
Abstract
5-methylcytosine (5mC) DNA methylation is an ancient epigenetic mechanism. Methyl-CpG-binding domain protein 2 or 3 (MBD2/3) is a component of the Nucleosome Remodeling and Deacetylase (NuRD) complex, essential for gene regulation in mammals. The MBD2/3 ortholog is surprisingly preserved even in 5mC-free invertebrates, such as nematodes and flies. We found that Caenorhabditis elegans MBD-2 (Cel-MBD-2), despite without the MBD domain, is a conserved component of the NuRD complex. Cel-MBD-2 null mutation or deletion of its NuRD-interacting coiled-coil domain causes morphological defects and infertility. While a minority of Cel-MBD-2 colocalizes with NuRD at active chromatin, the majority of Cel-MBD-2 localizes to repressive chromatin environments enriched with H3K27me3 and H3K9me2/3, not overlapping with NuRD. The mechanism by which Cel-MBD-2 localizes to the repressive chromatin is still unknown, but is independent of NuRD or MBD-5mC interaction. The potential compensation of DNA methylation by histone modifications highlights the adaptability of epigenetic regulation during evolution.
Similar content being viewed by others
Introduction
5-methylcytosine (5mC) DNA methylation is indispensable in vertebrate developmental processes, including imprinting, X-inactivation, and silencing of transposable elements. 5mC is recognized by methyl-CpG binding domain (MBD)-containing proteins, including MBD2, MBD3 and MeCP2. Vertebrate paralogs MBD2 and MBD3 are believed to have derived from duplication of the single ancestral invertebrate MBD2/31. Mammalian MBD2 isoforms all contain a functional MBD domain2. MBD3 isoforms possess either a mutation on a critical residue in MBD or a truncated MBD domain (Fig. 1A, B). Consequently, mammalian MBD3 can only weakly bind to 5mC3. Therefore, MBD3 primarily associates with hypomethylated CG-rich regions in the genome, whereas mammalian MBD2 predominantly binds to 5mC-dense CG-rich regions3,4.

A Sequences are aligned by ClustalOmega from top to bottom: Cel-MBD-2, human MBD2a, MBD2b, MBD2t (lacking part of the intrinsically disordered region (IDR) and coiled-coil domain (CC)), MBD3a, MBD3b and MBD3c. Gray box indicates the GR-rich domain present in human MBD2a and MBD2t. Orange box indicates the methyl-binding domain (MBD). The red box within the orange box indicates the essential residue for high affinity for methylated cytosine in MBD2, which is not conserved in MBD3. Blue box indicates the intrinsically disordered region (IDR) and the black box within the blue box indicates the conserved “SIFKQPV” sequence of IDR in other metazoans10. Green box indicates the coiled-coil domain (CC). B Schematic diagram (generated with Prosite MyDomains94) demonstrating the domains in Cel-MBD-2 and human MBD2 and 3 isoforms. Cel-MBD-2 only possesses the IDR and CC for interaction with NuRD complex components. Human MBD2 isoforms show high affinity for 5mC, but MBD3 possesses a point mutation in its MBD domain that leads to its diminished 5mC affinity. C Live images of GFP::3xFLAG::MBD-2 and mCherry::H2B expression in one-cell embryo. The time points indicated are relative to the metaphase time frame. D Live images of GFP::3xFLAG::MBD-2 and mCherry::H2B expression in young adult worms. The bottom panel shows close-up images of the germ nuclei (in which mCherry::H2B has strong germline expression, while GFP::3xFLAG::MBD-2 has minimal expression at the transition zone of germline), the ventral nerve cord nuclei (indicated with white arrowheads), and the intestinal nuclei (indicated with white arrowheads).
In mammals, MBD2 and MBD3 mutants show different phenotypes. Mammalian MBD2 knockout leads to behavioral phenotypes, including defects in maternal nurturing behavior and downregulation of autism-related genes in mice5,6. In contrast, knockout of MBD3 leads to preimplantation lethality in mice owing to its essential role in stem cell differentiation7. Interestingly, the longest isoform MBD2a (containing N-terminal MBD and GR-rich domain, both of which confer 5mC affinity) with the highest affinity for 5mC could only partially rescue the differentiation block in mbd3 knockout2. On the other hand, overexpression of MBD2 isoform b or mutants with a lower 5mC affinity (mutation at MBD or deletion of GR-rich domain) could fully rescue the differentiation defect of mbd3 knockout2. This suggests that MBD2 has retained the conserved MBD domain that confers 5mC affinity, allowing it to diversify its targets compared to MBD3. Without these domains, MBD2 is functionally redundant with MBD3. In addition, depletion of Nucleosome Remodeling and Deacetylase (NuRD) components leads to a similar differentiation block as seen in MBD3 mutant8. Therefore, these essential functions in stem cell differentiation and survival likely arise from the genes regulated by the MBD2/3-NuRD interaction. On the other hand, MBD2-specific gene targets mediated by 5mC-binding may confer cognitive ability, which is non-essential but may provide survival advantages5,6. In invertebrates with 5mC, for example in sponges, knockdown of the single MBD2/3 induces a highly penetrant developmental defect9.
On the other hand, the function of MBD2/3 in invertebrates that have lost 5mC methylation is less studied. For example, in the nematode P. pacificus, knockout of Ppa-mbd-2 leads to a fully penetrant egg-laying defect (Egl) related to a neurotransmitter defect and downregulation of eud-1, the master regulator of mouth form plasticity, but the mbd-2 mutants are viable10,11. Knockdown of C. briggsae Cbr-mbd-2 leads to strong uncoordinated movement (Unc) in > 50% of the F1 progenies and larval arrest in 35%12. In C. elegans, Cel-mbd-2 transcript has been detected at all larval stages and in adults12. Knockdown of Cel-mbd-2 led to 18% animals with mild Unc phenotype without paralysis, 11% with exploding vulva, and < 1% with multivulva (Muv)12. The mild, low penetrance of the RNA interference (RNAi) knockdown phenotypes was surprising, but no mutant analysis has been performed before current study.
Regardless of their different affinities to 5mC, mammalian MBD2 and MBD3 are mutually exclusive scaffolding components of the highly conserved Nucleosome Remodeling and Deacetylase (NuRD) complex. The NuRD complex is highly conserved across metazoans. The mammalian NuRD complex can be functionally split into two subcomplexes: (1) a histone deacetylase core consisting of MTA1/2/3, HDAC1/2, and RBBP4/7, and (2) a nucleosome remodeling core consisting of CHD3/4/5 (Mi-2) and GATAD2A/B (Table 1). MBD2/3 acts as a scaffolding component connecting the two subcomplexes. This holds true for the most ancestral metazoan sponges (which possess 5mC in their genomic DNA)9, Drosophila (in which the presence of genomic 5mC is disputed)13, planarians (which lack 5mC14), Xenopus15, mice16, and humans17.
In mice, MBD3 is confined to the nucleus and has been proposed to facilitate localization of the NuRD complex to chromatin. In mouse embryonic stem cells (mESCs), sequestering MBD3 in the cytoplasm produced transcriptomic changes similar to mbd3 knockout mutants, resulting in a loss of the NuRD component CHD4 binding to certain loci18. Mammalian MBD3’s binding sites are largely encompassed by the NuRD complex component binding sites, featuring transcriptional mark P300 and RNA polymerase II, and histone modifications of open chromatin, such as H3K27ac, H3K4me1, and H3K4me33,7,18,19. The general affinity of the NuRD complex to open chromatin is likely conserved from mammals18 to C. elegans20 based on ChIP-seq data of some NuRD components.
A conventional model posits that mammalian MBD2 recruits the NuRD complex to methylated promoters, silencing transcription through histone deacetylation. Supporting this view, histone deacetylase inhibitor trichostatin A leads to de-repression of certain MBD2-NuRD target genes21,22. However, recent evidence shows that only a small subset of vertebrate MBD2 binds to hypomethylated regions marked by active histone modifications4,23, overlapping with those of MBD3 and NuRD complex4,7,18. Consistently, MBD2t, a MBD2 isoform lacking the NuRD-interacting coiled-coil domain, does not bind to these MBD2-NuRD-overlapping sites4.
Surprisingly, MBD2 can also function independently of NuRD. In both normal and cancer cells, human MBD2 mainly binds to silent promoters enriched with methylated CpG3,24. Recent studies suggest that MBD2 binds to a large subset of methylated promoter regions without recruiting NuRD4. These 5mC-dense promoters bound by MBD2 are CG-rich, silent, and without correlation with any histone marks3,4,23,24. The above information leads to a new model of dual binding patterns of mammalian MBD2a: (1) a major subset of NuRD-independent binding to silent 5mC-dense regions, and (2) a minor subset of NuRD-overlapping binding to open chromatin, hypomethylated regions, similar to MBD3.
This ancient 5mC DNA methylation and its associated DNA methyltransferases (DNMT1/3) are lost in diverse invertebrate lineages, including C. elegans, Drosophila25, and planarians14. C. elegans is a well-validated 5mC-free model organism, as confirmed by various approaches, including high-performance liquid chromatography26,27, anti-5mC dot blotting of genomic DNA and immunofluorescence staining of germline28. The loss of genomic 5mC and the respective DNA methyltransferases (DNMT1 and DNMT3) in C. elegans has been proposed as a mechanism to evade DNMT-induced alkylation damage29. While basal members of marine nematodes29 and most insects30 retain functional 5mC methylation, the loss of 5mC has occurred independently in derived nematodes and insects, representing a striking case of convergent evolution. The convergent evolution of 5mC loss presents a critical question: how is the gene silencing function by DNA methylation upheld by compensatory epigenetic mechanisms? Alternative epigenetic tools such as repressive histone modifications (e.g., H3K27me3, H3K9me2) and noncoding RNAs may facilitate transcriptional silencing. However, the detailed mechanisms remain unresolved.
Intriguingly, despite the loss of 5mC, Methyl-CpG Binding Domain protein 2 (MBD2), a canonical 5mC reader historically linked to DNA methylation, is present in all these lineages that lack 5mC. For example, C. elegans MBD-2 (Cel-MBD-2) lacks the methyl-CpG binding domain. Despite limited information from previous RNA interference (RNAi) knockdown phenotypes, the function of Cel-MBD-2 in development remains largely unknown12, and whether it is part of the NuRD complex in C. elegans is unclear. Here, we have constructed Cel-mbd-2 null mutants and characterized their phenotypes and transcriptomes. By tagging Cel-MBD-2 with GFP, we observed its expression and localization during development and performed immunoprecipitation followed by mass spectrometry (IP-MS), which confirmed that Cel-MBD-2 indeed interacts with NuRD components. We also performed ChIP-seq on Cel-MBD-2, and compared the results with existing data on NuRD components and various histone modifications to determine its binding patterns. The results reveal dual binding patterns of Cel-MBD-2, resembling those of mammalian MBD2, in which MBD2 either binds its target genes together with or independently of the NuRD complex to modulate gene regulation.
Results
Cel-MBD-2 lacks the methyl-binding domain (MBD) and is expressed ubiquitously in the nucleus
Cel-MBD-2 does not possess the methyl-binding domain (MBD), but retains the coiled-coil (CC) domain and intrinsically disordered region (IDR) implicated in NuRD complex interactions (Fig. 1A, B). To determine the cellular localization and dynamics of Cel-MBD-2, we tagged it with GFP::3xFLAG at its N-terminus to avoid interference with the C-terminal coiled-coil domain essential for interacting with NuRD components. MBD-2 is highly expressed in the nuclei of embryos, and its localization is cell cycle-dependent (Fig. 1C). Specifically, during interphase, MBD-2 is localized in the nucleoplasm. In prometaphase to anaphase, MBD-2’s nuclear signal gradually decreases and is excluded from chromosomes. During cytokinesis, MBD-2’s signal gradually accumulates back in the nucleoplasm. MBD-2 is also enriched in the nucleoplasm ubiquitously in larvae and adult worms, including the germline and intestine (Fig. 1D, Supplementary Fig. 1). Such nuclear localization resembles some of the other NuRD components, including CHD-3, LET-418, and HDA-1 (Supplementary Fig. 2A–D). Expression of Cel-MBD-2 in the nuclei in all life stages is consistent with its potential role in gene regulation throughout development.
Cel-MBD-2 is essential for fertility and vulval development
To investigate Cel-MBD-2’s functions beyond the RNAi knockdown approach12 (Supplementary Fig. 3B, C), we constructed a mbd-2 null mutant. mbd-2 is located in an intron of pigo-1, which is on the opposite strand of mbd-2 (Fig. 2A). To generate the null mutation of MBD-2, we inserted a 43-nucleotide-long knock-in cassette (STOP-IN) 31, including five premature stop codons, in the first exon (Fig. 2A). Such an approach ensures premature termination of translation at all three reading frames31. This approach also alleviates the need to delete any DNA sequence in pigo-1’s intron, and thus should cause less effects on the expression of the overlapping gene pigo-1. Since homozygous mbd-2-/- mutant is sterile (Fig. 2B), the strain is maintained as heterozygous with a balancer chromosome, hT2, containing a mbd-2 wildtype copy (see Materials and methods). To verify that the observed phenotypes, including sterility, are due to mbd-2 loss-of-function mutation, but not related to the overlapping gene pigo-1, we expressed gfp::3xflag::mbd-2 single-copy transgene (tg) driven by mbd-2 endogenous promoter at the oxTi365 locus. The tg fully rescues the infertility and morphology defects of mbd-2-/- mutant (Fig. 2B).

A Genomic localization of mbd-2 visualized with JBrowse95. The design of mbd-2 null mutant (mbd-2-/-) and coiled-coil deletion mutant (gfp::3xflag::mbd-2∆CC/∆CC) are shown schematically. B Box-and-whiskers plot of brood size of wildtype (N2 strain) and mbd-2 null mutant worms. From left to right is the brood size of wildtype (N2 strain) worms (n = 31), P0 mbd-2-/- worms (from self-fertilization of mbd-2+/- carrying the genetic balancer hT2) (n = 18), F1 mbd-2-/- worms (from self-fertilization of P0 mbd-2-/-) (n = 37), a transgene-rescued strain [mbd-2-/-; tg/tg] (mbd-2-/- at the endogenous locus and carrying a single-copy insertion of mbd-2p::gfp::3xflag::mbd-2 transgene (tg)) (n = 16), P0 gfp::3xflag::mbd-2∆CC/∆CC worms (n = 13), and F1 gfp::3xflag::mbd-2∆CC/∆CC worms (n = 12). Each dot represents the brood size of a single worm. The brood size differences between mutants and wildtype, and between the two mutants (mbd-2∆CC/∆CC and mbd-2-/-), were analyzed using a two-tailed t-test. Box plots indicate median (middle line), 25th, 75th percentile (box) and minima, maxima (whiskers). Source data are provided as a Source Data file. C P0 mbd-2-/- worms with protruding vulva. D Bagging phenotype, in which hatched progenies stay in the uterus, is observed in P0 mbd-2-/- young adults (day 3 post L1). E Multivulva (indicated with white arrowheads) is observed in 32.4% F1 mbd-2-/- worms (n = 37). F Deformed anal region (Dar) phenotype (indicated with white arrowheads) is observed in 12.2% of F1 mbd-2-/- worms (n = 74).
Phenotypically, all of the P0 mbd-2-/- worms are small and relatively immobile adults with a fully penetrant protruding vulva (Pvl) phenotype (Fig. 2C). The P0 mbd-2-/- worms are able to produce on average 10 progenies (Fig. 2B). Almost all of the P0 mbd-2-/- worms died on day 3-4 of adult stage, likely from bagging, in which embryos that cannot be laid hatch inside the uterus and cause lethality to the mothers (Fig. 2D). Interestingly, the P0 mbd-2-/- mutants also frequently avoid the bacterial lawn.
The F1 mbd-2-/- worms, which also lack maternal mbd-2 transcripts, are even smaller and uncoordinated. The F1 mbd-2-/- worms have fully penetrant protruding vulva (Pvl) and sterility. Thirty-two percent (32.4%) of the F1 mbd-2-/- living adults have two vulvas (Fig. 2E). Thirty-seven percent (37.3%) of F1 mbd-2-/- mutants arrest in various developmental stages (L3-L4 larval stages), and 12.2% also show a deformed anal region (Dar) phenotype (Fig. 2F).
The severity of the phenotypes, including sterility and morphological defects, increases when both maternal and zygotic mbd-2 expression is depleted in F1 mbd-2-/-, as compared to P0 mbd-2-/-, suggesting that both maternal and zygotic mbd-2 contribute to the development and fertility of the offspring. On the other hand, heterozygous mbd-2+/- worms produced by mating P0 mbd-2-/- hermaphrodites with N2 wildtype males are fertile and morphologically indistinguishable from wildtype (Supplementary Fig. 3A), indicating that zygotic expression of mbd-2 is sufficient for its reproduction function. Thus, mbd-2 is not strictly maternal-effect sterile (mes), as is the case in PRC2 complex mutants32. The cause of sterility in mbd-2-/- is unclear, but it is unlikely to be caused solely by the protruding vulva (Pvl), as RNAi of mbd-2 resulted in a reduced brood size of F1 worms even without a protruding vulva (Supplementary Fig. 3B). Taken together, we have identified the role of Cel-MBD-2 in vulva development and fertility.
Drastic dysregulation of the transcriptome in C. elegans mbd-2 null mutants
To investigate the potential roles of MBD-2 in regulating genes involved in development and reproduction, we analyzed the transcriptomic changes in mbd-2-/- null mutants. We performed RNA-sequencing (RNA-seq) on young adult worms from N2 wildtype, P0, and F1 mbd-2-/- strains, with two biological replicates for each group. Using an adjusted p-value < 0.05 and Fold Change (FC) ≥ 2, we identified 5356 protein-coding genes that exhibited differential expression in P0 mbd-2-/- when compared to wildtype N2 worms (Supplementary Data 3). This represents more than a quarter of all protein-coding genes in C. elegans, indicating the importance of MBD-2 in master gene regulation. Specifically, 4206 and 1150 genes were significantly upregulated and downregulated, respectively, in the P0 mbd-2-/- worms (Supplementary Fig. 5A, Supplementary Data 3). The dysregulation is further enhanced in F1 mbd-2-/- worms with 5099 and 1547 genes significantly upregulated and downregulated, respectively (Supplementary Fig. 5B, Supplementary Data 4). The higher proportion of upregulated genes as compared to downregulated genes suggests that Cel-MBD-2 functions primarily as a gene repressor, similar to its mammalian ortholog MBD233.
As expected, the upregulated genes overlap extensively between the P0 and F1 mbd-2-/- mutants, but to a lesser extent for the downregulated genes (Fig. 3A, B). In general, the gene ontology (GO) term analysis suggests that P0 and F1 mbd-2-/- consistently led to upregulation of genes involved in nervous system development, and downregulation of genes involved in metabolic pathways (Fig. 3C, D, Supplementary Fig. 6A–D).

A, B Venn diagram showing the number of genes significantly (A) upregulated and (B) downregulated (FC ≥ 2, adjusted p-value < 0.05) that are overlapping in P0 and F1 mbd-2-/- mutant. C, D Top 5 most enriched gene ontology (GO) in the biological process (BP), Molecular Function (MF) and KEGG pathway terms for the genes commonly (C) upregulated (2949) and (D) downregulated (518) in P0 and F1 mbd-2-/- mutant. The -log10 Benjamini-adjusted two-tailed Wald test p-values from DESeq2 were plotted. Gene ratio represents the percentage of differentially expressed genes associated with that GO term. The gene count associated with the GO term is indicated by the size of the circle. Protein-coding genes with a non-zero read count in at least one RNA-seq sample (N2, P0 or F1 mbd-2-/-) were used as the background gene list. DAVID website was used with default parameters. E Expression of homozygous GFP::3xFLAG::MBD-2∆CC by live imaging, in differential interference contrast (DIC) and GFP channel. F, G Heatmap depicting the log2 fold change of differentially expressed protein-coding genes filtered by Benjamini-adjusted two-tailed Wald test p-value < 0.05. F 7714 differentially expressed genes overlap in mbd-2-/- and gfp::3xflag::mbd-2∆CC/∆CC and G 668 differentially expressed genes overlap in mbd-2-/-, gfp::3xflag::mbd-2∆CC/∆CC, hda-1 RNAi42 and dcp-66 RNAi43. Upregulated and downregulated genes are indicated in red and blue, respectively. Rows were clustered by complete Euclidean distance.
Interestingly, GO terms relevant to development, such as cell fate specification and Wnt signaling pathway, are enriched in genes that are exclusively downregulated in F1 mbd-2-/- (Supplementary Fig. 6F). Such differences may explain why completely penetrant sterility, additional morphological defects and growth arrest are only observed in F1 mbd-2-/-, but not in P0 mbd-2-/- which may still possess maternal mbd-2 transcripts in their early development.
On the other hand, 1691 of the P0 mbd-2-/- dysregulated genes are not significantly changed in F1 mbd-2-/-. Moreover, 198 genes are dysregulated in the opposite direction in F1 compared to P0 mbd-2-/-. For example, the biological process “DNA-binding transcription factor activity, RNA polymerase II-specific” (GO:0000981) is overrepresented in P0 mbd-2-/- upregulated and F1 mbd-2-/- downregulated GO terms (Supplementary Fig. 6A, D). The reason for such opposite-direction dysregulation is not clear, but could possibly be due to negative feedback loops or secondary effects, as the affected genes regulate transcription as well.
The coiled-coil domain contributes to major gene regulation function of C. elegans mbd-2
MBD2/3 is known to be a conserved component of the NuRD complex throughout metazoa9,34. The coiled-coil domain (CC) at C-terminus is highly conserved from C. elegans to human MBD2 and MBD3 (Fig. 1A)10, essential for interacting with the NuRD complex35,36,37,38. To further elucidate the function of its coiled-coil domain, we constructed a truncated MBD-2 without its C-terminal CC. To design the deletion of the CC of Cel-MBD-2, DeepCoil239 was used to detect the residues with a coiled-coil-forming probability of ≥ 20%, and identified 15 amino acids (residue 190-204) (Fig. 1A, B, gray label). These residues fall within the Cel-MBD-2 CC (residue 178-210) based on sequence alignment with CC of human MBD2 (residue 361–393)40 (Fig. 1A, gray bracket above green box). We reasoned that the deletion of these 15 amino acids within the CC domain would be sufficient to disrupt the domain’s structure and generated a strain of gfp::3xflag::mbd-2∆CC/∆CC mutant maintained with the same balancer as mbd-2-/- mutant. Interestingly, the truncated GFP::3xFLAG::MBD-2∆CC is stably expressed in embryo nuclei (Fig. 3E), consistent with a human MBD-2 study, which showed preserved nuclear location after deletion of the CC41.
Remarkably, gfp::3xflag::mbd-2∆CC/∆CC phenocopies the null mutation, as characterized by the low brood size in P0 gfp::3xflag::mbd-2∆CC/∆CC worms and complete sterility observed in F1 gfp::3xflag::mbd-2∆CC/∆CCworms (Fig. 2B). In addition, 14.7% of F1 gfp::3xflag::mbd-2∆CC/∆CC worms possess two vulvas (n = 170), and a fully penetrant protruding vulva phenotype is observed in both generations.
Next, we performed RNA-seq on young adult whole worms with three biological replicates for gfp::3xflag::mbd-2 (control) and P0 gfp::3xflag::mbd-2∆CC/∆CC mutants, and two biological replicates for F1 gfp::3xflag::mbd-2∆CC/∆CC. Interestingly, the transcriptomic changes in P0 and F1 gfp::3xflag::mbd-2∆CC/∆CC are highly similar to those in mbd-2-/- worms when compared to their respective controls (Fig. 3F, Supplementary Fig. 4G, 5A–F). The majority of upregulated (91%) and downregulated (52%) genes in P0 mbd-2-/- mutants are also dysregulated in the same direction in P0 gfp::3xflag::mbd-2∆CC/∆CC (Supplementary Fig. 5E).
To determine if MBD-2 and other NuRD components co-regulate the same genes, we have reanalyzed published RNA-seq data of the NuRD components hda-1 RNAi42 and dcp-66 RNAi43 in young adult worms together with our mbd-2-/- and gfp::3xflag::mbd-2∆CC/∆CC RNA-seq. The RNAi experiments42,43, likely resulting in partial protein depletion, yielded fewer differentially expressed genes (4972 for hda-1 RNAi and 1677 for dcp-66 RNAi) than our mutants ( > 11000 genes in P0 mbd-2-/- and P0 gfp::3xflag::mbd-2∆CC/∆CC), with a filtering criteria of adjusted p-value < 0.05. Among them, 668 differentially expressed genes overlap in all our P0 mbd-2-/-, P0 gfp::3xflag::mbd-2∆CC/∆CC, hda-1 RNAi, and dcp-66 RNAi, mostly showing expression change in the same direction (Fig. 3G). Specifically, 317 out of 668 genes are consistently upregulated in P0 mbd-2-/-, dcp-66 RNAi, and hda-1 RNAi. These genes, normally repressed by NuRD, are involved in nucleosome assembly, axon guidance and structural constituents of chromatin (Supplementary Fig. 6G). On the other hand, 171 out of 668 genes are consistently downregulated in the four conditions. These NuRD-activated genes are involved in the regulation of immune response and metabolic pathways (Supplementary Fig. 6H). Since most of the dysregulated genes are dysregulated in the same direction (488 out of 668) in mbd-2 null mutant, coiled-coil mutant, and NuRD complex component depletion, it suggests a functional association between MBD-2 and the NuRD complex.
MBD-2 is a scaffolding component of the NuRD complex through its coiled-coil (CC) domain
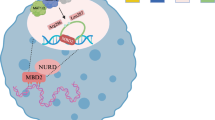
The NuRD complex can be functionally split into two subcomplexes, a histone deacetylase core and a nucleosome remodeling core9. Mammalian MBD2/3 serves as the scaffold of the NuRD complex. At one end, MBD2/3’s intrinsically disordered region (IDR) interacts with histone deacetylase core through the interface formed between MTA1/2/3 dimer-HDAC1/2 dimer35,37,40 (Fig. 4A). At the other end, MBD2/3’s C-terminal coiled-coil domain (CC) interacts with the coiled-coil domain of GATAD2A/B, which then recruits the chromatin remodeler CHD3/4/5 (Fig. 4A)35,36,37. The two subcomplexes that are connected via MBD-2’s IDR and CC are structurally conserved throughout metazoan. C. elegans possesses at least one ortholog for each component (Table 1 and Fig. 4A). Unexpectedly, a previous study could not detect any interaction between NuRD components and Cel-MBD-2, challenging such structural conservation of the NuRD complex in a methylation-free context44.

A Left: the known model of human/mouse NuRD complex. Right: our proposed model of Cel-NuRD complex. B Left: FLAG IP-MS of young adult worms expressing GFP::3xFLAG::MBD-2. Average iBAQ (Intensity-based absolute quantitation of protein peptides) in log2 scale of the 2 replicates are represented on the axes. FLAG IP of N2 (untagged MBD-2) worms was used as the negative control on X-axis. NuRD complex components and DVE-1 are annotated. Right: Schematic diagram of Cel-MBD-2 interacting with NuRD components, except LIN-40 could not be detected in IP-MS. C Venn diagram depicting overlapping interactors of MBD-2 (this study), HDA-146 and DVE-147 identified by IP-MS. “Confidently lost interactors” of MBD-2∆CC are in red. D Co-IP verification of MBD-2 interactors using anti-FLAG magnetic beads targeting GFP::3xFLAG::MBD-2. Left panel: Western blots of NuRD components (from top to bottom: HDA-1, LIN-53, and LET-418) detected from anti-FLAG IP. Right panel: Western blots of MBD-2 by GFP antibodies performed in parallel. All negative controls are of the same genetic background (N2 or tagged NuRD components) as GFP::3xFLAG::MBD-2, but with untagged MBD-2. Full blots of three biological replicates are available in Supplementary Fig. 7. E Left: FLAG IP-MS of young adult worms expressing GFP::3xFLAG::MBD-2∆CC. Average iBAQ in log2 scale of the 2 replicates are represented on the axes. The immunoprecipitation efficiency for ∆CC was lower than the full-length MBD-2 protein (Supplementary Data 7). To prevent this difference from being misinterpreted as a loss of specific protein interactions, the log2 iBAQ values of each interactor in the negative control (on the X-axis) were normalized before plotting (refer to Materials and methods for the normalization). The red dashed diagonal line indicates x = y. Right: Schematic diagram of Cel-MBD-2∆CC-containing NuRD complex. The coiled-coil interaction of MBD-2-DCP-66 is lost, leading to dissociation of the nucleosome remodeling core. Yet, LET-418 is still detected in ∆CC IP-MS. Source data for B and E are provided as Source Data files. The organism icons in panels A, B, and E are created in BioRender. Zheng, C. (2025) https://BioRender.com/fa05xlk.
To test if Cel-MBD-2 is indeed in the NuRD complex, we performed native Immunoprecipitation-Mass Spectrometry (IP-MS) on tagged MBD-2 (GFP::3xFLAG::MBD-2) in young adult worms. Immunoprecipitation was carried out on 0.5 ml worms using an anti-FLAG antibody with two biological replicates. Parallel IP-MS experiments were performed using untagged wildtype (N2) lysates as negative controls for background subtraction. High-confidence interactors were defined as proteins exhibiting ≥ 2-fold enrichment in iBAQ (Intensity-based absolute quantitation of protein peptides) intensity over the negative control (two-tailed t-test, p-value < 0.05). Using these criteria, we identified 140 MBD-2-interacting proteins (Fig. 4B, Supplementary Data 7), including NuRD components HDA-1/HDAC1/2, LIN-53/RBBP4/7, DCP-66/GATAD2A/B, CHD-3/Mi-2, and LET-418/Mi-2. The only exception is LIN-40/MTA2/3, which was not detected in our IP-MS results in adults and embryos.
In addition, DVE-1/SATB1/2, which is a NuRD-interacting transcription factor (TF) conserved from humans45 to C. elegans42, is also identified as a MBD-2 interactor. Notably, CHD-3 and DVE-1 are among the top 3 interactors of MBD-2 (Supplementary Data 7).
We also performed co-immunoprecipitation (Co-IP) followed by Western blot to confirm Cel-MBD-2 interactions with the NuRD histone deacetylase core components HDA-1/HDAC1/2 and LIN-53/RBBP4/7 and a nucleosome remodeling core component LET-418/Mi-2 (Fig. 4D, Supplementary Fig. 7). The above results reveal that MBD-2 is a conserved component of NuRD complex in C. elegans.
Next, we compared our MBD-2 IP-MS data with published IP-MS data on HDA-146 and DVE-147 of young adult worms (Fig. 4C). The 4 interacting proteins overlapping among the MBD-2, HDA-1 and DVE-1 interactors are NuRD complex components. Interestingly, HDA-1 IP-MS shows 32 interactors that are shared by MBD-2, but not by DVE-1. On the other hand, DVE-1 shares only 5 interactors with MBD-2 alone. This trend of having more interactors shared between MBD-2 and HDA-1 (total 36) than between MBD-2 and DVE-1 (total 9) is consistent with that DVE-1 is only one of the NuRD-interacting TFs but not a core component of the NuRD complex.
The CC of MBD2/3 in metazoans has been demonstrated as an essential anchor point for the nucleosome remodeling core of NuRD via its interaction with CC of GATAD2A/B35,36,37,38. We predicted that the deletion of MBD-2 CC would lead to the dissociation of the nucleosome remodeling core from the remaining MBD-2-histone deacetylase core of NuRD. Over a thousand gfp::3xflag::mbd-2∆CC/∆CC synchronized young adult worms were manually picked and collected for IP-MS of MBD-2∆CC using anti-FLAG antibody, and 2 biological replicates were prepared. As expected, 123 out of 141 of the confident MBD-2 interactors showed a decreased interaction in ∆CC (Fig. 4E, Supplementary Data 7). We further identified 35 “confidently lost interactors” (defined in Materials and Methods) that became completely undetectable in both replicates of ∆CC IP-MS (Fig. 4E, gray highlighted interactors in Supplementary Data 7). Intriguingly, CHD-3 and DVE-1 are among the “confidently lost interactors” in ∆CC, and DCP-66 shows reduced interaction with MBD-2∆CC (Supplementary Data 7), consistent with our hypothesis that MBD-2∆CC would lead to dissociation of the nucleosome remodeling core from the rest of the NuRD complex. However, LET-418, which also belongs to the nucleosome remodeling core, did not show a significant loss of interaction with MBD-2∆CC. This could also indicate that LET-418 may interact with MBD-2 independent of MBD-2 CC-DCP-66 CC interaction, possibly outside of the NuRD complex. Our verification of the Cel-MBD-2-NuRD interaction, specifically through its coiled-coil domain, has shed light on the structural and functional conservation despite the loss of the methyl-binding domain.
The majority of Cel-MBD-2 binding sites are non-NuRD-overlapping
Previous ChIP-seq data of some C. elegans NuRD components show general affinity for open chromatin20. We hypothesized that Cel-MBD-2 would bind mainly to open chromatin in association with the NuRD complex, similar to MBD3 in mammals3,4, as both do not have an intact MBD. To test this hypothesis, we performed ChIP-seq experiments on the strain expressing GFP::3xFLAG::MBD-2 at the young adult stage. The experiment included two biological replicates of 1.5 ml worms each, with matched input DNA controls collected for background normalization. Immunoprecipitation was performed using anti-FLAG antibody. To compare the genome-wide binding profile of Cel-MBD-2 to other NuRD components, we also obtained the published ChIP-seq data of the C. elegans NuRD components (HDA-1/HDAC1/242, CHD-3/Mi248 and LET-418/Mi248) and its known interacting transcription factor (DVE-1/SATB1/242) in the same life stage (synchronized young adult worms) (Supplementary Data 10). We discovered 32869 peaks bound by Cel-MBD-2. The number of MBD-2 peaks was much more abundant than those of other NuRD components (3731 for CHD-3, 3551 for LET-418, 10959 for HDA-1 and 7839 for DVE-1) when analyzed with the same peak-calling pipeline (see Materials and methods). This could be due to our deeper sequencing depth ( ≥ 11 GB) compared to published datasets for other Cel-NuRD components (2-5 GB). Alternatively, this may imply that MBD-2 has many more binding sites than other NuRD components.
By extracting the MBD-2 peaks that overlap with both HDA-1 and CHD-3 from the histone deacetylase core and nucleosome remodeling core, respectively, we have defined these peaks as “NuRD-overlapping MBD-2” peaks (6.6% of all MBD-2 peaks) (Fig. 5A, Supplementary Data 11). For this classification, we did not consider the data from LET-418, the mutually exclusive paralog for CHD-3 in the Cel-NuRD complex, even though LET-418 and CHD-3 share extensive overlapping targets in embryos and show partial functional redundancy in embryo development20. Indeed, HDA-1 and LET-418 are present not only in the NuRD complex but also in another chromatin remodeling complex called the MEC complex, which does not contain CHD-344. Thus, overlapping with both HDA-1 and CHD-3 is a stringent and reasonable criterion for “NuRD-overlapping MBD-2” peaks. The remaining MBD-2 peaks that do not overlap with both HDA-1 and CHD-3 are defined as “non-NuRD-overlapping” MBD-2 peaks (93.4% of all MBD-2 peaks) (Fig. 5A, Supplementary Data 12). Our classified NuRD-overlapping MBD-2 peaks and non-NuRD-overlapping MBD-2 peaks are depicted in heatmaps, with enrichment and depletion of other NuRD complex components, respectively (Fig. 5B).

A Venn diagram depicting the classification of “NuRD-overlapping MBD-2 peaks” by extracting MBD-2 peaks that overlap ≥ 1 bp with both HDA-142 and CHD-348 peaks; classification of “non-NuRD-overlapping MBD-2 peaks” by extracting MBD-2 peaks that are not overlapping with both HDA-1 and CHD-3 peaks. n = total number of peaks. B MBD-2 and the NuRD complex component ChIP-seq read count is normalized by count per million (CPM) for both ChIP and input. The ratios of the normalized read counts of ChIP vs. input are z-transformed and plotted as heatmap. The z-score of all the samples at NuRD-overlapping and non-NuRD-overlapping MBD-2 peak centers ± 3 kb are shown. Scale bars of the heatmaps depict the z-max and z-min of all panels. C From top to bottom: Top 5 motifs that are enriched at all MBD-2 ChIP-seq peaks, using C. elegans motif database (upper). A known motif bound by human MBD2 (from a human motif database) is also enriched at all Cel-MBD-2 ChIP-seq peaks (middle). Top 3 motifs that are enriched at NuRD-overlapping MBD-2 peaks, using C. elegans motif database (lower). D MBD-2 and transcription factors (DVE-1, DPL-1, and STA-1) ChIP-seq read count was normalized by count per million (CPM) for both ChIP and input. The ratios of the normalized read counts of ChIP vs. input are z-transformed and plotted as heatmap. The z-score of all the samples at NuRD-overlapping and non-NuRD-overlapping MBD-2 peak centers ± 3 kb are shown. Scale bars of the heatmaps depict the z-max and z-min. E Venn diagrams depicting the number of DPL-1 (top) and STA-1 (bottom) ChIP-seq peaks that overlap with non-NuRD-overlapping and NuRD-overlapping MBD-2 peaks, respectively.
We are aware that the deep sequencing coverage may allow us to discover many more MBD-2 peaks as compared to previous studies on other NuRD components. Therefore, when we increased the stringency in analysis and narrowed down MBD-2 peaks to those with fold enrichment ≥ 4 rather than the default 1.2 of MACS2 peak calling setting (refer to Materials and methods), it resulted in only 5820 MBD-2 peaks. Yet, consistent with the less stringent but default parameter, only 6.9% of these 5820 peaks could be classified as NuRD-overlapping MBD-2 peaks (Supplementary Fig. 8B). In addition, we performed unsupervised k-means clustering of all 32869 MBD-2 peaks using HDA-1 and CHD-3 ChIP signal (Supplementary Fig. 8C). This results in 5009 peaks in Cluster 1 (15.24% of total MBD-2 peaks), with relatively high enrichment with HDA-1 and CHD-3 binding, and 27860 peaks in Cluster 2 (84.76% of total MBD-2 peaks) which is relatively deprived of NuRD components. These results confirm that Cel-MBD-2 possesses many NuRD-independent binding sites across the genome.
When we searched in C. elegans motif database based on all MBD-2 peaks, the most significantly enriched motifs are C-rich and CG-rich helix-loop-helix (HLH)-family motifs (Fig. 5C). When we searched in human motif database, Cel-MBD-2 is surprisingly still enriched at the CG-rich motif bound by human MBD2 despite the loss of the MBD domain in Cel-MBD-2 (Fig. 5C). On the other hand, as expected, the NuRD-overlapping MBD-2 peaks are enriched with the top NuRD component HDA-1’s enriched motif42 (a TC-rich motif, Fig. 5C).
Predominant binding of Cel-MBD-2 to genomic sites independently of the NuRD complex is interesting as Cel-MBD-2 does not possess a MBD domain, and the genome itself is unmethylated. We speculate that MBD-2 could bind to non-NuRD-overlapping targets via interactions with transcription factors or histone mark readers, potentially through its protein-interacting CC domain or IDR.
Localization Cel-MBD-2 to repressive chromatin without strong overlap with other NuRD components and transcription factors
To identify potential DNA or chromatin recruiter for MBD2, we compared our Cel-MBD-2 IP-MS data with an existing C. elegans transcription factor (TF) database, WTF2.049. We have identified a few transcription factors interacting with Cel-MBD-2, including DPL-1/Dp and STA-1/ STATs, in addition to the mentioned NuRD-interacting TF DVE-1 (Supplementary Data 7). DP or STATs are not known as interactors of MBD2/3 in humans. Intriguingly, DPL-1 is a known interactor of HDA-150 and LIN-5351 in C. elegans, likely in the context of the DRM (DP/Rb/MuvB) complex50. On the other hand, STA-1/STATs is involved in the regulation of immune response in C. elegans and humans50. STA-1 possesses a conserved coiled-coil domain52, which may interact with Cel-MBD-2’s CC domain. Indeed, STA-1 is identified as a “confidently lost” interactor of MBD-2∆CC in our IP-MS. This may implicate that STA-1 could be competing with the NuRD remodeling core (DCP-66/GATAD2A/B) for MBD-2’s interaction through its CC.
However, published ChIP-seq data of these transcription factors show that their peaks have little overlap with non-NuRD-overlapping MBD-2 peaks. Particularly, DVE-1 is known to be a NuRD-interacting TF thus enriched at NuRD-overlapping MBD-2 peaks (Fig. 5D). In addition, DPL-1, which is known to interact with two of the NuRD components50,51, is also enriched mainly at NuRD-overlapping MBD-2 peaks (Fig. 5D, E, upper panel). Therefore, DPL-1 and DVE-1 may represent NuRD-interacting TFs that interact with MBD-2. STA-1 only shows weak enrichment at MBD-2 peaks (Fig. 5D). STA-1 peaks overlap slightly more with NuRD-overlapping MBD-2 peaks than with non-NuRD-overlapping MBD-2 peaks (Fig. 5E, lower panel). STA-1 recruitment may only contribute partially to non-NuRD-overlapping MBD-2 peaks. Therefore, it is unlikely that these TFs are the major recruiter for non-NuRD-overlapping MBD-2 targets.
In addition, the genome-wide binding pattern of MBD-2 shows characteristic enrichment at chromosome arms (Fig. 6A), which is a signature pattern of repressive histone marks, particularly H3K9me2, H3K9me3 and H3K27me348). In addition, MBD-2 shows a relatively more uniform distribution across chromosome X, similar to H3K27me3 (Fig. 6A). MBD-2 is more depleted in the central regions of chromosomes, which are enriched with H3K4me3, H3K9ac, and H3K27ac. Such enrichment at chromosome arms is unlikely to be observed in transcription factor-mediated recruitment, such as DVE-1, DPL-1, and STA-1, which show punctuated enrichment at transcribed gene promoters across the chromosome (Fig. 6A). Such genome-wide binding pattern suggests that MBD-2 localization could be transcription factor-independent, but related to local histone modification.

A Genome browser view showing log-transformed enrichment of MBD-2, histone marks, NuRD complex components, and transcription factors based on normalized read signals in the ChIP sample over the input, along all six C. elegans chromosomes. B MBD-2 and histone modification ChIP-seq read count is normalized by count per million (CPM) for both ChIP and input. The ratios of the normalized read counts of ChIP vs. input are z-transformed and plotted as heatmap. The z-score of all the samples at NuRD-overlapping and non-NuRD-overlapping MBD-2 peak centers ± 3 kb are shown. Scale bars of the heatmaps depict the z-max and z-min of all panels. Green color depicts active histone modifications, while red color indicates repressive histone modifications. C Heatmap illustrating the percentage of MBD-2 ChIP-seq peaks overlapping with various histone modifications (peaks with ≥ 1-bp overlap). All MBD-2 peaks, non-NuRD-overlapping and NuRD-overlapping MBD-2 peaks were plotted. n = total number of peaks. (D) Genome browser view showing examples of non-NuRD-overlapping MBD-2 and NuRD-overlapping MBD-2 binding regions on Chromosome X arm. Non-NuRD-overlapping MBD-2 peaks are enriched with silent histone modifications, H3K27me3, H3K9me2, and H3K9me3. On the other hand, NuRD-overlapping MBD-2 peaks are enriched with active histone modifications, H3K4me3 and H3K27ac (more examples in Supplementary Fig. 8D).
Non-NuRD-overlapping MBD-2 peaks are enriched with silent histone modifications, particularly H3K27me3 and H3K9me2 (Fig. 6B–D, Supplementary Fig. 8B, D). Forty-six percent (45.7%) and forty-one percent (41.1%) of the non-NuRD-overlapping MBD-2 peaks overlap with H3K27me3 and H3K9me2, respectively (Fig. 6C and Supplementary Data 13). Examples of non-NuRD-overlapping MBD-2 peaks and H3K27me3 peaks colocalization are also observed in genome browser (Fig. 6D, Supplementary Fig. 8D). Interestingly, H3K27me3 enrichment is exclusively at non-NuRD-overlapping MBD-2 peaks, and the binding patterns of them are similar (Fig. 6B–D, Supplementary Fig. 8D). On the other hand, H3K9me2 and H3K9me3 peaks are low in intensity and evenly distributed between non-NuRD and NuRD-overlapping regions (Fig. 6B–D, Supplementary Fig. 8D).
In contrast, NuRD-overlapping MBD-2 peaks are enriched with active histone modifications, including H3K4me3, H3K9ac and H3K27ac (Fig. 6B–D, Supplementary Fig. 8D). Our MBD-2 ChIP-seq data have confirmed the conservation of the enrichment of NuRD complex on active histone marks, consistent with the evidence by CHD-3 and LET-418 ChIP-seq data20. About eighty-seven percent (86.6%) of NuRD-overlapping MBD-2 peaks overlap with H3K27ac peaks (Fig. 6C), and NuRD-overlapping MBD-2 peaks frequently coincide with the “valleys” at H3K27ac peak centers (Figs. 6B, D, 8A, Supplementary Fig. 8D), consistent with NuRD complex enrichment at the nucleosome-depleted region of active promoters near TSS20.
When looking at the k-means clustered MBD-2 peaks based on HDA-1 and CHD-3 signals, Cluster 1 (15.24% of all MBD-2 peaks) is relatively enriched with NuRD components and active histone marks (H3K4me3 and H3K27ac), and Cluster 2 (84.76% of all MBD-2 peaks) is relatively deprived of NuRD components and enriched with H3K27me3 (Supplementary Fig. 8C). Consistently, we classified MBD-2 peaks based on whether they overlap with H3K27me3. Non-H3K27me3-overlapping MBD-2 peaks are enriched with NuRD components, while H3K27me3-overlapping MBD-2 peaks are not (Supplementary Fig. 9A). Reciprocally, MBD-2 is enriched at most H3K27me3 and H3K9me2 peaks, and partially on some H3K9me3 peak centers (Supplementary Fig. 9B–D). These features indicate the divergent epigenetic properties of MBD-2 non-NuRD-overlapping and NuRD-overlapping peaks.
NuRD-overlapping or MBD-2-activated gene targets are more highly expressed, and MBD-2 shows different binding patterns for gene repression and activation
In view of the divergent epigenetic properties of non-NuRD-overlapping and NuRD-overlapping targets, we perform gene-wise analysis of expression levels. We focused on the protein-coding genes identified in our RNA-seq analysis (10,779 genes with a non-zero read count in at least one sample (N2, P0 or F1 mbd-2-/-) that contain these MBD-2 peaks within 2 kb upstream or downstream of their transcription start sites (TSS), which usually covers the gene body. MBD-2 binds to 9262 protein-coding genes, almost half of the C. elegans genes (Supplementary Data 9). Interestingly, only around half of the differentially expressed genes in P0 and F1 mbd-2-/- RNA-seq are bound by MBD-2 at their transcription start site (TSS) ± 2 kb based on our MBD-2 ChIP-seq (Supplementary Fig. 10A, B). Surprisingly, the differentially expressed genes in P0 (1.03-fold; hypergeometric p-value = 0.00076) and F1 (1.01-fold; hypergeometric p-value = 0.23) are not enriched for MBD-2 binding, compared to expectation. This may suggest that MBD-2 binding at genes has unknown functions that cannot be revealed by gene dysregulation. Moreover, the remaining half of the differentially expressed genes unbound by MBD-2 could be its indirect or secondary targets.
We classified MBD-2 target genes based on the presence of non-NuRD-overlapping or NuRD-overlapping MBD-2 ChIP-seq peaks (peaks classified as in Fig. 5A), resulting in three categories (Fig. 7A), including: (1) 646 genes that have both NuRD-overlapping and non-NuRD-overlapping peaks near the TSS, (2) 9756 genes that exclusively only have non-NuRD-overlapping MBD-2 peaks near the TSS, and (3) 377 genes that exclusively only have NuRD-overlapping MBD-2 peaks near the TSS. We focus on the (2) exclusive non-NuRD-overlapping and (3) exclusive NuRD-overlapping MBD-2 gene targets for the following expression analysis.

A Venn diagrams depicting overlaps of genes identified from ChIP-seq data (single-end reads with a non-zero read count in at least one RNA-seq sample (N2, P0 or F1 mbd-2-/-) with non-NuRD-overlapping MBD-2 peaks (pink) and NuRD-overlapping MBD-2 peaks (red) at their TSS ± 2 kb, dysregulated genes ( ≥ 2 fold and Benjamini-adjusted two-tailed Wald test p-values < 0.05), either upregulated (purple) or downregulated (green) in P0 mbd-2-/- RNA-seq. Exclusive non-NuRD-overlapping and exclusive NuRD-overlapping MBD-2 gene targets are defined by genes with only non-NuRD-overlapping and NuRD-overlapping MBD-2 peaks, respectively. B, C Box-and-whiskers plots with median (middle line), 25th, 75th percentile (box) and minima, maxima (whiskers) depicting transcript per million (TPM) of protein-coding genes in wildtype (N2) worms with a non-zero read count in at least one RNA-seq sample (N2, P0 or F1 mbd-2-/-). B TPM of all protein-coding genes, exclusive non-NuRD-overlapping and exclusive NuRD-overlapping MBD-2 gene targets, C TPM of all protein-coding genes, genes with either exclusive non-NuRD-overlapping and exclusive NuRD-overlapping MBD2 peaks, which has either no change, downregulated or upregulated in P0 mbd-2-/-. Genes with TPM ≥ 500 are flattened on the red dotted line. The two-tailed t-test p-value of the TPM difference in different MBD-2 target types (exclusive non-NuRD-overlapping or exclusive NuRD-overlapping, and P0 mbd-2-/- upregulated or downregulated) versus all protein-coding genes was indicated on top. n = number of genes. Source data of B and C are provided as Source Data files. D, E Metaplot showing the binding patterns of MBD-2 (count per million (CPM) normalized reads over input) at its gene targets. All left panels show the binding pattern with TSS as the reference point ( ± 2 kb), while all right panels show the binding pattern across the gene, with a 1500-bp scaled region from TSS + 300 bp to TES-50 bp. (TSS-2 kb to TSS + 300 bp, and TES-50 bp to TES + 500 bp are excluded from scaling.) D MBD-2 binding pattern on non-NuRD-overlapping (blue) and NuRD-overlapping (green) targets. E MBD-2 binding pattern on genes upregulated (blue) and downregulated (green) ≥ 5-fold in P0 mbd-2-/- RNA-seq.
When looking at the expression levels of MBD-2 gene targets in wildtype (N2) worms, the median native expression level of the exclusive NuRD-overlapping MBD-2 gene targets is higher than that of the exclusive non-NuRD-overlapping MBD-2 targets and that of all protein-coding genes (Fig. 7B). We further subdivided the gene targets into upregulated and downregulated genes, and genes with unchanged expression, based on P0 mbd-2-/- RNA-seq (Fig. 7A). Surprisingly, only 23-31% of MBD-2-bound gene targets had significantly altered expression levels in both the null mutant and coiled-coil domain deletion mutant, no matter in the exclusive NuRD-overlapping MBD-2 gene targets or the exclusive non-NuRD-overlapping MBD-2 gene targets (Fig. 7A, Supplementary Fig. 10C, D). Nevertheless, exclusive non-NuRD-overlapping MBD-2 gene targets seem to show a higher degree of dysregulation than exclusive NuRD-overlapping ones, as observed in both P0 mbd-2-/- and gfp::3xflag::mbd-2∆CC/∆CC RNA-seq (Supplementary Fig. 10E, F). These results suggest that for the majority of the MBD-2 target genes, binding of MBD-2 to them does not significantly affect their expression levels in the adult worm analyzed. Yet, other developmental stages or stressed conditions have not been tested.
Interestingly, P0 mbd-2-/--downregulated genes, which are normally MBD-2-activated genes, have a higher median expression level than the no change or upregulated genes, both in the exclusive NuRD-overlapping and exclusive non-NuRD-overlapping MBD-2 gene targets (Fig. 7C). On the other hand, MBD-2-repressed genes have a lower median expression level. This indicates that MBD-2 reinforces the activation of its highly expressed targets or repression of its lowly expressed target genes.
Next, we investigated the binding pattern of MBD-2 on its gene targets. MBD-2 generally shows sharp enrichment at the TSS of its target genes. The binding is also extended to gene bodies with lower average signal compared to the TSS (Supplementary Fig. 11A). This holds true for both the exclusive non-NuRD-overlapping gene targets and the exclusive NuRD-overlapping gene targets (Fig. 7D). The binding upstream of the TSS is slightly higher in the exclusive NuRD-overlapping gene targets ( ~ 1.35 fold enrichment over input) compared to exclusive non-NuRD-overlapping gene targets ( ~ 1.21 fold). However, the overall gene binding patterns of MBD-2 show no dependency on its interaction with NuRD.
On the P0 mbd-2-/--upregulated genes, which are normally MBD-2-repressing genes, the average MBD-2 binding peak is at the TSS (Fig. 7E). In contrast, on the P0 mbd-2 -/--downregulated genes, which are normally MBD-2-activated genes, the average MBD-2 binding peak is more downstream of the TSS. It appears that the localization of MBD-2 at the gene promoter and TSS may be associated with repression of target genes, whereas MBD-2 at the gene body may promote gene activation. On the other hand, MBD-2 binding pattern on F1 mbd-2-/--downregulated genes is less distinct, and may represent secondary effects (Supplementary Fig. 11B). These analyses show that MBD-2 has different binding patterns at its repressing and activating targets.
H3K27me3 correlates specifically with non-NuRD-overlapping MBD-2 gene targets, but not with NuRD-overlapping MBD-2 gene targets
Consistently, active histone marks, including H3K4me3, H3K9ac, H3K27ac and H3K36me3, are more enriched at the gene body of exclusive NuRD-overlapping MBD-2 gene targets compared to exclusive non-NuRD-overlapping MBD-2 gene targets (Fig. 8A). This is consistent with NuRD’s tendency to bind to open chromatin regions marked with active histone marks, which likely include active genes. In contrast, repressive histone marks, including H3K9me2, H3K9me3, and H3K27me3, are more enriched at exclusive non-NuRD-overlapping genes (Fig. 8A).

A Metaplot showing the binding patterns of MBD-2, NuRD components and histone modifications (count per million (CPM) normalized reads over input) across MBD-2 gene targets, with a 1500-bp region from TSS to TES. 2 kb upstream of TSS and 500 bp downstream of TES are also displayed, all the regions are not scaled. Blue lines depict enrichment at 9756 exclusive non-NuRD-overlapping MBD-2 gene targets and green lines depict 377 exclusive NuRD-overlapping MBD-2 gene targets (as defined in Fig. 7A). B, C Heatmap depicting Spearman’s correlation coefficient of MBD-2, NuRD components and histone modifications on (B) exclusive non-NuRD-overlapping MBD-2 gene targets with MBD-2 peaks that do not overlap with H3K36me3 and (C) exclusive NuRD-overlapping MBD-2 gene targets. The color scale indicates the Spearman’s correlation coefficient.
Combining with the P0 mbd-2-/- RNA-seq data, we found that in wildtype, H3K9ac and H3K4me3 are more enriched at exclusive NuRD-overlapping MBD-2 gene targets that are downregulated, whereas H3K27ac and H3K36me3 are more enriched at MBD-2 gene targets with unchanged gene expression (Supplementary Fig. 12).
In agreement with our results that non-NuRD-overlapping MBD-2 peaks are enriched with H3K27me3 (Fig. 6B), H3K27me3 is enriched in the exclusive non-NuRD-overlapping MBD-2 gene targets, regardless of being upregulated or downregulated in P0 mbd-2-/- (Supplementary Fig. 12). However, H3K27me3 is relatively low in the exclusive non-NuRD-overlapping MBD-2 gene targets that have unchanged expression in P0 mbd-2-/- and exclusive NuRD-overlapping MBD-2 gene targets (Supplementary Fig. 12). H3K27me3 may cooperate with MBD-2 in the transcriptional regulation of non-NuRD-overlapping gene targets. Surprisingly, H3K36me3, which usually localizes at gene bodies, is enriched at MBD-2 gene targets with unchanged expression in P0 mbd-2-/- (Supplementary Fig. 12). Such an enrichment pattern of H3K36me3 seems to be opposite to that of H3K27me3 at MBD-2-regulated gene targets.
Consistent with the arm enrichment (Fig. 6A), MBD-2 exhibits a general positive correlation with repressive histone marks H3K9me2 and H3K9me3, irrespective of whether the target genes are non-NuRD- or NuRD-overlapping. In contrast, H3K27me3 is enriched at all non-NuRD MBD-2 peaks, except at locations where H3K36me3 is enriched (Fig. 6B). Indeed, H3K27me3 and H3K36me3 show a negative correlation (−0.67) at MBD-2 exclusive non-NuRD-overlapping gene targets (Supplementary Fig. 13A). When excluding the 3,913 non-NuRD-overlapping MBD-2 gene targets that have overlapping peaks with H3K36me3 (Supplementary Fig. 13B, C), we found that H3K27me3 reaches a correlation of 0.56 with MBD-2 at 5843 exclusive non-NuRD-overlapping MBD-2 gene targets (Fig. 8B). In contrast, H3K27me3 shows no significant correlation (0.01) with MBD-2 at NuRD-overlapping target genes (Fig. 8C, Supplementary Fig. 13C). This finding aligns with the observation in Fig. 6B, where H3K27me3 is exclusively enriched at non-NuRD-overlapping MBD-2 peaks, but not at NuRD-overlapping MBD-2 peaks. Conversely, when excluding non-NuRD-overlapping MBD-2 gene targets that overlap with H3K27me3, the correlation of H3K36me3 with MBD-2 increases from 0.03 to 0.31 (Supplementary Fig. 13D). Therefore, at non-NuRD-overlapping MBD-2 gene targets, the mutually exclusive enrichment of H3K27me3 or H3K36me3 could be related to its different gene regulatory functions (Supplementary Fig. 12).
These results suggest that while MBD-2 generally colocalizes with repressive histone marks, H3K27me3 is particularly distinctive in differentiating between its non-NuRD-overlapping and NuRD-overlapping gene targets. Further studies are needed to identify MBD-2’s recruiter(s) to repressed regions, particularly to elucidate the causal relationship between the enrichment of H3K27me3 and MBD-2 at its non-NuRD-overlapping gene targets.
Discussion
In this study, we have determined C. elegans MBD-2’s function in development and reproduction through characterization of mbd-2 null mutant and deletion of the coiled-coil domain. We have verified Cel-MBD-2 as a scaffolding NuRD complex component, with its coiled-coil domain required to connect with the nucleosome remodeling core. We have discovered that at least a quarter of C. elegans protein-coding genes are dysregulated in mbd-2 null mutant, suggesting MBD-2 is a crucial transcriptional regulator for both repression and activation. Particularly, MBD-2-activated genes have a relatively high median expression levels, as compared to all genes, in wildtype.
We have discovered dual binding patterns of MBD-2 on the genome. Specifically, NuRD-overlapping MBD-2 peaks are enriched with H3K4me3 and H3K27ac, whereas non-NuRD-overlapping MBD-2 peaks overlap extensively with H3K27me3. Such dual binding patterns of Cel-MBD-2 seem to resemble those of mammalian MBD2, and surprisingly do not match with those of mammalian MBD3, which lacks functional MBD and mostly overlaps with NuRD (Fig. 9). Our work suggests that H3K27me3 could be an alternative epigenetic mark of 5mC in 5mC-free organisms. Yet, whether and how H3K27me3 is involved in Cel-MBD-2 recruitment to non-NuRD-overlapping peaks warrants further investigation.

According to previous human and mouse studies, most MBD3 binding regions overlap with MBD2 and NuRD, but there are many unique MBD2 binding sites of high 5mC density (left). In a cytosine methylation-free context in C. elegans, we propose a model for the distribution of Cel-MBD-2 (right). Despite the absence of methylation in the C. elegans genome and the lack of methyl-binding domain in Cel-MBD-2, the dual binding patterns of Cel-MBD-2 are observed as in mammalian MBD-2, in which a small proportion of MBD-2 binds to open chromatin in association with NuRD complex and a large proportion binds to silent regions enriched with H3K27me3 and H3K9me2/3, independent of NuRD complex. The organism icons are created in BioRender. Zheng, C. (2025) https://BioRender.com/rfw8dhr.
mbd-2 null mutant phenotypes, such as reduced brood size, have been similarly observed in the mutant of the NuRD nucleosome remodeling core components, chd-3 and let-41853. The authors attributed the brood size reduction to their functions in maintaining a repressive chromatin environment for repairing meiotic double-strand breaks (DSBs)53. Based on our RNAi by injection at L4 stage, we found that the fertility of injected P0 mothers was surprisingly unaffected, in contrast to the affected F1 (Supplementary Fig. 3B). This difference suggests Cel-mbd-2 may be important for fertility during early development and before gametogenesis in L4 stage. Although vulval defects and bagging can also lead to reduced brood size, F1 mbd-2-/- mutant does not have bagging phenotype, but no embryos can be observed inside the body. Therefore, the sterility may not be caused by bagging.
let-41854, hda-154 mutants, F1 mbd-2-/- and F1 gfp::3xflag::mbd-2∆CC/∆CC mutants exhibit multivulva phenotype (Fig. 2E), suggesting that NuRD represses vulval induction signals. A gain-of-function of LIN-12/Notch, downstream of the Ras signaling pathway, is sufficient to induce the multivulva phenotype55. LET-418, CHD-3 and HDA-1 have been proposed to repress Notch signaling to prevent ectopic induction of vulva precursor cells56,57. Our RNA-seq data reveal lin-12 upregulation in both P0 and F1 mbd-2-/- and gfp::3xflag::mbd-2∆CC/∆CC (Supplementary Data 3-6), supporting that MBD-2, as part of NuRD, represses Notch signaling.
However, while MBD-2 affects the expression of over 5000 genes, only up to 31% of the MBD-2-bound gene targets are dysregulated in adults (Fig. 7A, Supplementary Fig. 10C, D). This is puzzling, but may suggest that Cel-MBD-2 fine-tunes the transcription of its gene targets instead of solely activates or represses, or that the dysregulation may occur in other developmental stage or untested conditions. Indeed, studies in mouse embryonic stem cells showed that although the NuRD complex is essential for stem cell differentiation, most of the dysregulated genes are up or downregulated by less than 2-fold upon NuRD component depletion18,58,59.
A prior study in C. elegans could not detect MBD-2 by Co-IP of LIN-53, LET-418, and CHD-3, components of both NuRD subcomplexes44. The absence of MBD-2 in earlier NuRD Co-IPs was attributed to MBD-2’s purported loss of function in recruiting NuRD to methylated DNA in the 5mC-free genome44, but can be related to different tagging strategies. Our MBD-2 IP-MS data have verified MBD-2 as a conserved NuRD subunit. Our findings reveal that MBD-2 retains its conserved scaffolding role even without MBD and cytosine methylation. This aligns with the essentiality of the NuRD complex across metazoans, from sponges to humans. We detected most of the NuRD subunits in our young adult Cel-MBD-2 IP-MS, except for LIN-40/MTA1/2/3 (Fig. 4B, D, Supplementary Data 7). This might indicate that MBD-2 and LIN-40 bind only weakly or transiently. Under our native IP condition without fixation, the interaction could be lost during the protein purification process.
MBD-2∆CC IP-MS further demonstrated that MBD-2’s coiled-coil domain is critical for tethering the NuRD remodeling core to the histone deacetylase core, mirroring mammalian NuRD architecture. This CC domain-driven interaction confirms Cel-MBD-2’s role as a conserved scaffold, independent of DNA methylation. Although the IDR domain also shares some conservation among humans and C. elegans (Fig. 1A, blue box), Cel-MBD-2 IDR was previously shown to lack a highly conserved “SIFKQPV” sequence (Fig. 1A, gray box within blue box), which is retained in Drosophila and human MBD2/310. The role of the IDR in Cel-MBD-2 needs to be further studied.
Based on the lack of MBD in C. elegans MBD-2, we originally expected that its binding patterns would be similar to those of mammalian MBD3, which has a mutated MBD and binds mainly with NuRD18. Surprisingly, our study has discovered that Cel-MBD-2 binds majorly to non-NuRD-overlapping sites (93%) (Fig. 5A, Supplementary Data 12), similar to that of mammalian MBD2 that possess NuRD-independent binding to methylated genes4.
In organisms with 5mC methylation, 5mC at transposons and gene promoters is associated with gene silencing, while methylation at gene bodies is associated with stable and active housekeeping gene transcription activity, with a proposed function in reducing transcriptional noise60,61. For example, X-inactivated genes have methylated promoters, whereas genes that escape from X-inactivation have gene body methylation62. In C. elegans, we have also observed that MBD-2 predominantly binds at the TSS in MBD-2-repressing genes, whereas MBD-2 binds more downstream of the TSS in MBD-2-activiating genes (Fig. 7E).
While mammalian MBD3 is essential for stem cell differentiation and development2, Cel-MBD-2’s essential role in fertility in C. elegans is potentially contributed by its conserved scaffolding function in the NuRD complex, through regulation of ~87 exclusive NuRD-overlapping MBD-2 targets dysregulated in P0 mbd-2 null mutant. On the other hand, mammalian MBD2 knockout is not lethal but leads to behavioral phenotype5,6. There are over 2800 exclusive non-NuRD-overlapping Cel-MBD-2 gene targets with dysregulation in P0 mbd-2 null mutant (Fig. 7A). Whether the regulation of non-NuRD-overlapping Cel-MBD-2 gene targets is essential or mainly affects nervous system development (the most enriched GO in upregulated genes) to confer survival advantages is unknown. Further investigation is required to understand the function of 5mC-independent and non-NuRD-overlapping MBD-2 gene targets.
Theoretically, analysis of MBD-2∆CC may provide a glimpse of MBD-2 binding without NuRD remodeling core and potential separation of function. However, RNA-seq analysis of the mbd-2∆CC/∆CC mutant found that deletion of the coiled-coil domain causes similar gene dysregulation as the mbd-2 null mutant (Fig. 3F, G, Supplementary Fig. 4G, 5A–F). The number and fold change of dysregulated non-NuRD-overlapping and NuRD-overlapping genes are similar in mbd-2-/- and mbd-2∆CC/∆CC (Supplementary Fig. 10C–F). This unexpected result suggests that the CC domain may not only bind to NuRD remodeling core, but also contribute to non-NuRD-overlapping MBD-2 binding and gene regulation. Alternatively, MBD-2 may bind to these non-NuRD-overlapping gene targets to fine-tune their expression. Thus, the minor transcriptional changes in these genes among wildtype, null mutation, or ∆CC may not be detected in the RNA-seq approach.
Further cross-metazoan comparisons will determine whether non-NuRD-overlapping MBD2 binding is a conserved feature in other 5mC-free organisms, and whether the non-NuRD-overlapping MBD2-bound genes and regulated genes have conserved, common functions.
The relationship of mammalian MBD2 with histone modifications is ambiguous3,4,6,23. A study found that the majority of regions bound by MBD2 are enriched with the repressive histone mark H3K9me3 in mouse hippocampus6, but the same could not be observed in MBD2 ChIP-seq studies in HeLa3 and MCF-7 (breast cancer) cells23. There seems to be no consensus on whether repressive histone marks are enriched in mammalian MBD2 binding sites.
Moreover, the relationship between 5mC DNA methylation and histone modifications, particularly the repressive histone mark H3K27me3, in mammals remains controversial. For example, a previous study found that H3K27me3 colocalizes extensively with 5mC outside of CpG islands in both human and mouse cells but is mutually exclusive in CpG islands63. However, a more recent study reported that H3K27me3 negatively regulates 5mC methylation via inhibiting H3 ubiquitination64.
Our observation on the enrichment of non-NuRD-overlapping MBD-2 peaks in H3K27me3 peaks is surprising, and it is tempting to speculate H3K27me3 is the “surrogate” epigenetic mark for 5mC based on their shared properties. Firstly, both 5mC and H3K27me3 are plastic epigenetic repressive marks, in which 5mC and H3K27me3 may be added or removed during different stages of development, responses to environmental stimulus and cancer metastasis. For instance, H3K27me3 marks bivalent genes that are repressed in the early stages of development. H3K27me3 on these genes are gradually removed, allowing cells to achieve terminal differentiation and maturation with age65. Moreover, there is evidence of H3K27me3 antagonizing DNA methylation in plants and mammals66,67,68,69,70,71. A triple knockout of DNA methyltransferases (DNMTs) in mouse embryonic stem cells (mESCs) has led to a gain of new H3K27me3 peaks across the genome72. Interestingly, 99% of these genes that have gained new H3K27me3 marks are methylated at the promoters in wildtype72. Such observations point to the possibility that when 5mC is lost in some species, the genome is more available for H3K27me3 deposition, possibly without hindrance from 5mC71. However, the lethality of the DNMT1/3a/3b knockout in mice73 suggests that the new H3K27me3 binding cannot fully compensate for the loss of 5mC.
Our study, together with others, raises an interesting question of whether 5mC-free animals have a higher proportion of genome marked by H3K27me3 as compared to that in methylated genomes, and this will require further cross-species evolutionary investigation. Alternatively, it could be possible that the proportion of genome marked with 5mC and H3K27me3 are conserved across species, and ancestral MBD-2/3 may show affinity to both 5mC and H3K27me3. In organisms that lose 5mC, MBD-2/3 could bind only to the retained H3K27me3 marks.
To maintain Cel-MBD-2 dual binding patterns in a 5mC-free organism, MBD-2 has to acquire non-canonical DNA binding mechanisms, such as possessing a novel DNA-recognizing domain or histone-binding domain itself, or interacting with a DNA- or chromatin-binding partner other than the NuRD components. As Cel-MBD-2 does not contain DNA- or histone-binding domains, but only the IDR and CC domain for protein interactions, Cel-MBD-2 is likely to be recruited to the DNA through interactions with transcription factors or histone mark readers.
Particularly, we could focus on those interactors that rely on MBD-2’s CC for interaction in IP-MS for future studies on the NuRD-independent recruiter of Cel-MBD-2. Since coiled-coil deletion phenocopies null mutation and drives similar transcriptomic dysregulation, MBD-2’s CC may contribute to the recruitment of non-NuRD-overlapping genes. It is further supported by the fact that the proportion of dysregulated genes in P0 mbd-2-/- overlapping with exclusive non-NuRD and NuRD-overlapping genes are highly similar to that of P0 gfp::3xflag::mbd-2∆CC/∆CC (Supplementary Fig. 10C, D). This suggests that the non-NuRD binding of MBD-2 could be contributed by coiled-coil interaction with certain chromatin factors. If this is the case, such recruiter may compete with NuRD complex for coiled-coil interaction with MBD-2, thus recruiting MBD-2 independently of the NuRD-overlapping binding sites. Indeed, the 5mC-mediated non-NuRD binding could compete with NuRD-dependent binding in mammals. This is supported by the fact that only MBD-2b, the isoform with a truncated N terminus GR rich domain and thus have minimal 5mC affinity, but not MBD-2a, the full length isoform with the highest 5mC affinity, could rescue MBD3 knockout in mouse embryonic stem cells2.
Future studies are needed to define the mechanism of MBD-2 genomic binding pattern. The localization of Cel-MBD-2 to repressive histone marks suggests that binding through a histone mark reader would be probable. MBD-2 recruitment to non-NuRD-overlapping regions could be related to interactions with repressive histone modifications, especially H3K27me3, which displays exclusive enrichment at non-NuRD-overlapping peaks. In our MBD-2 IP-MS, we did not identify any components of the Polycomb Repressive Complex (PRC2) components, which are known to be both the reader and writer of H3K27me374. However, LIN-53 in the NuRD histone deacetylase subcomplex has been predicted to interact with MES-3 in PRC275. We have identified potential MBD-2 interacting partners with DNA- or chromatin-binding activities, including TTLL-12, HAM-3/3/SMARCD1, and PME-1/PARP1. TTLL-12, tubulin tyrosine ligase-like, is the only MBD-2 interactor with a SET-like domain76, which has been implicated in histone lysine methylation. Yet, a previous study found that although human TTLL12 could bind to H3 and H4K20me3, its SET domain could not catalyze histone methylation in vitro77. HAM-3/SMARCD1-3 is an accessory subunit of the SWI/SNF chromatin remodeling complex78. HAM-3 is involved in the regulation of embryonic development, vulva specification, and intestinal cell proliferation78. Interestingly, PME-1/PARP1 is a MBD-2 interactor that is confidently lost in MBD-2∆CC IP-MS. PME-1/PARP1 has N-terminal zinc finger domains for DNA interaction and a C-terminal catalytic domain for protein poly ADP-ribosylation (PARylation)79. In humans, PARP1’s PARylation targets include histones, and PME-1/PARP can bind chromatin directly80.
In summary, the MBD-2 binding patterns with and without NuRD, the MBD-2 binding patterns within the gene, and the epigenetic histone mark association altogether help to explain the gene regulatory functions of MBD-2. The exclusive colocalization of MBD-2 with H3K27me3 at non-NuRD-overlapping gene targets supports that H3K27me3 maybe a possible candidate histone mark that confer NuRD-independent MBD-2 recruitment, similar to the role of 5mC methylation in mouse and humans (Fig. 9). Based on our MBD2 ChIP-seq data in C. elegans, as a representative model in invertebrates and 5mC-free genomes, our work has provided insights into alternative epigenetic mechanisms in DNA methylation-free genomes.
Methods
Worm maintenance
List of worm strains used in this study is available in Supplementary Data 1. All the newly constructed strains were outcrossed with N2 or hT2 balancer-containing males for at least 3 times. All experiments were then performed on young adult worms (3 days post-hatching) obtained by bleach synchronization. Briefly, bleach was used on adult worms to kill the worms and preserve the embryos. Upon hatching in M9 buffer, L1 larvae were fed with OP50 E. coli and grown on standard conditions at 20 °C unless specified.
Worm strain construction
Cas9-mediated tagging of GFP on MBD-2
The tagging of Cel-MBD-2 with GFP::[TEV]::3xFLAG N-terminal tag (strain identifier: WYY49; see Supplementary Data 1) was performed according to a published protocol81. Briefly, the plasmids PDD282 (with the tag and homology arms flanking MBD-2 transcription start site) and PDD162 (containing the Cas9 and sgRNA for MBD-2) were injected along with co-injection markers into the gonads of wildtype (N2) young adults. Successful integration of the tag was verified with live imaging of GFP expression, single worm PCR, Sanger sequencing, and Western blot. Sequences of primers (for amplifying the homology arms from genomic DNA) and sgRNA are provided in Supplementary Data 2.
Cas9-mediated mbd-2 loss-of-function, null mutation with premature stop codons
Insertion of stop codon cassette was performed as described31. Specifically, the cassette inserted contains stop codons in all three reading frames, and the cassette was inserted in the first exon of mbd-2. Using the co-CRISPR strategy, dpy-10 allele was used as the CRISPR positive control. The repair oligos for dpy-10 and mbd-2 (containing mbd-2 homology arms flanking the stop codon cassette), crRNA (specific for the mbd-2 and dpy-10) (sequences available in Supplementary Data 2), and universal tracrRNA were injected to wildtype (N2) young adult gonads. Successful null mutants were isolated based on the dumpy phenotype caused by the dpy-10 mutation, and knock-in of the stop codon cassette (STOP-IN) at mbd-2 was confirmed by single-worm PCR.
Given the homozygous sterility of mbd-2-/- null mutants, they were maintained as hT2[bli-4(e937) let-?(q782) qIs48]-balanced heterozygous stock (WYY74, with GFP signal at pharynx). Four rounds of outcross with hT2 males were performed to remove any background mutations and the dpy-10 mutation. We picked homozygous mbd-2-/- worms (with no pharyngal GFP) from the heterozygous stock. Then, we verified the genotype by single-worm PCR using primers flanking the insertion region, followed by Sanger sequencing. Primer sequences are available in Supplementary Data 2.
Single-copy transgenic expression of GFP::3xFLAG::MBD-2, at the oxTi365 locus
A single copy of mbd-2p::gfp::3xflag::mbd-2 transgene (tg) driven by the endogenous promoter (1408 bp upstream of its start codon) was inserted ectopically into the oxTi365 locus present in the EG8082 strain using Mos1-mediated Single Copy Insertion (MosSCI)82. Specifically, gfp::3xflag::mbd-2 containing its 3’UTR and endogenous promoter was amplified from the strain with endogenous gfp::3xflag::mbd-2 (WYY49) using single-worm PCR and cloned into the pCFJ150 vector (containing flanking sequence for oxTi365 locus to allow insertion via homologous recombination) using Gibson Assembly. The cloned vector was sequenced with M13F and M13R to verify correct insertion. The verified vector was then injected into the gonads of EG8082 worms. Selection for successful insertion was performed according to the protocol82. GFP::3XFLAG::MBD-2 expression was visualized by DeltaVision microscope. The strain was further crossed with mbd-2-/- null mutants at the endogenous mbd-2 locus carrying hT2 balancer (WYY74). Double homozygous mbd-2-/-; tg/tg worms were selected by nuclear GFP::3XFLAG::MBD-2 expression and the presence of a PCR band corresponding to the null mutant (one of the primers is overlapping with the stop codon cassette). At least 10 worms were screened for 3 generations.
Cas9-mediated deletion of the coiled-coil domain of mbd-2
CRISPR/Cas9 was performed for the construction of the mbd-2 null mutant strain31. Fifteen (15) amino acid residues predicted to confer formation of coiled-coil domain by DeepCoil239 (MBD-2 residue 190-204) were deleted via exclusion of these sequences in the repair template for gfp::3xflag::mbd-2 (WYY49). Homozygous CC deletion was verified by single-worm PCR covering the deletion region, followed by Sanger sequencing. Primer sequences are available in Supplementary Data 2. The expression of GFP::3xFLAG::MBD-2∆CC was verified by live imaging and Western blot.
Live imaging of GFP-tagged MBD-2
Preparation and mounting of GFP::3xFLAG::MBD-2 (WYY49) embryos were performed as described83. WYY49 was crossed with a strain containing mCherry-H2B (OD56), which enabled the visualization of chromatin in live-cell imaging. Time-lapse images were acquired using DeltaVision microscope at 22 °C, with a 60× oil objective. Embryos were captured in z-stacks of 12 sections at 0.2 μm intervals. Images were taken at 20-s intervals at 475-nm green (10% intensity) and 575-nm red (10% intensity) excitation filter channels; 535-nm green and 625-nm red emission filter, with exposure time of 0.05 s. For whole-worm live images, the worms were mounted in 2% sodium azide on 3% agarose pads and covered with a coverslip.
Knockdown of mbd-2
Knockdown of mbd-2 was performed according to the published protocol84. Briefly, total RNA was purified from wildtype (N2) worms. ~1.2 μg of the purified RNA was used for reverse transcription using High-capacity cDNA reverse transcription kit (Applied Biosystems, Cat. 4368813). Primers containing the T7 promoter were used to amplify the cDNA of mbd-2 covering part of exon 1 to part of the last exon (502 bp). Primer sequences are available in Supplementary Data 2. The amplified cDNA was purified with PCR Purification Kit (QIAGEN, Cat. 28106). dsRNA was synthesized with MegaScript T7 kit (Ambion, AM1334) following the manufacturer’s instructions. L4 stage N2 worms were injected with the dsRNA and singled out to new plates to allow for egg laying and brood size counting. For brood size counting, the worms were allowed to lay eggs and transferred to new plates every 24 h for 3 days. Only the F1 worms laid by injected worms between 24–48 h post-injection would be used for phenotype screening and brood size counting, as it was the time point at which the loss of GFP::3xFLAG::MBD-2 from embryonic nuclei was verified by live-cell imaging when gfp::3xflag::mbd-2 was injected with dsRNA in parallel.
RNA-sequencing of mbd-2 null and coiled-coil domain deletion mutant worms
mbd-2-/- and gfp::3xflag::mbd-2∆CC/∆CC mutant worms were maintained as hT2[bli-4(e937) let-?(q782) qIs48]-balanced heterozygous stock. At least 400 heterozygous worms were picked and bleach synchronized. Around 2500 hatched L1 larvae were then transferred onto 10 OP50 seeded plates to grow to adults. The selection of the homozygous P0 mbd-2-/- and gfp::3xflag::mbd-2∆CC/∆CC worms was achieved by first identifying the high penetrance phenotype of protruding vulva in small adults, and then verifying by the lack of the GFP in the pharynx, encoded by the balancer, under a fluorescent microscope. 100–150 P0 mbd-2-/- worms and 120-135 P0 gfp::3xflag::mbd-2∆CC/∆CC were used for RNA extraction. A portion of the P0 mbd-2-/- and P0 gfp::3xflag::mbd-2∆CC/∆CC worms were bleach synchronized to obtain homozygous F1 mbd-2-/- and F1 gfp::3xflag::mbd-2∆CC/∆CC embryos, respectively. Two biological replicates were prepared for each genotype. Only 65-90 F1 mbd-2-/-and 100-102 F1 gfp::3xflag::mbd-2∆CC/∆CC young adult worms were used due to the very low brood size of P0 mbd-2-/- and P0 gfp::3xflag::mbd-2∆CC/∆CC.
Total RNA was isolated using TRIzol-chloroform extraction, followed by isopropanol precipitation. To remove DNA, RNA was treated with TURBO DNA-free™ Kit (Ambion, Cat.#AM1907). The RNA was resuspended in 20 μl RNAse-free water. RNA quality was accessed with Bioanalyzer (Supplementary Fig. 4A, B) and concentration was measured by Qubit. A minimum amount of 196 ng of total RNA was obtained. The total RNA samples were then submitted to the Centre for PanorOmic Sciences (CPOS, HKU LKS Faculty of Medicine) for library preparation using KAPA mRNA HyperPrep Kit and Illumina pair-end sequencing using Illumina NovaSeq 6000.
RNA-sequencing analysis
Sequencing reads were filtered for adapter sequence, low quality sequence and retaining reads with read length ≥ 40 bp, using Cutadapt with parameter --max-n 0.05 and followed by fastp --unqualified_percent_limit 50 --qualified_quality_phred 12 --disable_adapter_trimming --n_base_limit 50 --length_required 40. Reads were mapped to the reference genome WBcel235 using STAR v2.7.9a (default parameters) with the following exception: --twopassMode Basic and gene and transcript counts were obtained with the built-in counting option --quantMode. Differential expression analysis was done using DESeq2 with the gene count output from STAR. Wald tests within DESeq2 were conducted to assess differential gene expression between groups. The two-tailed Wald test p-value, and the Benjamini-Hochberg adjusted Wald test p-value is displayed in Supplementary Data 3–6. Results from DESeq2 were then run through apeglm for LFC shrinkage based on effect size using the lfcShrink function from DEseq2 for volcano plot and MA plot visualization85. Differentially expressed protein-coding genes were filtered to possess Benjamini-Hochberg adjusted p-value < 0.05 and with log2 fold change (FC) shrinkage values ≥ 1.0 or ≤ -1.0. Genes with adjusted p-values of NA were excluded from downstream analysis. Gene ontology (GO) enrichment analysis was performed using DAVID. The Transcripts Per Million (TPM) value for each gene was calculated using RSEM v1.3.3 for plotting the heatmap (Supplementary Fig. 4E, F). MA plots and PCA plots are shown (Supplementary Fig. 4C, D, G).
Chromatin immunoprecipitation (ChIP)-sequencing of GFP::3xFLAG::MBD-2
ChIP-seq experiments were performed on 2 biological replicates. 3 rounds of 800 ml liquid culture was performed to obtain 1.5 ml of synchronized young adult worm pellet. The worm pellet was mixed with an equal volume of PBS supplemented with proteinase inhibitor, then snapped frozen as worm droplets by dripping into liquid nitrogen and stored at -80 °C. The frozen worm droplets were then grinded in liquid nitrogen for 10 minutes with a mortar and pestle into fine powder. The worm powder was then thawed in 3 ml of fixing solution (1.1% formaldehyde in PBS with proteinase inhibitor) and dounced with pestle B for 10 strokes to break the cuticle of the adult worms and allow penetration of the fixative. The 3 ml dounced mix was then transferred to a falcon. 2 ml of the fixing solution was used to flush the remaining dounced mix to the Falcon tube and 38 ml of fixing solution was further added. Fixation was carried out at room temperature with gentle rocking until the total fixation time reached 15 minutes. Next, the fixation was quenched by adding glycine to a final concentration of 125 mM and the sample was further rocked for 5 min at room temperature. The fixed worms were pelleted at 4000 × g, 4 °C for 5 min, and rinsed twice with ice-cold PBS supplemented with proteinase inhibitor, once with MNase buffer (NEB Cat# M0247S). The lysis and release of chromatin from worms were performed according to Tabuchi et al.86 with slight modifications. Firstly, the fixed worm pellet was partially lysed by resuspending in MNase buffer with lysis buffer (0.2% NP-40 and 0.5% sodium deoxycholate) for 5 min on ice with gentle tapping. The worm was re-pelleted at 4000 × g, 4 °C for 5 min and washed once with 1.5 ml of MNase buffer. It was then resuspended with 5 volumes of MNase buffer and snap frozen. A pilot test for MNase digestion was carried out on a small aliquot according to the manufacturer's protocol (NEB Cat# M0247S). In our hands, 37 °C for 20 min was enough to achieve mono- to di-nucleosome DNA fragment sizes. The remaining worm lysate (700-800 μl) was thawed for a larger scale MNase digestion. Then chromatin was solubilized by adding EDTA to a final 10 mM concentration, one protease inhibitor tablet (MCE Cat# HY-K0011) and Triton X-100 to a final 1% concentration, nutated at 4 °C for 2 hr. Two volumes of cold 150 mM NaCl FA buffer (1% Triton™ X-100 + 0.1% sodium deoxycholate + 0.1% sarkosyl + protease/phosphatase inhibitors) was added, followed by sonication with a Covaris M220 model (duty factor 20%, average power 8, cycles per burst 200, 60 s per cycle, total 5 cycles) to liberate chromatin. The lysate was centrifuged at 16,000 × g for 15 min at 4 °C to retrieve solubilized chromatin in the supernatant. A total of 5.25 mg total protein containing the soluble chromatin was used to perform immunoprecipitation with anti-FLAG antibody-conjugated magnetic beads (MCE Cat# HY-K0207) in the ratio of 10 μl magnetic beads per 500 μl lysate as suggested by the manufacturer. The lysate was aliquoted into 1.5 ml LoBind tubes (700 μg per tube) and the tubes were rotated with the magnetic beads overnight at 4 °C. 5% of the protein (262.5 μg) was stored as input. The washing of the beads was carried out under varying salt concentrations according to the protocol described by Askjaer et al.87. The antibody/protein/DNA complexes were then eluted by adding 125 μl fresh Elution buffer (1%SDS/0.1 M NaHCO3) and heated at 65 °C in a heat block for 15 min, and vortexed for 5 sec every 3 min. The beads were then magnetized and the eluate was retrieved into a low-binding 1.5 ml tube. The elution was repeated with 50 μl elution buffer and the eluates were combined. 1x volume of 2x elution buffer was added to the input and treated in the same condition in parallel with the immunoprecipitation samples. A Western blot was performed to verify the successful pull-down of GFP::3xFLAG::MBD-2.
RNase A (Thermo Scientific Cat# EN0531) was added in a 1:4 ratio to the volume of the ChIP-ed DNA samples and the inputs followed by incubation at 37 °C for 1 hr. 1 M TrisCl (pH 8) and 0.5 M EDTA were added to achieve a final concentration of 40 mM and 20 mM, respectively. Proteinase K (Qiagen Cat# 19131) (2.5 μl to every 150 μl) was added to eliminate the RNase and protein. Reverse crosslink was performed in 0.2 M NaCl salt at 800 rpm 65 °C, for 16 hours in the thermomixer. The DNA was purified with PCR Purification Kit (QIAGEN, Cat. 28106)87. Concentrations and quality for the input and ChIPed DNA were assessed with Qubit and the Agilent 2100 Bioanalyzer (Supplementary Fig. 8A). The samples were submitted to the Centre for PanorOmic Sciences (CPOS, HKU LKS Faculty of Medicine) for library preparation with KAPA Hyper Prep Kit and 151 bp paired-end DNA sequencing using Illumina NovaSeq 6000. We generated 20.9 and 17.3 GB raw data for the input replicate 1 and 2, respectively, and 12.9 and 11.5 GB for ChIP replicate 1 and 2, respectively.
ChIP-sequencing analysis
The reads were first filtered for adapter sequence and low-quality reads, retaining only reads with read length ≥ 30 bp. Low Quality was defined as reads with more than 5% unknown bases (“N”) and/or reads having more than 50% of bases with quality value <= 11 were filtered out. Clean reads were mapped to the reference genome WBcel235 using BWA (Version 0.7.17.5) with default parameters. Samtools view was used to remove unmapped reads and retain only properly paired mapped reads (-f 2 -F 4) with a minimum mapping quality of 30. Samtools sort was utilized to sort the alignments by coordinates. When comparing with single-end ChIP-seq data generated by other studies, including CHD-348, LET-41848, STA-188 and DPL-1 (ModENCODE) ChIP-seq, only the forward reads of the paired-end data of MBD-2 in this study, HDA-142 and DVE-142 were used for analysis for fair comparison (Figs. 5A, B, D, E, 6, 7A–C, D, 8, Supplementary Fig. 8B-D, 9, 10C, D, 12, and 13). On the other hand, when comparing the RNAseq and ChIP-seq data generated in this study, both reads of paired-end ChIP-seq data were used (Fig. 7E Supplementary Fig. 10A,B, 11A,B). For the above-mentioned ChIP-seq data, MACS2 was used to call narrow peaks with default settings except (--no model, --extsize 200, --SPMR (Save signal per million reads for fragment pileup profiles) and q-value cut-off of 0.01. For ChIP-seq data of histone marks, MACS2 was used to call broad peaks with default settings except (--no model, --extsize 200, --SPMR (Save signal per million reads for fragment pileup profiles) and q-value cut-off of 0.05. A complete list of ChIP-seq source data is available in Supplementary Data 8. Peaks are filtered for blacklisted regions supplied by ENCODE89 using bedtools. List of annotated MBD-2 paired-end peaks and single-end (using forward strand) are available in Supplementary Data 9 and 10, respectively.
The normalized input and ChIP bedgraph file generated by MACS2 with --SPMR (signal per million reads) was converted to bigwig file using bedGraphToBigWig. BigWigcompare was used to generate a BigWig file with Fold enrichment (-FE) of ChIP-seq signal over input for visualization on IGV (Fig. 6A, D and Supplementary Fig. 8D). For annotating the genes bound by MBD-2 peaks, ChIPseeker (V1.28.3) was used with default settings.
For visualization of ChIP-seq signal around transcription start sites (TSS) by plots or heatmaps, duplicated reads were marked and removed using Picard (V2.18.2) MarkDuplicates with “REMOVE_DUPLICATES=true”. Bigwig files with CPM (count per million) normalized read count was created with Deeptools bamCompare. Normalized ChIP read counts were divided by normalized input read counts to get the ratio of the number of reads. Deeptools ComputeMatrix was used to generate plots or heatmaps from the bamCompare output.
To compute correlations of MBD-2, NuRD components, and histone marks on exclusive non-NuRD- and NuRD-overlapping gene targets (as defined in Fig. 7A), we utilized DeepTools’ multiBigwig function. We calculated the Spearman’s correlation coefficient using previously generated BigWig files (SPMR-normalized ChIP signal fold enrichment over input) within the specified non-NuRD and NuRD-overlapping gene body.
Native co-immunoprecipitation (Co-IP) and immunoprecipitation-mass spectrometry (IP-MS) of GFP::3xFLAG::MBD-2
Ten 15-mm plates of mixed stage worms were bleached, followed by two rounds of 800 ml liquid culture were performed to obtain 0.5 ml of synchronized young adult worm pellet. The pellet was collected by washing three times with ice cold M9 and once with ice cold Co-IP lysis buffer (50 mM HEPES (pH 7.4), 1 mM EGTA, 1 mM MgCl2, 100 mM KCl, 10% glycerol, 0.05% NP-40, supplemented with proteinase inhibitor) according to a previous protocol90 and snapped frozen in liquid nitrogen. The lysate was thawed from -80 °C on ice, and an equal volume of Co-IP lysis buffer was added and subjected to sonication for 10 cycles of 30 sec on and 30 sec off using a Diagenode Bioruptor Pico. Highly purified total protein was obtained after two rounds of centrifugation: firstly at 20,000 × g for 10 min at 2 °C, and secondly at 100,000 × g for 20 min at 2 °C90.
For the IP-MS and Co-IP experiments of GFP::3xFLAG::MBD-2 worms, 0.8 mg of the total protein from whole worm lysate was incubated with 10 μl anti-FLAG antibody-conjugated magnetic beads (MCE Cat# HY-K0207) overnight at 4 °C. The beads were washed 6 times with washing buffer (TBST: 50 mM Tris-HCl, 150 mM NaCl, 0.5% Tween-20, pH 7.4) prior to elution by boiling at 1x SDS page loading buffer. For Western blot, only half of the elution per 0.8 mg IP experiment was loaded for the IP wells, while 120 μg (30% of protein used for IP) was loaded as input in Fig. 4D. Full Western blots for Co-IP done on three biological replicates are displayed in Supplementary Fig. 7.
For IP-MS, the IP samples were run on a SDS page gel until the total immunoprecipitated proteins stacked together as a tight band at the boundary between the stacking and resolving gel. Subsequently, the gel was stained with Coomassie Blue. The total immunoprecipitated protein bands were then excised and submitted to LKS Faculty of Medicine, Proteomics and Metabolomics Core Facility (PMcore), Centre for PanorOmic Sciences (CPOS), the University of Hong Kong for liquid chromatography-mass spectrometry (LC-MS/MS) analysis, using Bruker timsTOF pro mass spectrometer. Log2 transformed intensity-based absolute quantitation of protein peptides (iBAQ) value was used to assess the absolute abundance of each protein based on their intensity (normalized to the expected number of tryptic peptides, which was affected by protein length)91. The relative iBAQ value (riBAQ) was calculated by subtracting the average log2 iBAQ of each protein in the negative control of wildtype (untagged MBD-2) samples from the average log2 iBAQ of each protein identified in GFP::3xFLAG::MBD-2 IP-MS92,93. The interactor threshold cutoff is defined by riBAQ >= 1 and a two-tailed t-test p-value < 0.05 (Supplementary Data 7).
For the IP-MS of GFP::3xFLAG::MBD-2∆CC, ten 15-mm plates of mixed stage worms were bleached, followed by two rounds of 800 ml liquid culture. The worms were collected by centrifugation and transferred onto plates. 1000-2000 synchronized young adult worms were manually picked based on the protruding vulva and lack of balancer gfp into a LoBind tube containing 20 μl of cold M9. The selection of the homozygous ∆CC worms was achieved by identifying the high penetrance phenotype of protruding vulva in small adults, and verifying the lack of the GFP under a fluorescent microscope. After the worms had sunk to the bottom of the LoBind tube, the M9 was replaced with ice cold Co-IP lysis buffer and snapped frozen. Multiple rounds of worms picked in this approach were pooled together after thawing on ice, followed by measuring of protein concentration with a total of 280 and 490 μg for replicate 1 and 2, respectively. Co-IP lysis buffer was added to top up to 300 μl. Sonication and immunoprecipitation were performed with the same procedure as wildtype MBD-2. Same as the GFP::3xFLAG::MBD-2 IP-MS, riBAQ was calculated by subtracting the wildtype (untagged MBD-2) samples. The immunoprecipitation efficiency for the coiled-coil deletion mutated protein (MBD-2∆CC) (log2 iBAQ = 12.80) was lower than for the full-length MBD-2 protein (log2 iBAQ = 17.32). The log2 iBAQ difference is 4.52 (see Supplementary Data 7). To prevent this difference from being misinterpreted as a loss of specific protein interactions, a normalization step was performed before plotting Fig. 4E. The negative control (on the X-axis) log2 iBAQ values of each interactor were subtracted by the immunoprecipitation efficiency difference (4.52). For defining “confidently lost interactors” (35 interactors highlighted in gray in Supplementary Data 7), we select the criteria of (1) being significant interactors in GFP::3xFLAG::MBD-2-expressing strain (defined as above), (2) with relative log2 iBAQ (log2 iBAQ in GFP::3xFLAG::MBD-2-expressing strain over that in the negative control) greater than the MBD-2 IP efficiency difference between GFP::3xFLAG::MBD-2 and GFP::3xFLAG::MBD-2∆CC (4.52), and (3) being completely undetectable in both replicates of ∆CC IP-MS.
Statistics and reproducibility
The specific statistical tests used for data analysis are detailed in the corresponding figure legends. No statistical method was used to predetermine the sample size. Sequencing and IP-MS results are based on at least two biological replicates and associated significance values. For the Western blot, three biological replicates are included. For brood size counting and phenotype scoring, experiments were conducted with independent C. elegans animals with the indicated number of replicates (n).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The RNA-seq data generated in this study have been deposited in NCBI Gene Expression Omnibus (GEO) database under accession codes GSE290148. The ChIP-seq data generated in this study have been deposited in NCBI Gene Expression Omnibus (GEO) database under accession codes GSE290147. Source data are provided with this paper.
References
Hendrich, B. et al. Genomic structure and chromosomal mapping of the murine and human Mbd1, Mbd2, Mbd3, and Mbd4 genes. Mamm Genome 10, 906–912 (1999).
Schmolka, N. et al. Dissecting the roles of MBD2 isoforms and domains in regulating NuRD complex function during cellular differentiation. Nature Communications 14, 3848 (2023).
Günther, K. et al. Differential roles for MBD2 and MBD3 at methylated CpG islands, active promoters and binding to exon sequences. Nucleic Acids Res 41, 3010–3021 (2013).
Baubec, T., Ivánek, R., Lienert, F. & Schübeler, D. Methylation-dependent and -independent genomic targeting principles of the mbd protein family. Cell 153, 480–492 (2013).
Hendrich, B., Guy, J., Ramsahoye, B., Wilson, V. A. & Bird, A. Closely related proteins MBD2 and MBD3 play distinctive but interacting roles in mouse development. Genes Dev 15, 710–723 (2001).
Lax, E. et al. Methyl-CpG binding domain 2 (Mbd2) is an epigenetic regulator of autism-risk genes and cognition. Translational Psychiatry 13, 259 (2023).
Knock, E. et al. The methyl binding domain 3/nucleosome remodelling and deacetylase complex regulates neural cell fate determination and terminal differentiation in the cerebral cortex. Neural Dev 10, 13 (2015).
Hirota, A., Nakajima-Koyama, M., Ashida, Y. & Nishida, E. The nucleosome remodeling and deacetylase complex protein CHD4 regulates neural differentiation of mouse embryonic stem cells by down-regulating p53. J Biol Chem 294, 195–209 (2019).
Cramer, J. M. et al. Methylation specific targeting of a chromatin remodeling complex from sponges to humans. Scientific Reports 7, 40674 (2017).
Gutierrez, A. & Sommer, R. J. Functional diversification of the nematode mbd2/3 gene between Pristionchus pacificus and Caenorhabditis elegans. BMC Genet 8, 57 (2007).
Serobyan, V. et al. Chromatin remodelling and antisense-mediated up-regulation of the developmental switch gene eud-1 control predatory feeding plasticity. Nat Commun 7, 12337 (2016).
Gutierrez, A. & Sommer, R. J. Evolution of dnmt-2 and mbd-2-like genes in the free-living nematodes Pristionchus pacificus, Caenorhabditis elegans and Caenorhabditis briggsae. Nucleic Acids Res 32, 6388–6396 (2004).
Marhold, J., Brehm, A. & Kramer, K. The Drosophila methyl-DNA binding protein MBD2/3 interacts with the NuRD complex via p55 and MI-2. BMC Mol Biol 5, 20 (2004).
Jaber-Hijazi, F. et al. Planarian MBD2/3 is required for adult stem cell pluripotency independently of DNA methylation. Developmental Biology 384, 141–153 (2013).
Christov, C. P. et al. A NuRD complex from Xenopus laevis eggs is essential for dna replication during early embryogenesis. Cell Rep 22, 2265–2278 (2018).
Low, J. K. K. et al. The nucleosome remodeling and deacetylase complex has an asymmetric, dynamic, and modular architecture. Cell Rep 33, 108450 (2020).
Millard, C. J., Fairall, L., Ragan, T. J., Savva, C. G. & Schwabe, J. W. R. The topology of chromatin-binding domains in the NuRD deacetylase complex. Nucleic Acids Res 48, 12972–12982 (2020).
Bornelöv, S. et al. The Nucleosome Remodeling and Deacetylation Complex Modulates Chromatin Structure at Sites of Active Transcription to Fine-Tune Gene Expression. Mol Cell 71, 56–72.e54 (2018).
Shimbo, T. et al. MBD3 localizes at promoters, gene bodies and enhancers of active genes. PLoS Genet 9, e1004028 (2013).
Käser-Pébernard, S., Pfefferli, C., Aschinger, C. & Wicky, C. Fine-tuning of chromatin composition and Polycomb recruitment by two Mi2 homologues during C. elegans early embryonic development. Epigenetics Chromatin 9, 39 (2016).
Ng, H. H. et al. MBD2 is a transcriptional repressor belonging to the MeCP1 histone deacetylase complex. Nat Genet 23, 58–61 (1999).
Barr, H. et al. Mbd2 contributes to DNA methylation-directed repression of the Xist gene. Mol Cell Biol 27, 3750–3757 (2007).
Menafra, R. et al. Genome-wide binding of MBD2 reveals strong preference for highly methylated loci. PLoS One 9, e99603 (2014).
Chatagnon, A. et al. Preferential binding of the methyl-CpG binding domain protein 2 at methylated transcriptional start site regions. Epigenetics 6, 1295–1307 (2011).
Dunwell, T. L. & Pfeifer, G. P. Drosophila genomic methylation: new evidence and new questions. Epigenomics 6, 459–461 (2014).
Simpson, V. J., Johnson, T. E. & Hammen, R. F. Caenorhabditis elegans DNA does not contain 5-methylcytosine at any time during development or aging. Nucleic Acids Research 14, 6711–6719 (1986).
O’Brown, Z. K. et al. Sources of artifact in measurements of 6mA and 4mC abundance in eukaryotic genomic DNA. BMC Genomics 20, 445 (2019).
Greer, E. L. et al. DNA methylation on N6-adenine in C. elegans. Cell 161, 868–878 (2015).
Rošić, S. et al. Evolutionary analysis indicates that DNA alkylation damage is a byproduct of cytosine DNA methyltransferase activity. Nat Genet 50, 452–459 (2018).
Bewick, A. J., Vogel, K. J., Moore, A. J. & Schmitz, R. J. Evolution of DNA Methylation across Insects. Mol Biol Evol 34, 654–665 (2017).
Wang, H., Park, H., Liu, J. & Sternberg, P. W. An efficient genome editing strategy to generate putative null mutants in Caenorhabditis elegans Using CRISPR/Cas9. G3 (Bethesda) 8, 3607–3616 (2018).
Garvin, C., Holdeman, R. & Strome, S. The phenotype of mes-2, mes-3, mes-4 and mes-6, maternal-effect genes required for survival of the germline in Caenorhabditis elegans, is sensitive to chromosome dosage. Genetics 148, 167–185 (1998).
Boeke, J., Ammerpohl, O., Kegel, S., Moehren, U. & Renkawitz, R. The minimal repression domain of MBD2b overlaps with the methyl-CpG-binding domain and binds directly to Sin3A. J Biol Chem 275, 34963–34967 (2000).
Leighton, G. & Williams, D. C. Jr The Methyl-CpG-binding domain 2 and 3 proteins and formation of the nucleosome remodeling and deacetylase complex. J Mol Biol 432, 1624–1639 (2020).
Leighton, G. O., Shang, S., Hageman, S., Ginder, G. D. & Williams, D. C. Analysis of the complex between MBD2 and the histone deacetylase core of NuRD reveals key interactions critical for gene silencing. Proceedings of the National Academy of Sciences 120, e2307287120 (2023).
Gnanapragasam, M. N. et al. p66Alpha-MBD2 coiled-coil interaction and recruitment of Mi-2 are critical for globin gene silencing by the MBD2-NuRD complex. Proc Natl Acad Sci USA 108, 7487–7492 (2011).
Arvindekar, S. et al. Molecular architecture of nucleosome remodeling and deacetylase sub-complexes by integrative structure determination. Protein Science 31, e4387 (2022).
Low, J. K. et al. CHD4 is a peripheral component of the nucleosome remodeling and deacetylase complex. J Biol Chem 291, 15853–15866 (2016).
Ludwiczak, J., Winski, A., Szczepaniak, K., Alva, V. & Dunin-Horkawicz, S. DeepCoil-a fast and accurate prediction of coiled-coil domains in protein sequences. Bioinformatics 35, 2790–2795 (2019).
Desai, M. A. et al. An intrinsically disordered region of methyl-CpG binding domain protein 2 (MBD2) recruits the histone deacetylase core of the NuRD complex. Nucleic Acids Research 43, 3100–3113 (2015).
Lembo, F. et al. MBDin, a novel MBD2-interacting protein, relieves MBD2 repression potential and reactivates transcription from methylated promoters. Mol Cell Biol 23, 1656–1665 (2003).
Shao, L.-W. et al. Histone deacetylase HDA-1 modulates mitochondrial stress response and longevity. Nature Communications 11, 4639 (2020).
Golden, N. L., Foley, M. K., Kim Guisbert, K. S. & Guisbert, E. Divergent regulatory roles of NuRD chromatin remodeling complex subunits GATAD2 and CHD4 in Caenorhabditis elegans. Genetics 221, iyac046 (2022).
Passannante, M. et al. Different Mi-2 complexes for various developmental functions in Caenorhabditis elegans. PLoS One 5, e13681 (2010).
Britanova, O. et al. Satb2 is a postmitotic determinant for upper-layer neuron specification in the neocortex. Neuron 57, 378–392 (2008).
Kim, H. et al. HDAC1 SUMOylation promotes Argonaute-directed transcriptional silencing in C. elegans. eLife 10, e63299 (2021).
Zhu, D. et al. NuRD mediates mitochondrial stress–induced longevity via chromatin remodeling in response to acetyl-CoA level. Science Advances 6, eabb2529 (2020).
Hou, X. et al. Systematic characterization of chromodomain proteins reveals an H3K9me1/2 reader regulating aging in C. elegans. Nature Communications 14, 1254 (2023).
Reece-Hoyes, J. S. et al. A compendium of Caenorhabditis elegans regulatory transcription factors: a resource for mapping transcription regulatory networks. Genome Biol 6, R110 (2005).
Harrison, M. M., Ceol, C. J., Lu, X., Horvitz, H. R. & Some, C. elegans class B synthetic multivulva proteins encode a conserved LIN-35 Rb-containing complex distinct from a NuRD-like complex. Proc Natl Acad Sci USA 103, 16782–16787 (2006).
Zhong, W. & Sternberg, P. W. Genome-wide prediction of C. elegans genetic interactions. Science 311, 1481–1484 (2006).
Wang, Y. & Levy, D. E. C. elegans STAT: evolution of a regulatory switch. Faseb j 20, 1641–1652 (2006).
Turcotte, C. A. et al. Maintenance of genome integrity by Mi2 homologs CHD-3 and LET-418 in Caenorhabditis elegans. Genetics 208, 991–1007 (2018).
Dufourcq, P. et al. Functional requirement for histone deacetylase 1 in Caenorhabditis elegans gonadogenesis. Mol Cell Biol 22, 3024–3034 (2002).
Deng, Y., Luo, K. L., Shaye, D. D. & Greenwald, I. A Screen of the conserved kinome for negative regulators of LIN-12 negative regulatory region (“NRR”)-Missense Activity in Caenorhabditis elegans. G3 (Bethesda) 9, 3567–3574 (2019).
von Zelewsky, T. et al. The C. elegans Mi-2 chromatin-remodelling proteins function in vulval cell fate determination. Development 127, 5277–5284 (2000).
Ranawade, A. V., Cumbo, P. & Gupta, B. P. Caenorhabditis elegans histone deacetylase hda-1 is required for morphogenesis of the vulva and LIN-12/Notch-mediated specification of uterine cell fates. G3 (Bethesda) 3, 1363–1374 (2013).
Miller, A. et al. Sall4 controls differentiation of pluripotent cells independently of the Nucleosome Remodelling and Deacetylation (NuRD) complex. Development 143, 3074–3084 (2016).
Reynolds, N. et al. NuRD-mediated deacetylation of H3K27 facilitates recruitment of Polycomb Repressive Complex 2 to direct gene repression. Embo j 31, 593–605 (2012).
Zilberman, D. An evolutionary case for functional gene body methylation in plants and animals. Genome Biol 18, 87 (2017).
Huh, I., Zeng, J., Park, T. & Yi, S. V. DNA methylation and transcriptional noise. Epigenetics Chromatin 6, 9 (2013).
Sharp, A. J. et al. DNA methylation profiles of human active and inactive X chromosomes. Genome Res 21, 1592–1600 (2011).
Brinkman, A. B. et al. Sequential ChIP-bisulfite sequencing enables direct genome-scale investigation of chromatin and DNA methylation cross-talk. Genome Res 22, 1128–1138 (2012).
Zhang, H. et al. H3K27me3 shapes DNA methylome by inhibiting UHRF1-mediated H3 ubiquitination. Sci China Life Sci 65, 1685–1700 (2022).
Voigt, P., Tee, W. W. & Reinberg, D. A double take on bivalent promoters. Genes Dev 27, 1318–1338 (2013).
Mathieu, O., Probst, A. V. & Paszkowski, J. Distinct regulation of histone H3 methylation at lysines 27 and 9 by CpG methylation in Arabidopsis. Embo j 24, 2783–2791 (2005).
Lindroth, A. M. et al. Antagonism between DNA and H3K27 methylation at the imprinted Rasgrf1 locus. PLoS Genet 4, e1000145 (2008).
Reddington, J. P. et al. Redistribution of H3K27me3 upon DNA hypomethylation results in de-repression of Polycomb target genes. Genome Biol 14, R25 (2013).
Gal-Yam, E. N. et al. Frequent switching of Polycomb repressive marks and DNA hypermethylation in the PC3 prostate cancer cell line. Proc Natl Acad Sci USA 105, 12979–12984 (2008).
de la Calle Mustienes, E., Gómez-Skarmeta, J. L. & Bogdanović, O. Genome-wide epigenetic cross-talk between DNA methylation and H3K27me3 in zebrafish embryos. Genom Data 6, 7–9 (2015).
Richard Albert, J. et al. DNA methylation shapes the Polycomb landscape during the exit from naive pluripotency. Nat Struct Mol Biol 32, 346–357 (2025).
Hagarman, J. A., Motley, M. P., Kristjansdottir, K. & Soloway, P. D. Coordinate regulation of DNA methylation and H3K27me3 in mouse embryonic stem cells. PLoS One 8, e53880 (2013).
Liao, J. et al. Targeted disruption of DNMT1, DNMT3A and DNMT3B in human embryonic stem cells. Nat Genet 47, 469–478 (2015).
Blackledge, N. P., Rose, N. R. & Klose, R. J. Targeting Polycomb systems to regulate gene expression: modifications to a complex story. Nat Rev Mol Cell Biol 16, 643–649 (2015).
Snel, B., van den Heuvel, S. & Seidl, M. F. Caenorhabditis elegans MES-3 is a highly divergent ortholog of the canonical PRC2 component SUZ12. iScience 25, 104633 (2022).
Andersen, E. C. & Horvitz, H. R. Two C. elegans histone methyltransferases repress lin-3 EGF transcription to inhibit vulval development. Development 134, 2991–2999 (2007).
Brants, J. et al. Tubulin tyrosine ligase like 12, a TTLL family member with SET- and TTL-like domains and roles in histone and tubulin modifications and mitosis. PLoS One 7, e51258 (2012).
Ertl, I. et al. Functional Interplay of Two Paralogs Encoding SWI/SNF Chromatin-Remodeling Accessory Subunits During Caenorhabditis elegans Development. Genetics 202, 961–975 (2016).
Gagnon, S. N., Hengartner, M. O. & Desnoyers, S. The genes pme-1 and pme-2 encode two poly(ADP-ribose) polymerases in Caenorhabditis elegans. Biochem J 368, 263–271 (2002).
Zong, W., Gong, Y., Sun, W., Li, T. & Wang, Z. Q. PARP1: liaison of chromatin remodeling and transcription. Cancers (Basel) 14, 4162 (2022).
Dickinson, D. J., Pani, A. M., Heppert, J. K., Higgins, C. D. & Goldstein, B. Streamlined genome engineering with a self-excising drug selection cassette. Genetics 200, 1035–1049 (2015).
Frøkjær-Jensen, C. Transposon-assisted genetic engineering with mos1-mediated single-copy insertion (MosSCI). Methods Mol Biol 1327, 49–58 (2015).
Powers, J. A. Live-cell imaging of mitosis in Caenorhabditis elegans embryos. Methods 51, 197–205 (2010).
Conte, D., Jr., MacNeil, L. T., Walhout, A. J. M. & Mello, C. C. RNA Interference in Caenorhabditis elegans. Curr. Protoc. Mol. Biol. 109, 26.23.21–26.23.30 (2015).
Zhu, A., Ibrahim, J. G. & Love, M. I. Heavy-tailed prior distributions for sequence count data: removing the noise and preserving large differences. Bioinformatics 35, 2084–2092 (2019).
Tabuchi, T. M. et al. Caenorhabditis elegans sperm carry a histone-based epigenetic memory of both spermatogenesis and oogenesis. Nature Communications 9, 4310 (2018).
Askjaer, P., Ercan, S. & Meister, P. Modern techniques for the analysis of chromatin and nuclear organization in C. elegans. WormBook, 1–35 (2014).
Tanguy, M. et al. An Alternative STAT Signaling Pathway Acts in Viral Immunity in Caenorhabditis elegans. mBio 8, e00924-17 (2017).
Amemiya, H. M., Kundaje, A. & Boyle, A. P. The ENCODE blacklist: identification of problematic regions of the genome. Sci Rep 9, 9354 (2019).
Zanin, E. et al. Affinity purification of protein complexes in C. elegans. Methods Cell Biol 106, 289–322 (2011).
Millán-Oropeza, A., Blein-Nicolas, M., Monnet, V., Zivy, M. & Henry, C. Comparison of different label-free techniques for the semi-absolute quantification of protein abundance. Proteomes 10, (2022).
Zheng, J. et al. Deciphering intercellular signaling complexes by interaction-guided chemical proteomics. Nature Communications 14, 4138 (2023).
Shin, J. B. et al. Molecular architecture of the chick vestibular hair bundle. Nat Neurosci 16, 365–374 (2013).
Sigrist, C. J. et al. New and continuing developments at PROSITE. Nucleic Acids Res 41, D344–D347 (2013).
Diesh, C. et al. JBrowse 2: a modular genome browser with views of synteny and structural variation. Genome Biol 24, 74 (2023).
Acknowledgements
We thank Teresa Lee, Michael Huen, Ming Chan, Zhongying Zhao, and Dongying Xie for critically reading earlier versions of the manuscript. We also thank Shouhong Guang for providing 3xFLAG-GFP::LET-418 and 3xFLAG-GFP::CHD-3 worms, Ying Liu for providing HDA-1::GFP worms, and Chantal Wicky for providing the anti-LET-418 antibody. We are grateful to Dongying Xie for advice on strain generation, Yick Hin Ling and Lilian Wu for advice on data analysis, and Ling Jiang for assistance on worm picking. We thank Sharma Rakesh and Toto Chu from HKU Centre for PanorOmic Sciences (CPOS) for their help on sample processing and advice on IP-MS and RNA-seq analysis. This work is supported by the Ministry of Science and Technology of the People’s Republic of China (Chinese Ministry of Science and Technology) (2021YFA0909302) received by KWYY.
Author information
Authors and Affiliations
Contributions
H.N.T. performed all the experiments, planned the study, developed methods, analyzed data, and wrote the manuscript; C.Y.Y.W. developed methods; C.G.Z. advised on analysis and revised the manuscript; K.W.Y.Y. supervised the project, planned the study, wrote the manuscript, and acquired funding for the project.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Tsui, H.N., Wong, C.Y.Y., Zheng, C. et al. Dual genomic localizations and gene regulatory functions of MBD-2 with and without NuRD in Caenorhabditis elegans which lacks DNA methylation. Nat Commun 17, 1875 (2026). https://doi.org/10.1038/s41467-026-68592-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-68592-0
