
Abstract
Covering 40–50% of world’s arable lands, acidic soils pose a major constraint on global crop productivity by severely restricting root development and nutrient acquisition. The Arabidopsis C2H2-type transcription factor STOP1 plays a fundamental role in mitigating acid stress by activating H+/NO3− symport via NRT1.1, thereby driving rhizosphere alkalinization to protect root growth and improving nitrogen use efficiency (NUE). However, the upstream regulation of this pH-responsive STOP1-NRT1.1 pathway remains poorly defined. Here, we identify the central SWI2/SNF2-type ATPase BRAHMA (BRM) as a key suppressor of the STOP1 pathway. BRM physically interacts with STOP1 and occupies the genomic region of NRT1.1, repressing the STOP1-dependent activation of NRT1.1 expression and consequently limiting NO3− uptake and rhizosphere alkalization under chronic acidity. Genetic epistasis analysis using brm stop1 and brm nrt1.1 double mutants establish BRM as an upstream regulator of this signaling module. Notably, low pH rapidly triggers BRM degradation independent of NO3− availability, thereby relieving its repression on the STOP1-NRT1.1 pathway. This dynamic BRM disintegration enables robust induction of H+-coupled NO3− uptake, remodeling the rhizosphere pH landscape to foster optimal root growth under acidity. Collectively, our findings uncover the BRM-STOP1-NRT1.1 axis as a central regulatory module integrating NO3− acquisition with pH homeostasis, offering a dual-benefit strategy for enhancing crop resilience to acid soils and reducing fertilizer-driven acidification through improved NUE.
Similar content being viewed by others

Introduction
Acidic soils occupy 40–50% of the world’s total arable lands, and their acidification pace is accelerating due to the widespread application of urea and ammonium-based fertilizers1,2, posing a critical challenge to the sustainability of global agriculture. Acidic soils impose a vicious cycle: high proton (H+) concentrations in acidic soils severely limit crop productivity by inhibiting root growth and nutrient acquisition3,4, while plants’ inability to efficiently utilize N fertilizers exacerbates soil acidification through residual H+ accumulation and also the risk of environmental pollution5. Molecular breeding of plants to improve nitrogen acquisition and adaptation to acidic soils could be a cost-effective solution to these problems6.
The C2H2-type transcription factor, SENSITIVE-TO-PROTON-RHIZO TOXICITY1 (STOP1), was identified as the first key regulator of plant H+ tolerance, with its disruption leading to over 70% inhibition of root growth under acidic conditions4. Subsequent lines of evidence have suggested that STOP1 has pleiotropic roles in plant response to various environmental cues, including ammonium toxicity, Pi deficiency, N deficiency, Ca2+ deficiency, salt stress, drought, and hypoxia stresses7,8,9,10,11,12,13,14,15,16. Furthermore, STOP1 is functional conserved across land plant species17,18. Among critical STOP1-regulated modules, our previous study demonstrated that the STOP1-NITRATE TRANSPORTER 1.1 (NRT1.1) module emerges as a critical buffer in response to low pH: STOP1 activates NRT1.1-mediated H+/NO3− symport, alkalizing the rhizosphere to protect roots and enhance N use efficiency5. However, the upstream mechanisms that underlie STOP1-enhanced plant adaptation to acidity remain unclear.
Soil acidification is a dynamically deteriorating process: even in moderately acidic soils (pH 5.0–5.5), repeated urea application can lower rhizosphere pH by 0.17–0.72 units over one growing season1,19. If plant-induced H+ influx from root nutrient uptake could counteract (either fully or partially) H+ released during nitrification of ammonium-based fertilizers, this equilibrium could mitigate soil acidification, simultaneously benefiting agricultural ecosystem sustainability and crop productivity. In Arabidopsis, NRT1.1 serves as the major transporter for root NO3− uptake under adequate NO3− supplementation and acidic conditions20,21. The NRT1.1-mediated H+/NO3− symport drives rhizosphere alkalization to protect roots from acid toxicity and enhance nitrogen use efficiency (NUE)5. Thus, adaptation to acidic stress requires plants to orchestrate both immediate stress responses and sustained transcriptional programming that stabilizes NRT1.1 expression to ensure sufficient NO3− uptake to neutralize protons derived from ammonium nitrification22. It’s worth noting that STOP1-dependent H+ tolerance is functionally restricted by its post-translational modifications and dosage sensitivity, thereby stringently restricting its capacity to activate NRT1.1 transcription. Furthermore, STOP1 overexpression not only produces growth penalties but also fails to confer chronic stress adaptation, while its upstream regulators governing transcriptional activity under H+ stress are largely unknown. Chromatin remodeling, by enabling sustained transcriptional memory during prolonged stress23, emerges as a promising solution to circumvent these limitations through epigenetic stabilization of stress-responsive genes.
In this study, we identified BRAHMA (BRM), a switch defective/sucrose non-fermentable (SWI/SNF) ATPase, as a key repressor of the STOP1-mediated pathway. BRM directly occupies the genomic region of NRT1.1 and interacts with STOP1 to inhibit its trans-activation activity under acidity, thereby suppressing the STOP1-NRT1.1 module. Low pH rapidly triggers BRM degradation, relieving repression to augment H+-coupled nitrate influx and establish a rhizosphere pH optimum for root growth under acidic condition. Our findings may provide a strategy for simultaneously improving plant NUE to partially counter the H+ derived from nitrification of ammonium-based fertilizers and slowing soil acidification through the manipulation of the BRM-STOP1-NRT1.1 module.
Results
BRM physically interacts with STOP1
To identify potential regulators of STOP1 during H+ stress, we conducted a yeast two-hybrid (Y2H) screen using STOP1 as bait. Although pGBKT7-STOP1 exhibited weak autoactivation in the AH109 yeast strain, this background activity was effectively suppressed by 50 mM 3-amino-1,2,4-triazole (3-AT) (Supplementary Fig. 1). The screen yielded 75 positive colonies, corresponding to 23 putative STOP1-interacting proteins (Supplementary Data 1). Among these, BRAHMA (BRM), a core chromatin-remodeling subunit, was recovered in six independent clones, making it the most recurrent and therefore the highest-priority candidate among previously unreported STOP1 interactors. Region mapping assays showed that the N-terminal region of BRM (BRMn, amino acids 1 to 976) was both necessary and sufficient for their interaction (Fig. 1a–c). Since BRMn functions as a docking site for the recruitment of transcription factors24, we employed this fragment, rather than full-length BRM, in subsequent biochemical assays. In Nicotiana benthamiana leaf BiFC experiments, co-expression of STOP1 and BRMn produced a robust nuclear fluorescence signal, confirming their interaction in planta (Fig. 1d). Furthermore, co-immunoprecipitation assay using Arabidopsis plants co-expressing pBRM:BRM-FLAG and pSTOP1:GFP-STOP1 (GFP-STOP1/BRM-FLAG) confirmed that GFP-STOP1 was specifically co-immunoprecipitated with BRM-FLAG after a 6-h low-pH and MG132 treatment (Fig. 1e). These results firmly establish that BRM interacts with STOP1in the nucleus.

a Schematic diagram of full-length BRM and the truncated fragments used in this study. Numbers above the boxes indicate the amino acid positions corresponding to each fragment. b, c Yeast two-hybrid (Y2H) assays showing interactions between BRM and STOP1. The coding sequences of BRM and STOP1 were cloned in-frame into BD and AD vectors, respectively. Transformed yeast cells were grown on SD/-Trp-Leu (selection) and SD/-Trp-Leu-His-Ade (interaction) media. Serial dilutions (10−1, 10−2, 10−3) of saturated cultures were spotted onto the plates. Representative images were taken after 4 days. d Bimolecular fluorescence complementation (BiFC) assay showing BRM–STOP1 interaction in Nicotiana benthamiana leaves. Agrobacterium strains carrying the indicated constructs were co-infiltrated into N. benthamiana leaves and imaged after 48 h. Scale bar: 0.1 mm. e Co-immunoprecipitation (Co-IP) assay confirming the BRM–STOP1 interaction in Arabidopsis. Total proteins were extracted from 10-day-old seedlings of pSTOP1:GFP-STOP1/stop1 (GFP-STOP1) and pSTOP1:GFP-STOP1/pBRM:BRM-FLAG (GFP-STOP1/BRM-FLAG) after 6-h treatment with low pH and MG132. BRM-FLAG was immunoprecipitated using anti-FLAG beads, and co-precipitated GFP-STOP1 was detected by immunoblotting with anti-GFP antibodies. All experiments were repeated three times independently, with similar results.
Lack of BRM enhances plant growth and NO3 − acquisition under H+ stress
To investigate the role of BRM in response to acidic stress, we performed a phenotypic analysis of two hypomorphic alleles of BRM (brm-3 and brm-20) and a complementation line (pBRM:BRM-FLAG/brm-3, hereafter referred to as COM). Under low-pH conditions, loss of BRM function significantly enhanced root growth, with root elongation in brm-3 and brm-20 mutants increasing by approximately 60% compared with that of wild-type (Col-0) plants, while the complementation of BRM fully restored the root growth of brm-3 mutants (Fig. 2a, b). Even after prolonged exposure, brm-3 roots remained longer than those of Col-0 under acidic pH, whereas under neutral pH the brm-3 mutant displayed significantly shorter root elongation compared with Col-0 (Supplementary Fig. 2). These results indicate that BRM negatively regulates root tolerance to acidic stress.

a, b Root growth of Col-0, brm, and pBRM:BRM-FLAG/brm-3 (COM) plants on neutral- and low-pH agar media (n = 20 seedlings). Three-day-old seedlings were transferred to the indicated media and grown for 3 days. c–f Root growth of Col-0, brm, and stop1 mutants on media containing different concentrations of nitrate (n = 20 seedlings). Three-day-old seedlings were transferred to neutral- or low-pH media supplemented with either 0.06 mM or 6 mM KNO3 and grown for 3 days. Potassium concentrations were balanced to 6 mM by adjusting K2SO4 levels. g, h RT-qPCR analysis of NRT1.1 expression in the roots of Col-0, brm, and stop1 mutants (n = 4 biological replicates). i, j Root growth of single and double mutants of brm and nrt1.1 on neutral- and low-pH media. Three-day-old seedlings were transferred and grown for 5 days. Scale bar: 1 cm. In (b, g, h, j), centerlines in the boxplots show the medians, box limits indicate the 25th and 75th percentile, and whiskers indicate minimum and maximum values. In (d, f), data represent means ± SD of 20 seedlings. Different letters indicate statistically significant differences between means (p < 0.05), determined by Tukey’s multiple comparisons test (one-way ANOVA in (h); two-way ANOVA in (b, d, f, g, j)). Exact p values are provided in the Source Data file. All experiments were repeated at least three times independently, with similar results.
Given that BRM degradation is crucial for maintaining root apical meristem integrity by alleviating DNA double-stand breaks (DSBs) induced under high-B conditions25, we examined whether the enhanced H+ tolerance of brm-3 mutants is related to altered DSBs levels. However, the expression of DSBs-induced genes (GR1, RAD51, and BRCA1) was neither significantly induced by low-pH treatment nor affected by BRM or STOP1 activity (Supplementary Fig. 3). Consistently, 1-day low-pH treatment did not significantly alter WT root DSBs level, suggesting that plants tolerance to acidic stress is not closely related with DSBs. Furthermore, the DSBs level of the acid-tolerant brm-3 mutant showed similar level as that of the acid-sensitive stop1 mutant, though they were slightly suppressed when compared to Col-0 (Supplementary Fig. 4). These results indicate that BRM-regulated H+ tolerance is not primarily associated with its role in DNA damage repair. Given that STOP1-improved H+ tolerance and NUE are closely linked to enhanced root NO3− uptake5 and that BRM physically interacts with STOP1 (Fig. 1), we speculated that BRM may regulates H+ tolerance in a NO3−-depended manner. To test this notion, we compared the effects of different concentrations of NO3− supplies (0.06 and 6 mM) on root elongation of Col-0, brm-3, and stop1 plants, with 1 mM NH4+ supplied as an additional nitrogen source. Under low-pH treatments with 0.06 mM NO3−, root growth of Col-0, brm-3, and stop1 mutants was comparable (Fig. 2c, d). In contrast, brm-3 mutants showed significantly enhanced root growth compared to Col-0 under low-pH treatments with 6 mM NO3−, while stop1 mutants exhibited reduced root elongation (Fig. 2c, d). These results suggest that BRM-regulated H+ tolerance depends on NO3− availability.
Notably, both brm-3 and brm-20 are weak alleles. While brm-3 harbors a T-DNA insertion that disrupts the bromodomain in the C terminus, brm-20 is predicted to result in a truncation of a small segment at the C terminus. These mutations do not appear to be essential for chromatin remodeling activity, but may affect BRM stability or its tethering to chromatin26,27,28. To confirm the specific role of BRM in acid stress response, we further analyzed the phenotype of BRM-null mutant (brm-1), which is sterile and exhibits severe developmental defects27,29,30. As shown in Fig. 2e, f, root elongation in the brm-1 mutant was largely insensitive to low-pH stress, in stark contrast to the significant inhibition observed in Col-0 plants. Moreover, decreasing the concentration of NO3− from 6 mM to 0.06 mM significantly inhibited root growth of Col-0 but barely affected that of brm-1 mutants under low-pH treatments, demonstrating that BRM-repressed H+ tolerance depends on NO3− availability.
Since NRT1.1 mediates the majority of root NO3− uptake under low-pH condition and the low pH-induced transcription of NRT1.1 is controlled by STOP15,21, we analyzed the expression of NRT1.1 in the roots of Col-0, brm-3, and stop1 mutants under different pH conditions with 6 mM NO3− supplementation. As expected, low pH-induced NRT1.1 expression was remarkably enhanced in roots of brm-3 and brm-1 mutants compared with that in Col-0 plants (Fig. 2g, h). To further investigate whether BRM and NRT1.1 function in the same pathway in response to H+ stress, we generated a brm nrt1.1 double mutant by crossing the nrt1.1-1 and brm-20 mutants (Supplementary Fig. 5). We were unable to obtain brm-3 nrt1.1 double mutant due to the severely reduced fertility of brm-3, as recently reported31. In low-pH agar medium, brm nrt1.1 plants exhibited root growth comparable to that of the nrt1.1-1 single mutants (Fig. 2i, j), suggesting that NRT1.1 is epistatic to BRM in the regulation of H+ tolerance. Collectively, these results indicate that BRM inhibits H+ tolerance by repressing the low-pH-induced expression of NRT1.1.
Lack of BRM enhances STOP1-activated NRT1.1 expression and NRT1.1 chromatin accessibility
To elucidate how BRM regulates NRT1.1 expression in response to H+ stress, we first assessed whether BRM affects STOP1 protein accumulation, as the induced NRT1.1 expression by low pH is known to be STOP1-dependent5. Under low pH condition, comparative analysis of GFP-STOP1 fluorescence in Col-0 versus brm-20 mutants (GFP-STOP1/brm-20) revealed no significant difference in STOP1 protein levels (Supplementary Fig. 6), suggesting that BRM does not repressed NRT1.1 expression by altering STOP1 protein accumulation. We next examined whether BRM modulates STOP1-dependent transcriptional activation of NRT1.1. Using a transient expression system in N. benthamiana leaves, we co-expressed STOP1 and BRM along with luciferase gene (LUC) reporter constructs driven by either the NRT1.1 promoter region (P1) or the 4th intron (P2), both containing STOP1-binding sites5,14 (Fig. 3a). In addition, NLP7, another well-known transcription factor that activates NRT1.1 transcription32, and the NRT1.1 promoter fragment (P3), which contains the binding regions of NLP7 but not STOP1 or BRM, were used as negative controls. As shown in Fig. 3b, c, BRM co-expression substantially decreased the STOP1-activated P1- and P2-derived LUC signals to an almost undetectable levels, suggesting that BRM indeed affects the activation of STOP1 on NRT1.1 expression. However, BRM co-expression had no significant impact on either the NLP7-activated P3-drived LUC activity driven by the P3 fragment or EV-resulted P3-mediated LUC activity (Supplementary Fig. 7). These results suggest that BRM specifically targets the STOP1-dependent activation of NRT1.1, rather than exerting a non-specific inhibitory effect on LUC expression. To assess whether this regulatory interaction occur in vivo, we carried out chromatin immunoprecipitation (ChIP) experiments. STOP1 enrichment at endogenous NRT1.1 chromatin regions was notably higher in brm-3 mutants than in Col-0 plants (Fig. 3d), confirming that BRM inhibits the binding of STOP1 to NRT1.1 genomic regions.

a Schematic representation of the effector and reporter constructs used in the transient dual-luciferase assays. All constructs contain a NOS terminator (not shown). Different NRT1.1 genomic fragments were inserted upstream of the CaMV 35S minimal promoter (Mini; −46/0). b, c Effect of BRM on STOP1 binding to the NRT1.1 genomic regions assessed via transient dual-luciferase assays. The empty vector (EV, pCambia1300) was used as a negative control (set to 1). Effector and reporter constructs were co-transformed into N. benthamiana leaves for 48 h, after which the ratio of firefly to renilla luciferase activity was measured (n = 4 biological replicates). d ChIP-qPCR assay performed in Col-0 and brm-3 protoplasts transiently expressing HBT:STOP1-GFP or the control HBT:GFP construct. Immunoprecipitation was performed using an anti-GFP antibody (n = 3 biological replicates). e Genome browser snapshot showing BRM binding sites at the NRT1.1 locus. The BRM ChIP-seq data shown was downloaded from GEO under accession no. GSM4233035 and the ATAC-seq data were downloaded from GEO under accession no. GSM5799538. f ChIP-qPCR assay using 10-day-old Col-0 and pBRM:BRM-FLAG/brm-3 seedlings, with immunoprecipitation using an anti-FLAG antibody. Fold enrichment represents IP efficiency in transgenic plants normalized to that in Col-0 (n = 3 biological replicates). g FAIRE-qPCR analysis of chromatin accessibility in the P1-1 and P2-1 regions of NRT1.1, relative to the UBQ10 coding sequence. Whole 10-day-old Arabidopsis seedlings were used. Relative amplicon abundance represents signal in the mutants normalized to Col-0 (n = 3 biological replicates). h, i Comparison of root growth between Col-0 and axe1-5 mutants. The 3-day-old Arabidopsis seedlings were transferred to a neutral- or low-pH medium for 2 days. Scale bar: 1 cm. Centerlines in the boxplots show the medians, box limits indicate the 25th and 75th percentile, and whiskers indicate the minimum and maximum values (n = 20 seedlings). j Transcription of NRT1.1 in roots. The 6-day-old Arabidopsis seedlings were transferred to a neutral- or low-pH medium for 1 days (n = 3 biological replicates). In (c, d, f, g, j), data are shown as mean ± SD. Significant differences were determined by two-tailed Student’s t-test in (c, d, f, i, j), or two-way ANOVA in (c, g). Exact p values are provided in the Source Data file. All experiments were repeated at least three times independently, with similar results.
To further investigate BRM’s autonomous capacity to inhibit NRT1.1 expression, we analyzed publicly available datasets using WashU Epigenome Browser (https://epigenomegateway.wustl.edu/) and Plant Regulomics (https://plantregulomics.org/). This analysis revealed that BRM is associated with NRT1.1 genomic regions harboring STOP1-binding motifs (Fig. 3e; Supplementary Fig. 8). This association was further confirmed by ChIP-qPCR, which showed BRM enrichment at these sites (Fig. 3f). Together, these findings support the idea that BRM antagonistically regulates NRT1.1 expression by interfering with STOP1 activity. Given that BRM modulates gene expression by altering chromatin accessibility33,34,35, we next assess the chromatin state of the NRT1.1 locus using FAIRE-qPCR. Consistent with the ChIP-qPCR results, chromatin accessibility at the P1-1 and P2-1 regions of NRT1.1 was significantly increased in brm mutants compared to Col-0, with a stronger effect observed at P2-1 (Fig. 3g). These results indicate that BRM represses NRT1.1 expression by limiting chromatin accessibility at STOP1-binding regions, thereby reducing STOP1 occupancy and inhibiting NRT1.1 activation.
In addition, previous studies have shown that BRM interacts with the histone deacetylase HDA6 to repress transposons under various stresses by modulating DNA methylation and H3 acetylation levels26,36. Our data also implicate HDA6 in this regulatory circuit: loss of HDA6 function enhanced H+ tolerance and increased NRT1.1 expression in root tips—the zones most sensitive to acidic stress and the key sites where STOP1 acts to prevent H+ toxicity5 (Fig. 3h–j). These findings suggest that BRM and HDA6 may cooperatively repress NRT1.1 through chromatin remodeling mechanisms that likely involve H3 acetylation and DNA methylation.
BRM acts genetically upstream of STOP1 in regulation of NO3 − uptake and H+ tolerance
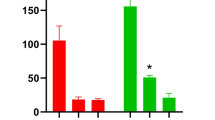
To clarify the genetic relationship between BRM and STOP1 in response to H+ stress, we generated a brm stop1 double mutant by crossing the brm-20 and stop1 mutants (Supplementary Fig. 9). When grown on low-pH agar medium, the brm stop1 double mutant displayed root growth similar to the stop1 mutant but notably shorter than that of Col-0 and brm mutant (Fig. 4a, b). In accord with the elevated NRT1.1 expression observed in roots of brm mutant, the brm plants also showed increased root NO3− uptake compared to Col-0 (Fig. 4c, d). However, both the enhancement of NRT1.1 expression and root NO3− uptake rate was completely abolished in the brm stop1 double mutant (Fig. 4c, d), indicating that the increased NRT1.1 expression and NO3− uptake in brm mutant requires functional STOP1. These results suggest that BRM exerts genetically upstream of STOP1 in regulating NO3− uptake and H+ tolerance under low pH conditions.

a, b Root growth of Col-0, stop1, brm, and stop1 brm double mutants. Three-day-old seedlings were transferred to either neutral or low-pH medium and grown for an additional 3 days (n = 20 seedlings). Scale bar: 1 cm. c, d Nitrate uptake rates and relative expression of NRT1.1 in Col-0, stop1, brm, and stop1 brm mutants under neutral and low-pH conditions. Data are shown as mean ± SD (n = 4 biological replicates per genotype and condition). In (b, d), centerlines in the boxplots show the medians, box limits indicate the 25th and 75th percentile, and whiskers indicate the minimum and maximum values. In (c), data represent mean ± SD. Different letters indicate statistically significant differences between means (p < 0.05), determined by Tukey’s multiple comparisons test (one-way ANOVA in (b, d); two-way ANOVA in (c)). Exact p values are provided in the Source Data file. All experiments were repeated at least three times independently, with similar results.
Lack of BRM enhances rhizosphere alkalization in response to H+ stress
Given the central role of the STOP1-NRT1.1 module in mediating H+-coupled NO3− uptake during acid stress, we hypothesized that targeted manipulation of BRM could promote a more favorable rhizosphere pH for root growth under low pH and curb N fertilizer-induced soil acidification. To test this, we first examined whether co-planting brm−3 and stop1 mutants could influence the root growth of stop1 mutants in low-pH agar media. Remarkably, when brm-3 and stop1 mutants were co-planted, stop1 root growth recovered to ~80% of brm-3 levels, versus only 57% in monoculture (Fig. 5a, b; Supplementary Fig. 10). Parallel experiments with Col-0 and brm-3 at pH 4.5 (accounting for inherent STOP1-NRT1.1 alkalinization capacity) similarly demonstrated improved Col-0 root growth in co-culture (Supplementary Fig. 10). These results suggest that loss of BRM function further promotes rhizosphere alkalinization, thereby enhancing root growth in response to acidic stress.

a, b Root growth of mono-planted and co-planted seedlings in acidic agar medium. Three-day-old brm-3 and stop1 seedlings were either mono-planted or co-planted on neutral or low-pH agar, as illustrated in Supplementary Fig. 6. Seedlings were photographed, and root elongation was measured 3–4 days after transfer (n = 20 seedlings). c, d Growth of Col-0, brm-3, and stop1 seedlings on agar medium with or without a pH buffer. Three-day-old seedlings were transferred to neutral or low-pH agar supplemented with or without 0.75 mM Homo-PIPES for 3 days (n = 20 seedlings). e Visualization of rhizospheric alkalization using bromocresol purple staining. Seedlings were grown for 12 h on basal agar medium with an initial pH of 4.9. f Measurement of pH in the rooting zone. Ten-day-old seedlings were pre-treated for 2 days on pH 4.7 agar before bromocresol purple staining and pH quantification. Data are shown as mean ± SD (n = 3 biological replicates with each replicate containing 30 seedlings). In (b, d), centerlines in the boxplots show the medians, box limits indicate the 25th and 75th percentile, and whiskers indicate the minimum and maximum values. Scale bar: 1 cm. Significant differences were determined using multi-way ANOVA in (b), and two-way ANOVA in (d), and one-way ANOVA in (f), followed by Tukey’s multiple comparisons test. Different letters indicate statistically significant differences (p < 0.05). Exact p values are provided in the Source Data file. All experiments were repeated at least three times independently, with similar results.
To further assess the role of BRM in regulating rhizosphere pH, seedlings were grown in acidic agar medium with or without 0.75 mM Homo-PIPES, a pH buffer used to minimize pH fluctuations. In the buffered acidic media, brm-3 mutants lost their growth advantage, exhibiting root elongation comparable to that of Col-0 and stop1 (Fig. 5c, d). Conversely, in unbuffered acidic media, brm-3 mutants showed dramatically improved root growth and enhanced rhizosphere alkalinization, as quantified by bromocresol purple staining and direct pH measurement after 6 h (Fig. 5e, f). In accordance with these results, the brm-1 mutant displayed significantly enhanced and rapid rhizosphere alkalinization compared to Col-0 (Supplementary Fig. 11). Taken together, these results indicate that loss of BRM enhances rhizosphere pH, thereby increasing H+ tolerance and offering a promising strategy to mitigate soil acidification under acidic conditions.
H+ stress induces BRM degradation
We next investigated the regulatory dynamics of BRM in roots under H+ stress. While low-pH treatment did not significantly alter the transcript levels of BRM (Fig. 6a), it substantially reduced the nuclear accumulation of BRM-GFP in root epidermal cells of pBRM:BRM-GFP/brm-20 transgenic plants (Fig. 6b). This decrease occurred independently of NO3− availability (Supplementary Fig. 12), and was detected as early as 2 h after low-pH exposure, preceding the induction of GFP-STOP1, which became evident at 4 h (Fig. 6b, c). This temporal hierarchy establishes BRM destabilization as an early event in the acid stress response, positioning BRM upstream of STOP1 signaling. Furthermore, the low pH-repressed BRM accumulation was restored by the treatment with the 26S proteasome inhibitor MG132 (Fig. 6d–f), suggesting that H+ stress likely promotes BRM degradation via the proteasome-dependent proteolysis. To investigate whether this degradation requires SnRK2 kinases, which are known to phosphorylate and inactivate BRM upon ABA treatment24, we assessed the growth of snrk2.2/2.3 and snrk2.6 mutants. However, these mutants exhibited root growth comparable to Col-0 under both neutral and low-pH conditions (Supplementary Fig. 13), ruling out SnRK2s participation in BRM-mediated H+ tolerance. Overall, these findings support a model in which low pH triggers proteasomal degradation of BRM, thereby enabling activation of the STOP1-NRT1.1 module to enhance H+ tolerance.

a RT-qPCR analysis of BRM transcript levels in roots of wild-type plants at 2, 4, and 24 h after low-pH treatment (n = 4 biological replicates). b, c Confocal imaging of BRM-GFP and GFP-STOP1 signals in roots of pBRM:BRM-GFP/brm-20 and pSTOP1:GFP-STOP1/stop1 seedlings under neutral or low-pH conditions at the indicated time points. Nuclear fluorescence intensity in the elongation zone was quantified using ImageJ and normalized to BRM-GFP signal at pH 6.5 (n = 20 seedlings). d, e Effect of low pH and MG132 treatment on BRM-GFP protein accumulation in roots of pBRM:BRM-GFP/brm-20 seedlings (n = 20 seedlings). f Western blot analysis of BRM-FLAG protein levels in roots of pBRM:BRM-FLAG/brm-3 seedlings after 12 h of treatment in neutral or low-pH medium, with or without MG132. Five-day-old seedlings were used. BRM-FLAG signal was quantified and normalized to ACTIN. In (a, c), data represent mean ± SD. In (e), centerlines in the boxplots show the medians, box limits indicate the 25th and 75th percentile, and whiskers indicate the minimum and maximum values. Exact p values are determined by two-tailed Student’s t-test in (a, e). Scale bar in (b, d): 0.1 mm. All experiments were repeated at least three times independently, with similar results. g Proposed model of the BRM–STOP1–NRT1.1 regulatory module in plant adaptation to acidic stress. BRM represses the STOP1–NRT1.1 pathway by binding the NRT1.1 genomic region and physically interacting with STOP1, thereby inhibiting STOP1’s transcriptional activity. Under acidic conditions, high H+ levels promote BRM degradation, releasing its repression of the STOP1–NRT1.1 module. This enhances H+-coupled nitrate uptake and promotes rhizospheric alkalization, ultimately improving root growth under low-pH stress. Created in BioRender. Ding, Z. (2026) https://BioRender.com/6pl2bd5.
Discussion
Understanding how plants adapt to H+ stress and acquire nutrients under acidic conditions is critical for improving nutrient use efficiency and crop production in acidic soils. The H+-coupled NO3− uptake by the root, mediated by the STOP1-NRT1.1 module, is a key biological process for improving plant H+ tolerance5,21,37,38. Here, we identified BRM as a pH-dependent repressor of STOP1-NRT1.1 module, revealing a vital regulatory layer in plant H+ adaptation (Fig. 6g). In this model, BRM physically interacts with STOP1 and binds directly to regulatory regions of NRT1.1, suppressing the STOP1’s transcriptional activation on NRT1.1. Low pH alleviates this repression by triggering BRM degradation, and thereby enables STOP1 to stimulate the H+-coupled NO3− uptake by NRT1.1, consequently enhancing rhizosphere alkalization, and establishing a favorable pH environment for root growth.
Despite the identification of STOP1’s role in H+ tolerance over a decade ago, the upstream regulatory mechanisms governing its activation and STOP1-improved H+ tolerance have long been enigmatic. Lines of evidences reveal that STOP1 is post-translationally modified in response to multiple stresses4,13,16,39,40,41,42. Unlike the Al3+-induced post-translational modification of STOP1(e.g., deSUMOylation/SUMOylation, phosphorylation, and ubiquitination39,40,42,43,44,45,46, our findings demonstrate that BRM physically interacts and suppresses STOP1 activity without affecting the STOP1 protein level, acting instead at the level of chromatin and transcriptional regulation (Fig. 3; Supplementary Fig. 3). Our identification of a BRM–STOP1 interaction is further supported by an independent high-throughput Y2H screen in which BRM was used as bait and STOP1 emerged as one of the strongest interactors47. Functionally, BRM specifically antagonizes STOP1-dependent activation of NRT1.1 (Fig. 3; Supplementary Fig. 7). Genetic evidence reinforces this relationship: the brm stop1 double mutant phenocopied stop1 in root growth, nitrate uptake, and NRT1.1 expression (Fig. 4), and was clearly distinct from the WT and brm single mutant. This non-additive genetic behavior demonstrates that STOP1 is epistatic to BRM in controlling H+ tolerance and NRT1.1 expression. BRM establishes a low-activity baseline for the STOP1–NRT1.1 system, preventing unwarranted rhizosphere alkalinization and nitrate uptake under non-stress conditions. Crucially, exposure to low pH triggers rapid BRM degradation (Fig. 6), creating a temporal activation window in which STOP1 and NRT1.1 can be robustly induced only under bona fide acidic stress. In this way, BRM acts as a pH-responsive gatekeeper that ensures both stability and sensitivity of the STOP1 regulatory network and dynamically coordinates pH homeostasis with nitrate acquisition. This pH-sensitive switch enables rapid metabolic and rhizosphere remodeling in response to fluctuating acidity—an essential strategy for soil adaptation. Future studies should focus on how plants sense acidic stress signals and elucidating the molecular cues that trigger BRM degradation in response to acidic stress. Possible upstream regulators include pH-sensing kinases or ion channels that could link environmental signals to BRM stability and functions. In parallel, the mechanisms underlying nucleus-localized STOP1 enrichment in response to H+ stress remain to be elucidated.
Beyond NRT1.1, BRM–STOP1 cooperativity extends to multiple STOP1 targets. Two canonical STOP1-regulated genes, ALMT1 and CIPK23, were significantly upregulated in brm mutants and this induction was fully abolished by STOP1 disruption (Supplementary Fig. 14). Phenotypically, brm mutants displayed enhanced Al tolerance and increased sensitivity to phosphate deficiency—both STOP1-regulated processes—while brm stop1 double mutants again resembled stop1 (Supplementary Figs. 15 and 16). These findings indicate that BRM acts as a broad-acting negative regulator of STOP1-mediated stress responses, supporting a direct mechanistic role of BRM–STOP1 interaction rather than a locus-specific effect at NRT1.1 alone. Moreover, while STOP1 is indispensable for multiple stress responses, sustained high STOP1 activity is detrimental under normal conditions (Supplementary Fig. 17). By physically interacting with STOP1 to dampen its transcriptional activity and by occupying chromatin regions of STOP1 target genes to restrict accessibility, BRM constrains both the intensity and spatial range of STOP1 function. This repression is essential to prevent inappropriate activation of STOP1-mediated pathways and the associated growth penalties. We therefore propose that the BRM–STOP1 module may operate as a generalized regulatory logic for managing trade-offs between growth and stress resilience. BRM-mediated repression keeps STOP1 activity at an optimal low level under non-stress or mild stress conditions, preserving normal development. When stress intensifies, BRM inhibition is relieved, enabling precise and timely activation of STOP1-mediated adaptation programs. This dynamic repression–activation mechanism likely underpins plant acclimation to diverse and rapidly changing environmental stresses.
BRM, a core SWI/SNF chromatin-remodeling ATPase, has been implicated in a variety of stress responses, such as drought, phosphate deficiency, and gibberellin acid (GA) signaling24,25,33,36,47,48,49,50,51. Our results revealed BRM directly represses NRT1.1 expression by modulating chromatin accessibility at its regulator loci and through physical interaction with its key activator STOP1. Importantly, this repression is relieved in brm mutants, which exhibit increased NO3− uptake and H+ tolerance under both neutral- and acidic conditions (Figs. 1 and 3). It’s worth noting that significant phenotypic divergence among brm alleles have been observed in response to low-pH treatment, which may due to the different functional domains affected among brm alleles. Using Y2H and BiFC assays, we showed that the N-terminus of BRM (BRMn, amino acids 1-976) is both necessary and sufficient for its physical interaction with STOP1 (Fig. 1). Notably, both brm-3 (harbors a T-DNA insertion disrupting the bromodomain in the C-terminus) and brm-20 (undergoes a small C-terminal truncation) mutants retain the intact N-terminal domain, thus maintaining the ability to interact with STOP1, which accounts for the partial retention of BRM’s repressive function on the STOP1-NRT1.1 module under low pH conditions. In contrast, the brm-1 null mutant, which lacks the entire protein including the N-terminal STOP1-interaction domain, exhibits a complete loss of BRM-mediated repression, leading to the full acid-tolerant phenotype. This underscores that the BRM N-terminus serves as the core domain for repressing the STOP1-NRT1.1 module. Furthermore, previous studies have showed that deletion of the C-terminal bromodomain (disrupted in brm-3) and the small C-terminal segment (truncated in brm-20) seems not to be essential for chromatin remodeling activity, but may affect BRM stability or its tethering to chromatin26,27,28. Therefore, although these regions are not required for BRM-STOP1 interaction (Fig. 1), their disruption does affect acid stress tolerance (Fig. 2). These results, together with the finding that low pH rapidly promotes BRM degradation (Fi.6), suggest that BRM C-terminus also play roles in modulating acid stress adaptation, likely via fine-tuning BRM stability. Further studies are needed to elucidate the function and mechanisms of these functional domains in acid stress tolerance. In addition, several studies showed that BRM interacts with histone deacetylase HDA6 to repress transposons in response to several stresses through modulation of DNA methylation and histone H3 acetylation levels26,36. Our data also point to a potential role for histone deacetylase HDA6 in cooperatively repressing NRT1.1 via chromatin remodeling mechanisms involving H3 acetylation and DNA methylation. Nevertheless, how BRM is recruited to the NRT1.1 loci and whether additional histone modifiers participate in this regulatory module require further investigation.
In agricultural contexts, excessive nitrogen fertilizers application, particularly urea-N fertilizer, has not only reduced NUE in crops but also significantly induced severe soil acidification1,52,53. This acidification further slows down microbe-driven nitrification processes54, prolonging the retention of fertilizer nitrogen in NH3 form and increasing NH3 volatilization potential. These sequential processes create a vicious cycle where continued acidification suppresses nitrification, while accumulated ammonium becomes increasingly prone to gaseous loss. Consequently, developing effective strategies for timely removal of H+ ions generated during nitrification represents a critical intervention point for improving NUE in agricultural systems. The BRM-STOP1-NRT1.1 pathway offers dual agronomic and ecological benefits by improving NUE and mitigating soil acidification driven by nitrogen fertilizers via enhancing NO3− uptake and rhizosphere alkalinization (Figs. 4 and 5). Theoretically, genetic manipulation of BRM to decouple its association with STOP1 and the genomic loci of NRT1.1 could optimize crop resilience in acidic soils while reducing environmental costs. Elucidating the evolutionary conservation of this pathway in staple crops, like rice or wheat, could unlock strategies for sustainable agriculture in acid-affected regions.
In summary, we identify BRM as a key regulator of the STOP1-NRT1.1 module, orchestrating rhizosphere pH homeostasis and nutrient acquisition under acidic conditions through modulation of H+-coupled NO3− uptake. This work bridges chromatin remodeling, H+ stress signaling, and nutrient acquisition, and provides a foundation for engineering crops with enhanced H+ tolerance and NUE. Further exploration of BRM’s degradation mechanism and broader its interaction network will further advance the development of resilient crop systems adapted to H+ stress and help address global soil acidification challenges.
Methods
Plant material
The Arabidopsis thaliana plants used in this work were all from Columbian (Col-0) background. The various mutants were stop1KO (SLAK_114108), nrt1.1-1 (SALK_097431), tfl2-1 (CS379655), axe1-536, brm-1 (chr2-1)56. The brm-3 (SALK_088462), brm-20 (SALK_002500), pBRM:BRM-GFP/brm-20, and pBRM:BRM-FLAG/brm-3 transgenic line used in this study has been previously described26. The brm stop1 and brm nrt1.1 double mutants were generated by crossing the brm-20 and stop1 mutants or nrt1.1-1 mutants, respectively, and homozygous lines were identified by PCR. The pSTOP1:GFP-STOP1/pBRM:BRM-FLAG (GFP-STOP1/BRM-FLAG) line was generated by crossing the pSTOP1:GFP-STOP1/stop1(#B10) transgenic line8 with pBRM:BRM-FLAG/brm-3, and the homozygous lines were selected in agar media containing basta and hygromycin. The primers used are shown in Supplementary Data 2.
Plant growth condition
Two days of stratification were conducted at 4 °C before sowing the seeds. Surface-sterilized Arabidopsis seeds were sown on basal agar medium containing KNO3 (6 mM), (NH4)2SO4 (1 mM), NaH2PO4 (1 mM), MgSO4 (500 μM), CaCl2 (1 mM), H3BO3 (10 μM), MnSO4 (0.5 μM), ZnSO4 (0.5 μM), CuSO4 (0.1 μM), (NH4)6Mo7O24 (0.1 μM), Fe-EDTA (50 μM), 1% agar (Sigma-Aldrich, A1296), 1% sucrose at pH 6.5 in a plant growth chamber (BPC600H, Fujian Jiupo Biotechnology, Co., Ltd.) at 22–23 °C under a 16-h light/8-h dark photoperiod cycle. Then, the 3-day-old plants were used for phenotype analysis in the above basal agar medium with an initial pH of either 4.7 or 6.5, and 0.75 mM Homo-PIPES were used for the buffering assay, as indicated in the figure legends. To analyze the effect of NO3− level on root growth in response to low pH, the media were supplemented with 0.06 or 6 mM KNO3, while the concentration of (NH4)2SO4 was reduced to 0.5 mM to avoid severe ammonium toxicity and the resulting differences in K concentrations were balanced by adjusting the K2SO4 concentration. To analyze the root growth of seedlings in response to low Pi, the Pi concentration in the media were reduced to 0.01 mM (low Pi). To analyze the root growth of seedlings in response to Al toxicity, seeds were directly sown on the control or Al treatment agar plates, prepared as previously described57.
Yeast two-hybrid screen and assays
For screening, the full CDS of STOP1 was amplified and inserted into pGBKT7 using the EcorI and BamHI restriction sites resulting in a pGBKT7-STOP1 constructs. This gene construct was transformed into yeast AH109 cells. Transformants were selected on SD/-Trp medium (Takara), after which the autoactivation activity of the bait vector pGBKT7-STOP1 was tested by plating on SD/-Trp-His-Ade medium (Takara) supplemented with different concentrations of 3-amino-1,2,4-triazole (3-AT). Then, the pGBKT7-STOP1 bait vector was cotransformed with an Arabidopsis cDNA library into AH109 yeast competent cells according to the manufacturer’s instructions (Clontech). Transformed cells were plated on SD/-Trp-Leu-His-Ade medium supplemented with 50 mM 3-AT, following incubation on 30 °C. Plasmids were isolated from positive yeast clones for analysis of insert and sequencing.
For targeted Y2H assays, the coding region of BRM and truncated version of BRM was amplified and inserted into pGBKT7 using the EcorI and BamHI restriction sites. The full CDS of STOP1 was amplified and inserted into pGADT7 using the EcorI and BamHI restriction sites. Yeast AH109 cells were co-transformed with specific bait and prey constructs. Yeast growth was shown on SD/-Trp/-Leu and SD/-Trp/-Leu/-His/-Ade medium, following incubation on 30 °C. The primers used for generating the various clones are listed in Supplementary Data 2.
Bimolecular fluorescence complementation (BiFC) assays
The full CDS of STOP1 and coding region of truncated BRMn was individually inserted into p2YN and p2YC vectors (46) to fuse with the N-terminal or C-terminal fragment of YFP. The primers used to generate the various clones are listed in Supplementary Data 2. The different combinations of recombinant plastmids were co-infiltrated into young Nicotinana benthamiana leaves for 48 h. The YFP fluorescence was detected using a confocal laser-scanning microscope (LSM880; Zeiss) and indicated in green color.
Co-IP assay
To better reflect the authentic in vivo interaction status between BRM and STOP1, we specifically used the double-tagged genetic material (BRM-FLAG/GFP-STOP1) for Co-IP assays. However, under neutral-pH conditions, the abundance of GFP-STOP1 protein in this transgenic line is extremely low, making it technically challenging to capture the GFP-STOP1 protein through immunoprecipitation. To address this issue, 10-d-old pSTOP1:GFP-STOP1 and pSTOP1:GFP-STOP1/pBRM:BRM-FLAG with 6-h low-pH treatment to induce GFP-STOP1 protein accumulation and with MG132 to inhibit the degradation of BRM-FLAG protein. Then, seedlings were harvested for total protein extraction in RIPA lysis buffer (FD011; Fdbio Science Biotech Co., Ltd). After centrifugating at 13,000 × g for 10 min, 50 µL of supernatants were kept as input controls, whilst the rest was incubated with 30 µL of anti-FLAG magnetic beads for 2 h. After the beads were washed five times with TBS buffer, the proteins were eluted by adding 50 µL 2× SDS sampling buffer at 95 °C for 8 min. Then the eluted proteins and input controls were separated on 7.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). The protein of interest was detected with α-GFP (1:10000; ABclonal) and α-FLAG (1:10000; ABclonal) antibodies.
Gene expression assay
Total RNA was extracted from plant roots using the FreeZol Reagent (R711-01; Vazyme Biotech Co., Ltd, Nanjing, PRC), and the first-strand cDNA was synthesized using ReverTra Ace qPCR RT Master Mix with gDNA Remover (TOYOBO, Osaka, Japan). The expression of the corresponding genes was determined using the SYBR Green Realtime PCR Master Mix (TOYOBO). The gene-specific primers used for quantitative PCR were listed in Supplementary Data 2. Expression levels of each mRNA were determined and normalized with the expression level of UBIQUITIN 10.
Analysis of green fluorescent protein (GFP)
After different time of low-pH treatment, as indicated in figure legends, BRM-GFP and GFP-STOP1 expression in the roots of pBRM:BRM-GFP/brm-20 and pSTOP1:GFP-STOP1 transgenic plants was detected using a confocal laser-scanning microscope (LSM880; Zeiss). The excitation and emission wavelengths were 488 nm/500–530 nm for GFP protein. Images were analyzed using the ZEN 2012 Blue Edition and ImageJ2X software.
Analysis of pH and visualization of rhizosphere alkalization
The 10-day-old Arabidopsis seedlings were pre-treated at pH 4.7 agar medium for 2 days before bromocresol purple staining and the pH value measurement. To visualize rhizosphere acidification, roots were transferred to a basal agar media with an initial pH of 4.9 and pH indicator bromocresol purple (0.008%, Sigma-Aldrich). The concentration of NaH2PO4 was reduced to 0.1 mM in the used agar media. Images were acquired 12 h after staining. The pH of the agar rooting medium was measured as previously described58. Briefly, the rooting medium was collected and frozen at –20°C for 4 h before thawed to free the aqueous phase from the agar. Then, the mixture was filtered at room temperature and the pH of the supernant was measured immediately using a pH electrode.
Chromatin immunoprecipitation (ChIP)-qPCR analysis
To assess the effect of BRM disruption on the binding ability of STOP1 to NRT1.1, protoplasts were isolated from the rosette leaves of 3-week-old WT or brm-3 plants, as described previously59. The effector plasmids pHBT:STOP1-GFP and pHBT:GFP used in this study have been previously described57. An effector or empty control plasmid (6 μg), and an internal control plasmid (pUBQ10:GUS; 2 μg) were co-transfected into 2 × 105 protoplasts using the polyethylene glycol (PEG)-mediated transformation method. At least 4 independent biological replicates were set for each transformation, and then the protoplasts were harvested together for the isolation of chromatin. The sheared DNA was immunoprecipitated with anti-FLAG magnetic beads overnight at 4 °C. In addition, ChIP assays were also carried out using 10-day-old Col-0 and pBRM:BRM-FLAG/brm-3 plants (approximately 2 g) and Pierce agarose ChIP Kit (26156; Thermo Fisher Scientific), according to the manufacturer’s instructions. The sheared DNA was immunoprecipitated with anti-FLAG magnetic beads overnight at 4 °C. The enrichment of specific DNA fragments in the immune precipitant was determined by qPCR analysis of the recovered DNA using specific primers (Supplementary Data 2). The fold enrichment represents the immune-precipitation efficiency in the transgenic plants normalized to that in the Col-0 plants.
Formaldehyde-assisted isolation of regulatory elements (FAIRE)-qPCR analysis
To assess the effect of BRM disruption on the open chromatin in the genome of NRT1.1, FAIRE was performed using the whole seedlings of 10-day-old WT, brm-3, and brm-20 plants, according to the protocol described previously with modifications60,61. Briefly, chromatins are cross-linked by 3% (final vol) formaldehyde treatment, followed by shearing chromatin by sonication, and reversing crosslinking. Then, a phenol/chloroform/isoamyl alcohol (PCI) extraction is performed, separating protein-free genomic DNA fragments into the aqueous phase and histone-DNA complexes to the interphase. The enrichment of specific DNA fragments in the recovered DNA was determined by qPCR analysis using specific primers (Supplementary Data 2). The relative enrichment of each DNA sample was first calculated using the delta cycle threshold (2−∆∆Ct) method by calculating the ratio of FAIRE signal to input signal. Relative amplicon abundance represents the results in the mutants normalized to that in the Col-0 plants.
Transient expression assay
To performed transient expression assays in Nicotiana benthamiana leaves, three DNA segments, selected from NRT1.1 genomic region (P1 and P2) that containing the identified binding site of STOP15,14 and (P3) that containing the identified binding site of NLP732 but not STOP or BRM, were individually inserted into the reporter vector pGreen0800-LUC using XhoI and BamHI restriction sites and using ClonExpress Ultra One Step Cloning Kit V3 (C117-01; Vazyme Biotech Co., Ltd, Nanjing, PRC). The primers used to generate the various clones are listed in Supplementary Data 2. In dual-luciferase reporter assays, reporter and effector constructs (35S:STOP113 or 35S:NLP7) were co-transformed into young N. benthamiana leaves for 48 h. Firefly and Renilla luciferase activities were quantified using a dual-luciferase assay kit (Beyotime Biotechnology, China). Some infiltrated leaves were also sprayed with 1 mM luciferin (A600577; Sangon Biotech, Shanghai, China), after which the images of luminescence were captured using a cooled charge-coupled device (CCD) imaging apparatus (LB985; Berthold).
Measurement of NO3 − uptake
For the assays of NO3− uptakes, 7-day-old seedlings were pre-treated in agar medium with a pH of 6.5 or 4.7, for 1 d. The plants were washed with 0.05 mM CaSO4 for 1 min and then transferred to a medium in which KNO3 was replaced with K15NO3 (atom % 15N, 99%) for 10 min under the same conditions used for pre-culturing seedlings. The plants were washed with 0.05 mM CaSO4 for 1 min, after which each plant was separated into shoots and roots, and the organs were dried at 65 °C for 3 d. The dried organs were weighed, and the roots and shoots of each plant were combined and ground into a fine powder. The samples were analyzed using an isotope ratio mass spectrometer (Isorime100; Elementar Analysensysteme, Hanau, Germany). The rate of 15NO3− uptake was calculated from the total 15N of the plant and the biomass of the roots.
Statistical analyses
Data were analyzed by one-way, two-way, or multi-way ANOVA with Tukey’s multiple comparison test.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data are available in the main text or the supplementary materials. Source data are provided with this paper.
References
Guo, J. H. et al. Significant acidification in major Chinese croplands. Science 327, 1008–1010 (2010).
Kochian, L. V., Pineros, M. A., Liu, J. & Magalhaes, J. V. Plant adaptation to acid soils: the molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 66, 571–598 (2015).
Schubert, S. S. E. & Mengel, K. Effect of low pH of the root medium on proton release, growth, and nutrient uptake of field beans (Vicia faba). Plant Soil 124, 239–244 (1990).
Iuchi, S. et al. Zinc finger protein STOP1 is critical for proton tolerance in Arabidopsis and coregulates a key gene in aluminum tolerance. Proc. Natl. Acad. Sci. USA 104, 9900–9905 (2007).
Ye, J. Y. et al. STOP1 activates NRT1.1-mediated nitrate uptake to create a favorable rhizospheric pH for plant adaptation to acidity. Plant Cell 33, 3658–3674 (2021).
Schroeder, J. I. et al. Using membrane transporters to improve crops for sustainable food production. Nature 497, 60–66 (2013).
Sawaki, Y. et al. STOP1 regulates multiple genes that protect Arabidopsis from proton and aluminum toxicities. Plant Physiol. 150, 281–294 (2009).
Balzergue, C. et al. Low phosphate activates STOP1-ALMT1 to rapidly inhibit root cell elongation. Nat. Commun. 8, 15300 (2017).
Mora-Macías, J. et al. Malate-dependent Fe accumulation is a critical checkpoint in the root developmental response to low phosphate. Proc. Natl. Acad. Sci. USA 114, 3563–3572 (2017).
Enomoto, T. et al. STOP1 regulates the expression of HsfA2 and GDHs that are critical for low-oxygen tolerance in Arabidopsis. J. Exp. Bot. 70, 3297–3311 (2019).
Godon, C. et al. Under phosphate starvation conditions, Fe and Al trigger accumulation of the transcription factor STOP1 in the nucleus of Arabidopsis root cells. Plant. J. 99, 937–949 (2019).
Sadhukhan, A. et al. Sensitive to Proton Rhizotoxicity1 regulates salt and drought tolerance of Arabidopsis thaliana through transcriptional regulation of CIPK23. Plant Cell Physiol. 60, 2113–2126 (2019).
Tian, W. H. et al. A transcription factor STOP1-centered pathway coordinates ammonium and phosphate acquisition in Arabidopsis. Mol. Plant. 14, 1554–1568 (2021).
Tokizawa, M. et al. The transcription factors, STOP1 and TCP20, are required for root system architecture alterations in response to nitrate deficiency. Proc. Natl. Acad. Sci. USA 120, e230044120 (2023).
Agrahari, R. K. et al. STOP1-regulatedSMALL AUXIN UP RNA55 (SAUR55) is involved in proton/malate co-secretion for Al tolerance in Arabidopsis. Plant Direct 8, 557–579 (2024).
Tian, W. H. et al. STOP1 regulates CCX1-mediated Ca2+ homeostasis for plant adaptation to Ca2+ deprivation. J. Integr. Plant Biol. 66, 2126–2139 (2024).
Ohyama, Y. et al. Characterization ofAtSTOP1 Orthologous Genes in Tobacco and Other Plant Species. Plant Physiol. 162, 1937–1946 (2013).
Dong, D. et al. Transcription factor SlSTOP1 regulates Small Auxin-Up RNA Genes for tomato root elongation under aluminum stress. Plant Physiol. 196, 2654–2668 (2024).
Shi, R. et al. Beneficial dual role of biochars in inhibiting soil acidification resulting from nitrification. Chemosphere 234, 43–51 (2019).
Huang, N. C., Chiang, C. S., Crawford, N. M. & Tsay, Y. F. CHL1 encodes a component of the low-affinity nitrate uptake system in Arabidopsis and shows cell type-specific expression in roots. Plant Cell 8, 2183–2191 (1996).
Liu, K., Huang, C. & Tsay, Y. CHL1 is a dual-affinity nitrate transporter of Arabidopsis involved in multiple phases of nitrate uptake. Plant Cell 11, 865–874 (1999).
Tian, D. S. & Niu, S. L. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 10, 024019 (2015).
Harris, C. J., Amtmann, A. & Ton, J. Epigenetic processes in plant stress priming: Open questions and new approaches. Curr. Opin. Plant Biol. 75, 102432 (2023).
Peirats-Llobet, M. et al. A direct link between abscisic acid sensing and the chromatin-remodeling ATPase BRAHMA via core ABA signaling pathway components. Mol. Plant 9, 136–147 (2016).
Sakamoto, T. et al. Proteasomal degradation of BRAHMA promotes Boron tolerance in Arabidopsis. Nat. Commun. 9, 5285 (2018).
Li, T. et al. The chromatin remodeler BRAHMA recruits HISTONE DEACETYLASE6 to regulate root growth inhibition in response to phosphate starvation in Arabidopsis. J. Integr. Plant Biol. 64, 2314–2326 (2022).
Hurtado, L., Farrona, S. & Reyes, J. C. The putative SWI/SNF complex subunit BRAHMA activates flower homeotic genes in Arabidopsis thaliana. Plant Mol. Biol. 62, 291–304 (2006).
Zhang, D., Li, Y., Zhang, X., Zha, P. & Lin, R. The SWI2/SNF2 Chromatin-Remodeling ATPase BRAHMA Regulates Chlorophyll Biosynthesis in Arabidopsis. Mol. Plant 10, 155–167 (2017).
Kwon, C. S. et al. A role for chromatin remodeling in regulation of CUC gene expression in the Arabidopsis cotyledon boundary. Development 133, 3223–3230 (2006).
Yang, S. G. et al. The Arabidopsis SWI2/SNF2 Chromatin Remodeling ATPase BRAHMA Targets Directly to PINs and Is Required for Root Stem Cell Niche Maintenance. The Plant Cell 27, 1670–1680 (2015).
Wrona, M. et al. The BRAHMA-associated SWI/SNF chromatin remodeling complex controls Arabidopsis seed quality and physiology. Plant Physiol. 197, kiae642 (2024).
Zhao, L. F. et al. The Arabidopsis NLP7 gene regulates nitrate signaling via NRT1.1–dependent pathway in the presence of ammonium. Sci. Rep. 8, 1487–1500 (2018).
Han, S. et al. The SWI2/SNF2 chromatin remodeling ATPase BRAHMA represses abscisic acid responses in the absence of the stress stimulus in Arabidopsis. Plant Cell 24, 4892–4906 (2012).
Ho, L. et al. Crabtree, esBAF facilitates pluripotency by conditioning the genome for LIF/STAT3 signalling and by regulating polycomb function. Nat. Cell Biol. 13, 903–913 (2011).
Hargreaves, D. C. & Crabtree, G. R. ATP-dependent chromatin remodeling: genetics, genomics and mechanisms. Cell Res. 21, 396–420 (2011).
Gao, L. et al. Abscisic acid–mediated autoregulation of the MYB41-BRAHMA module enhances drought tolerance in Arabidopsis. Plant Physiol. 196, 1608–1626 (2024).
Wang, Y., Hsu, P. & Tsay, Y. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 17, 458–467 (2012).
Fang, X. Z. et al. Alleviation of proton toxicity by nitrate uptake specifically depends on nitrate transporter 1.1 in Arabidopsis. New Phytol. 211, 149–158 (2016).
Zhang, Y. et al. F-box protein RAE1 regulates the stability of the aluminum-resistance transcription factor STOP1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 116, 319–327 (2019).
Fang, Q. et al. Regulation of aluminum resistance in Arabidopsis involves the SUMOylation of the zinc finger transcription factor STOP1. Plant Cell 32, 3921–3938 (2020).
Guo, J. et al. Mutation of HPR1 encoding a component of the THO/TREX complex reduces STOP1 accumulation and aluminium resistance in Arabidopsis thaliana. New phytol. 228, 179–193 (2020).
Zhou, F. et al. The MEKK1-MKK1/2-MPK4 cascade phosphorylates and stabilizes STOP1 to confer aluminum resistance in Arabidopsis. Mol. Plant 16, 337–353 (2023).
Fang, Q., Zhou, F., Zhang, Y., Singh, S. & Huang, C. F. Degradation of STOP1 mediated by the F-box proteins RAH1 and RAE1 balances aluminum resistance and plant growth in Arabidopsis thaliana. Plant J. 106, 493–506 (2021).
Xu, J. et al. SIZ1 negatively regulates aluminum resistance by mediating the STOP1–ALMT1 pathway in Arabidopsis. J. Integr. Plant Biol. 63, 1147–1160 (2021).
Cao, H. R. et al. Ca2+-dependent cytoplasmic and nuclear phosphorylation of STOP1 by CPK21 and CPK23 confers ALMT1-dependent aluminum resistance. Nat. Commun. 16, 5225 (2025).
Ma, Y. T. et al. CPK28-mediated Ca2+ signaling regulates STOP1 localization and accumulation to facilitate plant aluminum resistance. Nat. Commun. 16, 5224 (2025).
Efroni, I. et al. Regulation of leaf maturation by chromatin-mediated modulation of cytokinin responses. Dev. Cell 24, 438–445 (2013).
Archacki, R. et al. BRAHMA ATPase of the SWI/SNF chromatin remodeling complex acts as a positive regulator of gibberellin-mediated responses in Arabidopsis. PLoS One 8, 58588 (2013).
Clapier, C. R., Iwasa, J., Cairns, B. R. & Peterson, C. L. Mechanisms of action and regulation of ATP-dependent chromatin-remodelling complexes. Nat. Rev. Mol. Cell Biol. 18, 407–422 (2017).
Vain, T. et al. Selective auxin agonists induce specific AUX/IAA protein degradation to modulate plant development. Proc. Natl. Acad. Sci. USA 116, 6463–6472 (2019).
Zhang, C. et al. Gibberellin signaling modulates flowering via the DELLA-BRAHMA-NF-YC module in Arabidopsis. Plant Cell 35, 3470–3484 (2023).
Jadon, P. et al. Volatilization and leaching losses of nitrogen from different coated urea fertilizers. J. Soil Sci. Plant Nutr. 18, 1036–1047 (2018).
Guo, Y. et al. Air quality, nitrogen use efficiency and food security in China are improved by cost-effective agricultural nitrogen management. Nat. Food 1, 648–658 (2020).
Li, Y., Chapman, S. J., Nicol, G. W. & Yao, H. Nitrification and nitrifiers in acidic soils. Soil Biol. Biochem. 116, 290–301 (2018).
Li, Z. F. et al. Transcription factors AS1 and AS2 interact with LHP1 to repress KNOX genes in Arabidopsis. J. Integr. Plant Biol. 58, 959–970 (2016).
Wang, Z. Y. et al. SWI2/SNF2 ATPase CHR2 remodels pri-miRNAs via Serrate to impede miRNA production. Nature 557, 516–521 (2018).
Ding, Z. J. et al. The LRR receptor-like kinase ALR1 is a plant aluminum ion sensor. Cell Res. 34, 281–294 (2024).
Hachiya, T. et al. Nitrate addition alleviates ammonium toxicity without lessening ammonium accumulation, organic acid depletion and inorganic cation depletion in Arabidopsis thaliana shoots. Plant Cell Physiol. 53, 577–591 (2012).
Yoo, S., Cho, Y. & Sheen, J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat. Protoc. 2, 1565–1572 (2007).
Omidbakhshfard, M. A., Winck, F. V., Arvidsson, S., Riano-Pachon, D. M. & Mueller-Roeber, B. A step-by-step protocol for formaldehyde-assisted isolation of regulatory elements from Arabidopsis thaliana. J. Integr. Plant Biol. 56, 527–538 (2014).
Baum, S., Reimer-Michalski, E., Jaskiewicz, M. R. & Conrath, U. Formaldehyde-assisted isolation of regulatory DNA elements from Arabidopsis leaves. Nat. Protoc. 15, 713–733 (2020).
Acknowledgements
We thank Dr. She Long Zhang (the Instrument Platform of College of Life Sciences, Zhejiang University) for his assistant in Confocal measurement, Prof. Ming Guang Lei (Hangzhou Normal University, Hangzhou, China), Dr. Thierry Desnos (CEA Cadarache), Prof. Yi Kun He (Capital Normal University, Beijing, China), Prof. Zhi Ye Wang (Zhejiang University), and Prof. Ai Wu Dong (Fudan University, Shanghai, China) for providing seeds. This study was supported by the National Key Research and Development Program of China (grant no. 2022YFA1303402 to Z.J.D. and S.J.Z.), the National Natural Science Foundation of China (grant no. 31970272 to Z.J.D., 32500240 to J.Y.Y., and 32201702 to W.H.T.), the Postdoctoral Fellowship Program of CPSF (grant no. GZC20232327 to J.Y.Y.), the China Postdoctoral Science Foundation (2023M743048 to J.Y.Y.), the Ministry of Education and Bureau of Foreign Experts of China (grant no. B14027) and the Fundamental Research Funds for the Central Universities, the Central Public-interest Scientific Institution Basal Research Fund (grant no, Y2023QC21 to W.H.T.).
Author information
Authors and Affiliations
Contributions
S.J.Z., W.H.T., Z.J.D., and J.Y.Y. conceived and designed the experiments. J.Y.Y., W.H.T., and D.R.Z. performed the experiments. Q.Y.Y., J.M.X., G.X.L. and J.Y.Y. (Jing Ying Yan) assisted in plant material generation. J.Y.Y. and W.T.H. analyzed the data and generated the figures. W.N.D., C.W.J., and M.B. provided suggestions and revised the manuscript. J.Y.Y., Z.J.D., and S.J.Z. wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Tomasz Sarnowski, Naoki Yamaji and the other anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ye, J.Y., Tian, W.H., Zhang, D.R. et al. BRAHMA represses STOP1-NRT1.1 module to control plant rhizosphere alkalization and acid stress adaptation. Nat Commun 17, 3084 (2026). https://doi.org/10.1038/s41467-026-69905-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-69905-z
