
Abstract
The zeta inhibitory peptide (ZIP) interferes with memory maintenance and long-term potentiation (LTP)1 when administered to mice. However, mice lacking its putative target, protein kinase PKMζ, exhibit normal learning and memory as well as LTP2,3, making the mechanism of ZIP unclear. Here we show that ZIP disrupts LTP by removing surface AMPA receptors through its cationic charge alone. This effect requires endophilin-A2-mediated endocytosis and is fully blocked by drugs suppressing macropinocytosis. ZIP and other cationic peptides remove newly inserted AMPA receptor nanoclusters at potentiated synapses, providing a mechanism by which these peptides erase memories without altering basal synaptic function. When delivered in vivo, cationic peptides can modulate memories on local and brain-wide scales, and these mechanisms can be leveraged to prevent memory loss in a model of traumatic brain injury. Our findings uncover a previously unknown synaptic mechanism by which memories are maintained or lost.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others
Data availability
All data supporting the findings of this study are available within the paper and its Supplementary Information.
Code availability
Custom MATLAB code for analysing GluA1 puncta is available at Zenodo (https://doi.org/10.5281/zenodo.10199183)65 and code for analysis of STED and expansion microscopy data are available at GitHub (https://github.com/AotoLab/STED-and-ExM-Squassh-Analysis).
References
Pastalkova, E. et al. Storage of spatial information of LTP by the maintenance mechanism. Science 313, 1141–1144 (2006).
Lee, A. M. et al. Prkcz null mice show normal learning and memory. Nature 493, 416–419 (2013).
Volk, L. J., Bachman, J. L., Johnson, R., Yu, Y. & Huganir, R. L. PKM-ζ is not required for hippocampal synaptic plasticity, learning and memory. Nature 493, 420–423 (2013).
Kelly, M. T., Crary, J. F. & Sacktor, T. C. Regulation of protein kinase Mζ synthesis by multiple kinases in long-term potentiation. J. Neurosci. 27, 3439–3444 (2007).
Gao, P. P., Goodman, J. H., Sacktor, T. C. & Francis, J. T. Persistent increases of PKMζ in sensorimotor cortex maintain procedural long-term memory storage. iScience 5, 90–98 (2018).
Tsokas, P. et al. Compensation for PKMζ in long-term potentiation and spatial long-term memory in mutant mice. eLife 5, e14846 (2016).
Lin, J. & Alexander-Katz, A. Cell membranes open “doors” for cationic nanoparticles/biomolecules: insights into uptake kinetics. ACS Nano 7, 10799–10808 (2013).
Felgner, P. L. et al. Lipofection: a highly efficient, lipid-mediated DNA-transfection procedure. Proc. Natl Acad. Sci. USA 84, 7413–7417 (1987).
Kaplan, I. M., Wadia, J. S. & Dowdy, S. F. Cationic TAT peptide transduction domain enters cells by macropinocytosis. J. Control. Release 102, 247–253 (2005).
Sacktor, T. C. & Fenton, A. A. Appropriate application of ZIP for PKMζ inhibition, LTP reversal, and memory erasure. Hippocampus 22, 645–647 (2012).
Thompson, D. B., Villaseñor, R., Dorr, B. M., Zerial, M. & Liu, D. R. Cellular uptake mechanisms and endosomal trafficking of supercharged proteins. Chem. Biol. 19, 831–843 (2012).
Green, M. & Loewenstein, P. M. Autonomous functional domains of chemically synthesized human immunodeficiency virus tat trans-activator protein. Cell 55, 1179–1188 (1988).
Frankel, A. D. & Pabo, C. O. Cellular uptake of the tat protein from human immunodeficiency virus. Cell 55, 1189–1193 (1988).
Sadeh, N., Verbitsky, S., Dudai, Y. & Segal, M. Zeta inhibitory peptide, a candidate inhibitor of protein kinase M, is excitotoxic to cultured hippocampal neurons. J. Neurosci. 35, 12404–12411 (2015).
Serrano, P. et al. PKMζ maintains spatial, instrumental, and classically conditioned long-term memories. PLoS Biol. 6, 2698–2706 (2008).
Kopec, C. D., Li, B., Wei, W., Boehm, J. & Malinow, R. Glutamate receptor exocytosis and spine enlargement during chemically induced long-term potentiation. J. Neurosci. 26, 2000–2009 (2006).
Miesenbock, G., De Angelis, D. A. & Rothman, J. E. Visualizing secretionand synaptic transmission with pH-sensitive green fluorescent proteins. Nature 394, 192–195 (1998).
Bingor, A., Haham, T., Thornton, C., Stern-Bach, Y. & Yaka, R. Zeta inhibitory peptide attenuates learning and memory by inducing NO-mediated downregulation of AMPA receptors. Nat. Commun. 11, 3688 (2020).
Kawasaki-Nishi, S., Bowers, K., Nishi, T., Forgac, M. & Stevens, T. H. The amino-terminal domain of the vacuolar proton-translocating ATPase A subunit controls targeting and in vivo dissociation, and the carboxyl-terminal domain affects coupling of proton transport and ATP hydrolysis. J. Biol. Chem. 276, 47411–47420 (2001).
Banker, G. A. & Cowan, W. M. Rat hippocampal neurons in dispersed cell culture. Brain Res. 126, 397–425 (1977).
Turrigiano, G. G., Leslie, K. R., Desai, N. S., Rutherford, L. C. & Nelson, S. B. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature 391, 892–896 (1998).
Otmakhov, N. et al. Forskolin-induced LTP in the CA1 hippocampal region is NMDA receptor dependent. J. Neurophysiol. 91, 1955–1962 (2004).
Serrano, P., Yao, Y. & Sacktor, T. C. Persistent phosphorylation by protein kinase Mζ maintains late-phase long-term potentiation. J. Neurosci. 25, 1979–1984 (2005).
Crosby, K. C. et al. Nanoscale subsynaptic domains underlie the organization of the inhibitory synapse. Cell Rep. 26, 3284–3297.e3 (2019).
Swanson, J. A. & Watts, C. Macropinocytosis. Trends Cell Biol. 5, 424–428 (1995).
Casamento, A. & Boucrot, E. Molecular mechanism of fast endophilin-mediated endocytosis. Biochem. J. 477, 2327–2345 (2020).
Do-Monte, F. H., Quinõnes-Laracuente, K. & Quirk, G. J. A temporal shift in the circuits mediating retrieval of fear memory. Nature 519, 460–463 (2015).
Corcoran, K. A. & Quirk, G. J. Activity in prelimbic cortex is necessary for the expression of learned, but not innate, fears. J. Neurosci. 27, 840–844 (2007).
Sacco, T. & Sacchetti, B. Role of secondary sensory cortices in emotional memory storage and retrieval in rats. Science 329, 649–656 (2010).
Kourrich, S., Rothwell, P. E., Klug, J. R. & Thomas, M. J. Cocaine experience controls bidirectional synaptic plasticity in the nucleus accumbens. J. Neurosci. 27, 7921–7928 (2007).
Li, Y. et al. Inhibition of PKMz in nucleus accumbens core abolishes long-term drug reward memory. J. Neurosci. 31, 5436–5446 (2011).
Dumas, T. C. Late postnatal maturation of excitatory synaptic transmission permits adult-like expression of hippocampal-dependent behaviors. Hippocampus 15, 562–578 (2005).
Vives, E., Richard, J.-, Rispal, C. & Lebleu, B. TAT peptide internalization: seeking the mechanism of entry. Curr. Protein Pept. Sci. 4, 125–132 (2005).
Sloley, S. S. et al. High-frequency head impact causes chronic synaptic adaptation and long-term cognitive impairment in mice. Nat. Commun. 12, 2613 (2021).
Tye, K. M., Stuber, G. D., De Ridder, B., Bonci, A. & Janak, P. H. Rapid strengthening of thalamo-amygdala synapses mediates cue-reward learning. Nature 453, 1253–1257 (2008).
Everitt, B., Morris, K., O’Brien, A. & Robbins, T. The basolateral amygdala-ventral striatal system and conditioned place preference: further evidence of limbic-striatal interactions underlying reward-related processes. Neuroscience 42, 1–18 (1991).
Nabavi, S. et al. Engineering a memory with LTD and LTP. Nature 511, 348–352 (2014).
Stevens, C. F. A million dollar question: minireview does LTP = memory? Neuron 20, 1–2 (1998).
Compans, B., Choquet, D. & Hosy, E. Review on the role of AMPA receptor nano-organization and dynamic in the properties of synaptic transmission. Neurophotonics 3, 041811 (2016).
Hou, Q., Zhang, D., Jarzylo, L., Huganir, R. L. & Man, H.-Y. Homeostatic regulation of AMPA receptor expression at single hippocampal synapses. Proc. Natl Acad. Sci. USA 105, 775–780 (2008).
Sutton, G. & Chandler, L. J. Activity-dependent NMDA receptor-mediated activation of protein kinase B/Akt in cortical neuronal cultures. J. Neurochem. 82, 1097–1105 (2002).
Boucrot, E. et al. Endophilin marks and controls a clathrin-independent endocytic pathway. Nature 517, 460–465 (2015).
Triller, A. & Choquet, D. New concepts in synaptic biology derived from single-molecule imaging. Neuron 59, 359–374 (2008).
Choquet, D. & Triller, A. The dynamic synapse. Neuron 80, 691–703 (2013).
Tatavarty, V., Sun, Q. & Turrigiano, G. G. How to scale down postsynaptic strength. J. Neurosci. 33, 13179–13189 (2013).
Dong, Z. et al. Long-term potentiation decay and memory loss are mediated by AMPAR endocytosis. J. Clin. Invest. 125, 234–247 (2015).
Ge, Y. et al. Hippocampal long-term depression is required for the consolidation of spatial memory. Proc. Natl Acad. Sci. USA 107, 16697–16702 (2010).
Brebner, K. et al. Nucleus accumbens long-term depression and the expression of behavioral sensitization. Science 310, 1337–1340 (2005).
Buard, I. et al. CaMKII “autonomy” is required for initiating but not for maintaining neuronal long-term information storage. J. Neurosci. 30, 8214–8220 (2010).
Wu, Y. et al. Calmodulin kinase II is required for fight or flight sinoatrial node physiology. Proc. Natl Acad. Sci. USA 106, 5972–5977 (2009).
Temkin, P. et al. The retromer supports AMPA receptor trafficking during LTP. Neuron 94, 74–82.e5 (2017).
Uphoff, C. C. & Drexler, H. G. in Cancer Cell Culture: Methods and Protocols 2nd edn (ed. Cree, I. A.) Ch. 8 (Humana, 2011).
Aoto, J., Martinelli, D. C., Malenka, R. C., Tabuchi, K. & Südhof, T. C. Presynaptic neurexin-3 alternative splicing trans-synaptically controls postsynaptic AMPA receptor trafficking. Cell 154, 75 (2013).
Lloyd, B. A., Han, Y., Roth, R., Zhang, B. & Aoto, J. Neurexin-3 subsynaptic densities are spatially distinct from Neurexin-1 and essential for excitatory synapse nanoscale organization in the hippocampus. Nat. Commun. 14, 4706 (2023).
Yang, Y. et al. Endophilin A1 regulates dendritic spine morphogenesis and stability through interaction with p140Cap. Cell Res. 25, 496–516 (2015).
Restrepo, S., Langer, N. J., Nelson, K. A. & Aoto, J. Modeling a neurexin-3α human mutation in mouse neurons identifies a novel role in the regulation of transsynaptic signaling and neurotransmitter release at excitatory synapses. J. Neurosci. 39, 9065–9082 (2019).
Zhu, B., Eom, J. & Hunt, R. F. Transplanted interneurons improve memory precision after traumatic brain injury. Nat. Commun. 10, 5156 (2019).
Chen, Y. C., Mao, H., Yang, K. H., Abel, T. & Meaney, D. F. A modified controlled cortical impact technique to model mild traumatic brain injury mechanics in mice. Front. Neurol. 5, 100 (2014).
Purkey, A. M. et al. AKAP150 palmitoylation regulates synaptic incorporation of Ca2+-permeable AMPA receptors to control LTP. Cell Rep. 25, 974–987.e4 (2018).
Paul, G., Cardinale, J. & Sbalzarini, I. F. Coupling image restoration and segmentation: a generalized linear model/bregman perspective. Int. J. Comput. Vis. 104, 69–93 (2013).
Rizk, A. et al. Segmentation and quantification of subcellular structures in fluorescence microscopy images using Squassh. Nat. Protoc. 9, 586–596 (2014).
Frankowski, J. C. et al. Brain-wide reconstruction of inhibitory circuits after traumatic brain injury. Nat. Commun. 13, 3417 (2022).
Park, J. S. et al. Synthetic control of mammalian-cell motility by engineering chemotaxis to an orthogonal bioinert chemical signal. Proc. Natl Acad. Sci. USA 111, 5896–5901 (2014).
Stachniak, T. J., Ghosh, A. & Sternson, S. M. Chemogenetic synaptic silencing of neural circuits localizes a hypothalamus→midbrain pathway for feeding behavior. Neuron 82, 797–808 (2014).
Derdeyn, P. Quantifying number and density of neural puncta. Zenodo https://doi.org/10.5281/zenodo.10199183 (2023).
Acknowledgements
We thank K. Crosby, S. Olah and B. Lloyd for discussions on STED and expansion microscopy analysis, and M. Banghart and D. Thompson for discussions on the manuscript. This work was supported by the NIH (R00 DA041445, DP2 AG067666, R01NS130044, R01DA056599, 1R01DA054374, R01NS096012, F31NS132447), Tobacco Related Disease Research Program (T31KT1437, T31P1426), One Mind (OM-5596678), Brightfocus Foundation (A2022031S d), American Parkinson Disease Association (APDA-5589562), Alzheimer’s Association (AARG-NTF-20-685694), New Vision Research (CCAD2020-002), and Brain and Behavior Research Foundation (NARSAD 26845) to K.T.B.; 1F30DA056215 to M.H.; NIH R01 MH116901 and R21 MH129620 to J.A.; HHMI/Gilliam GT15852 to E.G.S.; and NIH (K99/R00GM126136, DP2GM150017) and Chan Zuckerberg Initiative (CZI) Advancing Imaging through Collaborative Projects award to Y.Z. and X.S. Y.Z. is also supported by an NSF-Simons grant, DMS1763272 (594598).
Author information
Authors and Affiliations
Contributions
Conceptualization: K.T.B. Investigation: K.T.B., E.G.S., J.J.V., G.A., M.N., A.T., Y.Z., V.M.G., M.T., I.T., J.A. Formal analysis: K.T.B., J.A. Visualization: K.T.B., M.H. Funding acquisition: K.T.B., J.A., X.S., R.F.H. Supervision: K.T.B., J.A., X.S., R.F.H. Writing—original draft: K.T.B. Writing—review and editing, K.T.B., J.A. and R.F.H.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Paul Frankland and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Concentration dependence of various cationic and non-cationic peptides on SEP-GluA1 fluorescence.
(a) Sample flow cytometry image indicating side scatter vs. forward scatter, and selection of gates to isolate cells. (b) Distribution of cells by FITC signal, normalized to the maximum number of cells found within a given FITC range bin. ZIP shifts the distribution of this curve to the left. (c) Effect of heparin on the ZIP-induced reduction in SEP-GluA1 fluorescence, added either to the same tube as ZIP before adding to cells, or added to the cell culture independently before ZIP. Heparin completely blocked ZIP or TAT’s effects on SEP-GluA1 fluorescence, whether heparin was premixed with peptide – vehicle vs. heparin/ZIP p = 0.39; vehicle vs. heparin/TAT p = 0.93 – or first added to the cells – vehicle vs. heparin/ZIP p = 0.17; vehicle vs. heparin/TAT p = 0.79. (d) Ratio of SEP-GluA1 internalization as a function of the log concentration of scrZIP and myr-scrZIP. scrZIP all n = 3, myr-scrZIP all n = 3. (e) Effect of 100 μM V5 or FLAG peptides relative to 100 μM ZIP on SEP-GluA1 fluorescence. Vehicle vs. V5 p = 0.87; vehicle vs. FLAG p < 0.0001. (f) Concentration dependence of the AIP peptide on SEP-GluA1 fluorescence. Vehicle vs. 10 μM AIP p < 0.0001; vehicle vs. 100 μM AIP p < 0.0001; vehicle vs. 1 mM AIP p < 0.0001. (g) Concentration dependence of the scrZIP peptide on SEP-GluA1 fluorescence. Vehicle vs. 0.1 μM scrZIP p = 0.32; vehicle vs. 1 μM scrZIP p < 0.0001; vehicle vs. 10 μM scrZIP p < 0.0001; vehicle vs. 100 μM scrZIP p < 0.0001; vehicle vs. 1 mM scrZIP p < 0.0001. (h) Concentration dependence of the Arg9 peptide on SEP-GluA1 fluorescence. Vehicle vs. 10 μM Arg9 p < 0.0001; vehicle vs. 100 μM Arg9 p < 0.0001. (i) Concentration dependence of the non-cationic AA3H and AA3H-PLP peptides on SEP-GluA1 fluorescence. Vehicle vs. 10 μM AA3H p < 0.0001; vehicle vs. 100 μM AA3H p < 0.0001; vehicle vs. 10 μM AA3H-PLP p < 0.0001; vehicle vs. 100 μM AA3H-PLP p < 0.0001. (j) Relationship between the net charge of each peptide and the relative change in SEP-GluA1 fluorescence (at 100 μM peptide), r2 = 0.70, p < 0.0001. Error bars are centered at the mean, ± 1 s.e.m., and full statistics are provided in Supplemental Table 1.
Extended Data Fig. 2 Effects of endocytosis-modulating drugs at different concentrations in HEK-SEP-GluA1 cells.
(a) Effects of various drugs on HEK cells without SEP-GluA1, relative to vehicle-treated HEK-SEP-GluA1 cells. Data are presented as mean fluorescence in the FITC channel per sample. (b) Effect of bafilomycin at different concentrations, with and without subsequent ZIP administration. As bafilomycin A1 canonically inhibits vacuolar H + -ATPase, which is the main proton pump responsible for endosome acidification, this likely prevents pH-induced changes in GFP fluorescence upon endocytosis. Vehicle (saline) vs. 1% DMSO p = 0.069; vehicle vs. 0.1 μM bafilomycin/ZIP p = 0.097; vehicle vs. 1 μM bafilomycin/ZIP p = 0.0007; vehicle vs. 0.1 μM bafilomycin p = 0.0023; vehicle vs. 1 μM bafilomycin p < 0.0001. This indicates our assay is working properly. (c) Dynasore (80 μM), chlorpromazine (5 μM) or nystatin (5 μg/μL) were applied to HEK SEP-GluA1 cells 4 h prior to ZIP or vehicle application. Dynasore and nystatin partially but incompletely blocked ZIP’s effects. Vehicle vs. Dynasore/ZIP p < 0.0001; vehicle vs. Dynasore p = 0.0012; vehicle vs. nystatin/ZIP p < 0.0001; vehicle vs. nystatin p < 0.0001. (d) Effect of chlorpromazine at different concentrations, with and without subsequent ZIP administration. Vehicle vs. 0.5 μM chlorpromazine/ZIP p < 0.0001; vehicle vs. 5 μM chlorpromazine/ZIP p < 0.0001; vehicle vs. 0.5 μM chlorpromazine p = 0.0026; vehicle vs. 5 μM chlorpromazine p < 0.0001. (e) Effect of Dynasore at different concentrations, with and without subsequent ZIP administration. Vehicle vs. 8 μM Dynasore + ZIP p = 0.03; vehicle vs. 80 μM Dynasore + ZIP p < 0.0001; vehicle vs. 8 μM Dynasore p = 0.0015; vehicle vs. 80 μM Dynasore p < 0.0001. (f) Effect of nystatin at different concentrations, with and without subsequent ZIP administration. Vehicle vs. 0.5 μg/mL nystatin + ZIP p < 0.0001; vehicle vs. 5 μg/mL nystatin + ZIP p < 0.0001; vehicle vs. 0.05 μg/mL nystatin p = 0.03; vehicle vs. 0.5 μg/mL nystatin p = 0.0003; vehicle vs. 5 μg/mL nystatin p < 0.0001. (g) Amiloride blocked ZIP’s effects at concentrations typically used to block macropinocytosis in cell culture, but not at lower concentrations. Vehicle vs. 40 μM amiloride/ZIP p < 0.0001; vehicle vs. 400 μM amiloride/ZIP p < 0.0001; vehicle vs. 4 mM amiloride/ZIP p = 0.16. (h) Effect of rottlerin at different concentrations, with and without subsequent ZIP administration. Vehicle vs. 50 μM rottlerin/ZIP p = 0.15; vehicle vs. 0.5 μM rottlerin p = 0.57; vehicle vs. 5 μM rottlerin p = 0.0051; Vehicle vs. 50 μM rottlerin, p < 0.0001. (i) Effect of Ly294002 at different concentrations, with and without subsequent ZIP administration. Vehicle vs. 0.5 μM Ly294002/ZIP p = 0.62; vehicle vs. 5 μM Ly294002/ZIP p = 0.01; vehicle vs. 50 μM Ly294002/ZIP p < 0.0001; vehicle vs. 0.5 μM Ly294002 p = 0.43; vehicle vs. 5 μM Ly294002 p < 0.0001; vehicle vs. 50 μM Ly294002 p < 0.0001. (j) Effect of EIPA at different concentrations, with and without subsequent ZIP administration. Vehicle vs. 40 μM EIPA/ZIP p = 0.01; vehicle vs. 400 μM EIPA/ZIP p = 0.0098; vehicle vs. 4 μM EIPA p < 0.0001; vehicle vs. 40 μM EIPA p < 0.0001; vehicle vs. 400 μM EIPA p < 0.0001. Error bars are centered at the mean, ± 1 s.e.m., and full statistics are provided in Supplemental Table 1.
Extended Data Fig. 3 Effect of positively charged and neutral molecules on SEP-GluA1 fluorescence.
(a) Effect of an equivalent concentration of free lysine (100 μM) and arginine (500 μM) as is present in 100 μM ZIP. A minor reduction in SEP-GluA1 fluorescence was observed. Vehicle vs. L-lys/L-arg p = 0.0003. (b) Effect of dextrans on SEP-GluA1 fluorescence. No change in SEP-GluA1 fluorescence was observed. Vehicle vs. 0.05 mg/mL p = 0.90; vehicle vs. 0.01 mg/mL p = 0.75; vehicle vs. 0.5 mg/mL p = 0.40; vehicle vs. 1 mg/mL p = 0.57; vehicle vs. 5 mg/mL p = 0.67. Error bars are centered at the mean, ± 1 s.e.m., and full statistics are provided in Supplemental Table 1.
Extended Data Fig. 4 Additional primary neuronal culture data describing metric used and demonstrating applicability for multiple cationic peptides.
(a) Confocal micrograph of representative neuronal structures for each condition selected for quantitative analysis. A GFP-based cell-fill used to highlight dendritic structure is shown in green. GluA1 immunostaining is shown in blue. Scale bar, 20 μm. (b) Density of GluA1 puncta for each condition (per 10 mm of dendrite). One-way ANOVA p = 0.42. (c) Mean intensity of GluA1 puncta for each condition (AU). One-way ANOVA p = 0.91. (d) Mean size of GluA1 puncta for each condition (mm2). One-way ANOVA p = 0.0017; control vs. TTX p = 0.0032. (e) Normalized integrated puncta intensity for each condition. One-way ANOVA p < 0.0001; Control vs. TTX p < 0.0001. (f) AIP induced GluA1 endocytosis that was blocked by amiloride. Vehicle vs. TTX p < 0.0001; Vehicle vs. TTX/amiloride/AIP p = 0.0004; TTX vs. TTX/AIP p = 0.019; TTX/AIP vs. TTX/amiloride/AIP p = 0.033. (g) scrZIP induced GluA1 endocytosis that was blocked by amiloride. Vehicle vs. TTX p < 0.0001; Vehicle vs. TTX/amiloride/scrZIP p < 0.0001; TTX vs. TTX/amiloride/scrZIP p < 0.0001; scrZIP vs. TTX/amiloride/scrZIP p < 0.0001; TTX/scrZIP vs. TTX/amiloride/scrZIP p < 0.0001. (h) Penetratin induced GluA1 endocytosis that was blocked by amiloride. Vehicle vs. TTX p < 0.0001; Vehicle vs. TTX/amiloride/penetratin p < 0.0001; TTX vs. TTX/amiloride/penetratin p < 0.0001; Penetratin vs. TTX/amiloride/penetratin p < 0.0001; TTX/penetratin vs. TTX/amiloride/penetratin p < 0.0001. (i) Arg9 induced GluA1 endocytosis that was blocked by amiloride. Vehicle vs. TTX p < 0.0001; Vehicle vs. TTX/amiloride/Arg9 p < 0.0001; TTX vs. TTX/Arg9 p = 0.0012; Arg9 vs. TTX/amiloride/Arg9 p = 0.046; TTX/Arg9 vs. TTX/amiloride/Arg9 p = 0.0009. Error bars are centered at the mean, ± 1 s.e.m., and full statistics are provided in Supplemental Table 1.
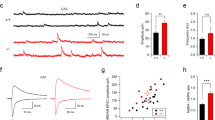
Extended Data Fig. 5 Effects of ZIP, TAT, and amiloride on EPSC frequency in primary cultured neurons.
(a) Cumulative probability graph for the inter-event interval for each condition. (b) Bar graph comparison of EPSC frequency for each condition. One-way ANOVA p = 0.011; no pairwise comparisons are significant. (c) Cumulative probability graph for the inter-event interval for each condition. (d) Bar graph comparison of EPSC frequency for each condition. One-way ANOVA p = 0.09. (e) Cumulative probability graph for the inter-event interval for each condition. (f) Bar graph comparison of EPSC frequency for each condition. One-way ANOVA p < 0.0001; control vs. TTX/amiloride p < 0.0001; control vs. TTX/amiloride/TAT p = 0.0003; TTX vs. TTX/amiloride p < 0.0001; TTX vs. TTX/amiloride/TAT p = 0.0034; TTX/amiloride vs. TTX/TAT p < 0.0001; TTX/TAT vs. TTX/TAT/amiloride p = 0.0001. Error bars are centered at the mean, ± 1 s.e.m., and full statistics are provided in Supplemental Table 1.
Extended Data Fig. 6 Electrophysiological properties of TTX, ZIP, TAT, and amiloride-treated cultured neurons.
(a) Membrane resistance of ZIP and/or TTX-treated cultures. (b) Membrane capacitance of ZIP and/or TTX-treated cultures. (c) Sample EPSCs of each of the ZIP and/or TTX-treated cultures. (d) Weighted EPSC decay constant for ZIP and/or TTX-treated cultures. (e) Membrane resistance of TAT and/or TTX-treated cultures. (f) Membrane capacitance of TAT and/or TTX-treated cultures. (g) Sample EPSCs of each of the TAT and/or TTX-treated cultures. (h) Weighted EPSC decay constant for TAT and/or TTX-treated cultures. (i) Membrane resistance of TAT, amiloride, and/or TTX- treated cultures. (j) Membrane capacitance of TAT, amiloride, and/or TTX-treated cultures. (k) Sample EPSCs of each of the TAT, amiloride, and/or TTX- treated cultures. (l) Weighted EPSC decay constant for TAT, amiloride, and/or TTX-treatment groups. One-way ANOVA p < 0.0001; control vs. TTX/amiloride p < 0.0001; control vs. TTX/amiloride/TAT p = 0.0002; TTX vs. TTX/amiloride p < 0.0001; TTX vs. TTX/amiloride/TAT p = 0.0007; TTX/amiloride vs. TTX/TAT p < 0.0001; TTX/TAT vs. TTX/amiloride/TAT p = 0.0032. All other non-noted comparisons in this figure were non-significant. Error bars are centered at the mean, ± 1 s.e.m., and full statistics are provided in Supplemental Table 1.
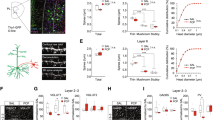
Extended Data Fig. 7 Investigation of effects of cationic peptides on GluA1 nanoclusters using expansion microscopy.
(a) Schematic of Expansion Microscopy. (b) Sample Expansion Microscopy confocal images of Homer1, GluA1, and merged for control, TTX, ZIP, TTX/ZIP, TAT, TTX/TAT. (c) Total volume occupied by GluA1 in the synapse. One-way ANOVA p < 0.0001; control vs. TTX p < 0.0001; TTX vs. ZIP p < 0.0001; TTX vs. TTX/ZIP p < 0.0001; TTX vs. TAT p < 0.0001; TTX vs. TTX/TAT p < 0.0001. (d) Numbers of GluA1 nanoclusters per synapse. One-way ANOVA p < 0.0001; control vs. TTX p < 0.0001; TTX vs. ZIP p < 0.0001; TTX vs. TTX/ZIP p < 0.0001; TTX vs. TAT p < 0.0001; TTX vs. TTX/TAT p < 0.0001. (e) Volume of GluA1 nanoclusters. One-way ANOVA p = 0.0056; control vs. TAT p = 0.0032. Error bars are centered at the mean, ± 1 s.e.m., and full statistics are provided in Supplemental Table 1.
Extended Data Fig. 8 Histograms of GluA1 and GABARγ2 (GABAAR) volumes following ZIP application.
(a) Histogram of GluA1 volume in the synapse for control, TTX, ZIP, and TTX/ZIP conditions. (b) Histogram of GluA1 volume in the synapse for control, TTX, TAT, and TTX/TAT conditions. (c) Histogram of GABAAR nanocluster volume for control, PTX, ZIP, and PTX/ZIP conditions. (d) Histogram of GABAAR nanocluster volume for control, PTX, TAT, and PTX/TAT conditions.
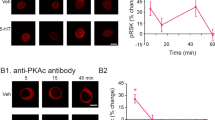
Extended Data Fig. 9 ZIP’s removal of AMPAR nanoclusters is dependent on endoA2.
(a) shRNA-mediated knockdown efficiency of endoA2 mRNA. (b) Sample STED microscopy images of Homer1, GluA1, and merged for control, TTX, TTX/ZIP, shRNA, shRNA/TTX/ZIP. (c) Schematic for hypothesized mechanism of action. TTX induces homeostatic plasticity, largely through increasing the number of nanoclusters. Cationic peptides trigger remodeling of the membrane through endoA2-mediated endocytosis, which is activated only upon cationic peptide-mediated stimulation. This preferentially removes newly inserted AMPAR nanoclusters.
Extended Data Fig. 10 Effect of peptide injection on auditory fear conditioning recall, as a function of the number of cationic charges on the peptide.
Saline n = 15, V5 n = 8, FLAG n = 10, AIP n = 7, ZIP n = 8, scrZIP n = 9, TAT n = 7, Arg9 n = 6.
Extended Data Fig. 11 Additional behavioral data detailing effects of cationic peptides on memory.
(a) Schematic of behavioral experiments testing whether TAT-mediated memory loss was permanent or not. (b) Following TAT injection, recall memory was impaired in the first and all subsequent recall sessions. Two-way ANOVA group factor p = 0.0004. (c) ZIP or TAT infusion impaired recall of the tone/shock association, and animals were able to re-learn this association equally well as during the first learning period. (d) Comparison of the time spent freezing during the third tone/shock association before and after ZIP or TAT administration. ZIP p = 0.61; TAT p = 0.33. (e) Bafilomycin administration had no effect on ZIP-mediated memory disruption. ZIP vs. bafilomycin/ZIP p = 0.95. All statistical comparisons are provided in Supplementary Table 1. Note that 1 point for the bafilomycin group is above the y-axis. (f) Clathrin-mediated endocytosis inhibitor chlorpromazine did not impair ZIP-mediated memory disruption. ZIP vs. ZIP/chlorpromazine p > 0.99. (g) Clathrin-mediated endocytosis inhibitor Dynasore did not significantly impair ZIP-mediated memory disruption. ZIP vs. ZIP/Dynasore p = 0.67. (h) The caveolin-mediated endocytosis inhibitor nystatin did not significantly impair ZIP-mediated memory disruption. ZIP vs. ZIP/nystatin p = 0.85. (i) A combination of bafilomycin, chlorpromazine, Dynasore, and nystatin had no effect on ZIP-mediated memory disruption. ZIP vs. 4 drug cocktail p = 0.91. (j) Administration of positively charged amino acids did not disrupt recall of the tone/shock association. Saline vs. AAs p = 0.029. Note that 2 points for the +AAs are above the y-axis. Error bars are centered at the mean, ± 1 s.e.m., and full statistics are provided in Supplemental Table 1.
Supplementary information
Supplementary Table 1 (download XLSX )
Detailed statistics for all data reported in this manuscript.
Supplementary Table 2 (download XLSX )
Information about peptides used in this manuscript, including sequence, net charge and molecular weight.
Supplementary Table 3 (download XLSX )
Information about reagents used in this manuscript.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Stokes, E.G., Vasquez, J.J., Azouz, G. et al. Cationic peptides cause memory loss through endophilin-mediated endocytosis. Nature 638, 479–489 (2025). https://doi.org/10.1038/s41586-024-08413-w
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41586-024-08413-w
This article is cited by
-
Bending the boundaries: the many facets of endophilin-As from membrane dynamics to disease
Cellular and Molecular Life Sciences (2025)
