
Abstract
Blood-stage malaria infection significantly increases the number of immune cells in the peripheral blood of vertebrate hosts, yet their roles in malaria transmission by mosquitoes remain poorly understood. In this study, we demonstrate that macrophages/monocytes—rather than neutrophils or NK/NKT cells—play a crucial role in enhancing malaria transmission to mosquitoes, both in rodent models and human. Mechanistically, these macrophages/monocytes that migrate into the mosquito midgut suppress the growth of transmission-blocking microbiota, including E. anophelis, S. sonnei, and K. oxytoca, primarily through non-opsonic phagocytosis mediated by scavenger receptor A (SR-A). Importantly, either the depletion of macrophages/monocytes or the use anti-SR-A neutralizing antibodies significantly enhances the transmission-blocking efficacy of anti-Pfs25 monoclonal antibodies by increasing the abundance of these bacteria. This combination approach can lead to the complete blockade of transmission of Pfs25-transgenic Plasmodium berghei (P. berghei-pfs25). These findings reveal a previously unrecognized mechanism that the parasite co-opts host immune cells to eliminate transmission-blocking bacteria, suggesting opportunities for using anti-SR-A antibodies to inhibit malaria transmission and improve the efficacy of anti-Pfs25 monoclonal antibodies.
Similar content being viewed by others

Introduction
Malaria continues to be one of the deadliest infectious diseases worldwide1. Although the use of insecticides and antimalarial drugs has significantly reduced malaria infections over the past few decades, the number of cases has plateaued and even increased in recent years2. Consequently, there is an urgent need for new intervention strategies to combat this infectious disease3. Malaria is transmitted through the bite of infected female mosquitoes. After a bite, both female and male gametocytes enter the mosquito’s midgut, where they fertilize to form zygotes. These zygotes then develop into ookinetes, which traverse the midgut epithelium and mature into oocysts. Once matured, the oocysts rupture, releasing sporozoites into the hemolymph. Finally, the sporozoites invade the salivary glands and are injected into the skin when the infected mosquito bites a person4. Thus, targeting mosquito infection represents a promising strategy to disrupt malaria transmission.
Following penetration of the midgut epithelium and subsequent development into oocysts, malaria parasites largely evade detection by the mosquito immune system, facilitating their maturation into sporozoites within the salivary glands5,6. In contrast, a substantial loss of malaria parasites occurs during their development in the midgut, rendering this stage a critical target for transmission-blocking interventions4. Several factors contribute to parasite attrition within the midgut. For instance, mosquito midgut microbiota can directly eliminate malaria sexual stages through the secretion of lipases or the generation of reactive oxygen species7,8,9. Additionally, mosquito midgut microbiota can also indirectly impede parasite development by modulating peritrophic matrix formation10 or mosquito immune responses11,12. Furthermore, the invasion of the midgut epithelium by ookinetes elicits both localized and systemic immune responses, leading to parasite destruction13,14,15,16.
Emerging evidence indicates that host-derived serum factors, including gametocyte-targeted antibodies and transforming growth factor (TGF)-β1, also enter the mosquito midgut during natural malaria transmission and can modulate malaria transmission17,18. It has been shown that malaria blood-stage infections induce a significant expansion of immune cells in the peripheral blood of mice19. However, it remains largely unknown whether immune cells that are taken into the mosquito midgut during bites from malaria patients can also influence malaria transmission20,21.
Here, we revealed that innate immune cells were expanded in the peripheral blood of the infected mice, and macrophages/monocytes enhanced malaria transmission by reducing the abundance of transmission-blocking microbiota, including E. anophelis, S. sonnei and K. oxytoca in the mosquito midgut via SR-A-mediated phagocytosis.
Results
Malaria infection primarily expands innate immune cells in peripheral blood of mice
Mosquitoes acquire malaria parasites by biting infected hosts. During these bites, immune cells from the host’s peripheral blood, along with the parasites, are ingested into the mosquito’s midgut. However, it remains unclear whether these ingested immune cells influence the development of the parasites within the mosquito. To explore the impact of vertebrate host-derived immune cells on malaria transmission, we first investigated whether blood-stage malaria infections could trigger the expansion of immune cells in the peripheral blood.
Four days post-infection with P. yoelii or P. berghei, blood samples were collected from the tails of infected mice, and the frequencies of innate immune cells, including DCs, macrophages, monocytes, NK/NKT Cells and neutrophils, as well as adaptive immune cells, such as CD4+/CD8+ T cells, B cells and DCs, were analyzed using flow cytometry (FACS) (Fig. 1a, Supplementary Fig. 1). Notably, we observed a significant increase in the frequency of neutrophils (CD11b⁺Ly6G⁺), macrophages (CD11b⁺F4/80⁺), and monocytes (CD11b⁺F4/80⁻) in the peripheral blood of parasite-infected mice. Initially, the frequency of DCs (CD11c⁺MHC II⁺) was also found to increase in response to either P. yoelii or P. berghei infection, but no significant change was observed for P. berghei infection after three replications (Fig. 1b–e). The frequencies of NK/NKT cells (NK1.1⁺), CD4⁺ T cells (CD3α⁺CD4α⁺), and B cells (CD19⁺CD220⁺) did not change in the peripheral blood of mice infected with either P. yoelii or P. berghei. However, parasite infection reduced the frequency of CD8⁺ T cells (CD3α⁺CD8α⁺) (Fig. 1f–i). These findings suggest that malaria infection primarily expands specific immune cell populations in the peripheral blood of mice.

a Representative FACS image of innate immune cells (upper panel, red square), including macrophages, monocytes, NK/NKT cells, DCs and neutrophils, and adaptive immune cells (lower panel, blue square), such as CD4+ T, CD8+ T and B cells, in peripheral blood at 4 days post P. yoelii or P. berghei infection. b–i Statistically analysis of the percentage of neutrophils (b), monocytes (c), macrophages (d), DC cells (e), NK/NKT cells (f), CD4+ T cells (g), CD8+ T cells (h), and B cells (i) in peripheral blood of mice (n = 9) 4 days post P. yoelii or P. berghei infection. Three independent experiments were performed for each experiment, and the data were pooled. The data were represented as the mean ± SD. Source data are provided as a Source Data file. Significance was determined by an unpaired t-test (two-tailed). Number between each group represented the P-value; P < 0.05 was considered as statistically significant.
Macrophages/monocytes, but not neutrophils and NK/NKT cells, significantly promote both rodent and human malaria transmission
To investigate the role of expanded innate immune cells in malaria transmission, we analyzed innate immune cell populations in the mosquito midgut three hours after the mosquitoes fed on parasite-infected mice using FACS. All major innate immune cell types were detected, with particularly high levels of NK/NKT cells, monocytes and neutrophils (Fig. 2a), suggesting a potential role for these cells in influencing malaria transmission.

a The percentage of neutrophils, monocytes, macrophages, DCs, and NK/NKT cells in the midguts of mosquitoes (n = 9) 3 h post-feeding on P. yoelii-infected mice was analyzed using FACS. Twenty midguts were pooled to create one sample, and each has three biological replicates. b Experimental scheme. Mice were treated with or without anti-Ly6G, anti-NK1.1, or CL at 3 days after infection with P. yoelii. Twenty-four hours later, mice were fed to mosquitoes. c, d Mice were treated with or without anti-Ly6G (c), or anti-NK1.1 (d) at 3 days after infection with P. yoelii. Twenty-four hours later, these mice were fed to mosquitoes, and 7 days later, the oocysts number were counted. Both infection intensity and prevalence were calculated and compared. e–j Mice were treated with or without CL at 3 days after infection with P. yoelii (e–g, i) or P. berghei (h, j). Twenty-four hours later, mice were fed to mosquitoes. The relative expression of ookinete-specific genes (n = 9), including SOAP (e) and WARP (f), to the internal control S7 was detected by qPCR in mosquitoes at 12 h post-feeding on CL-treated and control mice. Twenty mosquitoes constitute one sample, and three biological replicates were performed. Both the oocyst infection intensity and prevalence of P. yoelii (g) and P. berghei (h) at 7- or 9-days post-blood meal was compared, respectively. The individual dot represents oocysts number per mosquito, with a line indicating the median. The infection intensity and prevalence of P. yoelii (i) or P. berghei (j) sporozoites in the salivary gland of mosquitoes (n = 15) were counted and compared at 18 days post blood meal. Ten salivary glands constitute one sample, and five biological replicates were performed. k, l At 18 days post-feeding on P. yoelii- or P. berghei-infected mice (treated with or without CL), 5 mosquitoes were allowed to fed on naïve mice, and the onset of blood stage infection were monitored daily for 12 days. The onset of the blood stage of P. yoelii (k) or P. berghei (l) in mice (n = 15) was monitored, after infection by the bite of mosquitoes that fed on P. yoelii- or P. berghei-infected mice (treated with or without CL). m The infection intensity and prevalence of P. falciparum in mosquitoes that fed on P. falciparum-infected RBCs (red blood cells) with or without monocytes isolated from human peripheral blood. The individual dot in panels (c, d, g, h, m) represents oocysts number per mosquito, with a line indicating the median, and the pi chart under each scatter chart indicates the sample size and prevalence in each group. Three independent experiments were performed for each experiment, and the data were pooled. The data in panels (a, e, f, i, j) were presented as mean ± SD. Source data are provided as a Source Data file. Significance was determined by GLMMs test (two-sided) for (c, d, g, h, m); Unpaired t test (two-sided) for (e, f, i, j), Kaplan-Meier analysis was applied for (k, l), and Fisher’s exact test (two-sided) was applied for prevalence analysis. Number between each group represent the P-value, P < 0.05 was consider as statistically significant.
Since gametocytes are the only stage of the malaria parasite that can develop further in mosquitoes, factors influencing gametocytemia in mice are potentially important for malaria transmission. We monitored gametocytemia in mice treated with anti-Ly6G (to deplete neutrophils), anti-NK1.1 (to deplete NK/NKT cells), or clodronate liposomes (CL) (to deplete monocytes and macrophages). We found no significant changes in gametocytemia in either P. yoelii or P. berghei-infected mice pre-treated with anti-Ly6G, anti-NK1.1, or CL (Supplementary Fig. 2a, b). Furthermore, the in vitro transformation of gametocytes from anti-Ly6G, anti-NK1.1, and CL-treated infected mice into ookinetes was comparable to that of gametocytes from control mice (Supplementary Fig. 2c, d). This indicates that the depletion of neutrophils, NK/NKT cells, or macrophages/monocytes does not significantly impact gametocyte function in mice.
Next, mice infected with P. yoelii for three days were treated with anti-Ly6G or anti-NK1.1. Twenty-four hours later, mosquitoes were allowed to feed on these mice, and oocyst numbers were quantified 7 days post-blood meal (Fig. 2b). No significant difference in either infection intensity or prevalence was observed in mosquitoes fed on P. yoelii-infected mice depleted of neutrophils (anti-Ly6G-treated) or NK/NKT cells (anti-NK1.1-treated) compared to the control (Fig. 2c, d, Supplementary Fig. 3a–d). These results suggest that neither neutrophils nor NK/NKT cells significantly influence malaria transmission.
Then, mice were treated with a high dose of CL to selectively deplete monocytes and macrophages in peripheral blood. FACS confirmed that CL treatment significantly reduced the frequency of macrophages/monocytes but had no significant effect on CD4+ T cells, CD8+ T cells, or B cells (Supplementary Fig. 3e–h). These results demonstrate the specificity of CL-mediated depletion for macrophages/monocytes. Interestingly, we found that mosquitoes feed on CL-treated mice displayed significantly reduced expression of secreted ookinete adhesive protein (SOAP) and von Willebrand factor A domain-related protein (WARP), key markers of ookinete development22,23, at 12 h post-infection (Fig. 2e, f). This suggests that ookinete numbers are lower in mosquitoes fed on CL-treated mice compared to controls. In addition, the depletion of macrophages/monocytes using CL substantially reduced both infection intensity and prevalence of mosquitoes, regardless of whether they had fed on P. yoelii- or P. berghei-infected mice (Fig. 2g, h). By day 18, only a few sporozoites were detectable in mosquito salivary glands that had fed on P. yoelii-infected mice pretreated with CL (Fig. 2i), while no sporozoites were found in the salivary glands of mosquitoes feed on the CL-treated P. berghei-infected mice (Fig. 2j). Furthermore, the percentage of mice infected with P. yoelii was significantly reduced, and even no mice were infected with P. berghei following bites from mosquitoes that had fed on the infected mice pretreated with CL (Fig. 2k, l).
Evidence has shown that monocyte frequency in peripheral blood was also upregulated upon P. falciparum infection24. So, monocytes from healthy adult peripheral blood were isolated and then added to in vitro cultured P. falciparum gametocytes. The influence of human monocytes on the transmission of P. falciparum was evaluated by the standard membrane feeding assay (SMFA). SMFA revealed that the addition of human monocytes significantly increased both the intensity and prevalence of P. falciparum infection (Fig. 2m). These results demonstrate that macrophages/monocytes, rather than neutrophils and NK/NKT cells, play a crucial role in facilitating both rodent and human malaria transmission.
Macrophages/monocytes promote malaria transmission by reducing microbiota abundance in mosquito midgut
The depletion of macrophages/monocytes has no significant effect on gametocyte generation in mice, indicating that macrophages/monocytes exert their function in mosquitoes to facilitate malaria transmission. A previous study showed that monocytes present in the mosquito midgut might phagocytose and kill developing gametes25. However, our findings indicate that the depletion of macrophages/monocytes does not promote, but rather inhibits, malaria transmission (Fig. 2e–m). Therefore, the impact of monocytes on malaria transmission is not due to their direct action on malaria parasites within the mosquito midgut.
Recent evidence suggests that microbiota in the mosquito midgut can modulate malaria transmission by directly killing the parasite or inducing anti-malarial immune responses in mosquitoes26. This raises the question of whether macrophages/monocytes influence the microbial composition of the mosquito midgut. We found that mosquitoes fed on CL-treated mice exhibited a higher abundance of microbiota at 1- and 12-h post-blood meal, by colony-forming units (CFUs) quantification assay (Fig. 3a, b) and 16S rDNA qPCR (Fig. 3c). As microbiota proliferation after the blood meal has been found to affect mosquito lifespan27, we tested whether the increased microbial abundance in mosquitoes feed on CL-treated mice would reduce their survival. As expected, mosquitoes that fed on CL-treated mice exhibited a significantly lower survival rate compared to those that fed on control mice (Fig. 3d). To further investigate the role of midgut microbiota in the enhanced parasite infection observed in mosquitoes feeding on CL-treated mice, we pretreated mosquitoes with antibiotics to eliminate their native microbiota. Subsequently, mosquitoes with or without microbiota were fed on either control or CL-treated mice. Following antibiotic-mediated clearance of the midgut microbiota, oocyst numbers were significantly higher than in control mosquitoes that did not receive antibiotic treatment—a finding consistent with a previous study7. However, there was no significant difference in the number of oocysts or their prevalence between mosquitoes without microbiota, regardless of whether they were fed on the control or CL-treated mice (Fig. 3f). These findings suggest that macrophage/monocytes not only promote malaria transmission, but also enhance their longevity by reducing microbiota abundance in the mosquito midgut.

a, b CFUs were counted in the midguts of mosquitoes (n = 15) at 1 and 12 h after feeding on control or CL-treated mice. Ten mosquito midguts were pooled as one biological replicate, and five biological replicates were performed. c Total bacteria load in midguts of mosquitoes (n = 9) at 12 h post-feeding on control or CL-treated mice was detected by qPCR. Twenty mosquitoes were pooled as one biological replicate, and three biological replicates were performed. d The survival rate of mosquito (n = 90) feed on the P. yoelii-infected mice pre-treated with or without CL. e The bacterial burden in midguts of mosquitoes (n = 9) was determined by CFUs after being treated with antibiotics. Ten mosquitoes were pooled as one biological replicate, and each group has three biological replicates. f After treatment with or without antibiotics, mosquitoes were fed on infected mice pre-treated with or without CL. Seven days later, the infection intensity and prevalence of mosquitoes were determined and compared. Three independent experiments were performed for each experiment, and the data were pooled. The data in panels of (a–c, e) were presented as mean ± SD. The individual dot in panel (f) represents oocysts number per mosquito, with a line indicating the median, and the pi chart under each scatter chart indicates the sample size and prevalence in each group. Source data are provided as a Source Data file. Significance was determined by unpaired t-test (two-tailed) for (a–c, e); Kaplan-Meier analysis was applied for (d); GLMMs test (two-sided) for (f), and prevalence was analyzed by Fisher’s exact test (two-sided). Number between each group represent the P value, P < 0.05 was consider as statistically significant.
The depletion of macrophages/monocytes significantly increases the abundance of bacteria with transmission-blocking activity in mosquito midgut
The composition of the gut microbiota plays a crucial role in malaria infection. So, which specific bacterial differences contribute to this phenomenon? To address this question, midguts were collected from mosquitoes fed on control and CL-treated mice, and 16S rDNA sequencing was performed and compared. The midgut microbiota composition of mosquitoes fed on the CL-treated mice differed significantly from mosquitoes fed on the control mice (Fig. 4a). Notably, the abundances of E. anophelis—the dominant species in our laboratory An. stephensi strain—as well as S. sonnei and K. oxytoca, were significantly higher in mosquitoes that fed on CL-treated mice compared to controls. The increased abundance of E. anophelis, S. sonnei, and K. oxytoca in mosquitoes that fed on CL-treated mice was confirmed by qPCR (Fig. 4b–d). However, no significant changes were observed for other bacteria, such as Dietzia natronolimnaea, Nesteren salmonea, and Asaia spathodeae, which were also verified by qPCR (Supplementary Fig. 4a–c). This prompted us to investigate whether macrophages/monocytes enhance malaria transmission by decreasing the abundance of E. anophelis, S. sonnei or K. oxytoca in the mosquito midgut.

a The relative and absolute abundance of microbiota in the midgut of mosquitoes at 12 h post-feeding on P. yoelii infected mice pre-treated with CL or control liposomes. b–d The relative expression of E. anophelis, or S. sonnei and K. oxytoca in the midgut of mosquitoes (n = 9) was detected by qPCR at 12 h post-feeding on infected mice pre-treated with or without CL or control liposomes. Twenty mosquitoes were pooled as one biological replicate, and three biological replicates were performed. e A schematic representation of the experiment. Mosquito midgut microbiota was removed by antibiotics, then colonized with E. anophelis, S. sonnei, K. oxytoca or S. marcescens before fed with P. yoelii infected mice, oocyst numbers were counted and compared at 7 days post infection. f, g After colonization with E. anophelis (f), S. sonnei (g) and K. oxytoca (g), mosquitoes were fed on P. yoelii infected mice pretreated with or without CL. Seven days later, the infection intensity and prevalence of mosquitoes were determined. h After colonization with E. anophelis, S. sonnei, or K. oxytoca, individually or together, mosquitoes were fed on the P. yoelii-infected mice. Seven days later, the infection intensity and prevalence of mosquitoes were determined. i After colonization with S. marcescens, mosquitoes were fed on the P. yoelii-infected mice pretreated with or without CL. Seven days later, the infection intensity and prevalence of mosquitoes were determined. Three independent experiments were performed for experiments from b to i, and the data were pooled. The data in panels of (b–d) were presented as mean ± SD. The individual dot in panels of (f-i) represents oocysts number per mosquito, with a line indicating the median, and the pi chart under each scatter chart indicates the sample size and prevalence in each group. Source data are provided as a Source Data file. Significance was determined by an unpaired t-test (two-tailed) (b–d), GLMMs (two-sided) test for (f–i), and Fisher’s exact test (two-sided) was applied for prevalence analysis. Triple: Mosquito was colonized with Ea, Ss, and Ko at the same time; Number between each group represent the P value, P < 0.05 was consider as statistically significant.
The endogenous microbiota in mosquitoes was eliminated by antibiotic treatment. Following this, the mosquitoes were fed a sugar solution supplemented with isolated E. anophelis, S. sonnei or K. oxytoca for their colonization in the midgut. After colonization, mosquitoes were fed on the infected mice pretreated with CL (Fig. 4e, Supplementary Fig. 4d–g). E. anophelis, or S. sonnei or K. oxytoca colonization significantly reduced both infection intensity and prevalence in mosquitoes, demonstrating their suppressive effect on parasite development. These effects were further enhanced in mosquitoes fed on CL-treated mice (Fig. 4f, g). We also investigated whether these bacteria impair parasite development in the mosquito midgut individually or synergistically. Mosquitoes were colonized with each of the three bacterial species individually or in combination and then allowed to feed on the infected mice. However, we did not observe an additive or synergistic effect in mosquitoes colonized with multiple bacterial species compared to those harboring a single bacterium (Fig. 4h). Therefore, we conclude that macrophages/monocytes contribute to malaria transmission by reducing the abundance of E. anophelis, as well as S. sonnei and K. oxytoca, in the mosquito midgut, but no synergistical inhibitory effect was found for those three bacteria in malaria transmission. Noticeably, there is a difference between the gut microbiota of long-term laboratory-raised Anopheles mosquitoes and that of wild Anopheles mosquitoes. S. marcescens, a naturally occurring symbiotic bacterium found in field-caught mosquitoes, exhibits antimalarial activity in mosquito12. Consequently, we colonized the midguts of laboratory-reared Anopheles mosquitoes with this bacterium to evaluate whether CL treatment of infected mice could similarly influence malaria transmission. Consistent with our hypothesis, CL-treatment also significantly reduced both infection intensity and prevalence in S. marcescens-colonized mosquitoes (Fig. 4i, Supplementary Fig. 4h). These findings suggested that the enhancement of malaria transmission by macrophages/monocytes may extend beyond specific microbial communities, highlighting a broader ecological relevance.
Macrophages/monocytes phagocytose microbiota in mosquito midgut dependent on SR-A
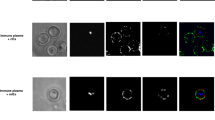
To investigate whether ingested macrophages/monocytes reduce midgut microbiota via phagocytosis, we first cleared the mosquito midgut microbiota with antibiotics and then colonized it with GFP-tagged E. anophelis. Mosquitoes were allowed to feed on P. yoelii-infected mice pre-treated with or without CL. Three hours post-feeding, midguts were dissected and analyzed by FACS to detect GFP-positive macrophages/monocytes. CL treatment significantly reduced the number of GFP-positive macrophages/monocytes (Fig. 5a). To distinguish between phagocytosed bacteria and surface-adherent GFP-tagged E. anophelis, extracellular bacteria were removed using antibiotic treatment as previously described28. Subsequent CFU assays revealed that macrophages/monocytes from mosquitoes fed on CL-treated mice phagocytosed significantly fewer bacteria than those from control-fed mosquitoes (Fig. 5b). We also assessed the survival of macrophages/monocytes in the mosquito midgut by FACS. A high frequency of macrophages/monocytes was detected at 1 and 3 h post–blood meal, but their numbers declined sharply by 12 h and were nearly undetectable by 24 h (Fig. 5c). These results indicate that macrophages/monocytes are present only transiently in the midgut and likely engage in active phagocytosis of midgut microbiota during the early phase following the blood meal.

a Mosquito midgut microbiota was removed by antibiotics and colonized with GFP-tagged-E. anophelis. Mosquitoes were allowed to fed on P. yoelii-infected mice treated with or without CL. Three hours post blood meal, the mosquito midgut was dissected, and FACS were used to detect GFP+ macrophages and monocytes (n = 9). Ten mosquitoes were pooled as one biological replicate, and three biological replicates were performed. b Three hours after mosquitoes fed on P. yoelii-infected mice pre-treated with CL or control liposomes, midguts were dissected and homogenized. The homogenate was incubated with antibiotics to eliminate extracellular bacteria, and then intracellular bacteria were released after lysis of macrophages/monocytes by Triton X. The released bacteria were cultured on LB agar plates for 24 h, and CFUs (n = 15) in each group were counted and compared. Ten mosquitoes were pooled as one biological replicate, and five biological replicates were performed. The right panel was the representative image of monocyte/macrophage phagocytosing bacteria in mosquito midgut at 3 h post blood meal, stained with Giemsa solution, scale bar: 5 μm. c The number of live macrophages/monocytes number at different time points in the mosquito midgut (n = 12) was detected by FACS. d The infection intensity and prevalence of mosquitoes fed on infected C3−/− mice pretreated with or without CL. e, f The infection intensity and prevalence of mosquitoes fed on infected mice pretreated with or without CL, or 10 μg (e) or 50 μg anti-SR-A (f). g The infected mice were pre-treated with 50 μg anti-SR-A at 1 day before, or 1 day post-infection, or treated with CL at 3 days post-infection, then the mice were fed to mosquitoes. 7 days post blood meal, oocysts number in mosquitoes was counted and compared. Three independent experiments were performed for each experiment, and the data were pooled. Data in (a–c) were presented as mean ± SD. In panels of (d–g), the individual dot represents oocysts number per mosquito, with a line indicating the median, and the pi chart under each scatter chart indicates the sample size and prevalence in each group. Source data are provided as a Source Data file. Significance was determined by an unpaired t-test (two-tailed) for (a, b), GLMMs (two-sided) test for (d–h), and Fisher’s exact test (two-sided) was applied for prevalence analysis. Number between each group represent the P-value, P < 0.05 was consider as statistically significant.
Since microbiota in the mosquito midgut do not induce the generation of antibodies, their phagocytosis is unlikely to be mediated by opsonization through Fc receptors on macrophages/monocytes. Furthermore, in the absence of antibodies, the classical pathway cannot activate C3. However, host-derived C3 ingested into the mosquito midgut may be activated via the alternative pathway to generate C3b29. This suggests that macrophages/monocytes might phagocytose bacteria opsonized by C3b. Notably, the depletion of macrophages/monocytes in C3−/− mice treated with CL still reduces malaria transmission (Fig. 5d). Therefore, it appears that macrophages/monocytes phagocytose bacteria independent on complement.
SR-A is one of the non-opsonic receptors involved in the phagocytosis of bacteria by macrophages/monocytes30. The treatment of parasite-infected mice with 10 ug anti-SR-A could significantly reduce malaria transmission (Fig. 5e), and 50 μg anti-SR-A could even reduce malaria transmission to a level comparable to CL treatment (Fig. 5f). We also observed the persistent blocking effect of anti-SR-A. The infected mice were pre-treated with or without anti-SR-A at −1, or 1 day, then allowed to fed to mosquitoes. As a result, the neutralization of SR-A at 1 day before infection (5 days before the blood meal) did not affect the number of oocysts in the mosquito midgut but reduced oocyst prevalence. In contrast, the administration of anti-SR-A antibody at 1-day post-infection (3 days before the blood meal) achieved a level of transmission reduction comparable to that observed in the CL-treated group (Fig. 5g), suggesting that antibody-mediated neutralization of SR-A can suppress malaria transmission for at least three days. Taken together, our data suggest that SR-A was responsible for the non-opsonic phagocytosis of bacteria in mosquito midgut by macrophages/monocytes, thereby promoting malaria transmission.
Anti-Pfs25 antibodies completely block malaria transmission in the presence of anti-SR-A antibodies
Malaria transmission blocking vaccines are designed to induce antibodies against surface proteins on sexual stages31. Pfs25, one of the leading transmission blocking vaccines, is thought to induce anti-Pfs25 antibodies to block the penetration of ookinetes through the midgut epithelium32,33. However, the regulation of the blocking efficacy of anti-Pfs25 antibodies is still largely unknown. Since monocyte/macrophage depletion suppressed natural malaria transmission by increasing transmission-blocking bacteria, we investigated whether it could also enhance the transmission-blocking efficacy of anti-Pfs25. Mice infected with P. berghei-pfs25 (Pfs25-transgenic P. berghei) were treated with or without CL and then administered anti-Pfs25 antibodies before mosquito feeding (Fig. 6a). While anti-Pfs25 antibodies alone significantly reduced transmission, complete blocking was achieved only when combined with CL-mediated monocyte/macrophage depletion (Fig. 6b–e). Furthermore, anti-Pfs25 antibodies also completely block the transmission of P. berghei-pfs25 in the presence of anti-SR-A antibodies (Fig. 6f–j). This effect was closely linked to the increased burden of E. anophelis, S. sonnei and K. oxytoca in the midgut of mosquitoes feed on the infected mice pretreated with anti-SR-A antibodies (Fig. 6k–m). Therefore, we demonstrated that the transmission of P. berghei-pfs25 can be completely blocked by anti-Pfs25 antibodies following the inhibition of SR-A-mediated phagocytosis of transmission-blocking bacteria by macrophages/monocytes.

a Experimental design. After mice were infected with P. berghei-pfs25 transgenic parasites for 3 days, mice were treated with or without CL, and injected with anti-Pfs25 antibodies one hour before fed to mosquitoes. Oocysts number of mosquitoes were counted 9 days post-blood meal. b–e After pretreated with or without CL, the P. berghei-pfs25-infected mice were then inoculated with anti-Pfs25; both the infection intensity (b) and prevalence (c) of mosquitoes, as well as the transmission reduction activity (d) and transmission blocking activity (e), were calculated. f Experimental design. After mice were infected with P. berghei-pfs25 transgenic parasites for 3 days, mice were treated with or without anti-SR-A, and injected with anti-Pfs25 antibodies one hour before fed to mosquitoes. Oocysts number of mosquitoes were counted 9 days post-blood meal. g, h After pretreated with or without anti-SR-A, the P. berghei-pfs25-infected mice were then inoculated with anti-Pfs25, both the infection intensity (g) and prevalence (h) of mosquitoes, as well as the transmission reduction activity (i) and transmission blocking activity (j), were calculated based on infection intensity and prevalence. k–m The relative expression of E. anophelis, or S. sonnei and K. oxytoca in the midgut of mosquitoes was detected by qPCR at 12 h post-feeding on infected mice pre-treated with anti-SR-A, anti-Pfs25 or both. Twenty mosquitoes were pooled as one biological replicate, and three biological replicates were performed. The individual dot in panels of (b, g) represents oocysts number per mosquito, with a line indicating the median, and the pi chart under each scatter chart indicates the sample size and prevalence in each group. Data were presented as mean ± SD (k–m). Three independent experiments were performed for each experiment, and the data were pooled. Source data are provided as a Source Data file. Significance was determined by GLMMs (two-sided) test for (b, g), and Fisher’s exact test (two-sided) was applied for (c, h); an unpaired t-test (two-tailed) was applied for (k–m). Number between each group represent the P-value, P < 0.05 was consider as statistically significant.
Discussion
Female adult mosquitoes require blood meals for egg development. During this process, they also ingest parasites and vertebrate host-derived factors into their midgut. While some vertebrate host-derived factors have been shown to regulate malaria transmission34,35,36,37,38,39. The effects of immune cells on malaria transmission remain largely unexplored. In this study, we report that immune cells expanded due to malaria infection are ingested into the mosquito midgut during a blood meal. Notably, only macrophages/monocytes—not neutrophils, and NK/NKT cells—facilitate malaria transmission by reducing transmission-blocking microbiota. The phagocytosis of microbiota, including E. anophelis, S. sonnei and K. oxytoca, by macrophages/monocytes primarily dependent on SR-A, rather than the ingestion of malaria parasites, ultimately contributes to an increase in malaria transmission.
The roles of immune cells have been investigated in the context of malaria transmission in the presence of immune serum20. CD4+ T cells activated by immunization with P. yoelii microgametes have been shown to inhibit malaria transmission by decreasing gametocytemia in the vertebrate host21. However, CD4+ T cells did not have a significant impact on malaria transmission following whole-killed blood-stage vaccination40. In vitro phagocytosis of gametes by macrophages/monocytes and neutrophils was positively correlated with levels of antibodies against the gamete surface proteins Pfs230 and Pfs48/45, but solid evidence is still lacking to support their roles in the reduction of malaria transmission25. To our knowledge, this is the first report demonstrating that macrophages/monocytes that enter the midgut promote, rather than suppress, natural transmission of both rodent and human malaria by reducing transmission-blocking bacteria. In a controlled human malaria infection study, classical monocytes (CD14+CD16−) were significantly elevated in peripheral blood, whereas non-classical monocytes (CD14−CD16+) were reduced41. This change suggests potential clinical implications for monocyte-mediated facilitation of malaria transmission, as supported by our findings on the enhanced effect of human monocytes on P. falciparum infection in mosquitoes. However, direct evidence is still needed to determine whether neutralization of SR-A can reverse the pro-transmission effect of human monocytes. Such evidence is essential to support the extrapolation of our findings from rodent models to human malaria. Notably, anti-Pfs25 antibodies completely blocked malaria transmission after the depletion of macrophages/monocytes, underscoring the critical role these immune cells play in regulating transmission-blocking interventions. These observations suggest that macrophages/monocytes may diminish the efficacy of anti-Pfs25 antibodies by modulating the composition of midgut microbiota. We speculate that macrophages and monocytes actively phagocytose microbiota during early blood digestion, while ookinetes opsonized by anti-Pfs25 antibodies might evade phagocytosis through Fc receptors. This immune evasion likely occurs because macrophages/monocytes do not persist into the later stages of infection when anti-Pfs25 antibodies bind to ookinetes. However, an experiment designed to evaluate the impact of the deficiency of monocyte/macrophage Fc receptor on the transmission-blocking efficacy of anti-Pfs25 has yet to be conducted to test this hypothesis.
Both macrophages/monocytes and neutrophils can phagocytose bacteria, yet it is noteworthy that neutrophils in the midgut have little effect on malaria transmission. In addition to bacteria, both cell types are capable of phagocytosing gametes, but neutrophils demonstrate a greater capacity for this process compared to macrophages/monocytes25. Therefore, while neutrophils may facilitate the development of the malaria sexual stage by phagocytosing bacteria, their role in phagocytosing gametes could counterbalance this effect, potentially explaining why their absence does not significantly impact malaria transmission.
A previous study demonstrated that CL treatment could trigger systemic cytokine responses in mice at 7 days post-infection with P. yoelii42. However, in our study, we did not evaluate the impact of CL treatment on cytokine dynamics. In addition, we identified SR-A as the primary phagocytic receptor mediating macrophage/monocyte uptake of gut microbiota, but the potential contributions from other receptors, such as CD36, DC-SIGN, and the mannose receptor, could not be completely ruled out. We recognize these limitations in our study.
We identified E. anophelis as the dominant symbiotic bacterium in the midgut of insectary-reared Mosquitoes, consistent with prior reports43,44. This predominance may stem from E. anophelis secreting complement membrane attack complex-like factors that eliminate competing microbes45. Notably, E. anophelis has been shown to inhibit sexual-stage development of Plasmodium in mosquitoes46,47. Based on these findings, it is reasonable for us to find that macrophages/monocytes could inadvertently promote malaria transmission by phagocytosing E. anophelis. Intriguingly, we also observed similar enhancement of transmission when macrophages/monocytes phagocytosed S. sonnei or K. oxytoca, both of which significantly inhibit malaria development in mosquitoes. However, the mechanisms underlying S. sonnei and K. oxytoca effects—whether through direct parasite killing or immune modulation—require further investigation.
In summary, we have uncovered a novel immune evasion strategy employed by malaria parasites, wherein they exploit host peripheral blood macrophages/monocytes to enhance transmission via SR-A-dependent phagocytosis of bacteria with transmission-blocking activity. Strikingly, depletion of these immune cells enabled anti-Pfs25 antibodies to achieve complete transmission blockade. These findings not only reveal a novel strategy for transmission-blocking interventions but also elucidate mechanistic insights that could significantly enhance the efficacy of next-generation malaria vaccines. Furthermore, this discovery may inform our understanding of transmission mechanisms in other vector-borne diseases.
Methods
Mosquito, parasites, and mice
An. stephensi (Hori strain) were maintained were reared and maintained under standard laboratory conditions at 27 °C with 75% RH (relative humidity) on a 12-h light-dark cycle. Rodent malaria parasites P. yoelii, P. berghei, P. berghei-pfs25 transgenic parasites (kindly donated from Xiamen University by Professor Jian Li) and the P. falciparum NF54 strain (kindly donated from the Chinese Academy of Sciences by Professor Lubin Jiang) were used in this study. Female C57BL/6 J mice, aged 6–8 weeks, were obtained from GemPharmat Animal Institute (Nanjing, China). C3−/− mice with a C57BL/6 J background were purchased from Jackson Laboratory. The mice were housed in a controlled environment with regulated temperature and humidity and maintained on a 12-h light/dark cycle, with lights off at 18:00 h. All animal procedures were reviewed and approved by the Animal Ethics Committee of the Army Medical University Institute of Medical Research under protocol number AMUWEC20218022.
Mice infection
C57BL/6 J mice female mice aged at 6-8 weeks were intravenously (i.v) injected with 1 × 106 (4 × 106 for P. berghei-pfs25) P. yoelii, P. berghei or P. berghei-pfs25 parasitized red blood cells (pRBCs). Parasitemia and gametocytemia were checked before fed to mosquitos by examining Giemsa-stained blood films under a light microscope.
Mosquito infection
An. stephensi were deprived of sugar meal for 24 h, then allowed to feed on P. yoelii-infected mice for 30 min. Not fully engorged mosquitoes were removed. Mosquitoes were maintained at 23–24 °C environment, with 70 % RH. At 7 days (P. yoelii oocysts), 9 days (P. berghei oocysts) or 18 days (P. yoelii and P. berghei salivary gland sporozoites) post infection, mosquitoes were dissected, and oocysts and salivary gland sporozoites were counted, respectively.
Flow cytometry analysis
Peripheral blood of mice infected with P. yoelii or P. berghei was collected from the orbital vein, and RBCs were lysed by Gey’s solution. Sample collected from mosquito midgut were gently homogenate with RPMI-1640 (Gibco, USA), by 1 ml pipette to release blood from its midgut, then RBCs were lysis by Gey’s solution after filter with a 70-micron mesh sieve. Then the cells were incubated with anti-CD16/CD32 antibodies (Biolegend, USA) for 10 min to block Fc receptors and then incubated with the antibodies of interest for 45 min. Antibodies used for flow cytometry analysis were as followed: anti-CD11b, anti-Ly6G, anti-MHC II, anti-CD3a, anti-CD4, anti-CD19, anti- CD8a, anti-CD3, and anti-NK1.1. Dead cells were excluded using LIVE/DEAD Fixable Dead Cell Stain Kit (Invitrogen, USA) according to the manufacturer’s instructions. Cells were collected on a BD FACS Canto II cytometer (BD Biosciences, USA) and analyzed by Flow Jo software v10.8.
Depletion of macrophages, NK/NK T cells, and neutrophils
After being infected with parasites for three days, mice were i.v injected with 200 μl CL (Liposoma, Netherlands), 200 μg anti-NK1.1 (Bio X Cell, USA) or 200 μg anti-Ly6G (Bio X Cell, USA) mono-antibody to deplete macrophages, NK/NKT cells, and neutrophils, respectively. The depletion efficacy was monitored by FACS analysis.
In vitro culture of ookinetes
After mice were infected with P. yoelii or P. berghei for 3 d, mice were i.v injected with 200 μl CL or control liposomes, 24 h later, blood was collected from orbit and immediately added to the ookinete culture medium which contains: RPMI-1640 (Gibco, USA), 10% fetal calf serum (Gibco, USA), 100 μM xanthurenic acid (MedChemExpress, USA), 25 mM Hepes (Sigma, USA); pH 8.0, 100 μg/ml streptomycin (Sangon, China), 100 U/ml penicillin (Sangon, China). The gametocytes were cultured at 19 °C (for P. berghei) and 23 °C (for P. yoelii) for 24 h to allow gametogenesis, fertilization, and ookinete differentiation. The generated ookinetes were calculated per 1000 RBCs in Giemsa-staining thin blood smears based on morphology; at least 50 fields were counted for each sample.
Detection of bacteria load in mosquito midgut
Colony forming unites (CFUs) assay was used to detect the bacterial load of mosquito’s midgut. Briefly, Mosquitoes were sterilization by spray 75 % ethyl alcohol, then washed three times with sterile PBS. Midguts were dissected under sterile conditions and homogenized. The homogenate was then diluted 10-fold and cultured on LB medium at 28 °C for 24 h. CFUs were calculated by the following equation:
Antibiotics treatment of mosquitoes
Three-days-old female mosquitoes were fed with 10 % sugar solution supplement with 10 U/ml penicillin (Sangon, China), 15 μg/ml streptomycin (Sangon, China), and 15 μg/ml gentamicin (Gibco, USA). Four days later, the bactericidal efficacy of antibiotics was accessed by the CFUs method.
Detection of bacterial phagocytosis by macrophages/monocytes
Method was midfield from described before28. Mice were infected with P. yoelii or for 3 days, followed by i.v injection of 200 µl of CL or control liposomes. Twenty-four hours post-injection, mosquitoes were allowed to feed on the mice for 30 min. Three hours after blood feeding, midguts were dissected, homogenized in 1 ml of PBS, and centrifuged (14,000 g, 5 min) to pellet bacteria and blood cells. The pellet was resuspended in 1 ml of PBS containing 100 U/ml penicillin, 100 µg/ml streptomycin, and 10 µg/ml gentamicin to eliminate extracellular bacteria. After 3 h of incubation, the homogenate was centrifuged (14,000 g, 5 min) and washed twice with sterile PBS to remove antibiotics. Macrophages and monocytes were lysed with 0.1% Triton X-100 (Sigma, USA) to release intracellular bacteria. Lysates were plated on LB agar, and bacterial CFUs were quantified after 24 h of incubation at 37 °C.
P. falciparum culture and mosquito infection
The P. falciparum NF54 strain was used to culture mature gametocytes, as described previously48. Matured gametocytes were diluted to 0.5% by adding fresh RBC, then resuspended with an equal volume of O+ serum and fed to mosquitoes by SMFA. To investigate the influence of human-derived monocytes on P. falciparum infection of mosquitoes, peripheral blood was collected from a healthy individual, and mononuclear cells were isolated by the Percoll method. Monocytes were then purified by Dynabeads™ Untouched™ Human Monocytes kit (Invitrogen, USA) according to the manufacture’s instructions. Isolated monocytes (5 × 105/ml) were added to in vitro cultured mature P. falciparum gametocytes. Nine days post blood meal, midguts of mosquitoes were dissected, and oocysts number was counted after being stained with 0.1% mercurochrome (MedChemExpress, USA) for 10 min.
Mosquito bite challenge
Mice infected with P. yoelii or P. berghei for 3 days were treated with CL or control liposomes, and then were allowed to fed to mosquitoes. Eighteen days later, 5 infected mosquitoes were fed on naïve mice; a fully engorged mosquito based on its abdomen was considered as a successful bite. Three days post mosquitoes’ bites, Giemsa-stained blood smears were made daily to check the on-set of blood stage infection for 12 days.
DNA extraction and 16S rRNA gene amplicon sequencing
Mosquitoes were surface-sterilized with 75% ethanol for 5 min and rinsed three times with sterile 1× PBS. Midguts were dissected aseptically, pooled into groups of 20 midguts per biological replicate, and homogenized. Total bacterial genomic DNA was isolated using the DNeasy PowerFecal Pro DNA Kit (QIAGEN, USA) according to the manufacturer’s protocol. DNA purity and concentration were determined spectrophotometrically (NanoDrop One, Thermo Fisher Scientific, USA) by measuring absorbance ratios at 260/280 nm and 260/230 nm. The hypervariable V3–V4 region of the bacterial 16S rRNA gene was amplified using universal primers flanked by Illumina adapters and sample-specific barcode sequences. Primary PCR products were purified with VAHTS™ DNA Clean Beads (Vazyme Biotech, China), and libraries were quantified fluorometrically using the Quant-iT™ dsDNA High-Sensitivity Assay Kit (Thermo Fisher Scientific, USA). Equimolar amounts of purified amplicons were pooled and subjected to paired-end sequencing (2 × 250 bp) on the Illumina NovaSeq 6000 platform (Illumina, USA). Absolute abundance was calculated by times the relative change of CFUs between the Control and CL-treated group.
Quantitative real-time PCR (qPCR)
To quantify the bacterial load, total DNA was extracted from mosquitoes that had fed on P. yoelii-infected mice treated with CL or control liposomes, using the DNeasy PowerFecal Pro DNA Kit (QIAGEN, USA) according to the manufacturer’s protocol. One hundred nanograms of DNA per sample were used as the template for qPCR to detect total and specific bacterial loads. For quantify the SOAP and WARP expression of ookinetes, RNA from mosquito midgut was extracted 12 h post blood meal, cDNA was obtained by using reverse transcription kit (Takara Bio Inc., Japan) following the manufacture’s protocol. Amplifications were performed with TB Green® Premix Ex Taq™ II (Takara Bio Inc., Japan) on a CFX96 Real-Time PCR Detection System (Bio-Rad, USA). All reactions included no-template controls to rule out contamination. Relative quantification was performed using the mosquito ribosomal protein S7 (S7) gene as an endogenous control. Data are expressed as fold-changes normalized to S7. Primers used in this study were listed in Supplemental Table 1.
Mosquito survival assay
Mice were infected with P. yoelii, three days later, mice were i.v injected with 200 μl CL or control liposomes, 24 h later, mosquitoes were allowed to fed on mice for 30 mins. Not fully engorged mosquitoes were removed. Mosquitoes were maintained at a 23–24 °C environment, with 70 % RH. Survival condition was recorded daily for 7 days.
Bacteria isolation and identification
Midguts were dissected, homogenized, and serially diluted (10-fold) on LB agar for 28 °C/24-h culture. Colonies underwent morphology-based classification (shape/color/size). Bacterial 16S rDNA was amplified via PCR (universal primers), purified, and sequenced (ABI 3730, USA). Sequences were BLAST-aligned against GenBank for identification.
Colonization of bacteria in mosquito midgut
After clearing the symbiont bacteria in the midgut with antibiotics, mosquitoes were fed a sugar solution containing 1 × 106/mL isolated E. anopheles, S. sonnei, K. oxytoca, or S. maecescens (kindly donated from the Chinese Academy of Sciences by Professor Sibao Wang) for 2 days to colonize E. anophelis into the mosquito midgut. A successful colonization was confirmed by qPCR (See S table 1 for primer information) or using a fluorescence microscope (for eGFP-E. anophelis) as previously described47.
Neutralization of SR-A receptor
Three days after P. yoelii infection, mice were i.v injected with 10 or 50 μg neutralizing anti-SR-A monoclonal antibody (R & D, USA). Twenty hours later, mice were fed to mosquitoes. To observe the longevity of SR-A blocking effect, the infected mice were pre-treated with 50 μg anti-SR-A at 1 day before infection, or 1 day post infection, and treated with CL 3 days post infection, 7 days post blood meal, oocysts number in mosquitoes were counted and compared.
Anti-Pfs25 transmission blocking assay
Anti-Pfs25 antibody was customized by GenScript (Shanghai, China) as previously described47. To evaluate the ability of anti-Pfs25 antibody to block P. berghei-pfs25 transmission, mice were i.v injected with 4 × 106 P. berghei-pfs25 pRBCs; Three days later, mice were i.v injected with 200 μl CL, 24 h later, 20 μg of anti-Pfs25 antibody was i.v injected 1 h before mosquito feeding. To determine the enchantment of anti-Pfs25 antibody block P. berghei-pfs25 transmission efficacy in the presence of anti-SR-A, mice were i.v injected with 4 × 106 P. berghei-pfs25 pRBCs; Three days later, mice were i.v injected with 50 μg anti-SR-A, 24 h later, 20 μg of anti-Pfs25 antibody was i.v injected 1 h before mosquito feeding. Nine days later, mosquitoes were dissected, and the oocysts were counted and compared. Transmission reduction activity (TRA) and transmission blocking activity (TRB) were calculated based on oocyst intensity and prevalence by following formula:
Statistics analysis
Analyses were performed using GraphPad Prism 10.0 (GraphPad Software, USA) and R Studio (version 2024.12.1 + 563). Differences of oocyst number between treatment and control groups were analyzed using the GLMMs (Generalized Linear Mixed Models) test. Specifically, if overdispersion is detected, a negative binomial regression is employed. Subsequently, the DHARMa package was used to test for zero inflation; if present, a zero-inflated model was applied for the final analysis. Fisher’s exact test (two-sided) was applied to compare infection rates. Kaplan-Meier test was used for comparing the ability of infected mosquito to transmit malaria and mosquito survival rate. Pooled data sets were determined by an unpaired t-test (two-tailed). Specific analysis methods were detailed in each figure legend. Numbers between each group represent the P-value; P < 0.05 was consider as statistically significant.
Ethics statement
Human blood was collected from healthy donors who provided written informed consent at Southwest Hospital of Army Medical University. Anonymous human blood and serum used for P. falciparum culture were obtained from the Chongqing Blood Bank. The in vitro culture of P. falciparum and subsequent infection of mosquitoes with the parasite were conducted in BSL-2+ laboratories, with oversight and approval from the Ethics Committee of the Army Medical University Institute of Medical Research (KY2025153).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The 16S rDNA sequence data generated in this study have been deposited in the National Center of Biotechnology Information Sequence Read Archive database under accession code PRJNA1209247. Source data are provided as a Source Data file. Source data are provided with this paper.
References
Malaria: (still) a global health priority. EClinicalMedicine 34, 100891. https://doi.org/10.1016/j.eclinm.2021.100891 (2021)
WHO. World Malaria Report 2024. (2025).
Alonso, P. L. & Tanner, M. Public health challenges and prospects for malaria control and elimination. Nat. Med. 19, 150–155 (2013).
Beier, J. C. Malaria parasite development in mosquitoes. Annu Rev. Entomol. 43, 519–543 (1998).
Zhu, F. et al. Malaria oocysts require circumsporozoite protein to evade mosquito immunity. Nat. Commun. 13, 3208 (2022).
Smith, R. C. & Barillas-Mury, C. Plasmodium Oocysts: Overlooked Targets Of Mosquito Immunity. Trends Parasitol. 32, 979–990 (2016).
Dong, Y., Manfredini, F. & Dimopoulos, G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 5, e1000423 (2009).
Moreira, L. A. et al. A Wolbachia Symbiont In Aedes Aegypti Limits Infection With Dengue, Chikungunya, And Plasmodium. Cell 139, 1268–1278 (2009).
Kumar, S., Molina-Cruz, A., Gupta, L., Rodrigues, J. & Barillas-Mury, C. A peroxidase/dual oxidase system modulates midgut epithelial immunity in Anopheles gambiae. Science 327, 1644–1648 (2010).
Feng, Y. et al. Anopheline mosquitoes are protected against parasite infection by tryptophan catabolism in gut microbiota. Nat. Microbiol. 7, 707–715 (2022).
Cirimotich, C. M. et al. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science 332, 855–858 (2011).
Gao, H. et al. A natural symbiotic bacterium drives mosquito refractoriness to Plasmodium infection via secretion of an antimalarial lipase. Nat. Microbiol. 6, 806–817 (2021).
Luckhart, S., Vodovotz, Y., Cui, L. & Rosenberg, R. The mosquito Anopheles stephensi limits malaria parasite development with inducible synthesis of nitric oxide. Proc. Natl. Acad. Sci. USA 95, 5700–5705 (1998).
Han, Y. S., Thompson, J., Kafatos, F. C. & Barillas-Mury, C. Molecular interactions between Anopheles stephensi midgut cells and Plasmodium berghei: the time bomb theory of ookinete invasion of mosquitoes. EMBO J. 19, 6030–6040 (2000).
Kumar, S., Gupta, L., Han, Y. S. & Barillas-Mury, C. Inducible peroxidases mediate nitration of anopheles midgut cells undergoing apoptosis in response to Plasmodium invasion. J. Biol. Chem. 279, 53475–53482 (2004).
Dimopoulos, G., Seeley, D., Wolf, A. & Kafatos, F. C. Malaria infection of the mosquito Anopheles gambiae activates immune-responsive genes during critical transition stages of the parasite life cycle. EMBO J. 17, 6115–6123 (1998).
Surachetpong, W., Singh, N., Cheung, K. W. & Luckhart, S. MAPK ERK signaling regulates the TGF-beta1-dependent mosquito response to Plasmodium falciparum. PLoS Pathog. 5, e1000366 (2009).
Dantzler, K. W. et al. Naturally acquired immunity against immature Plasmodium falciparum gametocytes. Sci. Transl. Med. 11, eaav3963 (2019).
Yazdani, S. S., Mukherjee, P., Chauhan, V. S. & Chitnis, C. E. Immune responses to asexual blood-stages of malaria parasites. Curr. Mol. Med 6, 187–203 (2006).
Lensen, A. H., Bolmer-Van de Vegte, M., van Gemert, G. J., Eling, W. M. & Sauerwein, R. W. Leukocytes in a Plasmodium falciparum-infected blood meal reduce transmission of malaria to Anopheles mosquitoes. Infect. Immun. 65, 3834–3837 (1997).
Harte, P. G., Rogers, N. C. & Targett, G. A. Role of T cells in preventing transmission of rodent malaria. Immunology 56, 1–7 (1985).
Dessens, J. T. et al. SOAP, a novel malaria ookinete protein involved in mosquito midgut invasion and oocyst development. Mol. Microbiol. 49, 319–329 (2003).
Abraham, E. G. et al. Analysis of the Plasmodium and Anopheles transcriptional repertoire during ookinete development and midgut invasion*. J. Biol. Chem. 279, 5573–5580 (2004).
Chimma, P. et al. A distinct peripheral blood monocyte phenotype is associated with parasite inhibitory activity in acute uncomplicated Plasmodium falciparum malaria. PLoS Pathog. 5, e1000631 (2009).
Healer, J., Graszynski, A. & Riley, E. Phagocytosis does not play a major role in naturally acquired transmission-blocking immunity to Plasmodium falciparum malaria. Infect. Immun. 67, 2334–2339 (1999).
Wang, J., Gao, L. & Aksoy, S. Microbiota in disease-transmitting vectors. Nat. Rev. Microbiol. 21, 604–618 (2023).
Gendrin, M. et al. Antibiotics in ingested human blood affect the mosquito microbiota and capacity to transmit malaria. Nat. Commun. 6, 5921 (2015).
Lu, S. M., Grinstein, S. & Fairn, G. D. in Phagocytosis and Phagosomes: Methods and Protocols. (ed R. Botelho) 79–91 (Springer New York, 2017).
Margos, G. et al. Interaction between host complement and mosquito-midgut-stage Plasmodium berghei. Infect. Immun. 69, 5064–5071 (2001).
Aderem, A. & Underhill, D. M. Mechanisms of phagocytosis in macrophages. Annu Rev. Immunol. 17, 593–623 (1999).
Acquah, F. K., Adjah, J., Williamson, K. C. & Amoah, L. E. Transmission-blocking vaccines: old friends and new prospects. Infect. Immun. 87, e00775–e00818 (2019).
Tomas, A. M. et al. P25 and P28 proteins of the malaria ookinete surface have multiple and partially redundant functions. EMBO J. 20, 3975–3983 (2001).
Kaslow, D. C. Efforts to develop Pfs25 vaccines. Am. J. Trop. Med Hyg. https://doi.org/10.4269/ajtmh.21-1326 (2022).
Lantero, E. et al. Heparin administered to anopheles in membrane feeding assays blocks plasmodium development in the mosquito. Biomolecules https://doi.org/10.3390/biom10081136 (2020).
Rodriguez, A. M. et al. Histamine ingestion by anopheles stephensi alters important vector transmission behaviors and infection success with diverse plasmodium species. Biomolecules https://doi.org/10.3390/biom11050719 (2021).
Simon, N. et al. Malaria parasites co-opt human factor H to prevent complement-mediated lysis in the mosquito midgut. Cell Host Microbe 13, 29–41 (2013).
Luckhart, S., Pakpour, N. & Giulivi, C. Host-pathogen interactions in malaria: cross-kingdom signaling and mitochondrial regulation. Curr. Opin. Immunol. 36, 73–79 (2015).
Pakpour, N., Riehle, M. A. & Luckhart, S. Effects of ingested vertebrate-derived factors on insect immune responses. Curr. Opin. insect Sci. 3, 1–5 (2014).
Luckhart, S. et al. Mammalian transforming growth factor beta1, activated after ingestion by Anopheles stephensi, modulates mosquito immunity. Infect. Immun. 71, 3000–3009 (2003).
Zhu, F. et al. Whole-killed blood-stage vaccine-induced immunity suppresses the development of malaria parasites in mosquitoes. J. Immunol. 198, 300–307 (2017).
Loughland, J. R. et al. Transcriptional profiling and immunophenotyping show sustained activation of blood monocytes in subpatent Plasmodium falciparum infection. Clin. Transl. Immunol. 9, e1144 (2020).
Terkawi, M. A., Nishimura, M., Furuoka, H. & Nishikawa, Y. Depletion of phagocytic cells during nonlethal plasmodium yoelii infection causes severe malaria characterized by acute renal failure in mice. Infect. Immun. 84, 845–855 (2016).
Ngwa, C. J. et al. 16S rRNA gene-based identification of Elizabethkingia meningoseptica (Flavobacteriales: Flavobacteriaceae) as a dominant midgut bacterium of the Asian malaria vector Anopheles stephensi (Dipteria: Culicidae) with antimicrobial activities. J. Med Entomol. 50, 404–414 (2013).
Kämpfer, P. et al. Elizabethkingia anophelis sp. nov., isolated from the midgut of the mosquito Anopheles gambiae. Int. J. Syst. Evolut. Microbiol. 61, 2670–2675 (2011).
Abrahamsen, H. L. et al. Distant relatives of a eukaryotic cell-specific toxin family evolved a complement-like mechanism to kill bacteria. Nat. Commun. 15, 5028 (2024).
Bahia, A. C. et al. Exploring Anopheles gut bacteria for Plasmodium blocking activity. Environ. Microbiol 16, 2980–2994 (2014).
He, B. et al. Host complement C3 promotes malaria transmission by killing symbiotic bacteria in the mosquito midgut. Proc. Natl. Acad. Sci. USA 122, e2424570122 (2025).
Tripathi, A. K., Mlambo G., Kanatani, S., Sinnis, P. & Dimopoulos, G. Plasmodium falciparum gametocyte culture and mosquito infection through artificial membrane feeding. J. Vis. Exp. https://doi.org/10.3791/61426 (2020).
Acknowledgments
This work was supported by the State Key Program of the National Natural Science Foundation of China 82430073 (W.X.), and the National Natural Science Foundation of China 82302570 (L.J.). Additionally, we stated that artificial intelligence was utilized to help us rephrase our manuscript.
Author information
Authors and Affiliations
Contributions
Conceptualization: X.W.; Methodology: H.B., M.L., J.F., Y.F., G.S., and T.N.; Investigation: H.B., M.L., J.F., Y.F., L.J., L.T., Z.F., and Z.J.; Funding acquisition: X.W. and L.J.; Supervision: X.W.; Writing – original draft: X.W. and H.B.; Writing – review & editing: X.W. and H.B.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
He, B., Li, M., Guo, S. et al. Host macrophages/monocytes promote malaria transmission by modulating mosquito microbiota via SR-A-mediated phagocytosis. Nat Commun 17, 4385 (2026). https://doi.org/10.1038/s41467-026-70966-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-70966-3
