
Abstract
Ergosteryl-3β-O-L-aspartate synthase (ErdS) catalyzes tRNA-dependent aspartylation of ergosterol, a lipid essential for fungal cell membrane integrity. However, the functional significance of ergosteryl-aspartate and the molecular mechanisms underlying its synthesis remain unclear. Here, we show that ErdS localization is highly dynamic and that Erg-Asp is required for proper hyphal growth, sporulation, and spore germination, and likely influences stress tolerance. The cryo-electron microscopy structure of ErdS revealed an unprecedented sterol-binding pocket. In addition, the structures in complex with a non-hydrolyzable Asp-N-tRNAAsp show a tRNA-guided intramolecular aminoacyl transfer mechanism between two functional domains of the enzyme. The CCA end of tRNAAsp undergoes a large displacement to reach the aa-tRNA transfer active site, while the tRNA elbow is clamped by a long extension an N-terminal α-helix. The present structural and mutational analyses demonstrate that domain fusion, dynamic repositioning, and tRNA-mediated substrate handover underlie the multifunctional catalytic efficiency of ErdS and participates in Erg-Asp synthesis independently from protein synthesis. These findings elucidate the regulatory mechanism of tRNA-dependent sterol modification and provide insights into fungal membrane dynamics, highlighting potential targets for antifungal therapies.
Similar content being viewed by others
Introduction
Aminoacylated transfer RNAs (aa-tRNAs) play an essential role in delivering amino acids (aa) to translating ribosomes. Recent studies have shown that aa-tRNAs also serve as a source of activated aa in a wide range of other cellular processes unrelated to protein synthesis1,2,3,4,5. In these alternate routes, the tRNA-esterified aa moiety is transferred onto hydroxyl- or amino-groups of diverse acceptor molecules by enzymes called aa-tRNA transferases (ATTs). One structural family of these ATTs shares a common double Gcn5-N-acetyltransferase (dGNAT)-like fold, and participates in a variety of antibiotic resistance and pathogenicity pathways via aminoacylations of peptidoglycan and glycerolipids in bacteria6,7,8,9,10. In higher fungi—including human and plant pathogens—we recently identified two new dGNAT-related ATTs, named ergosteryl-3β-O-(L)-amino acid (Erg-aa) synthases (ErxS), which perform aa-tRNA-dependent conjugation of aa to sterols5,11. The first discovered was the ergosteryl-3β-O-L-aspartate synthase (ErdS), which catalyzes the aspartylation of the 3β-OH group of ergosterol (Erg)11,12,13, yielding ergosteryl-3β-O-L-aspartate (Erg-Asp; Fig. 1A). Concomitantly, we found that an Erg-Asp hydrolase (ErdH) was discovered, which deacylates Erg-Asp back to Erg and aspartate (Asp)1. This latter enzyme is an ergosteryl-3β-O-glycine (Erg-Gly) synthase (ErgS), and is only found in ascomycota.

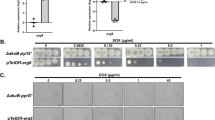
A ErdS is composed of an AspRS domain (blue) and an ATT domain (orange). The first binds tRNAAsp, L-Asp, and ATP to generate Asp-tRNAAsp. The acceptor arm then translocates to the ATT domain, where Asp is transferred onto the 3β-OH of Erg to produce Erg-Asp. ErdH hydrolyzes Erg-Asp to Erg and Asp. B Growth of colonies of the indicated Afm strains on malt agar supplemented with Glc (repressing condition) or Xyl (inducing condition). Pictures present colony morphologies after 48 h. Conidiation (green) areas were measured (fraction of total colony area) in each strain, in addition to colony diameters (see Supplementary Fig. 1C). Data were analyzed using a one-way ANOVA followed by Dunett’s multiple comparison test vs WT (***: p < 0.001, *: p < 0.05, ns: non-significant). C Growth of colonies of the indicated Afm strains on MM supplemented with Glc or Xyl. Colonies were imaged using a magnifier to visualize the fluffy phenotype (arrowheads), which is qualitatively described. D Growth of mycelial lawns of the indicated Afm strains on MM supplemented with Glc or Xyl in 12-well plates, after 20 or 48 h. Lawns were imaged with a magnifier at each time point to visualize the presence or absence of conidiophores. E Left. TLC analyses of total lipids from WT Mor and clones of erdH and erdS mutant strains. Right. TLC showing separation of total lipids from the WT and erdS strains of Afm and the WT and erdH strains of Ncr. In the latter, as in Mor, Erg-Asp is undetectable unless erdH is deleted. Asterisks (*) indicate Erg-Asp bands. F Colony morphologies of WT, erdS, and erdH strains of Mor on plates. Plates were illuminated from below (first row), or from above (second row). The third row presents colonies imaged from below. Bars represent the number of conidia counted per plate after 9 days of culture for Mor and after 5 days for Afm (N = 3 biological replicates). Data WT vs ∆erdS or ∆erdH were analyzed using a Welch’s t-test (***: p < 0.0001, ns: non-significant) for Mor and using a t-test for Afm. G Inoculation type.
ErdS is a bifunctional enzyme with distinctive structural features: the aspartyl-tRNA synthetase (AspRS) domain that activates Asp through adenylation and esterifies it to tRNAAsp, and an appended ATT domain10,14,15,16 that transfers the aspartyl moiety from Asp-tRNAAsp to the 3β-OH group of Erg16,17. As Aspergillus fumigatus (Afm) and Aspergillus oryzae (Aor) express a second AspRS5,11, which has a canonical structure and is dedicated to synthesizing Asp-tRNAs for protein synthesis, deletion of the erdS gene has no effect on growth. The bi-functional reaction catalyzed by ErdS thus suggests a mechanism in which Asp-tRNAAsp, synthesized after aspartylation, would remain bound to the AspRS domain of ErdS with only the tRNA acceptor arm shuttling directly into the ATT active site without release, which is reminiscent of other aa-tRNA utilizing reactions18,19. While this channeling mode may help fungi produce Erg-Asp efficiently and independently of protein synthesis, its molecular details have remained unknown. Importantly, ErdS is phylogenetically related to bacterial aaPGSs5,11, but aminoacylates sterols instead of glycerolipids, implying that the dGNAT module of ErdS contains an uncharacterized and unconventional sterol-recognition fold.
In fungi, Erg plays a crucial role in maintaining the structure and function of cell membranes, thereby directly affecting fungal viability and pathogenicity20,21. Aspartylation of Erg by ErdS alters the physical and chemical properties of the sterol, which might affect membranes and modify the responsiveness of cells to external stress, including antimicrobial stresses16,22,23. However, as Erg-aa species were only recently discovered, their functions in fungi remain elusive. We recently determined that Erg-Asp influences the regulation of asexual sporulation (conidiation) in Aspergillus oryzae (Aor), whereas Erg-Gly seems to promote the production of aerial hyphae24. Both Erg-aa species slightly modify the resistance of Aor towards two antifungal drugs, suggesting a direct or indirect role in modulating membrane properties. Additionally, ErdS is also present in a plethora of pathogenic fungi, such as in Afm, which can cause aspergillosis in immunocompromised individuals as a life-threatening condition associated with poor prognosis25, and in Magnaporthe oryzae (Mor) (rice blast fungus), a cereal pathogen that threatens global agriculture26, but the influence of Erg-Asp on their physiology remains unexplored.
Here, we show that Erg-Asp participates in asexual sporulation in Afm and in Mor, and that the deletion of the erdS gene alters the germination rate of conidia, whereas its deregulation or overexpression increases the production of aerial hyphae and delays conidiation. Both ErdS and the ErdH (the Erg-Asp hydrolase) appear to be expressed at all stages of the Afm life cycle, with dynamic localizations over the course of fungus’s development. Deletion of erdS also affects resistance to high salt stress and resistance to the cell wall-targeting compound Congo Red, suggesting that Erg-Asp might participate in a vast array of processes. In this context, deciphering the molecular underpinnings of tRNA-mediated modifications should be of both therapeutic and agricultural interest27,28. Using cryo-electron microscopy (cryo-EM), we determine the structures of the Afm ErdS apoenzyme and its complex with tRNA. We characterize its sterol binding pocket and highlight a mechanism in which the AspRS long N-terminal α-helix sequesters tRNAAsp, thereby facilitating Asp channeling between the AspRS and ATT active sites, with the RNA moiety acting in a manner mimicking swinging-arm prosthetic cofactors. Such a tRNA capture mechanism might facilitate Erg-Asp synthesis at appropriate locations in a context where ErdS is dynamically localized during fungal development.
Results
ErdS and ErdH are involved in asexual sporulation
To obtain insights into the physiological influence of Erg-Asp, we employed two fungal models: the human opportunistic pathogen Afm and the plant pathogen Mor. We used the previously described ∆erdS mutant1 of Afm, which does not produce Erg-Asp. Despite our best efforts, we could not obtain a ΔerdH mutant of Afm that accumulated Erg-Asp. We circumvented this issue by reintroducing the erdS gene at its locus under the control of the xylose promoter (PXyl) in the ∆erdS strain (∆erdS::PXyl-erdS strain). We also constructed a WT strain in which the PXyl-erdS construct directly replaced the original erdS copy (WT::PXyl-erdS strain). Erg-Asp strongly accumulates in those strains when PXyl-erdS is induced with xylose (Xyl) and is barely synthesized under repressive conditions in the presence of glucose (Glc) (Supplementary Fig. 1A). These results correlated with expression levels of ErdS (Supplementary Fig. 1A).
When Afm conidia (asexual spores) were point-inoculated on plates, they formed round-shaped filamentous colonies with green conidia in the oldest areas, and the maturing and actively growing mycelia formed a thin white circular margin (Supplementary Fig. 1B and S1C). On rich medium (malt agar), colonies of WT and ΔerdS strains showed no obvious morphological differences, whereas both PXyl-erdS strains (∆erdS::PXyl-erdS and WT::PXyl-erdS) presented a strong “fluffy” phenotype—dense aerial hyphae—under either repressing or inducing conditions (Fig. 1B–D), with much larger white margins and smaller conidiation areas. Colonies of all strains presented comparable diameters (Fig. 1B), suggesting that the larger margins reflected a delay in conidiation, rather than growth defects.
Rich media may interfere with repression or induction of the PXyl promoter. We thus point-inoculated conidia on minimal media (MM) supplemented with or without Glc or Xyl (Fig. 1C, Supplementary Fig. 1D). Both PXyl-erdS strains again presented a strong fluffy phenotype under repressing (Glc) conditions, which was reduced—although still visible—under inducing conditions (Xyl), when ErdS was over-expressed. The PXyl-erdS strains also presented far fewer conidiophores than WT or ΔerdS on the colony edges under repressing conditions, as observed by magnification (Fig. 2C, 3rd row, Supplementary Fig. 1D).

A Germination kinetics of WT, ∆erdS, and erdS::PXyl-erdS strains. The graph presents the percentages of swollen conidia (isotropic growth) observed between 3 and 9 h for two biological replicates. B Kaplan-Meier curves showing the distinct germination dynamics between strains in under MOI conditions. Confidence intervals (95%) illustrate the differences in variability and spore behavior across conditions. C Fluorescence microscopy of the Afm strain (N = 3 biological replicates, 10 fields were observed with ≥ 5 spores/field) carrying the erdH-mCherry—erdS-eGFP cluster at four stages of fungal development (illustrated on the right side and Supplementary Fig. 1B). D Congo Red (CR) sensitivity tests on MM + glucose plates supplemented with increasing concentrations of CR. Three strains (indicated on the left) were tested in three replicates (Rep. 1 to 3). Images were taken after 48 h of growth. In the absence of PXyl-erdS induction, the variability observed in complementation across replicates likely reflects the leakage due to the presence of the natural PErd promoter upstream of PXyl in our construct. E Stress sensitivity tests were conducted on the WT and erdS strains under the indicated stress conditions.
We next spread conidia of the four strains on MM (Fig. 1D) and observed the resulting mycelial lawns. The PXyl-erdS strains presented the fluffy phenotype at 20 h under either inducing or repressing conditions, and in contrast to the WT and ΔerdS strains, no obvious conidiation (greenish-colored mycelium) was visible to the naked eye. Magnification confirmed that the WT and ΔerdS strains produced a high density of conidiophores, while the PXyl-erdS strains formed very few (Fig. 1D, rows 1 and 2). Induction of erdS expression (Xyl) did not seem to reverse these phenotypes (Fig. 1D), in contrast to less intertwined and isolated (point-inoculated) colonies (Fig. 1C and Fig. S1D). However, lawns are built up of thousands convergent colonies; we thus interpreted that these mycelial layers likely cumulated residual fluffy hyphae to a level that masked any complementation effect under those growth conditions. After 48 h (Fig. 1D, 3rd and 4th rows), conidiation eventually occurred in PXyl-erdS strains, but the conidiophores were embedded in dense fluffy aerial hyphae (Supplementary Fig. 1E). This confirmed that changing the regulation of Erg-Asp synthesis leads to delayed conidiation in Afm.
In Mor, we not only deleted erdS (∆erdS, no Erg-Asp synthesis) but also erdH (∆erdH, no Erg-Asp hydrolase activity) (Supplementary Fig. 1F). We noted that the WT strain produced undetectable levels of Erg-Asp (Fig. 1E, left panel), as we had also observed in Neurospora crassa (Ncr)1 (Fig. 1E, right panel), another Sordariomycete. As in Ncr, the deletion of erdH resulted in Erg-Asp accumulation, although at low levels (Fig. 1E). Interestingly, the deletion of erdS in Mor triggered a one order of magnitude decrease in conidiation after 9 days (Fig. 1F, left panel, Mor), which paralleled the phenotype observed in Aor for the same mutant24. In contrast, in Afm, conidia production (after 5 days) remains unaffected upon erdS deletion (Fig. 1F, right panel). Colonies of both Mor mutants presented a larger white mycelium margin, although it was less prominent than those observed in Afm PXyl-erdS strains, also reflecting delayed conidiation throughout colony development. Surprisingly, the erdH deletion fully phenocopied that of erdS in Mor (Fig. 1F), suggesting that appropriately regulated levels of Erg-Asp are required for proper conidiation timing and/or levels in Mor. In Afm, conidiation timing and hyphal growth are affected by increased Erg-Asp synthesis, but not at detectable levels in its absence.
Erg-Asp influences germination rate
Beyond hyphal growth and conidiation, another important aspect of Afm physiology is the germination of conidia. We measured the germination kinetics of WT, ΔerdS, and ∆erdS::PXyl-erdS conidia (Fig. 2A) and observed that the initial isotropic swelling proceeded normally in all cases, with normal proportions of swollen spores in the three strains. However, the emergence of germ tubes was slower with the ∆erdS swollen conidia (mean rate 26.0 % h−1) compared to WT conidia (49.5 % h−1). As a result, at 9 h, while most WT conidia had advanced, as expected, to hyphal extension, ~25% of ΔerdS conidia remained non-germinated. For ∆erdS::PXyl-erdS conidia, germination started only after 6 h (3–4 h for WT conidia) post-inoculation, and the germination rate could not be evaluated under these conditions. This suggested that the lack of Erg-Asp synthesis either delays the germination of Afm conidia or alters synchronous germination29,30.
Other types of sterol conjugations are known to impact the immune recognition of fungal spores31. We thus decided to monitor the phagocytosis of FITC-stained dormant or swollen conidia by IC-21 murine peritoneal macrophages (IC-21 MPM), using fluorescence microscopy (Supplementary Fig. 2A). We did not observe significant differences in adhesion to internalization by macrophages between Afm WT, ∆erdS, or ∆erdS::PXyl-erdS conidia, suggesting that Erg-Asp has no major role in the recognition by these immune cells. The same conclusion was drawn from Nanolive microscopy (Supplementary Fig. 2B, Supplementary Movies 1 and 2). However, the Kaplan-Meier curves representing the proportion of non-germinated conidia (Fig. 2B) confirmed that the proportion of non-germinated conidia dropped sharply before 6 h with the WT strain (Multiplicities of infection [MOIs] of 5 and 10 showing no significant difference, p = 0.32, see Supplementary Fig. 2B for statistics), with >80% of germinated spores at 7–8 h post-inoculation (6–7 h after recording started). However, the ΔerdS conidia exhibited slower germination rates at an MOI of 5, and especially at an MOI of 10 (Fig. 2B). In addition, the final proportion of germinated conidia reached ~60% (MOI of 5) or only ~30% (MOI of 10) for ∆erdS conidia. This indicated that either the germination success was lower for the mutant, and/or the MOI had an influence on germination. Higher conidial densities are known to decrease germination rates29, but here the ΔerdS mutation or Erg-Asp synthesis deregulation (∆erdS::PXyl-erdS) might amplify this phenomenon or perturb synchronous germination within the population.
Dynamic localizations of ErdS and ErdH in A. fumigatus
Given that ErdS and ErdH—and thus Erg-Asp levels—appeared to impact conidiation, germination, and hyphal growth, we analyzed their subcellular localizations throughout the fungal life cycle (Fig. 2C). We thus replaced the erdH—erdS bigenic cluster of Afm with a version enabling the expression of ErdS and ErdH that were C-terminally fused to eGFP and mCherry, respectively. Fluorescence microscopy showed that Afm ErdS and ErdH were both expressed at all stages of the fungal life cycle and were already present in resting spores—consistent with the observation of Erg-Asp in total lipids extracted from dormant conidia (Supplementary Fig. 2C).
In resting conidia, ErdS-eGFP is diffuse and localized in the cytosol, while ErdH-mCherry is concentrated in cytosolic round-shaped structures that only partially colocalize with ErdS-eGFP (Fig. 2C, t = 0). In swollen conidia (t = 3 h), a portion of the ErdS-eGFP started to concentrate in puncta-like patches that seem to—at least apparently— colocalize with some of ErdH-mCherry puncta. In germinating conidia (t = 6 h) most of the ErdS-eGFP and ErdH-mCherry signals displayed granular distribution patterns that, on average, did not seem to colocalize. ErdS-eGFP seems evenly distributed and diffuse in the cytosol of growing mycelia, while ErdH was enriched on or within structures that resemble vacuoles (Fig. 2C, t = 16 h); however, fractions of both enzymes also appeared to colocalize at the plasma membrane, where most of the cellular aspartylable Erg resides. This localization was consistent with previous subcellular fractionation experiments on mycelia11. Finally, ErdS-eGFP and ErdH-mCherry were highly enriched at the plasma membranes of remnant germinating structures (Fig. 2C, t = 16 h).
These observations indicate that the localizations of ErdS and ErdH are highly dynamic throughout the fungal life cycle. It is therefore probable that Erg-Asp production temporally and spatially changes throughout the fungal lifecycle, and thus most likely regionally within colonies.
Erg-Asp might contribute to stress tolerance
Since both ErdS and ErdH colocalized at some point in fungal development to the plasm, this prompted us to test whether Erg-Asp could influence the sensitivity of Afm to the cell wall-disrupting agent Congo Red (CR). As shown in Fig. 2D, the ΔerdS strain was more sensitive to CR than the WT. This could indicate that the permeability of either the cell wall or plasma membrane32 is affected in this mutant and that Erg-Asp might contribute to cell wall and/or plasma membrane integrity, at least under stressful conditions. Interestingly, the ∆erdS::PXyl-erdS strain partially recovered CR resistance on MM medium with glucose. Deletion or overexpression of erdS, however, did not affect sensitivity to caspofungin, an echinocandin that targets ß-(1-3)-glucan synthases (Supplementary Fig. 2D, left panel), suggesting that the cell wall is likely not affected by Erg-Asp levels. High salt stress (NaCl and KCl) did not significantly affect growth (Fig. 2E), but the WT strain presented strong conidiation defects (Supplementary Fig. 2D), whereas the ΔerdS strain remained unaffected, suggesting that in the absence of Erg-Asp, osmotic stress tolerance was higher, at least in the context of conidiation. Taken together, these results might advocate for an effect of Erg-Asp on properties of the plasma membrane. However, this will require a more in-depth characterization that goes beyond the scope of this study.
The regulation of Erg-Asp synthesis and its levels under various conditions is currently unknown. The only condition under which we found a significant variation of Erg-Asp levels was when the WT Afm strain, first grown in liquid MM containing a nitrogen (N) source (ammonium), was transferred to a nitrogen-free MM medium for 5 h, which led to a ~ 1.9-fold decrease in Erg-Asp within the total lipids, as judged by TLC (Supplementary Fig. 2E), suggesting that N availability might be a signal for Erg-Asp synthesis. However, we did not detect significant differences in growth rates between WT and ΔerdS strains when grown in the presence of various N sources (Supplementary Fig. 2F) (ammonium, urea, bovine serum albumin, amino acids). When the N source was progressively reduced, however, the ∆erdS and ∆erdS::PXyl-erdS strains showed ~1.70-fold increased growth rates under N deprivation (0 mM ammonium) or limitation (1 mM), compared to N replete (10 mM) conditions, that the WT strain did not exhibit. These differences tended to decrease under inducing conditions (MM+Xyl) in the ∆erdS::PXyl-erdS strain, suggesting the influence of Erg-Asp in this phenotype (Supplementary Fig. 2G).
Given the diversity of effects induced by the absence and over-expression of ErdS in fungi, it seems that the temporally and spatially controlled synthesis of Erg-Asp is required for normal fungal development and propagation. This raised questions regarding how ErdS recognizes Erg and ensures proper Erg-Asp synthesis for normal germination, growth, and conidiation, as well as when the fungus copes with stress. We thus performed a structural characterization of ErdS to better understand Erg-Asp synthesis in Dikarya.
Structure and architecture of Afm ErdS: the aa-tRNA binding site
The closest homologs of ErdS in the dGNATα(+) ATT family are the bacterial aaPGSs that aminoacylate glycerolipids, which suggests that their lipid-binding pockets, which partly reside within the GNAT I subdomain, should differ structurally. No known sterol-binding domain was detected in ErdS, and, to our knowledge, no GNAT-related domain characterized thus far has been reported to bind sterols33. Similar to the other ATTs of the same structural fold, the GNAT II domain is expected to recognize the aa-tRNA moiety. Further, the mechanism by which the tRNAAsp acceptor stem travels between the AspRS and ATT active sites remains unknown. To identify the aa-tRNA and sterol binding pockets of ErdS, we expressed and purified an Afm ErdS mutant lacking the N-terminal 86 residues (ErdSΔN1-86) for more efficient expression, mixed it with an in vitro-transcribed Afm tRNAAsp and/or ergosterol, and performed single particle cryo-EM analyses.
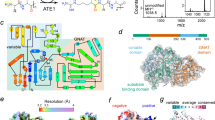
The first cryo-EM density map obtained showed that ErdSΔN1-86 bound neither tRNAAsp nor Erg, and that the apo-protein formed a homotetramer (Supplementary Fig. 3). This tetramer is composed of two ErdS dimers (Fig. 3A) formed by the interactions between two AspRS domains in a manner similar to that observed in the crystal structure of yeast AspRS15. We additionally determined an apo cryo-EM structure of full-length ErdS (Supplementary Fig. 4A–D). Superposition of the full-length ErdS dimer with the dimeric unit within the ErdSΔN1–86 tetramer, as well as with the Δ85 construct used in this study, revealed no appreciable differences in overall architecture (Supplementary Fig. 4E), indicating that N-terminal truncation does not measurably affect the dimer/tetramer assembly. Despite several attempts to obtain the structure of ErdS with bound Erg, we were unable to captured the sterol in the ATT domain, possibly due to its low solubility. No information on aa-tRNA or Erg binding could thus be obtained at this stage. Nevertheless, the structures of the two ATT domains found in the ErdS dimer could be resolved (Figs. 3A and 3B). Each of them had the dGNATα(+) architecture consisting of two GNAT folds linked by a positively-charged α helix (noted α(+)) (Fig. 3B, purple), similar to the architectures of the ATT domain of LysPGS10 and the bacterial Fem-ligases (FemX) (Supplementary Fig. 5A-D) involved in the tRNA-dependent aminoacylation of a peptidoglycan precursor8, with Cα RMSDs for the GNAT II core of 1.23 Å over 79 aligned residues (LysPGS) and 1.27 Å over 43 aligned residues (FemX).

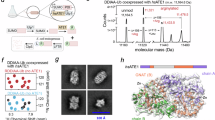
A. Structure of the ErdS dimer. For comparison, the architecture of ErdS is represented with the position of each domain numbered according to that for Afm. AspRSs are colored blue (AspRS 1) and light blue (AspRS 2), and ATTs are orange (ATT 1) and light orange (ATT 2). NTD: N-terminal domain; CD: catalytic domain of AspRS; ABD: anticodon-binding domain; α(+): positively charged alpha helix. B Structure of the ATT domain: GNAT I subdomain is dark gray, GNAT II is orange, and the α(+) helix is purple. C Structural comparison between the ATT domain of ErdS (orange) and the GNATⅡ domain of FemX (white) with its substrates (light gray) (PDB: 4II9). The tRNA CCA analog was superimposed. D Mutational analysis of ErdS. ErdS activity was determined (bars) for each mutant (indicated above) in total lipids of Sce separated by TLC (second row), by calculating the ratio of the Erg-Asp signal measured in the corresponding migration area and the signal of the phosphatidylethanolamine (PE) that migrates slightly above Erg-Asp (Erg-Asp/PE ratio (R)). In the absence of Erg-Asp, phosphatidylglycerol becomes visible, which results in an R ratio not equal to zero. The number of biological replicates (N) is indicated and data were analyzed using a t-test (***: p < 0.001, **: p < 0.01, *: p < 0.05, ns: non-significant). Immunoblots show the expression of ErdS using anti-ATT antibodies and the loading control (stain-free). Residues corresponding to those mutated in Afm are shown for the Aor, Ncr, and Mor ErdSs. E Superposition of the two starting ergosterol (Erg) poses used for MD simulations: SwissDock-derived docking-start pose (blue) and AlphaFold3-start pose (purple). F Organization of the Erg-binding domain based on the AlphaFold3 pose. G TLC showing products formed with in vitro sterol aminoacylation assays. Sterols used are indicated. The control TLC shows the Cho, Pregnenolone (Preg.), and DHA bands from stock sterols, and illustrates that no visible Cho seems to contaminate Preg. or DHA. Ctrl: no sterols. Histograms represent the mean value of steryl-[14C]Asp formed relative to Erg-[14C]Asp after 40 min of incubation with Afm ErdS (N = 2 replicates). H Structures of the sterols used in (G).
In the ErdS dimer, each ATT features a deep, large pocket in the GNAT II domain, next to the α(+) helix, with a surface predominantly covered by positively-charged or neutral residues (Supplementary Fig. 5E), which likely corresponds to the Asp-tRNA-binding site. As expected, superimposition of the structure of the ErdS ATT domain onto that of FemX in complex with a tRNA CCA-end analog9 (Supplementary Fig. 5C) revealed that the CCA-end analog fit well at the entrance of this cavity (Fig. 3C). In this superposition, several ErdS residues are positioned around the modeled CCA end, highlighting candidate contacts that could stabilize the CCA backbone at the cavity entrance. W785 appears in the proximity of the A76 and C75 phosphate groups, with which it could interact via anion-π interactions. R789 is positioned near the ribose of A76, and thus could interact via hydrogen bond interactions (Fig. 3C). Interestingly, the electrostatic potential of aa-tRNA–dependent lipid-modifying pockets has been reported to track the aminoacyl moiety to be transferred: the pocket of LysPGS, which transfers positively charged Lys, is overall acidic, whereas that of AlaPGS, which transfers neutral Ala, is comparatively neutral. In ErdS, this suggests that the overall positive/neutral character of the tunnel can favor accommodation of the polar Asp–CCA moiety, while ergosterol is concurrently accommodated in a hydrophobic, mildly cationic subpocket within the same cavity, thereby reconciling the physicochemical requirements of both substrates.
To study the contributions of several residues of ErdS to the Erg-Asp synthesis activity, we introduced point mutations into full-length ErdS and expressed them in the yeast Saccharomyces cerevisiae (Sce) as we showed before11. Notably, Sce possesses no ErxS homolog11. Then, Erg-Asp synthesis was assessed by thin-layer chromatography (TLC) after lipid extraction. To facilitate comparisons, the Erg-Asp levels were normalized to those of phosphatidylethanolamine (Erg-Asp/PE ratio, denoted as an R) (Fig. 3D). Of note, without Erg-Asp, phosphatidylglycerol becomes visible, which results in an R ratio above zero. First, we investigated the conserved GNAT II residues in the putative CCA pocket (Fig. 3C) and the Lys/Arg residues located in the α(+) helix, which were expected to further bind the tRNA moiety. R749, K750, R752 and R753 were mutated to Ala (ErdSα(0) mutant) or Glu (ErdSα(-)) (see Fig. 3B). As expected, ErdSα(-) and ErdSα(0) completely lost the Erg-Asp synthesis activity, with R values (0.12 ± 0.05 and 0.17 ± 0.06, respectively) comparable to that measured for an ErdS ∆ATT mutant lacking the entire ATT domain (0.17 ± 0.06), and well below that of WT ErdS (R = 1.42 ± 0.27) (Fig. 3D). We previously observed identical effects when the α(+) helix of the ergosteryl-glycine synthase (ErgS)5 was mutated. Single mutations in residues shaping the CCA pocket (W785H, W785A, R789A, K790E, G791A, Q793R, H795A, F913A and K916E) dramatically affected the ErdS activity, with all R < 0.5 for all. These results are consistent with their predicted involvement in CCA and tRNA binding (Fig. 3B, C). Interestingly, the K790E variant (R = 0.40 ± 0.08) is naturally present in Mor (Fig. 3D), which might explain the low ErdS activity in this fungus when erdH is deleted (no Erg-Asp hydrolase) (Fig. 1E).
ErdS GNAT I domain harbors a new sterol-binding fold
We next evaluated whether Erg can be stably accommodated within the putative sterol-binding cavity located at the GNAT I–GNAT II interface. To this end, we performed molecular dynamics (MD) simulations using two alternative starting poses: (i) a docking model positioned near residues highlighted by our mutagenesis results, and (ii) an AlphaFold3-predicted Erg-bound pose. For each starting model, we carried out four independent simulations (Fig. 3E and Supplementary Fig. 6). Across these trajectories, the AlphaFold3-starting pose exhibited greater stability, supporting the idea that Erg binds in a location close to this predicted site (Supplementary Fig. 5F).
Erg fits well in this cavity, with a conformation consistent with favorable steric and electrostatic interactions to orient the 3β-OH group towards the terminal A76 of tRNA, to which Asp is esterified. A structural comparison with the PG-bound form of the homolog LysPGS10 revealed, as expected, that the overall shape of the pocket differs substantially (Supplementary Fig. 5B), indicating that this predicted sterol-binding pocket (Fig. 3E, F) is indeed distinct in the dGNATα(+) ATT family and constitutes a non-conventional sterol-binding fold. Based on a multiple alignment of 98 ErdS orthologs, we selected 16 potentially critical residues within this cavity (Y637, T641, S642, T643, S644, W645, D647, R649, N677, E712, C728, E731, E798, W802, Q836 and K838) among which 13 are strictly or moderately conserved (Supplementary Fig. 7). The Q836R mutation completely abolished the ErdS activity (R = 0.14 ± 0.05), while other mutations significantly reduced the Erg-Asp synthesis activity. The S644A mutation had the strongest effect (R < 0.5), while the W645A, D647R and K838E mutations had milder consequences (0.5 < R < 1.0) (Fig. 3D). Notably, these residues cluster at the GNAT I–GNAT II boundary, suggesting that they contribute not only to direct sterol accommodation but also to maintaining the relative positioning of the two subdomains required to shape the pocket properly. Mutations of the remaining 11 residues had only slight or statistically nonsignificant effects. Since the ErdS variants were all expressed at comparable levels in Sce strains (Fig. 3D, immunoblot), we attributed the decreased ErdS activities to the introduced mutations (with the exception of F913A, which was expressed at low levels). We ruled out the possible long-range effects of these mutations on the AspRS activity, which would also reduce Erg-Asp synthesis, even if Asp-tRNA can be scavenged from the cellular pool11. This is because all these ErdS variants could replace the endogenous Sce AspRS (Dps1) by complementation of a dps1∆ strain34 (Supplementary Table 2). These results are in good agreement with our predicted location of the sterol-binding domain of ErdS.
Sterol aspartylation by ErdS is highly promiscuous
We previously observed that Afm ErdS aspartylates cholesterol (Cho) in addition to Erg, despite the structural differences between the two sterols11. Consequently, Cho fits well within ErdS’s sterol pocket in docking models (Supplementary Fig. 8A). Diosgenin, which features an unusual cyclized alkylyl side-chain (Supplementary Fig. 8A), also seems to bind productively, suggesting that ErdS might interact with a broad range of sterols. To investigate both the ErdS promiscuity towards sterols and the structural determinants for sterol recognition, we performed an in vitro sterol aminoacylation assay11 with a panel of structurally different sterols (Figs. 3G and 3H, Supplementary Fig. 8B and 8C).
Afm ErdS aspartylates sterols regardless of the alkylyl side-chain’s structure, since in addition to Erg and Cho (Fig. 3G, left panel, lanes 1, 2 and 4), it aspartylates the human hormones pregnenolone and trans-dehydroandrosterone (DHA), although they present much smaller polar side-chains (Fig. 3G, left panel, lanes 6 and 7). The lower aspartylation levels suggest that the alkylyl moiety could enhance the proper anchoring within the sterol pocket (Fig. 3G). Consequently, a variety of other sterols can be used as substrates, despite the fact that they differ in the numbers (0, 1, or 2) and positions of saturations in the B-ring or in the bulkiness, saturation, and methylation pattern of the alkylyl side chain (Supplementary Fig. 8B and S8C, lathosterol, zymosterol, β-sistosterol, stigmasterol, coprostanol, fucosterol, 5α-cholestan-3β-ol). As predicted above (Supplementary Fig. 8A), diosgenin is efficiently aspartylated by ErdS, indicating that a cyclized side chain does not prevent recognition and aminoacylation. Lanosterol, which features a 4-dimethylated position in the A-ring of the cyclopentanophenantrene (sterane) nucleus, however, was not aspartylated in vitro (Fig. 3G, left panel, lane 3, Supplementary Fig. 8B, lane 3), indicating that such bulky modifications may act as antideterminants for sterol recognition. Similarly, β-estradiol, with an aromatic A-ring, was not a substrate (Fig. 3G, left panel, lane 8), despite sharing common features with DHA, showing that a non-aromatic ring is required.
Interestingly, coprostanol and epicoprostanol are identical, except that the first possesses a 3-OH group in the β conformation, whereas the second is in the α conformation. These two steroids and 5α-cholestan-3β-ol have the same alkylyl side chain as Cho (but no unsaturation at position 5 in the steran nucleus). As expected, coprostanol and 5α-cholestan-3β-ol were aspartylated at similar levels as Cho, despite the absence of the Δ5 unsaturation and their different absolute conformation at position C5 (Fig. 3G, right panel). In contrast, epicoprostanol was not aminoacylated, most likely because the 3α-OH group is oriented away from the catalytic residues (see below) and the aminoacylated A76 residue of tRNAAsp. Consequently, cholic acid, which has a 3α-OH group, was also not a substrate (Fig. 3G, left panel, lane 5).
This promiscuous activity also existed in vivo, as demonstrated by the heterologous expression of Afm ErdS in Sce Erg biosynthesis pathway mutants35 (erg3∆, erg4∆,erg5∆, and erg6∆), which accumulate various Erg intermediates (Supplementary Fig. 8D and S8E). TLC analyses of total lipids from these strains revealed that ErdS aspartylates Erg intermediates that mostly differ in the saturation of the B-ring and the alkylyl side-chain, as expected from our in vitro results; notably, the erg6Δ strain accumulated zymosterol, which was also aminoacylated by ErdS in vitro (Supplementary Fig. 8B, lane 5). Therefore, ErdS is highly promiscuous for sterols, with the minimal requirements that the A-ring is non-aromatic and free of methyl groups and the 3-OH is in the β conformation.
Organization of the active site and catalytic mechanism of transesterification
Simultaneous docking of Erg and the CCA analog revealed that the 3-OH group of Erg, when in the β conformation, is oriented towards the 2´-OH of the ribose moiety of A76, in a geometry suitable for transesterification (Fig. 3F and Supplementary Fig. 8FS8G). The α conFigureuration evidently misoriented the 3-OH group of Erg (or any sterol) within the active site, preventing it from reacting with the Asp-tRNAAsp ester bond. In FemX in complex with a CCA analog and a peptidoglycan precursor, the nucleophile was provided by the ε-amino group of a Lys side chain from the peptidoglycan precursor before transpeptidation. By contrast, Erg must first be activated by ErdS. We propose (Supplementary Fig. 8F and S8G) that H795 acts as a general base to deprotonate the 3-OH group of Erg, thereby generating a nucleophilic alkoxide. This activated 3-alkoxide is positioned to attack the carbonyl carbon of the aminoacyl ester in Asp-tRNAAsp. During this reaction, the negative charge developing on the carbonyl oxygen is likely stabilized by the positively-charged guanidinium group of R914, facilitating the transesterification (Supplementary Fig. 8G). The K305 residue of FemX has an equivalent function and is located at the same position in superimposed models. This mechanism provides hints toward elucidating how the active site architecture of ErdS supports the selective transfer of the aspartyl group to only 3β-OH-containing sterol substrates. The presence of methyl groups at C4 of the sterol A-ring likely introduces steric hindrance, preventing proper accommodation within the binding pocket.
The N-terminal long α-helix of ErdS clamps the tRNA elbow to ensure enzymatic activity
The N-terminal extension in the Class IIb aaRSs of higher eukaryotes contains a conserved alpha-helix, which has been suggested to play a crucial role in enhancing their affinity for tRNA36,37,38,39. Although the crystal structure of AspRS has been determined, this N-terminal region was deleted in the resolved structure, and thus requires further investigation to elucidate its role in tRNA binding15,40. Notably, ErdS also possesses a similar N-terminal helix that includes the conserved xSKxxLKKxx motif (with the exception of Mor), which was previously proposed to be critical for tRNA binding38,39(Fig. 4A). The recently reported cryo-EM structure of human LysRS41 provides insights into the partial structure of this helix and its interactions with tRNA. However, the density corresponding to the anticodon-binding region remains unresolved, and the complete structural conFigureuration of the enzyme awaits elucidation. To investigate the role of the N-terminal helix, we prepared full length ErdS (FL-ErdS), incubated it with in vitro-transcribed Afm tRNAAsp, and performed a cryo-EM analysis. The structure revealed the dimeric conFigureuration of ErdS in complex with two tRNAAsp molecules (Fig. 4B; Figures S10and -S11).

A Alignment of the NTDs from the Afm, Aor, Ncr, and Mor ErdSs. Positively charged residues are blue, residues that interact with the anticodon stem are red, and the region involved in the interaction with the elbow region is highlighted in yellow. B Structure of ErdS with the NTD and tRNAAsp. C,D Close-up view of the contacts between the ErdS N-terminal region and tRNAAsp. E Electrostatic potential of the NTD (blue – basic; red – acidic; white - neutral). F Complementation of the Sce YAL3 dps1 strain with the indicated proteins and ErdS variants. Pictures represent Sce colonies on plates after 3 days of growth. G Complementation of the Sce YAL3 dps1 strain with the indicated Aor ErdS variants. Pictures represent Sce colonies on plates after 3 days of growth. Two clones of the Aor ErdS ΔN90 were used. Arrows indicate positions of several extremely slow-growing colonies.F,Glower panels: TLC profiling of total lipids from these strains to visualize Erg-Asp production (visualization under UV light). Asterisks (*) indicate Erg-Asp bands. Histograms represent the Erg-Asp/Phosphatidylethanolamine (PE) ratio of N = 2 biological replicates.
In the complex, each protomer of the ErdS dimer binds one tRNAAsp molecule (Fig. 4B–D). The anticodon of tRNAAsp interacts with the anticodon-binding domain (ABD) of the AspRS domain, and the acceptor stem is positioned within the catalytic domain of AspRS, consistent with the previously reported crystal structure of the Sce AspRS/tRNAAsp complex15. The ATT domain was disordered, suggesting that it moves flexibly relative to AspRS. Interestingly, residues 59–102 of the NTD form an extended linear pair of α-helices separated by a connective loop, which spans from the anticodon to the elbow region of tRNAAsp (Fig. 4B–E). This N-terminal region is rich in polar amino acids, which form multiple interactions with the phosphate backbone, ribose moieties, and nucleotide bases of tRNAAsp. Residues R61 and N62 interact with the phosphate backbone of residues G50 to G52 in the T-stem. The highly conserved K65 interacts with the ribose of G57 in the T-loop. C20a of tRNAAsp is flipped outward, with its base forming a salt bridge with R69 of the NTD. Lastly, E70 and S77 interact with the phosphate backbone of tRNAAsp’s variable loop. Further stabilization is provided by E81, Q84, and D87 within the connective loop between the α-helices, which, along with H92, which interact with either the ribose or the phosphate backbone of the anticodon stem.
We previously reported that full-length Afm ErdS can complement an Sce dps1Δ strain (AspRS gene deletion), although less efficiently than an Afm ErdS ΔATT variant. Notably, the ATT domain was removed from the variant, likely because Sce tRNAAsp was used primarily for Erg-Asp synthesis over protein synthesis in this context as no extra AspRS was present11. Based on our cryo-EM structure, we hypothesized that the NTD could also contribute to tRNAAsp sequestering within ErdS, so as to ensure its dedicated participation in Erg-Asp synthesis. To ascertain the contribution of the observed N-terminal domain in tRNAAsp sequestering, we designed mutants with progressive NTD truncations (ErdSΔN30, ΔN60, and ΔN84, in which the first 30, 60 or 84 residues were removed, respectively), allowing us to tested their complementation efficiency in the dps1Δ strain by plasmid shuffling. First, we confirmed that the ErdS ΔATT mutant complemented the dps1Δ strain as efficiently as Dps1 (SceAspRS). Strikingly, the growth progressively improved with the NTD truncations of Afm ErdS, with the ΔN84 mutant behaving like the ErdS ΔATT mutant, as judged by the Sce colony sizes (Fig. 4F) and by drop tests (Supplementary Fig. 9). This indicates that the region between residues 60 and 84, where the xSKxxLKKxx motif lies, has the strongest contribution to growth reduction, and is likely to tRNAAsp sequestering. The growth improvement was interpreted as the easier release of Asp-tRNAAsp from ErdS, in agreement with structural results.
In parallel, we visualized Erg-Asp production in those strains and observed that Erg-Asp synthesis dropped by 17.8-, 15.0-, and 12.1-fold, respectively, for the ErdSΔN30, ΔN60, and ΔN84 mutants, indicating that the efficiency of Erg-Asp synthesis is affected by even the smallest truncation (Fig. 4F). These results indicate that the NTD is crucial for the full activity. In the case of Aor ErdS, the complementation of the dps1Δ strain was highly inefficient (Fig. 4G), but the truncation of the NTD segment down to residue 90 (equivalent to Δ84 N in Afm ErdS) restored WT growth. Again, Erg-Asp synthesis was reduced (2.9-fold) in this mutant, compared to the full-length enzyme. These results suggest that, apart from the ATT domain, the NTD significantly contributes to tRNAAsp utilization in the in vivo context, likely through sequestering it within ErdS, making it unavailable for protein synthesis and thereby reducing growth, while strongly increasing the efficiency of Erg-Asp synthesis. This explains why all ErdS-containing fungi have a second, separated AspRS5,11 that ensures a sufficient supply of Asp-tRNAAsp for protein synthesis.
tRNAAsp behaves as a prosthetic swinging arm during the multi-enzymatic turnovers of ErdS
The cryo-EM structural analysis of the ErdS/tRNAAsp complex revealed 3D classes with density suggesting the ATT domain. Further analysis using a mask covering these weak densities improved the map, confirming that they extended from the C-termini of both AspRS domains, which validated them as the ATT domain (Fig. 5A, B and Fig. S10C). In one ErdS protomer (ErdS1), the ATT1 module is positioned adjacent to the elbow of the tRNA1Asp bound to the corresponding AspRS1 domain, on the same side where the NTD1 also interacts along the anticodon stem and the elbow region. Interestingly, tRNA1Asp is not only supported at the elbow region by the ATT1 domain and the NTD1, but its CCA end is located near the ATT2 module from the other protomer (ErdS2), effectively sandwiching tRNA1Asp (Fig. 5A). This explains why the dps1Δ strain expressing WT Afm ErdS grew more slowly than the strains expressing Dps1 (Sce AspRS) or the Afm ErdS ΔATT mutant (Fig. 4F). ErdS predominantly directs Asp-tRNAAsp towards Erg-Asp synthesis, resulting in slower growth in the absence of an independent AspRS, because both the NTD and ATTs prevent its aminoacylated tRNAAsp from being released. In light of this structural information, we propose that the ATT domains of ErdS dimers cooperate not only in Erg-Asp synthesis but also, together with the NTD, in sequestering tRNAAsp.

A Cryo-EM density map of the ErdS/tRNA tetramer with ATT domains. B Cryo-EM density map of ErdS/tRNA on the tRNA acceptor stem in the moved state. The other protomer in the ErdS/tRNA model is superimposed based on the AspRS position. C Superimposition of the tRNA structures when the acceptor arm is ATT-bound (purple) or AspRS-bound (gray). D Scheme of tRNA acceptor stem movement. The acceptor stem is translocated from AspRS to ATT in trans. E Structure of the ErdS/Asp-N-tRNA on the tRNA acceptor stem in the moved state. ATT2 is superimposed based on the AspRS position.
To further investigate the interaction between the ATT domain and the CCA end of tRNAAsp, we attempted cryo-EM structural analysis of ErdS bound to Asp-tRNA. We reasoned that the ATT domain requires the Asp moiety to bind tRNAAsp over the AspRS domain, which, in contrast, recognizes uncharged tRNAAsp. Since aa-tRNAs are unstable, we prepared a non-hydrolyzable Asp-tRNAAsp analog (Asp-N-tRNAAsp) by using a modified tRNAAsp transcript in which the 3´-hydroxy group of the terminal ribose is replaced by a 3´-amino group, so that the ester bond between Asp and tRNAAsp is replaced by an amide bond upon Flexizyme-mediated in vitro aspartylation42,43,44. We also prepared an ErdS variant with nine mutations in the AspRS domain (Q334E, S335 A, N362P, S363A, N364P, H366A, R367A, H368A, and Y494F) to reduce its tRNA binding affinity and allow the Asp-CCA end to enter the ATT catalytic site. We then incubated Asp-N-tRNAAsp with this purified ErdS variant and performed a cryo-EM structural analysis. This revealed the structure of an ErdS dimer bound with only one Asp-N-tRNAAsp (Fig. 5B, Supplementary Fig. 12). This Asp-N-tRNA1Asp was bound to the ABD of AspRS1 in a manner identical to that in the WT ErdS/tRNAAsp complex (Fig. 4B and Fig. 5A). Surprisingly, in this case, the CCA end protruded away from the AspRS1 active site (Figs. 5B to 5E, Fig. S13B–E). This structural rearrangement was accompanied by a 23 Å movement of the tRNAAsp CCA end and an 8 Å shift of the tRNA elbow (Fig. 5C). The ATT1 domain bound with Asp-N-tRNAAsp (Fig. 5B) was positioned similarly to that observed in the previous ErdS/tRNAAsp complex (Fig. 5A). It also interacted with the elbow of tRNAAsp, whereas the ATT2 domain from the tRNAAsp-free protomer (ATT2) showed no detectable density.
Surprisingly, these orientations of the acceptor stem and the CCA end of Asp-N-tRNA1Asp did not direct them towards the active site (CCA pocket) of ATT1, raising the question of the mechanism by which the Asp moiety could be used to modify Erg. We aligned the tRNAAsp-free AspRS1 domain with the AspRS domain of the ErdS/tRNAAsp complex characterized previously, in order to determine the positions of the missing tRNA2Asp and the ATT2 module. These models revealed that the CCA end of Asp-N-tRNA1Asp was even closer to the ATT2 domain (Fig. 5D). Notably, the α(+) helix of ATT2, which was predicted to interact with the tRNAAsp acceptor arm, was the closest to the CCA end (Fig. 5E). The W785 and K916 residues, which are crucial for the CCA interaction and the activity (Figs. 3A to 3D), were also close proximity to the CCA end. These results suggest that, in the ErdS dimer, after aspartylation by the AspRS1 domain, the CCA end of tRNA1Asp swings to the ATT2 domain of the other protomer, allowing the efficient aminoacylation of Erg (Fig. 6). Consistent with this trans-docking model, classification of the 2:2 ErdS–tRNA dataset revealed asymmetric tRNA conformations in which the CCA end remains associated with AspRS1 while the elbow region is displaced toward the ATT side (Supplementary Fig. 10D), supporting a model in which the second tRNA can bias the positioning of the partner ATT module to promote productive CCA docking in trans (Supplementary Fig. 14). Since tRNAAsp appeared to be trapped by its interactions with the NTD and both ATT domains, it is likely that it does not dissociate from ErdS, and that the acceptor arm would shuttle back and forth from one active site (AspRS1) to the next (ATT2), serving as a prosthetic swinging arm that shuttles an activated Asp moiety for Erg-Asp synthesis. This fully explains the low capacity of ErdS to replace the endogenous AspRS in Sce, unless either the ATT or NTD domain is removed to facilitate Asp-tRNAAsp release from ErdS to promote protein synthesis.

Screen captures from Supplementary Movie 3, showing ATT2 (light orange) and tRNA acceptor stem movements throughout the catalytic cycle of ErdS. (1) Description of the ErdS/tRNAAsp dimer complex with the tRNA1Asp (dark gray) acceptor arm in the AspRS1 (blue) active site. The anticodon of tRNA1Asp is bound to ABD1 and further stabilized by NTD1 (red) and ATT1 (orange). In (2) and (3) ATT2, initially bound to tRNA2Asp, rotates and approaches tRNA1Asp, with helix a(+) positioned on the acceptor arm. Then (4, 5), the acceptor arm of tRNA1Asp moves from the AspRS1 active site to ATT2 and is likely constrained by the α(+) helix. NTD1 retains tRNA1Asp and limits the dissociation of Asp-tRNA1Asp. Finally (6), the CCA extends into the GNAT II active site, where Asp is transferred to the 3-OH position of Erg. ATT2 therefore acts in trans. Finally, the acceptor stem of tRNA1Asp can shift back into the AspRS active site to be used for another round of aspartylation, and ATT2 can fold back to its initial position. Residues W785, R789, and K916, which are important for the ErdS activity, are indicated.
Discussion
Our study reveals a distinct mechanism by which an aa-tRNA is employed for small-molecule aminoacylation in eukaryotes. Through structural and biochemical analyses, we show how ErdS catalyzes the transfer of aspartate from Asp-tRNAAsp to the 3β-OH group of Erg—a lipid central to fungal membrane physiology. This transfer reaction is enabled by a previously uncharacterized sterol-binding pocket within the ATT domain and a tRNA-mediated handover mechanism that coordinates two spatially separate active sites. These findings provide new insights into how enzymes can co-opt aa-tRNAs to expand their catalytic repertoire beyond and independently of translation.
ErdS does not rely on the diffusion of intermediates. Instead, it uses the tRNA as a dynamic tether to direct the flow of the aminoacyl (aspartyl) group. First, the RNA moiety of the ErdS/tRNAAsp complex is sequestered within the ribonucleic particle, not only through interactions with the two ATT domains from the two ErdS protomers, but also by a long NTD that further interacts with the anticodon and elbow regions. This reinforces the interaction of the molecule with the AspRS part, thus stabilizing the tRNA through extensive interactions and guiding its movement across the two active site of the enzyme, which likely prevents its release upon aspartylation, as suggested by complementation experiments in Sce. The acceptor arm and CCA end of tRNAAsp can thus undergo a large-scale repositioning ( ~ 23 Å) to reach the ATT domain of the second protomer, while the domain itself is flexibly rearranged to facilitate this interaction. Unexpectedly, in the ErdS protomer, the tRNA acceptor arm does not simply shuttle from the AspRS active site to its appended (cis) ATT domain, as we initially thought (Fig. 1A). Instead, the cis ATT functions as a stabilizer of the ErdS/tRNAAsp interaction, whereas the second ATT domain, situated in the second ErdS protomer, captures the CCA-end of tRNAAsp through interactions with the α(+) helix, and functions as a trans-acting enzyme, using the aspartyl moiety to transfer it onto Erg. In this context, the tRNA functions as an architectural bridge—dynamically connecting the AspRS and ATT domains—in a manner reminiscent of the swinging arms seen in multienzyme complexes45 and of the shuttling between aminoacylation and amino acid editing functions observed in aminoacyl-tRNA synthetases46,47
Schematics of this molecular choreography are depicted in Fig. 6 and Supplementary Movie 3.
The domain architecture of ErdS, which fuses AspRS with a dGNAT-type ATT module, is essential for this coordinated reaction. Our structural and mutational analyses suggest that the extended NTD, which differs from those found in canonical cytoplasmic AspRSs, plays a critical role in clamping the tRNA to stably position it for productive handover. Shortening of the NTD reduces the Erg-Asp synthesis efficiency in vivo in the yeast heterologous model, and we previously observed that separating the AspRS and ATT domains also reduces the activity, which is exemplified when the ATT domain was expressed alone in Aor11, illustrating the catalytic advantages of a single bifunctional enzyme. This integration of synthetase and transferase activities into a single polypeptide represents a functional innovation, enabling direct control over substrate transfer and reactivity. Our complementation experiments in Sce illustrated that ErdS poorly releases Asp-tRNAAsp, which is thus efficiently trapped throughout the Erg-Asp synthesis cycle, leading to decreased protein synthesis. Hence, in nature, Dikarya harboring ErdS always possess a second, canonical AspRS to ensure efficient protein synthesis11. Sequestering of tRNAAsp within ErdS also provides an advantage by diverting a subset of tRNAAsp from the cytosolic pool for protein synthesis and dedicating it solely to Erg-Asp synthesis. This differs from free-standing ATTs, such as the Erg-Gly synthase also present in fungi. In the latter, an independent aa-tRNA synthetase is required for protein synthesis to produce their aa-tRNA substrate that would be used both for protein synthesis and for sterol aminoacylation. The fusion between the AspRS and ATT domains in ErdS and the molecular mechanism described here may ensure proper Erg-Asp levels under challenging conditions and at dedicated subcellular localizations, to enable proper mycelium development, conidiation, and likely plasma membrane properties in a manner independent of protein synthesis.
The deletion of erdS in Afm, Aor, or Mor leads to contrasting phenotypes. Conidiation is strongly reduced in ΔerdS mutants of Aor24 or Mor (this study), but not in Afm. This may reflect differences in regulatory circuits between species or in the direct or indirect contribution of Erg-Asp to such processes. Of note, in the ΔerdS strain of Afm, the ergS gene that encodes the Erg-Gly synthase (ErgS) is still present, and we cannot exclude the possibility that Erg-Gly has overlapping functions that could attenuate phenotypes. Conversely, Mor lacks ErgS.
In Afm, over-expression of ErdS—and thus of Erg-Asp—has much stronger effects than deletion of the erdS gene, especially on aerial hyphae production. Non-physiological and/or deregulated accumulation of Erg-Asp is thus more detrimental to Afm’s physiology than its absence. The Erg-Asp hydrolase ErdH is conserved at the erdS locus in the majority of Ascomycetes, including in Afm and Mor, suggesting that Erg-Asp homeostasis is important for these species. In support of this conjecture, we could never obtain ∆erdH mutants in Afm or Aor, despite extensive efforts. The ErdS/ErdH pair is thus likely central to Erg-Asp homeostasis. In Aor, it affects regulatory circuits that control sporulation and sclerotia (sexual structure) production 24.
Of note, the deletion of the Erg-Asp hydrolase gene erdH phenocopies the ΔerdS mutation in Mor, indeed suggesting that the deregulation of Erg-Asp levels plays significant roles in phenotypes. In this case, the lower activity of ErdS in Mor—i.e., low Erg-Asp accumulation, see Fig. 1E—is most likely much less toxic in a ∆erdH context than in Afm and Aor. A similar behavior was observed in Ncr before11.
Nevertheless, the deregulation or impaired synthesis of Erg-Asp always affects conidiation to various degrees, from decreased conidia production to differences in conidiation timing. In Afm it also affects vegetative growth (aerial hyphae production) and spore germination, and likely stress resistance as well.
These effects are expected to differ greatly between ErdS/H-containing fungi, as illustrated by the differences in Erg-Asp levels during vegetative growth between Afm and Aor on the one hand (Erg-Asp synthesis) and Ncr and Mor on the other hand (no detectable Erg-Asp). This may also be amplified by the presence or absence of ErgS, and thus, of Erg-Gly. In addition, the spatial and temporal regulations of Erg-Asp synthesis might also differ, not only between fungal species but also at the colony and cell levels within each species. Overall, Erg-Asp synthesis (ErdS) and regulation (ErdH) likely contribute to the fitness of fungi, as supported by their conservation across fungal clades in Dikarya (Supplementary Fig. 15). Further work is needed to understand the contributions of Erg-aa to the pathogenicity and stress resistance of these species, and to clarify their functions in reproduction and growth.
The sterol-binding pocket in the ATT domain adopts a geometry and electrostatic environment tailored to accommodate the rigid structure of Erg. The deep and hydrophobic cavity is lined with basic residues, providing both shape and charge complementarity to the ligand. This binding mode is structurally distinct from lipid-interacting enzymes such as LysPGS, and illustrates how the dGNAT fold can be repurposed for the selective recognition of sterol substrates. Given the essential role of Erg in fungal membrane integrity and drug resistance, this well-defined pocket may serve as a promising scaffold for antifungal drug development. It appears highly promiscuous for sterols, but it is worth mentioning that while Erg is the main sterol of numerous fungi, including Afm, Aor, Mor or Ncr, different types of sterols are used in other species48 that also possess ErdS, such as the Erg-related sterols in Agaricomycotina and the 24-ethylcholesterol-family molecules in Pucciniomycotina (Supplementary Fig. 15). This apparent promiscuity may reflect this molecular diversity of sterols, and could result from ErdS’s evolutionary history within Dikarya.
By integrating amino acid activation, tRNA coordination, and sterol modification within a single and spatially-organized framework, ErdS exemplifies a catalytic strategy built on architectural precision and dynamic domain interplay. Rather than simply combining functions, ErdS choreographs the positioning of its substrates through tRNA-guided movements and domain rearrangements. This mechanism not only reveals a distinct solution to non-ribosomal aminoacyl transfer but also offers a design principle for engineering synthetic enzymes with programmable spatial control. The structurally resolved sterol-binding pocket further provides a tractable and selective platform with potential relevance to antifungal drug development.
Methods
Preparation of ErdS for Cryo-EM
The gene encoding A. fumigatus ErdSΔ85 (residues 86-947) was cloned into a pE-SUMO vector. The N-terminally His6-SUMO-tagged ErdSΔN85 was expressed in Escherichia coli Rosetta2 (DE3). The E. coli cells were cultured at 37 °C until the A600 reached 0.6, and protein expression was induced by adding 0.2 mM isopropyl β-D-thiogalactopyranoside (NacalaiTesque). The E. coli cells were further cultured at 20 °C overnight, collected by centrifugation, resuspended in lysis buffer (20 mM Tris-HCl, pH 8.0, 300 mM NaCl, 20 mM imidazole, 50 µM TCEP), added 50 µM PMSF and then lysed by sonication. The lysate was centrifuged, and the supernatant was incubated with Ni-NTA Agarose resin (Qiagen) for 1 h. The resin was washed with lysis buffer, and the protein was then eluted with elution buffer (20 mM Tris-HCl, pH 8.0, 300 mM NaCl, 300 mM imidazole, 50 µM TCEP). The eluted protein was treated with the SUMO protease and dialyzed against dialysis buffer (20 mM Tris-HCl, pH 8.0, 50 mM NaCl, 40 mM imidazole, 50 µM TCEP). The protein sample was passed through a Ni-NTA Agarose column to remove the His6-SUMO and SUMO protease. The protein was then loaded onto a 5 mL HiTrap SP HP column (GE Healthcare), equilibrated with buffer (20 mM Tris-HCl, 50 mM NaCl, 50 µM TCEP). The protein was further purified by chromatography on a HiLoad 16/600 Superdex 200 column (GE Healthcare), equilibrated in elution buffer (20 mM Tris-HCl, pH 8.0, 150 mM NaCl, 5 mM MgCl2, 50 µM TCEP). The purified ErdSΔ85 was concentrated to 2 mg/mL by ultrafiltration, frozen in liquid nitrogen, and stored at −80 °C.
The full-length A. fumigatus ErdS (residues 1-947) was also expressed and purified in the same way as ErdSΔ85. Except that a 50 mM NaCl concentration was used as a buffer and an affinity chromatography column, HiTrap Heparin (GE Healthcare), was used instead of the HiTrap SP HP used during purification. The purified ErdS was concentrated to 3 mg/mL by ultrafiltration, frozen in liquid nitrogen, and stored at −80 °C.
ErdS variant harboring nine substitutions at the AspRS interaction interface was also expressed and purified in the same way as full-length ErdS. Based on our ErdS structure, we identified nine residues in AspRS domain that interact with the tRNAAsp acceptor stem. To weaken this interaction, these residues were mutated as follows: Q334E, S335A, N362P, S363A, N364P, H366A, R367A, H368A, and Y494F. The purified ErdS was concentrated to 3 mg/mL by ultrafiltration, frozen in liquid nitrogen, and stored at −80 °C.
Preparation of tRNAAsp and Asp-N-tRNAAsp for Cryo-EM
The gene encoding A. fumigatus tRNAAsp in which a hammerhead ribozyme sequence and the consensus T7 promoter sequence on its 5′ end was cloned into a pUC119 vector. tRNAAsp was transcribed in vitro at 37 °C for 16-20 h using T7 polymerase, followed by incubation at 60 °C for 16-20 h with the addition of MgCl2 to a final concentration of 30 mM. The target tRNA bands were separated by urea-PAGE, extracted by EluTrap, ethanol precipitated, buffer exchanged, and stored at −80 °C.
For the preparation of the Asp-N-tRNAAsp, 3′-Overlapping forward and reverse DNA oligonucleotides (Integrated DNA Technologies) were hybridized and enzymatically extended to form a dsDNA template in which a hammerhead ribozyme sequence was sandwiched between the consensus T7 promoter sequence and the coding sequence for AfmtRNAAsp as described49. To suppress non-templated transcription, the 5′-end of the reverse oligo contained two 2′-OMe nucleotides. tRNAAsp was transcribed in vitro at 37 °C for 6 h using T7 polymerase, followed by incubation at 60-65 °C for 1 h with the addition of MgCl2 to a final concentration of 30 mM. The target tRNA bands were separated by urea-PAGE, crushed with a glass rod, and extracted into TE by shaking overnight. After centrifugation the supernatant was passed through a 0.45 µm syringe filter (33 mm; Thermo Fisher) and the tRNA was ethanol precipitated.
3′-amino-tailing followed the protocol described in42. Briefly, reactions containing 100 µM tRNAAsp, 32 µM human CCA-adding enzyme,1.75 mM 3′-amino-ATP (BioLog Life Science Institute, Bremen, GER), 1 mM sodium pyrophosphate, 10 mM MgCl2, 1.0 mM DTT, 0.2 units/µL murine RNase inhibitor (NEB), and 50 mM glycine (pH 9.0) were incubated for 1 h at 37 °C. Prior to workup, pyrophosphate was hydrolyzed by adding inorganic pyrophosphatase (0.1 U/µl; New England Biolabs) to the reaction and incubating an additional 15 min at RT. Each reaction was quenched with 2.5 M NaOAc (pH 5.0), extracted with an equal volume of pH 5.0 phenol:chloroform-isoamyl alcohol (80:17:3), and the tRNA was ethanol precipitated. The extent of 3′-tailing was determined to be 60% by biotin-streptavidin gel shift assay50.
Amino-tailed tRNA was stably charged by incubation with flexizyme (dFx) and Asp-DBE43,44. Briefly, reactions containing 40 µM tRNA (60% amino-tailed), 31 µM flexizyme, 3 mM Asp-DBE, 375 mM MgCl2, 12.5% DMSO, and 0.1 M K-HEPES (pH 7.5) were incubated in a thermomixer at 25 °C with shaking for 72 h. After 24 and 48 hr each reaction was supplemented with fresh 200 mM Asp-DBE. Reactions were quenched with 0.1 volume 2.5 M NaOAc pH 5.0 followed by ethanol precipitation. The combined pellets were dissolved in 0.1 M glycine pH 9.0 and incubated 3 h at 37 °C to hydrolyze alkali-labile aminoacyl ester linkages. After ethanol precipitation, tRNA was purified away from Flexizyme by urea-PAGE. Since stably charged tRNA could not be resolved from amino-tailed tRNA by gel electrophoresis, mass spectrometry was used to determine the charging efficiency.
Oligonucleotide LC-MS/MS analysis was performed as previously described.4 Purified tRNA was digested with RNase T1 (Thermo Fisher Scientific; 50 units per µg RNA) in 100 µL of 20 mM NH4 OAc (pH 7.0) for 2 h at 37°C. The digestion mixture was lyophilized and reconstituted in mobile phase A (10 mM HFIP, 8.6 mM TEA). Digested oligonucleotides were separated on a Poroshell 120 EC-C18 column (2.1 × 50 mm, 1.9 µm; Agilent) at 250 µL/min using a linear gradient from 0% to 5% mobile phase B (acetonitrile) over 20 min. Mass spectrometric analysis was performed on an Agilent 6545 XT LC-Q-TOF in negative ion mode. Detected oligonucleotide fragments were assigned by comparing experimental m/z values with theoretical RNase T1 digestion products.
Both amino-tailed tRNA and stably charged Asp-N-tRNAAsp were analyzed in parallel. Following sequence-specific RNase T1 digestion, the charged 3’CCA fragment (CCA-NH-Asp) was identified by its characteristic 115.1 Da mass shift, observed exclusively in the sample treated with dFx and Asp-DBE, confirming specific charging. Charging efficiency was calculated as the integrated peak area of the charged CCA fragment divided by the total CCA peak area, yielding 40.2%.
Cryo-EM sample preparation, data collection, and image processing
For the ErdS tetramer structure, ErdSΔ85 and tRNAAsp were mixed at a molar ratio of 1:1 and incubated at 37 °C for 20 min. The 3 µL of the prepared sample was applied to a freshly glow-discharged Cu/Rh 300 mesh R1.2/1.3 grid (Quantifoil), in a Vitrobot Mark IV (FEI) at 4 °C, with a waiting time of 30 s and a blotting time of 4 s under 100% humidity conditions. The grid was plunge-frozen in liquid ethane cooled at liquid nitrogen temperature.
Cryo-EM data were collected using a Titan Krios G3i microscope (Thermo Fisher Scientific), running at 300 kV and equipped with a Gatan Quantum-LS Energy Filter (GIF) and a Gatan K3 Summit direct electron detector in the electron counting mode. Micrographs were recorded at a nominal magnification of 105,000×, corresponding to a calibrated pixel size of 0.83 Å, with a total dose of 49 electrons per Å2. The data were automatically collected by the image shift method using the SerialEM software, with a defocus range of −1.6 to −0.8 μm, and 3357 movies were obtained and processed using RELION-3.151,52,53,54,55. From the 3357 motion-corrected and dose-weighted micrographs, 2,021,179 particles were initially picked, and extracted at a pixel size of 3.32 Å. These particles were subjected to several rounds of 2D and 3D classifications. The selected 240,624 particles were re-extracted at a pixel size of 1.66 Å, and then subjected to 3D refinement, per-particle defocus refinement, beam-tilt refinement, Bayesian polishing, and 3D classification with the mask focusing on ErdS monomer56. Finally, a single class of 35,812particles was selected after signal subtraction and symmetry expansion to C1, yielding a map at 3.6 Å resolution, according to the Fourier shell correlation (FSC) = 0.143 criterion57.
As for the full-length ErdS dimer, ErdS was mixed with ergosterol to a final concentration of 5 mM were incubated at room temperature for 1 h. The sample was added to a previously glow-discharged 300 mesh Au R1.2/1.3 grid (Quantifoil) for cryo-EM observation and rapidly frozen.
Cryo-EM data were collected using a Titan Krios G4 microscope (Thermo Fisher Scientific), running at 300 kV and equipped with a Gatan Quantum-LS Energy Filter (GIF) and a Gatan K3 Summit direct electron detector in the electron counting mode. Micrographs were recorded at a nominal magnification of 105,000×, corresponding to a calibrated pixel size of 0.83 Å, with a total dose of 50 electrons per Å2. The data were automatically collected by the image shift method using the EPU software (Thermo Fisher Scientific), with a defocus range of −1.6 to −0.4 μm, and 9688 movies were obtained and processed using cryoSPARC v458. Initial particle picks were used as templates for repicking, yielding 10,026,841 particles from all micrographs, and extracted59. These particles were subjected to 2D classification, and the selected 2,346,793 particles were combined for multiple rounds of three-dimensional classification (ab initio model generation and 3D classifications). The best class containing 79,545 particles was refined using flex refinement, non-uniform refinement, and local refinement, yielding a map at 3.0 Å resolution60.
As for the ErdS/tRNAAsp complex, the tRNAAsp that had been aspartylated by AspRS was mixed with twice the molar volume of ErdS and ergosterol to a final concentration of 630 µM were incubated at room temperature for 1 h. The sample was added to a previously glow-discharged 300 mesh Au R1.2/1.3 grid (Quantifoil) for cryo-EM observation and rapidly frozen.
Cryo-EM data were collected using a Titan Krios G4 microscope (Thermo Fisher Scientific), running at 300 kV and equipped with a Gatan Quantum-LS Energy Filter (GIF) and a Gatan K3 Summit direct electron detector in the electron counting mode. Micrographs were recorded at a nominal magnification of 105,000×, corresponding to a calibrated pixel size of 0.83 Å, with a total dose of 48 electrons per Å2. The data were automatically collected by the image shift method using the EPU software (Thermo Fisher Scientific), with a defocus range of −1.6 to −0.4 μm, and 2334 movies were obtained and processed using cryoSPARC v458. Initial particle picks were used as templates for repicking using Topaz, yielding 885,089 particles from all micrographs, and extracted59. These particles were subjected to 2D classification, and the selected 510,043 particles were combined for multiple rounds of three-dimensional classification (ab initio model generation and heterogeneous refinement). The best class containing 20,590 particles was refined using non-uniform refinement after CTF refinement, yielding a map at 3.66 Å resolution60. To see the ATT domain, particles were subjected to several rounds of three-dimensional classification (3D refinement and heterogeneous refinement). The best class containing 15,808 particles was refined using Non-uniform Refinement, yielding a map at 3.43 Å resolution.
As for the ErdS/tRNAAsp complex in tRNA moved state, the Asp-N-tRNAAsp was mixed with ErdS variant at a molar ratio of 0.2:1 and incubated at room temperature for 1 h. The sample was added to a previously glow-discharged 300 mesh Au R1.2/1.3 grid (Quantifoil) for cryo-EM observation and rapidly frozen.
Cryo-EM data were collected using a Titan Krios G3i microscope (Thermo Fisher Scientific), running at 300 kV and equipped with a Gatan Quantum-LS Energy Filter (GIF) and a Gatan K3 Summit direct electron detector in the electron counting mode. Micrographs were recorded at a nominal magnification of 105,000×, corresponding to a calibrated pixel size of 0.83 Å, with a total dose of 51 electrons per Å2. The data were automatically collected by the image shift method using the EPU software (Thermo Fisher Scientific), with a defocus range of −1.6 to −0.4 μm, and 7,809 movies were obtained and processed using cryoSPARC v4. Initial particle picks were used as templates for repicking using Topaz, yielding 3,038,832 particles from all micrographs, and extracted. These particles were subjected to 2D classification, and the selected 354,720 particles were combined for multiple rounds of three-dimensional classification (ab initio model generation and 3D refinement). The best class containing 34,200 particles was refined using non-uniform refinement after CTF refinement, yielding a map at 3.65 Å resolution.
Model building, validation and superimpositions
For atomic model building, initial ErdS models were generated by AlphaFold261. The initial tRNAAsp was derived from the crystal structure of yeast tRNAAsp (PDB 1ASY)17. The models were manually docked into cryo-EM map and modeled in Coot and refined using phenix.real_space_refine62,63. The structure validation was performed using MolProbity in the PHENIX package64. The statistics of the 3D reconstruction and model refinement are summarized in Supplementary Data Table 1. The cryo-EM density map, figurers and molecular graphics were generated using UCSF ChimeraX65,66. Models were superimposed using the match-maker in ChimeraX. Ergosterol was positioned and relaxed by Sphere Refine on COOT.
Docking and molecular dynamics simulations
Ligand-bound models were first generated using AlphaFold3. For the ligand-bound predictions used in this study, the ATT domain (same sequence as in our cryo-EM construct) was provided to AlphaFold3 together with the ligand. Cholesterol (Cho) and diosgenin poses shown in the manuscript were obtained from AlphaFold3-predicted ligand-bound models. An AlphaFold3-predicted ergosterol (Erg)-bound pose was also used as one of the two starting models for molecular dynamics (MD) simulations. The backbone RMSD between the cryo-EM ATT domain structure and the corresponding AlphaFold3 model was 2.53 Å on average (variance 0.244).
In parallel, a docking-based Erg starting pose was generated using the SwissDock web server (EADock DSS). Docking was restricted to the putative sterol-binding cavity at the GNAT I–GNAT II interface, with the search region defined based on residues implicated by our structure-guided mutagenesis. SwissDock was run with default settings, and a representative pose located within the mutagenesis-supported cavity and free of steric clashes was selected. For comparison, LPG was docked into LysPGS using SwissDock with the search space restricted to the corresponding lipid-binding pocket.
MD simulations were then performed with GROMACS 2025.3 to evaluate the stability of sterol accommodation in the proposed cavity. Two alternative Erg starting poses were used: (i) the SwissDock-derived pose positioned near residues highlighted by our mutagenesis results and (ii) the AlphaFold3-predicted Erg-bound pose. For each starting model, ten independent simulations with different random seeds were performed for 100 ns. The AMBER99SB-ILDN force field was used for the protein, and the TIP3P model was used for water. Ergosterol was parameterized with GAFF2 (General AMBER Force Field 2), and partial charges were assigned using Antechamber/ACPYPE with the AM1-BCC scheme. Subsequently, water molecules were added to the system with a thickness of more than 10 Å, and counter-ions (Cl − ) were added to neutralize the net charge (no additional salt was added). The resulting simulation system contained 70,844 atoms in total (protein: 5,675; Erg: 73; water: 21,698 molecules[65,094 atoms]; ions: 2).
After construction of the initial structures, the systems were optimized until the maximum force was lower than 1000 kJ/mol/nm by using the steepest descent method. Subsequently, 50 ps NVT equilibration at 300 K was performed with position restraints applied to the protein backbone (1000 kJ/mol⁻¹/nm⁻²) and ligand heavy atoms (500 kJ/mol⁻¹/nm⁻²), followed by another 50 ps NVT equilibration at 300 K with backbone restraints only. During subsequent 100-ps NPT simulations at 300 K and 1 bar using the V-rescale thermostat and C-rescale barostat, position restraints were applied to the protein backbone (1000 kJ/mol⁻¹/nm⁻²). After equilibration, all restraints were removed, and ten independent production MD simulations with different random seeds were performed for 100 ns using a 2-fs time step with temperature controlled by the V-rescale thermostat and bonds constrained using LINCS.
For analysis, each trajectory frame was aligned on the protein backbone, and the ligand RMSD was calculated.
Fungal strains and growth media
All strains and derivatives used in this study are listed in Supplementary Table 1.S. cerevisiae strains used for plasmid shuffling are also presented in Supplementary Table 2.
S. cerevisiae strains BY4742 (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0) were used to express WT ErdS and mutants from pRS415 (LEU) plasmids. They were routinely grown on SC-Leu agar (MP™ Biomedicals) at 30 °C. The S. cerevisiae YAL3 (dps1Δ) shuffle strain (MATa ura3-52 lys2-801am trp1-63 his3-200 leu2-1 ade2-450 ade3-1483 dps1::HIS3) contained a rescue plasmid (TRP) and pRS415 (LEU) plasmids carrying WT of mutant versions of ErdS. Before shuffling, they were grown on SC-Leu-Trp at 30 °C, and after shuffling, when the rescue plasmid was chased, on SC-Leu agar (MP™ Biomedicals). For liquid cultures, the same media without agar were used in glass flasks, and cultures were incubated at 30 °C under shaking (200 rpm).
For Aspergillus fumigatus, the CEA17 ΔakuBKU80 strain (aka “WT KU80”) and derivatives were used A. fumigatus was maintained as follows: fresh conidia were spread on Malt agar (Thermo Scientific) plates or slants and incubated in the dark at 37 °C until they produced enough mature conidia. For phenotypic characterization, strains were grown on Malt agar or on Minimal Medium (MM) Agar plates (For 1 L, MM contained glucose (2 % w/v) or xylose (2 % w/v), ammonium tartrate dibasic (0.92 g, 5 mM, i.e., 10 mM ammonium), salts (10 mL of a 50 X solution containing KCl 26 g/L, MgSO4 7H2O 26 g/L, KH2PO4 76 g/L) and trace elements (0.5 mL of a 1000 X solution containing: FeSO4 7H2O 1 g, Na2EDTA 10 g, ZnSO4 7H2O 4.4 g, H3BO3 2.2 g, MnCl2 4H2O 1 g, CoCl2 6H2O 0.32 g, CuSO4 5H2O 0.32 g and Na2MoO40.8 g for 200 mL adjusted at pH 6.5; 1.5 % (w/v) agar was added). For liquid cultures, Afm was grown in liquid MM with 1 % (w/v) glucose. Liquid cultures were performed in glass flasks at 37 °C (Afm) under shaking (200 rpm) for 24 h (Afm, Sce) or 48 h (Ncr). For Neurospora crassa, the 74-OR23-1VA WT (Fungal Genetic Stock Center, strain FGSC#2489) and ΔerdH (FGSC#20235) strains were employed. N. crassa was grown on nutrient-rich plates (Glucose 4% w/v, Peptone 1% w/v, and Yeast extract 2% w/v), or in MM liquid medium at 30 °C. Magnaporthe oryzae strains (isolate Guy11) were routinely grown, maintained, and stored as previously described67.
Construction of A. fumigatus strains
Construction of the AfmΔerdS and ΔerdS::PXyl-erdS strains was described in ref. 11. The WT::PXyl-erdS strain was constructed using the same integration cassette as that used for the ΔerdS::PXyl-erdS strain. It was designed from the six-Pxyl-βrec-trpC-hygB-six resistance cassette (with the β-recombinase six sequences)68 surrounded by the 5’ and 3’ 1000-bp flanking regions of the erdS gene. The erdS gene was inserted in place of the β-recombinase gene between the PXyl promoter and TrpC terminator11. Transformations of A. fumigatus CEA17 ΔakuBKU80 were performed by electroporation of swollen conidia essentially as described69 and we used 10 μg of linearized (SmaI-digested) plasmid containing the replacement cassette. From the primary hygromycin-resistant colonies, conidia were isolated on individual Malt agar plates containing 150 μg/mL hygromycin B. After 3 days, conidia from single isolated colonies were scraped and transferred to Malt agar slants containing 150 μg/mL hygromycin B and incubated 5 days at 37 °C. Single isolated colonies DNA was extracted70 from mycelium cultures of mutants, and replacement of the Afm-erdS gene was confirmed by PCR (Primers FF#684 + NY#015 and NY#016 + FF#685).
Construction of Afm strains expressing ErdS-eGFP and ErdH-mCherry. With the E-Zyvec company (now Polyplus), we constructed plasmids containing the erdH—erdSlocus found in Afm that we modified by fusion codon-optimized ORFs of eGFP and mCherry, respectively, at the 3’-end of erdS and erdH. Both genes were thus under the control of their own natural promoter (Perd). This construct was inserted before the six-Pxyl-βrec-trpC-hygB-six cassette. The 5’- and 3’-flanking regions of the erdH—erdS locus were added to obtain the 5’-flank_erdH-mCherry—erdS-eGFP-six-Pxyl-βrec-trpC-hygB-six_3’_flankcassette, surrounded by SmaI restriction sites. The resulting plasmid was digested using SmaI and transformed in the A. fumigatus CEA17 ΔakuBKU80(WT KU80) strain by electroporation, as described. Hygromycin-resistant colonies were selected on MM medium (pH 6.8) containing 150 µg/mL hygromycin B and purified through two successive re-streaks on Malt agar containing 150 µg/mL hygromycin. Strains that presented red and green fluorescence were then grown on MM containing 2 % (w/v) xylose to excise the six-Pxyl-βrec-trpC-hygB-six cassette, and to ensure that only the 5’-flank_erdH-mCherry—erdS-eGFP-_3’_flank construct remained. Two independent clones were selected for epifluorescence observations. Strains were verified by PCR.
Construction of M. oryzae strains (CrispR)
The erdS (MGG_13783) and erdH (MGG_04680) genomic sequences were retrieved from EnsemblFungi [https://fungi.ensembl.org/]. erdS and erdH were deleted using the RNP-mediated CrispR editing strategy as previouslyadapted in M. oryzae71. To generate the donor DNAs, the BAR (glufosinate-ammonium resistance) cassette was amplified from the pCB1530 plasmid (FGSC) using approx. 100 bp-long primers leading to amplicons containing the BAR cassette flanked by erdS or erdH homologous arms. The sgRNA were obtained by in vitro transcription using the EnGen® sgRNA Synthesis Kit (Neb #3322) following the manufacturer’s instructions. All primers used to generate and verify the strains are listed in Supplementary Table 3.
Plasmids construction
The Afm erdS open reading frame (AFUA_1g02570) was synthesized (Genscript®) with codon optimization for expression in Sce, amplified by PCR and cloned in a pRS415 (LEU) yeast expression vector as described in ref. 11. The p415-Aor-erdS plasmid containing the erdS gene from Aor was constructed previously11. All ErdS mutants were obtained from these pRS415 plasmids through mutagenesis. Briefly, for site-directed mutagenesis, we proceeded as follows. A pair of overlapping primers containing the desired mutation were used (Supplementary Table 3). The “reverse” primer was used together with a “forward” primer matching the ampicillin resistance gene of the plasmid (FF#060) to amplify by PCR a first half of the pRS415-erdS plasmid, containing the desired mutation. The “forward” primer was used with the “reverse” primer (FF#061) matching the ampicillin resistance gene of the plasmid to amplify the second half of the plasmid, with the desired region mutated. Then, 1 µL of each PCR reaction were used in a Gibson Assembly reaction mixture to reunite the two plasmid halves. Then, 8 µL of the reaction mixture was used to transform XL-1 Blue chimiocompetent E. coli cells to select plasmids in which the ampicillin resistance gene was properly reconstituted. The same strategy was used to obtain the erdSΔ(1-90) mutant of Aor erdS from the pRS415-Aor erdS plasmid. The presence of the desired mutations in erdS was verified and validated by sequencing.
Plasmid-shuffling complementation assays in S. cerevisiae
Plasmid shuffling experiments were conducted in the YAL3Δdps1 Scestrain73rescued with a wild-type DPS1 gene copy cloned in an URA3-bearing plasmid. All Afmor Aor erdS constructs to be tested were cloned in pRS415 (LEU) plasmids, transformed into the Δdps1 strain, and shuffled essentially as described34,72.
Preparation of spores/conidia from A. fumigatus and N. crassa
Spores from 7 days-old Malt agar slants or plates were resuspended by the addition of 5 mL sterile Tween 20-H2O (0.05 % v/v) and vortexing, then the conidia were filtered with Cell Strainer filters (EASY strainerTM Greiner Bio-One), and the concentration was determined with a LUNA-FL cell counter (ref.59506, Dutscher). Conidia were stored in Tween 20-H2O (0.05 % v/v) at 4 °C for up to 2 weeks. For Ncr, conidia were harvested from 14 days-old Malt agar slants and treated similarly using 1 M sterile sorbitol. For M. oryzae, spores were harvested from 7-9 days old cultures grown on CM agar plates. 4 mL were poured on the Petri dish, and spores were released by scratching the mycelial surface using a sterile disposable plastic spreader. The spore solution was then filtered through Miracloth, centrifuged at 3000 x g for 5 min and finally resuspended in 1 mL of water. Cells were counted using a Bright-Line™ Hemacytometer (Merck #Z359629).
Phenotypic characterization of A. fumigatus strains
For phenotypic characterizations, 106Afm conidia in 5 µL were point-inoculated on squared agar plates from suspensions prepared as indicated below to obtain individual round-shaped colonies. Each plate was inoculated with the 4 Afm derivative (WT Ku80, ΔerdS, ΔerdS::PXyl-erdS and WT::PXyl-erdS), as to ensure proper comparison between strains. Colony diameter was measured at indicated time points and colonies were pictured using a cell phone or using an AxioVision magnifier. To produce mycelia lawns, 50 µL of a conidia suspension at 107 conidia/mL was spread on MM agar medium-containing wells of 12-well plates. Each plate featured 3 replicates of the 4 strains tested. Lawns were pictured using an AxioVision magnifier. All plates were incubated at 37 °C in the dark and documented at the indicated time points.
Liquid cultures and mycelia harvesting
Liquid cultures to produce mycelia for total lipids extraction were inoculated with 107 conidia/mL in 50 mL liquid MMG (MM + 1 % (w/v) glucose), incubated for 24 h at 37 °C (Afm) or 48 h at 30 °C(Ncr) in the dark under agitation (220 rpm). To test the overproduction of Erg-Asp, the 4 Afm strains were grown in liquid MM + 2 % (w/v) glucose (MMG) or MM + 2 % (w/v) xylose (MMX). Experiments were also conducted with cultures in the presence of various ratios of glucose and xylose. Mycelia were then filtered through two layers of gauze, rinsed twice with 50 mL sterile H2O, and squeezed to eliminate excess water. Mycelia were directly used to extract total lipids.
Germination assay
To investigate the role of Erg-Asp in the germination process, conidia suspension (1×106 conidia /mL) from WT KU80 and ΔerdS strains were prepared. Germination assays were performed in 1 mL MM medium in Eppendorf tubes, incubated at 37 °C with agitation at 200 rpm. Microscopic observation began 3 hours after incubation, then continued hourly. At each time point, samples were collected and centrifuged at maximum speed for 10 min at 4 °C. Supernatants were removed, and pellets were fixed in cold 4% paraformaldehyde (4% PFA) for 15 min at room temperature. After a second centrifugation (10 min, max speed, 4 °C) and removal of the fixative, conidia were resuspended in 1 mL, sterile 1xPBS (phosphate-buffered saline) and stored à 4 °C until microscopy observations. Bright field images were captured hourly using a Zeiss Axiovert 200 epifluorescence microscope with a 65x objective. Germination was manually assessed by examining several fields of view for each strain and each time point. Conidia were outlined manually, totaling approximately 100 fungal cells per strain per time point. Observations were recorded hourly 3–8 h post-incubation. Germination percentages at the germ tube stage were compared between WT KU80 and ΔerdS strains over time using a two-way ANOVA statistical test performed with GraphPad Prism. Multiple comparisons were conducted using the Sidak test, with a significance threshold set at 0.05.
Nanolive microscopy
Cell Culture. Murine macrophage IC-21 cells were cultured under standard conditions at 37 °C with 5 % CO2. They were maintained in complete growth medium (RPMI + FBS + PS) and seeded for two different experiments in 35 mm ibidi low border dishes. On July 18, 2023, five dishes were seeded with 60,000 cells each, while on July 24, 2023, two additional dishes were seeded with 80,000 cells each. Infection Preparation and Conditions: For the first experiment (WT KU80 conidia), one dish was used to assess cell viability following a 30-minute incubation at 4°C, a condition necessary for synchronizing phagocytosis. A second dish was used for cell counting to calculate the appropriate number of spores needed to reach the desired multiplicity of infection (MOI). The remaining three dishes were used for infection with WT Ku80 spores: one dish served as a negative control, one was infected at MOI 5, and the third at MOI 10. To achieve this, the stock solution (2.36 × 10⁹ spores in 4 mL) was first pre-diluted 1:200 (3 µL stock + 597 µL medium) to obtain 1.77 × 10⁶ spores in 600 µL. For an MOI of 10, 237 µL of this pre-dilution (corresponding to 700,000 spores) was added to 70,000 IC-21 cells in a final volume of 800 µL. For an MOI of 5, 119 µL (350,000 spores) was used under the same conditions. After 24 h of incubation, spores were added to the respective dishes. To synchronize phagocytosis, all infected dishes were placed at 4 °C for 30 min, followed by a medium change to remove unbound spores. In the second experiment (∆erdS spores) realized on July 24, 2023, IC-21 cells were infected with ∆erdS spores at MOI 5 and MOI 10. In this case, the stock solution (3.28 × 10⁹ spores in 4 mL) was also pre-diluted 1:200 (3 µL stock + 597 µL medium) to obtain 2.46 × 10⁶ spores in 600 µL. For an MOI of 10, 196 µL of this pre-dilution (corresponding to 800,000 spores) was added to 80,000 IC-21 cells per dish in a final volume of 800 µL. For an MOI of 5, 98 µL (400,000 spores) was used under the same conditions. The spore solution was taken out of the fridge for 15 min before dilution to reach room temperature and the cells were taken out of the incubator 5 min before adding the spores. As in the previous experiment, cells were incubated with spores at 4 °C for 30 min, then washed by changing the medium to eliminate unbound spores. All Afm spore preparations were performed one by one with complete disinfection in between to avoid any risk of cross-contamination between spore types. Image Acquisition: Following spore addition and washing, live-cell imaging was performed using a CX96-Focus (Nanolive SA, Tolochenaz, Switzerland) microscope. The microscope is equipped for long-term live cell imaging: temperature, humidity, and gas composition. The incubator chamber (Okolab) keeps the sample at 37 °C, is closed by a heating glass lid to prevent condensation, and is connected to a gas mixer (2GF-Mixer, Okolab) to maintain 5% of CO2. The humidity module ensures a 90% relative humidity within the chamber. The first experiment was done with a 3×3 grid scan with one image every 2min9s, while the second used a 4×4 grid scan with one image every 2min21s.
Fluorescence microscopy of A. fumigatus
Conidia from two independent Afm strains carrying the erdH-mCherry—erdS-eGFP locus were used to inoculate liquid YG medium (1% Yeast extract, 2% glucose) (105 conidia/mL). Development of the fungus was followed at the indicated time points. At each time point, 1 mL of culture was aliquoted, conidia or mycelia centrifugated and washed with phosphate buffer saline (PBS), resuspended in PBS and 10 µL of the resuspended strains were used to prepare slides. Epifluorescence images were taken with an AXIO Observer d1 (Carl Zeiss) epifluorescence microscope using a 100 × plan apochromatic objective (Carl Zeiss) and processed with the Image J software.
Total lipids extraction
Overnight Sce cultures were diluted to an OD600 = 0.1, grown (220 rpm) until the OD600 was ~1 and cells were harvested by centrifugation at 5000×g for 15 min at 4 °C. Afm or Ncr conidia were harvested as described and inoculated in glass flasks (50 mL) in the MMG medium at a concentration of 106 conidia/mL, shaken at 37 °C during 24 h (Afm) or 30 °C for 48 h (Ncr). Mycelia were collected as described above.
For total lipid extraction, 50 OD600a pellet of PBS-washed Sce cells were resuspended in 0.5 mL of 120 mM Na-Acetate, pH 4.5. Then, 3.75 volumes of CHCl3:CH3OH (2:1) and 1 mL of glass beads (ø 0.25-0.5 mm, Roth) were added and cells disrupted through mechanical lysis using a FastPrep Instrument (MP™ Biomedicals, Serial N° 10020698) at 1 min 5.5 m/s repeated 6 times with cooling on ice between each cycle. Cell lysates were incubated 3 h on a rotating wheel at 4 °C. Then, 1.25 volumes of CHCl3 and 1.25 volumes of 120 mM Na-Acetate pH 4.5 were added and the samples vortexed 1 min. Phases were separated by centrifugation (9000×g; 30 min; 4 °C) and the lower organic phase containing lipids was transferred into a clean glass tube and dried under vacuum (SpeedVac vacuum concentrator). Drying was finalized under an argon flow. Lipids were stored at −20 °C or resuspended in 50-100 µL of CHCl3:CH3OH (1:1, v:v) for analysis on TLC. In the case of Afm and Ncr strains, mechanical cell disruption was performed as follows: 2 g of fresh mycelia, dried on paper towel, were ground in a mortar with a pestle in the presence of liquid nitrogen and the resulting fine powder was resuspended in 1 mL (1 vol.) of 120 mM Na-Acetate pH 4.5 and treated as described above. For Afm conidia total lipids, 7-day old mycelial lawns grown on on malt agar plates were flooded with 10 mL of sterile H2O/0.05 % Tween-20 and conidia scrapped off the plate, filtered through a 40 µm cell strainer (ref. 0999225, Grosseron) and centrifuged 20 min, 8000×g at 4 °C. The supernatants were discarded and spores treated as described for Sce cells.
Lipid analysis by thin-layer chromatography (TLC)
TLC plates (Silica gel 60 aluminum foils, Sigma-Aldrich, 10 × 10 cm) were used. Lipids in the CHCl3:CH3OH (1:1, v:v) solvent were spotted on TLC, typically, 10–20 µL of total lipids or 25 µL of radiolabeled lipids extracted from in vitro reactions (see below). TLCs were developed with the CHCl3:CH3OH:H2O mobile phase (130:50:8) for 10 min, and air-dried. TLCs were stained with a sulfuric acid/MnCl2 solution (concentrated sulfuric acid 9 mL, MnCl2.4H2O 0.8 g, CH3OH 120 mL, H2O 120 mL) or with ninhydrin (Sigma-Aldrich, 0.4 % w/v in EtOH 100 %, v/v) and heated at 100 °C, 15 min. Plates were imaged under white light or at 254 nm. Radiolabeled lipid species were revealed by exposing TLC plates to a Fuji Imaging Plate and analyzed with a Typhoon TRIO, Variable mode imager (GE Healthcare).
In Vitro lipid aminoacylation assay
Reactions were performed in the following mix: 100 mM Na-Hepes pH 7.2 buffer containing 30 mM KCl, 12 mM MgCl2, 10 mM ATP, 0.1 mg/mL bovine serum albumin, pure yeast tRNAAsp (10 μM) (46), 10 µM [U-14C]-Asp (280 cpm/pmol, Perkin-Elmer, NEC268E050UC) in a final volume of 50 μL. To test the transfer of [14C]-Asp onto lipids, total lipids or pure sterols were added to a final concentration of 2 mg/mL, and commercial pure sterols (Merck, Avanti Lipids) were added to a final concentration of 0.5 mg/mL. Purified ErdSΔN86 was added to initiate the reaction before a 45-min-long incubation at 30 °C. After incubation, reactions were stopped with addition of 500 μL of CHCl3:CH3OH:120 mM Na-acetate pH 4.5 (130:50:8, v/v/v) and vortexing. Then, 130 μL of CHCl3 and 130 μL of Na-acetate 120 mM pH 4.5 were successively added, and the mixture was vortexed and centrifuged for 1 min at 5000 × g (RT). The lower organic phase was recovered and dried under vacuum. Reaction products were then dissolved in a CHCl3:CH3OH (1:1, v:v) mixture, spotted on TLC plates and separated with the CHCl3:CH3OH:H2O mobile phase. TLC plates were exposed onto an imaging plate (Fuji Imaging plate) for at least 2 h. Radioactivity was detected using a Typhoon TRIO variable Mode Imager (GE Healthcare). Quantification of radioactive spots or Erg-Asp and phosphatidylethanolamine (PE) bands was performed using the ImageJ software (number of pixels).
Protein extraction and Western blots
Yeast total proteins from Sce were extracted using 1 OD600nm of cells that were resuspended and incubated 10 min in 500 μL of pre-cooled NaOH 0.185 N, then precipitated by adding 50 μL of Trichloroacetic acid (TCA) 100 % and incubated 10 min on ice. Finally, the samples were centrifuged at 13,000 × g for 15 min and the resulting pellets were resuspended in 100 μL of Laemmli Sample Buffer. For Afm total protein extraction, fresh mycelia were harvested from 24 h-old liquid cultures, washed with sterile distilled H2O, suspended in 100 mM Tris-HCl pH 8.1, 30 mM KCl, 50 mM NaCl, glycerol 10 % (v/v), 0.25 % Tween-20 and a protease inhibitor cocktail (Roche Complete, EDTA-free). Mycelia were disrupted in a FastPrep-24 apparatus (MP Biomedicals, serial no. 10020698) with cooling on ice. Then, crude extracts were centrifugated at 800 × g and the supernatant precipitated with Trichloroacetic acid (TCA) 100% and incubated 10 min on ice, as described for yeast proteins. Then, 8 μL of each sample was resolved on 10% SDS-PAGE gels. Samples were separated by using a BioRad Mini-PROTEAN electrophoresis apparatus. For western blotting, proteins were transferred onto PVDF membranes that were blocked in 5% (w/v) skimmed milk in TBS-Tween (TBS 1X, Tween-20 0.3 % (v/v)) for 1 h at RT. Primary antibodies were polyclonal anti-ErdS, Covalab, France (dilution 1/10,000 for Sce extracts and 1/1000 for Afm extracts), and anti-PGK (dilution 1/10,000) and were incubated overnight at 4 °C with TBS-Tween. Membranes were then incubated for 1 h with HRP-conjugated secondary antibodies (Goat anti-rabbit for anti-ErdS and Goat anti-mouse for anti-PGK) at RT. Revelation was performed with the BioRad clarity western ECLKit and monitored in a BioRad ChemiDocTouch® apparatus.
ErdS sequence alignments
The sequence of AfmErdS was used as the probe to search homologs using BLAST with the default parameters. Then, 98 homologous sequences were selected across Dikarya and aligned using MUSCLE73, and the resulting alignment was used to produce a sequence logo at WebLogo 3 [https://weblogo.threeplusone.com/create.cgi].
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The source data underlying Figureures 1B, -E, -F, 2 A, -B, -C, 3D, -G, 4 F, and Supplementary Figure S1A, -F, S2A, -C, -D, -E, -F, -G, S8B, -D, are provided as a Source Data file. The atomic models have been deposited in the PDB under the accession codes: 9W9E (ErdS tetramer), 9W9F (ErdS/tRNA dimer on tRNA located AspRS) and 9W9G (ErdS/tRNA dimer on tRNA located ATT.). The cryo-EM density map has been deposited in the Electron Microscopy Data Bank under the accession codes: EMD-65772 (ErdS tetramer), EMD-65773 (ErdS/tRNA dimer on tRNA located AspRS) EMD-65774 (and ErdS/tRNA dimer on tRNA located ATT) Source data are provided with this paper.
References
Roy, H. & Ibba, M. RNA-dependent lipid remodeling by bacterial multiple peptide resistance factors. Proc. Natl. Acad. Sci. USA 105, 4667–4672 (2008).
Moutiez, M., Belin, P. & Gondry, M. Aminoacyl-tRNA-Utilizing Enzymes in Natural Product Biosynthesis. Chem. Rev. 117, 5578–5618 (2017).
Groenewold, M. K. et al. Virulence of Agrobacterium tumefaciens requires lipid homeostasis mediated by the lysyl-phosphatidylglycerol hydrolase AcvB. Mol. Microbiol. 111, 269–286 (2019).
Gill, C. P. et al. The MprF homolog LysX synthesizes lysyl-diacylglycerol contributing to antibiotic resistance and virulence. Microbiol. Spectr. 11, e0142923 (2023).
Yakobov, N. et al. RNA-dependent synthesis of ergosteryl-3β-O-glycine in Ascomycota expands the diversity of steryl-amino acids. J. Biol. Chem. 298, 101657 (2022).
Dyda, F., Klein, D. C. & Hickman, A. B. GCN5-related N-acetyltransferases: a structural overview. Annu. Rev. Biophys. Biomol. Struct. 29, 81–103 (2000).
Vetting, M. W. et al. Structure and functions of the GNAT superfamily of acetyltransferases. Arch. Biochem. Biophys. 433, 212–226 (2005).
Biarrotte-Sorin, S. et al. Crystal structures of Weissella viridescens FemX and its complex with UDP-MurNAc-pentapeptide: insights into FemABX family substrates recognition. Structure 12, 257–267 (2004).
Fonvielle, M. et al. The structure of FemXWv in complex with a peptidyl-RNA conjugate: mechanism of aminoacyl transfer from Ala-tRNAAla to peptidoglycan precursors. Angew. Chem. Int. Ed. 52, 7278–7281 (2013).
Hebecker, S. et al. Structures of two bacterial resistance factors mediating tRNA-dependent aminoacylation of phosphatidylglycerol with lysine or alanine. Proc. Natl. Acad. Sci. USA 112, 10691–10696 (2015).
Yakobov, N. et al. RNA-dependent sterol aspartylation in fungi. Proc. Natl. Acad. Sci. USA 117, 14948–14957 (2020).
Grob, G. et al. tRNA-dependent addition of amino acids to cell wall and membrane components. Biochimie 203, 93–105 (2022).
Yokokawa, D. et al. Synthesis of aminoacylated ergosterols: a new lipid component of fungi. Steroids 169, 108823 (2021).
Ibba, M. & Soll, D. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 69, 617–650 (2000).
Ruff, M. et al. Class II aminoacyl transfer RNA synthetases: crystal structure of yeast aspartyl-tRNA synthetase complexed with tRNA Asp. Science 252, 1682–1689 (1991).
Fields, R. N. & Roy, H. Deciphering the tRNA-dependent lipid aminoacylation systems in bacteria: Novel components and structural advances. RNA Biol. 15, 480–491 (2018).
Favrot, L., Blanchard, J. S. & Vergnolle, O. Bacterial GCN5-related N -Acetyltransferases: from resistance to regulation. Biochemistry 55, 989–1002 (2016).
Blaise, M. et al. Crystal structure of a transfer-ribonucleoprotein particle that promotes asparagine formation. EMBO J. 29, 3118–3129 (2010).
Suzuki, T. et al. Structure of the Pseudomonas aeruginosa transamidosome reveals unique aspects of bacterial tRNA-dependent asparagine biosynthesis. Proc. Natl. Acad. Sci. Usa. 112, 382–387 (2015).
Jordá, T. & Puig, S. Regulation of ergosterol biosynthesis in saccharomyces cerevisiae. Genes 11, 1–18 (2020).
Sun, Y. et al. Mevalonate diphosphate decarboxylase MVD/Erg19 is required for ergosterol biosynthesis, growth, sporulation and stress tolerance in aspergillus oryzae. Front. Microbiol. 10, 1074 (2019).
Singh, A. & Del Poeta, M. Lipid signalling in pathogesnic fungi. Cell. Microbiol. 13, 177–185 (2011).
Slavetinsky, C., Kuhn, S. & Peschel, A. Bacterial aminoacyl phospholipids – Biosynthesis and role in basic cellular processes and pathogenicity. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1862, 1310–1318 (2017).
Yokokawa, D. et al. Sterol aminoacylation regulates conidiation and sclerotia formation in Aspergillus oryzae. Fungal Biol. 129, 101609 (2025).
Latgé, J.-P. & Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 33, e00140-18 (2019).
Tan, J., Zhao, H., Li, J., Gong, Y. & Li, X. The devastating rice blast airborne pathogen magnaporthe oryzae—a review on genes studied with mutant analysis. Pathogens 12, 379 (2023).
Van De Veerdonk, F. L., Gresnigt, M. S., Romani, L., Netea, M. G. & Latgé, J. P. Aspergillus fumigatus morphology and dynamic host interactions. Nat. Rev. Microbiol. 15, 661–674 (2017).
Brown, G. D. et al. Hidden killers: human fungal infections. Sci. Transl. Med. 4, 3004404 (2012).
Araujo, R. & Rodrigues, A. G. Variability of germinative potential among pathogenic species of aspergillus. J. Clin. Microbiol. 42, 4335–4337 (2004).
Manavathu, E. K., Cutright, J. & Chandrasekar, P. H. Comparative study of susceptibilities of germinated and ungerminated conidia of aspergillus fumigatus to various antifungal agents. J. Clin. Microbiol. 37, 858–861 (1999).
Pereira de Sa, N. & Del Poeta, M. Sterylglucosides in Fungi. J. Fungi 8, 1130 (2022).
Liu, Z. et al. Functional genomic and biochemical analysis reveals pleiotropic effect of congo red on aspergillus fumigatus. mBio 12, e00863-21 (2021).
Salah Ud-Din, A., Tikhomirova, A. & Roujeinikova, A. Structure and functional diversity of GCN5-related N-Acetyltransferases (GNAT). Int. J. Mol. Sci. 17, 1018 (2016).
Ador, L. et al. Active site mapping of yeast aspartyl-tRNA synthetase by in vivo selection of enzyme mutations lethal for cell growth. J. Mol. Biol. 288, 231–242 (1999).
Heese-Peck, A. et al. Multiple functions of sterols in yeast endocytosis. Mol. Biol. Cell 13, 2664–2680 (2002).
Francin, M., Kaminska, M., Kerjan, P. & Mirande, M. The N-terminal domain of mammalian Lysyl-tRNA synthetase is a functional tRNA-binding domain. J. Biol. Chem. 277, 1762–1769 (2002).
Francin, M. & Mirande, M. Functional dissection of the eukaryotic-specific tRNA-interacting factor of lysyl-tRNA synthetase. J. Biol. Chem. 278, 1472–1479 (2003).
Frugier, M., Moulinier, L. & Giegé, R. A domain in the N-terminal extension of class IIb eukaryotic aminoacyl-tRNA synthetases is important for tRNA binding. EMBO J. 19, 2371–2380 (2000).
Ryckelynck, M., Giegé, R. & Frugier, M. Yeast tRNAAsp charging accuracy is threatened by the N-terminal extension of aspartyl-tRNA synthetase. J. Biol. Chem. 278, 9683–9690 (2003).
Moulinier, L. et al. The structure of an AspRS–tRNAAsp complex reveals a tRNA-dependent control mechanism. EMBO J. 20, 5290–5301 (2001).
Devarkar, S. C. et al. Structural basis for aminoacylation of cellular modified tRNALys3 by human lysyl-tRNA synthetase. Nucleic Acids Res. 53, 114 (2025).
Gamper, H. & Hou, Y.-M. tRNA 3′-amino-tailing for stable amino acid attachment. RNA 24, 1878–1885 (2018).
Murakami, H., Ohta, A., Ashigai, H. & Suga, H. A highly flexible tRNA acylation method for non-natural polypeptide synthesis. Nat. Methods 3, 357–359 (2006).
Katoh, T. & Suga, H. Flexizyme-catalyzed synthesis of 3′-aminoacyl-NH-tRNAs. Nucleic Acids Res. 47, 143 (2019).
Perham, R. N. Swinging arms and swinging domains in multifunctional enzymes: catalytic machines for multistep reactions. www.annualreviews.org (2000).
Nureki, O. et al. Enzyme structure with two catalytic sites for double-sieve selection of substrate. Science 280, 578–582 (1998).
Fukai, S. et al. Structural basis for double-sieve discrimination of L-Valine from L-Isoleucine and L-threonine by the complex of tRNAVal and Valyl-tRNA synthetase. Cell 103, 793–803 (2000).
Weete, J. D., Abril, M. & Blackwell, M. Phylogenetic distribution of fungal sterols. PLoS One 5, e10899 (2010).
Fechter, P., Rudinger, J., Giegé, R. & Théobald-Dietrich, A. Ribozyme processed tRNA transcripts with unfriendly internal promoter for T7 RNA polymerase: production and activity. FEBS Lett. 436, 99–103 (1998).
Gamper, H. & Hou, Y. M. A label-free assay for aminoacylation of tRNA. Genes (Basel) 11, 1–15 (2020).
Mastronarde, D. N. Automated electron microscope tomography using robust prediction of specimen movements. J. Struct. Biol. 152, 36–51 (2005).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Scheres, S. H. W. RELION: implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 180, 519–530 (2012).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. Elife 7, 42166 (2018).
Rohou, A. & Grigorieff, N. CTFFIND4: fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221 (2015).
Zivanov, J., Nakane, T. & Scheres, S. H. W. A Bayesian approach to beam-induced motion correction in cryo-EM single-particle analysis. IUCrJ 6, 5–17 (2019).
Rosenthal, P. B. & Henderson, R. Optimal determination of particle orientation, absolute hand, and contrast loss in single-particle electron cryomicroscopy. J. Mol. Biol. 333, 721–745 (2003).
Punjani, A., Rubinstein, J. L., Fleet, D. J. & Brubaker, M. A. CryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nat. Methods 14, 290–296 (2017).
Bepler, T. et al. Positive-unlabeled convolutional neural networks for particle picking in cryo-electron micrographs. Nat. Methods 16, 1153–1160 (2019).
Punjani, A., Zhang, H. & Fleet, D. J. Non-uniform refinement: adaptive regularization improves single-particle cryo-EM reconstruction. Nat. Methods 17, 1214–1221 (2020).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589 (2021).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D. Biol. Crystallogr. 66, 486–501 (2010).
Nicholls, R. A., Tykac, M., Kovalevskiy, O. & Murshudov, G. N. Current approaches for the fitting and refinement of atomic models into cryo-EM maps using CCP-EM. Acta Crystallogr. D. Struct. Biol. 74, 492–505 (2018).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D. Biol. Crystallogr. 66, 12–21 (2010).
Pettersen, E. F. et al. UCSF ChimeraX: structure visualization for researchers, educators, and developers. Protein Sci. 30, 70–82 (2021).
Goddard, T. D. et al. UCSF ChimeraX: meeting modern challenges in visualization and analysis. Protein Sci. 27, 14–25 (2018).
Talbot, N. J., Ebbole, D. J. & Hamer, J. E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell 5, 1575–1590 (1993).
Hartmann, T. et al. Validation of a self-excising marker in the human pathogen Aspergillus fumigatus by employing the β-Rec/ six site-specific recombination system. Appl. Environ. Microbiol. 76, 6313–6317 (2010).
Lambou, K., Lamarre, C., Beau, R., Dufour, N. & Latge, J. Functional analysis of the superoxide dismutase family in Aspergillus fumigatus. Mol. Microbiol. 75, 910–923 (2010).
Müller, F.-M. C. et al. Rapid extraction of genomic DNA from medically important yeasts and filamentous fungi by high-speed cell disruption. J. Clin. Microbiol. 36, 1625–1629 (1998).
Foster, A. J. et al. CRISPR-Cas9 ribonucleoprotein-mediated co-editing and counterselection in the rice blast fungus. Sci. Rep. 8, 14355 (2018).
Sikorski, R. S. & Boeke, J. D. [20] In vitro mutagenesis and plasmid shuffling: from cloned gene to mutant yeast. Methods Enzymol. 194, 302–318 (1991).
Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797 (2004).
Acknowledgements
We thank Dr. Y. Goto and Dr. H. Suga for providing the chemically synthesized Asp-DBE substrate and Dr. S. Friant for providing Sce erg∆ strains. Cryo-EM data were collected at the Cryo-EM facility of The University of Tokyo.
Funding
This work was supported by the “N-FLAMS” project from the Agence Nationale de la Recherche (ANR-20-CE44-0002), and by the Integrative Molecular and Cellular Biology (IMCBio) program as part of the Interdisciplinary Thematic Institutes (ITI) 2021–2028 initiative of the University of Strasbourg, CNRS, and INSERM, supported by IdEx Unistra (ANR-10-IDEX-0002) and EUR IMCBio (ANR-17-EURE-0023), under the framework of the French Investments for the Future Program (to N.Y., N.M., S.Z., B.S., F.F., and H.D.B.). Additional support was provided by the University of Strasbourg and CNRS (to N.Y., N.M., S.Z., B.S., F.F., and H.D.B.), by JSPS KAKENHI Grant Number 23KJ0722 (to H.M.), by JST CREST JPMJCR20E2 (to O.N.), by JSPS KAKENHI Grant Number 26H00007 (to O.N.), by NSF grant MCB-2448119 (to R.E.K.) and by NIH grant R35GM134931 (to Y.M.H.).
Author information
Authors and Affiliations
Contributions
H.M. performed structural analyses with assistance from M.N., Y.K., and Y.I. K.M. performed molecular dynamics simulations and analyzed the data. N.Y., N.M., N.F., and S.Z. conducted fungal genetics, phenotypic characterization, and biochemical assays, with assistance from B.S., F.F., and H.D.B. S.Z. and F.F. performed fluorescence microscopy. S.L. and M. F. performed the Nanolive imaging experiments. N.M. and L.S. also contributed equally to this work. L.H. performed PCR and cloning to generate overexpression constructs. H.B.G. and Y.M.H. prepared the Asp-tRNAAsp analog used for structural analysis. J.W.J. and R.E.K. performed mass spectrometry analysis of oligonucleotides, H.M., N.Y., H.D.B., Y.I., F.F., and O.N wrote the manuscript with input from all authors. F.F., Y.I., H.D.B., and O.N. supervised the research.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Lin Bai, Michael O’Connor, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Murayama, H., Yakobov, N., Mahmoudi, N. et al. Structure of ergosteryl-aspartate synthase reveals how an entrapped tRNA is used like a prosthetic swinging arm in the synthesis of aminoacylated sterols. Nat Commun 17, 4455 (2026). https://doi.org/10.1038/s41467-026-73135-8
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-73135-8
