
Abstract
The ocean’s biological carbon pump (BCP) plays a crucial role in climate regulation by facilitating long-term removal of atmospheric CO2. However, BCP changes during past hyperthermals remain poorly understood. Here we use the vertical carbon isotope (δ13C) gradients between surface and mid-waters to trace the BCP changes during the Permian−Triassic, early Toarcian, and Palaeocene−Eocene hyperthermals, detecting a two- to three-fold increase in the vertical δ13C gradients. We found that enhanced organic matter remineralization, driven by increased microbial respiration under warming, are the primary trigger for increased vertical δ13C gradients. Warming-induced enhancements in microbial metabolism caused a two- to six-fold reduction in the transfer efficiency of particulate organic carbon from the surface to the deep ocean during the three hyperthermals, potentially reducing carbon sequestration in the deeper ocean interior. This implies that the warming-induced enhancement of microbial metabolism may have amplified global warming during ancient hyperthermals via a positive feedback mechanism.
Similar content being viewed by others


Introduction
Rapid global warming events in the Phanerozoic, referred to as ancient hyperthermals, serve as analogues for predicting future climate changes1,2. These hyperthermal events3,4,5,6 are linked to remarkable perturbations in the global carbon cycle, as evidenced by large negative carbon isotope excursions (CIEs), rising atmospheric CO2 levels (pCO2)7,8,9, and biotic extinctions10. The rapid and substantial temperature rises are typically attributed to the release of massive quantities of CO2 from volcanic activities11,12, or biogenic methane emissions from the seafloor13,14. While these mechanisms emphasise carbon emissions as the primary driver of temperature increases, the role of the ocean’s biological carbon pump (BCP) in natural carbon sequestration and climate regulation is often overlooked.
The ocean’s BCP involves carbon fixation via photosynthesis at the surface, followed by the sinking of particulate organic carbon (POC) undergoing microbial respiration (remineralization) in the ocean interior, thereby cycling carbon and nutrients back to the surface15,16. This downward transport process stores carbon within the ocean interior where carbon can be sequestered on centennial and longer timescales17,18. Transfer efficiency, defined as the fraction of POC transported from the surface ocean (0 − 100 m) to the ocean interior, serves as a widely used proxy for estimating the biological carbon pump sequestration19,20,21. Consequently, projecting the transfer efficiency of POC within the BCP in response to warming is essential to understand future climate change21,22.
Previous studies on changes in the BCP during ancient hyperthermals have mainly focused on changes in primary productivity or POC export production in surface water in response to rapid temperature rises23,24,25,26,27. These studies suggest either a weakened biological pump due to ocean stagnation caused by intensified stratification23,24 or a much stronger biological pump with high productivity driven by increased nutrients due to warming-induced intensive weathering25,26,27. However, little is known about the transfer efficiency of POC from the surface to the deep ocean during ancient hyperthermals, which hinders a clear understanding of the feedbacks between oceanic carbon sequestration and climate change across the Phanerozoic.
Temperature is widely recognized as a key factor that influences the rates of microbial photosynthesis and respiration28,29. Previous studies have demonstrated that respiration rates can double or triple with a ~10 °C increase in ambient temperature28,29. Such temperature-driven changes in microbial remineralization strongly affect the degradation of sinking organic matter30,31. Together with ocean circulation and water mass ventilation, which regulate circulation timescales and pathways of sinking organic carbon, these processes collectively determine the transfer efficiency of POC21,32,33. Meanwhile, the vertical δ13C gradients of dissolved inorganic carbon (DIC), i.e., the decrease in DIC δ13C values (δ13CDIC) with increasing depth in the upper ocean (> 1000 m), are mainly controlled by the remineralization of particulate organic matter (POM) that releases isotopically lighter carbon at greater depths32. Therefore, vertical δ13C gradients are important geochemical indicators for tracing the transformation and remineralization of POC33,34,35. Here, we integrate proxy δ13CDIC and simulated δ13CDIC in an Earth system model to investigate changes in vertical δ13CDIC gradients during three key Phanerozoic hyperthermals4,10: the Permian−Triassic Thermal Maximum (PTTM; ~251.9 Ma), the early Toarcian Thermal Maximum in the Early Jurassic (ETTM; ~183.0 Ma), and the Palaeocene−Eocene Thermal Maximum (PETM; ~56.0 Ma). Utilising data-constrained cGENIE (carbon-centric Grid ENabled Integrated Earth system model), we evaluated the response of POC transfer efficiency to elevated temperatures during ancient hyperthermal events.
Results and Discussion
Increased vertical δ13CDIC gradients during ancient hyperthermals
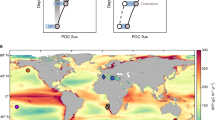
The three ancient hyperthermals examined in this study feature prominent and rapid global temperatures rises and well-documented carbon isotopes from globally distributed sites, which allows for the reconstruction of δ13CDIC depth profiles in response to warming. This is in contrast to other ancient hyperthermals with relatively smaller, slower temperature rate increases10 and/or poorly documented carbon isotope data (Fig. 1). The PTTM (also known as the Permian−Triassic mass extinction) is marked by ~10 °C increase in sea surface temperatures (SSTs) estimated from low-latitude conodont oxygen isotopes6 and coeval negative CIEs (~4‰) recorded in global marine carbonates (δ13Ccarb)7. During the ETTM (also known as the early Toarcian oceanic anoxic event), oxygen isotopes and Mg/Ca ratios of the belemnite from the middle-latitude European Shelf show a ~7 °C increase in SST, coinciding with a ~3‰ negative CIE in marine carbonates3,8. Biomarker proxy data based on TEX86 (TetraEther indeX of tetraethers consisting of 86 carbon atoms) from low-latitude regions indicate a ~4 °C increase in SST during the PETM, which is further supported by Mg/Ca and oxygen isotope data5. Moreover, foraminifera shells exhibit a ~2.5‰ negative CIE during the PETM9. The pronounced negative CIEs recorded in marine carbonates and foraminifera shells allow for the reconstruction of δ13CDIC depth profiles in the upper ocean.

a Global average temperature (GAT) across the last 300 million years4. b Stacked marine carbon isotopes of carbonate (δ13Ccarb) and foraminifera (δ13Cforam) and reconstructed sea surface temperature (SST) from oxygen isotopes of conodont, belemnite and TEX86 (TetraEther indeX of tetraethers consisting of 86 carbon atoms) during the Permian−Triassic Thermal Maximum (PTTM)7, early Toarcian Thermal Maximum (ETTM)8 and Palaeocene−Eocene Thermal Maximum (PETM)5,9. The blue and red lines represent the LOESS (locally estimated scatterplot smoothing) fit curve for δ13C and SST, respectively, while light blue and red shaded area represent 68% confidence intervals. c Simulated SST during the pre-CIE and peak CIE phases for the hyperthermal events from our simulations. Black dashed boxes show the locations of the proxy and model SST data shown in Fig. 1b. Abbreviations: CIE carbon isotope excursion; C. c Clarkina changxingensis; C. y Clarkina yini; C. m Clarkina meishanensis; H. c Hindeodus changxingensis; C. t Clarkina taylorae; H. p Hindeodus parvus; I. Isarcicella; D. Dactylioceras.
Here, we compiled published δ13Ccarb data from 174 locations (Supplementary Fig. 1; Supplementary Data 1; Supplementary Table 1) and identified three depth-dependent depositional settings, including shallow shelf (0−80 m, average ~40 m), deep shelf (80−180 m, average ~130 m), and basin (180−280 m, average ~230 m) facies. The compiled δ13CDIC data from the modern ocean demonstrate that δ13CDIC values from continental shelf environments provide a robust proxy of the global ocean’s carbon isotopic composition (Supplementary Fig. 2). These depths correspond to the depths of the three uppermost ocean grid layers in the cGENIE Earth system model36 for direct model-data comparison (a sensitivity test for different scenarios of assumed depth is provided in Supplementary Table 2). Then, we reconstructed δ13Ccarb depth profiles during the pre-CIE (pre-warming) and the peak CIE (peak warming) phases but focused on global stacked δ13Ccarb depth profiles (Fig. 2 and Supplementary Figs. 3–6), because δ13Ccarb data from some oceanic regions are insufficient to reconstruct local gradients. One inherent challenge in reconstructing the vertical δ13CDIC gradients is local carbonate diagenesis, including that induced by meteoric water and burial processes, which can produce anomalously large CIE37. We vetted our dataset by removing δ13Ccarb data altered by diagenesis and focused on relative changes to minimise the effects of diagenesis in our analysis (see “Methods”). Moreover, the average δ13C values in surface water (shallow shelf) were set as the reference standard and were adjusted to 0‰, while other proxy data were recalculated as offsets to eliminate isotopic differences between oceanic DIC and carbonate resulting from carbon isotope fractionation and carbonate diagenesis (see “Methods”). Surface water δ13CDIC values were normalised to zero for both proxy records and model scenarios to facilitate consistent intercomparisons. Planktonic foraminiferal shells can record oceanic δ13CDIC when using constrained shell size fractions to minimise physiological effects (see “Methods”), because smaller-size shells typically show lower δ13C and δ18O values due to vital effects34. Although planktonic foraminifera flourished after the Jurassic38 and, therefore, data are only available for the PETM, they provide additional evidence to support the observations obtained from δ13Ccarb data (see “Methods”).

Proxy data are from marine carbonate δ13C of three depth-dependent depositional settings (Supplementary Data 1) during the three hyperthermals: Permian−Triassic, early Toarcian and Palaeocene−Eocene Thermal Maximum. The average of simulated δ13C of dissolved inorganic carbon (δ13CDIC) is calculated from low to middle latitudes (48.6° S to 48.6° N), where most sections are located (Supplementary Fig. 1). Both proxy and modelled δ13CDIC values are adjusted as an offset from the average δ13CDIC in surface water (Methods). Error bars represent the one standard deviation. a–c Proxy (grey) and simulated δ13CDIC depth profiles at pre-carbon isotope excursion (CIE) phases with low temperatures for “microbial metabolism” (red), “eutrophication” (blue) and “circulation change” (green) scenarios. d–f The proxy and simulated δ13CDIC depth profiles at peak CIE phases with global warming for “microbial metabolism”, “eutrophication” and “circulation change” scenarios. g–i Vertical δ13CDIC gradients (mean ± 1 s.d.) are calculated from the δ13C difference between surface water (~40 m) and mid-water (~130 m), expressed as the δ13CDIC change per 100 m of water depth (‰/hm).
Compiled global δ13Ccarb records exhibit a pronounced negative carbon isotope excursion at three water depth levels (Supplementary Fig. 7; and Supplementary Table 1), which is commonly attributed to the injection of large amounts of 13C-depleted carbon into the atmosphere-ocean system11,12. Although δ13C variations in the ocean-atmosphere carbon inventory would theoretically induce equivalent isotopic shifts in both surface and deep waters, our data indicate that the CIE magnitude in the deep shelf or basin is greater than that in the shallow shelf (e.g., PTTM: 4.5‰ vs. 3.5‰). This discrepancy mainly attributed to an enhanced vertical δ13C gradient39,40. Our compiled global δ13Ccarb records show a two- to three-fold increase in the vertical δ13CDIC gradients between pre-CIE and peak CIE phases (Fig. 2; and Supplementary Table 1). Specifically, δ13Ccarb data indicate that the vertical δ13CDIC gradient rose from 0.5 ± 1.6‰/hm (pre-CIE) to 1.7 ± 1.7‰/hm (peak CIE) across the PTTM, from 1.7 ± 1.3‰/hm to 2.6 ± 1.5‰/hm during the ETTM, and from 1.0 ± 0.9‰/hm to 2.8 ± 1.9‰/hm during the PETM. An increase in the vertical δ13CDIC gradient was also recorded in the δ13C record of planktonic foraminifera during the PETM (Supplementary Figs. 8 and 9). Similar trends have also been observed in marine carbonates from depth-dependent depositional settings in South China during the PTTM40,41 as well as in planktonic foraminiferal shells from the warm early Eocene and mid-Miocene34,42, further supporting the global pattern of increasing vertical δ13CDIC gradients during these ancient hyperthermals.
Considerable δ13C variability exists even within the same depositional settings (e.g., 3.8‰ in shallow shelf during the P–Tr interval), exceeding the 2–3‰ variability observed in modern δ13CDIC (Supplementary Fig. 2). This greater variability may reflect mixing of DIC sources and fluctuating paleodepths when carbonate δ13C is used as a proxy. Both factors contribute to uncertainty in estimates of vertical δ13C gradients, potentially making changes in gradients statistically insignificant. Firstly, the δ13Ccarb values recorded in deep shelf and basin environments potentially represent a mixture of deep and shallow water DIC sources43. To evaluate potential mixing effects on gradient amplification, we implemented a simple isotopic mixing model (see “Methods”). Sensitivity tests reveal that increased gradient during the warming, as indicated by the δ13Ccarb data, show limited dependence on mixing source (Supplementary Fig. 10). The estimates (shown in Fig. 2) without regard to mixed sources provide conservative values, as the magnitude increase when raising the mixing proportion (Supplementary Fig. 10). Furthermore, in order to evaluate the potential uncertainty resulting from palaeodepth fluctuations (shallowing or deepening) at locations, we simulated 2000 different palaeodepth scenarios for each hyperthermal event to assess the vertical gradient responses to depth changes during the event (see “Methods”), as these changes could not be reliably quantified through existing proxy data. Results demonstrate that vertical gradients during the peak CIE stage remained significantly enhanced in most scenarios (Supplementary Figs. 11 and 12), though gradient magnitudes diminished, as more locations exhibited depth changes. Collectively, the amplified vertical δ13CDIC gradients documented across widely distributed sites and multiple geological intervals strongly suggest that local effects, including palaeodepth variations, mixed DIC sources, or diagenetic alterations, do not constitute the dominant control.
Testing the drivers of increased vertical δ13CDIC gradients during hyperthermals
To test the mechanistic drivers of the increased vertical δ13CDIC gradients during the three investigated hyperthermals, we conducted a series of experiments using the cGENIE Earth system model (Fig. 2; Supplementary Table 4). We prescribed a range of atmospheric pCO2 values (0.5–9× pre-industrial level, 140–2520 ppmv) and obtained simulated SSTs to match the proxy SST data from the low- to mid-latitudes (Fig. 1a). Previous studies suggest that vertical δ13CDIC gradients are mainly controlled by ocean nutrient conditions41, thermohaline circulation40, and temperature-induced changes in microbial metabolism39; however, a comprehensive assessment of their roles in driving δ13CDIC gradients during hyperthermals through modelling is lacking. Accordingly, we constructed three scenarios for each hyperthermal event—circulation change, eutrophication, and a warming-driven increase in microbial metabolism (hereafter, microbial metabolism). The “circulation change” scenario was simulated solely by temperature-driven changes in physical ocean circulation with invariant oceanic phosphate levels (1× the modern [PO4] level) between pre-CIE and peak CIE phases, whereas in the “eutrophication” scenario, increased [PO4] (2× the modern level following previous cGENIE experiments39) was set at the peak CIE. Both the “circulation change” and “eutrophication” scenarios were run using a static biological pump, in which global POM export production and remineralization are almost entirely controlled by nutrient availability (here, [PO4])26. In contrast, the “microbial metabolism” scenario used a constant 1× [PO4] and a dynamic biological pump, controlled by both nutrient availability and temperature-dependent metabolic rates. In particularly, the dynamic model incorporates a temperature-dependent scheme for microbial remineralization of organic matter to regulate POC flux in mid-waters44. In the “microbial metabolism” scenario, we reduced the POM sinking rate by ~22% in the PTTM experiments, following Hülse et al. based on marine animal body-size data39 (see “Methods”) to represent a shallower remineralization depth before the early Mesozoic. This is because modern marine phytoplankton (e.g., coccolithophorids, dinoflagellates, and diatoms) did not flourish until the Late Triassic38. The sinking rates for the ETTM and PETM were set at modern levels following previous modelling experiments22,34.
Our model shows that vertical δ13CDIC gradients increase significantly during the peak CIE in the “microbial metabolism” scenario, matching proxy data better than the other two scenarios (Fig. 2). In this scenario, the differences between proxy and model data at peak CIE phase are −0.3 ± 2.7‰/hm for the PTTM, 0.7‰/hm ± 2.4‰/hm for the ETTM, and −0.04 ± 2.9 ‰/hm for the PETM, while those for the other two scenarios are notably larger (1.0–2.3 ‰/hm) (Supplementary Table 4). This is further supported by root mean square error calculations that show the lowest deviation between proxy and model data under the “microbial metabolism” scenario (Supplementary Table 4). However, the modelled vertical δ13CDIC gradient in the “microbial metabolism” scenario does not fully account for the observed data, possibly due to uncertainties in model boundary conditions (e.g., set temperature values) and/or proxy data (e.g., local diagenesis or mixing of deep and shallow water DIC37).In particular, there are large discrepancies between the proxy data (most from European shelf) and model results during the peak CIE phase of the ETTM (Fig. 2b, e). This mismatch likely arises from the relatively low spatial resolution of the cGENIE model, which does not adequately capture the restricted conditions of the European shelf and therefore fails to reproduce the 13C-depleted DIC at mid-water depths observed in the proxy data.
Our best-fit model scenario (i.e., “microbial metabolism”) supports the proposition that temperature-dependent microbial metabolism has a significant impact on vertical δ13CDIC gradients while nutrients and ocean circulation are of secondary importance (Fig. 2; and Supplementary Fig. 6). Specifically, increased ocean temperatures during hyperthermals may have increased microbial respiration and resulted in a higher remineralization rate of sinking organic matter in mid-waters, generating more 13C-depleted DIC and larger vertical δ13CDIC gradients (Supplementary Fig. 13). Our simulation highlights the impacts of temperature-dependent microbial metabolism on vertical δ13CDIC gradients, which have been overlooked in previous studies (e.g., refs. 40,41), while not excluding the effects of nutrients and ocean circulation33,45. The “eutrophication” scenario demonstrates that doubled nutrient levels stimulate productivity (Supplementary Fig. 14), concurrently increasing the vertical δ13CDIC gradient. However, this effect remains substantially weaker than the gradient increase induced by the “microbial metabolism” scenario (Fig. 2). Only extremely high nutrient levels (e.g., 10× [PO4]41) or regionally limited vertical mixing (e.g., in the modern Black Sea40) could generate large vertical δ13CDIC gradients. Additionally, sensitivity experiments under the “microbial metabolism” scenario reveal that the reduced POC sinking rate in the cGENIE model exhibit limited influence on the gradients (Supplementary Fig. 15), reinforcing the primacy of temperature-dependent microbial metabolism.
Temperature-dependent microbial metabolism as a positive feedback
Our simulations indicate an increase in POC export production in surface water during the three studied hyperthermals, particularly in the “microbial metabolism” and “eutrophication” scenarios compared to a minimal change in the “circulation change” scenario (Fig. 3a and Supplementary Fig. 14). In the “microbial metabolism” scenario, 122%, 83%, and 40% increases in POC export production are associated with the PTTM, ETTM, and PETM, respectively (Fig. 3a). A doubling of oceanic phosphate concentrations in the “eutrophication” scenario results in a ~100% increase in POC export production (Supplementary Fig. 14). Some model26 and proxy25,27,46 studies also support a rise in primary production or export production during these hyperthermals as a result of increased nutrient input (e.g., phosphate), driven by intensified weathering, similar to our “eutrophication” scenario. However, in the “microbial metabolism” scenario—as the best-fit model scenario supported by vertical δ13CDIC gradients (Fig. 2)—the increased POC export production is primarily attributed to more recycled [PO4] returning to the surface water due to higher remineralization rates. This effect is further enhanced by warming-induced increases in photosynthetic rates, leading to higher productivity (Fig. 4). The “microbial metabolism” scenario incorporates the temperature dependency of the nutrient uptake rate in the model44. In contrast, the static model used in the “circulation change” and “eutrophication” scenarios does not account for temperature dependency (Supplementary Fig. 14). This implies that temperature-driven nutrient recycling plays a crucial role in stimulating POC export production.

a Global mean particulate organic carbon (POC) exported at surface water (40 m) from PTTM (red), ETTM (blue) and PETM (green). b Global mean POC flux at mid-water (1040 m). c POC transfer efficiency (i.e., the ratio of POC at the mid-water to those at the surface water). The pre-industrial levels (grey) estimated by cGENIE model are from previous work22. CIE carbon isotope excursion; PTTM Permian−Triassic Thermal Maximum; ETTM early Toarcian Thermal Maximum; PETM Palaeocene−Eocene Thermal Maximum.

a Ocean’s biological carbon pump processes during the pre-CIE phase. b A potential reduction in carbon sequestration by biological carbon pump serves as positive feedback to global warming triggered by massive carbon emission. CIE carbon isotope excursion; DIC dissolved inorganic carbon; POC particulate organic carbon.
Export production is an important metric for evaluating the sequestration capacity of the biological carbon pump15,47. However, quantifying export production alone is insufficient to assess the biological pump’s impact on atmospheric CO2, as its efficiency also depends on the residence time of exported carbon in the ocean17,18,19. Specifically, carbon in the upper ocean (< 1000 m) typically returns to the atmosphere on sub-centennial timescales, limiting its contribution to long-term CO2 storage, whereas carbon transported to the deep ocean (> 1000 m) or buried in sediments can remain sequestered for much longer periods20. Thus, the POC transfer efficiency at ~1000 m is critical for predicting ocean carbon sequestration20,21. Accordingly, we focused on the ratio of the POC flux at ~1040 m to that at the surface in the model as a proxy for the global mean transfer efficiency of POC21.
Despite the increased POC export production in the surface water, our results indicate that this compensatory flux is insufficient to fully offset the reduction in mid-water POC flux due to warming-induced increases in microbial respiration, leading to a significant decrease in POC transfer efficiency (Fig. 3b, c). Simulations showed a two- to six-fold decrease in POC transfer efficiency during warming, from 9.2% to 1.5% for the PTTM, 13.8% to 4.2% for the ETTM, and 1.9% to 0.9% for the PETM (Fig. 3c). Sensitivity analyses of other mid-water depths (~350 m and ~640 m) also revealed a significant decrease in transfer efficiency (Supplementary Fig. 16). Along with our sensitivity analyses of ocean temperature, the independence of transfer efficiency from [PO4] (Supplementary Fig. 17) and spatial pattern of higher transfer efficiency in the high latitudes in our simulations (Supplementary Fig. 18) further support the idea that transfer efficiency is inversely correlated with ocean temperature. Our spatial pattern is consistent with the results of data-assimilated biological pump models and modern field observations31, though quantitative discrepancies persist with direct sediment trap measurements20. This discrepancy potentially arises from the sole temperature-dependent remineralization process incorporated in our model, without considering zooplankton migration48 or size-dependent plankton communities49.
The primary triggers for rapid temperature rise during hyperthermals are typically massive volcanic CO2 and thermogenic methane release from large igneous provinces, with possible contributions from felsic volcanisms11,12,50, biogenic methane emissions from the seafloor13,14, and enhanced reverse weathering driven by the collapse of marine siliceous organisms51. Biological carbon pumps play a pivotal role in sequestering carbon emissions and reducing atmospheric pCO2. However, a reduction in POC transfer efficiency due to a warming-induced increase in microbial metabolism—as indicated by our model results (Figs. 3 and 4)—potentially lead to less CO2 sequestration in the ocean interior. This process may create potential positive feedback in amplifying the greenhouse effect during the three studied hyperthermal events, which persisted for hundreds of thousands to millions of years (Fig. 1).
We further perform “multi-factor” experiments varying [PO4] and temperature dependency simultaneously to investigate the interplay between nutrient-dependent productivity and temperature-dependent microbial metabolism on the BCP. Model results reveal that these competing drivers collectively reduce mid-water POC flux during the PTTM, while enhancing fluxes during the ETTM and PETM, despite maintaining consistently reduced POC transfer efficiency across all events (Supplementary Fig. 19). This opposite mid-water POC flux pattern may account for the contrasting OC burial responses observed among different hyperthermals52,53,54,55. Some hyperthermal events are associated with increased organic carbon (OC) burial, such as the widespread black shale deposition observed in the Toarcian OAEs52. However, there are also contrasting examples, such as the sudden decline in OC accumulation rates in South China during the PTTM53 and the suppression of global OC burial during mid-Miocene warming54. These contrasting observations of OC burial may imply that competitive factors control OC burial during hyperthermals56. Ocean anoxia, together with higher productivity triggered by increased weathering, has been widely used to explain elevated OC burial during OAEs52. In contrast, warming-induced increases in microbial metabolism led to reduced POC transfer efficiency and lower POC flux within the ocean interior, which likely have a countereffect on OC burial54. However, our simulated mid-water POC fluxes should not be conflated with OC burial, critical preservation processes of OC are not involved in our experiments57. A systematic integration of proxy data (e.g., TOC) with models explicitly incorporating OC burial processes will be essential to elucidate how these warming-induced competitive factors control OC burial across different hyperthermals.
Implications for BCP change and future climate
Our findings highlight that temperature-dependent microbial metabolism may significantly influence BCP changes and thus climate regulation. Specifically, a temperature increase of 4–10°C during three ancient hyperthermals could reduce transfer efficiency of BCP by 2–6 times, thereby lowering the flux of POC into the ocean interior and potentially counteracting OC burial (Fig. 3). This feedback likely resulted in reduced CO2 sequestration in the ocean interior, amplifying ancient warming (Fig. 4). Conversely, a cooling-induced decline in microbial metabolism increased the efficiency of BCP over the past 15 Myr34. It should be noted, however, this critical feedback becomes more effective over longer timescales, as the effects of organic carbon burial on carbon cycling typically become apparent over million-year timescales57. Consequently, the amplification effect during the PETM may have been relatively limited due to its shorter duration than those of the PTTM and ETTM (Fig. 1), which restricted the establishment of sustained positive feedbacks. Furthermore, a key limitation of our simulations is the inability to quantify how the 2–6-fold reduction in transfer efficiency decreases the amount of CO2 sequestered in the ocean by the BCP over geological timescales, which should be addressed in future work.
Despite this limitation of our simulation, changes in the BCP during ancient hyperthermals in the Phanerozoic provide a strong analogue for the future BCP and anthropogenic climate change. Projections of POC transfer efficiency in response to 21st century climate change exhibit conflicting results across the Coupled Model Intercomparison Project Phase 6 (CMIP6) ensembles21. These discrepancies reflect diverse physical and biogeochemical processes represented by the CMIP6 model. The cGENIE model incorporates temperature-dependent microbial processes, particularly temperature-dependent remineralization (see “Methods”)21. Based on this, previous study using the cGENIE model project that globally averaged transfer efficiencies at a depth of 1040 m will decline from ~11% to 4–9% under various carbon emission scenarios22. With a smaller fraction of organic matter reaching the ocean interior, the food supply and marine ecosystem in mid-waters are likely altered22. Moreover, a reduced amount of carbon transported to the seafloor and buried in sediments potentially creates a positive feedback loop, exacerbating global warming on long timescales, similar to the three ancient hyperthermals studied. However, this positive feedback on carbon sequestration depends on timescales. Model projections suggest a net increase in oceanic carbon storage by 2100 due to short timescales (< 100 years), despite a decrease in transfer efficiency21. Furthermore, this study does not account for other factors, including trait-based functional types of the plankton community49, oxygen-dependent processes58, and mineral ballasting effects59, all of which may influence transfer efficiency. Nevertheless, our integrated approach combining proxy data compilation with mechanistic modelling highlights that temperature-dependent microbial metabolism may play a significant role in modulating the BCP dynamics during both ancient hyperthermal events and future climate scenarios.
Methods
Carbonate carbon isotope compilation
We collected marine carbonate carbon isotope (δ13Ccarb) profiles from various water depth-dependent depositional settings on carbonate platforms or ramps to represent the δ13CDIC in the upper ocean (Supplementary Data 1 and references in Supplementary Information). Although our compiled δ13Ccarb data are derived primarily from continental shelf settings and lack pelagic records, modern δ13CDIC data demonstrate that δ13C values from continental shelf environments provide a robust representation of the global ocean (Supplementary Fig. 2). The selected δ13Ccarb profiles adhered to the following principles: (1) The δ13Ccarb data should predominantly be extracted from carbonate platforms or ramps to represent the upper ocean without drilling cores from the deep sea; (2) some of the extremely large CIE magnitudes, such as −26‰ caused by local authigenic carbonate at the Salisbury Embayment in the USA60, were excluded from our dataset as they are caused by regional factors; and (3) the high-resolution δ13Ccarb profiles preserve a clear expression of the negative CIE with steady pre-CIE, and prominent peak values were selected because this feature provides an unambiguous indicator and chronostratigraphic control of periods before and during hyperthermals. Biostratigraphy (conodonts for the PTTM, ammonites for the ETTM, and benthic foraminifer or nannofossils for the PETM) was also used to further ensure the age reliability of the carbon isotope profiles. Within the data compilation, we recognised three depositional settings based on lithology, special deposits (e.g., microbialites and oolites), depth-dependent fossils, and paleodepth calculations (Supplementary Data 1; Supplementary Information). These included shallow shelf (0–80 m), deep shelf (80–200 m), and basin facies (200–500 m), which represent the three distinct water depth levels within the upper ocean. For example, for the Permian–Triassic, shallow shelf deposits are represented by microbialites or oolites, whereas a deep shelf is indicated by marlstone intercalated with mudstone, and a deep basin is indicated by siliceous limestones or rocks with abundant radiolarians (see Supplementary Information). Our dataset comprises a total of 85 δ13Ccarb profiles from the PTTM, 56 profiles from the ETTM, and 33 profiles from the PETM (Supplementary Data 1; Supplementary Table 1). The δ13Ccarb dataset for the Permian–Triassic interval is based on our previous research7 with additional data updates, incorporating a total of 60 shallow shelf profiles, 17 deep shelf profiles, and 8 basin profiles. The δ13Ccarb dataset for the Toarcian encompasses 11 shallow shelf profiles, 31 deep shelf profiles, and 14 basin profiles. Additionally, the δ13Ccarb dataset for the PETM includes 10 shallow shelf profiles, 17 deep shelf profiles, and 6 basin profiles.
Estimates of vertical δ13CDIC gradients
To reconstruct the vertical δ13CDIC gradients before and during hyperthermals, the average δ13CDIC values (mean ± 1 s.d.) were calculated for each of the three palaeo-depths during the pre-CIE (pre-warming) and peak CIE (peak warming) phases. The Shapiro‒Wilk test (function “shapiro.test” in R) was performed to demonstrate that the δ13C values were statistically normally distributed. We then assumed the mean depths of the three depositional settings to be ~40 m, ~130 m, and ~230 m, which correspond to the three uppermost ocean levels in the cGENIE Earth system model36. There are multiple reasons for simplifying the water depth of depositional settings, as follows: (1) Our aim was to comprehend the relative change in the vertical δ13C gradient between pre-CIE and peak CIE phases, rather than reconstructing continuous temporal patterns; (2) photosynthesis and organic matter remineralization occur in the surface ocean (< ~100 m) and mid-water (> 100 m), respectively36, which correspond to our assumed surface depth of 40 m vs. mid-water depths of ~130 and ~230 m in the model; and (3) both proxy and modelled δ13C using the same water depth facilitate their direct comparisons. The gradients calculated by the δ13CDIC differences between the shallow shelf (40 m) and the deep shelf or basin (130 m or 230 m) were then calculated to determine the gradients. These gradients were expressed as δ13CDIC change per hundred metres in water depth (mean ± 1 s.d. ‰/hm). We concentrated specifically on the gradients calculated by the deep shelf because the data from the basin were insufficient for reliable gradient estimation (Supplementary Fig. 6). A two-sample t-test (function “t.test” in R) was performed to demonstrate the statistical significance of the difference in δ13CDIC between surface and mid-water (Supplementary Table 1). Additionally, we calculated the vertical δ13CDIC gradients under other two assumed water depth scenarios in depositional settings for comparison purposes (Supplementary Table 2).
The observed δ13Ccarb values at the deep shelf or basin may reflect a mixing signal of both deep and shallow water DIC. To assess the uncertainty in the mixing proportion, we randomly sampled 1000 different mixing proportions of shallow water DIC (0 ~ 90%). Using a simple isotopic mixing model of two end-members (Supplementary Information), then calculated the actual δ13C values of deep water (δ13CDIC-deep) and the vertical gradient (Supplementary Fig. 10). Furthermore, in order to assess the impact of depth uncertainty on the estimated vertical gradient, we consider two extreme scenarios: (1) the shallow shelf deepening to a deep shelf (“depth increase” scenario) and (2) the deep shelf becoming shallower to a shallow shelf (“depth decrease” scenario). In the “depth increase” scenario, we randomly sample several shallow-shelf sections and reclassify them as deep-shelf locations. In the “depth decrease” scenarios, we randomly sample several deep-shelf locations and reclassify them as shallow-shelf sections. For each scenario, 1000 random sampling iterations were performed (Supplementary Figs. 11 and 12).
Carbon isotopes of planktonic foraminifers
In addition to carbonate δ13C, the δ13C of planktonic foraminiferal shells can indicate oceanic δ13CDIC when considering shell size fractions and physiological disequilibrium effects61. Meanwhile, planktonic foraminiferal δ18O is a powerful proxy for sea temperature that decreases with depth, indicating paleodepth34,62. Therefore, the paired planktonic foraminiferal δ13C and δ18O profiles can be used to reconstruct the δ13CDIC depth profiles. In our dataset, the shells of 24 profiles used a constrained size fraction (212–355 μm) following previous work34, while the remaining 8 samples either lack size information or used a smaller size fraction (180–212 μm). Smaller shells typically show lower δ13C and δ18O values due to vital effects34, which could lead to lower estimated δ13CDIC and shallower inferred water depths. Consequently, the use of smaller shell sizes may slightly exaggerate vertical δ13CDIC gradients, although this bias is likely limited given the constrained size fractions applied to most samples. Deep-ocean benthic foraminifera are excluded from this approach, as previous studies have noted that the vertical δ13C gradient between planktonic and deep benthic foraminifera decreased during the PETM63,64. One possible explanation is that the smaller CIE magnitude observed at pelagic sites may result from incomplete preservation due to carbonate dissolution64, rendering them unsuitable for reconstructing the vertical gradient. However, this method is only possible for the PETM and, therefore, was considered a supplementary method in this study. A total of 32 paired planktonic foraminiferal δ13C and δ18O profiles from nine locations in the PETM were collected (Supplementary Data 1 and references in the Supplementary Information). The estimation of paleodepth was based on planktonic foraminiferal δ18O and modelled salinity and temperature, following a previous study34 (Supplementary Information).
Description of the Earth system model
We used ‘cGENIE’ in this study, an Earth system model of intermediate complexity composed of a 3-D dynamic ocean circulation model with a 2-D energy and moisture balance model of the atmosphere, a dynamic thermodynamic sea-ice model65, and a representation of the biogeochemical cycling of elements and isotopes in the ocean36. Our simulations integrated two types of biological pump schemes. First, a static model with a fixed double-exponential profile of POM flux has been applied in previous simulation works26. Second, an alternative approach is a dynamic model that incorporates temperature-dependent ocean microbial processes44. Specifically, the dynamic model incorporates Arrhenius-type temperature-dependent scheme for the microbial remineralization of organic matter to regulates POC flux in mid-waters44. Nutrient inventories, which are crucial to controlling the biological pump, have been simplified in our simulations by including only phosphate66. The biological pump parameters used in our simulations were based on modelling of middle Miocene cooling34. All the simulations were configured on a 36 × 36 grid with 16 vertical levels in the ocean. The palaeobathymetry, continental configuration, and wind stress of the three hyperthermal events were primarily derived from previous cGENIE simulations (PTTM67, ETTM68, and PETM12).
Description of the numerical experiments
The δ13C values of CO2 forcings were unchanged (−6.5‰) in the model during pre-CIE and peak CIE, as our simulations did not aim to investigate the impact of the massive light carbon release on the ocean δ13CDIC. The changes in ocean δ13CDIC are exclusively caused by various temperature-induced changes to the biological pump, including “circulation change”, “eutrophication”, “microbial metabolism” and “multi-factor” scenarios. A comparison of the static and dynamic biological pump models and the change in ocean phosphate concentration ([PO4]) between the pre-CIE (pre-warming) and peak CIE (peak warming) phases was designed to distinguish the proposed scenarios (Supplementary Table 4). Four scenarios are presented, as follows: (1) “circulation change” (temperature-driven physical ocean circulation change): 1.0× [PO4] during both pre-CIE and peak CIE with the static model; (2) “eutrophication” (nutrition-driven): 1.0× [PO4] at pre-CIE and 2.0× [PO4] at peak CIE with the static model; (3) “microbial metabolism” (warming-driven increase in microbial metabolism): 1.0× [PO4] at pre-CIE and peak CIE with the dynamic model; (4) “multi-factor”: 1.0× [PO4] at pre-CIE and 2.0× [PO4] at peak CIE with the dynamic model. A total of six experiments were designed for each hyperthermal, which included two pre-CIE experiments and four peak CIE experiments.
We prescribed a range of atmospheric pCO2 values (0.5–9× pre-industrial level, that is, 140–2520 ppmv) chosen to match the increase in SST from low or middle latitudes reconstructed from proxy records (Fig. 1). The modelled SST in the tropical Tethys Ocean for the PTTM rises from 27.3 ± 1.2 °C to 37.0 ± 1.1 °C; the modelled SST for the middle-latitude European Shelf for the ETTM increases from 20.8 ± 0.6 °C to 27.8 ± 0.6 °C; the simulated SST in the tropical Atlantic for the PETM increases from 33.1 ± 0.4 °C to 37.3 ± 0.4 °C. Furthermore, it has been observed that modern marine phytoplankton (e.g., coccolithophorids, dinoflagellates, and diatoms) occurred and flourished after the Mesozoic Marine Revolution38,69,70,71. Therefore, as for “microbial metabolism” scenario, we set the default sinking rate of the POM during the ETTM and the PETM after the Mesozoic Marine Revolution to be the same as the modern-day value (125 m yr-1). For the PTTM, a relatively slower sinking rate (98 m y-1) was determined based on changes in marine animal body sizes72, as suggested in a previous cGENIE simulation39.
Collectively, we performed three ensembles of 18 main experiments based on the Permian-Triassic, Toarcian, and late Palaeocene continental configurations. To further explore the contributions of environmental (temperature, phosphate, and continental configuration) and biotic (sinking rate) factors to the ocean carbon cycle, a series of sensitivity tests were conducted. We ran 50 model experiments using an ensemble of ocean temperatures (1, 2, 4, 8, and 16× the pre-industrial level of 280 ppmv), phosphate (1.0, 1.5, 2.0, 2.5, and 3.0× the modern [PO4] level of 2.159 μmol P kg−1) and a dynamic or static biological pump model with the Permian–Triassic configuration (Supplementary Fig. 13). An ensemble comprising five experiments was conducted with varying sinking rates (90, 100, 110, 120, and 130 m yr−1) based on the Permian–Triassic continental configuration using the dynamic model (Supplementary Fig. 15). All the simulations were run for 10 kyr to reach equilibrium.
Model and proxy data comparison
As vertical δ13C gradients vary across different latitudes according to observations from the modern ocean (Supplementary Fig. 2d-f). All averages of modelled δ13CDIC values were calculated based on modelled results gathered from low to middle latitudes (48.6° S to 48.6° N), where most of the sections are located (Supplementary Fig. 1). As sampling model results based on latitudinal regions helps avoid issues arising from spatial unevenness. Before any comparison between the proxy and model, δ13C values need to be adjusted. The average proxy and modelled δ13C values at the surface water (40 m) are designated as the reference standard and adjusted to 0‰, and then other proxy and model data were shifted accordingly by their corresponding reference standard. The proposed centring method eliminated the negative CIE in proxy data during peak CIE phases resulting from carbon emissions not included in the simulations. This also removed any discrepancy between the modelled δ13CDIC values and proxy δ13Ccarb values caused by fractionation between oceanic DIC and carbonate (invariant and independent of temperature) and diagenesis. Root mean square error calculations (function “rmse” within the package “Metrics” in R) were performed to compare the vertical δ13CDIC gradient among the three model scenarios and proxy data and determine the scenario that best fits the proxy data (Supplementary Table 2).
Data availability
All data generated in this study are provided in the Supplementary Information.
Code availability
The code necessary to run the cGENIE model is available at (https://github.com/derpycode/cgenie.muffin), and additional configuration codes to replicate the model results are accessible at (https://doi.org/10.5281/zenodo.10039080).
References
Tierney, J. E. et al. Past climates inform our future. Science 370, eaay3701 (2020).
Judd, E. J. et al. A 485-million-year history of Earth’s surface temperature. Science 385, eadk3705 (2024).
Gómez, J. J., Goy, A. & Canales, M. L. Seawater temperature and carbon isotope variations in belemnites linked to mass extinction during the Toarcian (early Jurassic) in central and northern Spain. Comparison with other European sections. Paleogeogr. Paleoclimatol. Paleoecol. 258, 28–58 (2008).
Scotese, C. R., Song, H. J., Mills, B. J. W. & van der Meer, D. G. Phanerozoic paleotemperatures: The earth’s changing climate during the last 540 million years. Earth-Sci. Rev. 215, 103503 (2021).
Cramwinckel, M. J. et al. Synchronous tropical and polar temperature evolution in the Eocene. Nature 559, 382–386 (2018).
Sun, Y. et al. Lethally hot temperatures during the Early Triassic greenhouse. Science 338, 366–370 (2012).
Wu, Y. Y. et al. Six-fold increase of atmospheric pCO2 during the Permian–Triassic mass extinction. Nat. Commun. 12, 2137 (2021).
Nordt, L., Breecker, D. & White, J. Jurassic greenhouse ice-sheet fluctuations sensitive to atmospheric CO2 dynamics. Nat. Geosci. 15, 54–59 (2022).
Cramer, B. S., Toggweiler, J. R., Wright, J. D., Katz, M. E. & Miller, K. G. Ocean overturning since the Late Cretaceous: Inferences from a new benthic foraminiferal isotope compilation. Paleoceanography 24, PA4216 (2009).
Song, H. J. et al. Thresholds of temperature change for mass extinctions. Nat. Commun. 12, 4694 (2021).
Wu, Y. Y. et al. Volcanic CO2 degassing postdates thermogenic carbon emission during the end-Permian mass extinction. Sci. Adv. 9, eabq4082 (2023).
Gutjahr, M. et al. Very large release of mostly volcanic carbon during the Palaeocene–Eocene Thermal Maximum. Nature 548, 573–577 (2017).
Dickens, G. R., O’Neil, J. R., Rea, D. K. & Owen, R. M. Dissociation of oceanic methane hydrate as a cause of the carbon isotope excursion at the end of the Paleocene. Paleoceanography 10, 965–971 (1995).
Berner, R. A. Examination of hypotheses for the Permo-Triassic boundary extinction by carbon cycle modeling. Proc. Natl Acad. Sci. Usa. 99, 4172–4177 (2002).
Ducklow, H. W., Steinberg, D. K. & Buesseler, K. O. Upper Ocean Carbon Export and the Biological Pump. Oceanography 14, 50 (2001).
Kwon, E. Y., Primeau, F. & Sarmiento, J. L. The impact of remineralization depth on the air–sea carbon balance. Nat. Geosci. 2, 630–635 (2009).
Ricour, F., Guidi, L., Gehlen, M., DeVries, T. & Legendre, L. Century-scale carbon sequestration flux throughout the ocean by the biological pump. Nat. Geosci. 16, 1105–1113 (2023).
Siegel, D. A., DeVries, T., Cetinić, I. & Bisson, K. M. Quantifying the ocean’s biological pump and its carbon cycle impacts on global scales. Annu. Rev. Mar. Sci. 15, 329–356 (2023).
DeVries, T., Primeau, F. & Deutsch, C. The sequestration efficiency of the biological pump. Geophys. Res. Lett. 39, L13601 (2012).
Henson, S. A., Sanders, R. & Madsen, E. Global patterns in efficiency of particulate organic carbon export and transfer to the deep ocean. Glob. Biogeochem. Cycle 26, GB1028 (2012).
Wilson, J. D. et al. The biological carbon pump in CMIP6 models: 21st century trends and uncertainties. Proc. Natl Acad. Sci. Usa. 119, e2204369119 (2022).
Crichton, K. A. et al. What the geological past can tell us about the future of the ocean’s twilight zone. Nat. Commun. 14, 2376 (2023).
Penn, J. L., Deutsch, C., Payne, J. L. & Sperling, E. A. Temperature-dependent hypoxia explains biogeography and severity of end-Permian marine mass extinction. Science 362, eaat1327 (2018).
Winguth, A. M. E., Thomas, E. & Winguth, C. Global decline in ocean ventilation, oxygenation, and productivity during the Paleocene-Eocene thermal maximum: implications for the benthic extinction. Geology 40, 263–266 (2012).
Ma, Z. et al. Carbon sequestration during the Palaeocene–Eocene thermal maximum by an efficient biological pump. Nat. Geosci. 7, 382–388 (2014).
Meyer, K. M., Kump, L. R. & Ridgwell, A. Biogeochemical controls on photic-zone euxinia during the end-Permian mass extinction. Geology 36, 747–750 (2008).
Schobben, M. et al. A nutrient control on marine anoxia during the end-Permian mass extinction. Nat. Geosci. 13, 640–646 (2020).
Gillooly, J. F., Brown, J. H., West, G. B., Savage, V. M. & Charnov, E. L. Effects of size and temperature on metabolic rate. Science 293, 2248–2251 (2001).
Boscolo-Galazzo, F., Crichton, K. A., Barker, S. & Pearson, P. N. Temperature dependency of metabolic rates in the upper ocean: A positive feedback to global climate change? Glob. Planet. Change 170, 201–212 (2018).
Cram, J. A. et al. The role of particle size, ballast, temperature, and oxygen in the sinking flux to the deep sea. Glob. Biogeochem. Cycle 32, 858–876 (2018).
DeVries, T. & Weber, T. The export and fate of organic matter in the ocean: New constraints from combining satellite and oceanographic tracer observations. Glob. Biogeochem. Cycle 31, 535–555 (2017).
Kroopnick, P. M. The distribution of 13C of ΣCO2 in the world oceans. Deep-Sea Res. 32, 57–84 (1985).
Morée, A. L. et al. Evaluating the biological pump efficiency of the Last Glacial Maximum ocean using δ13C. Clim. 17, 753–774 (2021).
Boscolo-Galazzo, F. et al. Temperature controls carbon cycling and biological evolution in the ocean twilight zone. Science 371, 1148–1152 (2021).
Hilting, A. K., Kump, L. R. & Bralower, T. J. Variations in the oceanic vertical carbon isotope gradient and their implications for the Paleocene-Eocene biological pump. Paleoceanography 23, PA3222 (2008).
Ridgwell, A. et al. Marine geochemical data assimilation in an efficient Earth system model of global biogeochemical cycling. Biogeosciences 4, 87–104 (2007).
Ahm, A.-S. C., Bjerrum, C. J., Blättler, C. L., Swart, P. K. & Higgins, J. A. Quantifying early marine diagenesis in shallow-water carbonate sediments. Geochim. Cosmochim. Acta. 236, 140–159 (2018).
Hull, P. M. Emergence of modern marine ecosystems. Curr. Biol. 27, R466–R469 (2017).
Hülse, D. et al. End-Permian marine extinction due to temperature-driven nutrient recycling and euxinia. Nat. Geosci. 14, 862–867 (2021).
Song, H. Y. et al. Large vertical δ13CDIC gradients in Early Triassic seas of the South China craton: implications for oceanographic changes related to Siberian Traps volcanism. Glob. Planet. Change 105, 7–20 (2013).
Meyer, K. M., Yu, M., Jost, A. B., Kelley, B. M. & Payne, J. L. δ13C evidence that high primary productivity delayed recovery from end-Permian mass extinction. Earth Planet. Sci. Lett. 302, 378–384 (2011).
John, E. H., Wilson, J. D., Pearson, P. N. & Ridgwell, A. Temperature-dependent remineralization and carbon cycling in the warm Eocene oceans. Paleogeogr. Paleoclimatol. Paleoecol. 413, 158–166 (2014).
Shen, B. et al. Carbon, sulfur, and oxygen isotope evidence for a strong depth gradient and oceanic oxidation after the Ediacaran Hankalchough glaciation. Geochim. Cosmochim. Acta. 75, 1357–1373 (2011).
Crichton, K. A., Wilson, J. D., Ridgwell, A. & Pearson, P. N. Calibration of temperature-dependent ocean microbial processes in the cGENIE.muffin (v0.9.13) Earth system model. Geosci. Model Dev. 14, 125–149 (2021).
Morée, A. L., Schwinger, J. & Heinze, C. Southern Ocean controls of the vertical marine δ13C gradient - a modelling study. Biogeosciences 15, 7205–7223 (2018).
Bridgestock, L., Hsieh, Y. T., Porcelli, D. & Henderson, G. M. Increased export production during recovery from the Paleocene-Eocene thermal maximum constrained by sedimentary Ba isotopes. Earth Planet. Sci. Lett. 510, 53–63 (2019).
Sarmiento, J. L. & Toggweiler, J. R. A new model for the role of the oceans in determining atmospheric Pco2. Nature 308, 621–624 (1984).
Gorgues, T., Aumont, O. & Memery, L. Simulated changes in the particulate carbon export efficiency due to diel vertical migration of zooplankton in the North Atlantic. Geophys. Res. Lett. 46, 5387–5395 (2019).
Ward, B. A., Dutkiewicz, S., Jahn, O. & Follows, M. J. A size-structured food-web model for the global ocean. Limnol. Oceanogr. 57, 1877–1891 (2012).
Wu, Y. Y., Song, H. J., Chu, D. L., Song, H. Y. & Tian, L. Environmental impacts and biotic responses to volcanism during the Permian⁃Triassic transition. Earth Sci. 50, 964–982 (2025).
Isson, T. T. et al. Marine siliceous ecosystem decline led to sustained anomalous Early Triassic warmth. Nat. Commun. 13, 3509 (2022).
Kemp, D. B., Suan, G., Fantasia, A., Jin, S. & Chen, W. Global organic carbon burial during the Toarcian oceanic anoxic event: Patterns and controls. Earth-Sci. Rev. 231, 104086 (2022).
Algeo, T. J. et al. Plankton and productivity during the Permian–Triassic boundary crisis: An analysis of organic carbon fluxes. Glob. Planet. Change 105, 52–67 (2013).
Li, Z., Zhang, Y. G., Torres, M. & Mills, B. J. W. Neogene burial of organic carbon in the global ocean. Nature 613, 90–95 (2023).
Papadomanolaki, N. M., Sluijs, A. & Slomp, C. P. Eutrophication and deoxygenation forcing of marginal marine organic carbon burial during the PETM. Paleoceanogr. Paleoclimatology 37, e2021PA004232 (2022).
Hülse, D., Arndt, S., Wilson, J. D., Munhoven, G. & Ridgwell, A. Understanding the causes and consequences of past marine carbon cycling variability through models. Earth-Sci. Rev. 171, 349–382 (2017).
Pika, P. H., Ulse, D. & Arndt, S. OMEN-SED(-RCM) (v1.1): a pseudo-reactive continuum representation of organic matter degradation dynamics for OMEN-SED. Geosci. Model Dev. 14, 7155–7174 (2021).
Laufkötter, C., John, J. G., Stock, C. A. & Dunne, J. P. Temperature and oxygen dependence of the remineralization of organic matter. Glob. Biogeochem. Cycle 31, 1038–1050 (2017).
Wilson, J. D., Barker, S. & Ridgwell, A. Assessment of the spatial variability in particulate organic matter and mineral sinking fluxes in the ocean interior: Implications for the ballast hypothesis. Glob. Biogeochem. Cycle 26, GB4011 (2012).
Self-Trail, J. M. et al. Shallow marine response to global climate change during the Paleocene-Eocene Thermal Maximum, Salisbury Embayment, USA. Paleoceanography 32, 710–728 (2017).
Birch, H., Coxall, H. K., Pearson, P. N., Kroon, D. & O’Regan, M. Planktonic foraminifera stable isotopes and water column structure: Disentangling ecological signals. Mar. Micropaleontol. 101, 127–145 (2013).
John, E. H. et al. Warm ocean processes and carbon cycling in the Eocene. Philos. Trans. R. Soc. A 371, 20130099 (2013).
McInerney, F. A. & Wing, S. L. The Paleocene-Eocene thermal maximum: a perturbation of carbon cycle, climate, and biosphere with implications for the future. Annu. Rev. Earth Pl. Sc. 39, 489–516 (2011).
Zachos, J. C. et al. The Palaeocene-Eocene carbon isotope excursion: constraints from individual shell planktonic foraminifer records. Philos. Trans. R. Soc. A 365, 1829–1842 (2007).
Edwards, N. & Marsh, R. Uncertainties due to transport-parameter sensitivity in an efficient 3-D ocean-climate model. Clim. Dyn. 24, 415–433 (2005).
Meyer, K. M., Ridgwell, A. & Payne, J. L. The influence of the biological pump on ocean chemistry: implications for long-term trends in marine redox chemistry, the global carbon cycle, and marine animal ecosystems. Geobiology 14, 207–219 (2016).
Cui, Y., Kump, L. R. & Ridgwell, A. Initial assessment of the carbon emission rate and climatic consequences during the end-Permian mass extinction. Paleogeogr. Paleoclimatol. Paleoecol. 389, 128–136 (2013).
Pohl, A. et al. Continental configuration controls ocean oxygenation during the Phanerozoic. Nature 608, 523–527 (2022).
Falkowski, P. G. et al. The evolution of modern eukaryotic phytoplankton. Science 305, 354 (2004).
Knoll, A. H. & Follows, M. J. A bottom-up perspective on ecosystem change in Mesozoic oceans. Proc. R. Soc. B-Biol. Sci. 283, 20161755 (2016).
Lu, W. et al. Late inception of a resiliently oxygenated upper ocean. Science 361, 174–177 (2018).
Heim, N. A., Knope, M. L., Schaal, E. K., Wang, S. C. & Payne, J. L. Cope’s rule in the evolution of marine animals. Science 347, 867–870 (2015).
Acknowledgements
We thank Andy Ridgwell for providing useful comments. This research was supported by the National Natural Science Foundation of China (42325202, 42293292, 92155201, 92255303, 42030513, 42202002), State Key R&D Project of China (2023YFF0804000), Natural Science Foundation of Hubei (2023AFA006), the Fundamental Research Funds for the Central Universities, China University of Geosciences (Wuhan).
Author information
Authors and Affiliations
Contributions
H.J.S. and Y.W. designed research. Y.W. and H.J.S. performed research. Y.W., H.J.S., D.C. and Y.C. analyzed data. Y.W., H.J.S., D.C., Y.C., J.D.C., G.L., H.Y.S., L.T., H.C.S., E.H.J. and J.T. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Christian Bjerrum, Jamie Wilson, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wu, Y., Song, H., Chu, D. et al. Microbial metabolism amplified warming in three Phanerozoic hyperthermal events. Nat Commun 16, 11372 (2025). https://doi.org/10.1038/s41467-025-66388-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-66388-2
