
Abstract
Collective cell migration during embryonic development, wound healing, and cancer metastasis requires the emergence of leader cells at the migration front. Despite their physiological relevance, the full mechanisms underlying the emergence of leader cells remain elusive. Here we report that leader cells display a peripheral accumulation of lysosomes in diverse model systems for wound healing, including cultured epithelial monolayer, mouse embryonic skin, and Drosophila embryos. This accumulation involves cellular contractile forces driving lysosomal transport along microtubules towards the leading edge. Indeed, we control leader cell emergence by manipulating lysosomal movement on microtubules. We further find that peripheral lysosomes associate with Rac1 molecules at the leading periphery, regulating local Rac1-activity, triggering actin polymerization and promoting lamellipodium formation. Taken together, we demonstrate that beyond their catabolic role, lysosomes act as an intracellular platform that links mechanical and biochemical signals to control the emergence of leader cells.
Similar content being viewed by others
Introduction
Collective cell migration is the coordinated movement of cells, which is essential for organogenesis, embryo development, wound healing, tissue regeneration, and cancer metastasis1,2,3,4,5,6,7,8,9,10,11,12,13. For successful migration of epithelial cell collectives, a few cells at the migration front undergo phenotypic transformation to form distinct lamellipodial structures and emerge as leader cells3,14. These cells provide guidance to collective cell migration13,15,16 and are critical for epithelial wound healing2,17, branching morphogenesis18,19, and cancer metastasis1,11,20,21. Despite this physiological importance, what factors selectively designate only a few cells at the migration front as the leader cells remain largely unknown. Nevertheless, recent works have suggested that both mechanical and biochemical cues are crucial for this process22,23. Mechanical cues, such as the spontaneous fluctuations in the cellular force field, which is defined by the cell-cell and cell-matrix forces, influence the leader cell formation22. Prior to the appearance of characteristic lamellipodial structures in leader cells, elevated traction forces and tensile stresses appear behind the would-be leader cells22. These results suggest a non-cell autonomous regulation of leader cell emergence. In addition to this mechanical regulation, spatial patterns of cellular signaling proteins such as p120-catenin24, Notch-Dll425, and RhoA23 distinguish the leader cells from other non-leader cells at the migration front and the follower cells behind the leaders. For example, the intrinsic polarized activity of Rac and RhoA GTPases is an integral signature of leader cell emergence23. Cues for leader cell emergence can also come from the extracellular milieu through the interaction of the migrating cells with the compliant extracellular matrix via focal adhesions3 and with the soluble signaling via self-generated chemokine gradient26. Furthermore, a recent study has identified the tumor suppressor protein p53 as a key marker for leader cells2. Elevated levels of p53 guide leader cell emergence and modulate the p21-CDK pathway to regulate collective migration2. But a connection between the mechanical cues and the biochemical events leading to leader cell emergence remains elusive.
As the sedentary cells become motile and develop a clear front-to-rear polarity, the intracellular organization undergoes a conspicuous transformation3. This transformation requires both breaking the existing structures and making new structures. In this respect, previous studies have primarily focused on elucidating the dynamics of the actin cytoskeleton during the emergence of leader cells. However, the role of other intracellular structures and cellular organelles in this process remains unexplored. Relevantly, one may presume that lysosomes, which are crucial for the catabolism and recycling of various biomolecules, might play a critical role in the cellular transformation to a motile state. Indeed, lysosomes can act as a signaling hub regulating cell growth, death, and differentiation27. They also house various proteins which regulate cell adhesion28,29 and migration30. At a single-cell level, lysosomes regulate the dynamics of focal adhesions in migrating fibroblasts29. During dendritic cell migration, lysosomes accumulate in the direction opposite of migration and locally release calcium to modulate actomyosin dynamics supporting cell propulsion31. Lysosomes also respond to the mechanical properties of the extracellular matrix by altering their intracellular position32. Interestingly, a parallel study demonstrates that peripheral lysosomes promote invadopodium-based matrix degradation and invasion of melanoma cells, highlighting the importance of lysosome positioning in carcinogenesis33. While it seems that lysosomes can integrate mechanical signals and biochemical signals, very little is known about the specific roles of this organelle in collective cell migration.
In this work, we explore the roles of lysosomes in the emergence of leader cells during collective cell migration during epithelial wound healing (or gap closure) and show that the peripheral accumulation of lysosomes facilitates leader cell emergence from the epithelial wound margin. Importantly, previous works have established that peripheral lysosomes contribute to cancer cell invasion primarily through exocytosis and ECM degradation34. In contrast, our study reveals a mechanosensitive role of lysosomes in collective epithelial migration, demonstrating that their peripheral accumulation is not just a consequence of motility but an active driver of leader cell emergence. We also show that lysosomes regulate Rac1 activation and lamellipodium formation, integrating mechanical cues with biochemical signaling to coordinate epithelial wound healing and morphogenesis. These findings redefine lysosomes as key mechanotransductive organelles that control collective migration beyond their conventional degradative functions.
Results
Lysosomes accumulate at the leading periphery of the leader cells in various wound healing models
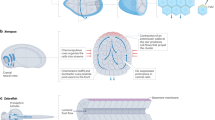
The biochemical, molecular, and cellular mechanisms underlying collective cell migration during the wound healing process have been studied employing both in vivo and ex vivo model systems. For instance, stage-15 Drosophila melanogaster embryos have been valuable in vivo models35, while mouse embryonic skin wounds have been utilized as ex vivo systems36,37. It is known that the epidermal wound closure in stage-15 Drosophila embryos occurs by a combination of pursed-string contraction of actin cables and lamellipodial crawling35. First, to study the localization dynamics of lysosomes in wound healing, we used glass needles (0.5 µm inner diameter and 100 µm outer diameter) mounted on a femtojet microinjection system to create wounds in the stage-15 embryos. Since the wound got closed completely in one-to-two hours, we fixed these embryos 15 min post-wounding to capture an intermediate stage of wound closure (Supplementary Fig. 1A). We then stained these embryos for lysosomes and actin, using an antibody against an established lysosome marker protein, LAMP1, and fluorescence-labeled phalloidin, respectively (Supplementary Fig. 1B) and observed a few cells at the migration front emerging as leader cells having lamellipodial structures (Supplementary Fig. 1B). Interestingly, the images revealed a distinct lysosomal accumulation at the cell periphery in these cells (Supplementary Fig. 1B). Next, we analysed the localization of lysosomes in mouse embryonic skin wounds. We subjected the mouse embryonic skin samples that were dissected out of day 16.5 embryos, to incisions and allowed the wound to heal for 30 min in an air-media interface in a humidified chamber maintained at 37 °C and 5% CO2 (Fig. 1A–C). We then fixed and stained these samples for lysosomes and actin (Fig. 1D, Supplementary Fig. 1C). Here, we also observed the movement of cells into the wound gap of the skin tissue (Fig. 1C) and peripheral lysosome distribution at the wound margin (Fig. 1D, Supplementary Fig. 1C). Although challenging, super-resolution imaging of mouse embryonic skin wounds revealed leader cells within the multilayered tissue. The leader cells with an extended lamellipodia have lysosomes polarized at the cell front, while a non-leader marginal cell did not show any peripheral lysosomal polarization (Fig. 1E). These results suggest that lysosome accumulation at the leading edge of wounds occurs in physiologically relevant model systems and may represent an evolutionarily conserved mechanism in collective cell migration.

A Diagrammatic representation of skin epithelium dissection, wounding and wound healing setup for day 14.5–16.5 mouse embryos. B Schematic representation of mouse embryonic skin wound showing the leading edge of collectively migrating cells. C 3-Dimensional projected image of mouse skin embryonic skin wound post 30 min of incision. Scale bar shows the z-slices of the tissue and the height. D Representative 3-D renders created on Imaris using the blend volume viewer, of a confocal image of mouse embryonic skin wound post 30 min of incision. The white arrowheads mark the lysosome accumulation in cells at the leading edge. Scale bar −15 µm. E Representative super-resolution images of leader and non-leader marginal cells in mouse embryonic skin wounds post 30 min of incision. F, G Representative confocal images of migrating monolayer of MDCK cells fixed at T-0 (F) and T-2 h (G) post confinement lift-off. Yellow arrowheads in (G) mark regions of lysosome accumulation in an emerging leader cell. H Schematic representation of Peripheral Occupancy Fraction (POF) analysis, used in this study to measure peripheral lysosome accumulation. ROI at the cell edge is marked by a black-dotted box. Junctional lysosomal fraction was measured by adding lysosome intensity in all junctional ROIs (marked by green-dotted box) and dividing by total lysosome intensity. I Dot-column plot showing POF for non-leader marginal and leader cells at T-0 and T-2 h in MDCK cells (marginal cells T-0 (n = 40) and T-2 h (n = 27); n = 26 leader cells analyzed from 3 independent experiments). J Junctional occupancy fraction of lysosomes was calculated for non-leader marginal (n = 40) and leader cells (n = 23), and is represented in the plot (Data from 3 independent experiments). Statistical significance was assessed using an unpaired Student’s t-test with Welch’s correction (two-tailed). K Dot-column plot showing POF for non-leader marginal and leader and non-leader marginal cells measured for mouse embryonic skin wounds (51 marginal cells and 44 leader cells were analyzed from 3 independent experiments). Statistical significance assessed using unpaired Student’s t-test with Welch’s correction (two-tailed). *** and **** denotes p-value < 0.001 and p < 0.0001, n.s is not significant. Scale bar-10 µm. All data are mean ± s.e.m.
However, the complex 3D architecture of these models makes it challenging to track individual leader cells and precisely quantify lysosome dynamics. To investigate the cellular mechanisms underlying lysosomal redistribution and leader cell emergence, we transitioned to a well-established gap closure assay2,8,12,22,23,38 using cultured epithelial monolayers. This model provided precise single-cell tracking and enabled rigorous quantifications, allowing for a more detailed mechanistic analysis of lysosome positioning during collective migration. We grew Madin-Darby canine kidney (MDCK) epithelial cells to confluence into the wells of a migration chamber (Supplementary Fig. 1D). In this system, after lifting off the confinement, cells migrated as two opposing epithelial sheets. Leader cells emerged from the collectively migrating monolayer (Supplementary Fig. 1E) approximately two hours after confinement lift-off. These cells, positioned at the forefront of the migrating sheet, extended large, actin-rich lamellipodia, which served as a key morphological feature for their identification25. To further investigate the dynamics of lysosomes in migrating epithelial monolayers, we fixed the migrating epithelial monolayer samples at 0, 15, 30, 60, 120, and 240 min post-confinement lift-off and immunostained them for lysosomes and actin (Supplementary Fig. 2A). We observed that in the cells at the migration front (marginal cells), lysosomes accumulated at the cell periphery, whereas the cells in the follower layer did not show any change in lysosome localization (Supplementary Fig. 2A) indicating that lysosome accumulation at the cell periphery in cells at the wound margin increases in a time-dependent manner. Interestingly, among the marginal cells, we observed further enhanced lysosome accumulation at the leading periphery of the leader cells, which was marked by their extended lamellipodial structures (Fig. 1F, G). We subsequently computed the fraction of peripherally localized lysosomes relative to the total lysosome intensity in a cell and reported it as Peripheral Occupancy Fraction (POF) (Fig. 1H). Our method of measuring POF takes into account the area of the cell, as the fraction is calculated by dividing the integrated densities (mean intensity*area). This quantitative analysis revealed a progressive increase in POF for lysosomes in cells at the margin as the migration progresses with time (Supplementary Fig. 2B). Further, a comparison of POF between the leader and non-leader marginal cells revealed a significantly higher fraction of lysosomes localized to the cell periphery in leader cells than in non-leader cells (Fig. 1I). Also, a closer examination of the distribution of lysosomes in leader and non-leader marginal cells revealed a lower junctional lysosome pool in leader cells as compared to non-leader marginal cells (Fig. 1J). In leader cells, lysosomes predominantly localize to the growing lamellipodia in the direction of migration, with minimal alignment along cell-cell junctions. In contrast, non-leader marginal cells exhibit lysosome enrichment along the actin cytoskeleton, but a significant fraction aligns with cell-cell junctions (Fig. 1G). This junctional occupancy was quantified as the fraction of lysosomal intensity at junctions relative to the total intensity (Fig. 1J). Lysosome accumulation at the cell periphery was measured using the POF analysis for leaders and non-leader marginal cells in mouse embryonic skin wounds (Fig. 1K). In our mouse embryonic skin wound assays, we observe a significantly higher lysosome accumulation at the cell periphery in leader cells compared to non-leader marginal cells. To further quantitatively validate lysosome accumulation to the cell periphery in leader versus non-leader marginal cells in the two-dimensional gap closure assay, we analyzed the images using a custom-written program that measures the mean distance of the weighted center of mass (organelle intensity) of the organelle from the cell edge. We called this parameter Mean Distribution from Edge or MDE. It revealed the distance of the organelle of interest, in our case, lysosomes, from the migrating, gap-exposed edge of the cell. Analysis of lysosomal distribution using MDE revealed that lysosomes are positioned closer to the cell edge in leader cells compared to non-leader marginal cells, indicating a greater peripheral accumulation of lysosomes in leader cells. (Supplementary Fig. 2C, left panel). Next, we quantified the asymmetry of lysosome distribution in collectively migrating MDCK cells at zero- and two-hours post-confinement lift-off. To measure this, we identified the long axis of each cell along the direction of migration (perpendicular to the wound edge) and divided the cell at its midpoint along this axis. We then measured the intensity of the lysosomal marker LAMP1 in the front and rear halves of the cell using FIJI (ImageJ) to assess asymmetrical lysosome distribution. Asymmetry analysis revealed a significant front-rear asymmetry in lysosome distribution in collectively migrating MDCK cells, particularly at two hours post-confinement lift-off, further reinforcing the occurrence of polarized lysosome positioning during migration and leader cell emergence (Supplementary Fig. 2C, right panel). To test the generality of our findings among cultured epithelial monolayer models, we next repeated the measurements in a murine mammary epithelial cell line, EpH4-Ev. Enhanced peripheral localization of lysosomes in leader cells also emerged in this model (Supplementary Fig. 2D, E). Since leader cell emergence and lysosomal accumulation in these cells occurred at a similar time scale, we wondered how these dynamics behaved at the onset of collective migration in epithelial monolayers. Subsequently, we asked whether there might be any correlation between lysosome accumulation at the cell periphery in the marginal cells and collective cell movement. The line plot of two events (Supplementary Fig. 2F) indeed showed a temporal correlation between collective cell movement and POF for lysosomes with different migration time points. As the epithelial cells migrated progressively, we observed an increase in lysosome accumulation starting at T-60 min onwards and peaking at T-120 min post-confinement lift-off.
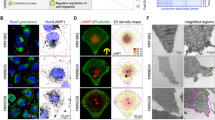
Finally, to understand how the lysosome dynamics are spatially correlated to actin dynamics within leader cells, we performed live-cell imaging of LifeAct-eGFP-expressing MDCK cells having Dextran-647 (Alexa Fluor 647-labeled Dextran) pre-labeled lysosomes (Fig. 2A, Supplementary video 1). Temporal color-coded map shows lysosome localization and initiation of lamellipodia as coordinated events (Fig. 2B). Live cell imaging of LifeAct-eGFP expressing MDCK cells having Dextran-647 labeled lysosomes, revealed differential lysosome dynamics in leaders and immediate non-leader marginal cells (Fig. 2C). Lysosomes in emerging leader cells showed peripheral accumulation followed by dispersal, faster bi-directional movement and subsequently, the lamellipodial protrusions started emerging (Fig. 2C, leader cell). Whereas in non-leader marginal cells, the lysosomes did not move dynamically as the migration progressed (Fig. 2C, marginal cell). Importantly, the live-cell imaging of these cells clearly showed that lamellipodia extension happens in lysosome-rich zones (Fig. 2D, E, and F-top panel, and Supplementary video 2). In contrast, the actin structure was retracted from lysosome-deficient regions (Fig. 2F, bottom panel, Supplementary video 2). The line plot in Fig. 2G shows the correlation of lamellipodia growth and lysosome accumulation. The increase in lamellipodia area and lysosome enrichment is positively correlated (Fig. 2G, Top-panel), while lamellipodia area decreases when there is little to no change in lysosome accumulation (Fig. 2G, Bottom-panel). Taken together, these results revealed that lysosome accumulation at the periphery of the leader cells is a ubiquitous event during collective cell migration across different model systems and is spatiotemporally correlated with the initiation and extension of lamellipodial structures in the leader cells.

A Live imaging snapshots of collectively migrating LifeAct-MDCK cells having Dextran-647 labeled lysosomes after confinement lift-off. Cells were imaged on a confocal microscope, and a zoom-in of a single cell is shown for the ease of understanding. B The dynamics of lysosomes and actin were monitored, and the representative temporal color-coded map of actin and lysosomes shows coordinated redistribution of actin cytoskeleton and lysosomes during collective migration and leader emergence. C Live-imaging snapshots of actin and lysosome (labeled using Dextran-647) dynamics in collectively migrating LifeAct-GFP MDCK cells. The red arrowhead marks an emerging leader cell, and the cyan arrowhead marks a non-leader marginal cell. Kymographs showing differential lysosome dynamics in leader cell (1) and marginal cell (2) marked by red and cyan arrowheads respectively in (C). D Schematic representation of the relation between lysosome localization and lamellipodial dynamics in a leader cell during collective epithelial migration. E Live imaging snapshots of lamellipodial and lysosome dynamics in a leader cell. The cells were allowed to migrate for T-2 h to allow leader cell emergence, followed by live imaging of lysosome dynamics in these cells. T-0 is the starting point of the imaging after T-2 h of confinement lift-off. The green and yellow arrowheads mark lysosome-rich and deficient zones within a leader cell. F Image montage lysosome-rich region (marked by green arrowhead in (E) and lysosome-deficient region (marked by yellow arrowhead in (E) captured as a time series on a confocal microscope. Lamellipodial growth is observed in the lysosome-rich region, whereas limited lamellipodia extension is observed in the lysosome-deficient region. G The correlation between lamellipodia dynamics (growth or no-growth) and lysosome enrichment or deficiency was measured for the leader cell imaged live at T-2 h post confinement lift-off and is represented in the graph here. Scale bar-10 µm. All time points are in minutes. Scale bar-10 µm.
Lysosome transport on microtubule tracks drives its peripheral accumulation
Next, we asked whether the peripheral accumulation of lysosomes in marginal cells, including the leader cells, was a result of increased lysosome biogenesis or of an escalated anterograde trafficking of the organelle. Considering that nuclear localization of Transcription Factor EB (TFEB) is an indicator of enhanced lysosome biogenesis39,40, we examined TFEB localization. However, we did not observe TFEB accumulation in the nuclei of marginal cells at T-120 min post confinement lift-off (Supplementary Fig. 3A). We further quantified the number of lysosomes in the marginal and follower cells, before and during migration and did not observe any significant change (Supplementary Fig. 3B). These results eliminated the possibility of de novo lysosome biogenesis and pointed towards the polarized transport of lysosomes in leader cells. Previous studies have shown that microtubules are the tracks for bidirectional molecular motor-mediated transport of lysosomes41. To gain insight into the organization of microtubules during collective epithelial migration, we performed a gap-closure assay with both MDCK and EpH4-Ev monolayers. We allowed the cells to migrate for 0 and 120 min and then fixed and immunostained them for microtubules with anti-Alpha-tubulin and lysosomes with anti-LAMP1. Microtubules show peripheral accumulation in marginal cells in both MDCK and EpH4-Ev cells (Fig. 3A, Supplementary Fig. 3C). Similar to computing the lysosomal POF, we also computed the POF (peripheral occupancy fraction) of microtubules to measure the microtubule accumulation at the cell periphery. We calculated the POF of microtubules by taking the ratio of the intensity of microtubules in the defined ROI to the total intensity of microtubules in the cell of interest. This analysis revealed a significant increase in POF of microtubules at T-2 h post confinement lift-off, like that was observed for lysosomes (Fig. 3B). We hypothesized that disrupting microtubules would abolish the dynamic peripheral accumulation of lysosomes. We, therefore, disrupted the microtubules by using nocodazole at a concentration of 10 µM and observed its effect on the collective migration of MDCK cells (Supplementary Fig. 3D, middle panel and supplementary video 3). Nocodazole treatment led to the loss of peripherally localized lysosomes (Fig. 3C, middle panel), and at the same time, we observed a reduced emergence of leader cells upon nocodazole treatment (Fig. 3D, Supplementary Fig. 3D, middle panel). Microtubules are dynamic structures and constantly undergo cycles of assembly and disassembly at the plus end42. Taxol (Paclitaxel) stabilizes the growing ends of microtubules, altering their dynamics. Treatment with 1 µM Taxol delayed collective migration of MDCK cells (Supplementary Fig. 3D, bottom panel; Supplementary video 3) and disrupted lysosome accumulation at the cell periphery, instead causing their even distribution throughout the cytoplasm of marginal cells (Fig. 3C, right panel). This disruption was accompanied by a significant reduction in leader cell emergence (Fig. 3D). We quantified POF for lysosomes in control, nocodazole-, and taxol-treated cells and found a significant drop in the peripheral accumulation of lysosomes in the drug-treated cells as compared to the control (Fig. 3E). Finally, we observed a temporally congruent polarization pattern for microtubules and lysosomes in MDCK cells, which were allowed to migrate for 0-, 15-, 30-, 60-, 120-, and 240-min post-confinement lift-off (Supplementary Fig. 3E).

A Representative confocal micrograph of collectively migrating MDCK cells, fixed at T-0 and T-2 h post confinement lift-off and immunostained for lysosomes, tubulin, actin and nucleus. Yellow arrowhead mark leader cell. B Dot-column plot of POF for microtubules and lysosomes, calculated using respective lysosome and tubulin staining intensity at T-0 and T-2 h for all cells at the margin (For T-0 microtubules n = 78, T-2 h microtubules n = 55, T-0 lysosomes n = 55, T-2 h lysosomes n = 74 for 3 independent experiments). Performed a one-way ANOVA test for statistics. C MDCK cells treated with nocodazole and taxol migrated for T-2 h post confinement lift-off, PFA fixed and immunostained for lysosomes (Anti-LAMP2), tubulin (Anti-β tubulin) and actin. The yellow arrowhead in control marks an emerging leader cell. D Dot-column plot of fraction of leader cell emergence in control (n = 29), nocodazole (n = 26) and taxol (n = 29) treated MDCK cells at T-2 h migration post confinement lift-off. Images were acquired for all treatments to cover the complete length of the margins of migrating MDCK cells (n represents the number of ROIs analysed for each treatment, each frame-2 migration fronts and each migration front consists of 65–75 cells per treatment). The statistical significance was calculated using one-way ANOVA. E The dot-column plot of POF-lysosomes in control, nocodazole and taxol-treated cells. Statistical analysis was done using one-way ANOVA (from left to right n = 67, 77, and 97 from 3 independent experiments). F, G Confocal micrographs of MDCK cells transiently transfected with either FLAG-tagged SKIP WT (F) or SKIP WD- > A mutant (G), migrated for T-2 h post confinement lift-off, fixed and immunostained with anti-FLAG tag, anti-LAMP1 and phalloidin to label actin. Yellow arrowheads mark transfected cells. H Dot-column graph showing the POF of lysosomes in cells expressing either FLAG-tagged SKIP WT (n = 25) or SKIP WD- > A (n = 34) mutant, migrated for T-2 h post confinement lift-off. The statistical significance was calculated using Student’s t-test (Unpaired with Welch’s correction). Scale bar-10 µm. **, ***, and **** indicates p-value < 0.01, 0.001, and 0.0001 respectively. All data are mean ± s.e.m.
We next investigated if cellular organelles such as late endosomes, early endosomes, endoplasmic reticulum (ER), and mitochondria, which also use microtubule tracks for their intracellular transport, show peripheral accumulation. We again allowed MDCK cells to migrate for 0- and 120-min post-confinement lift-off, fixed and immunostained for late-endosomes (Anti-Rab7), early endosomes (Anti-EEA1), endoplasmic reticulum (Anti-KDEL), and mitochondria (Anti-Tom20) (Supplementary Fig. 4A–E). We found that none of the other organelles exhibited enhanced peripheral localization compared to lysosomes. A comparison of the POF for all organelles (Supplementary Fig. 4F) revealed that lysosomes showed the highest peripheral enrichment at 120-min post-confinement lift-off. These results indicated that lysosomal accumulation at the periphery is a specific, active process rather than a passive consequence of microtubule re-orientation during collective cell migration.
However, nocodazole and taxol perturb microtubules, affecting multiple cellular processes and complicating the isolation of their specific role in lysosome relocalization during leader cell emergence. To overcome this, we used a strategy that selectively disrupts lysosome transport without altering microtubule structure. Downstream effectors of small GTPase Arl8b control the lysosome transport on the microtubule track. Among these proteins, lysosome adapter protein SKIP recruits the plus-end-directed kinesin motor to the lysosome43. We, therefore, hypothesized that transfecting the cells with a kinesin-binding defective mutant of SKIP (SKIP WD- > 2XA) can specifically disrupt the lysosome dynamics without affecting the microtubule structure. Interestingly, we found reduced peripheral accumulation of lysosomes in cells expressing the mutant SKIP (Fig. 3F, G). The POF of lysosomes SKIP-WT was significantly higher compared to SKIP WD- > 2XA mutant expressing cells (Fig. 3H). Taken together, these results indicate that the peripheral accumulation of lysosomes in marginal cells is the result of a microtubule-mediated dynamic localization of lysosomes.
Peripheral accumulation of lysosomes is necessary for leader cell emergence
Next, we sought to examine whether the peripheral accumulation of lysosomes is necessary for the emergence of leader cells. We subsequently hypothesized that if that were the case, preventing lysosomes from moving to the cell periphery would reduce the propensity of leader cell emergence. Conversely, forcing lysosomes to the cell periphery would increase this propensity. To test these hypotheses, we resorted to abrogating the interaction between kinesin motor, KLC2 and SKIP using the chemical inhibitor kinesore44 at a concentration of 12.5 µM. Lysosomes in MDCK cells treated with kinesore were repositioned to the perinuclear region compared to the control cells (Fig. 4A). The effect on lysosome positioning upon kinesore treatment was similar to the previous report44, thus confirming the inhibitor activity in our system. The POF of lysosomes measured in cells treated with kinesore revealed a significant reduction in the peripheral pool of lysosomes as compared to the control cells (Fig. 4B). The collective movement of the kinesore-treated MDCK cells was delayed, leading to diminished gap closure, and fewer leader cells emerged as compared to the control (Supplementary Fig. 5A, B, Supplementary video 4). Next, we treated mouse embryonic skin with kinesore and subjected these kinesore-treated skins to incision wounds. We then allowed the cells to migrate for 30 min, fixed the samples, and immunostained for lysosomes, actin, and microtubules (Fig. 4C). Here, also lysosome polarization was also lost in kinesore-treated skin wounds as compared to the control. POF of lysosomes measured in kinesore-treated skin wounds revealed a significantly lower accumulation of the organelle at the cell periphery compared to control conditions (Fig. 4D).

A Representative confocal micrograph of control and kinesore treated MDCK cells, migrated for T-2 h post confinement lift-off. Yellow arrowhead marks the leader cell. B Dot-column plot of POF-lysosomes in control (n = 101), and kinesore (n = 53) treated cells (3 independent experiments). Unpaired Student’s t-test with Welch’s correction (two-tailed) performed. C Mouse embryonic skin treated with kinesore subjected to wounding and migration post-incision on an air-liquid interface with the media containing the inhibitor. Yellow arrowheads mark lysosome enrichment in cells at the leading edge. Pink arrowhead marks the absence of lysosome polarization on kinesore treatment. Presence of enriched actin cables observed in kinesore treated mouse embryonic skin wounds at the migration front, indicating a lack of migratory cells. D Dot-column plot of POF-lysosomes in control (n = 144), and kinesore (n = 165) treated mouse embryonic skin wounds (3 independent experiments). Unpaired Student’s t-test with Welch’s correction (two-tailed). E–H Schematic and confocal micrographs of molecular basis and lysosome positioning bias to cell periphery (E, F) and cell center (G, H) using the RAMP system. I Fluorescence/bright-field time-lapse imaging snapshots of MDCK cells stably expressing mCherry-Kif5B*-Strep transiently transfected with LAMP1-SBP-GFP. Yellow arrowheads mark co-expression. J Time-lapse imaging snapshots of MDCK cells expressing Strep-KifC1*-mCherry transiently transfected with LAMP1-SBP-GFP. Yellow arrowheads mark co-expression. K Percentage of leader cells emerging in cells co-expressing mCherry-Kif5B*-Strep or Strep-KifC1*-mCherry and LAMP1-SBP-GFP (n = 12), Tom20-SBP-GFP (n = 28), and VAPA-SBP-GFP (n = 28) migrated for 4h (n - number of ROIs analysed, each ROI has multiple transfected cells, data shown is the average leader cell emergence percent from 3 independent experiments). One-Way ANOVA test. L MDCK cells co-expressing either mCherry-Kif5B*-Strep or Strep-KifC1*-mCherry and LAMP1-SBP-GFP, fixed at T-2 h and stained for focal adhesion protein Paxillin. M LAMP1-SBP-GFP and mCherry-Kif5B*-Strep or Strep-KifC1*-mCherry double-positive cells were imaged live for 24 h and quantified for the fraction of leader cells emerging. Inverted gray-scale images of cells fixed post-migration and stained for actin. n (ROIs) = 20 per condition, 3 independent experiments. Student’s t-test, unpaired, two-tailed. N Percentage of leader emergence in MDCK cells co-expressing KLC2 and SKIP-WT (n = 35) or SKIP WD- > A (n = 32) mutant and KLC2 alone (n = 37) (3 independent experiments). One-Way ANOVA test. Scale Bars-10 µm. Data are mean ± s.e.m.
Although the chemical inhibitor kinesore specifically abolishes lysosome transport on microtubule tracks by disrupting the interaction between KLC2 and SKIP, the effect is not limited to the cells at the migration margin. Hence, to precisely modulate lysosome distribution in cells at the migration margin and monitor its implications, we switched to a method that allowed us to manipulate lysosome localization in individual cells. To this end, we used the Reversible Association of Motor Protein (RAMP) system45. In this system, the interaction between streptavidin bound to motor protein and streptavidin-binding protein (SBP) fused to an organelle-localizing protein drives the preferential localization of the target organelle. To this end, we generated stable cell lines expressing streptavidin-bound to motor protein, mCh-Kif5b*-Strep or Strep-Kifc1*-mCh (Supplementary Fig. 5C, D, mCh: mCherry). mCh-Kif5b*-Strep generates plus-end-directed trafficking, accumulating the target organelle at the cell periphery (Fig. 4E, F, respectively). In contrast, Strep-Kifc1*-mCh generates minus-end-directed trafficking, accumulating the target organelle near the cell nucleus (Fig. 4G, H, respectively). Further, we transiently transfected these cells with LAMP-1-SBP-GFP, targeting lysosomes, and seeded them in the migration chamber. By the interaction between Streptavidin and SBP, lysosomes were either localized to the cell periphery in mch-Kif5b*-Strep-expressing cells or the perinuclear region in Strep-Kifc1*-mCh-expressing cells (Fig. 4F, H). To validate the preferential positioning bias of lysosomes using the RAMP system, we quantified the POF of lysosomes for LAMP1-SBP-GFP positive-cells in either mch-Kif5b*-Strep-expressing or Strep-Kifc1*-mCh-expressing MDCK cells (Supplementary Fig. 5E). Using time-lapse imaging, we then followed the trajectories of LAMP1-SBP-GFP-expressing cells at the migration front (Fig. 4I, J, Supplementary videos 5, 6). Interestingly, LAMP1-SBP-GFP-expressing cells in mCh-Kif5b*-Strep background, which had peripherally localized lysosomes (Fig. 4E, F, I), showed a significantly increased propensity for leader cell formation than LAMP1-SBP-GFP-expressing cells in Strep-Kifc1*-mCh background, which had lysosomes mostly localized to the perinuclear region (Fig. 4G, H, J). Next, we calculated the probability of leader emergence in the two scenarios by dividing the number of co-transfected cells at the margin with lamellipodia (identified as leader cells) by the total number of co-transfected cells at the migrating edge. The probability of cells with peripheral lysosomes becoming leaders was 55% ± 3% (mean ± SD), while the probability of cells with perinuclear lysosomes was 7% ± 2% (Fig. 4K). Relevantly, the baseline probability was 20% ± 5% in mCh-Kif5b*-Strep and 16% ± 2% in Strep-Kifc1*-mCh motor-only controls (Fig. 4K). We also examined whether the positional bias of mitochondria and the endoplasmic reticulum (ER) toward the cell periphery or center influences leader cell emergence during collective epithelial migration. Stable MDCK cell lines expressing mCh-Kif5b*-Strep or Strep-Kifc1*-mCh were transfected with Tom20-GFP-SBP (for mitochondria) or VAPA-GFP-SBP (for ER), and leader cell emergence was quantified (Fig. 4K). Peripheral or perinuclear repositioning of mitochondria and ER had no significant effect on leader emergence compared to Kif-only controls (Fig. 4K), indicating that lysosomal positioning plays a unique role in this process. Moreover, we found that cells co-expressing mCh-Kif5B*-strep and LAMP1-SBP-GFP possessed prominent lamellipodial structures and mature focal adhesions oriented perpendicular to the migrating front (Fig. 4L, top panel). In contrast, cells co-expressing Strep-KifC1*-mCh and LAMP1-SBP-GFP possessed peripheral actin cables and less prominent focal adhesions oriented parallel to the migrating front (Fig. 4L, bottom panel). These results suggested that mCh-Kif5B*-strep expression and subsequent peripheral localization of lysosomes promoted leader-like cell morphology, while mCh-Kifc1*-Strep expression and subsequent perinuclear localization of lysosomes promoted follower-like cell morphology22,23. We also compared the movement of cells transiently expressing either mCh-Kif5B*-strep+LAMP1-SBP-GFP or Strep-Kifc1*-mCh+LAMP1-SBP-GFP and observed no significant difference in cell motility between the two sets (Supplementary Fig. 5F). To further strengthen our findings, we monitored live collective cell migration in cells globally expressing LAMP1-SBP-GFP along with either Strep-Kifc1*-mCh or mCh-Kif5b*-Strep (Fig. 4M, Supplementary Fig. 5G, H). We observed the emergence of more pronounced leader cells in the LAMP1-SBP-GFP and mCh-Kif5b*-Strep condition, where lysosomes were globally biased toward the cell periphery, compared to the LAMP1-SBP-GFP and Strep-Kifc1*-mCh condition, which localized lysosomes toward the cell center (Supplementary Fig. 5G, H). The effect of globally manipulating lysosome distribution on leader cell emergence was quantified as the fraction of leader formation, as shown in Fig. 4M. These analyses together showed that while overall migration distance remains unchanged, cells with peripherally positioned lysosomes are more likely to form lamellipodia and emerge as leaders, whereas those with perinuclear lysosomes migrate within the cohort without extending lamellipodia. Leader emergence is thus a systems-level event22,23, allowing compensation by other cells, but global disruption of lysosome transport, such as with kinesore, impairs this compensatory mechanism, leading to slower wound closure.
Finally, we obtained similar results using EpH4-Ev cells, where cells transiently transfected with mCh-Kif5B*-strep and LAMP1-SBP-GFP or Strep-KifC1*-mCh and LAMP1-SBP-GFP showed lamellipodium or actin cables, respectively (Supplementary Fig. 5I). Noticeably, abrogating lysosomal association with kinesin motor using the kinesin binding defective mutant of SKIP also showed a similar phenotype of reduced lamellipodial formation and hence leader emergence when compared to cells expressing wildtype SKIP (Figs. 3F, G, 4N). In summary, we were able to regulate the propensity of leader cell emergence at the migrating front by altering lysosome localization. Collectively, these results suggested that peripheral localization of lysosomes is both necessary and sufficient for leader cell emergence.
Actomyosin contractility-generated cellular forces drive peripheral lysosome localization
In a cohesively migrating epithelium, leader cells emerge through an active mechanical process, where actomyosin contractility-generated cellular forces play critical roles4,6,9,46. We, therefore, asked whether the peripheral accumulation of lysosomes in leader cells might be driven by actomyosin contractility. To test this possibility, we reduced the actomyosin contractility by using a non-muscle myosin II inhibitor, blebbistatin and a Rho-associated protein kinase inhibitor, Y-27632. We used the concentrations of these drugs22 that are enough to disrupt the cytoskeletal structures without inducing cell toxicity. (Supplementary Fig. 6A, Supplementary video 7). We pre-treated the cells for 30 min with the desired concentration of these inhibitors. We then lifted off the confinement and allowed the cells to migrate in a media containing these inhibitors. Subsequently, we observed a reduced accumulation of lysosomes at the cell periphery upon drug treatment, analyzed by measuring POF for control and inhibitor-treated cells (Fig. 5A, B). In addition, there was no selective emergence of leader cells at the migrating front (Fig.5A). Time-lapse data of cells treated with actomyosin inhibitors revealed that the dynamics and speed of lysosomes were altered upon treatment (Supplementary Fig. 6B, C). Lysosomes in cells treated with actomyosin inhibitors showed enhanced lysosomal tubular extensions (Supplementary Fig. 6B). Together these results show that lysosome dynamics and localization are sensitive to the forces generated by actomyosin contractility. Loss of lysosome polarization upon blebbistatin and Y27632 treatment was corroborated in the ex vivo mouse embryonic skin wound model, which was treated with the inhibitors, analysed using confocal microscopy and lysosome accumulation analysed by measuring the POF (Fig. 5C, D). This set of experiments provided evidence supporting the mechano-sensitivity of lysosome localization during collective cell migration.

A Control (DMSO), Blebbistatin, and Y27632 treated MDCK cells allowed to migrate for 2h, fixed, and stained for lysosomes (anti-LAMP1), tubulin (Anti-β-tubulin) and actin (Alexa-fluorophore conjugated Phalloidin). Yellow arrowhead marks the leader cell. B The column-dot graph represents POF of lysosomes for Control (n = 107), Blebbistatin (n = 107), and Y27632 (n = 95) treated MDCK cells (3 independent experiments). Analyzed using a One-way ANOVA test. C Mouse embryonic skin treated with Blebbistatin and Y27632 were subjected to incision wounds, allowed to migrate for 30 min, followed by PFA fixation and immunostatining. D The column-dot graph represents POF of lysosomes for Control (n = 59), Blebbistatin (n = 46), and Y27632 (n = 71) treated mouse embryonic skin wounds (data from 3 independent experiments)). Analysed using a One-way ANOVA test. E Schematic representation of the forces acting on a cell-pair. F MDCK cells were seeded at a confluency to form cell pairs, followed by incubation with DMSO (control) or blebbistatin overnight, followed by blebbistatin washout for T-4 h. Lysosome distribution was analyzed using confocal microscopy post-fixation and immunostaining with anti-LAMP1, anti-LAMP2 to mark lysosomes, Alexa-fluorophore conjugated phalloidin marks actin, and DAPI stains the nucleus. Pink arrowheads mark the orientation of lysosomal staining. G POF of lysosomes was calculated for control (n = 35), blebbistatin (n = 31) and blebbistatin washout (n = 35) MDCK cells and represented as a dot-column graph (3 independent experiments). Statistical analysis for multiple comparisons was performed using the One-Way ANOVA test. H Schematic representing a micropatterned single beak mold used for defining leader cell-like characteristics in select cells. I Dynamics of dextran-647 labeled lysosomes were observed in LifeAct-GFP MDCK cells seeded in micropatterned chambers. Cells were allowed to migrate for up to 180 min post lifting off the confinement. Yellow arrowheads mark lysosomal accumulation in the leader cell. J Graph representing POF of lysosomes calculated for non-leader marginal (n = 54) and leader cells (n = 45) (data from 3 independent experiments). Scale bar-10 µm; *, **, *** and **** signifies p-value < 0.05, 0.01, <0.001 and <0.0001 respectively. All data are mean ± sem.
To further test the connection between the actomyosin contractility and the peripheral accumulation of lysosomes, we performed the following experiments. Previous studies have shown that increased traction forces and tensile stresses among the follower cells preceded leader cell emergence22. This observation is connected to the fundamental premise of contact inhibition of locomotion (CIL), where one can imagine a leader-follower doublet to be a connected pair of cells experiencing a force perpendicular to the cell-cell contact47,48 (Fig. 5E). Therefore, increased tension in one cell will trigger the formation of lamellipodium in the other cell, away from the first cell (Fig. 5E). Hence, we hypothesized that in such a cell-pair system, lysosomes would localize towards the cell periphery, away from the cell-cell interface. Moreover, this localization would depend on the actomyosin contractility. To test this hypothesis, we plated MDCK cells at a low density to allow them to form cell pairs and subjected them to overnight treatment with blebbistatin to reduce the actomyosin contractility (Fig. 5F). Compared to the control, where lysosomes were polarized away from the cell-cell interface, blebbistatin-treated cells exhibited a dispersed lysosomal distribution throughout the cell body and a reduced POF (Fig. 5F, G). To test whether lysosome polarization could be restored upon the revival of actomyosin contractility, we performed subsequent drug washout experiments. Cells maintained under drug-free conditions for four hours partially regained the lysosome polarization (Fig. 5F, G). These results indicated that the force experienced at the cell-cell interface of a leader-follower pair is crucial for lysosome localization.
We further examined whether locally enhanced contractile cellular forces within an epithelial monolayer might be sufficient to induce lysosome polarization away from the interface. To this end, we used an Opto-GEF-based strategy49 to enhance the actomyosin contractility of a specific cell within the monolayer (Supplementary Fig. 7A). This experiment involved blue light-mediated photo-stimulation of cryptochrome 2 (CRY2)-bound catalytic domain of a RhoA-activator, ARHGEF11. CIBN, one of the binding partners of CRY2, was engineered to localize either at the plasma membrane or mitochondria. Upon photo-stimulation, the CRY2-CIBN association triggered plasma membrane (Supplementary Fig. 7A–C, Supplementary video 8) or mitochondrial localization (Supplementary Fig. 7E, F, Supplementary video 9) of the RhoA-activator. When localized to the plasma membrane, RhoA-activator increased the actomyosin contractility49. We used the mitochondrial localization of the RhoA-activator as the negative control for this experiment.
Here, building on the leader-follower interactions observed during epithelial wound healing, we hypothesized that a highly contractile, photo-stimulated cell would generate pulling forces on its neighboring cells, similar to the tension exerted by contractile follower cells on leader cells in a migrating tissue22,47. This idea stems from the notion that mechanical cues, such as the forces generated by actomyosin contractility, can influence the behavior of neighboring cells22,47. We reasoned that in response to this mechanical tension, lysosomes in the adjacent cells would polarize toward the cell periphery, moving away from the contractile cell. This observation would suggest that lysosome repositioning is a mechanosensitive process, regulated by tension-mediated signaling rather than being a process exclusive to the leading edge of migrating cells (Supplementary Fig. 7A–C). Indeed, upon photo-stimulation, lysosomes in the cells adjacent to the excited cell moved towards the cell periphery, away from the stimulated cell (Supplementary Fig. 7C). However, in the negative control, where RhoA-activator localized to mitochondria, lysosomes in the cells adjacent to the stimulated cell did not undergo any such changes in localization (Supplementary Fig. 7E–G). One limitation of this experiment is that Opto-RhoA GEF activation induces a brief (~ 2 min) and localized contraction in a single cell, which distributes pulling forces across multiple neighbors. As a result, only subtle changes in lysosome positioning are observed, indicating that while mechanical forces can influence lysosome localization, this assay likely underestimates the magnitude of the effect seen during collective migration (Supplementary Fig. 7D, G). Consistent with this interpretation, the modest but reproducible outward shift of lysosomes in neighbors was observed only under membrane-targeted RhoA activation and not under the mitochondrial control, reinforcing that the assay does capture contractility-dependent polarization, albeit at a reduced magnitude. Thus, the opto-RhoA GEF results should be viewed as supportive, lower-bound evidence for force-driven lysosome repositioning rather than a full recapitulation of the robust polarization seen during collective migration. Nevertheless, together with the contractility inhibitor and cell doublet experiments, these results are consistent with the idea that forces transmitted through cell-cell interfaces can contribute to lysosome polarization. Lysosomes may therefore participate in coupling intercellular tension among follower cells to leader cell emergence.
Finally, we hypothesized that if the mechanical forces regulated peripheral lysosome localization during the emergence of leader cells, then any cue that spatially biases leader cell emergence should also spatially bias the peripheral localization of lysosomes. One such cue is wound geometry. Having highly positive-curved beaks at the migration front leads to the emergence of leader cells at the tip of the beak, due to strong force concentration at those points38,50,51. Would such beaks show enhanced peripheral localization of lysosomes? To answer this question, we used a micropatterning approach to fabricate silicone stencils made of polydimethylsiloxane (PDMS). These stencils would create wounds with a single beak to allow one or two cells to have the geometrical cue to become the leader cells (Fig. 5H, Supplementary Fig. 7H). We then seeded cells in these micropatterned chambers, labeled lysosomes using Dextran-647, lifted off the confinement, and performed live-cell imaging. The cells seeded in these micropatterned chambers moved collectively and covered the gap (Supplementary Fig. 7I, Supplementary video 10). Upon lifting the stencil, we indeed observed enhanced lysosome accumulation at the tip of these beaks as compared to their sides (Fig. 5I, J, Supplementary video 11). At the same time, lamellipodial structures emerged out of the beak region while actin fibers ran along both sides (Fig. 5I). These results suggested that the spatial concentration of mechanical force in the epithelial monolayer correlates with the enhanced peripheral localization of lysosomes, supporting the premise of mechanosensitive localization of lysosomes during leader cell emergence. It also strengthened the correlation (Fig. 2E) between peripheral localization of lysosomes and lamellipodia formation through the modification of the actin cytoskeleton.
Rac1 GTPase hitchhikes on lysosomal membranes
We then asked whether the peripheral localization of lysosomes promotes lamellipodia formation by locally modifying the actin cytoskeleton. In migrating epithelial cells, actin dynamics depend on the localization and activation of small Rho GTPases, including Rho, Rac1, and Cdc423,10,23,52. These proteins switch between the GTP-bound active state and the GDP-bound inactive state. To determine whether any Rho family GTPases associate with lysosomes, we performed a screen by transiently transfecting MDCK cells with constitutively active (CA, GTP-locked) and dominant-negative (DN, GDP-locked) forms of RhoA, Cdc42, and Rac1. After allowing migration for two hours post-confinement lift-off, we immunostained lysosomes using anti-LAMP1 and analyzed their association (Supplementary Fig. 8A–G). Interestingly, we observed that Rac1-GTPase associates with lysosomes (Supplementary Fig. 8E, G). Other Rho family GTPases did not show any appreciable colocalization with lysosomes (Supplementary Fig. 8A–D). Also, the wildtype (Rac1-WT) is majorly present on the plasma membrane, and a few punctate structures were observed, while Rac1-CA (Q61L) was primarily plasma membrane-localized, whereas Rac1-DN (T17N) was distributed as punctae, some of which were on lysosomes in both MDCK and EpH4-Ev epithelial cells (Supplementary Fig. 8E–I). Mander’s coefficient analysis between Rac1-WT, Rac1-CA, and Rac1-DN with the lysosomal membrane marker LAMP1 confirmed this localization pattern in both MDCK and EpH4-Ev cells (Supplementary Fig. 8H, I). Next, we examined the localization of endogenous Rac1 with lysosomes in collectively migrating MDCK cells. Endogenous Rac1 was majorly localized to the plasma membrane and a few punctate structures, some of which were LAMP1 positive (Fig. 6A, B). Co-localization of Rac1 and lysosome was recapitulated in our ex vivo mouse embryonic skin wound model system (Fig. 6C, D). To further validate our findings, we performed organelle fractionation from MDCK cells grown to 90% confluency using an OptiPrep density gradient and assessed Rac1 association across different organelle fractions (Fig. 6E). In the 15%–30% gradient, lysosomal signals were detected in fractions 1 and 3–7, with fraction 4 showing a strong lysosomal presence along with a faint mitochondrial contribution. Notably, endogenous Rac1 was predominantly found in the lysosome-enriched fractions 1 and 3, which were exclusively lysosomal, as well as in a few other fractions (Fig. 6E). Taken together, our experiments and data strongly point to the association of Rac1 with lysosomes in collective epithelial cells. To understand the dynamics of Rac1 and lysosome association, we performed live imaging of Dextran-647 labeled lysosomes and GFP-tagged Rac1 wildtype overexpressed in MDCK cells. Time-lapse imaging of a leader-like cell, having protrusive lamellipodial structures, revealed a subpopulation of lysosomes positive for Rac1 (Fig. 6F, supplementary video 12), thus supporting our previous findings.

A MDCK cells allowed to migrate for T-2 h were PFA fixed, immunostained for endogenous RAC1 and lysosomes (Anti-LAMP1), and imaged on an Airy-Scan super resolution microscope. Yellow arrowhead marks endogenous RAC1 co-localized with LAMP1. B Line plot showing the overlap of endogenous RAC1 and lysosomes. C Representative confocal micrograph of mouse embryonic skin wound allowed to migrate for 30 min, PFA fixed and immunostained for endogenous RAC1, lysosomes (Anti-LAMTOR-4) and Actin (Alexa-fluor conjugated phalloidin). Arrowheads mark colocalized RAC1 and LAMTOR-4. D The line plot shows the overlap between endogenous RAC1 and lysosomes in mouse embryonic skin wounds. E Representative immunoblot of organelle fractionated from MDCK cells on an optiprep gradient and probed for lysosomes (Anti-LAMP1), RAC1 (Anti-Rac1), mitochondria (Anti-Tom20), and early endosomes (Anti-EEA1). F1, F3 and F4 are exclusive lysosome fractions and are positive for endogenous RAC1. F Live imaging snapshots of MDCK cells expressing GFP-RAC1WT and Dextran-647 labeled lysosomes. The cells were allowed to migrate for 2 h, followed by live imaging of a cell at the migrating edge on a confocal microscope. The yellow arrowheads mark co-localized GFP-RAC1WT and Dextran-647 labeled lysosomes. G Schematic depicting the expression of mEOS-Rac1 and photoconversion using UV light, leading to a green to red shift in emission, thus allowing tracking of its localization dynamics. H MDCK cells transiently transfected with mEos-Rac1 and dextran-647 labeled lysosomes were allowed to migrate for 60 min before photoconversion to initiate lamellipodia formation. Dynamics of Rac1 puncta formation and trajectory were followed for photoconverted mEos-Rac1. Yellow arrowheads mark emerging Rac1 puncta and their localization to dextran-647 labeled lysosomes. Kymograph representing the colocalization dynamics of lysosomes and photoconverted mEos-Rac1. I–K Representative heat maps of FRET-based Rac1 activity sensor for control, RAC1 inhibitor (NSC), and kinesore treated MDCK cells. L Mean FRET index was calculated for control and kinesore treated cells at T-0 and T-2 h and represented as a scatter-column graph (from left to right n = 59, 47, 44 and 46 from 3 independent experiments). Statistical analysis was performed using the one-way ANOVA test. Scale bar-10 µm; *, ** and **** signify p-value < 0.05, 0.01 and 0.0001, respectively. All data are mean ± s.e.m.
These findings led us to investigate whether lysosomes actively shuttle Rac1 to the plasma membrane or vice versa. To examine the dynamic association between lysosomes and Rac1, we employed a photoconversion strategy. We expressed mEos-Rac1-WT in MDCK cells with lysosomes labeled using Dextran-647. Upon 405 nm illumination, mEos fluorescence shifted from green to red, enabling real-time tracking of Rac1 dynamics. In this experiment, migrating mEos-Rac1-expressing cells were photoconverted near the growing lamellipodial protrusions, and the redistribution of photoconverted Rac1 was monitored (Fig. 6G, H). Live imaging and kymograph analysis revealed punctate structures forming from photoconverted mEos-Rac1. Notably, these puncta frequently merged with nearby lysosomes, showing significant colocalization (Fig. 6H, Supplementary video 13), suggesting an active interaction between Rac1 and lysosomes during migration. We also tracked the photoconverted Rac1-DN (T17N) and Rac1-CA (Q61L) proteins for their redistribution within the cell (Supplementary Fig. 9A–C). As depicted by kymographs, live-tracking revealed puncta formation events for photoconverted mEos-Rac1-DN. Interestingly, these punctate structures merged with lysosomes positioned nearby, showing colocalization between the two (Supplementary Fig. 9A, C, Supplementary video 14). In contrast, photoconverted mEos-Rac1-CA diffused throughout the plasma membrane, and puncta formation was negligible in this case (Supplementary Fig. 9B, C, Supplementary video 15). These results indicate that Rac1 hitchhikes on peripheral lysosomes, which may play an essential role in branched actin formation and lamellipodia growth in an emerging leader cell.
Next, we investigated the influence of lysosome localization on Rac1 activity. Interestingly, imaging actin in control and kinesore-treated mouse embryonic skin wounds already indicated a potential link between lysosome positioning and Rac1 activity. In control samples, actin was distributed in a dynamic network at the leading edge, whereas in kinesore-treated wounds, we observed the formation of a prominent actin bundle (Fig. 4C), a hallmark of low or absent Rac1 activity23,52. This structural change correlated with a clear loss of lysosome polarization at the leading edge in kinesore-treated samples. However, to probe the Rac1 more directly, we measured the Rac1 activity with a FRET-based Rac1 sensor51, which was expressed in ~80-85% of cells due to high transfection efficiency (Supplementary Fig. 9D). We measured Rac1 activity in the presence or absence of kinesore, which induces perinuclear lysosome accumulation. We also used NSC23766 (Rac1 inhibitor)-treated cells as a negative control. As expected, NSC-treated cells showed no significant FRET signal compared to controls, confirming the functionality of the Rac1 activity sensor (Fig. 6J). Next, we analysed the mean FRET index in control and kinesore-treated cells, yielding three key findings. First, at T-0 (quiescent state), Rac1 activity levels were similar between untreated control and kinesore-treated cells, with no significant difference (Fig. 6I, K). Second, there was no significant change in Rac1 activity within kinesore-treated cells between T-0 and T-2 h. Third, at T-2 h, Rac1 activity was significantly lower in kinesore-treated cells compared to controls at the same migration time point (Fig. 6L). Together, these results indicate that peripherally localized lysosomes enhance Rac1 activity, linking force-induced lysosome accumulation at the cell periphery to lamellipodium formation and leader cell emergence during collective migration (Fig. 7). Interestingly, in migrating MDCK monolayers, Rac1+ lysosomes were frequently observed near early endosomes (Supplementary Fig. 9E–G), suggesting potential crosstalk between these compartments during leader cell emergence.

As epithelial cells start migrating collectively, lysosomes in the cells at the wound margin start accumulating at the cell periphery in the direction of migration. Two hours into migration, leader cells start emerging from the cell collective. As some cells emerge as leaders, lysosomes in these cells show significantly higher accumulation of lysosomes to the periphery. This peripheral pool of lysosomes facilitates lamellipodial extension in leader cells (A). Disruption of the peripheral lysosomal pool severely reduces leader cell emergence. The cue for peripheral distribution of lysosomes comes from the mechanical forces experienced by and emerging leader cell. Forces generated at the cell-cell and cell-surface interface are translated to changes in actomyosin contractility within a leader, which guides the peripheral accumulation of lysosomes (B). Lysosomes in leader cells regulate the growth of lamellipodium by shuttling the Rac1 from the plasma membrane, thus regulating its activity, which further regulates the formation of actin cross-links required for lamellipodial extension (C).
Discussion
Ever since Edward Ruth published his pioneering study on epithelial wound closure using the fragments of frog skin53, the ubiquity and importance of leader cells in collective cell migration have become apparent in various physiological contexts1,2,3,11,14. Yet, for more than a century, it has remained intriguing how these special cells, with their characteristic lamellipodial protrusions, emerge from a seemingly homogeneous population. Once the free edge is generated by wounding, cells at the wound margin experience an asymmetry. While cells within the epithelium are encircled by neighboring cells from all sides, cells at the migration front have at least one cell-free edge. Consequently, the geometric, physical, and molecular asymmetry arising at this free edge creates a spontaneous cue for polarized activation of small Rho GTPases, whose activity would otherwise be restricted by cell-cell junctions. In fact, asymmetric spatiotemporal regulation of Rho GTPases, including Rac1, is central to forming actin projections and stress fibers during collective cell migration, especially in the emergence of leader cells3. However, this asymmetry is equally likely to generate lamellipodial structures in all cells at the migration front, and it is not obvious at what level the selection of a leader cell is decided. Previous studies have shown that leader cell emergence is not an entirely stochastic biochemical event, as conventionally believed. Instead, the localization of highly tensile peaks in the stress landscape determines the precise location of the leaders. These studies have also suggested that enhanced tensions at the cell-cell junction promote the formation of lamellipodial protrusions away from the leader-follower interface, towards the cell-free space, in accordance with the fundamental principles of contact inhibition of locomotion47,48 (Supplementary Fig. 7A). However, the connection between this biophysical driving force and the eventual activation of Rac1 leading to lamellipodia formation remained elusive. It was not clear how these biophysical and biochemical paradigms could be unified under a single mechanistic framework. This study identifies lysosomes as an intracellular platform linking mechanical and biochemical signals towards the regulation of leader cell emergence. At one end, lysosomes respond to changes in the force landscape of the emerging leaders by localizing to the cell periphery (Fig. 7). On the other, they transiently colocalize with the small GTPase Rac1. This specific lysosome-Rac1 interaction supports lamellipodia formation by locally regulating Rac1 activity close to the plasma membrane (Fig. 6). Thus, lysosomes sense the local force field and regulate lamellipodia formation by modulating the Rac1 signaling. This forms a positive feedback loop where cell-cell forces determine a leader cell, in which the lysosome polarization enhancement leads to the regulation of Rac1 signaling in these emerging leaders.
Previous studies have shown that the position of lysosomes is critical for their function30. In this study, using the RAMP system, SKIP mutants, and kinesore, we precisely controlled lysosome localization to investigate its role in Rac1 activation and leader cell emergence. The RAMP system enabled targeted lysosome repositioning in individual cells, showing that peripheral lysosomes promote Rac1 activation and lamellipodia formation, while perinuclear lysosomes fail to do so. SKIP mutants disrupted kinesin-dependent lysosome transport, reducing peripheral accumulation and impairing leader cell formation without affecting microtubules. In contrast, kinesore, which globally inhibits SKIP-KLC interactions, prevented lysosome redistribution across all cells, suppressing Rac1 activation and slowing wound closure. These findings together establish lysosome positioning as a key determinant of leader cell selection. They also reveal that the leader cells emergence functions as a systems-level process, allowing collective migration to continue despite individual failures in leader cell formation. However, when lysosome transport is globally disrupted by kinesore, this compensatory mechanism is impaired across the entire migrating sheet, resulting in fewer leader cells and consequently slower wound closure. Relevantly, a major advance of our study is the ability to manipulate lysosome positioning in an ex vivo mouse embryonic skin wound model, providing direct evidence of how lysosome spatial organization impacts actin cytoskeletal remodeling in a physiologically relevant tissue. By pharmacologically perturbing lysosome positioning, we demonstrated that peripheral lysosome localization correlates with actin-rich protrusions at the leading edge, reinforcing the idea that lysosomes play a direct role in cytoskeletal reorganization. These findings suggest that lysosomal positioning is not only crucial for leader cell emergence in a living tissue context, where mechanical and biochemical cues dynamically regulate migration. This approach strengthens the physiological relevance of lysosome-driven leader cell selection. A limitation of our study is that we have not quantified the minimal proportion of cells that must regulate lysosome positioning for collective migration to proceed efficiently. This remains challenging because leader cells are highly dynamic and interchangeable, continuously emerging and being replaced during migration, making it difficult to define a fixed subset responsible for sustaining collective movement.
Related to lysosome-Rac1 interaction, it is important to note that Rac1 molecules organize into nanoclusters at the plasma membrane, form a gradient of spatial signaling and stay active for a few minutes in migrating cells54,55, recruiting actin remodeling proteins such as Arp2/3 and WAVE. Here, our findings demonstrate that lysosomes contribute to Rac1 spatial regulation during leader cell emergence, with Rac1-GDP preferentially associating with lysosomes and being transported away from the leading edge. They suggest a potential role for lysosomes in redistributing Rac1 to specific intracellular locations, influencing where activation may occur. However, our data do not support lysosomes as direct sites of Rac1 activation. Instead, a previous work56 has shown that early endosomes act as sites for Rac1 activation and transport activated Rac1 to the plasma membrane during single-cell migration. This raises the possibility of a two-step Rac1 activation model, in which lysosomes serve as transient carriers of Rac1-GDP, while early endosomes facilitate its reactivation. While our study provides strong evidence for lysosome-mediated Rac1 redistribution, further investigation is needed to determine whether lysosome-endosome interactions play a role in Rac1 activation or if additional regulatory mechanisms contribute to this process. Hence, it might be crucial to explore the crosstalk between lysosomes and early endosomes to further characterize the signaling pathways underlying leader cell emergence during epithelial cell migration10. Relevantly, we observed Rac1+ lysosomes juxta-positioned with early endosomes in migrating MDCK cells (Supplementary Fig. 9E–G). Taken together, despite the aforementioned limitations, these findings highlight the importance of intracellular organelle positioning in controlling signaling pathways during collective migration and open new avenues for exploring the spatial regulation of Rho GTPases in dynamic cell behaviors.
Finally, the coherent observation of lysosome accumulation in the leader cells of mammalian epithelial monolayer, ex vivo mouse embryonic skin and Drosophila epidermis indicates that the process we discovered could be evolutionarily conserved in epithelial wound healing. However, it remains unknown whether lysosome-Rac1 interaction is also critical for other kinds of single and collective cell migrations4, including neural crest cell migration in vertebrates4, border cell migration in Drosophila10, and collective migration of lateral line primordial cells in Zebrafish57. In addition, given that lysosomes play an important role in the continuous cross-layer migration and differentiation of keratinocytes within a mammalian epidermis58, the lysosome-Rac1 signaling axis might be critical for the development and maintenance of the multi-layered epidermal epithelium. Collectively, the results of our experiments and their implications place the functioning of lysosomes at a crucial junction of mechanochemical signals during collective cell migration and expand the scope of this organelle beyond its conventional role in the catabolism of cellular materials.
Methods
Cell culture
Madin-Darby canine kidney (MDCK) and EpH4 (ATCC; CRL-3063) epithelial cells were used in this study. Tetracycline-resistant wild-type MDCK (MDCK-WT) cell line was a gift from Yasuyuki Fujita. MDCK cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with GlutaMax (Gibco) with 5% fetal bovine serum (tetracycline-free FBS, Takara Bio) and 10 U mL−1 penicillin and 10 μg mL−1 streptomycin (Pen-Strep, Invitrogen) in an incubator maintained at 37 °C and 5% CO2, unless mentioned otherwise. EpH4-Ev cells (ATCC; CRL-3063) were cultured in Dulbecco’s modified Eagle’s medium supplemented with GlutaMax (Gibco) with 10% fetal bovine serum (tetracycline-free FBS, Takara Bio) and 1.2 μg ml−1 puromycin (Gibco) in an incubator maintained at 37 °C and 5% CO2.
MDCK cells stably expressing mCh-Kif5b*-Strep, Strep-KifC1*-mCh (mCh: mCherry), cells were transfected with the respective plasmids using Lipofectamine 2000 (Invitrogen) following the instructions provided by the manufacturer. 16–18 h- post-transfection, cells were seeded via serial dilution in a 96-well plate, such that each well had a single cell. Forty-eight hours post-cell seeding, the cells were subjected to selection media (DMEM-GlutaMax plus 5% FBS) containing 400 μg mL−1 of Geneticin (Invitrogen). The growth of single-cell-derived colonies was monitored over two weeks, following fluorescence confirmation. Colonies with a homogenous expression of the fluorescence-tagged protein of interest were expanded and further maintained in 100 μg mL−1 Genetic containing media. All transient transfections were done using Lipofectamine 2000 following the instructions provided by the manufacturer. To obtain LAMP1-SBP-GFP and mCh-Kif5b*-Strep or Strep-KifC1*-mCh double-positive cells, LAMP1-SBP-GFP was nucleofected using the SE-Cell Line 4D-Nucleofector™ X Kit (V4XC-1024, LONZA), followed by selection for GFP-positive cells. mCh-Kif5b*-Strep or Strep-KifC1*-mCh was then expressed in the LAMP1-SBP-GFP–expressing cells, and the cells were sorted by FACS for GFP- and mCh- double positive populations. The resulting double-positive cells were seeded into iBidi migration chambers to form a monolayer, followed by live imaging to monitor their collective motion.
Cell migration assays were performed using adhesive biocompatible silicone culture inserts from iBidi. 5 × 104 cells suspended in an 80 μL cell culture medium were seeded in a culture insert stuck to glass-bottom dishes and incubated at 37 °C in a 5% CO2 humidified chamber. After 18 h of cell seeding, the cells formed a monolayer, and the culture insert was lifted to allow the cells to migrate. All migration experiments were carried out at 37 °C and in a 5% CO2 humidified chamber (standalone or stage-top incubator).
Drosophila melanogaster embryo microinjection wounding assay
For fixation and immunostaining, a previously described protocol was followed59. Briefly, Stage 15 Drosophila melanogaster embryos were collected on an apple juice plate. These embryos were then aligned and stuck to a double-sided tape on a glass slide. They were left at room temperature for 5 min, followed by halocarbon oil application and subsequent wounding using glass needles mounted on an Eppendorf Femtojet system. At 15 min post-wounding, halocarbon oil was drained from the slide and flushed with heptane to release the embryos from the double-sided tape. Next, 0.8 mL 4% Formaldehyde (in PBS) was added to a fresh tube, and embryos were transferred with heptane to this tube, which was then placed on rotation in an orbital shaker for 25 min. The aqueous solution was then removed from the bottom of the tube, and subsequently, heptane was removed too. To devitellinize the embryos, heptane was rapidly added, followed by adding double to triple the volume of 90% ethanol and mixing it vigorously by vortexing. We then proceeded with embryos that settled down in the tube. The embryos were given 5 quick washes with 1X PBS and then blocked in PBS + 0.3%Triton-X100 + 0.5%BSA (PBTB) for 30 min at room temperature. A primary antibody against LAMP (Abcam) was prepared in PBTB. Embryos were incubated overnight at 4 °C, followed by three washes done in PBS + 0.3%TritonX-100 (PBT). Secondary antibody was prepared in PBTB and incubated for 2 h at room temperature, followed by 5 washes with PBT. DAPI and Phalloidin staining were done along with the secondary antibody. The embryos were mounted in Fluoromount (Sigma) on a glass slide and imaged using confocal microscopy.
Mouse embryonic skin wounding
C57/6 J strain mice, 8–12-week-old adult females, were used in the study for all experiments involving mice. No statistical methods were used to predetermine sample size. The results are independent of the sex of the animals. The development and physiology of the embryonic epidermal tissue is not known to be different in different sexes; therefore, sex of the animal was not considered a parameter in the study. The protocol was approved by the Institutional Animal Ethics Committee, TIFR Hyderabad, on behalf of the Committee for the Purpose of Control and Supervision of experiments on Animals, India.
For mouse embryonic wounding assays, E14.5 - E16.5 mouse embryos were dissected out. The dorsal skin from these embryos was then excised. The skin was placed on a filter paper (Whatman filter, 8 μm pore size) with the epidermal side up. Using a scalpel/fine scissor, incision wounds were made on the epidermis. The filter paper, along with the tissue, was kept floating in a 6-well plate containing 2 mL of DMEM with 5% fetal bovine serum (FBS) and 1% PenStrep. After the desired incubation period, the tissue was fixed in 4% paraformaldehyde (PFA) overnight at 4 °C. The tissue was washed in PBS and transferred to 90% ethanol in PBST (0.2% Triton X-100 in 1X PBS) overnight at −20 °C. The tissue was then rehydrated using graded ethanol (75%, 50% and 25% in PBST). After two more washes in PBST, the tissue was blocked using 10% normal goat serum (NGS) in PBST for 2 h. Primary antibodies were added and incubated overnight at 4 °C. This was followed by subsequent PBST washes, after which secondary antibodies were added and incubated for 2 h at room temperature. After washing, the tissue was mounted on a slide using Fluoroshield (Sigma Aldrich) mounting media. The slide was cured overnight and imaged using an upright confocal (Leica Stellaris). For actomyosin contractility inhibition in mouse embryonic skin wounds, 50 μM of Blebbistatin (Sigma) and 30 μM Y27632 (Sigma) were used. SKIP-Kinesin1 interaction inhibitor kinesore (Torcis) was used at a final concentration of 50 μM in the ex vivo mouse embryonic skin wounds. The concentration of inhibitors used for this model system is higher compared to their use for cell monolayers because of the extremely tight epithelial in the tissue, leading to no effect of these compounds at a lower concentration. 3-D renders were created using Imaris (Version 9.2.1).
Immunofluorescence
Cells were fixed in 4% formaldehyde diluted in 1X PBS for 15 min at room temperature. For most antibody staining, cells were permeabilized with 0.25% TritionX-100 in PBS for 10 min at room temperature, followed by three quick washes with 1X PBS to remove the detergent. Cells were treated with blocking/staining solution (0.1% TritonX-100in 1X PBS + 2% BSA) for 1 h at room temperature, washed thrice with 1X PBS, and further incubated with primary antibody prepared in blocking solution for 2 h at room temperature. Post-primary antibody incubation, cells were washed thrice with 1X PBS and incubated with secondary antibody, DAPI, and Phalloidin dilutions prepared in the blocking/staining solution for 1 h at room temperature. Finally, the samples were given 2 quick and 5 min wash with 1X PBS before proceeding for confocal microscopy.
For staining LAMP1 (Abcam), cells were incubated with a blocking solution (0.2% Saponin+5% BSA in 1X PBS) for 30 min at room temperature, followed by three washes with 1X PBS. Cells were incubated with the primary antibody in staining solution (PBS + 0.2%Saponin) for 2 h at room temperature, washed thrice with 1X PBS and further incubated for 1 h with secondary antibodies, DAPI (4′,6-diamidino-2-phenylindole, 1 μg mL−1 in PBS) and Phalloidin made in staining solution. Cells were washed thrice with 1X PBS and imaged using confocal microscopy.
Lysosome labeling for live cell imaging
Lysosomes of MDCK cells for live-cell imaging experiments were labeled with Alexa-fluor conjugated 10,000 MW Dextran (Dextran-647 from Invitrogen). Briefly, cells were incubated with Dextran-647 at a final concentration of 0.25 mg ml−1 in complete media for 6 h at 37 °C in a 5% CO2 humidified chamber. Media for cells was then replaced with complete media and incubated for another 12 h to completely chase Dextran-647 to lysosomes.
Confocal microscopy
Fluorescence images were acquired using a 60X oil objective (PlanApo N 60x Oil, N.A = 1.42, Olympus) and 100X oil objective (UPlanSApo, 100X/1.40 oil) mounted on an Olympus IX83 inverted microscope equipped with a scanning laser confocal head (Olympus FV3000). Time-lapse images of live samples were taken in the live-cell chamber provided with the microscopy setup.
Photoconversion studies were done using MDCK cells that had been transiently transfected with mEos-Rac1 WT, DN or mEos-Rac1 CA. Transiently transfected cells were seeded in culture inserts and proceeded for lysosome labeling using Dextran-647 as described above. Cells were allowed to migrate for 60 min to allow for lamellipodia formation in migrating cells. Stimulation was done on a point region of interest juxtaposed to the site of growing lamellipodia using a 405 nm laser at 2% intensity, looped over 25 times with a scan speed of 1000 μs/pixel. This was followed by LSM imaging of the green and red channels, and Rac1 dynamics were tracked for 5 min.
The photoactivation experiment was carried out using a 60X oil objective (PlanApo N 60x Oil, NA = 1.42, Olympus) mounted on an Olympus IX83 inverted microscope equipped with a scanning laser confocal head (Olympus FV3000), supported with a live-cell imaging setup. 20 mM HEPES (Gibco) was used to maintain CO2 levels. MDCK cells expressing optoGEF-RhoA and CIBN-GFP-CAAX or optoGEF-RhoA and Mito-CIBN-GFP (control)49, stained with lysosomal marker Dextran-647, were used for the experiment. Pulse activation (20 iterations) was performed in the selected rectangular ROIs (whole cell or cell-cell junction) using a 488 nm laser with 2.0% laser intensity at a 4.0 μsec/pixel scan rate. This step was immediately followed by time-lapse (5 min, 10-s intervals) LSM imaging in 561 nm and 647 nm channels, to visualize RhoA activation and monitor lysosomal movement, respectively. Endogenous Rac1 and lysosome localization images were acquired using a 100X oil objective (N.A. = 1.45) Zeiss Airy-Scan microscope (LSM 900) on SR mode, and fluorescence images of leader cells in mouse embryonic skin wounds were acquired using a Nikon inverted research microscope, Eclipse Ti2-E with Yokogawa CSU-W1 SoRa unit.
Förster resonance energy transfer (FRET)-based Rac1-activity measurements
FRET experiments for the Rac1 activity sensor were carried out in the live-cell confocal setup (Olympus FV3000). MDCK cells were plated in a 24-well plate and transiently transfected with Rac1 sensor plasmids52. After 6 h, cells were trypsinized and seeded in migration chambers and left overnight to settle and form confluent monolayers in the chamber. Cells were simultaneously labeled for lysosomes using Dextran-647. Control and AR treatment were given to cells as described in the Global perturbation of lysosome positioning using Ringer’s solution method section. The images were acquired for control and AR-treated cells just after lifting off the confinement (T-0) and post 2 h of migration (T-2 h). The exposure times for donor, acceptor, and FRET channels were kept constant. Images of Dextran-647-labeled lysosomes were also acquired. Each field yielded three 800 × 800-pixel images representing donor, acceptor, and FRET channels. Images were analysed using custom software written in MATLAB (Mathworks), described previously60.
Inhibition studies
For all inhibition studies, cells were pre-treated with the desired concentration of the inhibitor in opti-MEM for 30 min at 37 °C in a 5% CO2 humidified incubator, before lifting the confinement/culture insert. During migration, cells were kept in complete media containing the inhibitor at a given concentration for the required time. Actomyosin contractility was altered using Blebbistatin (Sigma), a myosin inhibitor and Y27632 (Sigma), a ROCK inhibitor, at concentrations of 5 μM and 10 μM, respectively, for MDCK cells. For microtubule disruption in MDCK cells, Nocodazole (Sigma) was used at 10 μM, and Taxol/Paclitaxel (Sigma) was used at 1 μM. For SKIP-Kinesin-1 interaction inhibition, Kinesore (Torcis) was used at 12.5 μM as recommended in the previous study44. Kinesore inhibition in MDCK cells was performed in OptiMEM for the complete duration of the treatment, and the control with these experiments were also kept in OptiMEM to maintain similar conditions. Cells were fixed and immunostained for the desired antibodies post-inhibitor treatment. For inhibitor studies in ex vivo mouse embryonic skin wounds, 50 μM of Blebbistatin (Sigma), 30 μM Y27632 (Sigma) were used, and SKIP-Kinesin1 interaction inhibitor kinesore (Torcis) was used at a final concentration of 50 μM. The concentration of inhibitors used for this model system is higher compared to their use for cell monolayers because of the extremely tight epithelial tissue, leading to no effect of these compounds at a lower concentration. Ex vivo mouse embryonic skin wounds were incubated with inhibitor-containing media for 30 min, fixed and immunostained.
Lysosome enrichment using density gradient separation
To perform lysosome enrichment, subcellular fractionation was carried out using the Lysosome Enrichment Kit (Thermo Scientific), using the manufacturer’s protocol. Briefly, the MDCK cells were grown to 90% confluency, followed by collecting cells on ice by cell scraping. The cell pellet was washed in ice-cold PBS, resuspended in Buffer-A, provided in the kit and homogenized with a dounce homogenizer on ice (~ 25 strokes). To confirm cell lysis, microscopic examination of the homogenate was done by adding 0.5% trypan blue dye. The homogenate was mixed with an equal volume of Buffer-B and subjected to centrifugation at 500 × g for 10 min at 4 °C, and post-nuclear supernatant (PNS) was diluted in OptiPrep gradient buffer to a final concentration of 15% OptiPrep. The sample was then carefully overlayered on the top of a discontinuous density gradient (17, 20, 23, 27, and 30%). The gradient was subjected to ultracentrifugation at 145,000 × g in a swinging bucket rotor (Beckman Colter) for 2 h at 4 °C. After the spin, ten fractions of 800 µl each were collected from top to bottom. The fractions were spun again at 18,000 × g for 20 min at 4 °C, and the resulting pellet was suspended in 4X SDS-sample buffer, boiled for 10 min, and analyzed by SDS-PAGE and immunoblotting.
Antibodies and plasmids
Source and dilution information for all primary and secondary antibodies used in immunofluorescence staining are given in Supplementary Table 1. Details of plasmids used in this study are listed in Supplementary Table 2 with their source.
Micropatterning
PDMS stencils with pillars of defined shapes were created using soft photolithography. Briefly, 150 um thick SU-8 2075 (Kayaku advanced materials, Y111074) was coated on a silicon wafer, followed by soft baking at 65 °C for 5 min and 95 °C for 20 min. SU-8 was exposed in the desired pattern using the MicroWriter ML 3 Pro (Durham Optics Magneto Ltd), followed by post-exposure baking at 65 °C for 7 min and 95 °C for 15 min. The unexposed photoresist was removed by immersing the wafers in the SU-8 developer (Kayaku advanced materials, Y020100) for 8–10 min. Patterned SU-8 wafers were then hard-baked at 170 °C for 45 min. PDMS (Sylgard 184, Dow Corning) was mixed in a ratio of 1:10, degassed, and poured over the patterned wafers. After curing at 85 °C for 2 h, stencils with pillars were peeled off. The stencils were then treated with 2% Pluronic acid for 2 h followed by 15 min of 1X PBS wash. PDMS stencil and a petri dish were plasma cleaned for 10 s, and the stencil was then placed upon the dish. Cells were seeded around the pillars, and after confluence, the stencils were removed, allowing the cells to migrate into the gaps.
Image analysis
All image analysis for this study was performed using Fiji. All images acquired as Z-stacks were converted to a single Z-projected ‘Sum of slices’ image before proceeding with analysis. Peripheral occupancy Fraction (POF) was measured for cells at the margin of a migrating collective in our two-dimensional cell culture and mouse embryonic skin wound models. A leader cell was selected as the cell that is in the front of a migrating monolayer of cells, having lamellipodial extensions. The ROI box with a fixed width of 2.5 μM from the cell edge facing the gap was drawn over the cell of interest. The box was duplicated for the lysosome channel, and lysosome marker intensity was measured. For each cell, the total lysosomal intensity was also calculated by manually marking the cell of interest. For calculating the POF, the lysosomal intensity in the ROI was divided by the total intensity of lysosomes in that cell. 20 marginal and ~10 leader cells per experiment were quantified from three independent experiments. For calculating the junctional occupancy fraction for lysosomes, we measured the lysosomal marker intensity in the cell-cell junctions of the leader cells and non-leader marginal cells by drawing an ROI of 2.5 μM from the junction, followed by dividing it by the total lysosomal intensity in the cell. For measuring the POF for different organelles, we followed the same approach.
To measure Mean Distribution from Edge (MDE), we used a custom-written MATLAB code to quantify the spatial distribution of lysosomes relative to the cell edge in migrating MDCK cells. Briefly, Z-projected sum-of-slices images were first processed to generate a separate cell boundary image from the actin cytoskeleton channel, which was then fed into MATLAB along with the lysosomal marker (LAMP1) image. The code detects the cell edge, allowing for precise spatial mapping of lysosome distribution. To compute MDE, the algorithm determines the center of mass (CoM) of lysosome fluorescence intensity within each cell, representing the weighted centroid of lysosome distribution, where pixel intensities contribute proportionally to their respective positions. The Euclidean distance between the CoM of lysosomal intensity and the detected cell edge is then calculated. A lower MDE value indicates greater lysosome enrichment at the periphery, while a higher MDE value reflects a more centrally distributed lysosome population. For each experiment, 20 marginal cells and approximately 10 leader cells were analyzed, with data collected from three independent experiments. This approach provides an unbiased, quantitative measure of lysosome positioning relative to the migration front, allowing for precise assessment of lysosome polarization dynamics.
Asymmetry of lysosome distribution in collectively migrating MDCK cells was measured at T-0 and T-2 h post-confinement lift-off. Specifically, we first identified the long axis of each cell along the direction of migration (perpendicular to the wound edge) and divided the cell at its midpoint along this axis, creating two distinct regions: the front half facing the migration direction, and the rear half positioned away from the leading edge. To quantify lysosome distribution, we measured the fluorescence intensity of the lysosomal marker LAMP1 in each half using ImageJ, followed by whole-cell LAMP1 intensity measurements. To compute the occupancy fraction for lysosomes at the front and rear ends of the cells, the intensity in each ROI was divided by the total LAMP1 intensity per cell. N = 3, with 15–20 cells per experiment analyzed for each time point. Statistical significance was determined using a one-way ANOVA test in GraphPad Prism.
To measure POF for experiments involving OptoGEF-RhoA, cells adjoining the photoactivated cell were marked manually using the DIC channel to mark cell boundaries. The image was duplicated for the lysosome channel. Lysosome intensity was determined using Fiji. This was followed by marking the cell periphery of the same cell by using the Edit → Selection → Enlarge tool of Fiji to reduce the outline by 2.5 μm. Using the Edit → Clear tool, the lysosomes in the perinuclear region were omitted, and the intensity of remaining lysosomes in the cell periphery was determined. To get the POF for cells, the intensity of lysosomes was divided by the total cell intensity of lysosomes for each cell. POF was determined for pre-activation and post-activation states. Change in POF was compared between pre- and post-activation for the same cell, and 60 cells were analysed per treatment from three independent experiments.
To measure leader emergence in experiments using the RAMP plasmid system, we quantified the total number of cells at the wound margin expressing both the motor (mCh-Kif5b*-Strep or Strep-KifC1*-mCh) and the cargo protein of interest (LAMP-, Tom20-, or VAPA-SBP-GFP), as well as the number of these transiently transfected cells that formed leader cells. The fraction of leader emergence was calculated by dividing the number of leader-forming cells by the total number of transfected cells at the wound margin. The method was used for measuring leader emergence fraction in cells expressing the KLC motor with SKIP-WT or SKIP WD- > 2XA mutant. For inhibitor treatments, the leader emergence was measured by dividing the number of leaders by the total number of cells facing the wound gap. Colocalization analysis of different forms of small GTPase Rac1 with lysosomal marker LAMP1 was performed using the JACoP tool of Fiji. For measuring Mander’s coefficient, cytoplasmic expression was analysed by marking the inside of the cell boundary using a freehand tool in Fiji. The cytoplasmic Rac1 expression includes its vesicular component, and its co-localization with lysosomes was measured.
Tracking of cells during collective migration was done using the Manual Tracking tool of Fiji. The displacement in X and Y was determined from the manual tracking approach for tracking cells with biased lysosome positioning (Cell lines expressing mch-Kif5b*-Strep, Strep-KifC1*-mCh co-transfected with LAMP1-SBP-GFP plasmids). To measure lysosome speed from time-lapse videos, the TrackMate tool of Fiji was used with the following parameters: Vesicle diameter-1 μm; Detector-DoG; Initial thresholding-none; Tracker-Simple LAP tracker; Linking max distance-2 μm; Gap-closing max distance-2 μm, Gap-closing max frame gap-2; Filters-none. The gap closure was measured by marking the gap area manually in Fiji at T-0, 4, 8, 12 and 16 h.
Statistical analysis
Statistical analyses were carried out in GraphPad Prism 9. Statistical significance was calculated by Unpaired t-test with Welch’s correction or One-way ANOVA as mentioned in the corresponding figure. Scatter-bar plots were displayed as mean ± s.e.m. P-values greater than 0.05 were considered to be statistically not significant. No statistical methods were used to set the sample size. Quantification was done using data from at least three independent biological replicates. For analysis involving live-imaging experiments, data were collected from three independent experiments. All the experiments with representative images were repeated at least thrice. Data distribution was assumed to be normal, but this was not formally tested. No statistical method was used to predetermine sample size, but our sample sizes are consistent with the standard of the field and are appropriate for the types of analyses performed. Samples were not allocated using randomization, as this study involved observational analysis of epithelial cells under defined experimental conditions. All groups were subjected to identical culture, treatment and imaging protocols to control for potential covariates, ensuring comparability across conditions. Data collection and analysis were not performed blind to the conditions of the experiments. No data points were excluded from the analyses.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data supporting the findings of this work are available within this article and its Supplementary Information or the Source data provided with this paper. Other data supporting the findings of this study, such as raw and processed microscopy images, which are large-sized files, are available from the corresponding author on request. For FRET analysis, we used MATLAB code described in an earlier publication (https://doi.org/10.1038/s41467-021-27896-z). The MDE MATLAB code has been made publicly available on GitHub (https://github.com/tamaltifrh/Mean-Distribution-from-Edge---MDE). Source data are provided with this paper.
References
Vilchez Mercedes, S. A. et al. Decoding leader cells in collective cancer invasion. Nat. Rev. Cancer 21, 592–604 (2021).
Kozyrska, K. et al. p53 directs leader cell behavior, migration, and clearance during epithelial repair. Science 375, eabl8876 (2022).
Mayor, R. & Etienne-Manneville, S. The front and rear of collective cell migration. Nat. Rev. Mol. Cell Biol. 17, 97–109 (2016).
Shellard, A. & Mayor, R. Collective durotaxis along a self-generated stiffness gradient in vivo. Nature 600, 690–694 (2021).
Pérez-González, C. et al. Mechanical compartmentalization of the intestinal organoid enables crypt folding and collective cell migration. Nat. Cell Biol. 23, 745–757 (2021).
Boocock, D., Hino, N., Ruzickova, N., Hirashima, T. & Hannezo, E. Theory of mechanochemical patterning and optimal migration in cell monolayers. Nat. Phys. 17, 267–274 (2021).
Barriga, E. H., Franze, K., Charras, G. & Mayor, R. Tissue stiffening coordinates morphogenesis by triggering collective cell migration in vivo. Nature 554, 523–527 (2018).
Das, T. et al. A molecular mechanotransduction pathway regulates collective migration of epithelial cells. Nat. Cell Biol. 17, 276–287 (2015).
Brugués, A. et al. Forces driving epithelial wound healing. Nat. Phys. 10, 683–690 (2014).
Ramel, D., Wang, X., Laflamme, C., Montell, D. J. & Emery, G. Rab11 regulates cell-cell communication during collective cell movements. Nat. Cell Biol. 15, 317–324 (2013).
Cheung, K. J., Gabrielson, E., Werb, Z. & Ewald, A. J. Collective invasion in breast cancer requires a conserved basal epithelial program. Cell 155, 1639–1651 (2013).
Tambe, D. T. et al. Collective cell guidance by cooperative intercellular forces. Nat. Mater. 10, 469–475 (2011).
Hayer, A. et al. Engulfed cadherin fingers are polarized junctional structures between collectively migrating endothelial cells. Nat. Cell Biol. 18, 1311–1323 (2016).
Rørth, P. Fellow travellers: emergent properties of collective cell migration. EMBO Rep. 13, 984–991 (2012).
Haeger, A., Wolf, K., Zegers, M. M. & Friedl, P. Collective cell migration: guidance principles and hierarchies. Trends Cell Biol. 25, 556–566 (2015).
Montell, D. J., Yoon, W. H. & Starz-Gaiano, M. Group choreography: mechanisms orchestrating the collective movement of border cells. Nat. Rev. Mol. Cell Biol. 13, 631–645 (2012).
Vishwakarma, M., Spatz, J. P. & Das, T. Mechanobiology of leader-follower dynamics in epithelial cell migration. Curr. Opin. Cell Biol. 66, 97–103 (2020).
Atsuta, Y. & Takahashi, Y. FGF8 coordinates tissue elongation and cell epithelialization during early kidney tubulogenesis. Development 142, 2329–2337 (2015).
Friedl, P. & Gilmour, D. Collective cell migration in morphogenesis, regeneration and cancer. Nat. Rev. Mol. Cell Biol. 10, 445–457 (2009).
Labernadie, A. et al. A mechanically active heterotypic E-cadherin/N-cadherin adhesion enables fibroblasts to drive cancer cell invasion. Nat. Cell Biol. 19, 224–237 (2017).
Ilina, O. et al. Cell-cell adhesion and 3D matrix confinement determine jamming transitions in breast cancer invasion. Nat. Cell Biol. 22, 1103–1115 (2020).
Vishwakarma, M. et al. Mechanical interactions among followers determine the emergence of leaders in migrating epithelial cell collectives. Nat. Commun. 9, 3469 (2018).
Reffay, M. et al. Interplay of RhoA and mechanical forces in collective cell migration driven by leader cells. Nat. Cell Biol. 16, 217–223 (2014).
Peglion, F., Llense, F. & Etienne-Manneville, S. Adherens junction treadmilling during collective migration. Nat. Cell Biol. 16, 639–651 (2014).
Riahi, R. et al. Notch1-Dll4 signalling and mechanical force regulate leader cell formation during collective cell migration. Nat. Commun. 6, 6556 (2015).
Donà, E. et al. Directional tissue migration through a self-generated chemokine gradient. Nature 503, 285–289 (2013).
Lawrence, R. E. & Zoncu, R. The lysosome as a cellular centre for signalling, metabolism and quality control. Nat. Cell Biol. 21, 133–142 (2019).
Ballabio, A. & Bonifacino, J. S. Lysosomes as dynamic regulators of cell and organismal homeostasis. Nat. Rev. Mol. Cell Biol. 21, 101–118 (2020).
Schiefermeier, N. et al. The late endosomal p14-MP1 (LAMTOR2/3) complex regulates focal adhesion dynamics during cell migration. J. Cell Biol. 205, 525–540 (2014).
Pu, J., Guardia, C. M., Keren-Kaplan, T. & Bonifacino, J. S. Mechanisms and functions of lysosome positioning. J. Cell Sci. 129, 4329–4339 (2016).
Bretou, M. et al. Lysosome signaling controls the migration of dendritic cells. Sci. Immunol. 2, eaak9573 (2017).
Wang, G. et al. Biomechanical control of lysosomal secretion via the VAMP7 Hub: a tug-of-war between VARP and LRRK1. iScience 4, 127–143 (2018).
Jerabkova-Roda, K. et al. Peripheral positioning of lysosomes supports melanoma aggressiveness. bioRxiv 2023, 548108 (2023).
Hämälistö, S. & Jäättelä, M. Lysosomes in cancer-living on the edge (of the cell). Curr. Opin. Cell Biol. 39, 69–76 (2016).
Abreu-Blanco, M. T., Verboon, J. M., Liu, R., Watts, J. J. & Parkhurst, S. M. Drosophila embryos close epithelial wounds using a combination of cellular protrusions and an actomyosin purse string. J. Cell Sci. 125, 5984–5997 (2012).
Aragona, M. et al. Defining stem cell dynamics and migration during wound healing in mouse skin epidermis. Nat. Commun. 8, 14684 (2017).
Takaya, K. et al. Actin cable formation and epidermis–dermis positional relationship during complete skin regeneration. Sci. Rep. 12, 15913 (2022).
Chen, T. et al. Large-scale curvature sensing by directional actin flow drives cellular migration mode switching. Nat. Phys. 15, 393–402 (2019).
Napolitano, G. & Ballabio, A. TFEB at a glance. J. Cell Sci. 129, 2475–2481 (2016).
Martina, J. A., Chen, Y., Gucek, M. & Puertollano, R. MTORC1 functions as a transcriptional regulator of autophagy by preventing nuclear transport of TFEB. Autophagy 8, 903–914 (2012).
Bálint, Š, Verdeny Vilanova, I., Sandoval Álvarez, Á. & Lakadamyali, M. Correlative live-cell and super-resolution microscopy reveals cargo transport dynamics at microtubule intersections. Proc. Natl. Acad. Sci. USA 110, 3375–3380 (2013).
de Forges, H., Bouissou, A. & Perez, F. Interplay between microtubule dynamics and intracellular organization. Int J. Biochem. Cell Biol. 44, 266–274 (2012).
Rosa-Ferreira, C. & Munro, S. Arl8 and SKIP act together to link lysosomes to kinesin-1. Dev. Cell 21, 1171–1178 (2011).
Randall, T. S. et al. A small-molecule activator of kinesin-1 drives remodeling of the microtubule network. Proc. Natl. Acad. Sci. USA 114, 13738–13743 (2017).
Guardia, C. M. et al. Reversible association with motor proteins (RAMP): A streptavidin-based method to manipulate organelle positioning. PLoS Biol. 17, e3000279 (2019).
Balasubramaniam, L. et al. Investigating the nature of active forces in tissues reveals how contractile cells can form extensile monolayers. Nat. Mater. 20, 1156–1166 (2021).
Weber, G. F., Bjerke, M. A. & DeSimone, D. W. A mechanoresponsive cadherin-keratin complex directs polarized protrusive behavior and collective cell migration. Dev. Cell 22, 104–115 (2012).
Zimmermann, J., Camley, B. A., Rappel, W. J. & Levine, H. Contact inhibition of locomotion determines cell-cell and cell-substrate forces in tissues. Proc. Natl. Acad. Sci. USA 113, 2660–2665 (2016).
Valon, L., Marín-Llauradó, A., Wyatt, T., Charras, G. & Trepat, X. Optogenetic control of cellular forces and mechanotransduction. Nat. Commun. 8, 14396 (2017).
Ravasio, A. et al. Gap geometry dictates epithelial closure efficiency. Nat. Commun. 6, 7683 (2015).
Rausch, S. et al. Polarizing cytoskeletal tension to induce leader cell formation during collective cell migration. Biointerphases 8, 32 (2013).
Machacek, M. et al. Coordination of Rho GTPase activities during cell protrusion. Nature 461, 99–103 (2009).
Ruth, E. S. Cicatrization of wounds in vitro. J. Exp. Med 13, 422–424 (1911).
Fritz, R. D. & Pertz, O. The dynamics of spatio-temporal Rho GTPase signaling: formation of signaling patterns. F1000Res https://doi.org/10.12688/f1000research.7370.1 (2016).
Remorino, A. et al. Gradients of Rac1 nanoclusters support spatial patterns of Rac1 signaling. Cell Rep. 21, 1922–1935 (2017).
Palamidessi, A. et al. Endocytic trafficking of Rac is required for the spatial restriction of signaling in cell migration. Cell 134, 135–147 (2008).
Valentin, G., Haas, P. & Gilmour, D. The chemokine SDF1a coordinates tissue migration through the spatially restricted activation of Cxcr7 and Cxcr4b. Curr. Biol. 17, 1026–1031 (2007).
Monteleon, C. L. et al. Lysosomes support the degradation, signaling, and mitochondrial metabolism necessary for human epidermal differentiation. J. Invest Dermatol 138, 1945–1954 (2018).
Juarez, M. T., Patterson, R. A., Li, W. & McGinnis, W. Microinjection wound assay and in vivo localization of epidermal wound response reporters in Drosophila embryos. J. Vis. Exp. https://doi.org/10.3791/50750 (2013).
Pothapragada, S. P., Gupta, P., Mukherjee, S. & Das, T. Matrix mechanics regulates epithelial defence against cancer by tuning dynamic localization of filamin. Nat. Commun. 13, 218 (2022).
Acknowledgments
We thank Aneesh T. Veetil for the critical discussion. R.M. and T.D. thank Prof. Dr. Joachim Spatz (MPI for Medical Research) for giving access to the Airy-Scan super-resolution microscope. We thank Dr. Mahak Sharma (IISER Mohali) for generously providing plasmids used in this study and Y. Fujita (Kyoto University) for the WT MDCK cell line. R.M. is a DBT/Wellcome Trust India Alliance Early Career Fellow. This project is funded by the DBT/Wellcome Trust India Alliance (Ref. No. IA/E/19/1/504967 to R.M.). In addition, the authors acknowledge generous support from the partner group grant of the Max Planck Society, the Human Frontier Science Program (HFSP) research grant (RGP0007/2022), and intramural funds at TIFR Hyderabad from the Department of Atomic Energy (DAE), India, under the Project Identification Number RTI 4007 to D.M., S.R., S.B., P.K., P.G., B.T., M.J. and T.D.
Author information
Authors and Affiliations
Contributions
R.M. and T.D. formulated the project. R.M. performed most of the experiments and analyses. D.M. and S.B. performed mouse embryonic skin wound assays, stained and acquired confocal micrographs for representation and analysis. S.R. performed micropatterning experiments and super-resolution microscopy of embryonic skin samples. P.K. standardized and performed initial opto-GEF experiments. P.G. contributed to KLC2-expressing stable cell line generation, acquiring representative videos for the Opto-GEF experiment, and helped in designing POF analysis. B.T. modified the MATLAB code for MDE calculation. M.J. provided critical inputs to the design of Drosophila embryo experiments. R.M., M.J., and T.D. wrote the manuscript. All authors agreed on the manuscript as in the submitted version.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Marwaha, R., Manoj, D., Rawal, S. et al. Mechanosensitive dynamics of lysosomes along microtubules regulate leader cell emergence during collective cell migration. Nat Commun 17, 921 (2026). https://doi.org/10.1038/s41467-025-67645-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-67645-0
