
Abstract
Studies on Bdellovibrio and like organisms (BALOs), obligate predatory bacteria, have highlighted the possibility of regulating bacteria and biofilms; however, yak-derived BALOs are yet to be reported. We aimed to characterize the BALOs isolated and identified from yak (Bos grunniens) feces and examine application potential. BALOs were isolated from healthy yak fecal samples, with Escherichia coli (ATCC 25922) as prey using the double-layer agar method, identified by transmission electron microscopy (TEM), and the specific 16S rDNA sequencing analysis. Sequencing of the 16S rDNA gene indicated that this isolate was 91% similar to the Bdellovibrio sp. NC01 reference strain and was named YBD-1. Proportion of YBD-1 lysed bacteria is 12/13. The YBD-1 showed best growth at 25–40°C, 0–0.25% (w/v) NaCl, and pH 6.5–7.5. YBD-1 significantly reduced the planktonic cells and biofilms of E.coli in co-culture compared to the E.coli group. Additionally, SEM analysis indicated that YBD-1 significantly reduced biofilm formation in E. coli. Furthermore, quantitative Real Time-polymerase chain reaction (qRT-PCR) showed that the expression of the virulence genes fim and iroN and the genes pgaABC involved in biofilm formation went down significantly. We concluded that YBD-1 may have the potential to control bacterial growth and biofilm-associated bacterial illnesses.
Similar content being viewed by others

Introduction
Antibiotics are extensively utilized to promote growth, illness prevention, and therapy in farmed animal industries1,2,3 Notably, the use of antibiotics often causes the emergence of drug resistance1, resulting in reduced or even extinguished efficacy of the therapeutic intervention against human and animal infections, causing a considerable challenge to public health, extensive food safety concerns, and environmental pollution4,5,6,7. Therefore, an urgent demand is for substitute antibiotic products to encourage animal development and avoid illnesses.
BALOs are Gram-stain negative bacteria that naturally invade and destroy Gram-stain negative and some Gram-stain positive pathogens8,9,10,11. They are widespread in natural and manufactured settings, including freshwater, seas, soils, plant rhizospheres, sewage, activated sludge, and human and animal intestines12. Since these predatory bacteria can eliminate a wide range of planktonic and biofilm bacteria without hurting higher eukaryotic species, they are being looked at as possible alternatives to antibiotics13 by significantly reducing their gene pools of antibiotic resistance14. Numerous in vitro toxicity studies and in vitro models have demonstrated that BALOs are not harmful13,15,16. Predatory bacteria are not only one of the most promising treatment options for multidrug-resistant diseases (MDR) infections16, but are also considered a significant mode of affecting bacterial community structure and microbial food webs15. Bacterial predation methods and characteristics have been thoroughly investigated in detail17,18. However, most current information is drawn from just a few model strains, resulting in a dearth of understanding of predatory bacterial species in the wild19.
Large ruminants known as yaks (Bos grunniens) are an essential source of life and income for highland farming communities in the Qinghai Tibet Plateau due to their strong ability to adapt to the region's high altitude, low oxygen concentration, low temperature, and lack of food20,21. Some researchers have even concluded that yaks have lived there for millions of years22. Yaks are not only used for farming and transportation but also provide residents with daily necessities, such as milk, meat, wool, leather, labor, and fuel, making them a vital mainstay industry for herders20,23,24. The ability of yaks to withstand harsh natural environments is related to their unique gastrointestinal micro-ecosystem21,22. Bacteria in the gastrointestinal system of yaks may have unique capacities due to changes in host species and living habitats21,25. Therefore, we were interested in isolating predatory bacterial BALOs from yaks.
The BALOs exhibit a lytic effect on many potential pathogens in nature, both planktonic cells and biofilms, by acting on specific virulence and biofilm-associated genes in the prey. Therefore, this study aimed to isolate and characterize BALOs from yak feces and to explore the prey range and lytic properties to assess the application potential of YBD-1.
Materials and methods
Collection, isolation, and purification of BALOs
Prey preparation
The bacterial strains employed in this investigation are enumerated in Table 1, and the strains without strain information are all laboratory isolates26. BALOs were isolated using E. coli ATCC 25,922 as prey. The prey bacterial (OD 2.0 at 600 nm) strains were cultivated for 1:100 in either Tryptic Soy Broth (TSB) or Luria–Bertani (LB) medium at 37°C with shaking at 220 rpm for 12–16 h (late exponential phase)27,28. After centrifuging the prey bacteria for 10 min at 4°C at 3000 g, the pellets were washed and resuspended in the diluted nutritional broth (DNB) (1:10 dilution of nutrient broth amended). After adjusting the prey cell suspensions to an optical density of around 2.0 at 600 nm (OD600) [~ 5 × 108 CFU/ml], they were kept in storage at 4°C28,29.
Sample collection, processing, and enrichment
Three samples of 20g each were collected from three-year-old adult healthy female yak (full of spirit, bright fur, free movement, no abnormality, and no signs of disease) fecal samples in Gannan of Gansu province, China, using sterile PE gloves, transferred to sterile 50 ml centrifuge tubes in dry ice, and sent immediately to the lab in 4h for further examination. One hundred milliliters of dilute nutrient broth (DNB) was added to 200 ml conical flask containing 20g samples, and to ensure that the organisms were distributed uniformly, the mixture was shaken with an orbital shaker set to 180 rpm for 1 h at 30°C29. Afterward, samples were incubated in an orbital shaker set to rotate at 180 rpm for an entire night at 30°C. The shaker was filled with 20 ml of washed E. coli prey cell suspension (prepared as mentioned above).
Isolation and purification with E.coli ATCC 25,922
Samples were enriched overnight and filtered with sterile gauze to remove partial residual prey cells and impurity particles. They were sequentially centrifuged at 1500 g for 5 min at 4°C and 3000 g for 5 min at 4°C. The supernatant efficiently concentrated Bdellovibrio28. The 10 ml supernatant was mixed with 20 ml DNB. Then, a sterile 10 ml centrifuge tube served to combine 0.3 ml of the supernatant with 0.5 ml of the prey (E.coli ATCC 25,922), followed by incubation at room temperature for 30 min. Subsequently, 4 ml of 0.7% molten diluted nutrient agar was added to the potential BALOs mixture. BALOs were isolated and purified using double-layer agar plating. The plates were incubated at 30°C for 12 days while plaque formation on DNB agar was observed29. Plaques visible on DNB plates after 2–3 d and progressively expanding for several days were thought to be probable BALOs. One of the biggest plaques was lifted (the plaque was first dug with the end of the 1 ml pipet tip and then picked with the tip of the pipet tip) from the plate to a sterile 5 ml centrifuge tube to combine 0.6 ml of the DNB with 0.6 ml of the prey (Suspension of prey cells with DNB), followed by incubation at room temperature for 30 min. Finally, 1 ml of the mixed incubation was pipetted into a sterile 10 ml test tube with 4 ml of 0.7% molten diluted nutrient agar, and purification was carried out following the same procedure with double-layer agar plating. The plaques obtained were purified by individual plaque isolation techniques in at least three successive platings until lytic plaques on an agar plate were homogeneous in size27,28,30,31, and not other bacterial colonies appeared on the plate. We believe that purified BALOs have been obtained.
Identification of the BALO
Transmission electron microscopy
10 µl recently lysed attack-phase (~ 108 PFU/ml) YBD-1 (the purified individual plaques were picked out and added to 100 ml of DNB containing 500 µl of E. coli ATCC 25,922 to incubate for 72 h at 30 °C, 180 rpm) suspension was placed on a Formvar carbon-coated 200-mesh copper microscope grid for 4 min at room temperature to determine the morphology of the BALOs. The excess liquid was removed using sterile filter paper. The samples were then counterstained with a 2% (w/v) phosphotungstic acid solution for 2 min before being viewed using a transmission electron microscope at a 100 kV accelerating voltage28 (Laboratory of Transmission Electron Microscopy, Lanzhou Veterinary Institute, Chinese Academy of Sciences).
16S rDNA
The BALOs' genomic DNA was extracted by centrifuging the co-culture twice at 3000 g for 5 min filtering the supernatant with a 0.45 µm bacterial filter to remove the remaining prey cells, then boiling the product for 10 min at 100°C and centrifuging at 12,000 g for 20 min. The supernatant obtained was used directly in the PCR reaction. PCR was used to amplify the Bdellovibrio 16S rDNA gene using primers specific to the Bdellovibrio 16S rDNA gene (63F primer: 5'-GAG GCC TAA CAC ATG CAA GTC-3'; 842R primer: 5'-CGW CAC TGA AGG GGT CAA-3')32. PCR amplification was performed by (Kodaq PCR MasterMix, Cat #G497) under the following conditions: predenaturation at 94°C for 10 min, 35 cycles of denaturation at 94 °C for 30s, annealing at 50°C for 30s, extension at 72°C for 48s, and final extension at 72°C for 10 s. The amplified products were analyzed in 1% agarose gel and identified with a UV transilluminator. The desired amplicon was extracted and purified using a Gel Purification Kit (TIANGEN, DP214-03) and sent for sequencing (Sangon Biotech, China, Shanghai).
Phylogenetic analysis
To clarify the specific evolutionary relationship of the isolate with other BALOs, the gene sequence of one representative BALO isolate was acquired and deposited in GenBank with the accession number OR186335 and named Bdellovibrio sp. strain YBD-1 (OR186335). The Basic Local Alignment Search Tool (BLAST)33 application compared data to the National Center for Biotechnology Information (NCBI) database. The YBD-1 isolate's sequences were aligned with reference strains using ClustalW34. MEGA 7.035 software was used to analyze the aligned sequences. The neighbor-joining method was used to build a phylogenetic tree. Bootstrap analysis with 1000 replications was used to determine the statistical significance levels of the inner nodes.
Prey range determination
A single pure plaque was excavated and mixed into a 100 ml culture containing bacterial prey cells (culture method was the same as prey preparation section, and LB medium was used) that had been washed with DNB using a pipette tip to assess the lytic range of YBD-1. The culture was shaken at 180 rpm for 6–7 days at 30°C on an orbital shaker until liquid clarity (predator stock for 5 × 108 and 5 × 109 PFU/ml). Then, the culture was centrifuged for 5 min at 3000 g, 4℃, discarded the pellet again, centrifugeed the supernatant, and followed the previous method. A few milliliters of DNB medium were resuspended from the pellet to a final concentration of 1 × 106 to 5 × 106 cells/ml (PFU/ml of the predator by double plate counting)36. The prey (Table 2) -to-predator ratio was adjusted to 10:1, and the double-layer plating technique was used to determine the prey range, as previously described26. Specific steps as shown in Fig. 1.

This is a flowchart for assessing of prey range of Bdellovibrio sp. YBD-1.
Lysis properties of YBD-1
Temperature, salinity, and pH effects on YBD-1 predation
The temperature impact on YBD-1 was evaluated using the procedure described in section "Prey range determination" to get YBD-1. 20 µl of YBD-1 was dropped onto an E.coli plate with filter paper, and temperatures were set to 15°C-50°C, with intervals of 5°C, and the control experiments were set up with the same temperature gradient substituting YBD-1 for PBS. The pH impact to YBD-1 was achieved at 30°C in the same way as mentioned for the determination of lytic to incubated on a shaker (180 rpm for 48 h at 30°C) to gain predator. Five milliliters of E. coli suspensions (~ 109 CFU/ml) contained 500 µl of predator in the attack phase (~ 108 PFU/ml). The mixed cultures were incubated in 50 ml of DNB for 6 d at 180 rpm at a range of pH values (5.0, 5.5, 6.0, 6.5, 7.0, 7.5, 8.0, 8.5, and 9.0)27,28, and the control experiment was set up with the same pH gradient in the absence of predators. MOPS was used as the medium buffer to maintain the experimental pH. The salinity effect was detected at 30℃ with varying salinity (0.0, 0.25%, 0.5%, and 1.0%)( ignore the salt concentration of the medium itself ) in the same procedure mentioned with 30 ml mixed cultures for the pH effect. At the same time, the PFU/ml of YBD-1 was detected at 0 h and 168 h. Specific steps as shown in Fig. 2.

This is the flowchart. The effect of temperature (A, 15℃–50℃, with intervals of 5℃), pH (B, 5.0–9.0, with intervals of 0.5), and . salinity(C, 0.0, 0.25%, 0.5%, and 1.0%) on lytic activity of Bdellovibrio sp. YBD-1.
Predation assays on planktonic cell cultures
As bacteria are mostly planktonic cells at the initial stage of infection, we measured the elimination of planktonic E. coli organisms by liquid culture with YBD-1, with slight modifications37. Briefly, 30 ml of DNB with 1 ml (1 × 108 CFU/ml) of E.coli and 1 ml of recently lysed attack-phase YBD-1 (suspend YBD-1 with phosphate-buffered saline (PBS)) suspension (1 × 107 PFU/ml) were incubated for 102 h at 30℃. A 30 ml DNB control culture containing E.coli at 1 × 108 CFU/ml and 1 ml of PBS. Aliquots (1 ml) of the cultures were collected at 12 h intervals starting at 0 h. In total, 900 µl was used to measure absorbance values at OD600, and the viable cells of E.coli on LB plates were calculated using 100 L of dilution.
Evaluation of YBD-1 impact on E. coli ATCC 25,922 biofilm
We evaluated the impact of YBD-1 on preformed biofilms and biofilm formation in E. coli. The effect of YBD-1 on preformed biofilms was determined, as described previously, with slight modifications38,39. Briefly, 200 µL of LB broth was added to a 96-well dish with an overnight culture of E. coli diluted 1:100 that was incubated at 30°C for 24, 48, and 72 h, and then washed using PBS to wipe out planktonic cells. Subsequently, 96-well dish was supplemented with 200 µl YBD-1 (1 × 108 PFU/ml) or DNB for 24h. Biofilm-formation experiments were also performed. Briefly, the 96-well plates were inoculated with 200 µl of E. coli as prey to resuspend YBD-1 (2 × 107 PFU/ml) as predator, and the plates were incubated for 24, 48, and 72 h. After incubation, the planktonic cell cultures and media components were removed using PBS. Then, the wells were dyed with 0.1% crystal violet (CV) for 20 min, and the unwashed dye was washed away with PBS by three washing steps. Finally, further determination was performed using the 30% acetic acid, and the OD600 was read as the absorbance of CV dye attached to the biofilm.
Scanning electron microscopy (SEM) characterization of E. coli ATCC 25,922 biofilms
As described previously, a biofilm of E, coli was established on a 28-mm cell crawler in a 6-well microtiter plate40,41. Briefly, the experimental wells of the treated biofilm group contained 1 ml of an overnight culture of E, coli diluted 1:100 in LB medium with 1 ml YBD-1 (1 × 108 PFU/ml). The control wells contained the same E, coli as above. The samples were fixed with 2.5% glutaraldehyde for 30 min at room temperature after 48 h of culture at 37°C. The samples were dehydrated gradient with ethanol (approximately 15 ml each), followed by drying at room temperature. Finally, the processed samples were sputtered with gold using an ion sputterer and viewed using an SEM at Wuhan Service Technology Co., Ltd. The voltage was adjusted to a value of 15 kilovolts (kV), and the samples were viewed with a magnification of 15,000 × .
Quantification of virulence and biofilm formation gene expression with qRT-PCR
The impact of YBD-1 on the expression of virulence and biofilm-forming genes in E. coli has been assessed using quantitative reverse transcription PCR (qRT-PCR), following the previously established method40, with slight modifications. E. coli were cultured in LB medium with or without YBD-1 for 24 h at 37°C to isolate RNA. Before extracting RNA, we centrifuged for 5 min at 3000 g to get sediment, then executed E. coli cell counting experiments, the final adjustment of E. coli concentration. TRIzol reagent (Thermo Fisher Scientific 15,596,026) was used to extract total RNA from 1ml of culture. According to the manufacturer's instructions, the extracted RNA samples were reverse transcribed into cDNA using a cDNA synthesis kit ( TaKaRa RR047A) for qRT-PCR analysis. qRT-PCR was used to measure gene expression levels using a Recycle 96 system (Light Cycler 96) and the ChammQ SYBR qPCR Master Mix (Vazyme Q311-02), with the primers mentioned in Table 3. The 16S rRNA gene of E.coli was employed as an internal control to standardize the data42.
Statistical analysis
MEGA 7 software was used to build the phylogenic tree, and Originpro 2019b and GraphPad Prism 8.0 were used to plot the diagram. The data was presented as mean ± standard deviation (SD) of at least three independent experiments. With SPSS 27.0, statistical analysis was carried out. One-way analysis of variance with the least significant difference (LSD) and multiple comparisons with Waller-Duncan treatment were carried out. It was deemed statistically significant when P < 0.05.
Results
Isolation and identification for B. bacteriovorus
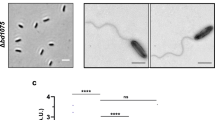
The YBD-1 was isolated from the Yak feces sample. Within 7 days, several lytic areas of E. coli prey were formed on the law of incubation at 30°C by the double agar plates technique (Supplementary Fig. 1). One of the larger plaques was chosen for further purification analysis in the same manner as for isolation. In purification experiments, we observed clear circular plaques of uniform size with uniform borders on the plates in Fig. 3A. TEM indicated that the isolated YBD-1 was a tiny comma cell (). The specific 16S rRNA gene was amplified using PCR with the primers (63F-842R) for further identification, and the amplified fragments (830 bp, Fig. 4A) were sequenced and revealed to be 91% similar in sequence to the Bdellovibrio sp. NC01 reference strain from GenBank33. A comparison of the phylogenetic tree analysis with the eleven BALO reference strains in GenBank revealed that the strain YBD-1 was near to distance as Bdellovibrio sp (Fig. 4B). The nucleotide sequence of 16S rDNA from the YBD-1 isolate has been deposited in the GenBank database with the following accession number: Bdellovibrio sp. strain YBD-1 (OR186335)45.

(A) Lytic plaques developed by YBD-1 on the lawn of E. coli prey cells. (B) Morphological identification of isolated YBD-1 in the attack phase using a transmission electron microscope(TEM). The scale bar represents 2.0μm.

(A) Agarose gel electrophoresis of PCR assay using universal specific 16S rRNA gene primers (63F-842R), 1: DNA ladder marker(2 k bp), 2: E.coli negative control, 3: YBD-1 genomic DNA. (B) The phylogenetic tree is based on the specific 16S rRNA gene sequences of Bdellovibrio. The tree was constructed using the Neighbour Joining method and the p-distance model. Bootstrap confidence was calculated from 1000 replicates.
Effect of temperature, salinity, and pH on lysis properties
Temperature (15°C–50°C), salinity (0.25–1%), and pH (5.5–9.0) were tested as typical environmental parameters on the predation activities of the isolated strains YBD-1. The method is to determine inhibition zone diameters with filter papers containing strain YBD-1 at 15°C, 20°C, 25°C, 30°C, 35°C, 40°C, 45°C, and 50°C. The largest diameter for the zone of inhibition was observed for strain YBD-1 at 25°C − 40°C and occurred within 5 d. At higher and lower temperatures, the circle of inhibition gradually decreased (Fig. 5, Table 2)28. Incubation experiments at the salinities ranging from 0%, and 0.25% salinity gradually decreased lytic ability; lytic ability is nearly aborted completely by 0.5% NaCl based on Fig. 6B, while at 1% NaCl, even YBD-1 viability based on PFU numbers (Fig. 6B) is decreased, indicating that lower salinity was optimal for YBD-1 (Fig. 6A, B). The strain YBD-1 exhibited its highest lytic activity at pH 6.5–7.5 within 96 h. There was no lytic effect at pH 5.5 and 9.0 (Fig. 7). We can see that E.coli & YBD-1 has always a higher OD600nm at 12h compared to E.coli alone.

The effect of temperature on the lytic activity of YBD-1 indicated by changes in the plaque diameter(mm) on the double-layer agar plating culture. (A: 15℃; B: 20℃; C: 25℃; D: 30℃; E: 35℃; F: 40℃; G: 45℃; H: 50℃).

(A) The effect of salinity on the lytic activity of YBD-1 indicated by changes in OD600 in the broth co-cultures(30℃, pH 7.0 for 168 h). (B) Number of YBD-1 cells (log10PFU/ml) for initial starting time point 0 h, and final point 168 h. Salinity concentrations were tested at 0, 0.25, 0.5, and 1% NaCl, it is the same for Fig. 4A and B. All experiments were conducted three times independently. Statistical analysis was performed using two-way ANOVA. The amount of asterisks(*) obtained from SPSS is directly related to statistical significance(***: p < 0.001, ns: not significant).

(A) The effect of pH on the and lytic activity of YBD-1 indicated by changes in OD600 in the broth co-cultures(30 ℃, pH 7.0 for 96 h). pH range was tested at 5.0, 5.5, 6.0, 6.5, 7.0, 7.5, 8.0, 8.5, and 9.0. (B) Number of E.coli prey cells(log10CFU/ml) with/ without YBD-1 for pH5.0,7.0,9.0 at 48 h. All experiments were conducted three times independently. Statistical analysis was performed using two-way ANOVA. The amount of asterisks(*) obtained from SPSS is directly related to statistical significance(***: p < 0.001).
Predation assays on planktonic cell cultures
Predation experiments on E. coli planktonic growth with and without YBD-1 indicated that YBD-1 exhibited predatory activity against E. coli. Predation viability was evaluated by determining the optical density and prey bacteria CFU/ml at 12 h intervals for 108 h. Figure 8 shows that after 12 h of co-culture with the predator YBD-1, there was a significant decrease in E. coli count compared to the control37.

(A) OD600 of E.coli in co-culture with YBD-1; (B) log 10 CFU/ml of E.coli in co-cultures with YBD-1. (C) log10FUP/ml of YBD-1 in co-cultures with E.coli. All experiments were conducted three times independently. Statistical analysis was performed using a paired t-test. The amount of asterisks(*) obtained from SPSS is directly related to statistical significance(***: p < 0.001).
Impact on preformed biofilms and biofilm formation
The impact of YBD-1 on preformed biofilms of E. coli was evaluated by dyeing with CVfor 24, 48, and 72 h biofilms incubated with a suspension of YBD-1 for 24 h. A significant reduction in biofilm biomass was observed compared to untreated control. Similarly, YBD-1 prevented the development of E. coli biofilms when co-cultured with predators and prey for 24, 48, and 72h, and compared to the control group, biofilm formation was significantly decreased (Fig. 9).

Bdellovibrio sp.YBD-1 predation assay on E. coli biofilm. (A) 24-h-preformed E.coli biofilm was treated with or without the predator(YBD-1) for 24 h; (B) 48-h-preformed E.coli biofilm was treated with or without the predator(YBD-1) for 48 h; (C) 72-h-preformed E.coli biofilm was treated with or without the predator(YBD-1) for 48 h. biofilm formation by E.coli after exposure to no or YBD-1 for 24 h (D), 48 h (E), or 72 h (F) All experiments were conducted three times independently. Statistical analysis was performed using a t-test. The amount of asterisks(*) obtained from SPSS is directly related to statistical significance(***: p < 0.001).
Scanning electron microscopy (SEM)
To determine if YBD-1 affected the structure of the biofilms, we performed SEM investigations. The results showed that E. coli treated without YBD-1 form biofilms than E. coli treated with YBD-1 (Fig. 10).

SEM image. (A) E.coli biofilm formation untreated with YBD-1 for 48-h; (B) E.coli after 48-h exposure to YBD-1.
qRT-PCR quantification of virulence and biofilm formation gene expression
It has been proven that virulence factors contribute to E. coli pathogenicity46, and the first step for BALOs to lyse E. coli is through the action of the bacterial outer membrane15. The expression of virulence and biofilm-forming genes was studied (Table 3) in E. coli incubated with YBD-1 using qRT-PCR. The findings demonstrated a significant reduction in the expression levels of virulence and biofilm-forming genes in cells from cultures treated with YBD-1 compared to those in untreated cells (Fig. 11).

Expression ratio of E. coli virulence and biofilm formation genes in the presence or absence of YBD-1. All experiments were conducted three times independently. Statistical analysis was performed using two-way ANOVA. The amount of asterisks(*) obtained from SPSS is directly related to statistical significance(***: p < 0.001; **: p > 0.001). The 16S rRNA gene of E.coli was used as an internal control to normalize the data.
Discussion
The emergence and use of antibiotics have provided enormous welfare for humans; however, their abuse also pollutes the human environment and causes significant health hazards. That is to say, the use of antibiotics does inhibit the occurrence of many diseases, but it has developed resistance to many pathogens. Therefore, researchers are interested in exploring alternative antibiotics, and predatory bacteria have become a focus of research.
Yaks, a rare species endemic to plateau regions, can adapt to harsh natural environments. Although predatory bacteria or BALOs have been studied in other animals13,47, they have not been reported in yak species. Therefore, this study aimed to isolate and characterize BALOs from yak feces and evaluate their potential application. We isolated a bacterial strain from yak feces that was consistent with the results of other studies and was also a new species. To assess the application potential of YBD-1, we conducted a few studies on lysis range, characteristics, biofilms, virulence, and biofilm formation gene expression.
Our procedure for successfully isolating and purifying YBD-1 is rather difficult, and various attempts were made in the early stages. In the first attempt the enrichment cultures passed subsequently through a 0.45 µm Millex filter37,44; The second try included centrifugation at different speeds, through 1.2 µm or 0.8 µm and 0.45 µm30,31; Finally, a protocol27,48 was performed involving centrifugation, filtration, and re-centrifugation. Transparent plaques were not observed on double-layer agar plates. After the enrichment culture, a sterile gauze was used to filter out large particles and centrifuged at 1500 g and 3000 g for 5 min. Eventually, we observed transparent plaques on the double-layer plate after approximately 5 d, which gradually increased in plaque size with the increase in days and finally formed transparent plaques with more regular borders (Fig. 3A). We suspect that other centrifugation and filtration methods may have removed the other prey while removing growth-dependent BALOs, leaving a minimal number of progeny BALOs. The viability of BALOs gradually disappeared during the purification process, resulting in the absence of BALO plaques. Similarly, we did not obtain transparent plaques when we selected single plaques for subculturing. We observed transparent plaques when we reduced the proportion of prey bacteria, and we hypothesized that the most important reason could be that the ratio of BALOs to the host reached a better ratio, as some earlier studies have shown that different ratios of prey can lead to different lysing affections48,49.
The strain identity was confirmed by electron microscopy (Fig. 3)and the Bdellovibrio-specific primers to amplify Bdellovibrio-specific 16S rDNA32. Evolutionary tree construction revealed that the strain YBD-1 was near to distance as Bdellovibrio sp, but is a new genus because its similarity is only 91% to the Bdellovibrio sp. NC01 (Fig. 4). The strain YBD-1 is 90.9% to Bdellovibrio bacteriovorus strain SDWB-DG8. Electron microscopy results showed that the isolated YBD-1 was comma-shaped.
The ability of Bdellovibrio sp. strain YBD-1 to work as a live antibiotic against infections is demonstrated by its bacteriolytic spectrum. We selected four Gram-stain negative and nine Gram-positive strains (Table 1) to determine the ability to lyse a wide range of pathogenic bacteria. S. aureus RN 4220 was absent from lysing plaques among the tested bacteria (Table 1, Supplementary Fig. 2). The predator must first approach and recognize suitable prey, then breach the outer membrane and colonize the prey’s periplasm. Once the breaching has occurred, prey and predator form a characteristic round-shaped structure called bdelloplast47. S.aureus RN 4220 has a mutation in the gene sau1 hsdR, which can cause RN 4220 to become a gene defect type of the restriction-modification system. Therefore, it is likely that the genetic defect of RN 4220 causes Bdellovibrio to fail to recognize prey or that the site on the outer membrane of RN 4220 that contacts Bdellovibrio fimbriae cannot be activated. Based on the size, transparency, and edge regularity of the lysis plaques, we noticed that the ability of YBD-1 to lyse Gram-stain negative bacteria was more potent than that of Gram-stain positive bacteria, and the lysis of E. coli ATCC 25,922 was the most effective. Therefore, subsequent experiments were conducted using E. coli ATCC 25,922. For strongly pathogenic bacteria, Streptococcus pyogenes ATCC 19,615, Acinetobacter baumannii, Escherichia coli ATCC 700,728, and Staphylococcus aureus TCH 1516 also have different lysis effects (Supplementary Fig. 2), which seem to have a more promising discovery for application, and we will focus on these critical pathogenic bacteria in future studies. Furthermore, researchers have concluded that BALOs are exclusive predators of Gram-negative bacteria50,51,52. However, in recent years, studies have rarely found that Gram-stain positive bacteria can also be lysed by BALOs11,53. There is some, but limited evidence that some Gram-stain positive organisms might be attached/killed. Therefore, YBD-1 lyses the tasted Gram-stain negative and partial-stain positive bacteria, according to our initial double-layer agar plating results. Our future research will mostly focus on studying Gram-stain positive species as prey. There have also been no reports of BALOs invading mammalian cells54,55. The proportion of YBD-1 lysed bacteria is 12/13, and the broad lysis spectrum supports the heterogeneity of BALOs from different sources. BALOs was isolated from different sources, which is the most trustworthy data confirming its safety and potential probiotic activity in animals and humans. Therefore, YBD-1 may be able to control a variety of bacterial infections.
The effects of temperature, salinity, and pH on YBD-1 predation efficiency were investigated. The observed temperature range of 25°C–45°C (Fig. 5, Table 2) and pH range of 6.0–8.5 are consistent with the results of earlier research28,31, but the ranges have expanded, which may be attributed to the different settings of the gradient for the determination of the temperature and pH as well as the difference 0–0.5% (Fig. 6A, B), in contrast to the optimal salinity of 2% reported in Kongrueng et al.36. However, the source of their samples was an aqueous environment, and it is reasonable that the salinity was also appropriately raised. The range of salinity detected by Li et al.29 was 1–4.5%, which was also shown to be isolated from a marine brine system.
Considering the final application pathway of YBD-1, we examined the lytic effect on planktonic E. coli in liquid culture. Compared to the control group, YBD-1 had a significant lytic effect on planktonic E. coli (Fig. 8). Biofilms increase bacterial tolerance to antibiotics, the environment, and host immune system attacks and provide a protective environment for pathogenic bacteria47; BALOs have been shown to act on biofilms40. Biofilm-forming bacteria and fungi have been estimated to cause 80% of all infections44. Therefore, we first proposed using BALOs to lyse E. coli biofilms. E. coli exhibited a decrease in biofilm biomass contrary to the control group. We determined the effectiveness of YBD-1 in disrupting preformed biofilms and biofilm formation using a 96-well plate, and YBD-1 showed a significant lytic effect on each treatment group (Fig. 9). In addition, to better visualize the biofilm pattern, we observed the predation behavior of YBD-1 using SEM (Fig. 8), and the results support and extend the results of earlier studies38,40,41,42,44,45,46,47.
It has been demonstrated that the pathogenicity of E. coli isolates is partially caused by virulence factors. IroN gene is a necessary component encoding iron acquisition systems for the survival of bacteria. During an infection, bacteria have evolved several methods for acquiring iron, including an iron acquisition system43,44. The type 1 fimbria necessary for bacterial adherence and bladder epithelial cell invasion is encoded by the fim gene cluster43,56. The pgaABC locus is necessary for biofilm formation in E. coli. We found a significant decrease in the expression of pgaA, pgaB, and pgaC after YBD-1 exposure in E. coli (Fig. 11)44,46. We again evaluated that BALO-related isolates inhibited the viability of E. coli when treated with YBD-140. Therefore, we hypothesized that YBD-1 kills E. coli by inhibiting virulence and biofilm formation genes.
Conclusion
The YBD-1 was isolated from yak feces and identified by TEM, PCR, and the 16Sr DNA gene sequencing analysis in this study, indicating the isolate as Bdellovibrio sp. YBD-1. Several experiments were conducted to investigate the application potential of the isolate by determining the range of prey, lysis characteristics of YBD-1, planktonic cells, and biofilm lysis, and identifying prey bacteria's virulence genes and biofilm formation-related genes. All results indicate that isolate YBD-1 has the potential to control bacterial growth and biofilm-associated bacterial infections. This work delivers in-depth insights into the differences in predation characteristics among wild-type environmental predation bacterial strains, which provides more possibilities for the development of alternative antibiotic products.
Data availability
The nucleotide sequence of 16S rDNA from the YBD-1 isolate has been deposited in the GenBank database with the following accession number: Bdellovibrio sp. strain YBD-1 (OR186335). The datasets presented in the study are available in the article/Supplementary Material. Further questions should be forwarded to the corresponding authors.
References
Xu, C. et al. A review of current bacterial resistance to antibiotics in food animals. Front. Microbiol. 13, 822689 (2022).
Brown, K. et al. Antimicrobial growth promoter use in livestock: a requirement to understand their modes of action to develop effective alternatives. Int. J. Antimicrob. Agents. 49, 12–24 (2017).
Rahman, M. et al. An overview of antimicrobial stewardship optimization: The use of antibiotics in humans and animals to prevent resistance. Antibiotics 11, 00667 (2022).
Allen, H. K. et al. Treatment, promotion, commotion: antibiotic alternatives in food-producing animals. Trends Microbiol. 21, 114–119 (2013).
Laxminarayan, R. et al. Antibiotic resistance—the need for global solutions. Lancet Infect. Dis. 13, 1057–1098 (2013).
Stanton, T. B. A call for antibiotic alternatives research. Trends Microbiol. 21, 111–113 (2013).
Laxminarayan, R. et al. The lancet infectious diseases commission on antimicrobial resistance: 6 years later. Lancet Infect. Dis. 20, e51–e60 (2020).
Vatansever, S. K. et al. Effect of predator bacteria bdellovibrio bacteriovorus on clinical pathogens and biofilms. Indian J Microbiol 63, 139–145 (2023).
Monnappa, A. K. et al. Bdellovibrio bacteriovorus inhibits staphylococcus aureus biofilm formation and invasion into human epithelial cells. Sci. Rep. 4, 3811 (2014).
Dashiff, A. & Kadouri, D. E. Predation of oral pathogens by Bdellovibrio bacteriovorus 109J. Mol. Oral Microbiol. 26, 19–34 (2010).
Im, H., Dwidar, M. & Mitchell, R. J. Bdellovibrio bacteriovorus HD100, a predator of Gram-negative bacteria, benefits energetically from Staphylococcus aureus biofilms without predation. ISME J. 12, 2090–2095 (2018).
Ezzedine, J. A., Desdevises, Y. & Jacquet, S. Bdellovibrio and like organisms: current understanding and knowledge gaps of the smallest cellular hunters of the microbial world. Crit. Rev. Microbiol. 48, 428–449 (2021).
Atterbury, R. J. & Tyson, J. Predatory bacteria as living antibiotics – where are we now?. Microbiol. 167, 001025 (2021).
Upatissa, S., Mun, W. & Mitchell, R. J. Pairing colicins B and E5 with bdellovibrio bacteriovorus to eradicate carbapenem- and colistin-resistant strains of Escherichia coli. Microbiol Spectr. 11, e0017323. https://doi.org/10.1128/spectrum.00173-23 (2023).
Mookherjee, A. et al. Interactions between Bdellovibrio and like organisms and bacteria in biofilms: Beyond predator–prey dynamics. Environ. Microbiol. 24, 998–1011 (2021).
Waso, M. et al. Insights into Bdellovibrio spp. mechanisms of action and potential applications. World J Microbiol. Biotechnol. 37, 85 (2021).
Negus, D. et al. Predator versus pathogen: How does predatory bdellovibrio bacteriovorus interface with the challenges of killing gram-negative pathogens in a host setting?. Annu. Rev. Microbiol. 71, 441–457 (2017).
Kaplan, M. et al. Bdellovibrio predation cycle characterized at nanometre-scale resolution with cryo-electron tomography. Nat. Microbiol. 8, 1267–1279 (2023).
Lai, T. F. et al. Advances in cellular and molecular predatory biology of Bdellovibrio bacteriovorus six decades after discovery. Front. Microbiol. 14, 1168709 (2023).
Goshu, H. et al. Copy number variations of KLF6 modulate gene transcription and growth traits in Chinese Datong Yak (Bos Grunniens). Anim. 8, 145 (2018).
Jiang, F. et al. Isoacids supplementation improves growth performance and feed fiber digestibility associated with ruminal bacterial community in yaks. Front. Microbiol. 14, 1175880 (2023).
Zeng, Z. et al. Probiotic properties of Bacillus proteolyticus Isolated From Tibetan Yaks. China. Front. Microbiol. 12, 649207 (2021).
Jing, X. et al. The adaptive strategies of yaks to live in the Asian highlands. Anim. Nutr. 9, 249–258 (2022).
Li, J. et al. Seroprevalence of Bluetongue virus in domestic yaks (Bos grunniens) in Tibetan regions of China based on circulating antibodies. Trop. Anim. Health Prod. 47, 1221–1223 (2015).
He, Y. et al. Probiotic Potential of Bacillus amyloliquefaciens Isolated from Tibetan Yaks. Probiotics Antimicrob Proteins https://doi.org/10.1007/s12602-022-10027-5 (2022).
Inoue, D. et al. Characterization of two novel predatory bacteria, Bacteriovorax stolpii HI3 and myxococcus sp MH1 Isolated from a freshwater pond: Prey range and predatory dynamics and efficiency. Microorganisms 10, 1816 (2022).
Lei, J. et al. Screening, identification, and probiotic properties of bacillus pumilus From Yak. Probiotics Antimicrob. Proteins. https://doi.org/10.1007/s12602-023-10054-w (2023).
Odooli, S. et al. Characterization of the first highly predatory Bdellovibrio bacteriovorus from Iran and its potential lytic activity against principal pathogenic Enterobacteriaceae. Iran. J. Basic Med. Sci. 23, 1275–1285 (2020).
Li, H., Liu, C., Chen, L., Zhang, X. & Cai, J. Biological characterization of two marine Bdellovibrio-and-like organisms isolated from Daya bay of Shenzhen, China and their application in the elimination of Vibrio parahaemolyticus in oyster. Int. J. Food Microbiol. 151, 36–43 (2011).
Stolp, H.a.S., M. P. Bdellovibrio bacteriovorus gen. etsp. n., a predatory, ectoparasitic, and bacteriolytic microorganism. Antonie Van Leeuwenhoek. 29, 217–248 (1963).
Chu, W. H. & Zhu, W. Isolation of bdellovibrio as biological therapeutic agents used for the treatment of aeromonas hydrophila infection in fish. Zoonoses Public Health. 57, 258–264 (2009).
Jurkevitch, E., Minz, D., Ramati, B. & Barel, G. Prey range characterization, ribotyping, and diversity of soil and rhizosphere Bdellovibrio spp. isolated on phytopathogenic bacteria. Appl. Environ. Microbiol. 66, 2365–2371 (2000).
Altschul, S. F. et al. Basic local alignment search tool. J Mol Biol. 215, 404–410 (1990).
Li, K.-B. ClustalW-MPI: ClustalW analysis using distributed and parallel computing. Bioinf. 19, 1585–1586 (2003).
Kumar, S., Stecher, G. & Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874 (2016).
Kongrueng, J. et al. Isolation of Bdellovibrio and like organisms and potential to reduce acute hepatopancreatic necrosis disease caused by Vibrio parahaemolyticus. Dis Aquat Organ 124, 223–232 (2017).
Jurkevitch, E. Isolation and classification of bdellovibrio and like organisms. Curr. Protoc. Microbiol. 26, 7B (2012).
He, H.-J. et al. Erythromycin resistance features and biofilm formation affected by subinhibitory erythromycin in clinical isolates of Staphylococcus epidermidis. J. Microbiol. Immunol. Infect. 49, 33–40 (2016).
Dashiff, A. et al. Predation of human pathogens by the predatory bacteria Micavibrio aeruginosavorus and Bdellovibrio bacteriovorus. J. Appl. Microbiol. 110, 431–444 (2011).
Bonfiglio, G. et al. Growth control of adherent-invasive Escherichia coli (AIEC) by the predator bacteria bdellovibrio bacteriovorus: A new therapeutic approach for crohn’s disease patients. Microorganisms 8, 17 (2019).
Gu, Y. et al. Development of resistance in Escherichia coli ATCC25922 under exposure of sub-inhibitory concentration of olaquindox. Antibiotics (Basel) 9, 791 (2020).
Liu, G. et al. Inhibition of virulence gene expression in salmonella dublin, Escherichia coli F5 and clostridium perfringens associated with neonatal calf diarrhea by factors produced by lactic acid bacteria during fermentation of cow milk. Front. Microbiol. 13, 828013 (2022).
Paniagua-Contreras, G. L. et al. Comprehensive expression analysis of pathogenicity genes in uropathogenic Escherichia coli strains. Microb. Pathog. 103, 1–7 (2017).
Dong, G. et al. Effects of sub-minimum inhibitory concentrations of ciprofloxacin on biofilm formation and virulence factors of Escherichia coli. Braz. J. Infect. Dis. 23, 15–21 (2019).
Zhang, Q. et al. In vitro investigation on lactic acid bacteria isolatedfrom Yak faeces for potential probiotics. Front. Cell. Infect. Microbiol. 12, 984537 (2022).
Wang, X., Preston, J. F. & Romeo, T. The pgaABCD locus of Escherichia coli promotes the synthesis of a polysaccharide adhesin required for biofilm formation. J. Bacteriol. 186, 2724–2734 (2004).
Cavallo, F. M. et al. Bdellovibrio bacteriovorus: A potential ‘living antibiotic’ to control bacterial pathogens. Crit. Rev. Microbiol. 47, 630–646 (2021).
Otto, S., Harms, H. & Wick, L. Y. Effects of predation and dispersal on bacterial abundance and contaminant biodegradation. FEMS Microbiol Ecol. 93(2), fwi241 (2017).
Sathyamoorthy, R. et al. Effects of the prey landscape on the fitness of the bacterial predators Bdellovibrio and like organisms. FEMS Microbiol Ecol. 97(5), fiab047 (2021).
Banks, E. J. et al. Asymmetric peptidoglycan editing generates cell curvature in Bdellovibrio predatory bacteria. Nat. Commun. 13, 1509 (2022).
Bratanis, E. et al. Biotechnological Potential of Bdellovibrio and Like Organisms and Their Secreted Enzymes. Front. Microbiol. 11, 662 (2020).
Duncan, M. C. et al. High-throughput analysis of gene function in the bacterial predator bdellovibrio bacteriovorus. mBio. https://doi.org/10.1128/mBio.01040-19 (2019).
Iebba, V. et al. Bdellovibrio bacteriovorus directly attacks Pseudomonas aeruginosa and Staphylococcus aureus Cystic fibrosis isolates. Front. Microbiol. 5, 280 (2014).
Shatzkes, K. et al. Examining the efficacy of intravenous administration of predatory bacteria in rats. Sci. Rep. 7, 1864 (2017).
Shatzkes, K. et al. Examining the safety of respiratory and intravenous inoculation of Bdellovibrio bacteriovorus and Micavibrio aeruginosavorus in a mouse model. Sci. Rep. 5, 12899 (2015).
Bower, J. M., Eto, D. S. & Mulvey, M. A. Covert operations of uropathogenic Escherichia coli within the urinary tract. Traffic. 6, 18–31 (2005).
Funding
This work was supported by the National Natural Science Foundation of China (32160850), Fuxi Foundation for Youth Talent at Gansu Agricultural University (No. Gaufx‐02Y10), the Innovative Talent Project of Gansu Province (2022LQTD59), the grants from the outstanding youth fund of Gansu Province (No. 20JR10RA561), and the Scientific research foundation for the new Scholars at Gansu Agricultural University (No. GSAU‐RCZX201701).
Author information
Authors and Affiliations
Contributions
YX, QyZ, MW, and YyP contributed to the conception and design of experiments. YX and ML participated in the preparation of materials, bacteria isolation, and data analysis. The author of the manuscript is YX. The manuscript was subject to QyZ, MW, and YyP's critical revision and final approval. All authors contributed to and authorized the submitted version of the article.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xi, Y., Pan, Y., Li, M. et al. Evaluation of the application potential of Bdellovibrio sp. YBD-1 isolated from Yak faeces. Sci Rep 14, 13010 (2024). https://doi.org/10.1038/s41598-024-63418-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-63418-9
