
Abstract
The potential risk posed by infectious agents (IAs) associated with netpen aquaculture to wild fishes is determined based on the “release” of IAs from netpens into the environment, the “exposure” of the wild fish to those released agents, and the “consequence” for wild fish experiencing infection by those agents. Information available to characterize these three factors is often lacking, and the occurrence of transmission from aquaculture to wild fish as well as potential consequences of such transmission are difficult to observe. In this study, we utilized environmental DNA (eDNA) to characterize the release of dozens of IAs from, and exposure of Pacific salmon to, Atlantic salmon aquaculture. We combined these factors with the consequence of infection, as determined by the literature, to identify IAs that may pose a risk to wild salmon exposed to aquaculture in British Columbia, Canada. Over an 18-month period, eDNA samples were collected from seven active and four inactive netpen aquaculture sites in the Broughton Archipelago, BC. A meta-analytical mean across 22 IAs showed that the odds of IA detection at active sites was 4.3 (95% confidence interval = 2.3:8.1) times higher than at inactive sites, with 11 IAs in particular demonstrating a pattern consistent with elevated release. Oncorhynchus tshawytscha was the only Pacific salmon species presenting eDNA detections more likely to occur around and within active netpens relative to inactive sites. After considering the evidence of negative consequences of infection (from previous literature) in tandem with release model results, we determined that Tenacibaculum maritimum, Tenacibaculum finnmarkense, Ichthyobodo spp., and Piscine orthoreovirus are potential risks to Pacific salmon exposed to marine netpen aquaculture. These IAs, and others demonstrating patterns consistent with release but with insufficient prior research to evaluate the consequences of infection, require further studies that identify the factors influencing the intensity of release, the spatial extent of release around netpens, and the prevalence of infection in wild fish within known distances from netpens.
Similar content being viewed by others
Introduction
Domestic animal production facilities can be difficult to isolate from their surrounding ecosystems. This is especially true in the case of infectious agents (IAs), which are sometimes carried into and out of facilities by wildlife vectors or even the flow of air or water. The introduction of IAs from the surrounding environment to animals within culture facilities is well-studied due to its highly visible (and sometimes economically catastrophic) impacts on cultured animal populations. Some examples of such “wild-domestic transmission” (sometimes referred to as “spillover”, a term we avoid here due to usage ambiguity) include Brucellosis transmitted from wild ungulates to cattle in North America1 and avian influenza transmitted from migratory aquatic birds to poultry2. The converse pattern, “domestic-wild transmission”, occurs when IAs amplified in dense, captive animal populations infect wild animals in ecosystems adjacent to culture facilities. Since wild animal populations are free-roaming, the impacts of domestic-wild transmission are much more difficult to observe, and any negative impacts, as well as the associated IAs, often go unidentified.
Compared to other types of animal production facilities, netpen aquaculture faces enhanced risk for IA transfer because nets provide no barrier against the flow of water, which can both convey agents long distances3 and provide a matrix within which they may persist4. In marine aquaculture regions, IAs can be transmitted horizontally between high density culture populations that are in close proximity to one another and through movement of equipment and personnel between facilities4,5,6. While it is well established that large numbers of cultured stock are lost to wild-domestic transmission7, the evidence of domestic-wild transmission impacting wild aquatic populations is much more sparse due to the difficulties in observing mortality and sub-lethal effects in wild populations8. However, studies have demonstrated that wild fish species (and in turn, their predators) are attracted to active aquaculture sites due to abundant and consistent nutrient subsidies in the form of feed and waste, as well as the presence of a physical structure9,10. Once attracted, wild species may convey IAs to the cultured fish, but are also likely to be exposed to abundant macro- and microparasites amplified by unnaturally high densities of cultured fish released from predation and experiencing reduced physiological demands for survival9.
In the presence of wild fish species of conservation concern and an absence of monitoring and research capable of adequately studying domestic-farm transmission from aquaculture, experts must synthesize available information to conduct risk assessments to inform regulation and management11,12. Considerations for whether IAs in aquaculture may pose a risk to wild fish include: whether or not cultured populations amplify IAs to concentrations beyond the baseline in the surrounding environment (referred to as “release”11), whether or not wild fish presence in proximity to aquaculture overlaps in space and time with release (“exposure”), and whether or not the IAs are virulent (“consequence”; Fig. 1). While consequence may be informed by previous literature, release and exposure require at least a modicum of real world data and the reliability of a risk assessment is improved as the quantity and relevance of data increases. In data-poor situations (i.e. those with elevated uncertainty) it is common to employ the “precautionary principle”, which holds that potential risk must be assumed, and protective action taken to prevent potential harm, until data satisfactory for evaluating risk are available13.

Risk to wild fish caused by infectious agent transmission from netpen aquaculture requires the intersection of release, exposure, and consequence. These three factors of risk analyses are represented in the three circles. Presence of two but not three of these factors does not indicate risk, and the outcome of two factors in the absence of the third is described in their overlaps in the figure.
The increasing adoption of modern molecular technologies in fisheries science is improving our understanding of wild-domestic and domestic-wild transmission by providing novel, informative datasets. Specifically, environmental DNA (eDNA), the extraction of nucleic acids from an environmental sample such as air, water, or soil, can offer insight into release and exposure. The presence of nucleic acid fragments specific to an organism in a water sample, while not necessarily indicating that organism’s viability, does signify the recent presence of that organism in the nearby surroundings, because eDNA degrades due to ultraviolet radiation and the metabolic action of microogranisms14,15. In the ocean, eDNA can be quickly advected or dispersed away from the farm source due to tidal and other currents; however, the balance of advective, diffusive, and decay processes can result in equilibrium distributions of eDNA around point sources, such as netpen farms16. Thus, a water sample collected in a matter of minutes can provide information regarding the presence of wild fish species and IA taxa. This technology has been used in recent years to monitor or study aquaculture facilities in order to anticipate wild-domestic transmission17, characterize domestic release18, and associate IA detections with subsequent mortalities in cultured fish19.
In coastal British Columbia, Atlantic salmon aquaculture facilities occupy waters where multiple species of Pacific salmon (a mixture of wild and hatchery-produced fish) reside and migrate as juveniles and adults. Many of these Pacific salmon populations have experienced declines in abundance over the last three decades20,21, with some populations showing some of the poorest returns on record in the most recent years20. While the factors leading to these declines are manifold (with climate change likely playing a major role), there has been vigorous debate regarding the impact of netpen aquaculture on Pacific salmon populations. In British Columbia, as elsewhere, wild-domestic transmission is better understood than domestic-wild transmission, although several recent studies have provided evidence suggesting that several pathogens may be transmitted from netpen aquaculture to free-roaming Pacific salmon18,22,23.
In this study we used a novel dataset featuring eDNA detections of Pacific salmon and IAs to inform the potential release of dozens of IA taxa from netpen aquaculture sites and exposure of Pacific salmon to these releases. Environmental DNA samples were collected at active and inactive netpen sites (fallow or decommissioned) to compare the presence of both Pacific salmon (exposure) and IAs (release) between active and inactive sites. We drew information regarding the impacts of these IAs from the literature to score the consequences of infection for each of the various taxa. While a formal and complete risk assessment is beyond the scope of this study, we use some components of the risk assessment process (release, exposure, consequence) to identify agents of concern in farm-wild interactions that warrant further attention and investigation.
Results
To determine whether contamination may have occurred at the time of eDNA collection, field blanks were collected prior to sampling at each site. Ninety-eight of 149 field blanks amplified at least one assay with a total of 208 sample/assay combinations that were positive, indicating some degree of contamination of the sampling equipment in the field. Contamination was likely due to splashing or aerosolization of ocean water during challenging (stormy, rough water) sampling conditions on an open boat deck. The amplified assays were from the most commonly detected taxa, with Salmo salar in 93% of positive field blanks, and Paranucleospora theridion (26%), and Candidatus Sygnamydia salmonis (23%) being the next most common. A total of 429 positive detections of pathogens or fish were nullified (coded as “NA”) due to positive detections in the field blanks from corresponding sample locations and sampling times (detections where Ct exceeded corresponding Ct in field blank). This included 227 positive detections of S. salar eDNA, 79 positive detections of P. theridion, and lower numbers (< 20) for 17 other assays.
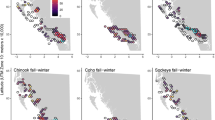
For some IAs, visualizing the raw data revealed stark differences between farms and inactive sites and provided evidence of both aquaculture-mediated IA release and exposure of a Pacific salmon species prior to the application of statistical analyses. A plot of Tenacibaulum maritimum (Fig. 2), serving as an exemplar, demonstrates that T. maritimum was almost never detected at inactive sites (including decommissioned and fallow farms), but was commonly detected in eDNA samples and S. salar tissues collected at active farms, often in the presence of Oncorhynchus tshawytscha eDNA. A similar pattern was observed for Atlantic salmon calicivirus (ASCV), Caligus clemensi, Cutthroat trout virus 2 (CTV-2), Lepeophtheirus salmonis, Moritella viscosa, Piscine orthoreovirus 1a (PRV), Tenacibaculum dicentrarchi, and Tenacibaculum finnmarkense. Plots with the same format as Fig. 2 but for all IAs, including those not modeled, are available in the supplementary material (Figures S1–S28).

Raw data from the Broughton Archipelago reveal that T. maritimum DNA (orange points) was more likely to be detected in water samples from active farms compared to inactive or fallow sites, T. maritimum was frequently detected in S. salar tissues (green points), and O. tshawytscha eDNA (blue points) was more likely to be detected in water samples from active farms and was commonly detected in the presence of T. maritimum. Points exceeding 40 (max Ct) indicate samples were collected but the target nucleic acid was not detected. Black vertical dashed lines indicate S. salar stocking dates and red vertical dashed lines indicate harvest dates (thus farms are fallow after red lines). Note that Cypress Harbour was a broodstock facility so that occasional, but never complete, harvest occurred and Doctor Islets was the only farm to be harvested and then restocked during the study period (fallow from November 2021 to March 2022).
Infectious agent prevalance in Salmo salar and eDNA samples
Infectious agent prevalences in S. salar ranged from zero prevalence (Neoparamoeba perurans, Erythrocytic necrosis virus (ENV), Salmon Pescarenavirus 1 (SPAV-1),Vibrio anguillarm, and Aliivibrio salmonicida) to high average prevalence (78–86%) across all collections (CTV-2, P. theridion, and PRV; Table 1). Note that T. finnmarkense was not assayed in S. salar tissues due to the timing of assay development.
In water samples, several IAs including SPAV-1, SPAV-2, and V. anguillarum, were never detected. Others including ASCV, CTV-2, Lepeophtheirus salmonis, Moritella viscosa, PRV, and T. maritimum, were detected at very low prevalance (or not at all) at inactive sites but several times higher at active farms. In contrast, IAs including Ca. S. salmonis, Flavobacterium psychrophilum, Ichthyophonus hoferi, Ichthyophthirius multifiliis, Kudoa thyrsites, Loma salmonae, and Piscirickettsia salmonis showed little difference in average prevalence between active farms and inactive sites (Table 1). Comparing prevalence between eDNA samples processed with the two different lysis buffers demonstrated the considerable impact of buffer type on detectability for most assays (Table 1), although this was confounded by season.
Association of Pacific salmon and other marine fish eDNA with active aquaculture
Environmental DNA from all salmon species assayed was detected over the course of the study (Table 1). Salmo salar eDNA was much more likely to be present in samples collected at active fish farms than those collected at inactive sites (Odds ratio = 234.0 (95% confidence interval = 59.5–921.3); Fig. 3A). Salmo salar eDNA was common across all samples (detected at an average prevalence of 95.3% across active farm visits and 43.4% across inactive site visits, Table 1). Of the Pacific salmon species, O. tshawytscha was the only species more likely to be detected at active farms than inactive sites (OR = 4.1 (2.1–7.9); Fig. 3A).
Of the non-salmonid marine fish taxa, Hypomesus pretiosus (surf smelt) was not detected in any water samples during the study. Engraulis spp. (anchovy) had the largest estimate in presence/absence models, indicating this genus was much more likely to be detected at active farms than inactive sites (OR = 273.1 (60.3–1237.1); Fig. 3B). Clupea spp. (herring) was the only other marine fish taxa for which eDNA was more likely to be detected at active farms relative to inactive sites (OR = 4.2 (1.9–9.2)).

Plots A–C depict odds ratios for sample location (inactive site versus active farm) in generalized linear mixed models of eDNA presence. Points indicate the odds ratios and horizontal lines represent 95% confidence intervals around the odds ratios. An odds ratio with a 95% confidence interval exceeding 1 (red vertical line) indicates that the eDNA of that taxa is more likely to be detected at an active farm than an inactive site.
Association of infectious agent eDNA with active aquaculture
The prevalences of Aeromonas salmonicida, Ca. S. salmonis, Lepeoptheirus salmonis, A. salmonicida, N. perurans, Parvicapsula pseudobranchicola, Viral Hemorrhagic Septicemia Virus (VHSV), and Yersinia ruckerii, were too low for modeling and that of Ca. S. salmonis, which was detected in almost every sample, was too high (i.e., models would not converge). The eDNA from 11 IAs (including four bacteria, three viruses, three microparasite taxa, and one macroparasite) was more likely to be detected at active farms than inactive sites (Fig. 3C). The meta-analytic mean of the models we ran for 22 IAs was significantly positive (Figure S29), indicating that overall, the odds of detecting pathogens at an active farm were 4.3 (95% CI = 2.3–8.1) times higher than the odds of detecting pathogens at an inactive site. Linear regression comparing model estimates for IAs in this study and those generated using the data from Shea et al.18 indicated that estimates for IAs that were modeled using data from both studies were more similar (\(\beta\) = 0.65, p = 0.003, \(R^2\) = 0.65) than would be expected due to chance alone (Figure S30). Model estimates were on average 1.5 times higher in this study compared to Shea et al.18. The meta-analytic mean for the models of the ten pathogens from the study by Shea et al.18 that overlapped with pathogens from our study was also positive with an odds ratio of 3.3 (95% C.I. = 1.5–:7.0, Figure S29). For reference, the corresponding odds ratio from the original analysis conducted by Shea et al.18, which used a different modeling approach and included a total of 19 pathogens, was 2.7 (1.5–5.0)18.
Infectious agent consequence scores from literature
Assigned “consequence of infection” scores and associated uncertainty based on evidence from the literature are provided in Fig. 4 (see Table S1 for score explanations and references for each IA). Thirteen of 22 agents for which we reviewed the literature lacked published studies of at least one of the categories (challenge studies, histological examinations, field epidemiology) we used to rank the evidence of consequence from infection. Putative narnavirus, in the most extreme example, has had no studies of the aforementioned categories conducted since its recent discovery24. Overall consequence scores ranged from zero (no evidence of impact on Pacific salmon) to six (evidence in each category of impact on at least one Pacific salmon species). Infectious agents with lower consequence scores tended to have higher uncertainty scores (Fig. 4).

Consequence scores (left panel), and accompanying uncertainty scores (right panel), were created for each modeled infectious agent based upon published literature. The consequence score was a composite of the weight of evidence from three categories: challenge studies, histological and/or clinical signs, and field or epidemiological studies (scoring rationale and references provided in Table S1). For each category where no information was available, or only pertained to S. salar, a point was added to the uncertainty score.
Integration of aquaculture association estimates and consequence scores
Tenacibaculum maritimum was the agent with the highest consequence score and largest odds ratio from the GLMMs with which we tested for an association with active aquaculture. Further, the model estimate for T. maritimum generated from reanalysis of data from Shea et al.18 was similar to the estimate from our study (Figure 5). Other agents with consequence scores of three or greater and odds ratios not overlapping one included F. psychrophilum, Ichthyobodo spp., T. finnmarkense, and PRV (Fig. 5).
The remaining six pathogens that were positively associated with active farms (as determined by GLMMs) all had consequence scores below 3. However, the literature available for composing the consequence scores was incomplete for these six pathogens (Fig. 4), in contrast to those with scores above three (Fig. 5).

Infectious agents are positioned to demonstrate the interplay between their likelihood of release associated with aquaculture (x-axis (log-scale)—odds ratios from models in this study and a reanalysis of data from Shea et al.18) and the consequence of infection (y-axis—consequence scores determined from the literature; see Figure 4, Table S1). The consequence axis is not linear, IAs are ordered by consequence score (for visibility) and horizontal dotted lines indicate unit divisions between those scores. For the consequence axis, overall scores are a composite of negative impacts demonstrated from challenge studies, clinical signs and/or histopathology, and field or epidemiological studies (Table S1). The font size of the infectious agent names indicates when literature is lacking for Pacific salmon (i.e., uncertainty)—the smallest text indicates no publications featuring Pacific salmon exist for any of the aforementioned topics, and the largest text indicates at least a single study featuring Pacific salmon exists for all components of the consequence score. Error bars indicate 95% confidence intervals around the estimate. The vertical dashed lines and surrounding ribbons represent the means and 95% confidence intervals from a meta-analysis (Figure S29) of the multiple models from each study, corresponding by color.
Nucleic acid presence in feed samples
Duplicate samples of pelleted feed from two different companies contained nucleic acids from salmonids, other marine fish, and IAs (Figure S31). Of the salmonids, S. salar (20–25 Ct) and O. nerka (23–29 Ct) nucleic acids were present in both duplicates for both feed samples. However, subsequent sequencing of the feed samples (unpublished data) indicated that salmonid nucleic acids were found at such low quantities that they were likely the result of environmental contamination occurring prior to our collection of the feed samples. In contrast, Clupea spp. and Engraulis spp. were abundant in both PCR results (Figure S31) and the subsequent sequencing analysis (unpublished data). Clupea spp. nucleic acids were highly abundant in Feed 2 (12–13 Ct) where Engraulis spp. was approximately 10 Ct higher, but the opposite was true in Feed 1 (Engraulis spp. = 13–14 Ct) (Figure S31). In Feed 2, the sample with a low Ct (high abundance) for Clupea spp., both duplicates were positive for Erythrocytic Necrosis virus, Ichthyophonus spp., K. thyrsites, and Loma spp. (all 20–30 Ct, Figure S31).
Comparison of industry sea lice counts and eDNA detections
Agreement between industry counts and eDNA samples (in terms of presence/absence) was low for L. salmonis (5–19%) and moderate for C. caligus (60–62%, Table S2). There were no positive eDNA detections for L. salmonis in the absence of industry count positives, but 16 for C. clemensii (Table S2). Lepeophtheirus salmonis was never detected in eDNA samples when the ReBead lysis buffer was used. The likelihood of C. clemensii detection in eDNA was positively correlated with industry counts of this copepod (Odds ratio = 2.8, 95% CI = 1.1–7.1, Figure S32B). There was no significant association between industry L. salmonis counts and either likelihood of detection or load in eDNA samples (Figure S32C, E), nor was Caligus clemensi industry count associated with eDNA sample load (Figure S32D). For C. clemensi, lysis buffer had a significant effect on likelihood of detection (OR = 21.7, 8.7–54.1, Figure S32B) and eDNA load (OR = 2.6, 1.8–3.8, Figure S32D). The binomial GLMM for C. clemensi predicted that when industry counts were zero and the Purelink lysis buffer was used, probability of detection would be 28–82% (Figure S32B).
Discussion
Marine aquaculture facilities are physically complex structures featuring high trophic subsidies and densities of fish, relative to the surrounding environment. The abundance of environmental nutrient inputs in the form of pelleted food and fish feces, as well as the physical structure itself, act as attractants for a large array of marine invertebrates and fish, some of which will in turn attract larger fish species9. The high density of cultured fish, which are protected from predation and starvation, facilitates the amplification and release of infectious agents. Together, these aspects set the stage for a scenario wherein wild fish are attracted to netpens and then exposed to potential infection (in addition to acting as potential sources of infection for the cultured population). By measuring eDNA of Pacific salmon, marine fish, and infectious agents, we found that such an attraction/exposure scenario likely occurred for Pacific salmon in the Broughton Archipelago, particularly for O. tschawytscha exposed to T. maritimum, a marine bacterium recently demonstrated to cause high mortality in O. tschawytscha25. Although virulence has been shown to vary considerably among T. maritimum strains25,26 and the impact of BC strains of this bacterium on wild Pacific salmon is still being determined, we suggest that the evidence from this study substantiates a potential risk to wild Pacific salmon. Our results regarding IA release were statistically consistent with those of a similar study conducted independently of ours18, indicating that our findings were robust and the patterns we identified appear to be consistent across space and time.
We interpreted elevated levels of O. tshawytscha eDNA at active salmon aquaculture sites as evidence that Chinook aggregate around farms. We hypothesized that this is the result of the attraction of this species to active aquaculture sites, perhaps due to trophic subsidies or physical structure27. While we have not seen previous publications describing the attraction of wild Pacific salmon to netpen aquaculture sites in BC, DFO maintains multiple databases of wild fish mortalities occurring during aquaculture operations (self-reported by aquaculture companies, https://open.canada.ca/data/en/dataset/0bf04c4e-d2b0-4188-9053-08dc4a7a2b03) and O. tshawytscha was documented in this database on seven occasions at Broughton Archipelago farms (including several we did not monitor) during our study period. Oncorhynchus kisutch (coho salmon) was observed on five occasions and O. gorbuscha (pink salmon) on eight, both at the farms we did not monitor. The hypothesis that some species of Pacific salmon are attracted to netpen aquaculture in BC is not unprecedented as this phenomenon has been documented with other fish species in aquaculture worldwide9,27,28. Several studies have described aquaculture feed appearing in stomach analyses of wild fish aggregating around netpens27. In the Mediterranean Sea, a study on wild fish attracted to netpen sites found that feed pellets were found in 66-89% of sampled stomachs for the five most abundant species captured around netpens29. However, one key caveat here, relevant to how we interpret the apparent aggregation of wild Chinook salmon around farms, particularly in the context of putative exposure to IAs, is that we cannot distinguish between adult and juvenile salmonids in our eDNA detections. This is important since susceptibility to some IAs may vary with age or previous exposure. Since eDNA detections are likely to indicate that fish are (or were recently) nearby14, we have frequent detections of Chinook in the winter period (Fig. 2) when adults would be uncommon in this region, and we have found no accounts of adult Oncorhynchus spp. in close proximity to netpens but several for juvenile fish (DFO incidental catch database and Johannes et al.30), we suspect that Oncorhynchus eDNA detections are more likely to represent juvenile fish. There are few local Chinook populations in the Broughton region, which means that detections of Chinook around farms are likely to come from coastal migrants from the Salish Sea and the central coast.
In addition to O. tshawytscha, we found elevated likelihood of detection at active farms for other non-salmonid marine fish taxa. However, the presence of large quantities of nucleic acids from Engraulis spp. (anchovy) and Clupea spp. (herring) in feed samples we tested complicated our interpretation of these results. Combining our water sample results, feed results, and DFO’s aquaculture incidental catch database provided the most complete interpretation. The abundant Engraulis nucleic acids of Engraulis spp. detected in pelleted feed, paired with high Engraulis eDNA prevalence at active farms (59.0%) but low prevalence at inactive sites (1.1%), and rare occurrence in the DFO incidental catch database (only observed on one occasion in the Broughton region during our study) suggests that the Engraulis spp. detections in water samples were most likely from feed. In contrast, while Clupea spp. occurred at high abundance in the feed samples, prevalence was high in water samples from both active (74.8%) and inactive (48.0%) farms, and Clupea pallasii was the most frequently observed species in incidental catch records in the Broughton region (140 occasions, with up to 490 000 individuals). Thus, we suspect that our results indicating elevated probability of Clupea spp. detections at active farms were truly driven by the presence of Clupea pallasii, although this interpretation remains uncertain due to the presence of Clupea spp. nucleic acids in the feed. Regardless, it is not disputed that C. pallasii are often present at netpen aquaculture sites in BC. The presence of C. pallasii at netpens could result in the secondary attraction of their predators9, including Pacific salmon (especially Chinook salmon). In addition to potentially facilitating wild-domestic infectious agent transmission when aggregating near netpens, C. pallasii may also be exposed to domestic-wild transmission and subsequently act as vectors to other wild fish or other aquaculture facilities, especially as herring entrained in netpens are released live, if possible, when farmed salmon are harvested.
Both S. salar and O. nerka (sockeye) nucleic acids were detected in both feed samples we tested but were not positive in a subsequent sequencing analysis of the feed samples (unpublished data). The feed samples were collected from DFO laboratories with these salmon species on site, and samples were taken from feed bags some time after they were opened, so we assume that positive PCR detections of salmonids were due to environmental contamination. In processing landings from some Pacific salmon fisheries, salmon by-products are converted to fish meal and fish oil which may be used in feeds31, but we are not aware of the use of these products in aquaculture feeds used in BC. In contrast, Clupea and Engraulis spp. were detected at very low Cts (i.e. high DNA copy numbers), also detected at abundance in the sequencing analysis (unpublished data), and these species were not cultured in the laboratories where the feed samples were obtained; therefore, we are more confident that herring and anchovy constitute real components of the feeds we tested.
Our analysis identified three bacteria species from the family Flavobacteriaceae that presented a high likelihood of eDNA release from active salmon aquaculture sites and high consequences of infection for Pacific salmon: F. psychrophilum, T. maritimum, and T. finnmarkense. Flavobacterium psychrophilum is well know from freshwater aquaculture, including Pacific salmon hatcheries32. While there is evidence that F. psychrophilum can persist in brackish water (6 ppt)33, we have seen no reports of transmission of F. psychrophilum or associated clinical disease in marine aquaculture or marine resident wild salmonids in BC. However, marine molecular detections of this bacterium have been observed in Pacific salmon34,35 and a geostatistical analysis indicated that early marine O. tshawytscha sampled closer to active netpen aquaculture were more likely to be positive for F. psychrophilum36. Nevertheless, molecular detection does not imply disease, and to confirm risk to Pacific salmon from F. psychrophilum in the marine environment, evidence of transmission in this environment is required.
In contrast to F. psychrophilum, the pathogenicity of both T. maritimum and T. finnmarkense has been well established for S. salar in the marine environment37. While clinical disease in Pacific salmon from T. maritimum has not been documented in BC per se, this cosmopolitan bacterium has been linked to mortality and clinical disease in Pacific salmon species elsewhere (O. mykiss in Chile38, in challenge studies using O. mykiss in Australia39 and O. tshawytscha in New Zealand25, in netpen O. tshawytscha in California40 (lesions), and in Alaska41). Virulence of T. maritimum varies among strains25,26, but strains that cause high mortality in S. salar are certainly present in BC aquaculture, where (excluding sea lice) T. maritimum is the infectious agent most often associated with pathogen-mediated mortality42. Therefore, while our qPCR assay did not discriminate among strains of T. maritimum, there is a substantial probability that virulent strains were represented in our results. In a correlative study, Bass et al.43 found negative associations between T. maritimum prevalence and load and population-level survival and body condition for O. tshawytscha and O. kisutch. Like T. maritimum, T. finnmarkense has been little studied in Oncorhynchus spp. in BC, but clinical disease has been documented in O. mykiss and O. kisutch in Chile44. Tenacibaculum dicentrarchi did not appear elevated at active farms in our study, but low prevalences across all samples led to high uncertainty in model estimates for this bacterium (Fig. 3C), and it was present in S. salar tissues at farms, although at half the average prevalence of T. maritimum. This is in contrast with the geographically overlapping work of Nowlan et al.45, where T. dicentrarchi typically occurred at higher levels than T. maritimum at two Broughton Archipelago farms and was considered the primary agent in Tenacibaculosis outbreaks. Despite several recent studies that have greatly expanded our understanding of the distribution and pathogenicity of Tenacibaculum spp. in BC and for S. salar37,45,46, there is a lack of studies addressing potential impacts to Oncorhynchus spp. Our study, in which T. maritimum provided the most striking intersection of release, exposure, and consequence across all pathogens tested, interpreted in concert with other recent works23,25,43,45,47, indicates that T. maritimum release from farms poses a potential risk for wild Chinook salmon in regions with active aquaculture.
Several viruses, including PRV, CTV-2, and ASCV, had positive associations with active farms, indicating elevated release. Piscine orthoreovirus 1a had the highest consequence score of these three viruses, principally because it has been studied more than the others. Recent research has found that likelihood of infection with PRV increases as the distance from active fish farms decreases22,36 and this has also been noted from non-statistical observations48,49. Other studies conducted in BC have indicated that PRV infection prevalence was negatively correlated with population-level survival for O. tshawytscha43 and PRV infection was associated with underweight O. tshawytscha and O. kisutch43,49. Wang et al.50 found that PRV-infected early marine juvenile O. tshawytscha had gene expression patterns consistent with a viral disease response and histopathological evidence of jaundice/anemia, a disease that has been associated with PRV infection on farms51, with similar disease manifestations in Pacific salmon and trout caused by other strains of PRV detected outside BC52,53. In contrast, laboratory challenge studies have led some researchers to conclude that PRV is not a risk to Pacific salmon54,55. The incongruous findings from laboratory challenge studies and observational field studies may indicate that other factors (e.g., temperature, predation) mediate PRV-related impacts. Taken together, the mounting evidence herein and across several published studies points to PRV posing the most impactful viral transmission risk posed to wild salmon from farms in BC.
Neither CTV-2 nor ASCV have been studied extensively in Oncorhynchus spp., and thus high consequence scores from the literature were not possible. Although ASCV has been shown capable of establishing systemic infection in S. salar56, there is no evidence that it causes clinical disease in that species and we found no studies focusing on the impacts of ASCV on Oncorhynchus spp. Challenge studies conducted on O. gorbuscha, O. tshawytscha, and O. nerka found that CTV-2 could be naturally transmitted from S. salar to O. tshawytscha57. Histological changes were found in the heart and kidney but the authors were unable to conclusively link this to infection with CTV-2 because control fish were not histopathologically assessed57. Nevertheless, the study by Long et al.57 demonstrated that in cell culture, CTV-2 had cytopathic effects on O. tshawytscha cells, viral loads persisted at higher levels in O. tshawytscha relative to other Oncorhynchus spp., and that endocarditis and intratubular protein casts in the kidney were more likely to occur in PCR-positive than PCR-negative O. tschawytscha. In-situ hybridization of CTV-2 in Atlantic salmon revealed particularly high level detections in the brain24, a tissue that was not examined in the Oncorhynchus challenge study57. Moreover, CTV-2 has been detected in both farmed and wild Chinook salmon, reaching prevalence on farms upwards of 12%24. Thus, further investigation of this virus in Pacific salmon is warranted and, given its strong release score, it should be considered a potential risk until more is known.
Two microparasite taxa, Ichthyobodo spp. and Paranucleospora theridion had positive associations with active farms. Ichthyobodo necator or I. salmonis are the species of this genus most likely to impact salmonids, but our genus-specific assay for Ichthyobodo cannot discriminate between these two species or others in the genus. Using the same assay as ours, Deeg et al.58 detected Ichthyobodo at prevalences of 14–30% in O. keta, O. gorbuscha and O. kisutch collected in the Gulf of Alaska, indicating that this genus of protozoan parasites may infect Oncorhynchus spp. in our region. Kent et al.59 observed heavy Ichthyobodo infections associated with gill damage in netpen O. tshawytscha in Sechelt, BC. Compared to BC, Ichthyobodo has been more thoroughly studied in Japan, where both I. necator and I. salmonis have been experimentally shown to cause high mortality in O. keta60,61. Given the high release score for Ichthyobodo in our analysis, the noteworthy lack of study on how this prevalent parasite genus impacts BC Oncorhynchus spp. populations, and its pathogenicity in other regions (which resulted in a relatively high consequence score in our analysis), we urge more research in the near future. Paranucleospora theridion is less understood than Ichthyobodo and its impact on physiology and the factors which regulate infections are understudied globally62. There is evidence that this marine microsporidian parasite contributes to gill disease62 and that infections are enhanced by elevated water temperatures36,63, suggesting it could become more impactful as ocean temperatures rise.
Based on the extensive body of literature associating sea lice presence and abundance with active netpen aquaculture worldwide and in our study area64,65, we were surprised to find very low prevalence of Lepeophtheirus salmonis (salmon louse) eDNA (too low to model). In contrast, the other sea louse assayed in our study, Caligus clemensi, showed a moderate prevalence and was more likely to be detected at active farms than inactive sites. The lower eDNA detection prevalence of L. salmonis relative to C. clemensi was unexpected given that the DFO industry sea lice count database indicated that L. salmonis was more commonly observed than C. clemensi. One factor that may help to explain this difference is the fact that C. clemensi are considered to be generalists but commonly attach to herring (indeed, one of their common names is “herring lice”), which we showed to be associated with active salmon farms. A three times higher rate of agreement for C. clemensi relative to L. salmonis, in terms of presence/absence between industry counts and eDNA detections, suggests that some aspect of our eDNA collection or laboratory analysis is better suited to the former species. Alternatively, the physiology or life cycle of C. clemensi somehow results in a greater rate of nucleic acid shedding or persistence in the environment. There is some evidence that crustaceans shed nucleic acids at low rates due to their hardened exoskeleton66, but both ectoparasite species in our study are copepod crustaceans. However, this could explain why eDNA concentrations of neither sea louse species was correlated with industry counts. Other researchers have reported unexpected dissimilarities between sea louse detections in eDNA and manual sea louse counts17,67 and this has been the case for other IAs as well15. Whatever the case, the low detection rate of L. salmonis in the presence of positive manual counts suggests that our study does not properly characterize the release of L. salmonis from active netpens and therefore we make no inference regarding the risk associated with this species. For C. clemensi, the presence (but not eDNA concentration) of which was positively correlated with industry counts, our results indicate that Oncorhynchus spp. are more likely to encounter this ectoparasite around active farms than inactive sites. We note also that, in addition to transmission via infective copepodid larvae, C. clemensi are more likely to transfer between hosts as adults and may be more common in the water column around infested farms. Few studies have investigated the potential impacts of C. clemensi on Oncorhynchus spp. but there is evidence of reduced growth68 and foraging ability69 for O. nerka infected with C. clemensi.
Records from one of the aquaculture companies in the study indicated that S. salar were vaccinated against Aliivibrio salmonicida, Vibrio anguillarum, Aeromonas salmonicida, Moritella viscosa, and Renibacterium salmoninarum. Indeed all of these infectious agents were detected not at all or at very low prevalence in S. salar tissues, and only M. viscosa and R. salmoninarum were detected at sufficient prevalence in eDNA to be statistically analysed. Moritella viscosa eDNA was more likely to be detected at active farms than inactive sites, which was surprising given its low prevalence in S. salar tissues. One potential explanation of this phenomenon is that other fish species attracted to netpens could be bringing this bacterium into the vicinity. Similarly, shedding from C. pallasii or other fish species attracted to netpens might also explain the elevated presence of erythrocytic necrosis virus (ENV) in eDNA collected at active farms. Although it was completely absent from S. salar tissues, ENV was more likely to be detected at active farms than at inactive sites (confidence intervals marginally overlapped zero, Fig. 3C) and C. pallasii is a well-known carrier of this virus70. Although our literature search indicated a low consequence score for ENV, the elevated detections despite absence of the virus in S. salar tissues illustrates a potential scenario where the attraction of one species (C. pallasii) results in pathogen exposure for another (e.g., Onchorhyncus spp.), a phenomenon described previously18,71. ENV was detected in both of the feed samples we tested (stronger detection in the sample with higher levels of C. pallasii) and therefore feeds administered at farms could be another source of elevated detection frequency.
Risk assessments regarding wild sockeye salmon and specific aquaculture interactions were conducted in British Columbia in 2019. These assessments were led by DFO to address the Cohen Commission recommendation that salmon aquaculture facilities be removed from the Discovery Islands unless they were demonstrated to pose minimal risk to Fraser River O. nerka72. The nine assessments covered risk from IHNV, A. salmonicida, P. salmonis, R. salmoninarum, Y. ruckerii, PRV, T. maritimum, M. viscosa, and VHSV (https://www.dfo-mpo.gc.ca/cohen/iles-discovery-islands-eng.html) potentially transferred from Discovery Island salmon farms. While our study included assays for all of these pathogen taxa aside from IHNV, we detected several at prevalences too low for statistical analysis (A. salmonicida, Y. ruckerii, VHSV), and found that two others did not show differences in detection prevalance between active farms and inactive sites (R. salmoninarum, P. salmonis - although reanalyzed data from Shea et al.18 indicated that P. salmonis was more likely to be detected at active farms). For the remaining three (PRV, T. maritimum, and M. viscosa) we found that release was elevated at active farms, findings consistent with the release assessment for these pathogens in the DFO risk assessments73,74,75. However, our interpretations concerning the likelihood of exposure and consequences of infection for these pathogens differ from the conclusions in the DFO assessments. This is partly due to the fact that, for the exposure element, the DFO risk assessments solely focused on Fraser River O. nerka, while we assayed all Oncorhynchus spp. occurring in our region and found that O. tshawytscha were more likely to be detected at active farms than inactive sites. For M. viscosa, the DFO risk assessment stated that this bacterium does not infect S. salar between the months of May and October, and therefore would not be amplified and released during the O. nerka migration period74. In contrast, we often detected M. viscosa eDNA at farm sites concurrently with O. tshawytscha eDNA, and the potential impacts of this bacterium on that or other Oncorhynchus spp. have not been studied. For PRV, consequences of infection were considered negligible in the DFO risk assessment73, but several studies published afterwards43,50, and others published prior51,76,77, indicate potential for consequences for O. tshawytscha and O. nerka that should be considered underexamined risks when operating under the precautionary principle. Infection with T. maritimum was considered unlikely to occur for Fraser River O. nerka by the DFO risk assessment75, but there have been no actual laboratory studies performed on sockeye salmon to determine their susceptibility to infection by this bacterium. Clinical disease from T. maritimum has, however, been observed in O. tshawytscha outside of BC25,40,41 and clinical disease and mortality in adult O. tshawytscha were recently observed in BC from a closely related species, T. dicentrarchi47.
Comparing our study and the DFO risk assessments for these latter three pathogens illustrates potential impacts on conclusions drawn from risk assessments due to the assessment’s scope (considering a single host species for exposure versus six species) and the availability of literature informing potential consequences of infection. Incomplete information from the scientific literature or regional field studies contributes to uncertainty in risk assessments, as illustrated by the case of T. maritimum. Challenge studies have not yet been conducted to determine whether BC strains of this bacterium can cause disease in BC wild salmon (although challenge trials are currently underway at DFO). In this context, the causal evidence of harm to Pacific salmon in BC remains, technically, outstanding. That said, the features of risk - and potential to mitigate that risk—will vary from context to context: region to region, species to species, and so on. Even when causality is known and risk—broadly defined—exists, environmental factors (e.g. temperature and salinity), pathogen factors (e.g. strain and dosage), and host factors (e.g. species, nutritional condition, predation context, immune state) all constitute “component causes”78, which may or may not create the conditions for an etiological agent to cause disease. Although we have not delved into all of the specifics - or component causes - required for a full-blown risk assessment, we have demonstrated necessary (if not sufficient) conditions for risk to wild Pacific salmon, particularly in the context of the precautionary principal. In a framework like the one we used here (a common framework based on the three key elements of release, exposure, and consequence), further information is much more likely to lead to more evidence of pathogenicity and risk of release, rather than less (since the default is minimal risk). Thus, our conclusions regarding which IAs may pose potential risks to Pacific salmon are likely conservative, particularly when we consider agents with strong evidence of release but insufficient scientific literature to accurately assess consequence, including M. viscosa, T. finnmarkense, P. theridion, CTV-2, C. clemensi, and ASCV.
Our definition of exposure in this study is based on elevated detections of wild fish eDNA within and around netpen facilities (within 50 m), and is thus more narrowly defined than what may truly be important ecologically. This is because our study was not designed to determine the spatial extent of IA release around active netpen facilities. Shea et al.16 found that S. salar eDNA could be detected up to 3.7 km from active farms, but the distance over which infectious agents remain viable is likely to vary across taxa15. Due to the biology of different IAs and the hydrodynamics around facilities, the highest infection pressure may not always occur in the immediate surroundings of an active farm79. Therefore, any individual Pacific salmon (not just O. tshawytscha) that migrates through areas in our study region where infection pressures are elevated could be exposed to agents released from netpens. Future studies, including eDNA collections, should feature an experimental design capable of determining the spatial extent of IA release from active farms, perhaps informed by biophysical models80.
A confirmation of exposure superior to co-occurrence of IA and host eDNA would be the observation of infections, above any background rate, in wild salmon collected in areas known to experience elevated release of IAs from netpen aquaculture (caged sentinels have been used to this end in the past81). Detection in eDNA cannot confirm infection of wild salmonid hosts15. The co-occurence of IA and host eDNA that we have interpreted here would also be consistent with scenarios in which wild hosts are exposed to IAs released from aquaculture but do not become infected. From the eDNA data we are unable to determine the precise location of Pacific salmon, the viability of a detected IA, or the infection pressure at a given location; therefore, we are unable to determine the likelihood of infection based on previously established challenge models (which are scarce for Pacific salmon for many of the IAs we assayed). However, because removal of free eDNA from the environment is fast and IA genetic material is relatively rare in the environment, the chances of detecting dead or extracellular nucleic acids is expected to be very low and thus detection in aquatic systems is highly likely to represent a viable life cycle stage14,15. Furthermore, the well-studied phenomenon of waterborne transmission of IAs between aquaculture sites that are kilometers apart (whether through free-ranging fish vectors or waterborne transmission)3,79 suggests that opportunities for transmission to susceptible wild fish around active farms should be high. Finally, previous studies have shown that fish collected at varying distances from netpen aquaculture were more likely to be infected with IAs and macroparasites as the distance from aquaculture decreased9,22,36,49,64,65,77,82. To resolve the data gap between co-occurrence of eDNA and potential infection, studies sampling wild salmon at varying distances from aquaculture, perhaps via nimble methods like “micro-trolling”83 or more experimentally controlled methods like caged sentinels81, are required and should be paired with eDNA collections. Challenge studies featuring Pacific salmon exposed to varying doses of IAs (and concurrent eDNA collection) would also aid in our interpretation of the risk represented by IA concentrations measured around netpen aquaculture via eDNA sampling. Until such laboratory challenge and field exposure studies are conducted, the combination of field data that characterize the risk of IA release from netpen aquaculture (as found in this study) coupled with consequence information from previous literature is the best means for evaluating the risk posed by netpen aquaculture to wild Pacific salmon. The precautionary principle suggests that the lack of laboratory challenge and field exposure studies (i.e. insufficient data for conclusively determining risk) does not warrant an assumption of minimal impact for wild fish.
The spatial (Broughton Archipelago region) and temporal (18 months) extents of our study may limit the applicability of its results. Variable abiotic and biotic factors among aquaculture regions in BC (and more broadly) are likely to influence the assemblages of salmonids and marine fish as well as the diversity of infectious agents35,36,42. Interannual variability can result in outbreak or absence of some IAs over time, as well as strong shifts in salmonid abundance (e.g., between the distinct odd versus even calendar-year O. gorbuscha populations). Nevertheless, we found striking similarities between our IA results and those of Shea et al.18, who sampled from a broader geographical region (Broughton Archipelago and Discovery Islands) and across three years prior to our study (2016-2018), indicating that many of our results are likely not exclusive to the spatiotemporal extent of our study. Furthermore, while we cannot use our results to generalize about what IAs might be problematic along the entire BC coast, we can use them to prioritize IAs for regulatory consideration, monitoring, and future research. Infectious agents that did not appear to be at a high risk of exposure for wild Pacific salmon in this study could be important elsewhere or under other (perhaps interannually varying) conditions15.
Potential sources of bias in our study include those that are specific to our field and laboratory methods, and those specific to the biology of the system that may favor detection of some IAs over others. Methodological biases may include the depth at which samples were collected, the volume of water filtered, filter size and type, and various aspects of the extraction procedure. If eDNA is stratified in the water column, our data might not fully characterize what fish species are attracted to farms because fish abundance and diversity have been shown to vary with depth around netpens27. The fact that we had to include the lysis buffer type in our models demonstrates the strong effect of this aspect of the extraction process and we recommend against such a switch mid-study, although it was necessary in our case due to limited availability of the Invitrogen lysis buffer at the time, in part due to the COVID-19 pandemic. Despite methodological differences between our study and Shea et al.18, including sampling at different depths, using different filters and filtering methods, and using different extraction techniques, we found very similar results, suggesting the biological patterns we observed were robust to variation in methods. However, Shea et al.18 almost never detected PRV, an RNA virus that we detected at moderate prevalence, and this was likely because those authors did not specifically extract RNA from filtrate material.
Agent-specific factors potentially biasing our characterisation of IA release include stability in seawater, variation in how particles disperse in the water column (e.g., parasite spores versus virus particles), and variability in life cycles that influence how IA eDNA is shed from hosts (free versus bound in faeces, mucous, urine, etc). By bias, here we mean among-IA differences in our ability to quantify release. For example, our analysis herein did not consider free viruses—those not bound in shed cellular material that could be captured on a filter. Given that eDNA shedding and decay rates vary with different animal forms66, it is likely that variability in eDNA persistence also exists within the IA taxa that constitute our assay panel (e.g., RNA viruses versus multicellular parasites). Pathogens that are transmitted between hosts by vectors and infect internal organs could be more difficult to detect than those transmitted by the fecal-oral route through the water column. Such sources of bias are likely to result in an underestimation of the presence of some infectious agents, relative to others. It is possible that if some taxa are underrepresented due to our methodology, we could be misinterpreting their potential for amplification and release associated with aquaculture.
The remaining aquaculture facilities in the Broughton Archipelago (those monitored in this study) were decommissioned in 2023, as decided by the First Nations included in the BATI agreement. However, the IAs identified as potential risks to wild salmon in this study, as well as the use of eDNA collections to inform risk assessments around domestic-wild pathogen transmission, may be applied to other regions in Canada where potential wild-aquaculture interactions exist. With results exceptionally consistent with those of a previous study18, despite methodological variations, our study presents robust evidence of IA release from marine netpen aquaculture. These data and others collected similarly should be considered as important elements to address data and knowledge gaps regarding domestic-wild transmission in future risk assessments.
Now that T. maritimum, T. finnmarkense, Ichthyobodo, and PRV have been identified as IAs that are released from active farms in BC and have known disease potential in Oncorhynchus spp., an important next step is to determine the environmental, biological, and operational factors that impact their shedding rates. Such a mechanistic understanding can be used to inform progressive regulations of industry to reduce impacts of these agents on wild salmon. As a hypothetical example, T. maritimum might be released from farms at elevated levels: on a seasonal pattern or when water temperatures are high, when fish are stressed by co-infections or in otherwise poor condition, or following farm operations that lead to acute stress such as mechanical delousing. This information would serve as the first biological input, the rate of IA particle release from a given location, into any prospective biophysical model80. Controlled laboratory studies could provide necessary information for other inputs, including the persistence of T. maritimum at various temperatures and salinities and the infection pressure required to cause infection80. With these parameters, biophysical models could predict the dispersal of T. maritimum around netpen aquaculture networks and collections of eDNA and wild fish could be used to ground truth model predictions80.
Regardless of the answers that future studies of pathogen release and dispersal will provide, no single approach is likely to provide comprehensive insight to the question of risk. For example, questions a biophysical dispersal model alone cannot answer are those that relate to the potentially infected hosts. Inter-species variability in susceptibility to infections, wild host migration details (in space and time), the complicating effects of cummulative stress, and indirect effects of sublethal infections will all factor into the true risk that any IA release creates to wild salmon. Judiciously combining all the relevant features, while also accounting for uncertainty, will be required to holistically gauge risk.
Methods
Methods overview
To approximate whether any IAs found in Atlantic salmon netpen aquaculture might pose a risk to wild Pacific salmon, we considered the likelihood of release and exposure as evidenced by eDNA detections of IAs and their hosts, coupled with the consequences of infection as determined by evidence from previous literature (Fig. 6). eDNA samples were collected over 18 months from seven active and 4 inactive (decommissioned or fallowed) netpen aquaculture sites and analyzed by qPCR for the presence of salmonids, marine fishes, and IAs (Table 1). To test for evidence of IA release, we modeled whether or not IAs were more likely to occur at active farms versus inactive sites. For exposure, we used the same model structure to determine which Pacific salmon species were more likely to be detected at active farms and thus more likely to be exposed (to agents that showed evidence of release). For consequences of infection we considered whether evidence existed in the form of negative impacts determined from challenge studies, histopathological and/or clinical evidence of disease, or evidence of population level impacts from field studies. Evidence from the literature was converted to qualitative scores. These scores and model estimates were then considered together to identify agents of concern (high probability of release and high consequence score, upper right of Fig. 6), particularly for Pacific salmon species with high exposure (Fig. 6).

Schematic outlining the study design. Environmental DNA collections were used to characterize infectious agent release at active farms and exposure of wild fish. Evidence from the literature was used to develop agent-specific consequence scores.
Study area
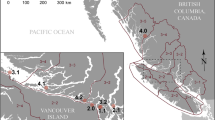
The Broughton Archipelago is a complex collection of islands between Vancouver Island and mainland British Columbia (Fig. 7). Salmon aquaculture was first introduced to this region in 1988. In 2018, three First Nations in this region, Kwikwasut’inuxw Haxwa’mis, Mamalilikulla, and ’Namgis, signed a Letter of Understanding with the Province of British Columbia regarding finfish aquaculture in the Broughton Archipelago. This led to an agreement between the Nations and tenure holders (MOWI Canada West and Cermaq Canada) around the transition of aquaculture practices in the region (Broughton Aquaculture Transition Initiative, BATI). The research described herein was conducted under the Indigenous Monitoring and Inspection Plan (IMIP), that was created alongside BATI.
Eleven farms sites were monitored during the course of this study (Fig. 7), with three levels of activity: “active” (fish on site), “fallow” (infrastructure present, no fish on site), and “decommissioned” (infrastructure removed). Sites were visited approximately once a month, weather permitting, from October 2021 to March 2023 (Table S3).

Environmental DNA samples were collected from Atlantic salmon netpen aquaculture sites in the Broughton Archipelago (A), located on Canada’s West Coast (B), on the north end of Vancouver Island in British Columbia (C). In panel A, red circles and text identify active farms and blue squares and text identify decommissioned farms. Panel B shows the Broughton Archipelago (red star) on a map of North America. Panel C shows all salmon aquaculture netpens on the British Columbia coast (at the time of the study) and around Vancouver Island (VI) as well as the Broughton Archipelago (red rectangle). This map was created by the authors in R version 4.3.084 (https://www.r-project.org/) using shoreline data from the Global Self-consistent, Hierarchical, High-resolution Geography Database Version 2.3.785 (https://www.soest.hawaii.edu/pwessel/gshhg/).
Environmental DNA collection
Environmental DNA (eDNA) was collected using the EZ-eDNA™ pump and filter system developed by RKS Laboratories Ltd. (Qualicum Beach, BC, Canada), described in86. This system consisted of a 12-volt pressure regulated diaphragm pump, coupled with a programmable flow controller set at 5 L. The sample water was pumped through two hollow membrane filter cartridges connected in parallel (field duplicates). Each filter cartridge has two luer locks to attach intake and discharge tubing. The hollow membrane filter consists of 120 Polyethersulfone (PES) membrane tubes with a nominal pore size of 0.1–0.45 \(\mu \hbox {m}\) (RKS Laboratories Ltd.). All eDNA samples used a total of 5 L (approximately 2.5 L passing through each filter cartridge). Once the filtration was concluded, an air pump included in the system was used to remove residual water from inside the filter housing. Next, 2 ml of RNA-Later™(ThermoFisher Scientific, Waltham, MA, USA) was injected by syringe into the filter cartridge for preservation, and luer lock end caps were attached for storage. The filter cartridge was then refrigerated for 24–48 h, and subsequently preserved in a -20 °C freezer, until ready to be extracted and analyzed.
At each visit to an active farm, eDNA samples were collected from within netpens and from transects surrounding the farm superstructure. Half of the total active pens on site were sampled at each farm visit (4–6 pens), including one pen (i.e. control pen) that was identified at the beginning of the production cycle and consistently sampled at each visit. Pens with recent mortality were prioritized for sampling. For samples collected from within netpens, the intake hose was submerged to 8 m depth and kept stationary using a weighted pole while 5 L of seawater was pumped (2.5 L per filter cartridge). At active or fallow farms (infrastructure present), transect samples were collected on each of the four sides of the farm superstructure, parallel to the side of the superstructure and at a distance of approximately 20–50 m from the structure’s edge, depending upon the presence of obstacles (boats and anchor lines) and conditions (weather and tides). Five L (2.5 L per cartridge) of seawater was pumped from a depth of approximately 8 m by attaching the intake hose to a downrigger weighted with a 11 kg lead ball. The rate at which each transect was completed by the boat was determined by the length of the side of the superstructure combined with the target of pumping 5 L (pumping took approximately 5–7 min). At decommissioned sites, transects were conducted by boat in four directions to roughly travel the shape of a square with 100 m sides (5 L of seawater collected per direction, to mirror farm samples).
If multiple sites were to be sampled for eDNA in a single day (fish were never sampled from multiple sites in a single day due to biosecurity concerns), a separate pump system was used at each site. When arriving at a sampling location, a field blank (i.e. negative control sample), consisting of 5 L of sterile distilled water was pumped and filtered through two parallel filter cartridges. When sampling at active farms, the transects outside the farm were conducted first, followed by the netpens, with the expectation that eDNA concentrations were likely to be greater inside netpens. When each netpen was sampled, the pump system was first flushed with 5 L of water from that netpen before a sample was taken. Such “flushes” were not conducted between transect samples. After each sampling day, 5 L of 10% bleach were run through pump systems, followed by 5 L of sodium thiosulfate, to neutralize the bleach. The exterior parts of the apparatus were also cleaned with these solutions.
As the composition of salmon feed can include a variety of marine fishes, and possibly nucleic acids from infectious agents present in fish used for feed, we recognized that salmon feed pellets could contribute to eDNA detections around farms, complicating the interpretation of the resulting qPCR data. Hence, we obtained samples of feed used in salmon hatcheries and for grow-out on farms and tested them across the same assays used on fish and eDNA in our study. While this was not an exhaustive effort, as feeds used by commercial farms likely include multiple brands, and can vary with life stage and husbandry concerns, it provided some insight to the interpretation of our results.
Atlantic salmon sampling
During visits to active farms, Atlantic salmon were collected from netpens for histological analysis and qPCR IA screening. Thirty live, apparently healthy fish as well as up to 10 moribund and/or dead fish were collected from each farm on the day of the visit. Fifteen of the thirty live fish were always collected from the control pen, while the other fifteen were usually collected from a secondary pen, decided by the veterinarian (EDC) on the day of the visit, based on the clinical conditions of the fish and the mortality data for the 30 days before the visit, provided by the facility staff. Collection of moribund/dead fish prioritised these same two pens; however, if very few fish were obtained from these two pens, additional pens were involved in the collection of moribund/dead fish. Moribund fish were either scooped up with a long dip net from inside the pen (possible due to a lack of startle reaction, typical of a lethargic state) or obtained by pumping up dead fish from the bottom of the pens. In one farm where no “mort pumps” were present, professional divers were hired to collect and provide morts from the bottom of the pens.
Small portions of the gills, liver and anterior kidney, to be used for molecular analysis, were sampled (in triplicate) from every fish (live, moribund or dead) and preserved in RNAlater (1.5 mL). Clinical observations were made during dissection. Tissues were stored at 4 °C, and transferred back to BATI’s laboratory facility in Nanaimo, BC. One set of gill, liver, and kidney samples was delivered to DFO’s Pacific Biological Station Molecular Genetics Laboratory within 1–2 days of sampling. Samples were processed for nucleic acid extraction upon arrival.
Ethics statement
Fish were collected and euthanized by aquaculture facility personnel (not authors or BATI technicians). Therefore, all work with animals was performed according to the Canadian Council on Animal Care’s (CCAC) Guide to the Care and Use of Experimental Animals, and protocols were approved by Fisheries and Oceans Canada (DFO) through its Pacific Region Animal Care Committee (conditions of license for salmon farm operations). Live-sampled fish were euthanised via overdose of tricaine methanesulfonate (Syndel laboratories Ltd., Nanaimo BC, Canada). All tissue samples involved were collected under the IMIP agreement between the aquaculture companies and BATI. Where applicable, methods herein are reported in accordance with the ARRIVE guidelines.
Molecular analysis
Environmental DNA extraction
DNA and RNA were extracted from hollow membrane filter cartridges using a modified version of the protocol (as described below) from the Invitrogen™ PureLink™ Viral RNA/DNA Mini Kit (Cat# 12280050, Thermo Fisher Scientific). We used two methods, which we label here as “manual extraction” and “semi-automated extraction.” Manual extraction was used for samples collected from September 2021 through April 2022 and semi-automated was used for samples collected thereafter, once the extraction machine became available for use.
For manual extraction, all components, aside from ethanol, used in extraction were included in the Invitrogen Mini Kit. First, RNAlater was removed from the filter cartridges using an electric air pump. Next, 2 ml of lysis buffer mixed with Proteinase K (2 mg/ml) and Carrier RNA (1 µg/sample) was pipetted into each filter cartridge. After thorough vortexing (2400 rpm, 5 min), the samples were incubated at 56 °C with slow shaking for 30 min to ensure complete lysis. The lysate was manually collected from each filter using 5 ml syringes and transferred to separate 5 ml tubes. The 5 ml tubes were then cetrifuged at maximum speed (2400 g) for 5 min to remove any remaining unlysed debris or particles. The supernatant was transferred to a new tube where ethanol was added to reach a final concentration of 37% ethanol. The resulting solution was then loaded onto a silica spin column, where a vacuum pulled the solution through. The column was then washed twice with Wash Buffer. Finally, RNA/DNA was eluted in 50 µl of sterile, RNase-free water.
For semi-automated extraction, RKS Laboratories Ltd. (Qualicum Beach, BC, Canada) developed a specialized system based on the manual extraction method. This semi-automated system is capable of processing 12 filters simultaneously. It removed RNAlater from the hollow membrane filters by pumping 5 ml of lysis buffer from Nanjing Rebeads Biotech Co., Ltd (Cat# RBX024-1000, Nanjing, China) into each filter cartridge. Subsequently, 2 ml of lysis buffer (Nanjing Rebeads) mixed with Proteinase K (2 mg/ml, Invitrogen) and Carrier RNA (1 µg/sample, Invitrogen) was introduced into each filter cartridge. The system shook and incubated the 12 filters at 40–50 °C for 30 minutes to ensure thorough lysis. After lysis, the system used air pumps to push the lysate out of the filters into separate 5 ml tubes for each filter. The remaining steps, from centrifuging in 5 ml tubes through elution, were conducted identically to those described above for manual extraction. Preliminary comparison of results from the extraction protocols was performed prior to the switch to semi-automated extraction, but we also discuss the effects on results below.
Feed pellets that were on hand at the experimental fish laboratory at DFO’s Pacific Biological Station and Pacific Science Enterprise Centre were extracted for analysis. The first sample, “feed 1,” was 6 mm BioBrood pellets (Bio-Oregon, Longview, WA, USA), which are intended for fish from 400 to 1000 g. Feed 2 was 1 mm pellets from EWOS (Surrey, BC, Canada). Both feeds were collected from bags that were previously opened, resulting in potential for environmental contamination. Duplicate samples were prepared from each feed type, each sample from feed 1 consisting of 2 pellets and each sample from feed 2 consisting of 6 pellets. The pellets were soaked in the Purelink lysis buffer at 40–50 °C for 30 min with slow shaking, homogenized at 30 Hz for 5 min, and then centrifuged at 3000 rpm for 10 min to allow for separation of liquid and solid layers. Ethanol was added to the supernatant to reach a final ethanol concentration of 37%. The remaining steps followed the same procedure as the eDNA manual extraction.
Salmo salar tissue extraction
Tissue preparation for qPCR followed Miller et al.87 for RNA extraction, but instead of also extracting DNA, DNA was not enzymatically removed from the RNA extraction. We found this procedure reduced dilution of the RNA without sacrificing DNA detections. Single tissues were homogenized on separate plates but equal aliquots for each of: gill, kidney, and liver tissue homogenates were combined for extraction.
High-throughput polymerase chain reaction
The BioMark™platform (Fluidigm Corporation, CA, USA; now Standard BioTools), a nanofluidic automated real-time quantitative PCR system, was used to test eDNA samples using 7 salmonid, 9 marine fish, and 32 IA assays (Tables 1, S4). The same platform was used to test S. salar mixed tissue samples using the same 32 IA assays run for eDNA. This section briefly describes how samples are prepared for use on the BioMark, but detailed methods are available in previous publications from the DFO Molecular Genetics Lab16,34,87.
Following extraction, RNA in the resultant nucleic acid (RNA and DNA) was reverse-transcribed to cDNA. For S. salar tissues, nucleic acids were first normalized to 62.5 ng \(\mu\)L−1 using a Biomek NXP™automated liquid-handling instrument, and 1 \(\mu\)g of normalized nucleic acid was converted to cDNA. The normalisation step for salmon samples ensures that IA results are reported relative to total nucleic acid, the vast majority of which comprises host DNA and RNA. For eDNA samples, because the eDNA itself comprises the nucleic acid in each sample, no such nucleic acid normalization was conducted prior to converting RNA to cDNA. Because reaction volumes used by the BioMark (7 nL) are so much smaller than those used in conventional qPCR (\(\sim\)25 \(\mu\)L), a pre-amplification step—as recommended by the manufacturer—was used to increase sensitivity with such small volume reaction wells. In this step, cDNA/DNA underwent 17 PCR cycles using a 1/10 dilution of all primers (no probes) targeting sequences to be assayed on the BioMark87. Miller et al.87 conducted extensive analyses regarding the impacts of this pre-amplification step, with no negative impacts on specificity of the final assays identified when post-amplification qPCR was carried out using TaqMan probes.
Following pre-amplification, samples and assays were pippetted into the respective loading wells of 96 x 96 well dynamic arrays (Standard BioTools). Serially diluted artificial probe constructs (APCs) and processing controls were included, as per Miller et al.87, and a second fluorescent dye was included in all reaction chambers to detect potential laboratory contamination by APCs. The purpose of APCs is to confirm assay function (positive control), calculate assay efficiency, and facilitate estimation of IA DNA/RNA copies. The dynamic arrays were run on the BioMark, individual runs were analyzed for cycle threshold using the Fluidigm Real-Time PCR Analysis software, and scored data were exported to a purpose-built SQL-database. Technicians were blind to the identity of the samples throughout the laboratory work and post-processing.
Consequences of infection literature search
To contextualize our IA release results, we created “consequence scores” for IAs based upon previously published, peer-reviewed literature (“consequence” in Figs. 1 and 6). For each IA that was detected during the study (Table 1) we searched the existing literature with the goal of developing scores representing the weight of evidence suggesting pathogenicity for a given agent. We considered whether negative impacts of infection have been observed for three categories including challenge studies, histological and/or clinical examinations, and field or epidemiological studies. Evidence for each consequence category was scored from 0 to 2, where 0 represents no evidence of a negative impact and 2 represents robust evidence of negative impacts in at least one species of Oncorhynchus. Scores of 2 required evidence of high mortality in challenge or epidemiological studies or severe lesions in histological studies. A score of 1 indicates some evidence of negative impact in Oncorhynchus or robust evidence of negative impacts in S. salar when evidence is lacking for Oncorhynchus spp. The scores for each category were summed so that consequence ranged from 0 to 6 with an associated uncertainty score ranging from 0 to 3 (Table S1). We prioritized studies featuring Oncorhynchus spp. but considered studies of S. salar where studies were lacking for Oncorhynchus spp. If no studies featuring Oncorhynchus spp. were found for a given agent for one of the categories, a point was added to the uncertainty score (uncertainty could range from 0 to 3, where 3 represents no studies conducted on Oncorhynchus spp. for any consequence categories). This approach gives each category equal opportunity to contribute to the overall consequence score, which is an imperfect simplification of the situation but we have provided the rationale behind all scores (Table S1). Furthermore, consequence scores do not reflect the complexity of contradictory findings amongst studies (e.g., an IA has negative impacts in one Oncorhynchus species but not another).
Comparison of sea louse counts with qPCR results
As a condition of their license to operate, aquaculture facilties in BC must routinely count parasitic copepods (i.e. “sea lice”, Lepeophtheirus salmonis and Caligus clemensi) in their netpens (https://open.canada.ca/data/en/dataset/3cafbe89-c98b-4b44-88f1-594e8d28838d/resource/f6a948f3-504c-34b0-ac30-58a6b06981e6). A thorough description of the counting process can be found in88. The DFO dataset of sea lice counts, collected independently from our eDNA samples, constitutes the only dataset that we could use to ground truth our results from eDNA sampling and laboratory analysis. The DFO Aquaculture Management Division (AMD) provided us with a netpen-level (online version is averaged by farm) dataset of industry sea louse counts from farms in the Broughton Archipelago. Industry sea louse counts could then be matched to eDNA samples for each specific netpen so that we could test for agreement (presence/absence) between the two methods and conduct other statistical analyses.
Statistical analysis
All IA assays, both for eDNA and S. salar tissues, were run in duplicate on the Biomark qPCR dynamic arrays. These analytical duplicates were averaged, and IAs not detected in duplicate were considered non-detections. Field replicates (samples collected simultaneously in parallel on the EZ-eDNA™ pump and filter system) were averaged when both were positive. If only one of two field replicates was positive, the sample was considered positive overall and the positive value was assigned (i.e., the positive value was not averaged with zero). We took this approach because the heterogenous distribution of some IAs (e.g., spore forming IAs) in the water column can lead to false negatives89. This approach of not averaging with zero would only have consequences for figures in this study, as models are all based on presence/absence. For IAs, processed results of qPCR data in Ct were converted to copy number calculated from the standard curve established by the APCs. For salmonid and marine fish assays, results were kept in Ct units. Although limit of detection (LOD) estimates have been developed for all IA assays run (Table S4), we did not apply LOD cutoffs to the data prior to analysis. If a field blank was positive for a given assay during a site visit we removed any detections from collections at the same site, date, and assay with a higher Ct value (weaker detection) from the dataset but left other detections unchanged. We did so under the assumption that detections stronger than those in the field blanks could not be caused by contamination.
During exploratory data analysis we determined that the type of lysis buffer used for eDNA filter extraction (Purelink versus Rebead) had an impact on qPCR results, with the Rebead buffer resulting in lower recovery of total nucleic acids, and particularly lower detection of many RNA viruses. We suspect that Rebead was less efficient at recovering RNA because it does not appear to contain Guanidine Thiocyanate, a potent inhibitor of RNase, which would reduce extraction efficiency, particulary for RNA viruses. Therefore, IA taxa more likely to be collected as RNA than DNA would be under-represented from samples collected from May 2022 onward. Other assays appeared less impacted by the lysis buffer used (e.g., salmonid and marine fish assays). When averaging across assays, we determined that Ct values were approximately 2.5 units greater when Rebead buffer was used instead of Purelink. Therefore, to account for this shift in extraction efficiency, the type of lysis buffer used for each sample was included as a fixed effect in all models. We avoided comparisons of prevalences or loads across time for eDNA detections as these would be impacted by the type of lysis buffer used, at least for some assays, given that the switch in lysis buffer was confounded with sampling date. The overall impact of the switch in lysis buffer midway through the project is that IA presence and load are likely to be underestimated, lending a conservative bias to IA detections.
To test whether the probability of detection was associated with the sample site (active farm versus inactive site) we used a generalized linear mixed model (GLMM) with an autoregressive (AR1) correlation structure to account for temporal autocorrelation at each sampling site. This GLMM structure was used both to characterize IA release and marine fish or salmon exposure to release (Fig. 6). The model included fixed effects for sample type (active farm versus inactive site—the variable of interest) and lysis buffer type (Purelink versus Rebead) as well as random intercepts for the sample month and year combination (spanning the 17 month study period) and the sample location (eleven sites). Contrary to the findings of Barrett et al.9, who found that inactive sites with infrastructure (“fallow” sites in this study) and those with no infrastructure (“decommissioned” sites in this study) had varying magnitudes of difference from active aquaculture sites, our initial models using these multiple categories of inactive sites did not reveal significant differences in eDNA concentrations between fallow and decommissioned sites; therefore, we grouped fallow and decommissioned sites together as inactive sites. Furthermore, fallow sites were not abundant in this study; most samples labeled as inactive sites were decommissioned farms (Table S3). In addition, we pooled samples collected from within netpens with those collected in transects around farms because no barriers prevent the movement of eDNA in and out of nets and transect samples were collected only a short distance (\(\sim\)50 m) away from netpen samples. The response variable was eDNA detection (IAs, marine fish, salmon species) and thus the GLMM was a binomial model with a logit link function. Separate models were run for each response variable using the glmmTMB package90 in R version 4.3.084. Models that did not converge were removed from analysis and not considered for interpretation. We tested for the uniform distribution of residuals by visualizing a qq-plot and a plot of the residuals against the predicted value using the DHARMa package91.
Our experimental design was similar to that used by Shea et al.18, who sampled eDNA adjacent to active and fallow aquaculture sites (although not within netpens) in the Broughton Archipelago and Discovery Islands during the summer months of 2016–2018. We used the publicly available data from Shea et al.18 (https://datadryad.org/stash/dataset/doi:10.5061/dryad.r7sqv9s98) to test for the repeatability of results for assays (n = 10) that overlapped between the two studies. Again, we used GLMMs fitted using the glmmTMB R package90. We did not use an AR1 correlation structure because samples were only collected during a three month period each summer in18. These GLMMs included a random intercept for study year, a random intercept for location, and the fixed variable of interest, sample location (active farms versus inactive sites). To determine whether or not the results were similar between the studies, we used simple linear regression.
To estimate a mean effect of farm status on detection probability in our study and in the reanalyzed data from Shea et al.18, we employed a meta-analytical approach (e.g., Worm et al.92) where each IA’s fitted model (within its respective data set) played the role of a “study” in a typical meta-analysis. We used the R package “meta”93 to conduct an inverse-variance weighted random-effects meta-analysis. The Paule-Mandel estimator was used as the method for estimating among-pathogen variance94,95.
To compare industry sea lice counts to sea lice loads in eDNA samples we used a similar GLMM with AR1 autocorrelation structure and random intercepts as described earlier. We created a binomial GLMM to test the relationship between industry counts (fixed effect of interest) and the likelihood of a detection in eDNA samples, for both C. caligus and L. salmonis. We also created a gaussian LMM to test for a positive correlation between industry counts and log-transformed copepod RNA/DNA copies in eDNA samples. As previously, lysis buffer was included as a fixed effect.
Data availability
The datasets generated during and/or analysed during the current study are available in the following GitLab repository, https://gitlab.com/mgl-published-repo/broughton-archipelago-edna-study-2024/-/tree/main.
References
O’Brien, M. P. et al. Brucellosis transmission between wildlife and livestock in the Greater Yellowstone Ecosystem: Inferences from DNA genotyping. J. Wildl. Dis. c53, 339–343 (2017).
Verhagen, J. H., Fouchier, R. A. & Lewis, N. Highly pathogenic avian influenza viruses at the wild-domestic bird interface in Europe: Future directions for research and surveillance. Viruses 13, 212 (2021).
Foreman, M. G. et al. Modelling infectious hematopoietic necrosis virus dispersion from marine salmon farms in the Discovery Islands, British Columbia, Canada. PLoS One 10, e0130951 (2015).
Oidtmann, B., Dixon, P., Way, K., Joiner, C. & Bayley, A. E. Risk of waterborne virus spread-review of survival of relevant fish and crustacean viruses in the aquatic environment and implications for control measures. Rev. Aquac. 10, 641–669 (2018).
Lyngstad, T. M., Høgåsen, H. R., Jansen, M. D. & Nilsen, A. Risk of disease transfer with wellboats in Norway–technical report. Veterinærinstituttets Oslo Rapportserie 15–2015 (2015).
Bouwmeester, M. M., Goedknegt, M. A., Poulin, R. & Thieltges, D. W. Collateral diseases: Aquaculture impacts on wildlife infections. J. Appl. Ecol. 58, 453–464 (2021).
Lafferty, K. D. et al. Infectious diseases affect marine fisheries and aquaculture economics. Ann. Rev. Mar. Sci. 7, 471–496 (2015).
Bakke, T. A. & Harris, P. D. Diseases and parasites in wild Atlantic salmon (Salmo salar) populations. Can. J. Fish. Aquat. Sci. 55, 247–266 (1998).
Barrett, L. T., Swearer, S. E. & Dempster, T. Impacts of marine and freshwater aquaculture on wildlife: A global meta-analysis. Rev. Aquac. 11, 1022–1044 (2019).
Uglem, I., Karlsen, Ø., Sanchez-Jerez, P. & Sæther, B.-S. Impacts of wild fishes attracted to open-cage salmonid farms in Norway. Aquac. Environ. Interact. 6, 91–103 (2014).
Phillips, M. J. & Subasinghe, R. P. Application of risk analysis to environmental issues in aquaculture. Understanding and applying risk analysis in aquaculture. FAO Fisheries and Aquaculture Technical Paper 101–119 (2008).
Taranger, G. L. et al. Risk assessment of the environmental impact of Norwegian Atlantic salmon farming. ICES J. Mar. Sci. 72, 997–1021 (2015).
Bondad-Reantaso, M. G. et al. Understanding and applying risk analysis in aquaculture (Food and Agriculture Organization of the United Nations, Rome, 2008).
Eichmiller, J. J., Bajer, P. G. & Sorensen, P. W. The relationship between the distribution of common carp and their environmental DNA in a small lake. PLoS ONE 9, e112611 (2014).
Bass, D., Christison, K. W., Stentiford, G. D., Cook, L. S. & Hartikainen, H. Environmental DNA/RNA for pathogen and parasite detection, surveillance, and ecology. Trends Parasitol. 39, 285–304 (2023).
Shea, D. et al. Environmental DNA dispersal from Atlantic salmon farms. Can. J. Fish. Aquat. Sci. 79, 1377–1388 (2022).
Krolicka, A. et al. Sea lice (Lepeophtherius salmonis) detection and quantification around aquaculture installations using environmental DNA. PLoS ONE 17, e0274736 (2022).
Shea, D. et al. Environmental DNA from multiple pathogens is elevated near active Atlantic salmon farms. Proc. R. Soc. B 287, 20202010 (2020).
Gomes, G. B. et al. Use of environmental DNA (eDNA) and water quality data to predict protozoan parasites outbreaks in fish farms. Aquaculture 479, 467–473 (2017).
Atlas, W. I. et al. Trends in Chinook salmon spawner abundance and total run size highlight linkages between life history, geography and decline. Fish Fish. 24, 595–617 (2023).
Peterman, R. M. & Dorner, B. A widespread decrease in productivity of sockeye salmon (Oncorhynchus nerka) populations in western North America. Can. J. Fish. Aquat. Sci. 69, 1255–1260 (2012).
Mordecai, G. J. et al. Aquaculture mediates global transmission of a viral pathogen to wild salmon. Sci. Adv. 7, eabe2592 (2021).
Bateman, A. W. et al. Atlantic salmon farms are a likely source of Tenacibaculum maritimum infection in migratory Fraser River sockeye salmon. Can. J. Fish. Aquat. Sci. 79, 1225–1240 (2022).
Mordecai, G. J. et al. Discovery and surveillance of viruses from salmon in British Columbia using viral immune-response biomarkers, metatranscriptomics, and high-throughput RT-PCR. Virus Evol. 7, veaa069 (2021).
Kumanan, K. et al. Experimental challenge of Chinook salmon with Tenacibaculum maritimum and Tenacibaculum dicentrarchi fulfils Koch’s postulates. bioRxiv 2024–03 (2024).
Frisch, K. et al. Experimental induction of mouthrot in Atlantic salmon smolts using Tenacibaculum maritimum from western Canada. J. Fish Dis. 41, 1247–1258 (2018).
Callier, M. D. et al. Attraction and repulsion of mobile wild organisms to finfish and shellfish aquaculture: A review. Rev. Aquac. 10, 924–949 (2018).
Fjelldal, P. G. et al. Wild atlantic salmon enter aquaculture sea-cages: A case study. Conserv. Sci. Pract. 3, e369 (2021).
Fernandez-Jover, D., Sanchez-Jerez, P., Bayle-Sempere, J. T., Valle, C. & Dempster, T. Seasonal patterns and diets of wild fish assemblages associated with Mediterranean coastal fish farms. ICES J. Mar. Sci. 65, 1153–1160 (2008).
Johannes, M. & Hay, D. Trophic interactions and consumption of wild fish and plankton by cage-reared salmon in British Columbia. Can. Tech. Rep. Fish. Aquat. Sci. 2634 vii + 22. (2006).
Ziegler, F. & Hilborn, R. Fished or farmed: Life cycle impacts of salmon consumer decisions and opportunities for reducing impacts. Sci. Total Environ. 854, 158591 (2023).
Nematollahi, A., Decostere, A., Pasmans, F. & Haesebrouck, F. Flavobacterium psychrophilum infections in salmonid fish. J. Fish Dis. 26, 563–574 (2003).
Madetoja, J., Nystedt, S. & Wiklund, T. Survival and virulence of Flavobacterium psychrophilum in water microcosms. FEMS Microbiol. Ecol. 43, 217–223 (2003).
Bass, A. L., Hinch, S. G., Teffer, A. K., Patterson, D. & Miller, K. S. A survey of microparasites present in adult migrating Chinook salmon (Oncorhynchus tshawytscha) in southwestern British Columbia determined by high-throughput quantitative polymerase chain reaction. J. Fish Dis. 40, 453–477 (2017).
Bass, A. L. et al. The spatial distribution of infectious agents in wild Pacific salmon along the British Columbia coast. Sci. Rep. 13, 5473 (2023).
Bass, A. L. et al. Intrinsic and extrinsic factors associated with the spatio-temporal distribution of infectious agents in early marine Chinook and Coho salmon. Mar. Ecol. Prog. Ser.[SPACE]https://doi.org/10.3354/meps14581 (2024).
Nowlan, J. P., Britney, S. R., Lumsden, J. S. & Russell, S. Experimental induction of Tenacibaculosis in Atlantic Salmon (Salmo salar L.) using Tenacibaculum maritimum, T. dicentrarchi, and T. finnmarkense. Pathogens 10, 1439 (2021).
Valdes, S. et al. First report and characterization of Tenacibaculum maritimum isolates recovered from rainbow trout (Oncorhynchus mykiss) farmed in Chile. J. Fish Dis. 44, 1481–1490 (2021).
Handlinger, J., Soltani, M. & Percival, S. The pathology of Flexibacter maritimus in aquaculture species in Tasmania, Australia. J. Fish Dis. 20, 159–168 (1997).
Chen, M., Henry-Ford, D. & Groff, J. Isolation and characterization of Flexibacter maritimus from marine fishes of California. J. Aquat. Anim. Health 7, 318–326 (1995).
Meyers, T. R. et al. Diseases of wild and cultured fishes in Alaska (Alaska Department of Fish and Game, Fish Pathology Laboratories, Anchorage, 2019).
Jyoti, S. et al. Utilization of publicly available data to summarize spatio-temporal patterns of fish health events of Atlantic salmon (Salmo salar) reported by marine finfish industries in British Columbia (BC), Canada. J. Fish Dis. e14022 (2024).
Bass, A. L. et al. Identification of infectious agents in early marine Chinook and Coho salmon associated with cohort survival. Facets 7, 742–773 (2022).
Avendaño-Herrera, R., Collarte, C., Saldarriaga-Córdoba, M. & Irgang, R. New salmonid hosts for Tenacibaculum species: Expansion of tenacibaculosis in Chilean aquaculture. J. Fish Dis. 43, 1077–1085 (2020).
Nowlan, J. P., Britney, S. R., Lumsden, J. S. & Russell, S. Application of quantitative-PCR to monitor netpen sites in British Columbia (Canada) for Tenacibaculum species. Pathogens 10, 414 (2021).
Nowlan, J. P., Lumsden, J. S. & Russell, S. Advancements in characterizing Tenacibaculum infections in Canada. Pathogens 9, 1029 (2020).
DiCicco, E. et al. Tenacibaculosis in wild-caught, captive Chinook salmon (Oncorhynchus tshawytscha) in British Columbia, Canada. bioRxiv 2023–02 (2023).
Vendramin, N. et al. Presence and genetic variability of Piscine orthoreovirus genotype 1 (PRV-1) in wild salmonids in Northern Europe and North Atlantic Ocean. J. Fish Dis. 42, 1107–1118 (2019).
Turcotte, L. D., Bradshaw, J. C., Polinski, M. P. & Johnson, S. C. Piscine orthoreovirus Genotype-1 (PRV-1) in Wild Pacific Salmon of British Columbia, Canada: 2011–2020. Fishes 8, 252 (2023).
Wang, Y. et al. Infectious agents and their physiological correlates in early marine Chinook salmon (Oncorhynchus tshawytscha). Conserv. Physiol. 11, coad031 (2023).
Di Cicco, E. et al. The same strain of Piscine orthoreovirus (PRV-1) is involved in the development of different, but related, diseases in Atlantic and Pacific Salmon in British Columbia. Facets 3, 599–641 (2018).
Vendramin, N. et al. Piscine orthoreovirus subtype 3 (PRV-3) causes heart inflammation in rainbow trout (Oncorhynchus mykiss). Vet. Res. 50, 1–13 (2019).
Eckstrand, C. D. et al. Detection, sequencing, and tissue distribution of Piscine orthoreovirus 2-like virus in diseased coho salmon in Alaska. J. Vet. Diagnos. Invest. 36, 338–345 (2024).
Garver, K. A. et al. Piscine reovirus, but not jaundice syndrome, was transmissible to Chinook salmon, Oncorhynchus tshawytscha (walbaum), sockeye salmon, Oncorhynchus nerka (walbaum), and Atlantic salmon, Salmo salar l. J. Fish Dis. 39, 117–128 (2016).
Purcell, M. K. et al. Consequences of Piscine orthoreovirus genotype 1 (PRV-1) infections in Chinook salmon (Oncorhynchus tshawytscha), coho salmon (O. kisutch) and rainbow trout (O. mykiss). Journal of fish diseases 43, 719–728 (2020).
Mikalsen, A. B. et al. Characterization of a novel calicivirus causing systemic infection in Atlantic salmon (Salmo salar L.): Proposal for a new genus of Caliciviridae. PloS One 9, e107132 (2014).
Long, A. et al. Distribution and pathogenicity of two cutthroat trout virus (CTV) genotypes in Canada. Viruses 13, 1730 (2021).
Deeg, C. M. et al. Way out there: Pathogens, health, and condition of overwintering salmon in the Gulf of Alaska. Facets 7, 247–285 (2022).
Kent, M. L. et al. Diseases of seawater netpen-reared salmonid fishes in the Pacific Northwest (Department of Fisheries and Oceans Canada, Nanaimo, BC, 1992).
Urawa, S. Effects of Ichthyobodo necator infections on seawater survival of juvenile chum salmon (Oncorhynchus keta). Aquaculture 110, 101–110 (1993).
Mizuno, S. et al. Epizootiology of the ectoparasitic protozoans Ichthyobodo salmonis and Trichodina truttae on wild chum salmon Oncorhynchus keta. Dis. Aquat. Org. 126, 99–109 (2017).
Boerlage, A. S. et al. Epidemiology of marine gill diseases in Atlantic salmon (Salmo salar) aquaculture: A review. Rev. Aquac. 12, 2140–2159 (2020).
Gunnarsson, G. et al. Desmozoon lepeophtherii (microsporidian) infections and pancreas disease (PD) outbreaks in farmed Atlantic salmon (Salmo salar L.). Aquaculture 468, 141–148 (2017).
Serra-Llinares, R. M. et al. Salmon lice infection on wild salmonids in marine protected areas: An evaluation of the Norwegian “National Salmon Fjords”. Aquac. Environ. Interact. 5, 1–16 (2014).
Nekouei, O. et al. Association between sea lice (Lepeophtheirus salmonis) infestation on Atlantic salmon farms and wild Pacific salmon in Muchalat Inlet, Canada. Sci. Rep. 8, 4023 (2018).
Andruszkiewicz Allan, E., Zhang, W. G., Lavery, A. C. & Govindarajan, A. F. Environmental DNA shedding and decay rates from diverse animal forms and thermal regimes. Environ. DNA 3, 492–514 (2021).
Nygaard, M. Environmental DNA for assessing abundance of salmon lice (Lepeophtheirus salmonis) in salmon (Salmo salar) farms. Master’s thesis, UiT The Arctic University of Norway (2018).
Godwin, S., Dill, L., Krkošek, M., Price, M. & Reynolds, J. Reduced growth in wild juvenile sockeye salmon Oncorhynchus nerka infected with sea lice. J. Fish Biol. 91, 41–57 (2017).
Godwin, S. C., Dill, L. M., Reynolds, J. D. & Krkošek, M. Sea lice, sockeye salmon, and foraging competition: Lousy fish are lousy competitors. Can. J. Fish. Aquat. Sci. 72, 1113–1120 (2015).
Hershberger, P., Rhodes, L., Kurath, G. & Winton, J. Infectious diseases of fishes in the Salish Sea. Fisheries 38, 402–409 (2013).
Dempster, T. et al. Coastal salmon farms attract large and persistent aggregations of wild fish: An ecosystem effect. Mar. Ecol. Prog. Ser. 385, 1–14 (2009).
Cohen, B. Commission of inquiry into the decline of Sockeye salmon in the Fraser River (Minister of Public Works and Government Services, Ottawa, Canada, 2012).
Mimeault, C. et al. Assessment of the risk to Fraser River Sockeye Salmon due to piscine orthoreovirus (PRV) transfer from Atlantic Salmon farms in the Discovery Islands area, British Columbia (Canadian Science Advisory Secretariat (CSAS), 2019).
Mimeault, C. et al. Assessment of the risk to Fraser River Sockeye Salmon due to Moritella viscosa transfer from Atlantic Salmon farms in the Discovery Islands area, British Columbia (Canadian Science Advisory Secretariat (CSAS), 2020).
Mimeault, C. et al. Assessment of the risk to Fraser River Sockeye Salmon due to Tenacibaculum maritimum transfer from Atlantic Salmon farms in the Discovery Islands area, British Columbia (Canadian Science Advisory Secretariat, 2020).
Miller, K. M. et al. Infectious disease, shifting climates, and opportunistic predators: Cumulative factors potentially impacting wild salmon declines. Evol. Appl. 7, 812–855 (2014).
Morton, A., Routledge, R., Hrushowy, S., Kibenge, M. & Kibenge, F. The effect of exposure to farmed salmon on piscine orthoreovirus infection and fitness in wild Pacific salmon in British Columbia, Canada. PloS One 12, e0188793 (2017).
Rothman, K. J. & Greenland, S. Causation and causal inference in epidemiology. Am. J. Public Health 95, S144–S150 (2005).
Cantrell, D. L. et al. The use of kernel density estimation with a bio-physical model provides a method to quantify connectivity among salmon farms: spatial planning and management with epidemiological relevance. Front. Vet. Sci. 5, 269 (2018).
Cantrell, D. L., Groner, M. L., Ben-Horin, T., Grant, J. & Revie, C. W. Modeling pathogen dispersal in marine fish and shellfish. Trends Parasitol. 36, 239–249 (2020).
Pert, C. C. et al. Using sentinel cages to estimate infestation pressure on salmonids from sea lice in Loch Shieldaig, Scotland. Aquac. Environ. Interact. 5, 49–59 (2014).
Wallace, I., Gregory, A., Murray, A., Munro, E. & Raynard, R. Distribution of infectious pancreatic necrosis virus (IPNV) in wild marine fish from Scottish waters with respect to clinically infected aquaculture sites producing Atlantic salmon, Salmo salar L. J. Fish Dis. 31, 177–186 (2008).
Duguid, W. D. & Juanes, F. Microtrolling: An economical method to nonlethally sample and tag juvenile pacific salmon at sea. Trans. Am. Fish. Soc. 146, 359–369 (2017).
R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria (2022).
Wessel, P. & Smith, W. H. A global, self-consistent, hierarchical, high-resolution shoreline database. J. Geophys. Res. Solid Earth 101, 8741–8743 (1996).
Deeg, C. M. et al. eDNA sampling systems for salmon ecosystem monitoring. bioRxiv (2024).
Miller, K. M. et al. Report on the performance evaluation of the Fluidigm BioMark platform for high-throughput microbe monitoring in salmon. DFO Can. Sci. Advis. Sec. Res. Doc. 2016/038. xi + 282 p. Tech. Rep., Fisheries and Oceans Canada Science Advisory Secretariat Research Document 2016/038 (2016).
Godwin, S. C., Krkošek, M., Reynolds, J. D. & Bateman, A. W. Bias in self-reported parasite data from the salmon farming industry. Ecol. Appl. 31, e02226 (2021).
Sieber, N., Hartikainen, H. & Vorburger, C. Validation of an eDNA-based method for the detection of wildlife pathogens in water. Dis. Aquat. Org. 141, 171–184 (2020).
Brooks, M. E. et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. https://doi.org/10.32614/RJ-2017-066 (2017).
Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models (2022). R package version 0.4.6.
Worm, B. & Myers, R. A. Meta-analysis of cod-shrimp interactions reveals top-down control in oceanic food webs. Ecology 84, 162–173 (2003).
Balduzzi, S., Rücker, G. & Schwarzer, G. How to perform a meta-analysis with R: a practical tutorial. Evidence-Based Mental Health (2019).
Paule, R. & Mandel, J. Consensus values and weighting factors. J. Res. Natl. Bur. Stand. 87, 377. https://doi.org/10.6028/jres.087.022 (1982).
Veroniki, A. A. et al. Methods to estimate the between-study variance and its uncertainty in meta-analysis. Res. Synth. Methods 7, 55–79. https://doi.org/10.1002/jrsm.1164 (2016).
Acknowledgements
The authors would like to acknowledge that this work was conducted within the territories of the Mamalilikulla, ‘Namgis, and Kwikwasut’inuxw Haxwa’mis First Nations and are thankful for the opportunity to collaborate. In addition to several of the co-authors, BATI leadership included Brad Puglas (MFN), Don Svanvik (NFN), Matt Ambers (NFN), and Eric Joseph (KHFN). BATI staff and field technicians included Andrew Wadhams, Jack Alfred, Jonah Johnson, Peter Mountain, Roy Mountain-Wadhams, Joey Webber, Dennis Johnson, Gerald Alfred, Scott Rufus, Daniel Wadhams, Tre Alfred, Emily Wisden-Seaweed, and Una Kim. Diana Drozdzik and Christopher Tam conducted many eDNA extractions. Kalya Zielke, Jonathan Archembault, Kaleb Mantha-Rensi sampled eDNA in and around the farms, dissected fish, managed data, and completed many other logistical tasks. Will Bugg, Gideon Mordecai, Sean Godwin, and Christoph Deeg provided valuable feedback on the analytical approach. This project was funded through the Indigenous Monitoring and Inspection Plan (IMIP) negotiated with the Aquaculture Industry to oversee operations of the Fish Farms by First Nations in the Broughton Archipelago.
Author information
Authors and Affiliations
Contributions
E.D.C., R.J., J.P., V.I., N.B.D., A.W.B., and K.M.M. conceived the study design, E.D.C. conducted the field work, K.H.K. and S.L. conducted laboratory analysis, A.L.B. and A.W.B. undertook statistical analysis. A.L.B led writing. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bass, A.L., DiCicco, E., Kaukinen, K.H. et al. Infectious agent release and Pacific salmon exposure at Atlantic salmon farms revealed by environmental DNA. Sci Rep 14, 31488 (2024). https://doi.org/10.1038/s41598-024-83250-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-83250-5
