
Abstract
Gene fusion products involving protein kinases are known drivers in human cancers and actionable targets for personalized therapy, yet the structural and molecular determinants that control their function are largely unexplored. Here we show that a CCDC6-RET fusion protein, a driver and therapeutic target in lung and thyroid cancers, is a highly active dimeric kinase. Time-resolved mass spectrometry together with a robust biochemical and biophysical characterization reveal that CCDC6-RET functions as a dual ATP- and ADP-dependent kinase able to bind both nucleotides and to use them as phosphoryl donors. We also identify a crosstalk between the C-terminal and the activation segments controlling both the processing and the catalytic activity of the fusion protein. Furthermore, a 3D-structural assembly of a CCDC6-RET homodimer was generated combining single particle electron microscopy, small-angle X-ray scattering and in silico molecular dynamics simulations. Our structural model together with cross-linking mass spectrometry data demonstrated that CCDC6-RET in the inactive state forms a face-to-face dimer characterized by intermolecular-crosslinked activation segments. Upon nucleotide binding the catalytic domains swing apart and fast activation loop phosphorylation could be driven by a mechanism in cis. Our work uncovers the molecular and structural determinants that control the mechanism of CCDC6-RET autoactivation.
Similar content being viewed by others


Introduction
Following the paradigm of the Philadelphia chromosome and the BCR-ABL1 fusion in chronic myelogenous leukemia (CML)1,2, gene fusion products involving protein kinases are known drivers in human cancers and actionable targets for personalized therapy3. Yet, the structural and molecular determinants that control their function and (oncogenic) mechanism of action are paradoxically elusive in most of the cases. In 1985, a proto-oncogene named Rearranged during Transfection (RET) was discovered in NIH3T3 cells transfected with DNA isolated from human lymphoma cells, to cause malignant transformation4. The characterization of the isolated RET cDNA clones revealed a carboxy-terminal domain with high homology with members of the tyrosine kinase gene family preceded by a hydrophobic sequence characteristic of a transmembrane domain, suggesting that the RET oncogene encoded a cell-surface receptor5. The characterization of the human6 and mouse7,8 proto-oncogene orthologs revealed the amino acid sequence (primary structure) of the RET protein. Shortly after, a RET fusion product was discovered in papillary thyroid carcinoma (PTC), it was named RET/PTC1 and its N-terminal partner protein identified as H4(D10S170)9,10. The RET/PTC-1 oncogene was a fusion product containing the N-terminus of the H4 gene, currently known as CCDC6 (Coiled-Coil Domain Containing protein 6, aa 1-101) fused to the tyrosine kinase domain of c-RET10. CCDC6-RET fusions are detected in up to 30% of papillary thyroid carcinomas10,11. Although the prevalence of CCDC6-RET fusions in PTC shows significant geographic variation12, there is a strong correlation with radiation exposure. In 1986, after the Chernobyl nuclear power disaster, a sharp increase in the incidence of PTC was observed in children and adolescents of contaminated regions of Belarus, Russia, and Ukraine with near 60% of post-Chernobyl PTCs children patients harboring a RET fusion13. Up to date, more than twenty RET fusions products have been described in PTC3. The most common RET fusions products are CCDC6-RET (RET/PTC1) and NCOA4-RET (RET/PTC3), which together account for about 90% of RET fusion-positive cases in thyroid cancer12. In all these cases, the DNA-rearrangements generate the fusion of the tyrosine kinase domain of RET to the 5’ portion of different genes. Breakpoints in the RET gene are consistently located at intron 11, so the fusion starts from exon 12 to the stop codon, resulting in a protein chimera containing the tyrosine kinase domain but lacking the juxtamembrane segment and extracellular domains of RET12. A common molecular hallmark in PTC is the constitutive activation of the MAPK (Mitogen-Activated Protein Kinase) cascade. Indeed, besides RET and other RTKs, common PTC-drivers are gain-of-function mutations of RAS and BRAF kinase, respectively. The mutually exclusive nature of these lesions supports the notion that they act along a common signaling axis driving the oncogenic phenotype in PTC14. RET fusions are also found in 1–2% of lung adenocarcinoma (LADC)15,16,17. The most common RET fusion products in non-small cell lung carcinoma (NSCLC) are KIF5B-RET, NCOA4-RET, and CCDC6-RET respectively. KIF5B (Kinesin-1 heavy chain) is by far the most common RET fusion partner in NSCLC, being detected in up to 70–90% of the cases15. In NSCLC, RET fusions are mutually exclusive with other driver mutations, such as ALK or ROS1 rearrangements or EGFR or KRAS mutations, suggesting again common signaling mechanisms driving the oncogenic phenotype, and the definition of a potential molecular subtype17,18.
Clinically, there is a strong interest in the development of next-generations RET tyrosine kinase inhibitors for the development of personalized therapies in NSCLC and PTC19,20. Crucially RET fusions (CCDC6-RET, NCOA4-RET, and the newly described CDC123-RET) have been shown to mediate acquired resistance mechanism in lung adenocarcinomas (LADC) to EGFR or ALK tyrosine kinase inhibitors21,22. Understanding the structural, molecular and dynamic determinants that control the catalytic function and signaling of these oncogenic RET fusion products will uncover druggable vulnerabilities. Such vulnerabilities can be therapeutically exploited for the design and development of next-generation inhibitors for targeted and personalized therapies that are able to overcome acquired resistance by secondary mutations in RET-driven cancers19,20,23,24,25,26. In this study we produced a recombinant CCDC6-RET gene fusion product using a baculovirus expression system. We performed a detailed functional and structural characterization of the CCDC6-RET fusion protein by state-of-the-art biochemistry, mass spectrometry and structural biology tools that allowed us to generate a 3D-structural model of a CCDC6-RET homodimer for single particle analyses and mechanistic validation. Our findings uncover the structural and molecular determinants that control a RET oncogenic fusion´s mechanism of autoactivation, revealing unexpected and striking features not previously envisioned with important implications for drug discovery. Our data provides at the same time a solid framework for the structural and functional study of other RET fusion products.
Results
Expression, purification and quality check of a recombinant CCDC6-RET fusion product
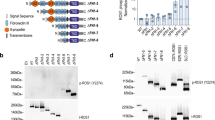
We have produced a recombinant CCDC6-RET fusion product (Fig. 1a) composed by fragments of residues 1-101 of CCDC6 and residues 713-1072 of RET using a baculovirus expression system in Sf9 insect cells. Protein purification was achieved by tandem immobilized metal affinity (Ni2+) and glutathione-conjugated gravity flow chromatography and in-gel 3C-protease digestion followed by size-exclusion chromatography, if required (Fig. 1b). This is an updated version of a previously published single-step affinity protocol using glutathione-conjugated resin for the purification of recombinant RET kinase and intracellular domains expressed in insect cells using a baculovirus system27,28,29. The tandem double affinity chromatographic step allowed us to obtain highly pure and monodisperse protein with no traces of DNA or nucleotide contamination as indicated by the values of absorbance at 280 and 260 nm and their optimal ratio (Fig. 1c), resulting in a recombinant protein yield of 0.3-0.4 mg per liter of culture. Further quality check over a range of different protein concentrations by mass photometry shows that CCDC6-RET is a 101.2 ± 2.2 kDa dimer in solution (Fig. 1d). Binding of RET tyrosine kinase inhibitors BLU-66720 and Ponatinib30 was evaluated by DSF measuring changes in intrinsic fluorescence (IF) upon a temperature gradient (Fig. 1e, f). Binding of both compounds to CCDC6-RET resulted in a significant shift of the inflection temperature (Ti) of 12.2 and 19.8 °C, respectively, indicative of a robust thermal stabilization (Fig. 1f). Altogether, these results assured the quality, monodispersion and catalytic integrity of the protein sample prior to its functional and enzymatic characterization. Initial functional characterization showed that CCDC6-RET functions as a highly-active dimer in solution displaying fast autophosphorylation kinetics (Fig. 1g) in agreement with the KM value for ATP 139.3 ± 11 μM compared with the RET kinase domain (KD, 429 ± 43 μM) construct27, see Fig. 1h, i.

a Schematic diagram of the CCDC6-RET fusion product depicting functional domains with the indicated phospho-sites and aa number. b In tandem immobilized metal affinity (Ni2+), glutathione-conjugated gravity flow chromatography and in-gel 3C-protease digestion. Indicated fractions were run on an SDS-PAGE and stained with Coomassie: 1–4 elution fractions with 100% buffer B, 5 recombinant protein bound to the glutathione conjugated resin before proteolytic cleavage, 6–7 elution fractions after cleavage with 3C-PreScission protease (GST-tagged, see lane 8), 8 glutathione conjugated resin after proteolytic cleavage. Size exclusion chromatography (sec) using a Superdex 200 3.2/300 column. Indicated fractions were run on an SDS-PAGE and stained with Coomassie (lower panel). Data are representative of n ≥ 10 experiments. c Absorbance spectra (260–310 nm) with max at 280 nm and indicated (260/280 nm) ratio. d Mass photometry profiles (counts vs mass in kDa) of CCDC6-RET at different concentrations in color code: 50 nM (magenta), 25 nM (blue), 10 nM (green), 5 nM (orange). e DSF profile (IF) of CCDC6-RET (2 μM) in the apo state and bound to BLU-667 (10 μM) and Ponatinib (10 μM). f Ti (°C) and ΔTi (°C) values from (f). Data (mean ± SEM) of three experiments, n = 3. g WB of samples from a time-course autophosphorylation experiment with CCDC6-RET (1 μM) in the presence of ATP (1 mM) and MgCl2 (2 mM) for 0–30 min using a total anti-phospho-tyrosine antibody. The total amount of protein was visualized by Coomassie staining. Data are representative of n ≥ 10 experiments. h Enzymatic assay performed with CCDC6-RET (0.25 μM) incubated with increasing concentrations of ATP at a fixed concentration (1.35 mM) of a c-Abl-derived peptide (EAIYAAPFAKKK). Data represent the mean ± SEM of three experiments in duplicate (n = 6). Enzymatic activity (ODs−1 × 10−2) versus ATP concentration and KM values are depicted. i Consumption curves OD (λ 243 nm) versus time (in seconds) at increasing concentrations of ATP (0–2.5 mM) of a representative enzymatic assay from (h). Source data are provided as a Source data file.
Mass spectrometric characterization of CCDC6-RET autophosphorylation
Next, we applied high-resolution time-resolved liquid chromatography tandem mass spectrometry (LC-MS/MS) to monitor the dynamic changes of the autophosphorylation reaction (0–30 min) on CCDC6-RET in vitro. Fast autophosphorylation kinetics were observed in most of the identified phospho-sites, which recapitulated the results from the WB data using phospho-specific antibodies (Fig. 2a). A total of seventeen phospho-sites were mapped, of which fourteen were the facto autophosphorylation sites as indicated by their time-dependent saturation kinetics during the reaction (Fig. 2b, c and Table 1). CCDC6-RET autophosphorylates on both serine (e.g., S904 and S909) and tyrosine residues (Fig. 2c, f and Table 1). CCDC6-RET autophosphorylates not only on sites located on the catalytic core of the fusion, but also on discrete sites of the N-terminal and coiled-coil region of CCDC6, some of which were not previously described. We identify among other several phospho-sites on the amino terminal and coiled-coil region of the gene fusion product: Ser 46, Ser 52, Tyr 80 and Ser 98 (Fig. 2c, d). Mutating Ser 46, Ser 52 and both residues simultaneously to alanine does not cause any defects on the phospho-tyrosine activity as seen in in vitro autophosphorylation reactions (supplementary Fig. 1a-c). In the same line, a CCDC6-RET fusion with a Y80F or a S98A mutation appeared to be catalytic competent (supplementary Fig. 1d, e). Autophosphorylation sites on the catalytic core of the RET fusion were also identified (Table 1 and supplementary tables 1-4). These phospho-sites are located at different structural and functional motifs of the RET kinase domain: Tyr 806 and Tyr 809 (hinge), Tyr 826 (kinase insert), Ser 896, Tyr 900, Ser 904, Tyr 905, Ser 909 (activation loop) and Tyr 1015, Tyr 1029, Ser 1021, Ser 1034 and Tyr 1062 (C-terminal segment), see also Fig. 2c, e, f. Some of these phospho-sites showed high levels of phosphorylation in the basal state which decreased over time during the autophosphorylation reaction. This is the case for activation loop Tyr 900 and C-terminal Ser 1034; both phospho-sites are located within peptides containing in addition other bona fide autophosphorylated residue whose levels increase one order of magnitude during the reaction. See the example of activation loop tyrosines 900 and 905 (Fig. 2c, f). In other cases, phospho-sites appeared only in doubly or triply phosphorylated peptides (e.g., Ser 896 and Ser 1021). A complete list of the CCDC6-RET phospho sites consistently identified across the four independent experiments is shown on main Table 1 (see also supplementary tables 1-4). Quantifications of representative high occupancy autophosphorylation sites from three independent experiments are shown in supplementary Fig. 2. Next, we targeted C-terminal phospho-sites for site-directed mutagenesis to test whether they are required for catalytic activity. The mutation of residue Tyr 1015 to phenylalanine (Y1015F) did not cause any detrimental effect on tyrosine phosphorylation as seen in the time course autophosphorylation assays (supplementary Fig. 1g). The same applied to a Y1062F and S1034A mutants (supplementary Fig. 1g, h).

a WB of samples from a time-course autophosphorylation experiment with CCDC6-RET (1 μM) in the presence of ATP (1 mM) and MgCl2 (2 mM) for 0–30 min using the indicated total and phospho-specific antibodies. The total amount of protein was visualized by Coomassie staining. WBs images are representatives of ≥10 independent experiments. b Schematic diagram of the CCDC6-RET fusion product with the identified phospho-sites and aa number (note hereafter that sites of the fusion after aa 101 of CCDC6 follow the RET aa numbering) that resulted from the LC/MS-MS analyses. c Heat map (intensity, white-blue scale arbitrary units) representation of the phospho-proteomic quantification from the CCDC6-RET autophosphorylation reaction in vitro (0–30 min, ATP) identified by LC/MS-MS. d–f Phosphorylation kinetics (standarized and scaled phospho-signal) of the indicated CCDC6-RET phospho-sites (highlighted in red, with the different tryptic peptides and charges) measured by LC-MS/MS from c. Data shown are two technical replicates of a single experiment. Data are representative of four independent experiments. Source data are provided as a Source data file.
As negative controls in the phospho-proteomic experiments we used adenosine monophosphate (AMP) and biphosphate (ADP) nucleotides (supplementary Fig. 3). We found a striking increase of CCDC6-RET tyrosine phosphorylation in the presence of ADP and not by AMP. These unexpected results were a clear sign of ADP-dependency by the CCDC6-RET fusion product, which we explored further (see next “Result” section).
CCDC6-RET is a dual ATP- and ADP-dependent kinase
Next, we performed a direct functional comparison of CCDC6-RET with a RET 713-1072 aa fragment, the catalytic core of the fusion product, as a control (Fig. 3a). First, we performed enzymatic assays to calculate the catalytic rates and enzymatic constants for ATP using an excess of peptide as an exogenous substrate (Fig. 3b). Under these experimental conditions, CCDC6-RET was a more active enzyme than the catalytic core of the RET fusion (aa 713-1072) as indicated by their corresponding KM (133.25 ± 13 μM vs 429.28 ± 77 μM) and a six-fold increase in the catalytic efficiency constant (\({k}_{cat}/{K}_{M}\)) by the oncogenic chimera (Fig. 3b–d). We next performed in vitro time course autophosphorylation assays (0–30 min) using ATP (1 mM), MgCl2 (2 mM) with 1 μM final protein concentration. Autophosphorylation was monitored by Western blotting (WB) using RET total and phospho-specific antibodies against Tyr 905, Tyr 1015, Tyr 1062 and a total phospho-tyrosine antibody (Fig. 3e). Fast CCDC6-RET autophosphorylation kinetics reaching saturating levels at early time points (1 min) contrasted with slower rates of autophosphorylation by the 713-1072 construct, see e.g., Tyr 905 and Tyr 1015 phosphorylation (Fig. 3e). We also evaluated the binding of ATP and ADP to both constructs respectively by monitoring changes of IF (Fig. 3h–j). Titration experiments using a range of increasing concentrations of both nucleotides allowed us to calculate an apparent KD for ATP (186.8 ± 38 μM) and ADP (1057.0 ± 157 μM) respectively (Fig. 3j). These results together with the MS data (supplementary table 2) prompted us to explore further whether CCDC6-RET is a dual ATP- and ADP-dependent kinase that can use in addition to ATP, ADP as a phosphoryl donor. We found that in contrast to a 713-1072 construct, CCDC6-RET efficiently auto-phosphorylates in the presence of ADP (Fig. 3f, g), although with slower rates than with ATP, in accordance with the affinity values obtained (Fig. 3j). Together these data demonstrate that CCDC6-RET: i) is a highly active dimeric kinase in solution with fast kinetics of autophosphorylation, and ii) functions as a dual ATP- and ADP-dependent kinase able to bind to and use both nucleotides as phosphoryl donors.

a Schematic diagram of the CCDC6-RET fusion product and RET catalytic core (aa 713-1072) depicting functional domains with the indicated phospho-sites and aa number. b Enzymatic assay performed with CCDC6-RET (0.25 μM) and RET 713-1072 (0.25 μM) incubated with increasing concentrations of ATP at a fixed concentration (1.35 mM) of a c-Abl-derived peptide (EAIYAAPFAKKK). Data represent the mean ± SEM of three experiments with two duplicates each, n = 6. c Catalytic efficiency constant (\({k}_{cat}/{K}_{M}\)) fold-change versus control (i.e., RET aa 713-1072) from (b). d Consumption curves OD (λ 243 nm) versus time at increasing concentrations of ATP (0–2.5 mM) of a representative enzymatic assay from b. Legend: Dotted (RET aa 713-1072) and straight (CCDC6-RET) lines are depicted. e–g WB of samples of an in vitro time-course autophosphorylation experiment with CCDC6-RET (1 μM) and RET aa 713-1072 (1 μM) in the presence of ATP or ADP (1 mM) and MgCl2 (2 mM) for 0–30 min using the indicated antibodies. Data represent two experiments (n = 2). h, i Representatives DSF profiles (IF, emission 330 nm, first derivative) of CCDC6-RET upon titration with increasing concentrations of nucleotides (ATP and ADP), showing the transitions of the inflexion temperature (Ti). j Apparent KD values for ATP and ADP (μM). Increment of the Ti (ΔTi, °C) versus the concentration of nucleotide (μM). Data shown is the mean value ± SEM of three experiments (n = 3). Source data are provided as a Source data file.
ADP regulates CCDC6-RET phospho-tyrosine activity in vitro
To explore further the ADP dependent activity of CCDC6-RET, we performed ATP and ADP titration experiments in which the phospho-tyrosine activity of CCDC6-RET was monitored under increasing concentrations of nucleotide (range 10–5000 μM) using a total phospho-tyrosine and a RET Tyr 905 phospho-specific antibodies (Fig. 4a). Increasing CCDC6-RET phospho-tyrosine activity was observed at low ATP concentrations (10–50 μM) reaching saturation levels at 1000 μM (Fig. 4a). In the case of ADP, increasing CCDC6-RET phospho-tyrosine activity reached a maximum at 1000 μM, which followed a marked decrease in activity at 2500 and 5000 μM concentrations. These data suggested a potential inhibitory role of ADP at high concentrations, which we explore further in ADP-ATP competition assays (Fig. 4b). Pre-treatment of CCDC6-RET with increasing concentrations of ADP (10–5000 μM) followed by treatment with ATP (1 mM) for 15 min resulted again in a detrimental effect on the activity induced by ATP at higher ADP concentrations (2500 and 5000 μM). These data again demonstrate a regulatory and competing role of ADP at higher concentrations, resulting in the inhibition of the CCDC6-RET activity induced by ATP. These data were also recapitulated in time course autophosphorylation experiments (0–30 min) where the activation of CCDC6-RET by ATP was completely abrogated over the complete time course (even at late time points) by pretreatment with 5 mM ADP (Fig. 4c). In the case of pretreatment with 1 mM ADP, a consistent detrimental impact at early time points (1 min) with 1 mM ATP stimulation was observed (Fig. 4d). In these sets of experiments, we used a catalytic dead HRD/A mutant (D874A) of CCDC6-RET which did not respond to either ATP or ADP (Fig. 4b and c). To prove direct β phosphate transfer of ADP by CCDC6-RET in the autoactivation mechanism, we used ADP radiolabeled on the β-phosphate (β-ADP-P33) in autophosphorylation experiments (Fig. 4e). In parallel, we used radiolabeled ATP (γ-ATP-P32) as an internal control. In these in vitro experiments using radioactive nucleotides, we found that CCDC6-RET efficiently autophosphorylates in the presence of ADP, although with slower kinetics than with ATP, as anticipated (Fig. 4e).

a Nucleotide dose-dependent experiment of CCDC6-RET (WT, 1 μM) under increasing concentrations of both ATP and ADP (0–5000 μM, 5 min) was monitored by WB using the indicated phospho-specific antibodies. The total amount of protein was visualized by Coomassie staining. Quantification of CCDC6-RET phospho-tyrosine activity; data (phospho-signal RET Tyr 905, fold-change) represent the mean ± SEM of five (n = 5) experiments for ADP and four (n = 4) for ATP. b An ADP-ATP competition assay with CCDC6-RET (WT, 1 μM) pre-treated with increasing concentrations of ADP (0–5000 μM, 20 min) followed by ATP stimulation (1 mM, 15 min) was monitored by WB using the indicated phospho-specific antibodies. The total amount of protein was visualized by Coomassie staining. Data (total phospho-tyrosine signal, ratio) represent the mean ± SEM of three experiments, n = 3. A CCDC6-RET HRD/A mutant (D874A) was used as a negative control, n = 2. c An ADP-ATP competition experiment with CCDC6-RET WT (1 μM) unstimulated and pre-treated with 5 mM ADP (20 min), after which it was subjected to a time course (0–30 min) with ATP (1 mM). CCDC6-RET phospho-tyrosine activity was monitored by WB using the indicated phospho-specific antibodies. The total amount of protein was visualized by Coomassie staining. Data (total phospho-tyrosine signal, ratio) represent the mean ± SEM of three experiments, n = 3. A CCDC6-RET HRA mutant (D874A) was used as a negative control, n = 2. d An ADP-ATP competition experiment with CCDC6-RET WT (1 μM) unstimulated and pre-treated with 1 and 5 mM ADP (20 min), after which it was subjected to a time course (0–30 min) with ATP (1 mM). CCDC6-RET phospho-tyrosine activity was monitored by WB using the indicated phospho-specific antibodies. The total amount of protein was visualized by Coomassie staining. Data (total phospho-tyrosine signal, ratio) represent the mean ± SEM of three experiments, n = 3. e In vitro time course autophosphorylation experiment of CCDC6-RET stimulated with radioactive ATP-P32 and ADP-P33 as described in material in methods. Data are representative of two independent experiments. f An active CCDC6-RET dimer MD model (accession code: ma-pcoh8 and supplementary file CCDC6-RET_ADP2.pdb) in complex with ADP. Coiled-coil (green) and catalytic core (blue), activation loop phospho-sites are shown together with nucleotide (purple) and peptide substrate (EAIYAAPFAKKK) in wheat color code. In the right panel, a magnification view of the active site with key residues, secondary structural elements and functional motifs are depicted and labeled. Source data are provided as a Source data file.
We generated structural models for an active CCDC6-RET homodimer (accession codes: ma-pcoh8 and supplementary file CCDC6-RET_ADP2.pdb) in complex with ADP, Mg2+ and a Tyr-containing peptide substrate that revealed a plausible structural basis for ADP-dependent catalysis (Fig. 4f). In this process, activation loop Arg 912 would play a pivotal role fixing the β-phosphate group of ADP by displaying an extended sidechain that is fully compatible with peptide binding and adoption of a near-attack conformation for the phospho-transfer reaction. Importantly, the backbone amides from activation-loop Iso 913 and Pro 914 form the necessary hydrogen bonds with +1 and +2 residues of the phosphorylatable tyrosine-containing peptide substrate. In addition to the prototypical interactions required for ATP γ-phosphate transfer, namely, the salt bridge between β3 K758 and αC E775, the coordination of Asp 892 (DFG motif) with Mg2+, and the positioning of Asp 874 in the HRD motif for proton abstraction from the tyrosine hydroxyl group; a key cation-pi interaction between Arg 878 and Trp 917 is also in place for the correct arrangement of the Michaelis complex. Taken together, our biochemical and structural data support the notion of an ADP-dependent activity by CCDC6-RET.
Activation loop Tyr 905 is required for CCDC6-RET catalytic activity
It is known that the activation loop of protein kinases in the unphosphorylated state can function as an autoinhibitory module by sterically blocking substrate access into the active site31. Based on our own results indicating that the C-terminal segment can also restrict the access of the substrate into the catalytic cleft (supplementary Fig. 4), we aimed to explore further a potential crosstalk between the activation loop and the C-terminal segments in the mechanism of CCDC6-RET autoactivation and signaling. For this purpose, we generated a series of activation loop mutants in full-length and C-terminal deleted constructs (ΔCT). First, we produced single and double activation loop tyrosine mutants in a full-length construct (Y900F, Y905F and Y900/905F) and evaluated their catalytic activity in time course autophosphorylation experiments (Fig. 5a). None of the evaluated mutants had a significant detrimental impact on the tyrosine phosphorylation activity compared with the WT control in autophosphorylation experiments (Fig. 5a). However, in enzymatic assays using a peptide substrate we found that Tyr 905 was required for enzymatic activity (Fig. 5c). Of note, a full-length CCDC6-RET Y900F mutant construct resulted consistently in very low yield of soluble protein after purification (supplementary Fig. 5). The very low levels of expression made it difficult to work at comparable levels due to its short life and that it was not possible to concentrate it to reach workable concentrations. Intriguingly, such detrimental effect on protein yield was rescued by the concomitant mutation on Tyr 905 in the case of a double activation loop mutant (Y900/905F), see Fig. 5a. On the contrary, when we evaluated the function of the complete series of activation loop mutants in a C-terminal deleted context (ΔCT), we observed that the Y900F mutant was efficiently expressed to the same levels as the WT protein. Interestingly, we also observed a catalytic dependency on activation loop Tyr 905 in the ΔCT constructs (Fig. 5b). We found that in autophosphorylation and enzyme kinetics experiments, where we evaluated the KM for ATP in the presence of an excess of peptide substrate, a Y905F and double Y900/905F mutants have a loss of function effect, contrary to the Y900F, which was catalytic competent (Fig. 5d). Hence, these data uncovered a catalytic dependency on Tyr 905 by the CCDC6-RET fusion product. Interestingly, these data were recapitulated although to a lower extent by a S909A mutant, which also has a compromised enzymatic activity towards a peptide substrate (supplementary Fig. 6). Taking these data together, we conclude that the CCDC6-RET activation loop Tyr 905 controls the catalytic activity and signaling of the CCDC6-RET fusion protein.

a Schematic diagram of the CCDC6-RET fusion product depicting functional domains with the indicated phospho-sites and aa number (upper inset). WB of samples from a time-course autophosphorylation experiment with CCDC6-RET WT and activation loop mutants Y905F and Y900/905F (1 μM) in the presence of ATP (1 mM) and MgCl2 (2 mM) for 0–30 min using the indicated antibodies. Note that a CCDC6-RET mutant Y900F is not viable and cannot be properly evaluated. Data represent n = 4 experiments. b Schematic diagram of the CCDC6-RET ΔCT fusion product depicting functional domains with the indicated phospho-sites and aa number (upper inset). WB of samples from a time-course autophosphorylation experiment with CCDC6-RET ΔCT WT and activation loop mutants Y900F, Y905F and Y900/905F (1 μM) in the presence of ATP or ADP (1 mM) and MgCl2 (2 mM) for 0–30 min using the indicated antibodies. Note that a CCDC6-RET ΔCT Y900F is viable and functional. Data represent n = 5 experiments. c, d Enzymatic assay performed with CCDC6-RET and ΔCCDC6-RET CT, WT, and indicated activation loop mutants Y900F, Y905, and Y900/905 F (0.2 μM) incubated with increasing concentrations of ATP at a fixed concentration (1.35 mM) of a c-Abl-derived peptide (EAIYAAPFAKKK). Enzymatic activity data (ODs−1 × 10−2) represent the mean ± SEM of three experiments in duplicate (n = 6). Catalytic efficiency constant (\({k}_{cat}/{K}_{M}\)) fold-change versus control (WT). Source data are provided as a Source data file.
In silico reconstruction of a CCDC6-RET homodimer
We generated in silico a high-confidence structural reconstruction of a CCDC6-RET homodimer. First, we used the Waggawagga coiled-coil prediction server32. Coiled-coil predictions are based on the identification of periodic heptad repeats, which can be represented by a net- or helical wheel-diagrams. This bioinformatic tool enables the discrimination between coiled-coil domains and single α-helices (SAH). In the case of CCDC6-RET there is a high confidence parallel homodimer coiled-coil interface from residues 56-101 of each protomer (Fig. 6a, d), as indicated by both the coiled-coil probability marcoil (p-value 100) and very low SAH-score (0.0333). The coiled-coil inner hydrophobic core was clearly defined by the heptad repeats (Fig. 6b, c), and several salt-bridge interactions (strong) involving Glu 58 and Arg 62, Lys 74 and Glu 78, and Lys 85 and Glu 89. These contacts also involved Glu 78 and Lys 81 and Glu 89 and Arg 92, and weak coordination between Asp 93 and Lys’96, as depicted in the helical wheel- and heptad-net-diagrams (Fig. 6b, c). We visualized in 3D a parallel homodimeric coiled-coil interface (Fig. 6d) using CCbuilder 2.033. In an attempt to disrupt the dimerization interface, we generated a series of mutants at key positions within the coiled-coil, for example Cys 84 (C84A) that can form a disulfide bridge with its counterpart (Fig. 6d), and Ser 98 (S98A) which we found was an unexpected autophosphorylation site, see main Table 1 and supplementary tables 1- 4. None of these mutants was able to dissociate the dimer as indicated by the elution volumes observed in SEC analyses compared with the WT (data not shown). These coiled-coil mutants did not affect the activity of the fusion products as detected in autophosphorylation experiments (Supplementary Fig. 1e and f), suggesting a tight dimerization constant by the interface. Next, we applied artificial intelligence to obtain a high-confidence 3D-predictive model of a monomeric CCDC6-RET fusion protomer using AlphaFold (AF id: J7M8C2). By the superimposition of two monomers of the predicted fusion protein onto the dimeric coiled-coil interface (Fig. 6d) we generated a two-fold rotational symmetry dimer (Fig. 6e) in which the catalytic domains are facing away symmetrically from the coiled-coil stem. The high confidence and feasibility of the predictive 3D-model was supported by careful inspection of key structural features, see supplementary Fig. 7a-d: i) the correct position and configuration of secondary structural elements and functional motifs of an active kinase defined overall by the correct alignment of the regulatory (R-) and catalytic (C-) spines and DFG-in configuration28; ii) the hydrophobic patch formed by contacts between the αC helix (i.e., PIF-like pocket) and the coiled-coil stem; iii) analogy to the domain arrangement found in bacterial histidine kinases where the catalytic domain is place to the left of the symmetry plane of the coiled-coil interface defined in those cases by a four helix bundle instead34, and iv) in good agreement with two independent and high-confidence (pLDDT scores of 79.08 and 87.74, respectively) solutions provided by AlphaFold multimer (ColabFold) and AlphaFold3, respectively. These represent different configurations and functional states depending on the specific situation of the catalytic domains with respect to the coiled-coil interface (supplementary Fig. 7e, f).

a Coiled-coil probability (p-score) and single alpha helix (SAH) score for Marcoil prediction of the N-terminal sequence of CCDC6-RET. b Identification of the heptad repeats depicted in the form of a net-diagram for the coiled-coil region (aa 54-101) of CCDC6-RET. c Heptad repeats represented in a helical wheel view, from (a, b). d Cartoon representation of the coiled-coil region (aa 54-101) of CCDC6-RET with the side chains of the represented residues in sticks. Frontal and lateral views are depicted with indicated side chain residues. e A high-confidence theoretical 3D-model of a monomeric CCDC6-RET fusion protomer comprising the short C-terminal isoform of RET using Alpha-Fold (AF id: J7M8C2). Superimposition of two monomers of the predicted fusion product onto the parallel homodimeric coiled-coil interface generates a two-fold rotational symmetry dimer in which the catalytic domains are facing away from the coiled-coil stem. Frontal and upper views are depicted and schematic diagram of the domain organization showing the 2-fold rotational symmetry arrangement of the domains (top view).
Architecture of a CCDC6-RET homodimer by single particle EM
For EM, freshly purified CCDC6-RET was placed on homemade carbon-coated 400 mesh copper grids and negatively stained. An analysis of the EM field showed a relatively uniform distribution of particles (Fig. 7a). Reference-free 2D averages obtained with Relion35 revealed particles of similar size but varying shapes and orientations, clearly indicating the presence of a two-fold symmetry (Fig. 7b). A similar 2D-classes were obtained with cryoSPARC36, see supplementary Fig. 8b). To compare the in silico reconstructed CCDC6-RET homodimer with the experimental data, we filtered the in silico model to 15 Å and generated its retroprojections (supplementary Fig. 8a). The generated 2D projections show good agreement in size and shape with the reference-free 2D averages of a CCDC6-RET homodimer (Fig. 7 b, c, and supplementary Fig. 8b), with a thinner stem (coiled-coil interface) in the middle and two globular arms segregated from the stem facing at opposite almost perpendicular directions. An initial 3D model was built and further refined using cryoSPARC to produce the low-resolution EM map (Fig. 7d, e, and supplementary Fig. 8c). The reliability of the map is supported by the strong agreement between the reference free 2D classes and the projections of the map and the even distribution of Euler angles (supplementary Fig. 8d). The 3D-map confirms the dimeric and twisted arrangement of the globular catalytic domains towards the left of the 2-fold rotational axis from the coiled-coil stem. This comes in line with the catalytic domain configuration seen in bacterial histidine kinases34, where the catalytic core of the DHp-CA module is placed to the left of helix b. When the density map is contoured at a threshold chosen to encompass a molecular weight of approximately 110 kDa (calculated molecular weight of CCDC6-RET), the map generally has good connections between adjoining domains but lacks density for the whole coiled-coil interface and N-terminal part of the fusion. Importantly, the independent dimeric models from AlphaFold fit well into the low-resolution EM map, supporting the accuracy of the AlphaFold prediction (see Fig. 7e and supplementary Fig. 7a-f and 8c).

a Representative micrograph of an NS grid, scale bar 100 nm. Magnified view on the right panel. Data represent n ≥ 10 images. b Reference free 2D-classification showing different views, sizes and orientations of CCDC6-RET. c Orthogonal views of an in-silico 3D-reconstructed CCDC6-RET dimer showing particle size. Note that N- and C-terminal CCDC6-RET IDRs are not included. The coiled-coil region is colored in green and catalytic domains in blue. d 3D-maps (surface representation) of CCDC6-RET, in different orthogonal orientations (side, top, and bottom views). e Superimpositions of 3D-maps (surface representation) from d, onto the in-silico 3D-reconstructed CCDC6-RET dimer from (c), in different orthogonal orientations (side, top, and bottom views).
To obtain further information on the molecular shape of a CCDC6-RET homodimer in solution, low-angle X-ray scattering (SAXS) data of a CCDC6-RET-ΔCT construct were collected (supplementary Fig. 9a). Pair distance distributions exhibited fine features consistent with a multidomain protein sample. Ab initio envelopes were generated for CCDC6-RET-ΔCT using DAMAVER consistent with the pair distance distributions showing an elongated shape at the root of the coiled-coil and a more globular architecture at the catalytic portion of the fusion resulting in an inverted T-shape object (Supplementary Fig. 9b and c) with a radius of gyration (Rg) of 55.07 ± 0.18 Å and Dmax of 195 Å. These measurements are in accordance with the reference-free 2D- particles and 3D-envelop obtained by electron microscopy and also the 3D-in silico reconstructed particles considering the IDRs (see Figs. 6 and 7). Of note, the CCDC6-RET-ΔCT construct generated an extra volume or ¨bump¨ adjacent to one of the two catalytic domains of the fusion, which was tentatively attributed to the dynamic motion of both catalytic domains, as we can fit in another catalytic domain molecule within the extra volume. The CCDC6-RET-ΔCT structure was modeled using a high-confidence Alpha-Fold (AF id: J7M8C2) prediction of a monomeric chimera without the amino- and carboxy-terminal IDRs. The initial 54 amino acids were then manually built into the SAXS envelope (supplementary Fig. 9d). The fitting of the theoretical curves derived from this CCDC6-RET-ΔCT model against the experimental curves gave a χ2 value of 2.2, indicating a good quality of the data, see Supplementary Table 5. The high-quality SAXS data were therefore sufficient to generate a preliminary model for a CCDC6-RET dimer assembly, defining clear pairwise interdomain angles of approximately 90° between the coiled-coil interface and the catalytic domains and a potential solution for the extra volume observed in the SAXS envelop (supplementary Fig. 9d). The coiled-coil and catalytic domains angle appears to be in the range observed for the DHp-and CA domains in bacterial histidine kinases34,37,38. The SAXS envelops fits well all the in silico-generated CCDC6-RET assemblies and the 3D-map from the EM single particle analyses, capturing the dynamic transitions between the different assemblies’ states and the dynamic motion of the catalytic domains from the coiled-coil stem. These data also suggest some degree of asymmetry between catalytic domains and the dimer interface (supplementary Fig. 9d).
Cross-linking MS analyses of inactive and autophosphorylated CCDC6-RET
Cross-linking MS is a technique that captures conformational changes in solution by mapping structural details of individual proteins and their complex assemblies based on the coordination of amino acid pairs that are positioned in close proximity to each other by using chemical crosslinkers39. These covalent connections then serve as basis for deriving distance constraints within the protein(s) different conformational and functional states. To provide experimental restraints for the 3D-reconstruction of the different functional states of a CCDC6-RET homodimer (i.e., unstimulated and autophosphorylated forms), we performed in-solution cross-linking with the homobifunctional cross-linker DSBU (disuccinimidyl dibutyric urea)39. CCDC6-RET cross-linking was performed under unstimulated and saturating ATP and MgCl2 (60 min) conditions. A non-crosslinked sample was used as an internal control (Fig. 8a). Three independent replicates of each set of samples were subjected to high-resolution LC-MS analysis. In parallel, an LC-MS analysis of non-crosslinked sample was performed to assess the purity and homogeneity of the CCD6-RET preparation. This analysis demonstrated a highly pure and homogenous recombinant protein preparation (Fig. 8a). Size exclusion chromatography of the crosslinked samples (both unstimulated and autophosphorylated) revealed the same molecular size (as indicated by the same elution volume) than the non-crosslinked one, which excludes the formation of artefactual oligomeric and/or high molecular weight species during the treatment with the crosslinker (Fig. 8a, inset). We identified a total of 148 and 131 crosslinked counts by Merox in the resting (i.e., inactive, unstimulated) and phosphorylated states, which resulted in the final identification of 55 and 46 crosslinked peptides, respectively (Fig. 8b). Next, we classified the specific crosslinked peptides for each of the functional states resulting in the identification of 13 specific crosslinked peptides for the unstimulated form (i.e., resting) and 9 for the autophosphorylated (i.e., active) protein, respectively, as shown in the Venn diagram (Fig. 8c). Overall assessment of cross-linked residues in both states revealed significant changes upon phosphorylation (Fig. 8d). Initial inspection of the identified cross-links for each of the functional states represented in circular views (Fig. 8d) pointed at the activation loop as a hotspot defining the unstimulated or resting state of CCDC6-RET, which involved a series of crosslinked peptides containing activation loop Lys 907 that disappear in the active phosphorylated state of the protein (Fig. 8d, e). In particular, we pay attention to three crosslinked peptides that coordinated Lys 907 (peptide DVYEEDSYVK*R) from the activation loop with other residues from the activation loop itself or its vicinity, for example Ser 909 (S*QGRIPVK), Ser 896 (ISDFGLS*R) and also Lys 889 (MK*ISDFGLSR) just at the beginning of the activation segment defined by the DFG motif (Fig. 8d, e and Supplementary Fig. 11). When we look at the distances between pairs at the activation segments (e.g., K907-S909 and K907-S896) in a CCDC6-RET dimer assembly corresponding to the unstimulated (i.e., resting) estate (Fig. 8f), we observed that distances between pairs were compatible with intermolecular crosslinks rather than intramolecular ones considering the size and range of the DSBU crosslinker molecule (12.5-27.5 Å). These results are in line with a face-to-face arrangement of both catalytic domains defined by intermolecular crosslinked activation segments within the dimer in the inactive state (Fig. 8e, f). In the presence of nucleotide and MgCl2, the catalytic domains swing apart, as indicated by the loss of activation loop crosslinked peptides, adopting a configuration in which both kinase domains are facing towards opposite directions from the stem of the coiled-coil. Furthermore, we showed on the two 3D-structural models the specific crosslinked peptides that defined the two functional and structural states, and the percentage of crosslinks within distance for each of them (Fig. 8g). The crosslinked peptides that defined the topology of the two functional and conformational states were clearly distinct, with one main central node of crosslinked peptides arising from activation loop Lys 907 in the inactive (unstimulated) state. In the case of the active (auto-phosphorylated) state, there were two main nodes identified, one at Lys 965 just below the substrate binding pocket, and another at a Lys 747 on the β2 of the G-loop. Mapping all the detected crosslinks in both structural models revealed a percentage of crosslinks within distance (27.5 Å cutoff) of 63.5% and 61.9% for the unstimulated and phosphorylated states, respectively (Supplementary Fig. 11d, e). The subset of distance-compatible crosslinks and their clustering on specific structural elements is consistently observed across replicates and functional conditions, further supporting the internal consistency of the dataset. These data together demonstrate significant conformational changes during the mechanisms of CCDC6-RET autoactivation, and at the same time validate the internal consistency of the XL-MS dataset and its suitability as a structural restraint for the proposed models.

a Top 20 precursor id list from a tryptic non-crosslinked CCDC6-RET sample represented in column chart. Size exclusion chromatography using a Superdex 200 3.2/300 of non-crosslinked CCDC6-RET sample (unstimulated, blue), and crosslinked (XL) unstimulated (green) and autophosphorylated (red) CCDC6-RET samples, inset. b Crosslinks Merox count for both peptide spectrum match (PMS) and peptides identified in the unstimulated and phosphorylated samples of CCDC6-RET. c Venn diagram showing specific crosslinked peptides for each of the functional states. d Identified cross-links specific for the unstimulated and phosphorylated states are represented in circular views. In green, highlighted cross-links involving residues from the activation loop specific for the resting state. In dark orange, highlighted crosslinks involving residues from the activation loop specific to the phosphorylated state. e Heat map (Z-scores) of crosslinked peptides involving activation segment residues for both states in triplicate samples (n = 3). Crosslinked residues are highlighted in red. f Distances between identified activation loop crosslinked pairs (K907-S896) and (K907-S909) filtered on the structural Alpha-Fold model for the unstimualted form of a CCDC6-RET dimer. Residue numbering corresponds to CCDC6 (UniProt: Q16204) and RET (UniProt: P07949-2) aa sequences. g Specific crosslinked peptides for the unstimulated and phosphorylated states of CCDC6-RET mapped onto their 3D-structural models with the indicated key nodes and percentage of crosslinks within distance (27.5 Å). Intermolecular (magenta) and intramolecular (yellow) crosslinked peptides are depicted.
MD simulation analyses of inactive and active (phosphorylated) CCDC6-RET
We generated structural models for an active CCDC6-RET homodimer (accession codes: ma-pcoh8 and ma-b7la0) in complex with ATP, ADP, Mg2+ and a Tyr-containing peptide substrate (Fig. 9b) that provided a plausible structural explanation for ADP-dependent catalysis (Fig. 4f). To assess the dynamic feasibility of the proposed structural models and peptide binding modes, MD simulations (300 ns) were run for the unstimulated (resting) and phosphorylated (active) states of CCDC6-RET. In the latter case, either ATP or ADP was placed on each of the active sites in complex with Mg2+ in addition to the phosphorylatable tyrosine-containing peptide (Fig. 9a). The results obtained support our proposed model in terms of stabilizing interactions and conformational changes that define both states. Importantly, the results also allowed us to identify the key residues and secondary structural elements implicated in catalysis that define the dual-specificity on Tyr and Ser substrates in the same active site (Fig. 9b). For the Tyr-containing substrate, we find the prototypical interactions required for catalysis with an open G-loop, defined by the side chain of Glu 734 forming a salt bridge with the side chain of A-loop Arg 912 (Fig. 9b, upper inset). In addition to the catalytic salt bridge (β3 Lys 758- αC Glu 775), the carboxylate of Asp 874 (HRD motif) is well positioned for proton abstraction from the hydroxyl of the phosphorylatable Tyr residue. This key interaction in complemented by the side chains of Arg 878 (held in position by means of a key cation-π interaction with the indole ring of Trp 917) and Asp 879 (β6-β7 loop) to ensure proper γ-phosphate transfer (Fig. 9b, upper inset). In the case of the Ser specificity, we observed that for the Ser residue in the phosphorylatable peptide to reach deeper into the active site, the activation loop has to undergo a large conformational change that appears to be driven by a critical electrostatic interaction between the ATP’s γ-phosphate and the side-chain guanidinium of Arg 912 (Fig. 9b, lower inset). In this setting, Ile 913, Pro 914 and Lys 916 form key contacts with main and side-chain residues flanking the phosphorylatable Ser site.

a Ensemble of overlaid CCDC6-RET dimers obtained from the MD simulation (300 ns) for both the unstimulated (inactive) and active (phosphorylated) states, respectively. For simplicity, nucleotides ATP, ADP, and Mg2+ are not depicted. Representative frames (6) were chosen to recapitulate each of the functional states based on initial AlphaFold models, in (b). Rainbow-colored ribbon representation. b Initial frames from refined AlphaFold models (see material and methods) for the inactive and active states of CCDC6-RET (accession code: ma-b7la0). In the active state, nucleotides ATP and ADP, Mg2+, and phosphorylatable peptide (EAIYAAPFAKKK, in wheat color) are shown for each chain. Snapshot of a tyrosine-phosphorylatable peptide in the active site of CCDC6-RET in complex with ATP and Mg2+ showing key implicated residues in sticks (upper inset). The equivalent snapshot in the active site of RET kinase (Supplementary Table 6) with a serine-phosphorylatable peptide (EEDSYV, pink) is also shown, lower inset. c Protein-protein Interface analyses between chain A and B for the unstimulated and active (phosphorylated) states. Schematic diagram of interactions between protein chains A and B. Interacting side chains residues are joined by colored lines, each representing a different type of interaction, as per the indicated key. The highlighted area of each circle is proportional to the surface area of the corresponding protein chain interface. The extent of the interface region on each chain is represented by the black wedge whose size is proportional to the interface surface area. Statistics for this interface are given in the small table below, together with the indicated color-coded corresponding to the type of interaction. d Schematic representation of the proposed model for the mechanisms of CCDC6-RET autoactivation, see also Fig. 8g. Note, both IDRs N- and C-terminal sequences are not included in the models.
Next, to identify and quantify the key contacts between chains that define the two specific configurations of CCDC6-RET, we used the PDBsum server40. Because of a more compacted arrangement of both kinase domains and their interface, a larger number of amino acids interacting from both chains A-B were found in the case of the inactive form (60–59 aa) than the phosphorylated state (37–36 aa). This was the result of a much tighter interface found in the inactive (3084–3136 Å) than in the phosphorylated form (2106–2121 Å), as indicated also by the larger number of salt-bridges and hydrogen bonds found (Fig. 9c and supplementary Fig. 12). These MD simulation data together with the biochemical and structural information supported our proposed model and conformational switch that defines a transition from the inactive to the active (phosphorylated) state in CCDC6-RET during the process of autoactivation and signaling (Fig. 9d).
Discussion
In this work we uncover the molecular and structural determinants that control the autoactivation mechanism of the CCDC6-RET fusion protein, a driver and therapeutic target in lung adenocarcinoma and papillary thyroid cancer15,16,17. The RET/PTC1 (i.e., CCDC6-RET) oncogene was discovered more than 30 years ago by the lab of Giancarlo Vecchio9, and despite the existence of a significant number of published studies on the genetics and signaling of this oncogene, the molecular and structural bases for its (oncogenic) mechanism of action have remained largely elusive. Here, we biochemically dissected its autoactivation mechanism and found that CCDC6-RET is a highly active dimeric kinase in solution (Fig. 1). The very high autophosphorylation kinetics observed for many of the identified CCDC6-RET phospho-sites, together with the low KM values for ATP, are indicative of a lack of specificity. These data contrast with the sequential and ordered pattern of RET ICD autophosphorylation observed for the wild-type protein29. Furthermore, the CCDC6-RET fusion product functions not only as a kinase with dual specificity for tyrosine and serine residues (Fig. 2, Table 1 and Supplementary Tables 1–4); but also, strikingly as a dual ATP- and ADP-dependent kinase, able to bind and use both nucleotides as phosphoryl donors (Figs. 3 and 4). We also found a crosstalk between the C-terminal (short isoform), and the activation segment, uncovering a mutually exclusive dependency on activation loop phospho-sites controlling both the processing (via Tyr 900) and the catalytic activity (via Tyr 905) of the fusion protein (Fig. 5 and Supplementary Fig. 5). Our data suggest that RET activation loop Tyr 900 phospho-site is required for the adequate synthesis and folding of the protein in a process that appears to be dependent on the integrity of the whole catalytic core of the fusion, and in particular on the C-terminal segment (Supplementary Figs. 4 and 5). These findings suggest that a yet unidentified element from the C-terminal segment (specific to the short C-terminal isoform 9) makes key contacts with (phosphorylated) Tyr 900 at the activation loop in the unstimulated (resting) state; and these contacts appear to be critical for the stability and folding of the fusion protein. The detrimental impact caused by a non-phosphorylatable Tyr 900 mutant (Y900F) is strikingly rescued by the concomitant mutation of the other activation loop Tyr 905 autophosphorylation site. Note that activation loop Tyr 900 appears already phosphorylated at time zero in the unstimulated state, and upon CCDC6-RET autoactivation its phosphorylation levels correlate inversely with the phosphorylation state of activation-loop Tyr 905, which undergoes fast and robust autophosphorylation in a time dependent manner (Fig. 2 and supplementary Fig. 2). This site (Tyr 905) is catalytically required to phosphorylate a substrate surrogate (i.e., peptide), see Fig. 5. In this active setting, phosphorylation on C-terminal sites will release the C-tail to act as a scaffold or signaling platform to assemble signaling complexes. This is also supported by the fact that a C-terminal deleted construct appears to be more efficient catalytically (supplementary Fig. 4). We hypothesize that the lack of availability of the C-terminal segment mimics its own engagement in the signaling assembly and transduction, where the activation loop Tyr 905 is catalytically required.
Furthermore, we generated a 3D-assembly of a CCDC6-RET homodimer combining an integrated structural biology and XL-MS approach. We demonstrated that, in accordance to our structural reconstruction, CCDC6-RET adopts a face-to-face dimer configuration in the unstimulated state, and upon nucleotide binding and conformational readjustment, activation loop autophosphorylation could be driven by a mechanism in cis (Figs. 8 and 9). These data were unexpected in the first place, as we anticipated that a constitutive (forced) protein kinase dimer would undergo direct intermolecular (i.e., from one chain to the other) autophosphorylation; but at the same time compatible with the domain arrangement seen in bacterial histidine kinases, where a catalytic domain situated to the left of the coiled-coil helix b (i.e., to the left of the four-helix bundle and 2-fold symmetry plane) undergoes cis-autophosphorylation34. Recent studies demonstrated that PKD autoinhibition in trans is compatible with and controlled by an activation loop autophosphorylation mechanisms in cis (as a monomer)41. In another relevant study, they show that PDK1 is autoinhibited by its PH domain intramolecularly (in cis) in the inactive state and that, upon PIP3 binding, positive cooperativity promotes a switch-like activation of PDK1 driving intermolecular PDK1 autophosphorylation by the assembly of a face-to-face dimer42. Our results are in line with these recent studies that highlight altogether the existence of complex regulatory elements during the autoactivation mechanism, which are controlled by intra- and inter-molecular components that appear to be rather specific and private for each kinase43.
The dual ATP- and ADP-dependency seen by CCDC6-RET in vitro is a striking mechanistic feature that appears to be rather specific compared with the wild-type kinase (Fig. 3f), and with other RET fusion products (RIα-RET and KIF5B-RET, work in progress), because ADP-dependent kinases are usually very restrictive to their ligands being unable to use tri-phosphorylated nucleotides such as ATP44,45,46. From a biological point of view, the dual ATP and ADP dependency seen for the CCDC6-RET fusion product would result in a gain of functionality by being able to perform two catalytic cycles from one single ATP molecule; transferring both the γ- and β-phosphate groups of the initial ATP and its immediate product ADP, to itself (autophosphorylation) and/or to different substrates surrogates. In normal cells, ATP concentrations are maintained between 1–10 mM, while ADP levels are lower (20–100 µM), yielding an ATP/ADP ratio of ~100–50047,48. In contrast, cancer cells often exhibit suppressed mitochondrial metabolism and enhanced glycolysis (i.e., the Warburg effect), leading to reduced ATP levels and ATP/ADP ratios as low as 5–20, with ADP reaching 100–200 µM or more49. Thus, tumor cells display ADP/ATP ratios of 0.05–0.2, much higher than in normal cells. Under significant ATP-limiting conditions, ADP can accumulate to comparable concentrations to those of ATP (≈500 μM)50 and under these conditions, it is plausible that ADP can be utilized as an alternative phosphate donor by certain protein kinases.
We further hypothesize that local concentration effects, such as the formation of CCDC6–RET condensates or lipid phase separation, could locally enrich ADP and thereby enable ADP-dependent phosphorylation. Similar mechanisms have been reported for oncogenic kinase fusions like EML4–ALK, which forms condensates that compartmentalize signaling51, and for ATP-driven phase separation of disordered basic proteins (Kota et al., bioRxiv 2023.08.20.554035). To test this working hypothesis, we are developing cellular systems that allow direct manipulation of the ATP/ADP ratio using genetically encoded tools such as ATPGobble (Ekvik et al., bioRxiv 2025.08.12.670003). These models will enable us to reduce the ATP/ADP ratio even further and assess ADP-dependent phosphorylation under controlled, physiological conditions (work in progress). Finally, recent studies describe similar ADP-driven phosphorylation in MAP2Ks and IKK52,53, supporting the emerging concept that ADP-dependent activity, though secondary under normal conditions, may act as an adaptive mechanism during energy limitation. This could have important implications for how (cancerous) cells maintain signaling functions under stressful conditions such as low oxygen or ischemia, which are common in diseases like cancer and atherosclerosis. In this line, we are currently undertaking substrate identification in cells54 using a whole-cell lysate kinase assay coupled with LC/MS-MS under saturating concentrations of both nucleotides ATP and ADP (work in preparation). Our preliminary results indicate that CCDC6-RET exert a different pattern of phosphorylated substrates in cells; also showing a higher activity (i.e., higher number of phosphorylated substrates identified) when ADP was used to trigger the phosphorylation reaction as compared to ATP. The incremental number of phosphorylated peptide substrates is explained, however, not by an increase in the phospho-tyrosine activity of CCDC6-RET, but by a large increment in the appearance of Ser-containing phosphorylated peptides. These data suggest that ADP-dependent activity by the CCDC6-RET fusion product could be coupled to a change in substrate specificity towards Ser and Thr rather than Tyr.
There is a striking lack of homology and fold-conservation between RET (and human protein kinases in general) and human ADP-dependent kinases, taking for example, the ADP-dependent glucokinase (ADPGK) as a main paradigm46. ADP-dependent kinases are classified as members of the ribokinase superfamily44,45. The ribokinase-like fold is basically composed of an eight-stranded β-sheet surrounded by eight α-helices, three on one side and five on the other which constitutes a single domain (the large domain). This superfamily initially included ATP-dependent kinases of adenosine, fructose, tagatose-6-P, fructose-6-P, and fructose-1-P among others, besides ribokinase, the canonical enzyme55. Later, the ribokinase superfamily also included enzymes that can transfer the γ-phosphate of ATP to some vitamins involved in B6 synthesis, such as pyridoxal kinase56. This superfamily can be subdivided into three major groups: the ATP-dependent vitamin kinases, the ATP-dependent sugar kinases, and the ADP-dependent sugar kinases. The main structural difference between these groups is related to the presence of a second (small) domain composed of a β-sheet, with some α-helical insertions in the case of the ADP-dependent kinases and ATP-dependent kinases, while the vitamin kinase enzymes present only the αβα ribokinase-like fold (the large domain). In the case of a CCDC6-RET homodimer, the arrangement of both catalytic domains in the active configuration of the homodimer facing into opposite directions, can explain a higher degree of structural malleability within the active site. The lack of steric impediments by the other chain in the case of the intermolecular process will allow a deeper access into the active site by the substrate to be in range from the β-phosphate group of the ADP molecule (Fig. 4f and Fig. 9b). Leucine-zipper motifs are one of the main structural motifs known to interact with DNA elements. Note that the coiled-coil interface of CCDC6-RET resembles highly of a leucine-zipper motif characterized by a Leu in position d of the heptad repeat, which in the case of the RET fusion is formed by Leu 59, Leu 66, Leu 73, Tyr 80, Leu 87, Leu 94 and Ile 101. The connection of CCDC6-RET with the DNA-damage response pathway, its interaction with DNA-elements and protein kinases and other effectors implicated in this fundamental process are yet to be elucidated. Interestingly, the RET S909 autophosphorylation site is within an ATR/ATM28 consensus site (pS/TQG), being potentially an intermolecular regulatory element. Contrary to the case of the wild-type RET28, this residue when mutated appears to have a significant effect on the catalytic activity of the RET fusion product towards a substrate despite lacking any effect on autophosphorylation (supplementary Figs. 1 and 6). This is in fact clear evidence that autophosphorylation and the phosphorylation of an exogenous substrate (e.g., a peptide, or an intact substrate surrogate) are not controlled by the same mechanistic determinants. Thus, even though both are used as readouts for catalytic activity, they should be treated as independent mechanisms or processes28,43,57.
Our biochemical data allowed us to discriminate between protein kinases that can (e.g., c-Src) or cannot (e.g., CCDC6-RET) phosphorylate an inactive substrate surrogate that mimics its own kinase domain (Supplementary Fig. 10). However, while this excludes interdimer phosphorylation, the exact cis/trans relationship within the dimer remains unresolved. In addition, the crosslinking mass spectrometry data filtered under our reconstructed 3D-structural models recapitulate the different conformational and functional states in the autoactivation mechanisms of CCDC6-RET (Fig. 8d–f). Despite the resolution limitation of our EM data (Figs. 6 and 7), our integrative approach, including now MD simulation data over 300 ns provided a feasible model compatible with a face-to-face arrangement of both RET kinase domains within the homodimer in the inactive state (Fig. 8 and Fig. 9). In the presence of nucleotide and MgCl2, the catalytic domains swing apart (as indicated by the breaking up of the activation loop crosslinked peptides) orienting their active sites away from each other-compatible with each kinase phosphorylating its own activation loop. The reorientation of the catalytic domains is coordinated with the motion of the coiled-coil interface in a way that resembles a ¨chopstick¨ model, where a tighter base at the coiled-coil grasps the two catalytic domains together into a compacted face-to-face dimer (Figs. 8 and 9). In the active state, the contacts at the base of the coiled-coil interface are loosened, resulting consequently in a more relaxed and open interface at the base of the coiled-coil and a more proximal amino-terminal sections of the dimerization interface compatible with the motion (swinging) of the catalytic domains (Figs. 8e–g and 9, and supplementary Fig. 12). However, we acknowledge that the transition from the inactive (i.e., resting state) to the active (phosphorylated state) may not be driven by nucleotide binding only, and that at the very early stages, an intermolecular phosphorylation process may be causing the transition. To overcome this limitation, we are currently trying to obtain higher resolution information by cryo-EM studies and X-ray crystallography that will allow us to solve with near atomic-resolution structures of the different functional and conformational states of CCDC6-RET, including the transition state, to fully support our mechanistic model. All in all, our work uncovers for the first time the molecular and structural determinants that control the mechanisms of CCDC6-RET autoactivation and provides at the same time a solid framework for the structural and functional analyses of other RET fusion products.
Methods
Plasmids
A modified pBac-PAK3 plasmid (Invitrogen) containing an N-terminal 6 x His and a GST (Glutathione S-transferase) tags followed by a Human Rhinovirus (HRV)-3C protease cleavage site was used to express a codon optimized CCDC6-RET fusion product composed by fragments of residues 1-101 of human CCDC6 and 713-1072 of human RET27,28,29. A construct lacking the C-terminal segment of RET (RET aa 713-1013, i.e., ΔCT) was also generated. These plasmids were used to create different point mutants by site-directed mutagenesis.
Site-directed mutagenesis
Site-directed mutagenesis was performed by the Q5-site directed mutagenesis kit (New England Biolabs) following manufacturer instructions and the indicated non-overlapping primers:
RET_Y900F- forward 5’- CCGTGACGTGTTCGAGGAGGACT -3’
RET_Y900F- reverse 5’- CTCAGTCCGAAGTCGGAG -3’
RET_Y905F- forward 5’- GGAGGACTCCTTCGTCAAGCGTT -3’
RET_Y905F- reverse 5’- TCGTACACGTCACGGCTC -3’
RET_Y1015F- forward 5’- CGTCGTGACTTCCTGGACCTG -3’
RET_Y1015F- reverse 5’- CTTGACCATCATCTTCTCCAG -3’
RET_Y1062F- forward 5’- AACAAGCTCTTCGGTCGTATCTC -3´
RET_Y1062F- reverse 5’- CTCGATCCAGGTGGAGGG -3´
RET_S1034A_forward 5´- CGACGGACTGGCCGAGGAGGAGA -3´
RET_S1034A_reverse 5´- TCGTAGATCAGGGAGTCGGAGG -3´
RET_S98A_forward 5’-GCGCAAGGCTGCGGTGACTATCG -3´
RET_S98A_reverse 5’- AGGTCGCGGTTTTCC -3
CCDC6_S52A forward 5’- AATCGTGATCGCCCCATTCCGTC -3´
CCDC6_S52A reverse 5’- CCACCAGATTTTCCACCG -3´
CCDC6_C84A_forward 5’- CAAGCTGAAGGCCAAGGCTCTGC -3´
CCDC6_C84A_reverse 5’- TAGGTTTCCAGCTCGATC -3´
CCDC6_Y80F_forward 5´- GCTGGAAACCTTCAAGCTGAAGT -3´
CCDC6_Y80F_reverse 5´- TCGATCTTCAGCACCTTG -3´
RET_D874A forward 5´- GGTGCACAGAGACCTGGCCGCCA -3´
RET_D874 reverse 5´- AGCTTCATCTCGGCCAGGTACTGCATG -3´
Alternatively, other mutations were generated by using overlapping primers:
RET_S909A forward 5’- CGTCAAGCGTGCCCAGGGTCGTA -3
RET_S909A reverse 5’- TACGACCCTGGGCACGCTTGACG -3´
CCDC6_S46A forward 5’- CGGTGGAAAAGCTGGTGGAATCGTGATCTC -3´
CCDC6_S46A reverse 5’- GAGATCACGATTCCACCAGCTTTTCCACCG -3´
Heterologous protein expression in insect cells
We used the flashBACTM system (Oxford Expression Technologies) for recombinant protein expression using a baculovirus system27,28,29 following the manufacturer recommendations. Briefly, Sf9 insect cells (Oxford Expression Technologies) were seeded in a 6-wells/plate at a density of 0.3–0.5 × 106 cells/mL in serum-free Sf900-II media (Gibco) supplemented with 200 μL of Gentamycin (50 mg/mL) per liter. Next, the transfection reaction was prepared in a sterile Eppendorf tube containing antibiotic-free media Sf900-II (up-to 1 mL) and the transfection mixture composed of: 2.5 μL of flashBacTM linear viral DNA (20 ng/ μL), 1 μL (500 ng) of transfer plasmid (pBacPAK3) and 5 μL of transfection reagent (Fugene) following manufacturer instructions. After 5–6 days, the supernatant of the transfected cells was collected to have the recombinant baculovirus stock (P1). The P1 baculovirus was inoculated (1:25) in a suspension culture of 25 mL of complete Sf900-II media at a density of 1.5–2 × 106 cells/ml and incubated shaking at 90–95 rpm for 3–4 days. After this time, the supernatant was recovered, obtaining the P2 baculovirus stock. After testing the expression of the protein, the P2 baculovirus was used to inoculate another 100 mL suspension culture of Sf9 cells as indicated before at a 1:100 dilution. After 3-4 days (72–96 h), the supernatant became the P3 baculovirus stock. Both P2 and P3 baculovirus were used as baculovirus stocks for protein production.
Protein purification
Protein purification was achieved by in tandem immobilized metal ion affinity (Ni+2) and Glutathione-conjugated gravity flow chromatography and in-gel 3C-protease digestion followed by size exclusion chromatography if required. This optimized protocol resulted in a highly pure and monodisperse recombinant fusion product. Inoculated insect cell cultures (72–96 h) were harvested by centrifugation at 1800 g (Eppendorf Centrifuge 5810R) during 10 min at 4 °C. Pellets were stored at −80 °C or used directly for protein purification. Pellets were resuspended in lysis buffer (50 mM Tris pH 7.5, 500 mM NaCl, 10 mM Benzamidine, 0.2 mM AEBSF, 2 mM TCEP). After sonication on ice (20 cycles 9 s on/3 s off and 37% amplitude), crude lysates were centrifuged at 33,000 g/48,000 g (JA 25-50 rotor, Beckman Coulter Avanti J-25) at 4 °C for 45 min. Clarified lysate underwent a final sonication step of 10 s followed by filtration with a 45 μm filter (Jet Biofilter) with a syringe. Crude lysate, clarified lysate (supernatant) and insoluble material (pellet) samples were mixed with 5× Laemmli sample buffer (Thermo Fisher Scientific) and run on SDS-Polyacrylamide gel electrophoresis (SDS-PAGE). First, the clarified lysate was incubated with a previously equilibrated Nickel (Ni2+) chelate resin. After 2 h incubation on rotation in a cold room, the flow through (FT) was separated and beads were washed with Buffer A (20 mM Tris pH 7.5, 500 mM NaCl, 2 mM TCEP) with an additional 20 mM Imidazole. Protein elution was done with Buffer B (20 mM Tris pH 7.5, 500 mM NaCl, 400 mM Imidazole, 2 mM TCEP) using a filter spin column (SC 1000-1KT, Lot SLCD4489, SIGMA-ALDRICH) by centrifugation at 6000 g (Eppendorf centrifuge 5415D) during 2 min. Fractions were checked by SDS-PAGE stained with Coomassie previously to the second purification step. Positive fractions eluted from the His-affinity step were incubated with Glutathione conjugated Sepharose beads (Glutathione Sepharose® 4 Fast Flow, Cytiva) for several hours at 4 °C. Next, the GST-tagged protein was separated by centrifugation at 160 g (Eppendorf Centrifuge 5810 R) and washed 4–5 times with 10 mL of GST-Buffer (20 mM Tris pH 7.5, 500 mM NaCl, 2 mM DTT) prior to in gel digestion (3-4 hours) with an GST-3C protease (200 μg/L culture) and elution (2–3) using a filter spin column at 6000 g for 2 min (Eppendorf centrifuge 5415D). Eluted fractions were checked by SDS-PAGE gel stained with Coomassie. Protein concentration and purity were measured by absorbance using a Nanodrop. When required, a size exclusion chromatography (SEC) step using AKTA pure protein purification system (GE Healthcare) was performed. The sample was concentrated up to 2 mL with a Millipore concentrator with a 30 kDa cut-off by centrifugation at 800 g (Eppendorf Centrifuge 5810 R) at 4 °C. The concentrated sample was injected into a Superdex 200 16/60, 10/300 or 3.2/300 columns (GE Healthcare). The input of the column together with the collected fractions were analyzed by SDS-PAGE followed by Coomassie staining, and the protein concentration was measured by absorbance.
SDS-PAGE, western blotting, and antibodies
Proteins were electrophoretically separated on 12% SDS-PAGE gels run in the Mini-PROTEAN system with electrophoresis buffer (25 mM Tris pH 8.3, 192 mM glycine, 0.1% SDS) by using a BIO RAD PowerPac Basic at a constant voltage of 150–200 mV for 30–45 min. Pre-stained dual color markers (Precision Plus Protein Standards 10-250 kDa, BIO-RAD and Protein Ladder PS10 Plus 11-180 kDa, GeneOn) were used as molecular weight standards. For protein staining, gels were gently shacked in Coomassie staining solution (10% acetic acid, 40% absolute ethanol and 50% MilliQ water with 1 g/L Brilliant blue 250R) for 30–60 min. Then the gels were distained for 1–2 h with a distaining solution (10% acetic acid, 50% absolute ethanol, 40% MilliQ water). Proteins were transferred from SDS-PAGE gels onto nitrocellulose 0.2 μm (Amersham) or PVDF 0.45 μm (Millipore) membranes using the Mini-PROTEAN system (BIO RAD). Before transference, PVDF membranes were activated first in ethanol 100% for 10 min. Membrane were wet-transferred in transfer buffer (25 mM Tris pH 8.8, Glycine 190 mM and ethanol 10%) on ice at a constant 100-200 V for 60 min. Next, transferred membranes were blocked in TBS (10 mM Tris pH 8, 150 mM NaCl) supplemented with 5% (w/v) skimmed powder milk and gently shacked for 30 min. After blocking step, membranes were washed in TBS-T (10 mM Tris pH 8, 150 mM NaCl Tween-20 0.1% v/v) for 3-4 times. Then they were incubated with constant agitation (Duomax 1030, Heidolph) O/N at 4 °C with the following primary antibodies diluted in TBS-T supplemented with 5% BSA (w/v): phospho-Tyr (pTyr-100, CST #9411), RET (D3D8R, CST #14698), phospho-Ret Tyr 905 (CST #3221), phospho-Ret Tyr 1015 (Abcam # 74154) and phospho-Ret Tyr 1062 (Abcam # 51103) at 1:5.000 or 1:10.000 dilution. After overnight incubation with the primary antibody, membranes were washed with TBS-T for 3-4 times and incubated with the secondary antibody for 60 min with gently shaking protected from light. Secondary antibodies used were anti-rabbit IgG (H + L) conjugated at 800 nm (DyLight 800 Conjugate #5151, CST) and anti-mouse IgG (H + L) conjugated at 680 nm (DyLight 680 Conjugate #5470, CST). These secondary antibodies were diluted 1:20.000 in TBS-T with 5% (w/v) skimmed powder milk. After 60 min of secondary antibody incubation, membranes were washed with 20 mL of TBS-T for 3-4 times. Finally, proteins were detected in the membranes with the Odyssey CLx scanner (LI-COR Biosciences) and the images were analyzed with the Image Studio lite software.
Differential scanning fluorimetry
To determine the thermal stability of the recombinants proteins we apply a direct method of Differential Scanning Fluorimetry (DSF) based on intrinsic fluorescence (IF) detection using a Tycho NT.6 instrument (NanoTemper Technologies). Briefly, a protein sample (1–2 μM, 12 μL) was loaded into a glass capillary tube and heated from 35 °C to 95 °C at a rate of 30 °C/min. The inflection temperatures (Ti) were calculated by the instrument based on the 350/330 nm fluorescence ratio. We measure the increment in thermal stability in the absence or presence of different ligands and inhibitors, as indicated in the text.
Mass Photometry
To determine the molecular mass distributions (40 kDa to 5 MDa) of single molecules we applied light scattering to detect individual, unlabeled molecules in dilute solutions using a MPone instrument (Refeyn). All the events were recorded with Refeyn Acquire MP software and analyzed with Refeyn Discover MP software to obtain mass distribution of protein samples.
In vitro phosphorylation assays
For autophosphorylation experiments, recombinant protein 1–2 μM (final concentration) was used in 20 mM Tris pH 7.5, 500 mM NaCl, 2 mM DTT and 2 mM MgCl2. We performed a time-points reaction triggered by the addition of ATP (1 mM): unstimulated, 1, 5, 15, 30 min. To stop the phosphorylation reaction samples were mixed with 5X Laemmli sample buffer and denaturalized at 95 °C during 2–5 min in a thermoblock (AccuBlock, Labnet). In the case of phosphorylation experiments using intact substrate surrogates (catalytic dead) a 1:3 molar excess of the substrate to the enzyme was employed in the reaction. For the nucleotide titration experiments the phospho-tyrosine activity of CCDC6-RET was monitored under increasing concentrations of nucleotide (from 10 to 5000 μM) after 1–15 min stimulation.
For the time course autophosphorylation assays using radiolabeled nucleotides we prepared an in vitro time course autophosphorylation reaction as follow: for ATP (γP32, Revvity) we performed a 0–30 min time course assay in elution buffer 20 mM Tris pH 7.5, 500 mM NaCl, 2 mM DTT and 4 mM MgCl2 complemented with kinase (2.5 μM), ATP (2 mM) and ATP γP32 (50 μCi). In the case of the radioactive assay with ADP (βP33, Hartmann Analytics), for the time course of the autophosphorylation reaction (0–60 min) buffer was complemented with kinase (10 μM), ADP (1 mM) and ADP βP33 (60 μCi). Samples were mixed with 5X Laemmli sample buffer and denaturalized at 95 °C during 2–5 min before being run on a 4–12% precast bis-tris polyacrylamide gels (Invitrogen, Thermo Fisher Scientific). Electrophoresis was run at 200 V, after which the gels were introduced in a exposure cassette with a phosphor screen for 2–4 h first, and then overnight for a longer exposure if required. Screens were developed in an Amersham Typhoon Biomolecular scanner using the phosphor imaging option using 4000 V and 100 µM of pixel size resolution.
Enzyme kinetic assays
Rates of phospho-tyrosine activity by recombinant CCDC6-RET and its variants were determined by a continuous ATP-coupled pyruvate kinase essay based on the consumption of NADH in the absence and presence of different peptide substrates at increasing concentrations of ATP28,29,58. The enzyme-substrate solution (1 mM Tris-Cl pH 7.5, 1 mM MgSO4, 400 mM phosphoenol pyruvate (PEP), 100 mM NADH, 2450 U/mL Pyruvate kinase, 2.26 mg/mL Lactate dehydrogenase) was prepared at different concentrations of ATP (mM): 0, 0.08, 0.16, 0.32, 0.65, 1.25, 2.5, and 5. Unless otherwise stated, the final protein concentration used in these assays was 0.2–1 μM, and the concentration of the peptides used EAIYAAPFAKKK and the RET A-loop DVYEEDSYV were 1.35 and 0.55 mM, respectively. The experiments were performed in a 384 well plate (Greiner bio-one) and the NADH consumption was read at 340 nm wavelength during 120 cycles of 30 s each by using a microplate reader (CLARIOstar Plus, BMG LABTECH). NADH consumption curves were obtained, and slopes were calculated by using de CLARIOstar Data Analysis software. Finally, by using Prism software, catalytic rates and kinetic constants were obtained by fitting the data to the Michaelis-Menten equation.
Mass spectrometry
For liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS) samples (2.5–5 μM) were reduced and alkylated in 15 mM TCEP and 30 mM CAA for 30 min at room temperature in the dark. Proteins were then digested overnight at 37 °C with trypsin (Promega) in 1 M urea, 50 mM Tris-HCl pH 8.0, using an estimated protein-to-enzyme ratio of 100:1. Digestion was quenched with 0.1% TFA and the resulting peptides were desalted using C18 stage-tips. LC-MS/MS analysis was performed on a Q Exactive Plus mass spectrometer (Thermo Fisher Scientific) coupled to an UltiMate 3000 RSLCnano system. Digested peptides were loaded into a trap column (Acclaim™ PepMap™ 100 C18 LC Columns 5 µm, 20 mm length) for 3 min at a flow rate of 10 µL/min in 0.1% formic acid, and then separated on an EASY-Spray PepMap RSLC C18 column (2 µm, 75 µm x 50 cm, Thermo Fisher Scientific) operated at 45 °C using a 60 min effective gradient (buffer A: 0.1% FA; buffer B: 100% ACN, 0.1% FA) at a flow rate of 250 nL/min. The gradient used was, from 4% to 6% B in 2 min, from 6% to 33% B in 58 min, plus 10 additional min at 98% B.
Peptides were ionized using an EASY-Spray source at 1.5 kV with a capillary temperature of 300 °C. The mass spectrometer was operated in a data-dependent acquisition mode (DDA using a top 15 method with an intensity threshold of ≥4.5 × 104, a dynamic exclusion of 5 or 10 s and excluding unassigned, +1 and >+6 charged ions. Full MS scans (350–1500 m/z) were acquired at 70,000 resolution (FWHM at 200 m/z), followed by MS/MS scans at 35,000 resolution using higher-energy collisional dissociation (HCD, NCE 27). The isolation window was 2.0 Th. Ion targets were 3 × 10⁶ for MS (max IT 25 ms) and 1 × 10⁵ for MS/MS (max IT 110 ms).
Raw files were processed with Maxquant (v 1.6 and higher) using the default settings against a database containing the sequences of the recombinant proteins. Carbamidomethylation of cysteines was set as a fixed modification whereas oxidation of methionines, acetylation and phosphorylation of serines, threonines and tyrosines were set as variable modifications. Minimal peptide length of seven amino acids and a maximum of two tryptic missed cleavages were allowed. Results were filtered at 1% FDR at both peptide and protein levels. Raw data were imported into Skyline. Quantitative analysis was performed in Skyline. Label free quantification of phosphopeptides was performed by integrating the extracted ion chromatogram (XIC) of the isotopic envelope. Only interference-free peaks were used for quantification. Phosphopeptides intensities were normalized by the intensity of the non-modified peptides of the target protein. Data shown are from four independent autophosphorylation experiments with two technical replicates each (see supplementary Tables 1–4). Phospho sites that are consistent across the different datasets are summarized in Table 1.
Crosslinking mass spectrometry
Both phosphorylated (1 mM ATP, 2 mM MgCl2, 30 min) and un-phosphorylated CCDC6-RET (6 µM) were crosslinked with 200-fold molar excess of disuccinimidyl dibutyric urea (DSBU, Thermo Fisher Scientific) in 20 mM HEPES pH 7.5, 500 mM NaCl, 1 mM TCEP. All the crosslinking reactions were performed at RT for 50 min and then quenched with 20 nM NH4HCO3. The cross-linked samples in triplicate were vacuum-dried and resuspended in 4% SDS in 50 mM Tris/HCl pH 8.0. Reduction and alkylation were performed with 15 mM TCEP and 25 mM chloroacetamide 30 min at 45 °C, respectively. Samples were then trypsinized (16 h, 37 °C) at an enzyme-to-substrate ratio of 1:50 following an automated SP3 protocol59. Digested samples were resuspended in 1% (v/v) formic acid, 0.5% DMSO for LC-MS/MS analysis. Peptides were analyzed by LC-MS/MS in an Exploris480 mass spectrometer (Thermo) coupled to a Vanquish Neo LC system. Peptides were loaded into a trap column (PepMap™ Neo, 5 µm C18, 300 µm x 25 mm) and then separated on an Easy-Spary™ PepMap Neo™ analytical column (C18 2 µm 75 µm × 500 mm) maintained at 50 °C. An 85-min gradient was used at a flow rate of 400 nL/min, with solvent A consisting of 0.1% formic acid (FA) and solvent B consisting of 100% acetonitrile (ACN) with 0.1% FA. The gradient ramped from 2% to 25% solvent B over 50 minutes, then from 25% B to 45% solvent B over 20 min, followed by a 9-min wash at 98% Buffer B.
The mass spectrometer was operated in a data-dependent mode, with an automatic switch between MS and MS/MS scans using a top 15 method. Intensity threshold ≥5 × 103, dynamic exclusion of 20 s and excluding charges (+1 and ≥+8). MS spectra were acquired from 400 to 1500 m/z with a resolution of 60,000 FWHM (200 m/z). Ion peptides were isolated using a 0.7 Th window and fragmented using higher-energy collisional dissociation (HCD) with a stepped normalized collision energy of 29, 31 and 33. MS/MS spectra resolution was set to 60,000 (200 m/z). The ion target values were 3 × 106 for MS (maximum IT of 25 ms) and 1 × 105 for MS/MS (maximum IT set to auto, according to the transient length). Aliquots of both the cross-linked and pre-cross-linking samples were digested with trypsin in parallel. Two hundred nanograms of the resulting non-cross-linked peptides were analyzed using a 37-min gradient on a Vanquis Neo, coupled with an Orbitrap Astral. The Orbitrap Astral was operated under a standard Data-Independent Acquisition (DIA) protocol to optimize protein detection and enhance quantification. Subsequently, DIANN analysis was performed on the pre-cross-linking sample, querying a custom-built FASTA database that included the host expression organism (Sf9 and the baculovirus), the CCD6-RET fusion protein construct, and the UniProt sequences of 525 proteins commonly identified as contaminants over the past decade in proteomic analyses of diverse samples. The proteins identified by DIANN were sorted by intensity and examined for potential contamination. For the cross-linked samples, the top 313 most abundant proteins in the non-cross-linked sample, based on the sum of all precursor intensities as calculated by DIANN, were selected. These proteins were then used to construct a customized FASTA database for MeroX searches. This number was deemed sufficient to model decoy hits and control the false discovery rate (FDR), as MeroX uses a built-in target-decoy strategy at the crosslink spectrum match level for reliable error estimation. For MeroX analysis of cross-linked samples, raw files acquired from the Orbitrap Exploris 480 were converted to MZML files using the ProteoWizard ms.convert program. The search parameters for MeroX were configured as follows: protease sites R/P and K/P, with a maximum of three missed cleavages, with K blocked as XL-site. Carbamidomethylation of cysteine (C) was set as a fixed modification, while oxidation of methionine (M) and phosphorylation of Ser, Thr, and Tyr were treated as a variable modification. DSBU was chosen as the cross-linker, and MeroX’s default parameters were used, considering dead-end reactions with H2O and NH3, since the cross-linking reaction was quenched with excess ammonium bicarbonate. The precursor and fragment ion precision were set to 5 ppm (MS1) and 10 ppm (MS2), respectively. Identification focused on b and y ion types, with a minimum of three fragments per peptide. Charge states of +2 and higher were selected, and deisotoping was enabled. Precursor mass correction was disabled. RISEUP was chosen as the analysis mode, allowing for one missing ion. Neutral losses were enabled, with a maximum of one loss per fragment. For scoring and FDR calculations, a prescore of 10%, an FDR cutoff of 1%, and a score cutoff of 50 were applied. Given the use of a customized database based on comprehensive sample analysis, the MeroX cRAP database was excluded from the searches.
Finally, each fragmentation spectra were manually inspected and validated. Circular views and further processing of crosslinked residues were done using XiVIEW60. For cross-linked peptide quantification, raw data were imported into Skyline. Label-free quantification of identified cross-linked peptides was performed using the extracted ion chromatogram of the isotopic distribution. Only peaks without interference were used for quantification and manually inspected for correct area integration. The sum of the ion areas was collapsed for each cross-linked peptide and normalized using the total ion intensity of each sample. Data shown are from one experiment with triplicate samples with two technical replicate each. Crosslinked samples were tested by SEC to monitor the formation of dimers versus high-molecular-weight species.
Negative staining and electron microscopy
CCDC6-RET (0.7 μM) was placed onto freshly glow-discharged (GloQube, Quorum Technologies) home-made carbon-coated 400 mesh copper electron microscopy (EM) grids. The conditions for glow discharge were: 15 mA, 45“, 0.1 bar. The samples were then incubated for 5 s at RT. The grids were consecutively placed on top of three distinct 50 μL drops of MilliQ water, striped gently with filter paper for 2 s and laid on the top of two different 50 μL drops of 1% uranyl acetate and stained for 1 min. Finally, they were stripped gently for 5 s and air dried. Prepared grids were visualized on a Tecnai G2 Spirit transmission EM (Thermo Fisher Scientific) with Lanthanum hexaboride cathode operated at 120 keV. Images were recorded at 61,320 × nominal magnifications on a 4k × 4k TemCam-F416 CMOS camera (TVIPS), with pixel size of 2.5 Å/pix. For data collection, the open-source software SerialEM (bio3d.colorado.edu/SerialEM) was used under a Massachusetts Institute of Technology (MIT) license. For data processing, open-source software cryoSPARC and Relion were used.
Small angle X-ray scattering
SAXS experiments were conducted at the beamline B21 of the Diamond Light Source (Didcot, UK)61. CCDC6-RET∆CT (40 μL, 5 mg/mL) was injected at room temperature into a Superdex 200 Increase 3.2 column connected to an in-line Agilent 1200 HPLC system. The buffer used for the SEC column was the same protein buffer (20 mM Tris pH 7.5, 500 mM NaCl, 1 mM DTT). The eluted samples were exposed for 300 s in 10 s acquisition blocks using an X-ray wavelength of 1 Å, and a sample-to-detector (Eiger 4 M) distance of 3.7 m. The data covered a momentum transfer range of 0.0032 <q < 0.34 Å−1. The frames recorded immediately before elution of the sample were subtracted from the protein scattering profiles. The Scåtter software package (www.bioisis.net) was used to analyze data, buffer-subtraction, scaling, merging and checking possible radiation damage of the samples. The Rg value was calculated with the Guinier approximation assuming that at very small angles q < 1.3/Rg. The particle distance distribution, Dmax, was calculated from the scattering pattern with GNOM and shape estimation was carried out with DAMMIF/DAMMIN, both programs included in the ATSAS package62. The protein’s molecular mass was estimated with GNOM. Interactively generated PDB-based homology models were made using the program COOT by manually adjusting them into the envelope given by SAXS that could match the experimental scattering data. This was computed with the program FoXS63.
Structural modeling and molecular dynamics simulations
Publicly available X-ray crystal structures of different RET kinase constructs, as well as those of related tyrosine kinase domains from catalytic receptors such as EGFR, FGFR (1–3), and the human insulin receptor, were suitably aligned using the Dali server. The RCD+ server64 was employed to fill in existing gaps in the structurally disordered regions due to a lack of electron density. The ATP molecule and the two coordinating, partially hydrated, magnesium ions (Mg²⁺) were carefully placed in an arrangement compatible with transfer of the γ-phosphate to a peptide substrate (near-attack conformation), as found in the prototypical cyclic AMP-dependent protein kinase A (PDB ids. IL3R and 4HPU). This experimental information was complemented with models generated by AlphaFold 365 for wild-type RET and the oncogenic CCDC6-RET homodimer, both in the absence and presence of adenosine nucleotides, Mg²⁺, and peptide substrates (accession codes: ma-pcoh8 and ma-b7la0). Additional models of CCDC6-RET in the unstimulated form were obtained with AlphaFold (AF id: J7M8C2, for the single chain in the manual model) and AlphaFold multimer Colab (RETPTC1DIMER_a902d_unrelaxed_rank_001_alphafold2_multimer_v3_model_2_seed_000), in the case of the homodimer. The SWISS-MODEL homology modeling server66 was also used for validation. Energy minimization and molecular dynamics (MD) simulations at 300 K of the systems studied were run in explicit physiological saline solution under periodic boundary conditions using the ff14SB AMBER force field67 with updated parameters for phosphates68, TIP3P water molecules and the pmemd.cuda_SPFP engine, as implemented in AMBER 18 running on Nvidia GTX 1080 RTX2080Ti GPUs, essentially as described69. The cutoff distance for calculating the non-bonded interactions was 9 Å and the smooth particle mesh Ewald method with a grid spacing of 1 Å was used to account for electrostatic interactions. Application of the SHAKE algorithm to all bonds involving hydrogens allowed an integration step of 2.0 fs to be used. Each solvated system was simulated in the absence of any restraints for at least 300 ns during which system coordinates were collected every 0.5 ns for further analysis. Complementarily, for the respective Michaelis complexes, a time-dependent biasing harmonic potential of 5 kcal mol−1 Å−2 was included -as an extra term in the form of a flat well with parabolic sides- to impose pulsating distance restraints within a range compatible with nucleophilic attack (3.9–4.0 Å) and hydrogen bond formation (2.9–3.1 Å). The molecular graphics program PyMOL v. 2.6 was employed for molecular visualization and figure preparation.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The mass spectrometry data have been deposited in the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD053907 (https://proteomecentral.proteomexchange.org/cgi/GetDataset?ID=PXD053907). The SAXS data were deposited in the small-angle scattering biological data bank (SASBDB) with ID: SASDVB5 (https://www.sasbdb.org/data/SASDSVB5). Protein model for the active configuration of CCDC6-RET has been deposited in ModelArchive with accession code ma-b7la0 (DOI: 10.5452/ma-b7la0) and ma-pcoh8 (DOI: 10.5452/ma-pcoh8). EM maps and other CCDC6-RET models used in this study are provided in the supplementary data files. Source data are provided with this paper.
References
Nowell PCaH, D. A. A minute chromosome in human chronic granulocytic leukemia. Science 142, 1497 (1960).
de Klein, A. et al. A cellular oncogene is translocated to the Philadelphia chromosome in chronic myelocytic leukaemia. Nature 300, 765–767 (1982).
Santoro, M., Moccia, M., Federico, G. & Carlomagno, F. RET gene fusions in malignancies of the thyroid and other tissues. Genes 11, 424 (2020).
Takahashi, M., Ritz, J. & Cooper, G. M. Activation of a novel human transforming gene, ret, by DNA rearrangement. Cell 42, 581–588 (1985).
Takahashi, M. & Cooper, G. M. ret transforming gene encodes a fusion protein homologous to tyrosine kinases. Mol. Cell Biol. 7, 1378–1385 (1987).
Takahashi, M. Structure and expression of the ret transforming gene. IARC Sci. Publ. 92, 189–197 (1988).
Iwamoto, T. et al. cDNA cloning of mouse ret proto-oncogene and its sequence similarity to the cadherin superfamily. Oncogene 8, 1087–1091 (1993).
Pachnis, V., Mankoo, B. & Costantini, F. Expression of the c-ret proto-oncogene during mouse embryogenesis. Development 119, 1005–1017 (1993).
Fusco, A. et al. A new oncogene in human thyroid papillary carcinomas and their lymph-nodal metastases. Nature 328, 170–172 (1987).
Grieco, M. et al. PTC is a novel rearranged form of the ret proto-oncogene and is frequently detected in vivo in human thyroid papillary carcinomas. Cell 60, 557–563 (1990).
Jhiang, S. M. et al. Detection of the PTC/retTPC oncogene in human thyroid cancers. Oncogene 7, 1331–1337 (1992).
Nikiforov, Y. E. RET/PTC rearrangement in thyroid tumors. Endocr. Pathol. 13, 3–16 (2002).
Ricarte-Filho, J. C. et al. Identification of kinase fusion oncogenes in post-Chornobyl radiation-induced thyroid cancers. J. Clin. Investig. 123, 4935–4944 (2013).
Fagin, J. A. & Wells, S. A. Jr. Biologic and clinical perspectives on thyroid cancer. N. Engl. J. Med. 375, 2307 (2016).
Kohno, T. et al. KIF5B-RET fusions in lung adenocarcinoma. Nat. Med. 18, 375–377 (2012).
Lipson, D. et al. Identification of new ALK and RET gene fusions from colorectal and lung cancer biopsies. Nat. Med. 18, 382–384 (2012).
Wang, R. et al. RET fusions define a unique molecular and clinicopathologic subtype of non-small-cell lung cancer. J. Clin. Oncol. 30, 4352–4359 (2012).
Klempner, S. J. et al. Emergence of RET rearrangement co-existing with activated EGFR mutation in EGFR-mutated NSCLC patients who had progressed on first- or second-generation EGFR TKI. Lung Cancer 89, 357–359 (2015).
Redaelli, S., Plaza-Menacho, I. & Mologni, L. Novel targeted therapeutics for MEN2. Endocr. Relat. Cancer 25, T53–T68 (2018).
Shehata, M. A. et al. Structural and dynamic determinants for highly selective RET kinase inhibition reveal cryptic druggability. J. Adv. Res. 45, 87–100 (2023).
McCoach, C. E. et al. Resistance mechanisms to targeted therapies in ROS1(+) and ALK(+) non-small cell lung cancer. Clin. Cancer Res. 24, 3334–3347 (2018).
Piotrowska, Z. et al. Landscape of acquired resistance to Osimertinib in EGFR-mutant NSCLC and clinical validation of combined EGFR and RET inhibition with Osimertinib and BLU-667 for acquired RET fusion. Cancer Discov. 8, 1529–1539 (2018).
Lin, J. J. et al. Mechanisms of resistance to selective RET tyrosine kinase inhibitors in RET fusion-positive non-small-cell lung cancer. Ann. Oncol. 31, 1725–1733 (2020).
Solomon, B. J. et al. RET solvent front mutations mediate acquired resistance to selective RET inhibition in RET-driven malignancies. J. Thorac. Oncol. 15, 541–549 (2020).
Tan, L. & Solomon, B. J. Defining resistance mechanisms to selective RET tyrosine kinase inhibitors in RET fusion-positive non-small-cell lung cancer. Ann. Oncol. 31, 1599–1600 (2020).
Terzyan, S. S. et al. Structural basis of resistance of mutant RET protein-tyrosine kinase to its inhibitors nintedanib and vandetanib. J. Biol. Chem. 294, 10428–10437 (2019).
Knowles, P. P. et al. Structure and chemical inhibition of the RET tyrosine kinase domain. J. Biol. Chem. 281, 33577–33587 (2006).
Plaza-Menacho, I. et al. RET functions as a dual-specificity kinase that requires allosteric inputs from juxtamembrane elements. Cell Rep. 17, 3319–3332 (2016).
Plaza-Menacho, I. et al. Oncogenic RET kinase domain mutations perturb the autophosphorylation trajectory by enhancing substrate presentation in trans. Mol. Cell 53, 738–751 (2014).
Mologni, L., Redaelli, S., Morandi, A., Plaza-Menacho, I. & Gambacorti-Passerini, C. Ponatinib is a potent inhibitor of wild-type and drug-resistant gatekeeper mutant RET kinase. Mol. Cell Endocrinol. 377, 1–6 (2013).
Hubbard, S. R., Wei, L., Hendrickson, W. A. & Ellis, L. WA. Crystal structure of the tyrosine kinase domain of the human insulin receptor. Nature 372, 746–754 (1994).
Simm, D., Hatje, K. & Kollmar, M. Waggawagga: comparative visualization of coiled-coil predictions and detection of stable single alpha-helices (SAH domains). Bioinformatics 31, 767–769 (2015).
Wood, C. W. & Woolfson, D. N. CCBuilder 2.0: powerful and accessible coiled-coil modeling. Protein Sci. 27, 103–111 (2018).
Bhate, M. P., Molnar, K. S., Goulian, M. & DeGrado, W. F. Signal transduction in histidine kinases: insights from new structures. Structure 23, 981–994 (2015).
Kimanius, D., Dong, L., Sharov, G., Nakane, T. & Scheres, S. H. W. New tools for automated cryo-EM single-particle analysis in RELION-4.0. Biochem. J. 478, 4169–4185 (2021).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Dubey, B. N. et al. Hybrid histidine kinase activation by cyclic di-GMP-mediated domain liberation. Proc. Natl. Acad. Sci. USA. 117, 1000–1008 (2020).
Dubey, B. N. et al. Cyclic di-GMP mediates a histidine kinase/phosphatase switch by noncovalent domain cross-linking. Sci. Adv. 2, e1600823 (2016).
Iacobucci, C. et al. A cross-linking/mass spectrometry workflow based on MS-cleavable cross-linkers and the MeroX software for studying protein structures and protein-protein interactions. Nat. Protoc. 13, 2864–2889 (2018).
Laskowski, R. A., Jablonska, J., Pravda, L., Varekova, R. S. & Thornton, J. M. PDBsum: Structural summaries of PDB entries. Protein Sci. 27, 129–134 (2018).
Reinhardt, R. et al. PKD autoinhibition in trans regulates activation loop autophosphorylation in cis. Proc. Natl. Acad. Sci. USA. 120, e2212909120 (2023).
Levina, A., Fleming, K. D., Burke, J. E. & Leonard, T. A. Activation of the essential kinase PDK1 by phosphoinositide-driven trans-autophosphorylation. Nat. Commun. 13, 1874 (2022).
Reinhardt, R. & Leonard, T. A. A critical evaluation of protein kinase regulation by activation loop autophosphorylation. Elife 12, e88210 (2023).
Guixe, V. & Merino, F. The ADP-dependent sugar kinase family: kinetic and evolutionary aspects. IUBMB Life 61, 753–761 (2009).
Ito, S. et al. Structural basis for the ADP-specificity of a novel glucokinase from a hyperthermophilic archaeon. Structure 9, 205–214 (2001).
Richter, J. P., Goroncy, A. K., Ronimus, R. S. & Sutherland-Smith, A. J. The structural and functional characterization of mammalian ADP-dependent glucokinase. J. Biol. Chem. 291, 3694–3704 (2016).
Gout, E., Rebeille, F., Douce, R. & Bligny, R. Interplay of Mg2+, ADP, and ATP in the cytosol and mitochondria: unravelling the role of Mg2+ in cell respiration. Proc. Natl. Acad. Sci. USA 111, E4560–E4567 (2014).
Tantama, M., Martinez-Francois, J. R., Mongeon, R. & Yellen, G. Imaging energy status in live cells with a fluorescent biosensor of the intracellular ATP-to-ADP ratio. Nat. Commun. 4, 2550 (2013).
Maldonado, E. N. & Lemasters, J. J. ATP/ADP ratio, the missed connection between mitochondria and the Warburg effect. Mitochondrion 19, 78–84 (2014).
Coccimiglio, I. F. & Clarke, D. C. ADP is the dominant controller of AMP-activated protein kinase activity dynamics in skeletal muscle during exercise. PLoS Comput. Biol. 16, e1008079 (2020).
Sampson, J., Richards, M. W., Choi, J., Fry, A. M. & Bayliss, R. Phase-separated foci of EML4-ALK facilitate signalling and depend upon an active kinase conformation. EMBO Rep. 22, e53693 (2021).
Juyoux, P., von Velsen, J., Pellegrini, E. & Bowler, M. W. Dual-specificity mitogen-activated protein kinase kinases can use ADP to phosphorylate MAP kinases in vitro. J. Biol. Chem. 301, 110578 (2025).
Borar, P. et al. Dual-specific autophosphorylation of kinase IKK2 enables phosphorylation of substrate IkappaBalpha through a phosphoenzyme intermediate. Elife 13, RP98009 (2025).
Knight, J. D. et al. A novel whole-cell lysate kinase assay identifies substrates of the p38 MAPK in differentiating myoblasts. Skelet. Muscle 2, 5 (2012).
Bork, P., Sander, C. & Valencia, A. Convergent evolution of similar enzymatic function on different protein folds: the hexokinase, ribokinase, and galactokinase families of sugar kinases. Protein Sci. 2, 31–40 (1993).
Li, M. H. et al. Crystal structure of brain pyridoxal kinase, a novel member of the ribokinase superfamily. J. Biol. Chem. 277, 46385–46390 (2002).
Plaza-Menacho, I., Mologni, L. & McDonald, N. Q. Mechanisms of RET signaling in cancer: current and future implications for targeted therapy. Cell Signal. 26, 1743–1752 (2014).
Cuesta-Hernandez, H. N. et al. An allosteric switch between the activation loop and a c-terminal palindromic phospho-motif controls c-Src function. Nat. Commun. 14, 6548 (2023).
Hughes, C. S. et al. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 14, 68–85 (2019).
Combe, C. W., Graham, M., Kolbowski, L., Fischer, L. & Rappsilber, J. xiVIEW: Visualisation of Crosslinking Mass Spectrometry Data. J. Mol. Biol. 436, 168656 (2024).
Cowieson, N. P. et al. Beamline B21: high-throughput small-angle X-ray scattering at Diamond Light Source. J. Synchrotron. Radiat. 27, 1438–1446 (2020).
Petoukhov, M. V. et al. New developments in the ATSAS program package for small-angle scattering data analysis. J. Appl. Crystallogr. 45, 342–350 (2012).
Schneidman-Duhovny, D., Hammel, M., Tainer, J. A. & Sali, A. FoXS, FoXSDock and MultiFoXS: Single-state and multi-state structural modeling of proteins and their complexes based on SAXS profiles. Nucleic Acids Res. 44, W424–429 (2016).
Lopez-Blanco, J. R., Canosa-Valls, A. J., Li, Y. & Chacon, P. RCD+: Fast loop modeling server. Nucleic Acids Res. 44, W395–400 (2016).
Abramson, J. et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 630, 493–500 (2024).
Waterhouse, A. et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 46, W296–W303 (2018).
Maier, J. A. et al. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 11, 3696–3713 (2015).
Raguette L. E., et al. phosaa14SB and phosaa19SB: Updated Amber Force Field Parameters for Phosphorylated Amino Acids. J. Chem. Theory Comput. 20, 7199–7209 (2024).
Mills, A. & Gago, F. Atomistic insight into sequence-directed DNA bending and minicircle formation propensity in the absence and presence of phased A-tracts. J. Comput. Aided Mol. Des. 34, 253–265 (2020).
Acknowledgements
We are grateful to the Genomics and Spectroscopy and NMR Units (CNIO) for technical assistance, to Marta San Román and Mariano Barbacid from the Experimental Oncology Group (CNIO) for helping with the radioactive experiments, and to former members of the Protein Phosphorylation and Cancer Group for their contribution at the very early stages of the project (Pablo Soriano, Nicolás Cuesta y Julio Martínez-Torres). We thank Nabil Djouder for helpful comments and advice on the manuscript and Michael Tress for helping with the.json files. The authors would like to thank Diamond Light Source for beamtime (beamline B21, proposal mx30297) and their staff for assistance during data collection. We thank the Centro Nacional de Investigaciones Oncológicas (CNIO), which is supported by the Instituto de Salud Carlos III and recognized as a “Severo Ochoa” Centre of Excellence (ref. CEX2019-000891-S, awarded by MCIN/AEI/ 10.13039/5 01100011033) for core funding and supporting this study. This work was further supported by projects from the Agencia Estatal de Investigación (AEI), Ministerio de Ciencia e Innovación (MCIN): BFU2017-86710-R (MCIN/AEI/10.13039/501100011033 and ERDF “A way of making Europe”), PID2020-117580RB-I00 (MCIN/AEI/10.13039/501100011033), RYC-2016-1938 (MCIN/AEI/10.13039/501100011033 and ESF “Investing in your future”), CNS2023-145340 “Ayudas para Incentivar la Consolidación Investigadora” (MCIU/AEI/10.13039/501100011033 and the European Union, NextGenerationEU/PRTR) and a FP7-PEOPLE-2013-COFUND-Marie-Curie Action: “Co-funding of regional, National and International Programmes” International grant (number 608765) to I.P-M. Work at the University of Alcalá was supported by grant PID2022-136307OB-C22/ AEI/10.13039/501100011033.
Author information
Authors and Affiliations
Contributions
Conception of the study, experimental design, supervision of the study, funding acquisition and preparation of first draft (I.P.-M.), manuscript writing, data analysis and figures preparation (I.P.-M., A.M.-H.), mass spectrometry including data analysis (J.S.-W., J.M., E.Z, F. García and M.I.), EM studies and data analysis (J.B. and J.LC.), SAXS studies including data analysis and figure preparation (I.G.M.), molecular dynamics simulations studies and data analysis (F. Gago), experimental work and data acquisition (A.M.-H., J.C., I.P.-M.).
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Xiabin Chen and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Martín-Hurtado, A., Contreras, J., Sánchez-Wandelmer, J. et al. The oncogenic CCDC6-RET fusion protein is a dual ATP- and ADP-dependent kinase. Nat Commun 17, 3595 (2026). https://doi.org/10.1038/s41467-026-69833-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-69833-y
