
Abstract
Sequential immunization is a promising approach to elicit broadly neutralizing antibodies (bNAbs) against the HIV-1 Envelope (Env). However, available protocols are inefficient and involve multiple immunizations over long periods of time. Here, we present WIN332, a new engineered Env immunogen that induces a new class of Asn332-glycan-independent antibodies to the conserved V3-glycan epitope of Env with low inhibitory activity indicative of a neutralization activity after a single bolus immunization in nonhuman primates. WIN332 binds to precursors of canonical human Asn332-glycan-dependent (type-I) V3-glycan bNAbs but also of a first-of-its-class Asn332-glycan-independent (type-II) V3-glycan bNAb. A single immunization elicits low inhibitory serum and monoclonal antibodies that are boosted and affinity matured with a heterologous immunogen. Electron microscopy polyclonal epitope mapping analysis of serum antibodies, antibody cloning and cryogenic electron microscopy analysis reveals that WIN332 elicits Asn332-glycan-independent antibodies with striking sequence and binding similarities with the most potent human type-I and type-II V3-glycan bNAbs. Thus, WIN332 is a promising vaccine candidate to streamline V3-glycan bNAb elicitation.
Similar content being viewed by others

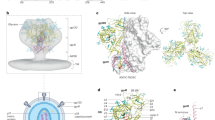
Main
Sequential immunization using a series of engineered and native-like HIV-1 Env proteins is a promising approach to eliciting bNAbs against HIV-1 (ref. 1). Sequential immunization aims to activate naive B cells expressing bNAb precursor antibodies and subsequently induce affinity maturation to increase antibody neutralization potency and breadth. To date, only two sequential immunization protocols designed to induce antibodies against the fusion peptide and CD4 binding site of Env have consistently induced bNAbs in nonhuman primates (NHPs)2,3. However, the elicited responses showed limited potency and breadth and required multiple immunizations over long periods of time.
Different bNAb lineages have different constraints for maturation. The activation of ‘premium’ antibody lineages that require less maturation to acquire neutralization potency and breadth would simplify sequential immunization protocols. In this regard, the conserved V3-glycan epitope of Env is the most common specificity for broadly neutralizing activity in studies of cohorts with bNAb responses4,5. The V3-glycan epitope comprises a peptide motif, GDIR and several surrounding glycans including those at positions Asn133, Asn137, Asn156, Asn301 and Asn332 (ref. 6). Interaction with the Asn332 glycan is a defining signature of V3-glycan bNAbs7,8,9 and a requirement for V3-glycan bNAbs to neutralize HIV-1, thus all currently available immunogens to elicit V3-glycan bNAbs are designed to retain this glycan10,11,12,13,14. However, whether Asn332-glycan dependency is intrinsic to the unmutated precursors of these bNAbs or alternatively Asn332-glycan dependency develops through somatic hypermutation (SHM) of Asn332-glycan-independent precursors is unknown. Supporting the latter, the Asn332 glycan is underrepresented in transmitted/founder (T/F) viruses15, and infection with Asn332-glycan-deficient T/F viruses led to the development of Asn332-glycan-dependent V3-glycan bNAbs in two humans15 suggesting that Asn332-glycan-deficient Env proteins may have played a role in Asn332-glycan-dependent bNAb lineage activation. Thus, we hypothesized that Asn332-glycan dependency develops during bNAb maturation and that unmutated precursors and early intermediates of V3-glycan bNAbs may not require this glycan for binding and activation. We further hypothesized that Asn332-glycan-independent antibody lineages can mature into Asn332-dependent (type-I) and Asn332-glycan-independent (type-II) V3-glycan bNAbs.
Notably, two new human V3-glycan bNAbs, EPTC112 and 007, have been recently isolated from HIV-infected individuals16,17. EPTC112 and 007 target the V3-glycan epitope of Env; however, their cryogenic electron microscopy (cryo-EM) structures showed no contacts with the Asn332 glycan. 007 is among the most potent V3-glycan bNAbs isolated to date17. These bNAbs are, therefore, evidence that Asn332-glycan-independent bNAbs can develop in humans.
Here, we present WIN332, an Asn332-glycan-deficient priming immunogen rationally designed to broadly activate precursors of both types of V3-glycan bNAbs, Asn332-glycan dependent (type-I) and Asn332-glycan independent (type-II). WIN332 prime elicited antibodies that closely resembled human type-I and type-II V3-glycan bNAbs in NHPs. Remarkably, one single bolus immunization with WIN332-elicited serum and monoclonal antibodies (mAbs) that neutralized wild-type fully glycosylated viruses and were further boosted by a heterologous boosting immunogen. WIN332 accelerated the development of neutralization by activating a new class of ‘premium’ Asn332-glycan-independent antibody lineages and arises as a promising vaccine candidate to simplify sequential immunization protocols against HIV-1.
Results
WIN332, an HIV-1 Env immunogen for broad elicitation of V3-glycan antibodies
WIN332 is an engineered native-like Env SOSIP trimer derived from the Env protein of the clade A/E HIV-1 strain BG505 (ref. 18; Fig. 1a). To design WIN332, we reevaluated the canonical definition of the V3-glycan epitope. We hypothesized that antibody recognition of the V3-glycan epitope may not always require interaction with the Asn332 glycan and that Asn332-glycan-independent antibody lineages can be activated and matured through immunization. Supporting this hypothesis, two studies have recently reported two human Asn332-glycan-independent bNAbs, EPTC112 (ref. 16), and 007 (ref. 17), which confirm that this class of antibodies is elicited in humans. Thus, here we considered two subtypes of V3-glycan antibody lineages: type-I, Asn332-glycan dependent7,8,19,20,21 and type-II, Asn332-glycan independent16. To test our hypothesis, we designed WIN332, a derivative of the previously described RC1 priming immunogen10 that lacks the signature potential N-linked glycosylation site (PNGS) at position Asn332 in addition to those previously removed in RC1 (Asn133, Asn137 and Asn156; Fig. 1a and Extended Data Table 1). WIN332 also naturally lacks a glycan at position Asn334, as it is derived from BG505 (ref. 18).

a, Top images show the three V3-glycan epitopes of the native-like BG505 and WIN332 Env trimers on a top view of the trimers. Bottom images show closer views of one of the V3-glycan epitopes of BG505 (left) and WIN332 (right). Env gp120 is shown in gray, glycans are represented with colored spheres, and GDIR is shown in salmon. Pie charts including colored slices are used for schematic representation of the V3-glycan epitopes. White slices indicate absent glycans. b, Representative BLI (OCTET) curves showing binding of WIN332 to the indicated iGL and UCA human bNAb precursors. Fabs were tested at 20 µM, 10 µM, 5 µM and 2.5 µM. c, Immunization protocol in NHPs. LN, Lymph node biopsy. d, ELISA results showing binding of the serum collected from NHPs before immunization (naive) and 3 weeks after one immunization with WIN332 (Prime; n = 4) to WIN332 (P = 0.0015). The human V3-glycan bNAb, PGT121, was used as a positive control at 5 µg ml−1. e, ELISA results showing the binding of the serum from WIN332-primed NHPs (n = 4) to WIN332 and a version of WIN332 with GAIA mutations in the GDIR motif (WIN332-GAIA; P = 0.0153). f, Endpoint titers (n = 4) for ELISA in d. g, ELISA results showing the binding of the serum from WIN332-primed NHPs (n = 4) to Env proteins with serially more native and glycosylated V3-glycan epitopes. h, Results of competition ELISAs using the serum from WIN332-immunized NHPs (n = 4) on the indicated Env proteins (P = 0.0052, 0.0036, 0.0037, 0.0082, 0.0206, 0.0673 and 0.0284, respectively). Human bNAbs PGT121 (V3-glycan epitope) and 3BNC117 (CD4bs) were used as competing antibodies. i, ELISA results showing binding of the macaque serum (n = 4) to different native-like Env trimers of different clades. Data were analyzed using two-sided Student’s t-test, presented as mean values + s.d. and considered statistically significant at *P ≤ 0.05 and **P ≤ 0.01. AUC, area under the curve. Illustrations in c were created using BioRender.com.
WIN332 elutes from size exclusion chromatography (SEC) columns at the expected fractions for gp140 trimers (Extended Data Fig. 1a) and shows the expected propeller shape by negative staining electron microscopy (Extended Data Fig. 1b). Glycan site occupancy for each PNGS was determined by mass spectrometry confirming the absence of glycosylation at Asn133, Asn137, Asn156 and Asn332 and the presence of glycosylation at Asn301 (Extended Data Fig. 1c). WIN332 shows high binding to human bNAbs targeting the CD4 binding site (CD4bs; VRC01), V1V2 (PGDM1400 and PGT145), V3-glycan (PGT121 and BG18) and the gp120–gp41 interface (PGT151) epitopes and undetectable or low binding to non-neutralizing antibodies including 3074, A32, F105, F425, 4025 and 17b (Extended Data Fig. 1d).
We conclude that WIN332 is a well-formed SOSIP trimer that retains the relevant antigenic properties of the HIV-1 Env.
WIN332 binds to unmutated precursors of type-I and type-II V3-glycan bNAbs
To evaluate whether WIN332 could activate precursors of known human V3-glycan bNAbs and initiate bNAb maturation, we interrogated the binding of WIN332 to inferred germline (iGL) precursors and unmutated common ancestors (UCAs) of human type-I and type-II V3-glycan bNAbs by biolayer interferometry (BLI; Fig. 1b). Interestingly, despite the absence of the Asn332 glycan, WIN332 showed quantifiable affinity for the precursors of human type-I bNAbs PGT121 (KD = 9.4 × 10−2 M), BG18 (KD = 4.5 × 10−4 M), DH270 (KD = 4.6 × 10−5 M), PCDN76 (KD = 2.7 × 10−5 M) and BF520 (KD = 1.5 × 10−6 M). Moreover, WIN332 showed quantifiable affinity for the precursor of the type-II EPTC112 (KD = 1 × 10−4 M) bNAb. We observed a wide range of affinities for the different human bNAb precursors. BF520 UCA showed an affinity of 1.493 × 10−6 M for WIN332 despite the BF520 bNAb being Asn332-glycan dependent. To investigate the role of the Asn332 glycan in the interaction of BF520 UCA with WIN332, we performed BLI experiments to compare the affinities of BF520 UCA to WIN332 and RC1 (WIN332 + Asn332 glycan)10. BF520 UCA showed an affinity of 1.48 × 10−5 M for RC1 (Extended Data Fig. 1e). Thus, BF520 UCA shows higher affinity to WIN332 than to RC1, indicating that the absence of the Asn332 glycan facilitates binding to Env.
To evaluate whether the low affinity of WIN332 for precursors of type-I bNAbs is biologically relevant, we immunized immunoglobulin knockin mice expressing the iGL PGT121 antibody. WIN332 was able to initiate antibody responses in these mice, confirming that the low affinity for this precursor is sufficient to activate B cells in vivo (Extended Data Fig. 1f).
We demonstrate that WIN332 binds to precursors of V3-glycan bNAbs from both Asn332-glycan-dependent (type-I) and Asn332-glycan-independent (type-II) subtypes, suggesting that WIN332 could activate these bNAb precursors in vivo.
WIN332 elicits Asn332-glycan-independent V3-glycan antibodies in NHPs
To characterize the ability of WIN332 to broadly elicit V3-glycan antibodies, we immunized rhesus macaques with one subcutaneous bolus injection of WIN332 in SMNP adjuvant22 distributed between the four limbs. Three and seven weeks after immunization, blood and draining lymph node (dLN) biopsy samples were collected (Fig. 1c). ELISA analysis of the serum from naive and immunized macaques revealed a robust antibody response to WIN332 in the immunized macaques (average endpoint titer: 956; Fig. 1d–f). The serum from WIN332-primed NHPs showed strongly reduced binding to a version of WIN332 with a mutated GDIR (WIN332-GAIA), indicating that the serum antibodies require an intact GDIR motif for binding to WIN332 (Fig. 1e and Extended Data Fig. 1g). The week-3 serum showed comparable binding to WIN332 and RC1, indicating that the serum antibodies could accommodate the Asn332 glycan. In addition, the serum showed gradually reduced binding to WIN332 variants with serially more native and glycosylated V3-glycan epitopes, including low but detectable binding to the fully glycosylated BG505 SOSIP1,11 (Engineering: WIN332 > RC1 > 11MutB > 10Mut > 7Mut > 5Mut > BG505; Fig. 1g and Extended Data Table 1).
To map the serologic response, we performed competition ELISAs on WIN332 using the V3-glycan bNAb, PGT121 and the CD4bs bNAb, 3BNC117 as competing antibodies (Fig. 1h). Binding of the serum to WIN332 was significantly reduced (41.2%) in the presence of PGT121, suggesting that a fraction of the serum antibodies targeted the V3-glycan epitope. Competition by PGT121 was also detectable on the serially more native RC1, 11MutB, 10Mut, 7Mut and native BG505 Env proteins (Fig.1h). The serum from week 7 bound to a panel of native multi-clade trimers, including BG505 Thr332Asn, CH119, Q23.17, AD8, CNE55, 001428, CE0217 and BJOX002000 (Fig. 1i).
To further confirm the presence of V3-glycan antibodies in the serum of WIN332-immunized macaques, we performed electron microscopy polyclonal epitope mapping (EMPEM). EMPEM analysis using WIN332 (Fig. 2a and Extended Data Fig. 2a,b) and RC1 (Fig.2b) revealed the presence of antibodies targeting the V3-glycan epitope in the serum of the three macaques independently analyzed. Overlapping the electron density of WIN332–Fab and RC1–Fab complexes with the WIN332 and RC1 structures, respectively10, confirmed V3-glycan specificity (Fig. 2c,d). EMPEM analysis using RC1 confirmed the presence of V3-glycan antibodies that could accommodate the Asn332 glycan (Fig. 2b,d,e–g). Comparison of the RC1–Fab and WIN332–Fab electron densities with published structures of Env-bound BG18, PGT121 and EPTC112 (refs. 12,16,23) showed structural alignment with all three antibodies (Fig. 2e–g and Extended Data Fig. 2c,d). In addition to V3-glycan antibodies, we detected Fabs binding the base of the Env trimer and the gp120–gp41 interface (Fig. 2a,b).

a,b, EMPEM results showing the binding of Fabs (colored) purified from the serum of WIN332-primed NHP 1 and 3 to WIN332 (gray; a) and NHP 1, 2 and 3 to RC1 (gray; b). c, Close-up views highlighting the interactions of the Fabs from NHP 1 and 3 (blue and red) with the Asn160 glycan of WIN332. WIN332 Env density is shown in gray with the model of RC1 (Protein Data Bank (PDB) 6ORO) with Asn332 glycan removed fit in (RC1 trimer ΔAsn332 glycan). d, Densities of NHP 1 Fab (green) bound to RC1 (gray) with RC1 model fit in (PDB 6ORO). Close-up views highlighting accommodation of Asn332 and Asn301 glycans on RC1. e–g, Comparison of Fab densities from NHP 1 (green), NHP 2 (dark blue) and NHP 3 (purple) with EPTC112 (pink), BG18 (orange) and PGT121 (cyan) Fab densities. h, Representative results of TZM-bl neutralization assays using purified immunoglobulins from the serum of WIN332-primed NHPs (n = 4) against pseudoviruses carrying gradually more native-looking Env proteins. 5MUT and BG505 (BG505 Thr332Asn) are fully glycosylated pseudoviruses. BG505 Asn301Ala is a modified version of the BG505 pseudovirus with a mutation that knocks out the V3-glycan epitope by removing the Asn301 glycan. Pie charts represent the V3-glycan epitope, and colored slices indicate present glycans.
We demonstrate that one immunization with WIN332 elicits serologic antibodies targeting the V3-glycan epitope in an Asn332-glycan-independent manner that cross-react with sequentially more native-like Envs in NHPs.
WIN332 elicits neutralizing serologic responses in NHPs
To evaluate the capacity of WIN332-elicited serum antibodies to neutralize HIV-1, we purified immunoglobulins from the serum from week 7 and tested them in TZM-bl assays against a panel of pseudoviruses including WIN332, RC1, 11MutB, 10Mut, 7Mut, 5Mut, fully glycosylated BG505 (BG505 Thr332Asn), a V3-glycan epitope knockout (BG505 Asn301Ala) and the murine leukemia virus (MLV) as a control (Fig. 2h and Extended Data Fig. 2e). Strong neutralization activity was observed against WIN332 and RC1 and gradually reduced activity against 11MutB, 10Mut, 7Mut and 5Mut. Remarkably, the serum showed low but detectable and reproducible inhibitory activity indicative of neutralization against the autologous fully glycosylated BG505 pseudovirus (Fig. 2h). No detectable activity was observed against the V3-glycan knockout pseudovirus (BG505 Asn301Ala) and MLV control, suggesting that the serum neutralizing response was entirely immunofocused to the V3-glycan epitope (Fig. 2h and Extended Data Fig. 2e).
We conclude that one immunization with WIN332 elicits serum Asn332-glycan-independent V3-glycan antibodies that neutralize autologous BG505 pseudovirus in NHPs.
WIN332 elicits Asn332-glycan-independent antibodies that resemble type-I and type-II human V3-glycan bNAbs in NHPs
To further characterize the V3-glycan antibodies elicited by WIN332 in NHPs, we isolated single WIN332-specific or BG505-specific germinal center B cells from dLNs collected 3 weeks after WIN332 immunization and cloned their immunoglobulin genes (Fig. 3a and Supplementary Fig. 1). Analysis of the immunoglobulin genes revealed extensive B cell clonal expansion (Fig. 3b). The average number of SHMs was 5.7 nucleotides for VH, 2.8 for VK and 6.1 nucleotides for VL genes (Fig. 3c). The average CDRH3 length was 14.4 amino acids ranging from 7 to 26 amino acids (Fig. 3d).

a, Representative flow cytometry plot showing WIN332-binding B cells in the dLNs of WIN332-primed NHPs. b, Pie charts show clonal expansion of WIN332-specific B cells isolated 3 weeks after WIN332 prime from NHP 2 and NHP 3. Heavy chains (HC), kappa light chains (KC) and lambda light chains (LC) are shown in purple, green and blue, respectively. White slices correspond to singlet sequences. The number in the middle of the pie chart indicates the number of sequences analyzed. c, Nucleotide SHMs in HC, KC and LC sequences from b. d, CDRH3 length of sequences from b. e, Pie charts show the frequency of sequences with CDR3 motifs of different human type-I and type-II V3-glycan bNAbs. Data for HC, KC and LC are shown in purple, green and blue, respectively. f, Graphs show the diversity and frequency of genes used by sequences from e. g, Heat map summarizes results of screening ELISAs to determine the binding of 43 WIN332-binding mAbs to the gradually more glycosylated Envs RC1, 11MutB and native BG505. Darker blue indicates higher ELISA binding. PGT121 was used as a positive control. h, Graph summarizes the ELISA binding in g, including the binding to WIN332-GAIA. BG505-binding antibodies are marked in pink in h–j. i, V3-glycan specificity of antibodies in g as determined by competition ELISAs on WIN332 using PGT121 (V3-glycan bNAb) and 3BNC117 (CD4bs bNAb) as competing antibodies. j, Affinities of antibodies in g for WIN332. OD, optical density.
Around 73% of the lambda light chains (IgL) presented CDRL3 motifs previously identified in human type-I and type-II V3-glycan bNAbs. These motifs included the ‘QxDSS’ motif of type-I PGT121/10-1074/BG18 bNAbs7,21 and of previously reported NHP V3-glycan antibodies10 and the ‘SYxG’ motif of the type-II EPTC112 bNAb (ref. 16; Fig. 3e). Around 43% of the kappa light chains (IgK) presented CDRL3 motifs of human bNAbs including DH1030, BF520 and PCDN.76 (refs. 24,25). CDRH3 analysis showed that 12% of the 392 sequences analyzed had the key ‘SxW’ motif of EPTC112 and 10% had the ‘SxYY’ motif of DH270. In addition, we identified CDRH3s with the ‘RxY’ motif of PGT121, the FGVV motif of BG18 and the ‘WSG’ motif of AIIMS-P01 and PCDN76 (refs. 7,9,12,16,19,25,26,27; Fig. 3e and Extended Data Table 2). Antibodies with these motifs use diverse VH, VK and VL genes (Fig. 3f)
We conclude that WIN332 elicits antibodies that resemble human type-I and type-II V3-glycan bNAbs in NHPs.
Macaque mAbs elicited by WIN332
We produced a selection of 43 mAbs that had signature motifs of human V3-glycan bNAbs (Extended Data Table 3). To characterize the binding of these mAbs, we performed a battery of ELISAs against different engineered and native Envs (Fig. 3g,h). The ELISA results showed that 43/43 antibodies bound WIN332, 40/43 WIN332-binding antibodies bound RC1, which reintroduces the Asn332 glycan, and 32/40 bound to 11MutB, which reintroduces the Asn156 glycan. Thirteen mAbs bound to the fully glycosylated BG505 SOSIP trimer (Fig. 3g,h). We performed competition ELISAs on WIN332 using the V3-glycan bNAb, PGT121 and the CD4bs bNAb, 3BNC117 as competing antibodies. PGT121 competed with the binding of 30/43 mAbs with an average competition of 74.8% and ranging from 50% to 90% (Fig. 3i). Moreover, mAbs showed significantly reduced binding to WIN332 with a GDIR motif mutated to GAIA (WIN332-GAIA; Fig. 3h) further supporting V3-glycan epitope specificity. The affinity of the 43 WIN332-binding mAbs for WIN332 had an average KD of 10−9 M (Fig. 3j).
We conclude that WIN332 elicits mAbs that cross-react with the fully glycosylated BG505 trimer and are competed by the V3-glycan bNAb PGT121.
WIN332 elicits neutralizing BG18-like antibodies in NHPs
We selected two NHP mAbs for further characterization. Ab1999 was selected for its high similarity with the human type-I V3-glycan bNAb, BG18. The HC of Ab1999 has a 76.7% amino acid identity with the HC of iGL BG18 and 60.4% with the mature BG18 HC (Fig. 4a and Extended Data Fig. 3a). Ab1999 has the defining ‘FGVV’ CDRH3 motif of BG18-like antibodies, which is involved in the binding and neutralization activity of BG18 (refs. 7,28). Ab1999 uses macaque VH4-106, DH3-3, JH5-1, VK1-74 and JK2*01 genes and has a 22-amino-acid-long CDRH3, only one amino acid shorter than BG18 (Fig. 4a). Ab1999 uses a kappa LC with 65.8% amino acid homology with the kappa LC of PCDN.76 UCA (Extended Data Fig. 3b). It has 2 and 14 nucleotide SHMs in the VH and VK genes, respectively (Fig. 4a, Extended Data Fig. 3a and Extended Data Table 3).

a, Alignment of the amino acid sequences of Ab1999 and iGL BG18 HC. Dots represent amino acids that are different between sequences. The key TxFGVV CDRH3 motif for neutralization and binding of BG18 is highlighted in pink. Kabat nomenclature is used. b, ELISA binding of Ab1999 to the indicated engineered Envs. c, ELISA binding of Ab1999 to WIN332 and two mutant versions of WIN332 that abrogate the V3-glycan epitope: WIN332glycanKO, which differs from WIN332 in that it lacks the N301 glycan; and WIN332-GAIA with a mutated GDIR motif. PGT121, BG18 and 3BNC117 were used as controls in b and c at 5 µg ml−1. Data are representative of at least two independent experiments (P = 0.0207). d, BLI (OCTET) curves showing binding of Ab1999 to fully glycosylated native-like BG505 Env (BG505 Thr332Asn). e, Results of TZM-bl assays showing neutralization activity of Ab1999 against pseudoviruses carrying Envs with the gradually more native V3-glycan epitopes WIN332, RC1, 11MutB, 10Mut and 5Mut. Data are representative of at least two independent experiments. f, Results of TZM-bl assays showing neutralization of Ab1999 against the fully glycosylated BG505 pseudovirus and a mutant version of this that abrogates the V3-glycan epitope (BG505 Asn301Ala). MLV was used as a negative control in e and f. Data are representative of at least two independent experiments. g, Atomic model of the RC1 Env–Ab1999 complex, showing interactions between Env (gp120 in blue, gp41 in gray) and Ab1999’s variable domains (salmon and pink). h, Cryo-EM density map of the RC1 Env trimer (gp120 in blue and gp41 in gray) bound to a single Ab1999 Fab (resolution is 3.9 Å). Three views highlight the Fab’s orientation and binding site. i, A close-up view highlights the Fab’s complementarity-determining regions (CDRs). Gp120 is shown as surface with contact residues colored according to Ab1999 CDR loop. j, Overview of the interactions of Ab1999 with the V1 (green), V2 (olive) and V3 (pink) loops. k–n, Detailed views of interactions with each loop including h-bonds and hydrophobic interactions. TxFGVV motif is colored in magenta. Antibody residues are numbered using Kabat nomenclature. k, h-bonds (dashed lines) involved in V1 loop contacts with Ab1999 Fab. l, Hydrophobic interactions on the V1 loop. m, Van der Waal’s contacts on the V2 loop. n, Hydrophobic core formed with the tip of TxFGVV motif. o, Comparison of the binding interfaces of Ab1999 (salmon), BG18 (gray) and PGT121 (purple) CDRH3s to V1 (green), V2 (olive) and V3 (pink). GDIR motifs are colored in red. p, Comparison of BG18 mature (mt) antibody bound to gp120 (gray; PDB 6DFG) with BG18 iGL precursor antibody bound to gp120 (orange, PDB 6DFH). Zoom-in view of the antibody–gp120 interfaces for both antibodies. q, Gp120 surfaces (BG18 mt engaged gp120 in gray and BG18 iGL engaged gp120 in orange) and buried surface areas interacting with BG18 mt and BG18 iGL shown in green for V1 and pink for V3. Measured buried area numbers are labeled accordingly. Data were analyzed by two-sided Student’s t-test, presented as mean values + s.d. and considered statistically significant at *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001 and ****P ≤ 0.0001.
Ab1999 binds to WIN332, to RC1, which reintroduces the Asn332 glycan, and to 11MUTB, 10MUT and 7MUT, which reintroduce the Asn332 and Asn156 glycans (Fig. 4b). Ab1999 does not bind to WIN332glycanKO, a mutant version of WIN332 that abrogates the V3-glycan epitope (Fig. 4c). WIN332glycanKO is different from WIN332 in that it lacks the V3 glycan at position 301, the only signature glycan remaining in WIN332. Ab1999 also shows reduced binding to WIN332-GAIA (Fig. 4c), strongly indicating V3-glycan epitope specificity and binds to fully glycosylated BG505 (Fig. 4b,d and Extended Data Fig. 3c).
To evaluate the capacity of Ab1999 to neutralize HIV-1, we performed TZM-bl assays against a series of HIV-1 pseudoviruses carrying the autologous WIN332 Env and a series of gradually more native-looking Envs including RC1, 11MutB, 10Mut, 5Mut, BG505 and the mutant BG505 Asn301Ala to map V3-glycan specificity. We observed high neutralization activity against the WIN332, RC1 and 11MutB pseudoviruses and serially reduced neutralization activity against more native-like pseudoviruses (Fig. 4e). Ab1999 showed low but reproducible inhibitory activity indicative of neutralization against the fully glycosylated BG505 pseudovirus and showed no neutralization for the BG505 Asn301Ala pseudovirus and MLV control (Fig. 4f), recapitulating the low activity observed in the serum and consistent with low levels of SHM. As an additional control, a different NHP antibody isolated from a WIN332-primed macaque (Ab2045) did not show any detectable neutralization activity against any of these pseudoviruses (Extended Data Fig. 3d). Ab1999 did not show neutralization activity in TZM-bl assays against a panel of multi-clade tier-2 HIV-1 pseudoviruses (Supplementary Tables 1 and 2).
These data demonstrate that a single immunization with WIN332 elicits antibodies that neutralize HIV-1 pseudoviruses, including low levels of neutralization against the fully glycosylated autologous BG505 virus.
Cryo-EM structure of Ab1999
To elucidate the molecular basis of Ab1999 recognition of Env, we determined a cryo-EM structure of Ab1999 Fab in complex with RC1 at a resolution of 3.9 Å (Fig.4g,h, Supplementary Fig. 2 and Supplementary Table 3). The structure revealed a single Ab1999 Fab bound to RC1, with the interaction predominantly mediated by the antibody’s long CDRH3 loop (Fig. 4h,i). Ab1999 engages the V1, V2 and V3 regions of RC1 (Fig.4j), and we did not observe antibody–glycan interactions (Fig. 4j–n). We observed hydrogen bond (h-bond) interactions between CDRH3 and CDRL1 with the V1 loop. In CDRH3, E100H and V100A interact with Lys151 and Asn149, respectively. In CDRL1, Tyr32 interacts with Ala135 through a backbone h-bond. (Fig. 4k). We also perceived hydrophobic contacts formed by the CDRL1, CDRL2 and CDRH3 with V1. Tyr32 and His34 of CDRL1, together with Ala50 and Tyr49 of CDRL2 form a hydrophobic patch with Leu138 and Leu139 of RC1. In the CDRH3, V100A and T100F interact with Asn149 and Arg151 of RC1 (Fig. 4l). Interactions with V2 are less extensive but include van der Waal’s interactions between V100D of CDRH3 and Arg178 of RC1 (Fig. 4m). Similarly, Ab1999 engages V3 through the CDRH3 tip via hydrophobic interactions, where residues V100A and F100B of CDRH3 make contacts with Met328. In a previous study, the original Gln328 residue on V3 was mutated to methionine (Gln328Met) to increase affinity for BG18 precursors11. We observed the CDRH3 tip, especially the TxFGVV motif, engages with a critical hydrophobic core consisting of Asn149, Met150, Met328, Pro417 and Arg419 (Fig. 4n). Although the CDRH3 of Ab1999 shares the same TxFGVV motif with BG18, the interaction with V3 and the structural comparison with BG18 and PGT121 show a distinct binding mechanism. Notably, Ab1999 binds RC1 by recognizing an alternative hydrophobic pocket consisting of V1, V2, V3 (Met328) and C4 without the involvement of the Asn332 glycan (Fig. 4o).
Since Ab1999 was an early elicited antibody, we compared its epitope with the epitope of BG18 iGL. We compared the Fab-Env structures of iGL and mature BG18 and observed that the buried areas for BG18 iGL at V1 and V3 were 488 Å2 and 188 Å2, respectively, whereas for mature BG18 they were 372 Å2 and 356 Å2 (Fig. 4p,q). These data indicated that the BG18 precursor more strongly relied on the V1 loop over the V3 loop for binding and that upon maturation, V1 and V3 dependencies were reduced and increased, respectively. At this stage of maturation, the engagement of Ab1999 was similarly more dependent on V1 than V3. In fact, WIN332 and related previously described immunogens include several mutations on V1 introduced to increase affinity for the V3-glycan epitope11 (Extended Data Table 1). We conclude that Ab1999 binds to the V3-glycan epitope of Env in an Asn332-glycan-independent manner.
WIN332 elicits EPTC112-like antibodies in NHPs
Ab1983 was selected for further characterization for its extraordinarily high similarity with the human type-II V3-glycan bNAb, EPTC112 (ref. 16; Fig. 5a). The HC of Ab1983 has 83.3% amino acid identity with the HC of iGL EPTC112, 61.9% with the mature EPTC112 HC (Extended Data Fig. 3e) and contains all the amino acid motifs critical for the neutralization activity of EPTC112 (ref. 16; Fig. 5a). These motifs include two serines and one tyrosine in the CDRH1, one serine in the CDRH2 and an ‘SGW’ motif in the CDRH3 (Fig. 5a). Ab1983 uses macaque VH4S13, DH6-31, JH4*01, VL9-84 and JL2A*01 genes (Extended Data Table 4). VH4S13 and DH6-31 share 91.7% and 95.2% identity with the human VH4-59 and DH6-19 genes of EPTC112 (Extended Data Table 4), respectively. Ab1983 has three nucleotide and one nucleotide SHMs in the VH and VL genes, respectively, and a 14-amino-acid-long CDRH3, which is comparable in length to the 15-amino-acid-long CDRH3 of EPTC112 (Fig. 5a and Extended Data Table 3).

a, Alignment of the amino acid sequences of Ab1983 and iGL EPTC112 HC. Dots represent amino acids that are different between sequences. Key motifs for neutralization and binding of EPTC112 are highlighted in pink. b, ELISA binding of Ab1983 to the indicated engineered Envs. EPTC112 was used as a positive control at 5 µg ml−1. Data are representative of at least two independent experiments. c, ELISA binding of Ab1983 to WIN332 and two mutant versions of WIN332 that abrogate the V3-glycan epitope: WIN332glycanKO and WIN332-GAIA. Data are representative of at least two independent experiments (P = 0.0086) d, Atomic model of the WIN332 Env–Ab1983 complex. e, Cryo-EM density map of the WIN332 Env trimer (gp120 in blue, gp41 in gray) bound to two Ab1983 Fabs (resolution of 3.8 Å). Three views highlight the Fab’s orientation and binding site. f, Close-up view of the Ab1983 paratope, with annotated CDRs that interact with the Env epitope. Gp120 is shown as surface with contact residues colored according to Ab1983 CDR loops. g, Interactions of Ab1983 with V1 (green), V2 (olive) and V3 (pink). h,i, Detailed hydrogen bonds (h) and hydrophobic interactions (i) with the V3 loop. Hydrogen bonds (dashed lines) between Ab1983 residues and Env residues are shown, along with Asn301 glycan-mediated interactions. j,k, Interactions with V1. Hydrogen bonding (dashed lines; j) and hydrophobic interactions (k) are shown. l, Limited interaction with V2 and the hydrophobic core formed by Trp100B. m, Comparison of the mechanisms used by Ab1983, EPTC112, BG18 and PGT121 to bind the GDIR motif. GDIR motifs are colored in red. Data were analyzed by two-sided Student’s t-test, presented as mean values + s.d. and considered statistically significant at *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001 and ****P ≤ 0.0001.
Ab1983 showed high binding to WIN332 and RC1 in ELISA (Fig. 5b) and did not bind to 11MutB, native BG505 and WIN332 glycan KO (WIN332 with an Asn301-glycan deletion) suggesting that Ab1983 requires the Asn301 glycan for binding (Fig. 5b,c and Extended Data Fig. 3f). Ab1983 showed reduced binding to WIN332-GAIA, suggesting that antibody binding depends on an intact GDIR motif (Fig.5c).
Cryo-EM structure of Ab1983
To elucidate the molecular basis of Ab1983 recognition of WIN332, a cryo-EM density map was reconstructed at a resolution of 3.8 Å (Fig. 5f,g, Extended Data Fig. 3 and Extended Data Table 5). The structure revealed two copies of Ab1983 bound to WIN332 Env (Fig. 5d–f). Ab1983 Fab engages with V1, V2 and V3 regions, with its heavy chain forming most contacts with V3 (Fig. 5g–i). The Ab1983–WIN332 structure revealed a complex interaction network involving h-bonds and hydrophobic interactions across the V1, V2 and V3 loops. We observed that the CDRL3, CDRH1, CDRH2 and CDRH3 loops of Ab1983 interact with V3 through both h-bonds and hydrophobic contacts (Fig. 5h,i). In CDRL3, the side chain of Thr94 forms both hydrogen and hydrophobic interactions with Asp325. In CDRH1, Tyr33 engages the Gly324 of the GDIR motif through an h-bond and Ile323 through hydrophobic contacts, whereas Ser31 forms an h-bond with the side chain of Lys305. In CDRH2, Ile53 forms hydrophobic contacts with Ile323. In CDRH3, Tyr98 forms h-bonds with the backbone of Ile322 and the side chain of D321A, while simultaneously making hydrophobic contacts with Ile326. (Fig. 5h). Ab1983, similarly to EPTC112 and PGT121, binds the Asn301 glycan, although using different binding mechanisms16,23. Ile53, Ser54 and Glu55 in the CDRH2 of Ab1983 are critical for the contact with the Asn301 glycan (Fig. 5i). Furthermore, the CDRH3, CDRL1, CDRL2 and CDRL3 loops engage with V1 (Fig. 5j,k). Ab1983 forms hydrophobic contacts between Trp100B of CDRH3 and Thr132, Tyr134 and Gln156 of V1. Also, hydrophobic interactions are formed between Tyr31 of CDRL1, G52E of CDRL2 and Thr94 of CDRL3 and Leu138, Thr132 and Asn137, respectively (Fig. 5j,k). In detail, in CDRH3, Trp100B binds with the backbone of Asn133 via an h-bond, as well as interacts with a hydrophobic core consisting of Thr132, Tyr134, Gln156 and Leu175 (Fig. 5j–l). In addition, Ser99 and Ser100 form h-bonds with the backbone of Asn133, Ala135 and Asn137 of WIN332 (Fig. 5j). In CDRL1 and CDRL3, Ala32 (CDRL1) and Asp91 (CDRL3) backbones interact with the Asn137 side chain through h-bond interactions, whereas Tyr31 (CDRL1) and Thr94 (CDRL3) form hydrophobic contacts with Leu138 and Asn137, respectively. In CDRL2, G52E interacts with Thr132 via hydrophobic contacts (Fig. 5k). At V2, only limited contacts were formed by the backbone of I52C in the CDRL2 with Ser187 of WIN332 (Fig. 5l). Notably, Ab1983 engages with the GDIR motif through a combination of h-bonds and hydrophobic interactions between CDRH1 (Tyr33), CDRH3 (Tyr98), CDRL3 (Thr94) and Gly324, Ile326 and Asp325, respectively (Fig. 5h,i). When comparing with other V3-glycan bNAbs, we observed the heavy chain of Ab1983 adopts a similar angle of approach to the GDIR motif to EPTC112, but engages the GDIR motif differently from BG18 and PGT121 (Fig. 5m)
We conclude that Ab1983 is an EPTC112-like antibody that recognizes the V3-glycan epitope in an Asn332-glycan-independent manner and through interactions with V1, V3 and the Asn301 glycan.
Sequential immunization boosts the neutralization activity elicited by WIN332 in NHPs
To boost the neutralizing response elicited by WIN332, we sequentially immunized these NHPs with an escalating dose of 7Mut-ST2-Asn332Tyr, a modified version of 7Mut11 where several glycan holes and the trimer base were masked through the introduction of PNGS sites, and a tyrosine was added at position 332 to contribute a bulky side chain that mimics the presence of the Asn332 glycan (Fig. 6a). We isolated WIN332-specific germinal center B cells from dLNs collected 3 weeks after the boost from the sequentially immunized NHPs (Fig. 6b and Extended Data Fig. 4a). Antibody cloning revealed extensive clonal expansion (Fig. 6c) and affinity maturation of WIN332-specific antibodies. Average numbers of mutations were increased to 12.7 nucleotides for HC and 6.9 nucleotides for LC (7.7 nucleotides for VL and 6.2 for VK; Fig. 6d,e)

a, Immunization protocol. b, Flow cytometry plot showing WIN332-specific B cells (purple) in pooled LN homogenates from sequentially immunized NHPs. c, Pie charts showing B cell clonal expansion in sequentially immunized NHPs. Pie charts include HC, KC and LC from B cells isolated from NHPs 2 and 3. d,e, Nucleotide mutations in HC (P = < 0.00001; d) and LC (P = < 0.00001; e) of B cells isolated after WIN332 prime (Prime) and 7Mut-ST2-Asn332Tyr boost (Boost). f, ELISA binding of the serum (n = 4) collected after WIN332-prime and 7Mut-ST2-Asn332Tyr boost to the boosting immunogen 7Mut-ST2-Asn332Tyr (P = 0.0007). g, ELISA results showing the binding of the serum (n = 4) collected after the boost to WIN332 and WIN332-GAIA (P = 0.0017). h, ELISA binding of the prime and boost serum (n = 4) to different native Envs (P = 0.0006, 0.0028, 0.0169 and 0.0017, respectively). i, Results of TZM-bl neutralization assays using purified immunoglobulins from the serum of the sequentially immunized NHPs against the fully glycosylated BG505 pseudovirus (BG505 Thr332Asn) and three mutant versions with glycan deletions at the V3-glycan epitope (BG505 Asn137Asp, BG505 Asn156Ala and BG505 Asn301Ala). PGT121 was used as a control. Data are representative of at least two independent experiments. j, Graph shows IC50 values for the neutralization activity in h (P = 0.0036, 0.0076). k, Results of TZM-bl neutralization assays using purified serum immunoglobulins (300 µg ml−1) from the four NHPs against a panel of 24 tier-2 HIV-1 pseudoviruses. Each dot corresponds to a macaque. The dotted line corresponds to MLV background. Data were analyzed by Student’s t-test, presented as mean values + s.d. and considered statistically significant at *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001 and ****P ≤ 0.0001. Illustrations in a were created using BioRender.com.
7Mut-ST2-Asn332Tyr strongly boosted the serologic response to the boosting immunogen (Fig. 6f), and the response remained partially dependent on an intact GDIR motif (Fig. 6g). This boost immunization induced significantly stronger binding to the fully glycosylated BG505 trimer and to the heterologous trimers Q23.17, AD8 and 001428 (Fig. 6h)
Remarkably, boost immunization with 7Mut-ST2-Asn332Tyr significantly increased the serum neutralization activity against the fully glycosylated BG505 pseudovirus (Fig. 6i) and the neutralization activity was partially specific to the V3-glycan epitope as determined through mapping TZM-bl assays using purified immunoglobulins against the BG505-based mutant pseudoviruses BG505 Asn137Asp, BG505 Asn156Ala and BG505 Asn301Ala and MLV as a control (Fig. 6i,j and Extended Data Fig. 4b). To evaluate whether these early serum antibodies showed any neutralization breadth, we tested purified serum immunoglobulins for neutralization against a panel of 24 tier-2 HIV-1 pseudoviruses from different clades in TZM-bl assays. This panel was selected based on sensitivity to human type-I and type-II V3-glycan bNAbs (Supplementary Table 2)17. Although we did not observe neutralization activity above 50% for any of the pseudoviruses in this panel, we observed low-level inhibition above the MLV control that may indicate the onset of neutralization activity against seven of the pseudoviruses (Fig. 6k). These pseudoviruses naturally lack glycans at positions Asn133 or Asn137 and/or Asn332, in agreement with elicitation of these antibodies by immunogens lacking these glycans.
We conclude that WIN332 followed by sequential immunization with 7Mut-ST2-Asn332Tyr elicits neutralizing antibodies that target the V3-glycan epitope of Env.
Discussion
Sequential immunization is a promising strategy to elicit anti-HIV-1 bNAbs. Important immunogen design efforts are focused on germline-targeting approaches aiming to activate and mature a few selected human bNAb lineages. Several germline-targeting immunogens have been successful at activating precursors of selected human type-I V3-glycan bNAb lineages1,10,12,14. Sequential immunization was effective at maturing the precursor of PGT121 into highly mutated, broad and potent PGT121-like antibodies in an immunoglobulin knockin mouse expressing iGL PGT121 (ref. 1). Subsequent studies have attempted to achieve neutralization potency and breadth against the V3-glycan epitope through immunization in outbred animals; however, success has been modest, with sporadic elicitation of neutralizing responses of limited potency and breadth29 or no neutralization activity28.
Reported sequential immunization protocols designed to elicit bNAbs involve numerous immunizations or sustained antigen release over long periods of time2,13,29,30. Identifying ‘premium’ antibody lineages that require less convoluted maturation pathways to become bNAbs and designing immunogens that activate them could simplify sequential immunization protocols by reducing the number of immunization steps. Here we used a lineage ‘agnostic’ approach for immunogen design to activate a broad diversity of antibody lineages to the conserved V3-glycan epitope. This approach increases the probability of activating V3-glycan antibody lineages with the potential to mature into bNAbs and broadens the repertoire of antibody lineages to consider for vaccine design purposes.
Human V3-glycan bNAbs show promiscuous binding to the V3-glycan epitope involving different angles of approach and glycan usage; however, all V3-glycan bNAbs isolated until very recently seem to establish key contacts with the Asn332 glycan7,8,9,21. Multiple observations by us and others led us to hypothesize that there may be antibody lineages that bind the V3-glycan epitope in an Asn332-glycan-independent manner, that Asn332-glycan dependence may develop during bNAb maturation, and that Asn332-glycan deficient Env immunogens may be more efficient at activating V3-glycan bNAb precursors. Among our observations, we found antibodies that bound the V3-glycan epitope in an Asn332-glycan-independent manner in previous mouse and NHP studies. Moreover, the Asn332 glycan is underrepresented in T/F viruses15 and infection with Asn332-glycan-deficient T/F viruses led to the development of Asn332-glycan dependent V3-glycan bNAbs in two humans15 suggesting that Asn332-glycan-deficient Env proteins may play a role in bNAb lineage activation. In line with our hypothesis, the UCA of the human V3-glycan bNAb DH270 bound to a synthetic V3-glycopeptide (Man9-V3) containing the Asn332 and Asn301 glycans but also bound to its aglycone version31. Even though DH270 UCA establishes contacts with the Asn332 glycan in the context of the CH848 d949 Env protein14, glycan dependence appears when the antibody is ~6% mutated19, suggesting that the Asn332 glycan may not be necessary to activate this lineage32. Our immunization experiments with WIN332 together with the recent isolation and characterization of two human bNAbs that target the V3-glycan epitope in an Asn332-glycan-independent manner16,17 demonstrate that in addition to type-I Asn332-glycan-dependent lineages, Asn332-glycan-independent V3-glycan antibodies (type II) should be considered for vaccine design purposes. Noteworthy, WIN332 lacks not only the Asn332 glycan, which was rationally removed, but also the Asn334 glycan, which is naturally absent in the BG505 strain. Thus, the antibodies elicited by WIN332 are not only Asn332-glycan independent, but also Asn334-glycan independent.
WIN332 bound to iGLs and UCAs of both type-I and type-II human bNAbs. WIN332 showed high affinity for BF520 UCA despite the BF520 bNAb being an Asn332-glycan-dependent type-I bNAb. Interestingly, BF520 UCA showed higher affinity to WIN332 than to RC1 (WIN332 + Asn332 glycan) supporting our hypothesis that the precursors of type-I bNAbs may not need the Asn332 glycan for activation and that Asn332-glycan dependency may develop through SHM of Asn332-glycan-independent precursors. Moreover, we show that WIN332 is able to activate antibody responses in iGL PGT121 immunoglobulin knockin mice, altogether suggesting that WIN332 could be efficient at broadly activating V3-glycan bNAb precursors in humans. In NHPs, WIN332 induced a diverse repertoire of Asn332-glycan-independent V3-glycan antibodies with high homology with human type-I bNAbs such as PGT121, BG18 or DH270 and also the recently isolated type-II bNAb EPTC112.
Remarkably, WIN332-elicited serum and mAbs with low inhibitory activity indicative of neutralization against the autologous fully glycosylated HIV-1 pseudovirus upon one single immunization and at times as short as 3 weeks after immunization. The activity observed after the prime is low, not reaching IC5O levels; however, high levels of neutralization activity cannot be expected after one single immunization. This low activity is reproducible under different experimental conditions and recapitulated by a mAb (Ab1999) isolated from the macaques. Furthermore, a sequential boost immunization induced affinity maturation of V3-glycan lineages activated by WIN332 and increased the magnitude of the autologous neutralizing activity of the serum. This serum also showed low heterologous neutralization activity (lower than 50%) against seven pseudoviruses from a panel of 24 multi-clade tier-2 viruses selected based on sensitivity to human type-I and type-II V3-glycan bNAbs. This early binding to the native V3-glycan epitope of Env and neutralization activity has not been previously observed in response to immunization with any reported immunogen or immunization strategy.
Of note, the mAbs isolated from the immunized NHPs, including the neutralizing Ab1999, had very low amounts of SHM, suggesting that moderate levels of SHM could be sufficient to achieve potency and breadth. Ab1999 binding to RC1 Env involves important contacts with V1, which parallels the higher involvement of V1 versus V3 in the mechanism of binding of BG18 iGL and other BG18-like antibodies12. The maturation of BG18 resulted in increased contacts of V3 to strengthen the binding to the epitope and reduced engagements of V1 suggesting that a similar maturation path could be possible for Ab1999.
A striking feature of the reported type-II EPTC112 bNAb was its short CDRH3 of 15 amino acids, which is the average length found in the human antibody repertoire. Ab1983, the EPTC112-like NHP antibody characterized here, has a similar CDRH3 of 14 amino acids. Without the requirement of a long CDRH3, this class of antibody is expected to be easier to elicit through immunization.
Altogether, here we establish two classes of V3-glycan antibody lineages: type-I, Asn332-glycan-dependent and type-II, Asn332-glycan-independent antibodies and report WIN332, an Env immunogen rationally engineered to activate and mature antibody lineages that do not require the Asn332 glycan at early stages of maturation. The early neutralization induced by WIN332, presents this immunogen as a promising candidate to shorten sequential immunization protocols and streamline HIV-1 vaccine development.
Methods
Ethics and inclusion
This research complies with all relevant ethical regulations (Tulane National Primate Research Center IACUC protocol P0498 (3698) and The Wistar Institute IACUC protocols 201518 and 201522). Research has been conducted following the recommendations set out in the Global Code of Conduct for Research in Resource-PoorSettings including local researchers throughout the research process.
Envelope proteins
The newly engineered Env SOSIP trimers, WIN332, WIN332-GAIA–AviTag, WIN332–AviTag, 7MUT-ST2-Asn332Tyr and other previously reported engineered SOSIP Env trimers, RC1, RC1-glycan knockout (same as WIN332 glycan KO)10, 11MUTB, 10MUT, 7MUT, 5MUT11 and native-like BG505 Thr332Asn18, CEO217 (clade C), Q23 (clade A), AD8 (clade B), BJOX2000 (clade AE), CNE55 (clade C), 001428 (clade B), CH119 (clade BC) and AMC011 (clade B) were cloned in the pPPI4 or pVAX expression vectors using synthetic gene fragments (Integrated DNA technologies) or were produced by GenScript. Specific mutations for each protein including glycan deletions are listed in Extended Data Table 1.
All the SOSIP trimers used in this study include MD39 mutations for stabilization11 (Extended Data Table 1).
Env trimers were expressed as soluble native-like gp140 trimers18. Soluble Env trimers were expressed by transient transfection in Expi293 cells (Thermo Fisher Scientific) and purified from cell supernatants by Lectin chromatography and SEC as previously described33. Proteins were stored at −80 °C in PBS until the time of use.
Analysis of site-specific N-linked glycopeptides by liquid chromatography–mass spectrometry
Aliquots of purified WIN332 protein were reduced by incubating with 10 mM dithiothreitol (Sigma) at 56 °C and alkylated by 27.5 mM iodoacetamide (Sigma) at room temperature in the dark. The aliquots were then digested by using a combination of alpha lytic protease (New England BioLabs), AspN (Promega), chymotrypsin (Athens Research and Technology), Lys-C (Promega), Glu-C (Promega) and trypsin (Promega). The resulting peptides were separated on an Acclaim PepMap 100 C18 column (75 μm × 15 cm) and eluted into the nano-electrospray ion source of an Orbitrap Eclipse Tribrid mass spectrometer (Thermo Scientific) at a flow rate of 200 nl min−1. The elution gradient consisted of 1–40% acetonitrile in 0.1% formic acid over 370 min followed by 10 min of 80% acetonitrile in 0.1% formic acid. The spray voltage was set to 2.2 kV, and the temperature of the heated capillary was set to 275 °C. Full mass spectrometry scans were acquired from m/z 200 to 2,000 at a resolution of 60,000, and tandem mass spectrometry scans following higher-energy collisional dissociation with stepped collision energy (15%, 25%, 35%) were collected in the orbitrap at a resolution of 15,000. pGlyco3 (ref. 34) was used for database searches with mass tolerance set as 20 ppm for both precursors and fragments. The database search output was filtered to reach a 1% false discovery rate for glycans and 10% for peptides. The glycan and peptide assignment for each spectra was then manually validated after filtering. Quantification was performed by calculating spectral counts for each glycan composition at each site. Any N-linked glycan compositions identified by only one spectra were removed from quantification. N-linked glycan compositions were categorized into 19 classes (including ‘Unoccupied’ as class 19): HexNAc(2)Hex(9 ~ 5)Fuc(0 ~ 1) was classified as M9 to M5, respectively; HexNAc(2)Hex(4 ~ 1)Fuc(0 ~ 1) was classified as M1-M4; HexNAc(3 ~ 6)Hex(5 ~ 9)Fuc(0)NeuAc(0 ~ 1) was classified as Hybrid with HexNAc(3 ~ 6)Hex(5 ~ 9)Fuc(1 ~ 2)NeuAc(0 ~ 1) classified as F-Hybrid; Complex-type glycans are classified based on the number of antenna and fucosylation: HexNAc(3)Hex(3 ~ 4)Fuc(0)NeuAc(0 ~ 1) is assigned as A1 with HexNAc(3)Hex(3 ~ 4)Fuc(1 ~ 2)NeuAc(0 ~ 1) assigned as F-A1; HexNAc(4)Hex(3 ~ 5)Fuc(0)NeuAc(0 ~ 2) is assigned as A2/A1B with HexNAc(4)Hex(3 ~ 5)Fuc(1 ~ 5)NeuAc(0 ~ 2) assigned as F-A2/A1B; HexNAc(5)Hex(3 ~ 6)Fuc(0)NeuAc(0 ~ 3) is assigned as A3/A2B with HexNAc(5)Hex(3 ~ 6)Fuc(1 ~ 3)NeuAc(0 ~ 3) assigned as F-A3/A2B; HexNAc(6)Hex(3 ~ 7)Fuc(0)NeuAc(0 ~ 4) is assigned as A4/A3B with HexNAc(6)Hex(3 ~ 7)Fuc(1 ~ 3)NeuAc(0 ~ 4) assigned as F-A4/A3B; HexNAc(7)Hex(3 ~ 8)Fuc(0)NeuAc(0 ~ 1) is assigned as A5/A4B with HexNAc(7)Hex(3 ~ 8)Fuc(1 ~ 3)NeuAc(0 ~ 1) assigned as F-A5/A4B.
Analysis of deglycosylated HIV-1 envelope protein by liquid chromatography–mass spectrometry
Aliquots of the purified WIN332 protein were reduced by incubating with 10 mM dithiothreitol (Sigma) at 56 °C and alkylated by 27.5 mM iodoacetamide (Sigma) at room temperature in dark. The aliquots were then digested using chymotrypsin (Athens Research and Technology), Glu-C (Promega), Lys-C (Promega), Arg-C (Promega) and trypsin (Promega). Following digestion, the extracted peptides were deglycosylated by endoglycosidase H (Promega) followed by PNGaseF (Promega) treatment in the presence of 18O water (Cambridge Isotope Laboratories). The resulting peptides were separated on an Acclaim PepMap 100 C18 column (75 μm × 15 cm) and eluted into the nano-electrospray ion source of an Orbitrap Eclipse Tribrid mass spectrometer (Thermo Scientific) at a flow rate of 200 nl min−1. The elution gradient consisted of 1–40% acetonitrile in 0.1% formic acid over 370 min followed by 10 min of 80% acetonitrile in 0.1% formic acid. The spray voltage was set to 2.2 kV, and the temperature of the heated capillary was set to 275 °C. Full MS scans were acquired from m/z 200 to 2,000 at a resolution of 60,000, and tandem mass spectrometry scans following collision-induced dissociation at 38% collision energy and higher-energy collisional dissociation with stepped collision energy (15%, 25%, 35%) were collected in the ion trap. The spectra were analyzed using SEQUEST (Proteome Discoverer 2.5, Thermo Fisher Scientific) as well as Byonic (v4.1.10, Protein Metrics)35 with mass tolerance set as 20 ppm for precursors and 0.5 kDa for fragments. The search output was filtered to reach a 1% false discovery rate at the protein level and 10% at the peptide level. The site assignment for each spectra was then manually validated after filtering. Occupancy of each N-linked glycosylation site was calculated using spectral counts assigned to the 18O-Asp-containing (PNGaseF-cleaved) and/or HexNAc-modified (EndoH-cleaved) peptides and their unmodified counterparts.
Animals
Male rhesus macaques (Macaca mulatta) of Indian genetic origin, 4 years of age, were housed and cared for in a biosafety level-2 facility at the Tulane National Primate Research Center. All animal procedures and experiments were performed according to protocols approved by the Tulane National Primate Research Center Institutional Animal Care and Use Committee.
SMNP was produced as previously described22. At day 0, macaques received one subcutaneous bolus immunization with a total of ~200 μg of soluble WIN332 SOSIP trimer adjuvanted in 375 μg of SMNP distributed between the four legs (~50 µg per foreleg and ~50 µg per hind leg). Macaques were boosted with a total amount of 200 μg of soluble 7MUT-ST2-Asn332Tyr in 375 μg of SMNP adjuvant using an escalating dose delivery approach36 at week 8 (one dose of 30 µg of WIN332 in 56.5 µg SMNP at day 1, one dose of 60 µg of WIN332 in 112 µg SMNP at day 3 and one dose of 110 µg of WIN332 in 206.5 µg SMNP at day 5) and distributed between the four legs like the prime. Soluble trimers were selected based on previous unpublished immunization experiments in the humanized DH3-3/JH6 mice37. Blood samples were obtained from the macaques before immunization, 3 and 7 weeks after WIN332 prime and 3 weeks after 7MUT-ST2-Asn332Tyr boost immunization (week 11). Lymph node biopy samples were obtained from the immunized macaques 3 weeks after WIN332 prime and 3 weeks after boost immunization with 7MUT-ST2-Asn332Tyr.
Mice expressing the iGL precursor of the human V3-glycan bNAb PGT121 (C57BL/6 strain background) were provided by M. Nussenzweig (The Rockefeller University). Male and female mice were immunized intraperitoneally with 10 µg of WIN332 soluble Env trimer and three units of SMNP. Serum was collected 14 days later and used for ELISA analysis.
All mouse cages were on ventilated, ducted single-use racks. Cages were pre-bedded with alpha-dry bedding. Water bottles and cage supplies were irradiated. Cages were changed every 2 weeks, and lids and feeders were changed once a month inside a class-II biosafety cabinet using aseptic technique. Animals receive HCL water with a 3.0 pH. The Wistar Institute’s animal facility is operated as a modified barrier and with room-level protection. Holding rooms are negative to the corridors. Up to five adult mice or two adult mice plus one female with a litter may be housed in a cage (81 square inches). Temperature is within 64 to 79 degrees Fahrenheit and humidity is within 30% to 70%. Light cycle is 12-h on starting at 6:00 and 12-h off starting at 18:00.
ELISA
ELISAs with SOSIP Env trimers WIN332, RC1, WIN332glycanKO, 11MUTB, 10MUT, 7MUT, 5MUT and native Env trimers were performed as previously described10. Briefly, 96-well plates were coated with 50 μl of a solution of the trimer at 4 μg ml−1 in 1× PBS (Corning, 20-030-CV) and incubated at 4 °C overnight. Plates were washed with washing buffer (1× PBS with 0.05% Tween-20; Sigma-Aldrich, P7949) and blocked (1× PBS with 5% milk) for 1 h at room temperature. Serum samples were assayed at a 1:100 or 1:30 starting dilution, supernatants from Expi293 cells transfections were assayed at a 1:3 starting dilution and the antibody controls were added at 5 μg ml−1 all followed by seven additional threefold serial dilutions for 1 h at room temperature. Plates were washed three times and incubated with an anti-human IgG secondary antibody conjugated to horseradish peroxidase (HRP; Jackson ImmunoResearch, 115-035-071) at a 1:5,000 dilution in washing buffer for 1 h at room temperature. Plates were developed by adding ABTS, and optical density was measured at 405 nm.
Alternatively, 96-well plates were coated with 50 μl of a solution of PGT121 or 3BNC117 antibodies engineered to have a mouse Fc at 4 μg ml−1 in 1× PBS and incubated at 4 °C overnight. Plates were washed with washing buffer (1× PBS with 0.05% Tween-20), blocked (1× PBS with 5% milk) for 1 h at room temperature and incubated in 50 μl of a solution of an Env trimer at 4 μg ml−1 in blocking buffer for 1 h at room temperature. Plates were washed three times and incubated with 5 μg ml−1 of an antibody with human Fc and threefold serial dilutions for 1 h. Plates were washed three times and incubated with an anti-human IgG secondary antibody conjugated to HRP at a 1:5,000 dilution in washing buffer. Plates were developed by adding ABTS, and optical density was measured at 405 nm.
For competition ELISAs, 96-well plates were directly coated with 50 μl of a solution of an Env trimer at 4 μg ml−1 in 1× PBS and incubated at 4 °C overnight. Plates were washed three times in washing buffer and blocked in blocking buffer for 1 h at room temperature. Plates were then incubated in 50 μl of a 15 μg ml−1 solution of PGT121 or 3BNC117 with a mouse Fc in blocking buffer for 1.5 h at room temperature. Plates were washed three times and NHP serum or mAbs were added at the indicated starting dilutions and six additional threefold serial dilutions for 30 min at room temperature. Plates were washed and developed using anti-human IgG secondary antibody conjugated to HRP at a 1:5,000 dilution in washing buffer. Plates were developed by adding ABTS, and optical density was measured at 405 nm.
Endpoint titers were calculated as the reciprocal number of the dilution at which the sample optical density was three times the background.
Flow cytometry and single B cell sorting
Frozen cell homogenates from lymph node biopsy samples obtained from immunized macaques were thawed and washed in RPMI medium 1640 (1×; Gibco, 11875-093).
Lymph node cells were incubated with 100 μl of FACS buffer (PBS 1× with 2% fetal bovine serum and 1 mM EDTA) with human Fc Block (BD Biosciences, 564219, clone Fc1) at a 1:500 dilution for 30 min on ice.
WIN332, WIN332-GAIA and BG505 tetramers were prepared by incubating 5 μg of AviTagged and biotinylated WIN332 (WIN332–AviBio), WIN332-GAIA (WIN332-GAIA–AviBio) or BG505 (BG505–AviBio) with fluorophore-conjugated streptavidin at a 1:200 dilution in 1× PBS for 30 min on ice. WIN332++ WIN332-GAIA− or BG505++ lymph node B cells were isolated using WIN332–AviBio or BG505–AviBio conjugated to streptavidin AF647 (BioLegend, 405237), WIN332–AviBio or B41–AviBio conjugated to streptavidin PE (BioLegend, 405204) and WIN332-GAIA–AviBio conjugated to streptavidin BV605 (BioLegend, 405229) as baits. Tetramers were mixed with the antibody cocktails indicated below to a final concentration of 5 μg ml−1 each.
Macaque LN cells were stained with: anti-CD3 APC-eFluor 780 (Invitrogen, 47-0037-41, clone OKT3), anti-CD14 APC-eFluor 780 (Invitrogen, 47-0149-42, clone 61D3), anti-CD16-APC-eFluor 780 (Invitrogen, 47-0168-4, clone CB16), anti-CD8 APC-eFluor 780 (Invitrogen, 47-0086-42, clone OKT8), anti-CD20 PE-Cy7 (BD Biosciences, 335793, clone L27) and anti-CD38 FITC (Stem Cell, 60131FI, clone AT-1) at a 1:200 dilution and the LIVE/DEAD marker Zombie NIR (BioLegend, 423106) at a 1:400 dilution. Cells were incubated with the mixture of tetramers and antibody cocktail for 30 min on ice.
Zombie NIR−/CD16−/CD8a−/CD3−/CD14−/CD20+/CD38+/WIN332++/WIN332-GAIA− or BG505++/WIN332-GAIA− single cells were isolated from the macaque lymph node cell homogenates using a FACS Aria III (Becton Dickinson). Single B cells were sorted into individual wells of 96-well plates containing 5 μl of lysis buffer (TCL buffer (Qiagen, 1031576) with 1% 2-β-mercaptoethanol). The cell lysates were stored at −80 °C or immediately used for subsequent mRNA purification.
Alternatively, trimer-specific B cells were sorted in RPMI with 5% FBS for 10x genomics.
Antibody sequencing
RNA was purified from single cells using paramagnetic beads (2.2× ratio; RNAClean XP, A63987 Beckman Coulter). RNA was eluted from the magnetic beads with 11 μl of a solution containing 14.5 ng μl−1 of random primers (Invitrogen, 48190-011), 0.5% of tergitol, (type NP-40, 70% in H2O, Sigma-Aldrich, NP40S-100ML), 0.6 U μl−1 of RNAse inhibitor (Promega, N2615) in nuclease-free water (Qiagen), and incubated at 65 °C for 3 min. cDNA was synthesized by reverse transcription (SuperScript III Reverse Transcriptase, Invitrogen, 18080-044, 10,000 U)38. cDNA was stored at −20 °C or 10 μl of nuclease-free water was added to the cDNA before subsequent use for antibody gene amplification by nested PCR.
Macaque IgH and IgK/L genes were amplified and cloned38 using the primers and PCR protocols in Extended Data Tables 3 and 4 (ref. 39). Briefly, after a first PCR reaction to amplify the antibody genes, a second PCR (cloning PCR) introduced nucleotide tails that were used for subsequent ligation-independent cloning (LIC) in expression vectors39 (see ‘Antibody cloning’). The cloning PCR products were sequenced by Sanger sequencing using the primers in Extended Data Table 3.
10x Genomics
Single WIN332-specific germinal center B cells were bulk-sorted in RPMI with 5% FBS, processed by Wistar Genomics Core following the manufacturer protocol (Chromium Next GEM Single Cell V(D)J protocol v3) with modifications40 and macaque V(D)J libraries were generated.
Illumina NGS-generated raw data files were processed using the Cell Ranger single cell gene expression software provided by 10x Genomics. Sequences were quality filtered, followed by assembly and annotation using Cell Ranger adapted to contain rhesus macaque Ig genes from a custom reference. Contigs assembled by Cell Ranger were reannotated. To assign antibody sequences to each barcode, a custom Bash pipeline was developed to generate a CSV file that organizes query sequences by heavy chain (VH), light chain kappa (VK), and light chain lambda (VL). This pipeline incorporates a Python script that automatically aligns query sequences to germline V(D)J segments using the NCBI IgBLAST database. The resulting annotations are parsed and structured into separate columns based on their corresponding variable regions (VH, VK, VL). Finally, sequences sharing the same identifier are matched and paired by row to represent complete antibody repertoires. After pairing, sequences were analyzed with a custom pipeline with macaque immunoglobulin libraries to determine B cell immunogenetics and clonality41.
Antibody cloning
Antibody cloning was performed by GenScript or in house through LIC cloning. For LIC cloning, the cloning PCR products were purified using the PCR clean-up kit (Qiagen, 28183). The purified products were cloned into the corresponding expression vectors encoding human IgG, IgL and IgK constant regions digested with the restriction enzymes AgeI and SalI for IgH, AgeI and XhoI for IgL and AgeI and BsiwI for IgK as previously described39.
Antibody production and purification
Macaque IgGs were produced by GenScript or in house through transient transfection in Expi293T cells. For transfections, DNA mixtures containing IgH and IgL/IgK antibody genes at a 1:1 ratio were used for transfections following the manufacturer’s instructions (Thermo Fisher Scientific, A14635). IgGs were purified from the supernatants of transfected Expi293T cells using Protein G (Cytiva, 28-4083-47).
For structural studies, macaque Fab was obtained by digesting IgG at 1–5 mg ml−1 with ficin (Sigma). Fab was purified by SEC42, followed by monoQ 5/50 (GE Healthcare) ion exchange chromatography.
Serum immunoglobulins were purified from 200 μl of macaque serum using Ab Spin Trap Protein G Sepharose columns (GE Healthcare, 28-4083-47). Immunoglobulin-containing fractions were buffer exchanged with PBS using a 30-kDa molecular-weight-cutoff concentrator (Milipore Sigma, UFC903024).
In vitro neutralization assay
Env-pseudotyped viruses were produced following the standardized protocol43. Briefly, HEK 293T cells were seeded in 100-mm plates at a density of 4 × 10⁶ cells per plate and incubated overnight in complete growth medium. Cells were transfected using FuGENE 6 (Promega) with 4 µg of Env plasmid and 8 µg of backbone plasmid per plate. After a brief incubation (4–5 h), the medium was replaced, and the culture was maintained for 48 h. Virus-containing supernatants were harvested, supplemented with fetal bovine serum to a final concentration of 20%, filtered through 0.45-µm membranes, aliquoted into cryovials and stored at −80 °C.
Pseudovirus titers were determined using a tissue culture infectious dose assay in TZM-bl cells, a reporter cell line engineered to express luciferase under control of the HIV-1 Tat protein43. Serial fivefold dilutions of the pseudovirus stock were prepared in 96-well plates, and 10,000 TZM-bl cells per well were added in the presence of DEAE-dextran (Final concentration of 40 µg ml−1). After 48 h of incubation at 37 °C, luminescence was measured using the Firefly Luciferase Assay Kit (Merck). Virus input was considered optimal when luminescence signals fell within the range of 15,000 to 50,000 relative luminescence units.
TZM-bl assays were performed as described44. For the assay setup, 96-well flat-bottom plates were used. To reduce edge effects and minimize variability due to evaporation, all peripheral wells (columns 1, 11 and 12; rows A and H) were filled with sterile PBS and were not used for experimental conditions. The assay was conducted using the remaining internal wells. Column 2 served as the virus control (cells and virus without antibody), while columns 3 to 10 were used for serial dilutions of the test antibody or serum.
Serial threefold dilutions of the test antibody or purified IgGs from macaque serum were performed in duplicate, using wells from bottom to top (row G to row B) with a starting concentration of 100 µg ml−1 for WIN332, RC1 and 11MutB pseudoviruses, 200 µg ml−1 for 10Mut and 7Mut and 400 µg ml−1 for 5Mut, BG505 Thr332Asn, BG505 Asn301Ala and MLV. Serial threefold dilutions of purified IgGs from macaque serum were performed in duplicate, using wells from bottom to top (row G to row B) with a starting concentration of 300 µg ml−1. After dilutions were prepared, 50 μl of pseudovirus diluted in growth medium was added to all experimental wells (columns 2–10).
Plates were incubated at 37 °C for 1 h to allow virus–antibody interaction. During this time, a suspension of TZM-bl cells was prepared at 1 × 10⁵ cells per milliliter in growth medium containing DEAE-dextran (final concentration of 40 µg ml−1). Of this suspension, 100 μl was added to each well. Plates were incubated for 48 h at 37 °C in a humidified 5% CO2 atmosphere.
Neutralization activity was calculated as a function of the reduction in Tat-induced luciferase expression.
BLI (OCTET) binding studies
BLI experiments were performed using the OCTET Red96 system to determine affinities of antibodies (Fabs) for WIN332. Biotinylated WIN332–AviTag (WIN332–AviBio) was immobilized on high-precision streptavidin (SAX) biosensors (FORTÉBIO) using a solution of WIN332–AviBio at 5 µg ml−1 in dilution buffer (FORTÉBIO). Four serial dilutions (20, 10, 5 and 2.5 µg ml−1) of each experimental Fab or antibody and PGT121 antibody were prepared in dilution buffer (FORTÉBIO). Alternatively, the experimental antibody (5 µg ml−1) was immobilized on Anti-Human Fc Capture (AHC) Biosensors (FORTÉBIO) and assayed against four serial dilutions (20, 10, 5 and 2.5 µg ml−1) of WIN332. The binding experiment was performed at 30 °C using the following protocol: baseline 1 (60 s), WIN332 or antibody/Fab loading (300 s), baseline 2 (200 s), association (300 s) and dissociation (600 s). Analysis was performed using OCTET software Data Analysis HT 10.0 (FORTÉBIO).
EMPEM
Serum from each immunized macaque was tenfold diluted in PBS. Diluted sera were incubated with protein G sepharose 4 Fast Flow resin (Cytiva) overnight at 4 °C. Polyclonal IgG was purified by eluting in 0.1 M Glycine-HCl (pH 3.5) and neutralized immediately in 1 M Tris-HCl (pH 8). Purifed polyclonal IgG samples were further digested with 2% papain (Thermo Fisher). Around 2 mg of digested polyclonal Fab was incubated with 15 µg of WIN332 or RC1 trimer on ice or overnight at 4 °C. The complexes were purified by SEC through a Superose 6 Increase 10/300 GL column (Cytiva). Fractions corresponding to trimer–Fab complexes were concentrated. Four microliters of purified complexes at 0.01 mg ml−1 were applied to a glow-discharged, carbon-coated 400 mesh copper grid (Electron Microscopy Sciences). After blotting extra samples, grids were stained with 2% uranyl formate for 2 min followed by blotting extra staining buffer. Grids were imaged using a 100 kV FEI Tecnai T12 microscope at a magnification of ×57,000 equipped with a Gatan CCD camera. Collected micrographs were processed in Relion45. Specifically, Laplacian-of-Gaussian-picked particles were cleaned up by two-dimensional classification. Subset classes showing a clear trimer or Fab bound trimer were selected for three-dimensional (3D) classification. Data analysis and figure generation were performed in UCSF Chimera46.
Cryo-EM sample and grid preparation
RC1 and Ab1999 Fab or WIN332 and Ab1983 Fab were mixed together using a molar ratio of 1:9 (Env:Fab) and incubated for 1 h on ice. Complexes were purified by SEC through a Superose 6 increase 10/300 GL column (Cytiva). Fractions containing the Env–Fab complexes were pooled and concentrated down using 100-kDa molecular-weight-cutoff concentrators. Samples were flash frozen in liquid nitrogen and stored at −80 °C.
Complexes were thawed and diluted to around 0.085 g l−1. UltrAuFoil 1.2/1.3 300 mesh grids were glow discharged for 1 min and immediately used for graphene oxide application using a previously published protocol47. Complexes were vitrified by applying 4 μl of the diluted sample to the graphene oxide-coated grids using vitrobot Mark IV (FEI) at 4 °C and 100% humidity followed by plunging into liquid ethane.
Cryo-EM data collection and processing
Cryo-EM grids containing RC1–Ab1999 or WIN332–Ab1983 complexes were loaded onto a 200-kV Thermo Fisher Glacios Microscope equipped with a Falcon 4 detector. Dose-fractionated movies were collected using a total dose 40 e−/Å2 across 40 frames or 50 e−/Å2 across 49 frames for WIN332–Ab1983 or RC1–Ab1999, respectively. Both datasets were collected with a defocus range of −0.8 μm to −2.2 μm.
Data processing for both datasets was done similarly. In summary, raw movies were imported into CryoSPARC and preprocessed using Patch Motion Correction and Patch CTF Estimation. Particles were picked using a blob picker and then extracted with binning by 4 resulting in pixel size of 3.79 Å per pixel. Particles were first cleaned up using two-dimensional classification and particles in the junk classes were discarded. Remaining particles were used for ab initio reconstruction, and the resulting 3D volumes were used for subsequent heterogeneous refinement. Particles belonging to the good 3D classes were selected and reextracted with binning by 2. This was followed by another round of heterogeneous refinement. The selected particles were then reextracted with the original pixel size and used for non-uniform refinement. The resulting auto-sharpened maps were used for model building and analysis.
Model building
Initial models of Ab1999 and Ab1983 Fabs were generated using SWISS-MODEL48 without reference structures. The previously published model of RC1 (PDB 6ORO) was used as the initial model for model building for both RC1–Ab1999 and WIN332–Ab1983. The WIN332 Env model was modified by deleting the Asn332 glycan and mutating the asparagine into glutamine. Reference models were docked into the density maps in UCSF ChimeraX49 and manually real-space refined in Coot50. N-linked glycans were added to the models manually in Coot50. FModels were refined in Rosetta and real-space refined in Coot iteratively. Model geometry was validated by MolProbity51, glycan conformation was validated by Privateer52, and model fit-to-map was validated by EMRinger53.
Analysis
Geneious Prime was used for sequence analysis. Flow cytometry data were processed using FlowJo v.10.5.0. GraphPad Prism 10 was used for data analysis. Structural analysis was performed using the software indicated above. For antibody analysis, a custom V(D)J database was used with modifications41.
Quantification and statistical analysis
Statistical information, including sample size (n), mean, standard deviation and statistical significance values, are indicated in the text or the figure legends. GraphPad Prism 10 was used for statistical analysis by paired Student’s t-test. Data were considered statistically significant at *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001 and ****P ≤ 0.0001.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Cryo-EM maps and models have been deposited to the PDB under accession codes PDM 9OED and PDM 9O8M. The mass spectrometry data have been deposited to the ProteomeXchange Consortium via the MassIVE partner repository (https://massive.ucsd.edu/ProteoSAFe/static/massive.jsp) under dataset identifier MSV000097767. Source data are provided with this paper.
Code availability
Computer code for macaque immunoglobulin gene analysis is publicly available on GitHub via https://github.com/stratust/igpipeline/tree/igpipeline2_timepoint_v2/ and https://github.com/MartaTarquis/blastN_pairs/.
References
Escolano, A. et al. Sequential immunization elicits broadly neutralizing anti-HIV-1 antibodies in Ig knockin mice. Cell 166, 1445–1458 (2016).
Kong, R. et al. Antibody lineages with vaccine-induced antigen-binding hotspots develop broad HIV neutralization. Cell 178, 567–584 (2019).
Caniels, T. G. et al. Germline-targeting HIV vaccination induces neutralizing antibodies to the CD4 binding site. Sci. Immunol. 9, eadk9550 (2024).
Landais, E. et al. Broadly neutralizing antibody responses in a large longitudinal sub-Saharan HIV primary infection cohort. PLoS Pathog. 12, e1005369 (2016).
Rusert, P. et al. Determinants of HIV-1 broadly neutralizing antibody induction. Nat. Med. 22, 1260–1267 (2016).
Kong, L. et al. Supersite of immune vulnerability on the glycosylated face of HIV-1 envelope glycoprotein gp120. Nat. Struct. Mol. Biol. 20, 796–803 (2013).
Freund, N. T. et al. Coexistence of potent HIV-1 broadly neutralizing antibodies and antibody-sensitive viruses in a viremic controller. Sci. Transl. Med. 9, eaal2144 (2017).
Mouquet, H. et al. Complex-type N-glycan recognition by potent broadly neutralizing HIV antibodies. Proc. Natl Acad. Sci. USA 109, E3268–E3277 (2012).
Garces, F. et al. Structural evolution of glycan recognition by a family of potent HIV antibodies. Cell 159, 69–79 (2014).
Escolano, A. et al. Immunization expands B cells specific to HIV-1 V3 glycan in mice and macaques. Nature 570, 468–473 (2019).
Steichen, J. M. et al. HIV vaccine design to target germline precursors of glycan-dependent broadly neutralizing antibodies. Immunity 45, 483–496 (2016).
Steichen, J. M. et al. A generalized HIV vaccine design strategy for priming of broadly neutralizing antibody responses. Science 366, eaax4380 (2019).
Saunders, K. O. et al. Vaccine elicitation of high mannose-dependent neutralizing antibodies against the V3-glycan broadly neutralizing epitope in nonhuman primates. Cell Rep. 18, 2175–2188 (2017).
Saunders, K. O. et al. Targeted selection of HIV-specific antibody mutations by engineering B cell maturation. Science 366, eaay7199 (2019).
Moore, P. L. et al. Evolution of an HIV glycan-dependent broadly neutralizing antibody epitope through immune escape. Nat. Med. 18, 1688–1692 (2012).
Molinos-Albert, L. M. et al. Anti-V1/V3-glycan broadly HIV-1 neutralizing antibodies in a post-treatment controller. Cell Host Microbe 31, 1275–1287 (2023).
Gieselmann, L. et al. Identification of a potent V3 glycan site broadly neutralizing antibody targeting an N332gp120 glycan-independent epitope. Nat. Immunol. https://doi.org/10.1038/s41590-025-02385-3 (2026).
Sanders, R. W. et al. A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS Pathog. 9, e1003618 (2013).
Bonsignori, M. et al. Staged induction of HIV-1 glycan-dependent broadly neutralizing antibodies. Sci. Transl. Med. 9, eaai7514 (2017).
Walker, L. M. et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 477, 466–470 (2011).
Barnes, C. O. et al. Structural characterization of a highly-potent V3-glycan broadly neutralizing antibody bound to natively-glycosylated HIV-1 envelope. Nat. Commun. 9, 1251 (2018).
Silva, M. et al. A particulate saponin/TLR agonist vaccine adjuvant alters lymph flow and modulates adaptive immunity. Sci. Immunol. 6, eabf1152 (2021).
Sajadi, M. M. et al. A comprehensive engineering strategy improves potency and manufacturability of a near pan-neutralizing antibody against HIV. Structure 33, 1150–1164.e8 (2025).
Simonich, C. A. et al. Kappa chain maturation helps drive rapid development of an infant HIV-1 broadly neutralizing antibody lineage. Nat. Commun. 10, 2190 (2019).
MacLeod, D. T. et al. Early antibody lineage diversification and independent limb maturation lead to broad HIV-1 neutralization targeting the Env high-mannose patch. Immunity 44, 1215–1226 (2016).
Swanson, O. et al. Rapid selection of HIV envelopes that bind to neutralizing antibody B cell lineage members with functional improbable mutations. Cell Rep. 36, 109561 (2021).
Kumar, S. et al. An HIV-1 broadly neutralizing antibody from a clade C-infected pediatric elite neutralizer potently neutralizes the contemporaneous and autologous evolving viruses. J. Virol. 93, e01495-18 (2019).
Steichen, J. M. et al. Vaccine priming of rare HIV broadly neutralizing antibody precursors in nonhuman primates. Science 384, eadj8321 (2024).
Escolano, A. et al. Sequential immunization of macaques elicits heterologous neutralizing antibodies targeting the V3-glycan patch of HIV-1 Env. Sci. Transl. Med. 13, eabk1533 (2021).
Cirelli, K. M. et al. Slow delivery immunization enhances HIV neutralizing antibody and germinal center responses via modulation of immunodominance. Cell 180, 206 (2020).
Alam, S. M. et al. Mimicry of an HIV broadly neutralizing antibody epitope with a synthetic glycopeptide. Sci. Transl. Med. 9, eaai7521 (2017).
Henderson, R. et al. Structural basis for breadth development in the HIV-1 V3-glycan targeting DH270 antibody clonal lineage. Nat. Commun. 14, 2782 (2023).
Wang, H. et al. Asymmetric recognition of HIV-1 Envelope trimer by V1V2 loop-targeting antibodies. eLife 6, e27389 (2017).
Zeng, W. F., Cao, W. Q., Liu, M. Q., He, S. M. & Yang, P. Y. Precise, fast and comprehensive analysis of intact glycopeptides and modified glycans with pGlyco3. Nat. Methods 18, 1515–1523 (2021).
Bern, M., Kil, Y. J. & Becker, C. Byonic: advanced peptide and protein identification software. Curr. Protoc. Bioinformatics 13, 13.20.1–13.20.14 (2012).
Lee, J. H. et al. Long-primed germinal centres with enduring affinity maturation and clonal migration. Nature 609, 998–1004 (2022).
Schiffner, T. et al. Vaccination induces broadly neutralizing antibody precursors to HIV gp41. Nat. Immunol. 25, 1073–1082 (2024).
von Boehmer, L. et al. Sequencing and cloning of antigen-specific antibodies from mouse memory B cells. Nat. Protoc. 11, 1908–1923 (2016).
Viant, C., Escolano, A., Chen, S. T. & Nussenzweig, M. C. Sequencing, cloning, and antigen binding analysis of monoclonal antibodies isolated from single mouse B cells. STAR Protoc. 2, 100389 (2021).
Williams, W. B. et al. Fab-dimerized glycan-reactive antibodies are a structural category of natural antibodies. Cell 184, 2955–2972 e2925 (2021).
Gaebler, C. et al. Evolution of antibody immunity to SARS-CoV-2. Nature 591, 639–644 (2021).
Diskin, R., Marcovecchio, P. M. & Bjorkman, P. J. Structure of a clade C HIV-1 gp120 bound to CD4 and CD4-induced antibody reveals anti-CD4 polyreactivity. Nat. Struct. Mol. Biol. 17, 608–613 (2010).
Sarzotti-Kelsoe, M. et al. Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV-1. J. Immunol. Methods 409, 131–146 (2014).
Montefiori, D. C. & Measuring, H. I. V. neutralization in a luciferase reporter gene assay. Methods Mol. Biol. 485, 395–405 (2009).
Scheres, S. H. RELION: implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Biol. 180, 519–530 (2012).
Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Patel, A., Toso, D., Litvak, A. & Nogales, E. Efficient graphene oxide coating improves cryo-EM sample preparation and data collection from tilted grids. Preprint at bioRxiv https://doi.org/10.1101/2021.03.08.434344 (2021).
Waterhouse, A. et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 46, W296–W303 (2018).
Meng, E. C. et al. UCSF ChimeraX: tools for structure building and analysis. Protein Sci. 32, e4792 (2023).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132 (2004).
Chen, V. B. et al. MolProbity: all-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66, 12–21 (2010).
Agirre, J. et al. Privateer: software for the conformational validation of carbohydrate structures. Nat. Struct. Mol. Biol. 22, 833–834 (2015).
Barad, B. A. et al. EMRinger: side chain-directed model and map validation for 3D cryo-electron microscopy. Nat. Methods 12, 943–946 (2015).
Acknowledgements
We thank members of the Escolano, Pallesen, Kulp and Weiner laboratories for valuable discussions; J. Faust, J. Fundyga and S. Majumdar at The Wistar Institute’s Flow Cytometry and Genomics cores for assistance; The Wistar Institute’s Animal Facility for outstanding mouse care services; M. Nussenzweig (The Rockefeller University) for providing the iGL PGT121 immunoglobulin knockin mice; and H. Mouquet (Institut Pasteur) for providing the nucleotide sequence of EPTC112 and advice to infer its germline precursor. We thank all the authors for critically reading the manuscript and providing valuable feedback. This work was supported by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health (NIH) Grant R00 AI140770-03 (to A.E.), 5 P30 AI045008-23 (to A.E.), P30 AI045008-24 (to A.E.), SAP no. 4100090384 (to A.E.), R01 AI172627-01A1 (to A.E.) and by the Gates Foundation INV-036995 (to A.E.). J.P. is supported by 5 U19 AI166916-03. D.B.W. is supported by U19 AI166916, BEAT-HIV UM1AI64570, and the W.W. Smith Charitable Trust Distinguished Professorship in Cancer Research. Mass spectrometry studies were supported by NIH grants R01GM130915 (to L.W.) and R01AI157854 (to L.W.), and a National Science Foundation Biofoundry: Glycoscience Research, Education, and Training Grant 2400220 (to L.W.). M.S.S. is supported by Gates Foundation grant INV-036842. Additional support included the Ching Jer Chern Postdoctoral Fellowship (to I.R.R.).
Author information
Authors and Affiliations
Contributions
I.R.R. designed, performed and analyzed experiments, and coordinated all experimental aspects of the project. J.D., Z.L., J.C. and J.P. designed, performed and analyzed Cryo-EM and EMPEM experiments. M.K., M.T.-M., E.U., C.B., S.G. and C.A. designed and performed experiments. M.W. coordinated immunizations in rhesus macaques, collected and prepared tissue and cell samples. C.L.L. performed and analyzed TZM-bl neutralization assays. P.Z. designed, performed and analyzed mass spectrometry experiments. R.H. provided proteins and advice. A.A.W. and M.B.M. produced SMNP adjuvant. N.S. created the WIN332 illustrations. G.M.S. and B.H.H. provided UCA and iGL antibodies, pseudoviruses, a macaque immunoglobulin gene reference and valuable advice for TZM-bl neutralization assays. D.I. provided SMNP adjuvant. L.W. supervised mass spectrometry experiments. D.W. supervised experiments, reviewed specific serological analysis with S.G. and provided valuable reagents. R.S.V. supervised macaque treatments and sample collection. M.S.S. supervised and analyzed TZM-bl neutralization assays. D.W.K. supervised the design and production of 7MUT-ST2-Asn332Tyr and provided valuable reagents and advice. J.P. supervised Cryo-EM and EMPEM studies. A.E. conceived the study, designed WIN332 and supervised and coordinated the study. I.R.R. and A.E. wrote the original draft, with contributions from P.Z., L.W., J.D., Z.L. and J.P. All authors contributed to draft reviewing and editing.
Corresponding author
Ethics declarations
Competing interests
A patent application including reagents and concepts of this study has been submitted by The Wistar Institute, with A.E. listed as inventor. Additionally, A.E. is named inventor on other patent applications related to the RC1 HIV-1 immunogen and antibodies elicited and isolated using RC1. A.E. has received financial compensation from The Rockefeller University for the licensed patent of RC1. The remaining authors declare no competing interests.
Peer review
Peer review information
Nature Immunology thanks Julià Blanco and the other, anonymous, reviewers for their contribution to the peer review of this work. Primary Handling Editor: P. Jauregui, in collaboration with the Nature Immunology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Biophysical and antigenic characterization of WIN332.
a. Size exclusion chromatography (SEC) showing elution volume for fractions containing WIN332. b. Negative staining electron microscopy (nsEM) 2D classes of WIN332 showing the expected propeller shape. c. Mass spectrometry analysis of WIN332 glycosylation. Pie charts show the glycan composition of WIN332 Potential N-glycosylation sites (PNGS). Gray pie charts indicate unoccupied PNGSs. The frequencies of high mannose, hybrid and complex-type glycans found at each PNGS are shown in green, dashed green-blue, and blue respectively. d. Antigenic profiling of WIN332 by ELISA. ELISA plots show the binding of different human bNAbs and the lack of binding of non-neutralizing antibodies to WIN332. Data is representative of at least 2 independent experiments. e. BLI experiment to evaluate the affinity of BF520 UCA for RC1. f. ELISA results showing WIN332 binding of the serum (n = 5) collected from iGL PGT121 Ig KI mice immunized with WIN332. PGT121 and serum from naïve mice were used as positive and negative controls respectively. g. ELISA controls for the experiments in Fig.1f. Data are presented as mean values + SD.
Extended Data Fig. 2 EMPEM analysis of the serum from WIN332-primed NHPs.
a, b. Negative staining electron microscopy (nsEM) densities of WIN332 Env (gray) bound to Fabs from NHP2 (blue and green) (a) and NHP3 (red) (b). WIN332 density is modelled on PDB6ORO. c,d. Comparison of the NHP2 (c) and NHP3 (d) top binder Fab densities with published Fabs EPTC112 (pink), BG18 (orange), and PGT121 (cyan). e. TZM-bl neutralization assay to determine the activity of the NHP serum in Fig. 2h against the control Murine Leukemia Virus (MLV).
Extended Data Fig. 3 Sequence identity between Ab1999 and Ab1983 with human V3-glycan bNAbs and prediction of glycan clashes.
a,b. Amino acid sequence alignment of Ab1999 HC with BG18 HC mt (a) and of Ab1999 LC with PCDN.76 UCA LC (b). c. Alignment of the BG505 SOSIP.664 trimer structure (PDB 8FR6) with the Ab1999-RC1 complex to predict potential clashes of Ab1999 with glycans. Potential clashes were primarily observed with glycans at Asn133 and Asn137, involving residues from CDRL1, CDRL2, CDRH2 and CDRH3. Specifically, Asn31 and Asn32 on CDRL1 and G68 on CDRL2 clashed with the N137 glycan, while Y52 on CDRH2 and E100E on CDRH3 showed clashes with the N133 glycan. d. Control TZM-bl assay using a non-neutralizing NHP antibody isolated from one of the WIN332-primed NHPs. e. Amino acid sequence alignment of Ab1983 HC with EPTC112 iGL HC. Dots represent identical amino acids. Dashes represent absent amino acids. f. Alignment of the BG505 SOSIP.664 trimer structure (PDB 8FR6) with the Ab1983-WIN332 complex to predict potential clashes of Ab1983 with glycans. Potential clashes were primarily associated with the N133, N137, and N156 glycans. Specifically, the CDRL2, CDRL3, CDRH1, CDRH2 and CDRH3 regions contributed to these clashes. The V52D in CDRL2 clashed with the N133 glycan. Residues G93, T94, and F95C in CDRL3, as well as Y33 in CDRH1 and Y52 in CDRH2, clashed with the N137 glycan. Furthermore, 15 residues within CDRH1, CDRH3 and framework1 exhibited steric overlap with the N156 glycan.
Extended Data Fig. 4 Gating strategy to isolate single WIN332-specific B cells from NHP LN biopsies after sequential immunization.
a. Flow cytometry plots showing the gating strategy used to isolate single WIN332-specific B cells from a pool of cell homogenates prepared from LN biopsies from sequentially immunized NHPs. b. TZM-bl neutralization assay to determine the activity of the NHP serum in Fig. 6h against the control Murine Leukemia Virus (MLV). c. Neutralization curves for the seven HIV-1 pseudoviruses showing low neutralization activity by the serum (n = 4) and for three representative pseudoviruses that were not neutralized. Data are presented as mean values + SD. Pie charts represent the V3-glycan epitope. Colored slices indicate present glycans.
Supplementary information
Supplementary Information (download PDF )
Supplementary Figs. 1 and 2 and Tables 1–5.
Source data
Source Data Fig. 1 (download XLSX )
Numerical source data.
Source Data Fig. 2 (download XLSX )
Numerical source data.
Source Data Fig. 3 (download XLSX )
Numerical source data.
Source Data Fig. 4 (download XLSX )
Numerical source data.
Source Data Fig. 5 (download XLSX )
Numerical source data.
Source Data Fig. 6 (download XLSX )
Numerical source data.
Source Data Extended Data Fig. 1 (download XLSX )
Numerical source data.
Source Data Extended Data Fig. 2 (download XLSX )
Numerical source data.
Source Data Extended Data Fig. 3 (download XLSX )
Numerical source data.
Source Data Extended Data Fig. 4 (download XLSX )
Numerical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Relano-Rodriguez, I., Du, J., Lin, Z.J. et al. Rapid elicitation of neutralizing Asn332-glycan-independent antibodies to the V3-glycan epitope of HIV-1 Env in nonhuman primates. Nat Immunol 27, 586–599 (2026). https://doi.org/10.1038/s41590-025-02408-z
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41590-025-02408-z
This article is cited by
-
Better and closer glycan-independent anti-V3 antibodies to help HIV-1 vaccine
Nature Immunology (2026)
