
Abstract
The brainstem houses numerous nuclei and tracts that serve vital functions. Genome-wide associations with brainstem substructure volumes have been explored in European individuals, yet other ancestries remain under-represented. Here, we conduct cross-ancestry genome-wide association meta-analyses in 103,098 individuals for brainstem and 78,062 individuals for eight substructure volumes, including 7094 Chinese Han individuals. We identify 713 locus-trait associations with brainstem and substructure volumes at P < 5.56 ×10−9, comprising 569 new associations. Two associations show different effect sizes, while 496 associations have similar effect sizes between ancestries. We prioritize 186 genes associated with brainstem volumetric traits. We find both shared and distinct genetic loci, genes, and pathways for midbrain, pons, and medulla volumes, along with the shared genetic architectures related to disease phenotypes and physiological functions. The results provide new insights into the genetic architectures of brainstem and substructure volumes and their genetic associations with brainstem physiologies and pathologies.
Similar content being viewed by others


Introduction
The brainstem anatomically connects the cerebrum to cerebellum and spinal cord and consists of the medulla, pons, and midbrain. It contains numerous grey matter nuclei and white matter tracts that are essential for our survival. For instance, the vital center in the medulla is responsible for autonomic control of breathing, blood pressure, and heart rate1,2,3. The locus coeruleus in the pons is the primary source of noradrenaline, playing key roles in vigilance and cognition; dysfunction of this nucleus may lead to arousal-related disorders, such as attention deficit hyperactivity disorder (ADHD) and depression4. The substantia nigra in the midbrain contains many dopaminergic neurons, generating dopamine (DA) crucial for controlling voluntary movement and regulating emotion. Damage to this structure is associated with schizophrenia, Parkinson’s disease (PD), depression, and ADHD5,6. In addition to the grey matter nuclei, white matter fibers in the brainstem are also vital for various physiological functions. For example, the reticular formation is essential for maintaining consciousness7 and the corticospinal tract is critical for controlling limb movement.
Based on individual’s brain structural magnetic resonance imaging (MRI) data, the volumes of the brainstem and its substructures (medulla, pons, and midbrain) could be estimated using various methods such as Bayesian brainstem segmentation8. Significant differences in the volumes of brainstem substructures have been observed between healthy controls and patients with neuropsychiatric disorders8,9,10,11, such as Alzheimer’s disease (AD), amyotrophic lateral sclerosis (ALS), migraine, and autism spectrum disorder (ASD). This suggests that the brainstem and substructure volumes may serve as potential neuroimaging markers for these neuropsychiatric disorders. To explore the genetic architectures of these brainstem volumetric traits, six genome-wide association studies (GWAS) were conducted on whole brainstem volume involving up to 61,859 individuals12,13,14,15,16,17. Two of these studies also performed GWAS for brainstem substructure volumes in up to 33,224 individuals12,13. Although one study involving the GWAS for whole brainstem volume included 7058 individuals with East Asian ancestry (EAS)17, other studies focused only on individuals with European ancestry (EUR)12,13,14,15,16, hindering the ability to distinguish between ancestry-specific and ancestry-shared associations for brainstem substructure volumes.
In this work, we conduct the largest GWAS for whole brainstem volume in 103,098 non-overlapping individuals. We also perform the largest GWAS for brainstem substructure volumes in 78,062 individuals. We identify ancestry-shared and ancestry-specific genetic associations with brainstem volumetric traits based on the EAS-GWAS and EUR-GWAS summary statistics. We detect potential causal variants via statistical fine-mapping and prioritize relevant genes. We examine shared and distinct genetic loci, genes, and biological pathways related to brainstem substructure volumes, and explore overlaps with other phenotypes by conducting phenome-wide association study (PheWAS), genetic correlation, genetic colocalization, and conjunctional false discovery rate (conjFDR) analyses. The study design is presented in Supplementary Fig. 1. By increasing sample size and population diversity, we uncover more associations, enhancing our understanding of the genetic architecture of brainstem volumetric traits.
Results
Study overview
We included three sets of raw neuroimaging genetics datasets (CHIMGEN, ABCD, and UKB) and a set of GWAS summary data for whole brainstem volume from ENIGMA, CHARGE, and UKB (first release) datasets14. Consistent with the prior GWAS12,13 for brainstem substructure volumes, we applied the same Bayesian approach8 to obtain the absolute volumes of the whole brainstem, medulla, pons, midbrain, and superior cerebellar peduncle (SCP) (Fig. 1a) for each participant from CHIMGEN, ABCD, and UKB, although SCP is not a classical brainstem substructure. The total brainstem volume (including SCP) was included as an additional covariate to perform GWAS for the relative volumes of the medulla, pons, midbrain, and SCP12. As the prior GWAS14 applied the automatic subcortical segmentation method to obtain the whole brainstem (excluding SCP) volume, we also employed the same approach to calculate the whole brainstem volume for each participant from CHIMGEN, ABCD, or UKB, which was used in GWAS for the trait. The demographic and volumetric data are presented in Supplementary Table 1. The participants and genetic variants included in each GWAS are detailed in Supplementary Table 2. The genomic control inflation factor (λGC) and linkage disequilibrium score regression (LDSC) intercepts18 were utilized to assess population stratification for each GWAS, and no population stratification was observed (Supplementary Table 3).

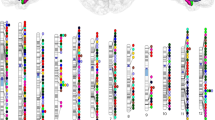
a Diagrams illustrate the brainstem and its four substructures (medulla, pons, midbrain, and SCP). We conduct GWAS for nine brainstem volumetric traits, including the whole brainstem volume as well as the absolute and relative volumes of each substructure. b Pie plots display the counts and ratios of replicated associations (two-sided P < 0.05) among the discovered significant associations (two-sided P < 5.56 × 10−9, Bonferroni corrected) for the nine brainstem volumetric traits identified by single-ancestry (linear regression model) or cross-ancestry (fixed-effect model) GWAS. c The ideogram demonstrates study-wide significant (two-sided P < 5.56 × 10−9, Bonferroni corrected) locus-trait associations in the total sample (fixed-effect model). d The regional plot shows an example of locus-trait associations between 2p11.2 and pons absolute volume in EAS-GWAS. The red and blue lines indicate genome-wide (two-sided P = 5 × 10−8) and study-wide (two-sided P = 5.56 × 10−9, Bonferroni corrected) significance thresholds (linear regression model). e The ideogram shows known (rhombus) and new (circle) locus-trait associations (two-sided P < 5.56 × 10−9, Bonferroni corrected) compared to prior GWAS identified by single-ancestry (linear regression model) or cross-ancestry (fixed-effect model) GWAS. SCP superior cerebellar peduncle. Source data are provided as a Source Data file.
Genetic architecture of brainstem volumetric traits
In 103,098 participants from three raw datasets and a set of GWAS summary data, we conducted two-stage GWAS for nine brainstem volumetric traits at both autosomal and X-chromosomal variants with information score (info) > 0.9, minor allele frequency (MAF) ≥ 0.5%, and P ≥ 1 × 10−7 in Hardy–Weinberg equilibrium. In the discovery stage, we found 250, 251, and 352 independent variant-trait associations (P < 5.56 × 10−9, Bonferroni corrected) in EUR-GWAS, EUR-EAS-GWAS and admixed-GWAS meta-analyses, respectively (Supplementary Fig. 2). Out of all 853 associations, 748 (87.7%) were replicated at nominal P < 0.05 (Fig. 1b), 738 (86.5%) at Benjamini–Hochberg false positive rate (BH-FDR)-corrected Pc < 0.05, and 423 (49.6%) at Bonferroni-corrected Pc < 0.05 in the replication stage (Supplementary Data 1).
We also conducted EAS-GWAS, EUR-GWAS, EUR-EAS-GWAS, and admixed-GWAS for each brainstem volumetric trait in the total sample to identify additional variant-trait and locus-trait associations. Using a study-wide significance threshold (P < 5.56 × 10−9, Bonferroni corrected), we identified 128, 129, and 153 loci associated with whole brainstem volume in EUR-GWAS, EUR-EAS-GWAS, and admixed-GWAS. We also found 63/66/82 loci for medulla absolute volume, 38/36/46 for medulla relative volume, 94/97/123 for pons absolute volume, 61/58/70 for pons relative volume, 65/66/89 for midbrain absolute volume, 48/47/69 for midbrain relative volume, 18/18/28 for SCP absolute volume, and 13/14/16 for SCP relative volume in the three categories of GWAS (Fig. 1c, Supplementary Fig. 3 and Supplementary Data 2). In EAS-GWAS, we identified a single locus-trait association (P = 2.61 × 10−9; Fig. 1d and Supplementary Data 2) between rs6547787 (2p11.2) and pons absolute volume. This variant is an expression quantitative trait locus (eQTL) for EIF2AK3, a risk gene linked to progressive supranuclear palsy characterized by brainstem pathology19.
As most prior GWAS for brainstem and substructure volumes were conducted in EUR individuals, and since EUR samples were at least seven times larger than EAS samples in this study, we used the EUR-LD reference to pool all significant locus-trait associations (P < 5.56 × 10−9, Bonferroni corrected). This allowed us to integrate genetic discovery within and across GWAS. We identified 713 locus-trait associations, which included 161 loci for whole brainstem volume, 90 for medulla absolute volume, 50 for medulla relative volume, 128 for pons absolute volume, 74 for pons relative volume, 92 for midbrain absolute volume, 71 for midbrain relative volume, 29 for SCP absolute volume, and 18 for SCP relative volume (Fig. 1e and Supplementary Data 3). To identify new associations, we integrated all previous GWAS for brainstem and substructure volumes12,13,14,15,16,17 to define the reference (P < 5.56 × 10−9) of known locus-trait associations (n = 174). We identified 569 new locus-trait associations, including 105 new loci for whole brainstem volume, 72 for medulla absolute volume, 50 for medulla relative volume, 89 for pons absolute volume, 66 for pons relative volume, 78 for midbrain absolute volume, 67 for midbrain relative volume, 24 for SCP absolute volume, 18 for SCP relative volume (Fig. 1e and Supplementary Data 3). For example, the new locus at 3p14.1 was associated with SCP absolute volume (a white matter tract) identified in EUR-GWAS (P = 3.80 × 10−9), EUR-EAS-GWAS (P = 3.37 × 10−9), admixed-GWAS (P = 1.75 × 10−10). Its lead variant, rs2306272, is a missense variant of LRIG1, which controls axonal extension, guidance, and branching by regulating growth factor signals during neural development20. Additionally, we found an association between rs7029693 at 9q33.1 and medulla relative volume in EUR-GWAS (P = 1.15 × 10−20), EUR-EAS-GWAS (P = 3.47 × 10−22), and admixed-GWAS (P = 4.38 × 10−26). The lead variant is mapped to ASTN2, which is involved in neuronal migration and linked to migraine without aura21, characterized by tractus solitarius impairment in the medulla22.
We conducted sensitivity analyses to assess the influence of adjusting for total intracranial volume (TIV) and total brainstem volume on the GWAS for brainstem volumetric traits, as recommended in a previous study23. We calculated correlations among TIV, whole brainstem volume, and brainstem substructure volumes, revealing moderate to high correlations (ρ = 0.330–0.982; Supplementary Fig. 4). We repeated the analyses for 1009 pooled variant-trait associations (Supplementary Data 3), excluding TIV and total brainstem volume as covariates. We found that 689 (68.3%) associations were no longer significant in the sensitivity analyses. However, we found a strong correlation (ρ = 0.88; Supplementary Fig. 5) of effect sizes for the 1009 associations derived from both the main and sensitivity analyses. For example, we identified a significant association between rs59371099 (22q12.2) and pons relative volume (P = 5.15 × 10−12) in the main analysis, while it was not significant in the sensitivity analysis (P = 2.43 × 10−1) (Supplementary Data 3). This variant is a missense variant of NEFH, which is highly expressed in pons24 and identified as a biomarker of neural damage25.
After excluding the major histocompatibility complex (MHC) genomic region, we employed LDSC18,26 to estimate the single-nucleotide polymorphism (SNP)-based heritability of these nine brainstem volumetric traits in EAS, EUR, and cross-ancestry populations, utilizing the EAS, EUR, and cross-ancestry GWAS summary data and LD references, respectively. All traits exhibited significant SNP-based heritability (h2 = 0.18–0.39, SE = 0.01–0.03, all P < 1.42 × 10−43; Supplementary Table 4) in the EUR and cross-ancestry individuals. However, the SNP-based heritability (h2 = 0.15–0.40, SE = 0.09–0.12) of two traits was not significant (P > 0.05) in EAS individuals.
After excluding the genetic variants in the MHC region, we used Popcorn27 to calculate the genetic-effect correlation (without considering MAF information) for each brainstem volumetric trait between EAS and EUR, identifying moderate to high correlations (rg = 0.53–0.90, SE = 0.11–0.16, all P < 1.18 × 10−3; Supplementary Table 5). We also calculated genetic-impact correlations (considering MAF information) for these brainstem volumetric traits between EAS and EUR, and similarly found moderate to high correlations (rg = 0.50–0.85, SE = 0.11–0.15, all P < 1.29 × 10−3; Supplementary Table 5).
To identify ancestry-shared and ancestry-specific variant-trait associations for nine brainstem volumetric traits, we used the Cochran’s Q test (CQ-test) to quantify effect size differences between EAS and EUR for 606 pooled variant-trait associations (P < 5.56 × 10−9) from EAS-GWAS, EUR-GWAS, and cross-ancestry GWAS. Given the disparity in sample size between EAS-GWAS and EUR-GWAS, we performed the CQ-test in the discovery and replication stages using EUR-GWAS data from the respective stages. However, we included the EAS-GWAS data in the total sample throughout both stages. We defined associations with P ≥ 0.05 as ancestry-shared associations and those with P < 8.25 × 10−5 (Bonferroni corrected) as ancestry-specific associations in the discovery stage, during which we found two ancestry-specific and 525 ancestry-shared associations (Fig. 2a, b and Supplementary Data 4). In the replication stage, we validated the two ancestry-specific (CQ-test: P = 0.0023 and P = 1.42 × 10−6) and 496 ancestry-shared associations (CQ-test: P ≥ 0.05) (Fig. 2b and Supplementary Data 4). The two ancestry-specific associations were found between rs111883632 and pons absolute volume, and rs151057105 and midbrain absolute volume (Fig. 2c). We found 119 ancestry-shared associations for whole brainstem volume, 56 for medulla absolute volume, 34 for medulla relative volume, 98 for pons absolute volume, 62 for pons relative volume, 60 for midbrain absolute volume, 41 for midbrain relative volume, 15 for SCP absolute volume, 11 for SCP relative volume (Fig. 2d). We identified 107 new ancestry-shared associations for whole brainstem volume, compared to a prior study17. As an example of new ancestry-shared associations, the association between rs2206656 and whole brainstem volume was observed in both EUR and EAS (CQ-test: P = 0.84 for discovery and P = 0.77 for replication). This variant is associated with PTPN1, a mechanism-based therapeutic target for Rett syndrome, a neurological developmental disease related to the brainstem28.

a The bar chart shows the counts of ancestry-specific (blue; Cochran’s Q test: two-sided P < 8.25 × 10−5, Bonferroni correction for 606 variant-trait associations included in both EAS-GWAS and EUR-GWAS) and ancestry-shared (red; Cochran’s Q test: P ≥ 0.05) associations between EUR-GWAS in the discovery stage and EAS-GWAS. b Pie plots illustrate the counts and ratios of the discovered (left: EAS-GWAS vs EUR-GWAS in the discovery stage) and replicated (right: EAS-GWAS vs EUR-GWAS in the replication stage) ancestry-specific and ancestry-shared associations. c The z-score differences in two ancestry-specific associations between EAS-GWAS and EUR-GWAS in discovery (top: EAS-GWAS vs EUR-GWAS in the discovery stage) and replication (bottom: EAS-GWAS vs EUR-GWAS in the replication stage) stages. d The lollipop plot displays the counts of ancestry-shared associations for nine brainstem volumetric traits. The inserted Venn diagram shows the relationships of ancestry-shared associations between this study and previous research. Source data are provided as a Source Data file.
We conducted GWAS for the volumes of 58 brainstem nuclei in 47,887 admixed-ancestry participants from three datasets, from which we identified 591 independent locus-trait associations (P < 8.62 × 10−10; Supplementary Fig. 6, Supplementary Data 5). For example, we identified significant associations between rs11539157 (Xq13.1) and the volumes of 15 brainstem nuclei (all P < 8.58 × 10−10). This variant is a missense mutation of PJA1, a gene specifically expressed in the cytoplasm of scattered cells within developing brainstem nuclei29.
Statistical fine-mapping and functional annotation
For each locus of the pooled locus-trait associations, we employed the matched LD reference to perform statistical fine-mapping by estimating the posterior probability (PP) of each variant being a causal variant (PP > 0.8) using the probabilistic annotation integrator (PAINTOR) tool30,31 with the Markov chain Monte Carlo (MCMC) model that accommodates multiple causal variants. In the 713 pooled locus-trait associations, one was significant across all three categories (EAS, EUR, and cross-ancestry) of GWAS, 49 were significant only in EUR-GWAS, 48 only in cross-ancestry GWAS, 476 in both EUR-GWAS and cross-ancestry GWAS (Supplementary Data 6). We conducted fine-mapping for one locus based on EAS-GWAS using the EAS-LD reference, 526 loci based on EUR-GWAS using the EUR-LD reference, and 525 loci based on cross-ancestry GWAS using the merged LD reference, respectively. We found 699 unique causal variants for brainstem volumetric traits, of which 223 were identified only in EUR analyses, 455 only in cross-ancestry analyses, and 21 in both EUR and cross-ancestry analyses (Fig. 3a and Supplementary Data 6). For the 476 locus-trait associations demonstrating significant in both EUR and cross-ancestry GWAS, we further conducted PAINTOR under the assumption of one causal variant and reported the findings in Supplementary Data 7. We then used the Wilcoxon rank-sum test (P < 0.05) to investigate whether cross-ancestry fine-mapping can reduce the 95% credible sets for these trait-associated loci compared to EUR-specific fine-mapping. We discovered that the 95% credible sets (median = 4) from cross-ancestry analyses were significantly smaller (P = 7.74 × 10−21; Fig. 3b) than those (median = 10) from EUR analyses. For example, while the locus-trait association between 16q22.1 and midbrain absolute volume was significant in both EUR-GWAS and EUR-EAS-GWAS, we were unable to identify any causal variants based on the EUR-GWAS, but identified one causal variant (rs1837016: PP = 1.00; Supplementary Data 6) through the cross-ancestry GWAS (Fig. 3c). This variant is an eQTL variant of NQO1, which is expressed in both astroglial and neuronal cells. Its expression increases in the substantia nigra pars compacta of patients with PD32.

a The ideogram shows the causal variants (PP > 0.8) identified through statistical fine-mapping of 574 pooled locus-trait associations related to brainstem volumetric traits. We identify 699 unique causal variants, comprising 223 identified solely from EUR analyses, 455 exclusively from cross-ancestry analyses, and 21 from both analyses. b Comparison of the sizes of 95% credible sets for the 476 trait-associated loci identified through statistical fine-mapping for cross-ancestry GWAS (orange) and EUR-GWAS (blue). Two-sided P value is calculated using the Wilcoxon rank-sum test. The sizes of the 95% credible sets are log-transformed and reported as medians and interquartile ranges. c The regional plots show a locus-trait association between 16q22.1 and midbrain absolute volume, demonstrating improved resolution in cross-ancestry fine-mapping. d Categorizing the causal variants (PP > 0.1) from fine-mapping for EUR-GWAS (top) and cross-ancestry GWAS (bottom) based on genomic location and functional consequence. Source data are provided as a Source Data file.
To investigate the functional consequences of variants associated with brainstem volumetric traits, we used FUMA33 to perform functional annotations for the unique variants with PP > 0.1 in fine-mapping. Based on genomic location and functional consequence, we categorized the four variants with PP > 0.1 in the only locus-trait association identified by EAS-GWAS. We found one variant in the intronic region, two variants in the intergenic region, and one variant as a missense variant of EIF2AK3 (Supplementary Data 8). We also categorized 7507 unique variants with PP > 0.1 in the 526 locus-trait associations identified by EUR-GWAS and found that they were mainly in the intronic (47.8%) and intergenic (33.1%) regions (Supplementary Data 8). Additionally, we found 67 variants (0.9%) in the coding sequence and 153 variants (2.0%) in the untranslated region (UTR), which included one stop-loss variant, three stop-gained variants, and 30 missense variants. For instance, rs2231142 (4q22.1), associated with brainstem (P = 4.37 × 10−14, PP = 0.16) and pons absolute (P = 8.70 × 10−11, PP = 0.18) volumes, is a stop-gained variant of ABCG2, a regulator of neural stem cell self-renewal34. As for the 4993 unique genetic variants (PP > 0.1) in 525 locus-trait associations identified by cross-ancestry GWAS, they were also primarily found in the intronic (44.8%) and intergenic (35.6%) regions (Supplementary Data 8). We identified 59 (1.2%) coding and 91 (1.8%) UTR variants, including two stop-gained and 35 missense variants. We found that cross-ancestry fine-mapping could identify a greater proportion of meaningful variants with PP > 0.1 (Fig. 3d). For the 11,621 unique variants with PP > 0.1, we also used combined annotation-dependent depletion (CADD) score to assess the deleteriousness of 11,586 variants included in the database, from which we identified 594 pathogenic variants (CADD score > 12.37; Supplementary Data 8). Furthermore, we used RegulomeDB (RDB) score to identify variants with regulatory function and found 217 regulatory variants (RDB ≤ 1 f; Supplementary Data 8) among the 10,266 fine-mapped variants included in the RDB database.
Prioritized genes and pathways
We used the fine-mapped locus assessment model of effector genes (FLAMES)35 to prioritize genes based on the fine-mapping results from EUR-GWAS summary statistics (discovery, replication, and combined stages) for nine brainstem volumetric traits. In the discovery stage, we found 127 gene-trait associations (69 genes) with a cumulative precision exceeding 75%. Of these, 45 were present in the replication dataset. Among the 45 associations, 28 were validated with 75% cumulative precision in the replication stage (Supplementary Data 9). Using the EUR-GWAS data from the total sample, we prioritized 186 unique genes (Fig. 4a), including 79 genes for whole brainstem volume, 45 for medulla absolute volume, 28 for medulla relative volume, 63 for pons absolute volume, 42 for pons relative volume, 46 for midbrain absolute volume, 35 for midbrain relative volume, 15 for SCP absolute volume, and 11 for SCP relative volume (Fig. 4b and Supplementary Data 10).

a The ideogram shows the locations of prioritized genes for nine brainstem volumetric traits. b The bar chart (top) shows the counts of prioritized genes for these nine traits. The lollipop plot (bottom) displays the counts of prioritized genes for the brainstem and its substructures (regardless of absolute or relative volume). c The enrichment map illustrates the hierarchical relationships of biological pathways enriched by 186 prioritized genes associated with brainstem volumetric traits. Each circle represents an enriched pathway, while the thickness of the lines indicates the number of genes shared by the two pathways. Source data are provided as a Source Data file.
Based on biological pathways from Gene Ontology (GO)36 and Reactome37, we performed pathway enrichment analyses for the prioritized 186 genes associated with brainstem volumetric traits using g:Profiler38 (https://biit.cs.ut.ee/gprofiler/gost). We identified 418 enrichment pathways (Pc < 0.05, g:SCS corrected for set counts and sizes; Supplementary Data 11), including RNA transcription processes, intracellular signal transduction, and neuron projection. (Fig. 4c). Additionally, we found one significant pathway in the Reactome dataset, Netrin-1 signaling (P = 8.31 × 10−3; Fig. 4c), which supports the notion that Netrin-1 influences the regulation of migration of rhombic lip-derived neurons during brainstem development39.
Shared and distinct genetic architectures between brainstem substructures
Although sharing some white matter tracts, the three brainstem substructures (medulla, pons, and midbrain) originate from different embryonic structures (medulla and pons from the hindbrain and midbrain from the mesencephalon) and contain distinct gray matter nuclei and white matter tracts, indicating the coexistence of shared and distinct genetic architectures. Therefore, we investigated the shared and distinct genetic loci, genes, and enriched pathways between each pair of the three brainstem substructures.
We pooled the 505 locus-trait associations with brainstem substructures (medulla, pons, and midbrain) into 244 LD-independent loci. Among these 244 loci, we searched for those associated with the volumes of one, two, or three substructures, regardless of absolute or relative volume. We found 127 substructure-specific loci (38 for medulla, 49 for pons, and 40 for midbrain) and 117 substructure-shared loci (57 shared by all substructures, 19 by medulla and pons, five by medulla and midbrain, and 36 by pons and midbrain) (Fig. 5a and Supplementary Data 12). For example, we found a locus (rs7523102) at 1q43 shared by all brainstem substructures, a medulla-specific locus (rs4888145) at 16q23.2, a pons-specific locus (rs637990) at 1p36.12, and a midbrain-specific locus (rs1000371) at 21q22.2 (Fig. 5b).

a The ideogram displays shared (sphere: shared by three substructures; triangle: shared by two substructures) and distinct (diamond) loci (two-sided P < 5.56 × 10−9, Bonferroni corrected, linear regression model for single-ancestry GWAS and fixed-effect model for cross-ancestry GWAS) among the medulla (red), pons (purple), and midbrain (blue) volumes. b The radar chart shows an example of a locus with lead variant rs7523102, which is shared by all three substructures, along with three examples of substructure-specific loci: rs4888145 for the medulla; rs637990 for the pons; and rs1000371 for the midbrain. The colors represent different brainstem substructures (medulla: red; pons: purple; midbrain: blue), while the scale indicates the significance (two-sided P-value, linear regression model, and fixed-effect model) of the most significant variant-trait association for each variant, in relation to its corresponding absolute and relative substructure volumes across all GWASs. c The Venn diagram illustrates the shared and distinct prioritized genes for medulla (red), pons (purple), and midbrain (blue) volumes. d The enrichment map depicts the hierarchical relationships of significant biological pathways enriched by 68 prioritized genes for medulla volumes (red), 89 for pons volumes (purple), and 76 for midbrain volumes (blue). Each circle denotes an enriched pathway, while the line thickness indicates pathway similarity scores measured by overlap genes between two pathways. Source data are provided as a Source Data file.
Among the 186 prioritized genes associated with brainstem volumetric traits, 154 were associated with the absolute or relative volume of medulla, pons, or midbrain. We divided the 154 genes into those specific to one, two, or three substructures, irrespective of absolute or relative volume. We identified 100 substructure-specific genes (32 for medulla, 38 for pons, and 30 for midbrain) and 54 substructure-shared genes (25 shared by all substructures, eight shared by medulla and pons, three shared by medulla and midbrain, and 18 shared by pons and midbrain) (Fig. 5c and Supplementary Data 13). For example, CNTN2 was identified as a medulla-specific gene that plays a key role in axonal outgrowth and cell migration in the caudal medulla40; HES1 was a pons-specific gene involved in the neurogenesis process41; and DSCAM was a midbrain-specific gene that regulates the delamination of neurons in the developing midbrain42.
We pooled the prioritized genes for medulla, pons, and midbrain without regard to absolute or relative volume, resulting in 68 genes for medulla, 89 for pons, and 76 for midbrain. We then used g:Profiler to conduct pathway enrichment analyses for the three gene groups, respectively. We found 306 unique enrichment pathways (Pc < 0.05, g:SCS corrected), comprising 182 pathways for medulla, 170 for pons, and 208 for midbrain (Supplementary Data 14). Next, we examined the pathways enriched by one, two, or three substructures, discovering 139 substructure-specific pathways (53 for medulla, 33 for pons, and 53 for pons) and 167 substructure-shared pathways (87 shared by all substructures, 12 shared by medulla and pons, 30 shared by medulla and midbrain, and 38 shared by pons and midbrain) (Supplementary Data 14). Some pathways shared by all substructures were involved in anatomical structure generation, artery morphogenesis, programmed cell death (Fig. 5d). One medulla-specific pathway was PIK3/AKT signaling, whose activation in rostral ventrolateral medulla could lead to neurogenic hypertension43. Some midbrain-specific pathways were clustered into the process of calcineurin-mediated signaling, which is involved in regulating cell shape changes during midbrain-hindbrain boundary formation44.
Overlap with other phenotypes
For each of the 713 lead variants from pooled locus-trait associations (578 unique variants), we conducted PheWAS using the FinnGen-MVP-UK Biobank meta-analysis database (https://mvp-ukbb.finngen.fi/), encompassing 330 diseases. We found 228 variants showing significant associations (P < 5 × 10−8) with 113 diseases across 18 categories (Fig. 6). Type 2 diabetes exhibited the highest number of associations with variants (n = 65) associated with brainstem volumetric traits. The diseases of the circulatory system demonstrated the highest number of associations with variants (n = 83) associated with brainstem volumetric traits, with hypertension being the disease in this category that had the most associations (n = 42). For instance, rs11877758 was a new locus-trait association with pons relative volume, which was associated with many diseases, such as migraine (P = 4.18 × 10−12; Supplementary Data 15).

The compound diagram illustrates the PheWAS results of the lead variant from pooled significant locus-trait associations with brainstem volumetric traits. Each point denotes a disease phenotype plotted based on its disease category (x-axis) and the count of significant associations (y-axis). The bar indicates the total number of association counts within each disease category. Neoplasms* neoplasms from hospital discharges, Neoplasms** neoplasms from cancer register, PheWAS phenome-wide association study, T2D type 2 diabetes. Source data are provided as a Source Data file.
We used LDSC18,45 to calculate the genetic correlations between nine brainstem volumetric traits and 26 non-imaging phenotypes based on the EUR-GWAS summary data (Supplementary Data 16). We identified 37 significant genetic correlations at P < 0.05, ten at BH-FDR-corrected Pc < 0.05, and four at Bonferroni-corrected Pc < 0.05. The four genetic correlations that passed Bonferroni correction were between ADHD and whole brainstem volume (rg = −0.13, P = 1.32 × 10−6), pons absolute volume (rg = −0.12, P = 2.93 × 10−5), midbrain absolute volume (rg = −0.18, P = 5.18 × 10−9), and midbrain relative volume (rg = −0.11, P = 2.87 × 10−5). Compared to the genetic correlations identified from previous GWAS summary data for brainstem volumetric traits12,13,16, we discovered 20 new genetic correlations at P < 0.05, seven at BH-FDR-corrected Pc < 0.05 (Fig. 7a), two at Bonferroni-corrected Pc < 0.05 (Supplementary Data 17).

a The left heat plot shows genetic correlations between brainstem volumetric traits and non-imaging phenotypes. An asterisk (*) means a genetic correlation significant at two-sided P < 0.05, a double asterisk (**) signifies significance at two-sided Pc < 0.05 (BH-FDR-corrected), and a triple asterisk (***) denotes significance at Bonferroni-corrected two-sided Pc < 0.05. The right Venn diagram depicts the relationship of genetic correlation findings (two-sided Pc < 0.05, BH-FDR-corrected) between this study and previous research. b The left ideogram shows genetic colocalizations (PP.H4 > 0.8) between brainstem volumetric traits and non-imaging phenotypes. The right Venn diagram illustrates the identified colocalizations between this study and previous research. c The left heat plot displays the counts of loci shared by each pair of brainstem volumetric traits and non-imaging phenotypes in conjunctional FDR (FDR < 0.05) analyses. The right Venn diagram illustrates the shared loci identified in this study and previous research. d The regional plots show a locus (7q21.2) that is shared by brainstem volumetric traits and blood pressure-related phenotypes, as identified by both conjunctional FDR and colocalization analyses. AD Alzheimer’s disease, ADHD attention deficit hyperactivity disorder, ALS amyotrophic lateral sclerosis, AN anorexia nervosa, ANX anxiety disorder, ASD autism spectrum disorders, BD bipolar disorder, DBP diastolic blood pressure, DEP depression, GGE genetic generalized epilepsy, iRBD isolated rapid eye movement sleep behavior disorder, LBD Lewy body dementia, MP morning person, PD Parkinson’s disease, PA Physical activity, PP pulse pressure, PP.H4 posterior probability of shared causal variant, PTSD posttraumatic stress disorder, RHR resting hear rate, SA sleep apnea, SBP systolic blood pressure, SCZ schizophrenia, SCP superior cerebellar peduncle, SWB subject well-being. Source data are provided as a Source Data file.
We employed coloc46 to identify the genetic loci shared by the nine brainstem volumetric traits and the 26 non-imaging phenotypes based on the EUR-GWAS summary data. Among the 528 locus-trait associations in EUR-GWAS (P < 5.56 × 10−9), we identified 113 genetic colocalizations (PP.H4 > 0.8) between all nine brainstem volumetric traits and 13 non-imaging phenotypes at 52 genetic loci (Fig. 7b and Supplementary Data 18). These included non-imaging phenotypes related to brain disorders (AD, ASD, bipolar disorder, depression, schizophrenia, and stroke), cardiovascular functions (diastolic and systolic blood pressure, pulse pressure, and resting heart rate), circadian rhythms (chronotype and morning person), and subjective well-being. For example, schizophrenia showed colocalizations at 2q33.1 with whole brainstem volume (PP.H4 = 0.91). The lead variants in this locus are eQTLs of TYW5, a regulator of neurodevelopment47. Compared to the genetic colocalizations identified from previous GWAS summary data for brainstem volumetric traits12,13,16, we found 90 new colocalizations (Fig. 7b). For instance, the two circadian rhythms phenotypes exhibited new colocalizations (PP.H4 = 0.89–0.99) at 19p13.11 with whole brainstem volume, medulla absolute volume, medulla relative volume, and pons relative volume.
Based on the non-overlapping EUR-GWAS summary data (Supplementary Data 19), we conducted conjFDR analyses using pleioFDR48 to identify shared genetic variants (conjFDR < 0.05) between each pair of nine brainstem volumetric traits and 26 non-imaging phenotypes. Among the 234 trait-phenotype pairs, 186 pairs showed shared genetic variants (conjFDR < 0.05; Supplementary Fig. 7), encompassing nine brainstem volumetric traits and all 26 non-imaging phenotypes. These trait-phenotype pairs shared 4846 loci (Fig. 7c and Supplementary Data 20), corresponding to 883 unique pleiotropic loci (Supplementary Data 21). Compared to conjFDR results identified from previous GWAS summary data12,13,16, we found 3352 new loci shared by brainstem volumetric traits and non-imaging phenotypes (Fig. 7c and Supplementary Data 20), corresponding to 171 new pleiotropic loci (Supplementary Data 21). For instance, we found a new locus (4q24) shared between schizophrenia and pons relative volume, where the lead SNP rs13107325 is a missense variant of SLC39A8 associated with schizophrenia49 and manganese deposition in the dorsal pons50. Additionally, numerous genetic sharing findings were consistent with colocalization analyses. For example, both conjFDR and colocalization analyses indicated that the locus (7q21.2) was shared between brainstem volumetric traits (absolute volumes of medulla and midbrain) and blood pressure-related phenotypes (systolic blood pressure and stroke) (Fig. 7d). Its lead SNP rs42039 is located in the UTR3 region of CDK6, a key player in cell cycle progression51. We also found genetic sharing between brainstem volumetric traits and six non-imaging phenotypes that were not significant in either genetic correlation or genetic colocalization analyses.
Discussion
By incorporating all available neuroimaging genetics data, particularly from the ABCD and CHIMGEN studies, we conducted cross-ancestry GWAS meta-analyses for whole brainstem volume in 103,098 individuals and for its substructure volumes in 78,062 individuals. We identified 713 locus-trait associations and 186 genes associated with nine brainstem volumetric traits, including 569 new associations. We discovered two ancestry-specific and 496 ancestry-shared associations, consistent with moderate to high cross-ancestry genetic correlations between EAS and EUR. We also provided new evidence for the merit of cross-ancestry fine-mapping by identifying 476 causal variants (PP > 0.8), surpassing the 244 identified from EUR fine-mapping. We revealed shared and distinct loci, genes, and pathways for midbrain, pons, and medulla volumes. We uncovered significant associations between 228 lead variants of loci linked to brainstem volumetric traits and 113 diseases, as well as shared genetic architectures between these brainstem volumetric traits and 26 physiological and pathological phenotypes related to the brainstem. These results align with the brainstem’s role in regulating heart rate, blood pressure, and circadian rhythms, along with its links to neuropsychiatric disorders such as AD, ADHD, depression, PD, and schizophrenia.
The first contribution of this study is the identification of over four times as many locus-trait associations compared to those significant at P < 5.56 × 10−9 in previous GWASs for brainstem volumetric traits12,13,14,15,16,17. We found 569 new locus-trait associations. This substantial number of new genetic findings may enhance the understanding of the genetic architectures of brainstem and substructure volumes. For instance, we found a new locus-trait association at 18q23 with the absolute volumes of the pons and midbrain, with rs470797 as the lead variant, a stop-gained variant of MBP. The protein encoded by the gene is a key component of the myelin sheath52. Therefore, the locus may influence the volumes of the pons and midbrain through regulating the development of the myelin sheath. Another new locus-trait association was identified between 12q13.13 and pons relative volume, with its lead variant rs56210711 serving as an eQTL for HOX3. The family of HOX genes may specify rhombomere identity in the developing hindbrain53. We also found a locus at Xp11.22, the only new locus on X-chromosome, which was linked to whole brainstem volume and pons absolute volume. Its lead variant rs12388258 is an eQTL of CINAP. This gene influences brain development54 and has been linked to an X-linked brain malformation with brainstem hypoplasia55.
The second contribution is the novel genetic findings derived from cross-ancestry analyses. We revealed moderate to high cross-ancestry genetic correlations (rg = 0.53–0.90) in brainstem volumetric traits between EUR and EAS, largely consistent with 496 ancestry-share associations as 248 times the number of ancestry-specific associations (n = 2). The volumes of the medulla and SCP exhibited lower cross-ancestry genetic correlations compared to other brainstem substructure volumes, likely due to imperfect segmentation resulting from the small size and complex routing and adjacency of the SCP, as well as the poorly defined boundary between the medulla and spinal cord. These results indicate that EAS and EUR individuals share similar genetic architectures for brainstem and substructure volumes. We also conducted statistical fine-mapping for the locus-trait associations of brainstem volumetric traits and identified 699 causal variants, of which 455 (65.1%) were detected solely through cross-ancestry fine-mapping. These findings further highlight the value of cross-ancestry fine-mapping in uncovering causal variants for brain imaging phenotypes56. For instance, rs55831773 was identified as a causal variant (PP = 1.00) for whole brainstem volume in both EUR and cross-ancestry fine-mapping. This variant is mapped to ATP1B2, linking to astrocyte degeneration in the brainstem57.
The third contribution of this study is the discovery of shared and distinct genetic architectures (loci, genes, and pathways) among midbrain, pons, and medulla volumes. We found 57 loci and 25 genes shared by all brainstem substructures, primarily related to neural development processes shared by brain structures. For example, we found a locus at 1q43 that was shared by all brainstem substructures. Its lead SNP, rs7523102, is located in the intronic region of AKT3, an AKT kinase involved a variety of biological processes, including brain development58, neuronal survival59, and myelination60. In addition, we identified 127 loci and 100 genes that were specific to individual brainstem substructures, providing more detailed insights into the genetic architecture of each substructure. For instance, a locus (16q23.2) was recognized as a medulla-specific locus, with its lead variant rs4888145 serving as an eQTL for the giant axonal neuropathy (GAN) gene. GAN encodes gigaxonin, which affects neuronal survival by regulating the degradation of the light chain of microtubule-associated protein 1B61. GAN mutation may lead to the disorganization of the neurofilament network62. These findings align with the compact axon arrangement in the medulla.
The last contribution is the identification of overlaps with other phenotypes related to brainstem volumetric traits. A previous study has conducted genetic correlation and conjunctional FDR analyses to investigate the shared genetic architectures between brainstem volumetric traits and eight common brain disorders12, but it failed to identify any significant genetic correlations after multiple testing correction, despite observing overlapping genetic loci with all disorders. Compared to the prior study12, this research performed additional analyses (PheWAS and genetic colocalization) and incorporated more phenotypes (330 in PheWAS and 26 in other analyses), uncovering numerous novel findings. In PheWAS, we identified significant associations between 228 variants related to brainstem volumetric traits and 113 diseases, which are involved in various organic systems. These results indicate a general role of brainstem in human health. We also identified ten significant genetic correlations (Pc < 0.05, FDR corrected), of which seven cannot be identified based on the previous GWAS12,13,16. For example, we found genetic correlations between midbrain absolute volume and conditions such as ADHD, depression, and PD, all of which are associated with abnormal DA levels. These results agree with the role of midbrain DA neurons in regulating voluntary movement, reward, salience, motivation, and emotion5,6. In addition to colocalization with brain disorders, we also found genetic colocalizations of brainstem volumetric traits with physiological phenotypes (heart rate, blood pressure, chronotype, and morning person) associated with cardiovascular function and circadian rhythm, which are well-known functions of the brainstem2,3,63. The shared genetic architectures with physiological phenotypes were then confirmed by the conjunctional FDR analyses, where brainstem volumetric traits shared more loci with physiological phenotypes than with any other phenotypes.
Several limitations should be noted when interpreting our findings. First, although we included 7094 Chinese Han individuals from the CHIMGEN study, we found only one association in EAS-GWAS. The smaller sample size of EAS individuals compared to that of EUR individuals may limit the power of our genetic findings in EAS-GWAS and could also bias the cross-ancestry GWAS for brainstem volumetric traits, as well as the effect size comparisons between EAS and EUR populations. Future GWAS should include more non-EUR individuals from EAS and other ancestral populations. Second, despite we controlled for the age effect in GWAS, we cannot rule out the bias from age differences among participants from ABCD (aged 8–11), CHIMGEN (aged 18–30), and UKB (aged 40–70). Third, consistent with previous GWAS for brainstem volumetric traits12,13,14,15,16,17, we accounted for variations in brain size among participants by including TIV as a covariate for the GWAS of absolute brainstem volumetric traits and adding total brainstem volume as an additional covariate for the GWAS of relative brainstem volumetric traits. However, the inclusion of heritable covariates in GWAS can introduce unintended bias, resulting in false positives23. Although we conducted sensitivity analyses and reported genetic association findings both with and without adjustment for TIV and total brainstem volume, a leave-one-chromosome-out polygenic score (LOCO-PGS) method64 could be a potential approach to better account for possible collider bias.
In conclusion, we presented the largest GWAS of brainstem and substructure volumes in an admixed-ancestry population. We identified 713 locus-trait associations, including 569 new associations. We also prioritized 186 genes and 699 causal variants. We further revealed shared and specific genetic architectures of brainstem volumetric traits across different ancestries and distinct substructures. We uncovered the overlaps of brainstem volumetric traits with other phenotypes. Our results may provide valuable insights into the genetic regulation and functional implication of the whole brainstem and its substructure volumes.
Methods
Participants and data preparation for GWAS
In GWAS for brainstem volumetric traits, we included three sets of raw neuroimaging genetics datasets (CHIMGEN, ABCD, and UKB) along with a set of GWAS summary data for whole brainstem volume14 from the ENIGMA, CHARGE, and UKB datasets. The CHIMGEN study was approved by the Medical Research Ethics Committees of Tianjin Medical University General Hospital and all other institutions, and written informed consent was obtained from each participant. The procedures of the ABCD study at most research centers were approved by a central Institutional Review Board (IRB) at the University of California, San Diego, and by local IRBs at a few centers65. All parents or caregivers provided written informed consent, and children provided written assent. We accessed the data under application ID 17607. The UKB study was approved by the National Health Service Research Ethics Service (21/NW/0157), and written informed consent was obtained from each participant. We accessed the data under application number 75556. Consistent with previous GWAS for brainstem substructure volumes12,13, we used the same Bayesian approach8 to calculate the absolute volumes of the whole brainstem, medulla, pons, midbrain, and SCP from CHIMGEN, ABCD, and UKB participants. The total brainstem volume (including SCP) was included as an additional covariate to perform GWAS for the relative volumes of the medulla, pons, midbrain, and SCP. As the previous GWAS14 used the automatic subcortical segmentation method to obtain the whole brainstem volume (excluding SCP), we also applied the same technique to calculate the whole brainstem volume for each participant in CHIMGEN, UKB, or ABCD, which was used in GWAS for whole brainstem volume throughout our study.
CHIMGEN participants and data preparation
All EAS participants were recruited from the CHIMGEN study (http://chimgen.tmu.edu.cn/), which collected genomic and neuroimaging data from 7306 healthy Chinese Han participants aged 18–30 years from 32 centers66. Among the 7306 participants, 7195 participants with DNA samples were genotyped using the Illumina ASA-750K (Asian Screening Array), specially designed for Asian individuals. We conducted sample-level and variant-level quality control, principal component analysis, and genetic data imputation17,56. After quality control, we included 7163 participants and 5,950,145 imputed autosomal and 171,620 X-chromosomal bi-allelic variants (MAF ≥ 0.5%, info > 0.9, and PHWE ≥ 1 × 10−7) in EAS-GWAS. The brain structural MRI data were acquired using ten types of 3.0-Tesla MRI scanners and 12 sets of scanning parameters (Supplementary Table 6). After excluding 61 participants without qualified structural MRI data, we calculated the volumes of the whole brainstem, medulla, pons, midbrain, and SCP for the remaining 7102 participants using the Bayesian segmentation8 implemented in FreeSurfer v7.0 (https://surfer.nmr.mgh.harvard.edu). With FreeSurfer v7.0, we also used the automatic subcortical segmentation to obtain the whole brainstem volume (excluding SCP). For each of the five brainstem volumetric traits, we removed the participants with volumes greater than five times the median absolute deviation (MAD) from the median value, finally included 7094 participants in EAS-GWAS. The quality control procedures for CHIMGEN data are presented in Supplementary Fig. 8.
ABCD participants and data preparation
The ABCD study (https://abcdstudy.org/) is a publicly available longitudinal dataset comprising over 10,000 participants aged 9–10 years at their baseline assessment from 21 research centers67. From the 11,099 participants with qualified imputed genotype data, we included 6605 EUR participants whose genetic ancestry was predominantly (> 80%, a recommended threshold) European, as estimated by SNPweights v2.168 based on SNP weights for European, West African, East Asian, and Native American populations. We applied MAF ≥ 0.5%, r2 > 0.6, and PHWE ≥ 1 × 10−7 to filter the variants and transformed them from GRCh38/hg38 to GRCh37/hg19 to align with the genetic data from CHIMGEN and UKB. In the ABCD-GWAS, we included 7,506,033 autosomal and 225,593 X-chromosomal bi-allelic variants. For the remaining 5060 admixed-ancestry participants, we used the same procedure to retain 5,853,740 autosomal and 184,972 X-chromosomal bi-allelic variants (MAF ≥ 0.5%, r2 > 0.6, and PHWE ≥ 1 × 10−7). Among the 6605 participants, 6060 had qualified whole-brainstem (excluding SCP) volume data obtained through automatic subcortical segmentation using FreeSurfer v7.0. Of the 6060 participants, 5815 had raw brain structural MRI data, from which we calculated the volumes of the whole brainstem, medulla, pons, midbrain, and SCP using the Bayesian segmentation algorithm8 in FreeSurfer v7.0. After removing participants with volumes exceeding five times the MAD from the median, we included 6060 and 5804 participants for whole brainstem volume and brainstem substructure volumes in ABCDEUR-GWAS. For the ABCDnon-EUR-GWAS, we included 4650 and 4491 participants for whole brainstem volume and brainstem substructure volumes (Supplementary Fig. 9).
UKB participants and data preparation
UKB (https://www.ukbiobank.ac.uk/) collected approximately 500,000 participants aged 40–69 years at recruitment from 22 research centers across the United Kingdom69. After initial genetic data quality control and imputation, the remaining 487,207 participants were included in further sample-level quality control. After excluding 651 participants with sex chromosome aneuploidy, 186 with sex mismatch, and 78,257 non-Caucasians, we included 408,113 EUR participants with qualified genomic data. Using the filtering criteria of MAF ≥ 0.5%, info > 0.9, and PHWE ≥ 1 × 10−7, we finally included 8,587,798 autosomal and 293,886 X-chromosomal bi-allelic variants in UKBEUR-GWAS. We also used these filtering criteria to perform variant-level quality control (MAF ≥ 0.5%, info > 0.9, and PHWE ≥ 1 × 10−7) to obtain 8,581,457 autosomal and 301,737 X-chromosomal bi-allelic variants in UKBnon-EUR-GWAS. Among the 408,113 participants, 53,448 had volumetric data of the brainstem and substructures obtained by the two segmentation approaches using FreeSurfer 7.0. We then visually checked raw brain structural images and the brainstem segmentation images, further excluding 914 participants with brain tumors, incomplete brainstem coverage, imaging artifacts, or incorrected brainstem segmentation. After removing participants with volumes greater than five times the MAD from the median, we finally included 48,204 participants for brainstem substructure volumes in UKBEUR-GWAS. We also excluded participants from the first released UKB neuroimaging data who were included in prior GWAS14 for whole brainstem volume, ultimately including 52,392 participants for the UKBEUR-GWAS of whole brainstem volume. Following imaging quality control (Supplementary Fig. 10), we also included 8281 participants in UKBnon-EUR-GWAS.
Reproducibility of brainstem segmentation
In 7096 CHIMGEN, 5815 ABCD, and 35,611 UKB participants, we calculated the intraclass correlation coefficient (ICC) of whole brainstem volumes obtained from the two brainstem segmentation methods. Although the automatic subcortical segmentation generated whole brainstem volume excluding SCP, while the Bayesian segmentation generated whole brainstem volume including SCP, we found high ICCs (0.956–0.997; Supplementary Table 7) in whole brainstem volumes obtained by the two brainstem segmentation methods. Among these participants, 24 CHIMGEN, 4077 ABCD, and 2698 UKB participants had brain structural MRI data acquired at two time points. For each participant, we calculated the brainstem and substructure volumes using the Bayesian brainstem segmentation based on the MRI data acquired at the two time points. For each dataset, we calculated the ICC of each volumetric trait obtained from the two time points to assess the test-retest reliability of the Bayesian brainstem segmentation, and found high ICCs (0.868–0.992; Supplementary Table 7).
Harmonization and normalization of brainstem and substructure volumes
The brain structural MRI data of the CHIMGEN, ABCD, and UKB participants were acquired using different MRI scanners, which may introduce bias into the integrated analyses of brainstem and substructure volumetric data derived from these scanners. To mitigate this bias, the Combat method was employed for each brainstem volumetric trait from each dataset to harmonize the volume data derived from MRI scans obtained using different scanners. This approach can eliminate between-scanner variation while preserving biological variability70. We tested the effect of ComBat harmonization in two participants who traveled to different centers and were scanned at 28 MRI scanners. For each participant, we segmented and calculated the volumes of the whole brainstem and four substructures based on the MRI data acquired from each scanner, using the coefficient of variation (CV) to assess between-scanner variations of the five traits. In both participants, we found that the CVs of these volumetric traits significantly reduced (Wilcoxon rank-sum test: P = 0.018) after harmonization (Supplementary Fig. 11). As the skewed data distribution would violate the assumption of normality when utilizing a linear regression model for GWAS, quantile normalization was subsequently applied to the harmonized brainstem and substructure volumetric data.
Covariates for GWAS
We controlled for age at imaging, genetically determined sex, age × sex, TIV, and the first genetic principal components (PCs) in CHIMGEN-GWAS, ABCD-GWAS, and UKB-GWAS, and additionally controlled for genotyping batches in ABCD-GWAS and UKB-GWAS. For each participant, TIV was estimated using FreeSurfer 7.0 and underwent ComBat harmonization and quantile normalization. We accounted for the first ten genetic PCs in CHIMGEN-GWAS, 32 in ABCD-GWAS, and 40 in UKB-GWAS, selected based on the complexity of the population.
GWAS for brainstem and substructure volumes
GWAS strategies
We performed a two-stage GWAS, followed by a combined GWAS in all participants. For each dataset with raw data, we randomly divided the participants into discovery (two-thirds of participants) and replication (one-third of participants) samples, which formed the basis for the discovery and replication stages, respectively. The prior GWAS summary data for whole brainstem volume14 were allocated solely to the replication stage. In both discovery and combined analyses, we adjusted for the nine brainstem volumetric traits, resulting in a Bonferroni-corrected P < 5.56 × 10−9. In the replication stage, we validated the variant-trait associations that were significant in the discovery stage and reported their significance at nominal P < 0.05 and Pc < 0.05 using both the BH-FDR and Bonferroni correction. As the UKB and ABCD cohorts, particularly ABCD, included twins and siblings, we excluded related participants from the replication samples of both datasets to ensure no related participants exist within each replication cohort or between the discovery and replication cohorts. For each dataset sharing the same ancestry and originating from the same cohort, we performed GWAS using fastGWA with a mixed linear model implemented in GCTA71. For non-EUR participants with diverse ancestries from ABCD (ABCDnon-EUR-GWAS) and UKB (UKBnon-EUR-GWAS), we used GENESIS Bioconductor package in R to evaluate genetic the relationship matrix. This package was designed for large-scale genetic analyses in samples with complex structures, including relatedness, population structure, and ancestry admixture based on PC-Air and PC-Relate72.
EUR-GWAS meta-analyses
Based on the summary statistics of ABCDEUR-GWAS and UKBEUR-GWAS for eight brainstem substructure volumes, we employed the inverse variance weighted (IVW) fixed-effect model in METAL73 to conduct EUR-GWAS meta-analyses for these brainstem volumetric traits. In the EUR-GWAS meta-analysis for whole brainstem volume, to fully use the available data resources, we also included the GWAS summary data for whole brainstem volume (n = 28,809) from ENIGMA, CHARGE, and UKB (first release) datasets14. The participants and genetic variants included in the EUR-GWAS meta-analyses for brainstem volumetric traits are detailed in Supplementary Table 2.
Cross-ancestry GWAS meta-analyses
Based on the summary statistics obtained from CHIMGEN-GWAS and EUR-GWAS meta-analyses, we conducted cross-ancestry GWAS meta-analyses (EUR-EAS-GWAS) for these nine brainstem volumetric traits using the IVW fixed-effect model in METAL73. Based on EUR-EAS-GWAS, ABCDnon-EUR-GWAS, and UKBnon-EUR-GWAS, we also conducted the admixed-ancestry GWAS meta-analyses (admixed-GWAS) for these brainstem volumetric traits. The participants and genetic variants included in the cross-ancestry GWAS meta-analyses are presented in Supplementary Table 2.
Population stratification estimation
For each GWAS, we used the genomic control inflation factor (λGC) and LDSC intercepts18 to estimate population stratification. λGC was calculated as the median of the resulting chi-squared (χ2) test statistics (z scores) divided by 0.4549, the expected median of the χ2 distribution with one degree of freedom. As a high λGC suggests either genomic inflation or polygenicity, we employed the LDSC intercept to distinguish between genomic inflation and polygenicity based on LD scores.
Defining LD references and independent associations and loci
We utilized imputed genotype data from 7094 EAS participants from CHIMGEN and 52,392 EUR participants from UKB, both with qualified genetic data and brainstem volumetric data, to construct EAS-LD and EUR-LD references, respectively. We also developed a cross-ancestry LD reference using a sample-weighted method based on the two datasets. Additionally, we employed imputed genotype data from 8281 non-EUR participants in the UKB to create a non-EUR-LD reference. Based on the summary statistics from CHIMGEN-GWAS, ABCDEUR-GWAS, UKBEUR-GWAS, EUR-GWAS, EUR-EAS-GWAS, or admixed-GWAS for each brainstem volumetric trait, we used the matched LD reference to identify independent variant-trait associations through PLINK clumping74 with the following steps: (1) all significant variants were compiled into a list of candidate variants; (2) the most significant variant was defined as the first lead variant (independent variant), and variants within 500 kb from and in LD with (r2 > 0.1) the lead variant were clumped; (3) the remaining variants formed a new list of candidate variants, and step (2) was repeated; and (4) the iterative process continued until the list was empty. We identified independent locus-trait associations by: (1) creating loci for independent variants by adding 500 kb to both sides of each variant; (2) merging loci within 500 kb; (3) merging loci if any independent variant of one locus was in LD (r2 > 0.1) with any independent variant of another locus; and (4) merging loci overlapped with the MHC or 8p23.1 region into a single locus. The associations of the remaining loci with this trait were defined as independent locus-trait associations.
As EUR samples were at least seven times larger than EAS samples in this study, we used the EUR-LD reference to pool GWAS results from different ancestries for each trait. We used the aforementioned strategies to identify independent variant-trait and locus-trait associations for each trait.
As the prior GWAS for brainstem and substructure volumes12,13,14,15,16,17 were primarily conducted in EUR participants, we used the EUR-LD reference to pool these GWAS to identify known independent variant-trait and locus-trait associations (P < 5.56 × 10−9). Using the EUR-LD reference and each known list of associations, we defined a new variant-trait association if the variant was 500 kb away from and not in LD (r2 < 0.1) with any variants of the same trait in the list of known variant-trait associations; and a new locus-trait association when the locus was 500 kb away from all known locus-trait associations and all lead variants in the locus were not in LD (r2 < 0.1) with any lead variants in the loci of all known locus-trait associations.
Sensitivity analyses
Previous study suggested that including highly heritable phenotypes as covariates in GWAS could introduce unintended bias. Therefore, we performed a series of analyses as the prior study suggested23. First, we evaluated phenotype correlation by calculating Spearman correlation coefficients among TIV, whole brainstem volume, and brainstem substructure volumes. Second, we repeated association analyses for the pooled variant-trait associations from the main analyses, excluding TIV and whole brainstem volume (only for relative volumes of substructures) as covariates. Third, we calculated the Spearman correlation of effect sizes for these variant-trait associations derived from the main and sensitivity analyses.
GWAS for brainstem nuclei volumes
We also conducted admixed-GWAS on the volumes of 58 brainstem nuclei. For each participant, we calculated the volumes of brainstem nuclei by applying the Brainstem Navigator Atlas75 to the preprocessed brain structural images of that participant. An automated processing pipeline76 was used to preprocess brain structural MRI data from UKB77, primarily based on the FSL software (https://fsl.fmrib.ox.ac.uk/fsl). The brain structural MRI data from CHIMGEN and ABCD were preprocessed using the pipeline for CHIMGEN66, primarily using the computational anatomy toolbox (CAT 12, version r1364, http://dbm.neuro.uni-jena.de/cat). The volume data for the 58 brainstem nuclei underwent the same quality control and normalization procedures as the volume data for the nine brainstem volumetric traits. Finally, only the 47,887 participants with both qualified genetic data and volumetric data for all brainstem nuclei were included in the admixed-GWAS meta-analyses. This group comprised 6845 EAS participants from CHIMGEN, 5828 EUR, and 4584 non-EUR participants from ABCD, and 30,630 EUR participants from UKB. However, we excluded non-EUR participants from the UKB in this analysis due to missing data, as only 457 (5.5%) of the 8281 participants had raw brain structural MRI data necessary for the analysis.
SNP-based heritability and cross-ancestry genetic correlation
We used LDSC18 to estimate the SNP-based heritability for brainstem volumetric traits in EAS and EUR based on the ancestry-specific LD reference and the GWAS summary statistics for autosomal variants. We applied the covariate-adjusted LDSC26 method to estimate the cross-ancestry SNP-based heritability for these traits, using the merged LD reference and GWAS summary statistics for autosomal variants, while adjusting for the top 40 genetic PCs derived from both CHIMGEN and UKB genetic data. After excluding variants in the MHC region, based on EAS-LD and EUR-LD references and cross-ancestry GWAS summary statistics for autosomal variants, we employed Popcorn (https://github.com/brielin/Popcorn)27 to estimate the cross-ancestry genetic-effect (not considering MAF information) and genetic-impact (considering MAF information) correlations between EAS and EUR for each brainstem volumetric trait.
Allele effect heterogeneity between EAS and EUR
We used the CQ-test to assess the allelic effect heterogeneity between EAS and EUR by comparing the differences in effect sizes of candidate variants derived from EAS-GWAS and EUR-GWAS. Cochran’s Q was estimated based on the χ2 test, with the null hypothesis positing homogeneity between the two ancestral populations and the alternative hypothesis suggesting heterogeneity between ancestral populations. The candidate variants were those included in the pooled variant-trait associations (P < 5.56 × 10−9) from EAS, EUR, and cross-ancestry GWAS in the total sample. Given the disparity in sample size between EAS-GWAS and EUR-GWAS, we performed the CQ-test in the discovery and replication stages using EUR-GWAS data from the respective stages. However, we included the EAS-GWAS data in the total sample throughout both stages. In the discovery stage, we defined the associations with CQ-test P ≥ 0.05 as ancestry-shared associations, while those with Pc < 0.05 (Bonferroni correction for the total number of associations tested) as ancestry-specific associations. In the replication stage, we validated the discovered ancestry-shared and ancestry-specific associations. A successful verification for ancestry-shared associations was defined as CQ-test P ≥ 0.05, while a successful verification for ancestry-specific associations was defined as CQ-test Pc < 0.05 (Bonferroni correction for the number of discovered ancestry-specific associations).
Statistical fine-mapping
For each locus of the pooled locus-trait association, we used ancestry-matched LD references and GWAS summary data to perform statistical fine-mapping by estimating the PP of each variant being a causal variant (PP > 0.8) using the PAINTOR tool30,31 with the MCMC model that allows multiple causal variants. We conducted fine-mapping using the ancestry-specific LD reference for pooled locus-trait associations significant only in EAS-GWAS or EUR-GWAS, and the cross-ancestry LD reference for pooled locus-trait associations significant only in cross-ancestry GWAS. When a locus-trait association was significant in two or more categories, we performed fine-mapping using the corresponding matched LD references. For pooled locus-trait associations significant in both EUR and cross-ancestry GWAS, we also conducted PAINTOR under the assumption of one causal variant and used the Wilcoxon rank-sum test to assess whether cross-ancestry fine-mapping can reduce the 95% credible sets for these loci compared to EUR-specific fine-mapping.
Functional annotations
We used FUMA33 to perform functional annotations for unique variants with PP > 0.1 in statistical fine-mapping for the pooled locus-trait associations identified by EAS-GWAS, EUR-GWAS, and cross-ancestry GWAS, respectively. We categorized these variants based on genic position, such as exon, intron, UTR, and intergenic region. We used the CADD score to prioritize deleterious and pathogenic variants, considering a variant pathogenic when the CADD score exceeded 12.37. Additionally, we employed the RDB score to prioritize variants in non-coding regions, with a classification scheme based on known and predicted regulatory elements.
Prioritized genes and pathways
We prioritized genes using the FLAMES framework35, which is based on the statistical fine-mapping results and pathway-naive Polygenic Priority Score (PoPS) results from the EUR-GWAS for brainstem volumetric traits. FLAMES integrates SNP-to-gene evidence with convergence-based evidence to produce a single XGBoost score for each gene. We prioritized genes based on the EUR-GWAS for the discovery, replication, and combined stages, respectively. The prioritized genes were defined by a cumulative precision exceeding 75%. We submitted the prioritized genes to g:Profiler38 (https://biit.cs.ut.ee/gprofiler/gost), a web server for enrichment analysis, to perform pathway enrichment analyses based on pre-specified pathways from GO36 (15,472 biological processes) and Reactome37 (2562 terms) databases. We corrected for multiple testing using the method (Pc < 0.05, g:SCS corrected) recommended by the tool. The significant GO biological processes and Reactome terms were visualized as a graph using Cytoscape78.
Shared and distinct genetic architectures between brainstem substructures
Shared and distinct genetic loci between brainstem substructures
From the pooled LD-independent loci associated with brainstem volumetric traits, we searched for loci that were associated with the volumes of one, two, or three substructures (medulla, pons, and midbrain), irrespective of absolute or relative volume. In this manner, we identified loci specific to the medulla, pons, and midbrain, along with loci shared by any two substructures or all three substructures.
Shared and distinct genes between brainstem substructures
From the prioritized genes associated with brainstem volumetric traits, we searched for genes prioritized for the volumes of one, two, or three brainstem substructures (medulla, pons, and midbrain), regardless of absolute or relative volume. Consequently, we identified genes specific to the medulla, pons, and midbrain, as well as those shared by any two substructures or all three substructures.
Shared and distinct enrichment pathways between brainstem substructures
Based on the prioritized genes for medulla, pons, and midbrain, regardless of absolute or relative volume, we used the above-mentioned method to perform pathway enrichment analyses for genes associated with medulla, pons, and midbrain volumes, respectively. Then, we searched for pathways enriched by the volumes of one, two, or three substructures to identify pathways specific to the medulla, pons, and midbrain, as well as those shared by any two substructures or all three.
Phenome-wide association studies
To provide insights into the genetic overlaps between brainstem volumetric traits and diseases, we performed a PheWAS to determine whether the lead variant of each pooled locus-trait associations with brainstem volumetric traits also demonstrated significant associations (P < 5 × 10−8) with human diseases. The GWAS summary statistics for 330 diseases were derived from the database of the FinnGen-MVP-UKB meta-analyses (https://mvp-ukbb.finngen.fi/).
Genetic sharing between brainstem volumetric traits and non-imaging phenotypes
Based on the available GWAS summary statistics for non-imaging phenotypes that are potentially associated with brainstem functions, we used three complementary methods (genetic correlation, genetic colocalization, and conjFDR) to identify shared genetic architectures between brainstem volumetric traits and non-imaging phenotypes. As GWASs for both traits and phenotypes are mainly conducted in the EUR population and most approaches require GWAS samples from the same ancestry, we included only 26 non-imaging phenotypes (Supplementary Data 16 and 19) with EUR-GWAS summary statistics. We then conducted the three genetic sharing analyses based on the EUR-GWAS summary data and the EUR-LD reference.
Genetic correlation analyses
We used LDSC18,45 to calculate the genetic correlations between nine brainstem volumetric traits and 26 non-imaging phenotypes based on the EUR-GWAS summary statistics and the EUR-LD reference.
Genetic colocalization analyses
We utilized a Bayesian colocalization method called coloc (https://chr1swallace.github.io/coloc/)46 to identify loci shared by each pair of nine brainstem volumetric traits (locus-trait associations with P < 5.56 × 10−9) and 26 non-imaging phenotypes (locus-phenotype associations with P < 5 × 10−8) based on the EUR-GWAS summary statistics. Using the default priors (P1 = 1 × 10−4, P2 = 1 × 10−4, and P12 = 1 × 10−5), we considered evidence for colocalization if PP.H4 (the posterior probability of shared causal variant) was greater than 0.8.
ConjFDR analyses
Based on the EUR-GWAS summary data without sample overlap and EUR-LD reference from the 1000 Genomes Project, we conducted the conjFDR analyses using pleioFDR (https://github.com/precimed/pleiofdr)48. The analysis was to identify shared variants (conjFDR < 0.05) between each pair of nine brainstem volumetric traits and 26 non-imaging phenotypes. The conjFDR value for each variant was defined as the maximum FDR value of that variant in the two mutual conditional FDR analyses. In our analyses, we excluded variants located in the MHC and 8p23.1 genomic regions. We pooled the shared variants for each trait-phenotype pair with the same criteria (<500 kb and LD r2 > 0.1) based on the EUR-LD reference and identified unique pleiotropic loci by merging loci within 500 kb across all trait-phenotype pairs. We annotated the lead variants of each shared loci using the same strategies.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Individual-level data were accessed under application number 75556 from the UK Biobank and 17607 from the ABCD. The GWAS summary statistics used in this work from following publicly available dataset: the ENIGMA study (https://enigma.ini.usc.edu/research/download-enigma-gwas-results/). All GWAS summary statistics of the brainstem and substructure volumes (EAS-GWAS, EUR-GWAS, EUR-EAS-GWAS, and admixed-GWAS) are publicly available at https://doi.org/10.5281/zenodo.13382122. The CHIMGEN raw data are not publicly available as this could compromise research participant privacy/consent. Source data are provided with this paper.
Code availability
We made use of publicly available software and tools. All codes used to generate results reported in this paper are publicly available at https://doi.org/10.5281/zenodo.17404660. The following software packages were used for data analyses: FreeSurfer (v.7.0.0): https://surfer.nmr.mgh.harvard.edu; CAT12 (r1364): http://dbm.neuro.uni-jena.de/cat; Combat Harmonization: https://github.com/Jfortin1/ComBatHarmonization; PLINK software (v.1.9 and v.2.0): www.cog-genomics.org/plink; SNPweights (v2.1): https://hsph.harvard.edu/research/price-lab/software/; GCTA software (v1.94.0): https://yanglab.westlake.edu.cn/software/gcta/; R (v4.2.2): https://cran.r-project.org/; GENESIS (v2.38.0): https://www.bioconductor.org/packages/release/bioc/html/GENESIS.html; PhenoGram: https://visualization.ritchielab.org/phenograms/plot/; LocusZoom: https://genome.sph.umich.edu/wiki/LocusZoom; CMplot: https://github.com/YinLiLin/; LDSC software: https://github.com/bulik/ldsc; cov-LDSC software: https://github.com/yang-luo-lab/cov-ldsc; METAL software: http://csg.sph.umich.edu/abecasis/Metal/; Popcorn software: https://github.com/brielin/Popcorn; PAINTOR tool (v.3.0): https://github.com/gkichaev/PAINTOR_V3.0; FUMA website: https://fuma.ctglab.nl/; FLAMES: https://github.com/Marijn-Schipper/FLAMES; g:profiler: https://biit.cs.ut.ee/gprofiler_beta/gost; coloc: https://chr1swallace.github.io/coloc/; conjFDR: https://github.com/precimed/pleiofdr.
References
Del Negro, C. A., Funk, G. D. & Feldman, J. L. Breathing matters. Nat. Rev. Neurosci. 19, 351–367 (2018).
Guyenet, P. G. The sympathetic control of blood pressure. Nat. Rev. Neurosci. 7, 335–346 (2006).
Herring, N., Kalla, M. & Paterson, D. J. The autonomic nervous system and cardiac arrhythmias: current concepts and emerging therapies. Nat. Rev. Cardiol. 16, 707–726 (2019).
Poe, G. R. et al. Locus coeruleus: a new look at the blue spot. Nat. Rev. Neurosci. 21, 644–659 (2020).
Smidt, M. P. & Burbach, J. P. H. How to make a mesodiencephalic dopaminergic neuron. Nat. Rev. Neurosci. 8, 21–32 (2007).
Castellanos, F. X. & Tannock, R. Neuroscience of attention-deficit/hyperactivity disorder: the search for endophenotypes. Nat. Rev. Neurosci. 3, 617–628 (2002).
Steriade, M. Arousal: revisiting the reticular activating system. Science 272, 225–226 (1996).
Iglesias, J. E. et al. Bayesian segmentation of brainstem structures in MRI. Neuroimage 113, 184–195 (2015).
Milella, G. et al. Medulla oblongata volume as a promising predictor of survival in amyotrophic lateral sclerosis. Neuroimage-Clin. 34, 103015 (2022).
Chong, C. D., Plasencia, J. D., Frakes, D. H. & Schwedt, T. J. Structural alterations of the brainstem in migraine. Neuroimage-Clin. 13, 223–227 (2017).
Bosco, P. et al. Brainstem enlargement in preschool children with autism: results from an intermethod agreement study of segmentation algorithms. Hum. Brain Mapp. 40, 7–19 (2019).
Elvsåshagen, T. et al. The genetic architecture of human brainstem structures and their involvement in common brain disorders. Nat. Commun. 11, 4016 (2020).
Smith, S. M. et al. An expanded set of genome-wide association studies of brain imaging phenotypes in UK Biobank. Nat. Neurosci. 24, 737–745 (2021).
Satizabal, C. L. et al. Genetic architecture of subcortical brain structures in 38,851 individuals. Nat. Genet. 51, 1624–1636 (2019).
Zhao, B. et al. Genome-wide association analysis of 19,629 individuals identifies variants influencing regional brain volumes and refines their genetic co-architecture with cognitive and mental health traits. Nat. Genet. 51, 1637–1644 (2019).
García-Marín, L. M. et al. Genomic analysis of intracranial and subcortical brain volumes yields polygenic scores accounting for variation across ancestries. Nat. Genet. 56, 2333–2344 (2024).
Fu, J. L. et al. Cross-ancestry genome-wide association studies of brain imaging phenotypes. Nat. Genet. 56, 1110–1120 (2024).
Bulik-Sullivan, B. K. et al. LD Score regression distinguishes confounding from polygenicity in genome-wide association studies. Nat. Genet. 47, 291–295 (2015).
Stutzbach, L. D. et al. The unfolded protein response is activated in disease-affected brain regions in progressive supranuclear palsy and Alzheimer’s disease. Acta Neuropathol. Commun. 1, 31 (2013).
Mandai, K. et al. LIG family receptor tyrosine kinase-associated proteins modulate growth factor signals during neural development. Neuron 63, 614–627 (2009).
Freilinger, T. et al. Genome-wide association analysis identifies susceptibility loci for migraine without aura. Nat. Genet. 44, 777–782 (2012).
Nyholt, D. R., Borsook, D. & Griffiths, L. R. Migrainomics - identifying brain and genetic markers of migraine. Nat. Rev. Neurol. 13, 725–741 (2017).
Aschard, H., Vilhjálmsson, B. J., Joshi, A. D., Price, A. L. & Kraft, P. Adjusting for heritable covariates can bias effect estimates in genome-wide association studies. Am. J. Hum. Genet. 96, 329–339 (2015).
Lieberburg, I. et al. Cloning of a cDNA encoding the rat high molecular weight neurofilament peptide (NF-H): developmental and tissue expression in the rat, and mapping of its human homologue to chromosomes 1 and 22. Proc. Natl. Acad. Sci. USA 86, 2463–2467 (1989).
Jacquier, A. et al. Cryptic amyloidogenic elements in mutant NEFH causing Charcot-Marie-Tooth 2 trigger aggresome formation and neuronal death. Acta Neuropathol. Commun. 5, 55 (2017).
Luo, Y. et al. Estimating heritability and its enrichment in tissue-specific gene sets in admixed populations. Hum. Mol. Genet. 30, 1521–1534 (2021).
Brown, B. C., Asian Genetic Epidemiology Network Type 2 Diabetes Consortium, Ye, C. J., Price, A. L. & Zaitlen, N. Transethnic genetic-correlation estimates from summary statistics. Am. J. Hum. Genet. 99, 76–88 (2016).
Krishnan, N. et al. PTP1B inhibition suggests a therapeutic strategy for Rett syndrome. J. Clin. Invest. 125, 3163–3177 (2015).
Shin, J. et al. PRAJA is overexpressed in glioblastoma and contributes to neural precursor development. Genes Cancer 8, 640–649 (2017).
Kichaev, G. et al. Integrating functional data to prioritize causal variants in statistical fine-mapping studies. PLoS Genet. 10, e1004722 (2014).
Kichaev, G. et al. Improved methods for multi-trait fine mapping of pleiotropic risk loci. Bioinformatics 33, 248–255 (2017).
van Muiswinkel, F. L. et al. Expression of NAD(P)H:quinone oxidoreductase in the normal and Parkinsonian substantia nigra. Neurobiol. Aging 25, 1253–1262 (2004).
Watanabe, K., Taskesen, E., van Bochoven, A. & Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 8, 1826 (2017).
Lin, T. T., Islam, O. & Heese, K. ABC transporters, neural stem cells and neurogenesis - a different perspective. Cell Res. 16, 857–871 (2006).
Schipper, M. et al. Prioritizing effector genes at trait-associated loci using multimodal evidence. Nat. Genet. 57, 323–333 (2025).
Ashburner, M. et al. Gene Ontology: tool for the unification of biology. Nat. Genet. 25, 25–29 (2000).
Gillespie, M. et al. The Reactome Pathway Knowledgebase 2022. Nucleic Acids Res. 50, D687–D692 (2022).
Kolberg, L. et al. g:Profiler-interoperable web service for functional enrichment analysis and gene identifier mapping (2023 update). Nucleic Acids Res. 51, W207–W212 (2023).
Yung, A. R. et al. Netrin-1 confines rhombic lip-derived neurons to the CNS. Cell Rep. 22, 1666–1680 (2018).
Kyriakopoulou, K., de Diego, I., Wassef, M. & Karagogeos, D. A combination of chain and neurophilic migration involving the adhesion molecule TAG-1 in the caudal medulla. Development 129, 287–296 (2002).
Hirata, H., Tomita, K., Bessho, Y. & Kageyama, R. Hes1 and Hes3 regulate maintenance of the isthmic organizer and development of the mid/hindbrain. Embo J. 20, 4454–4466 (2001).
Arimura, N. et al. DSCAM regulates delamination of neurons in the developing midbrain. Sci. Adv. 6, eaba1693 (2020).
Wu, K. L. et al. Redox-sensitive oxidation and phosphorylation of PTEN contribute to enhanced activation of PI3K/Akt signaling in rostral ventrolateral medulla and neurogenic hypertension in spontaneously hypertensive rats. Antioxid. Redox Signal 18, 36–50 (2013).
Sahu, S. U. et al. Calcium signals drive cell shape changes during zebrafish midbrain-hindbrain boundary formation. Mol. Biol. Cell 28, 875–882 (2017).
Bulik-Sullivan, B. et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 47, 1236–1241 (2015).
Wallace, C. Eliciting priors and relaxing the single causal variant assumption in colocalisation analyses. PLoS Genet. 16, e1008720 (2020).
Li, S. et al. Regulatory variants at 2q33.1 confer schizophrenia risk by modulating distal gene TYW5 expression. Brain 145, 770–786 (2022).
Andreassen, O. A. et al. Improved detection of common variants associated with schizophrenia and bipolar disorder using pleiotropy-informed conditional false discovery rate. PLoS Genet. 9, e1003455 (2013).
Mealer, R. G. et al. The schizophrenia-associated variant in SLC39A8 alters protein glycosylation in the mouse brain. Mol. Psychiatry 27, 1405–1415 (2022).
Anagianni, S. & Tuschl, K. Genetic disorders of manganese metabolism. Curr. Neurol. Neurosci. Rep. 19, 33 (2019).
Fassl, A., Geng, Y. & Sicinski, P. CDK4 and CDK6 kinases: from basic science to cancer therapy. Science 375, eabc1495 (2022).
Lindquist, R. A. et al. Identification of proliferative progenitors associated with prominent postnatal growth of the pons. Nat. Commun. 7, 11628 (2016).
Gaufo, G. O., Thomas, K. R. & Capecchi, M. R. Hox3 genes coordinate mechanisms of genetic suppression and activation in the generation of branchial and somatic motoneurons. Development 130, 5191–5201 (2003).
Wang, G. S. et al. Transcriptional modification by a CASK-interacting nucleosome assembly protein. Neuron 42, 113–128 (2004).
Najm, J. et al. Mutations of CASK cause an X-linked brain malformation phenotype with microcephaly and hypoplasia of the brainstem and cerebellum. Nat. Genet. 40, 1065–1067 (2008).
Liu, N. et al. Cross-ancestry genome-wide association meta-analyses of hippocampal and subfield volumes. Nat. Genet. 55, 1126–1137 (2023).
Magyar, J. P. et al. Degeneration of neural cells in the central nervous system of mice deficient in the gene for the adhesion molecule on Glia, the beta 2 subunit of murine Na,K-ATPase. J. Cell Biol. 127, 835–845 (1994).
Baek, S. T. et al. An AKT3-FOXG1-reelin network underlies defective migration in human focal malformations of cortical development. Nat. Med. 21, 1445–1454 (2015).
Dudek, H. et al. Regulation of neuronal survival by the serine-threonine protein kinase Akt. Science 275, 661–665 (1997).
Flores, A. I. et al. Constitutively active Akt induces enhanced myelination in the CNS. J. Neurosci. 28, 7174–7183 (2008).
Allen, E. et al. Gigaxonin-controlled degradation of MAP1B light chain is critical to neuronal survival. Nature 438, 224–228 (2005).
Mahammad, S. et al. Giant axonal neuropathy-associated gigaxonin mutations impair intermediate filament protein degradation. J. Clin. Investig. 123, 1964–1975 (2013).
Hastings, M. H., Maywood, E. S. & Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 19, 453–469 (2018).
McCaw, Z. R. et al. Pitfalls in performing genome-wide association studies on ratio traits. HGG Adv. 6, 100406 (2025).
Auchter, A. M. et al. A description of the ABCD organizational structure and communication framework. Dev. Cogn. Neuros-Neth. 32, 8–15 (2018).
Xu, Q. et al. CHIMGEN: a Chinese imaging genetics cohort to enhance cross-ethnic and cross-geographic brain research. Mol. Psychiatry 25, 517–529 (2020).
Casey, B. J. et al. The adolescent brain cognitive development (ABCD) study: imaging acquisition across 21 sites. Dev. Cogn. Neuros-Neth. 32, 43–54 (2018).
Chen, C. Y. et al. Improved ancestry inference using weights from external reference panels. Bioinformatics 29, 1399–1406 (2013).
Bycroft, C. et al. The UK Biobank resource with deep phenotyping and genomic data. Nature 562, 203–209 (2018).
Fortin, J. P. et al. Harmonization of cortical thickness measurements across scanners and sites. Neuroimage 167, 104–120 (2018).
Jiang, L. D. et al. A resource-efficient tool for mixed model association analysis of large-scale data. Nat. Genet. 51, 1749–1755 (2019).
Gogarten, S. M. et al. Genetic association testing using the GENESIS R/Bioconductor package. Bioinformatics 35, 5346–5348 (2019).
Willer, C. J., Li, Y. & Abecasis, G. R. METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics 26, 2190–2191 (2010).
Purcell, S. et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575 (2007).
Bianciardi, M. et al. Toward an in vivo neuroimaging template of human brainstem nuclei of the ascending arousal, autonomic, and motor systems. Brain Connect. 5, 597–607 (2015).
Zhang, Y., Brady, M. & Smith, S. Segmentation of brain MR images through a hidden Markov random field model and the expectation-maximization algorithm. IEEE Trans. Med. Imaging 20, 45–57 (2001).
Miller, K. L. et al. Multimodal population brain imaging in the UK Biobank prospective epidemiological study. Nat. Neurosci. 19, 1523–1536 (2016).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504 (2003).
Acknowledgements
We are grateful to all participants and researchers from CHIMGEN, UKB, and ABCD, who generously donated their time to make these resources available. We acknowledge funding from the National Natural Science Foundation of China (82430063, 82030053, 81425013 to C.Y., 82330058, T2341014 to S.Q., and 82402253 to J.F.), National Key Research and Development Program of China (2018YFC1314300 to C.Y.), Tianjin Key Medical Discipline Construction Project (TJYXZDXK-3-008C to C.Y.), and Tianjin Natural Science Foundation (25JCQNJC00020 to J.F.). We are grateful to the ENIGMA for providing the GWAS summary statistics of whole brainstem volume.
Author information
Authors and Affiliations
Consortia
Contributions
C.Y., H.X., and J.F. designed the study. C.Y. and H.X. wrote the article. H.X. analyzed the data. C.Y., Z.G., S.Q., and W.Z. supervised this work. C.Y., Z.G., J.C., M.W., L.Z., G.C., Y.Q.Y., W.H.L., H.Z., B.G., X.X., T.H., Z.W.Y., Q.Z., W.Q., F.L., M.L., S.W., Q.X., J.Y.X., C.W., N.L., Y.J., H.X., P.Z., W.L., W.W., D.S., S.L., Z.H.Y., F.C., J.Z., W.S., Y.M., D.W., J.-H.G., Y.J.Y., K.X., J.F.X., B.Z., X.Z., X.-N.Z., M.J.L., Z.X.Y., S.Q., W.Z. acquired the data. All authors critically reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Elleke Tissink and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Xue, H., Fu, J., Geng, Z. et al. The genetic architecture of brainstem structures. Nat Commun 17, 526 (2026). https://doi.org/10.1038/s41467-025-67221-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-67221-6
