinical samples. Z.R.Z., B.Y., J.W.T., L.L., A.H.G., F.C., D.J.W., and J.P. provided suggestions about the study.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Baohui Xu, who co-reviewed with Jiayu Ye; Lars Maegdefessel and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Luo, S., Chen, J., Zhong, K. et al. Ras-related protein 2 limits vascular smooth muscle cell phenotypic switching and abdominal aortic aneurysm development. Nat Commun 16, 9460 (2025). https://doi.org/10.1038/s41467-025-64496-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-64496-7
