
Abstract
Heart failure with preserved ejection fraction (HFpEF) is increasingly common but its pathogenesis is poorly understood. The ability to assess genetic and pharmacologic interventions is hampered by the lack of robust preclinical mouse models of HFpEF. We developed a novel “two-hit” model, which combines obesity and insulin resistance with chronic pressure overload to recapitulate clinical features of HFpEF. C57Bl6/NJ mice fed a high-fat diet (HFD) for > 10 weeks were administered an AAV8-driven vector resulting in constitutive overexpression of mouse Renin1d. HFD-Renin (aka “HFpEF”) mice demonstrated obesity and insulin resistance, moderate left ventricular hypertrophy, preserved systolic function, and diastolic dysfunction indicated by echocardiographic measurements; increased left atrial mass; elevated natriuretic peptides; and exercise intolerance. Transcriptomic and metabolomic profiling of HFD-Renin myocardium demonstrated upregulation of pro-fibrotic pathways and downregulation of metabolic pathways, in particular branched chain amino acid catabolism, similar to human HFpEF. Treatment with empagliflozin, an effective but incompletely understood HFpEF therapy, improved multiple endpoints. The HFD-Renin mouse model recapitulates key features of human HFpEF and will enable studies dissecting the contribution of individual pathogenic drivers to this complex syndrome. Additional preclinical HFpEF models allow for orthogonal studies to increase validity in assessment of interventions.
Similar content being viewed by others

Introduction
Heart failure with preserved ejection fraction (HFpEF) is a complex disease, and despite many advances in research, its underlying mechanisms are not yet fully understood1. This is due, in part, to the lack of pre-clinical small animal models that faithfully recapitulate the human condition. There is no consensus on the required elements to define rodent HFpEF2,3,4,5. Consequently, current rodent models have notable downsides, including significant lead time hindering rapid experimentation6, need for surgical expertise7,8, predominant hypertensive phenotype without metabolic dysregulation9, or inability to use genetic manipulation, as in the ZSF-1 rat or leptin signaling mutant models10,11,12,13. Additionally, the best characterized mouse model to date using L-NAME and high-fat diet (HFD)14, induces endothelial dysfunction using an artificial, pharmacological inhibitor of vascular NO signaling and some, including our lab, have shown that this model is not consistently reproducible15,16.
We sought to devise a complementary “two-hit” model of mouse HFpEF that combines the most common human drivers of the metabolic phenogroup of human HFpEF, diet-induced obesity (DIO), and chronic hypertension. Among HFpEF patients, the phenogroups with obesity/diabetes, abnormal systemic metabolism, liver and renal dysfunction, and high renin level exhibit the worst overall prognosis and lowest survival probability17,18. Our goal was to establish a mouse model that exhibits both cardiac and extra-cardiac manifestations of HFpEF and meets the following criteria: (1) left ventricular (LV) hypertrophy (LVH) driven by a pathogenic driver relevant to the human condition, (2) obesity and insulin resistance, (3) LV diastolic dysfunction, (4) preserved LV systolic function, and (5) exercise intolerance. We also sought to develop a model with relatively balanced contributions of increased peripheral vascular resistance and obesity with insulin resistance. The model described here, termed “HFD-Renin” mice, involves modest, chronic hypertension achieved by adeno-associated virus overexpression of renin (AAV8-Renin) to activate the renin-angiotensin-aldosterone system (RAAS) in the setting of diet-induced obesity producing a modest but significant LVH, LV diastolic dysfunction with stable systolic function, exercise intolerance, and obesity-related glucose intolerance and insulin resistance. This model utilizes common pathogenic drivers resulting in features of human HFpEF and allows for assessment of the relative contribution of each driver independently18,19. Given concerns of reproducibility with prior published models, we developed and conducted complementary studies at two sites in a coordinated manner to verify reproducibility. Renin was chosen as a tool for inducing modest renovascular hypertension, a critical driver of HFpEF. The relevance of this approach relates to the observation that the RAAS system is activated in most pathophysiological states that contribute to HFpEF. In contrast, L-NAME is an artificial pharmacologic intervention. Admittedly, AAV-mediated delivery of renin is not physiologic. No small animal model will be a perfect representation of this complex human disease, including the one presented here. However, the availability of multiple verified HFpEF models will be valuable to the field, allowing for more rigorous assessment of therapeutic interventions via orthogonal studies.
Results
Development and optimization of AAV-mediated renin delivery to achieve hypertension-related LV hypertrophy and remodeling in mice
We sought to use an AAV8 delivery system to achieve a mild degree of hypertension in mice. An AAV-Renin vector (details of the construct provided in Methods) or null backbone vector (control) were administered in order to constitutively express a cleavable form of renin in the liver20. Dose-ranging studies demonstrated a dose-dependent increase in circulating mouse renin concentration (Supplementary Fig. S1A). No change in body weight was observed during the six-week study period (Supplementary Fig. S1B). Implantable telemetry testing revealed a dose-dependent, modest (20-40mmHg) increase in systolic blood pressure (Supplementary Fig. S1C), which was amenable to treatment with standard of care angiotensin converting enzyme (ACE) inhibitor enalapril. Consistent with the hypertensive state, increased normalized biventricular heart weight, left atrial enlargement, and modest lung congestion developed in the AAV-renin-treated mice (Supplementary Fig. S1D). Gene markers of LV remodeling were also induced, including a decrease in the myosin heavy chain 6 (Myh6)/Myh7 ratio (Supplementary Fig. S1E).
The dose-ranging studies were repeated in HFD-treated mice. Mice were started on HFD at ~ 8 weeks of age prior to AAV administration at ~ 20 weeks of age. Dose-dependent LVH was achieved in the context, but required higher viral doses presumably due to presumed decreased viral transduction in a fatty liver (Supplementary Fig. S1F). Notably, at the highest doses, significant loss of body weight was observed (Supplementary Fig. S1G). The dose-ranging studies allowed for the selection of similar, though not identical, doses at both institutions (lower for chow-fed animals, higher for HFD-fed animals) that resulted in the optimal 2-3x increase in circulating renin and LVH with minimal perturbance in body weight. Potential effects of viral-induced inflammation were controlled for by using a null virus in the control arms. Plasma aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were measured 6 weeks post-AAV8 injection (Supplementary Fig. S1H) and demonstrated no elevation in the chow-fed animals, and mild elevation (2-3x upper limit of normal) in HFD null and HFD-Renin conditions, suggesting that the transaminitis is a result of diet-induced steatohepatitis.
The HFD-Renin model results in features of HFpEF, including systemic metabolic derangements, LV hypertrophy, diastolic dysfunction, and exercise intolerance
Having established that AAV-Renin produces modest hypertension and cardiac hypertrophic remodeling, this effect was overlayed on HFD-fed animals. For the HFpEF model, 8- to 10-week-old mice fed chow (control) or HFD for 10 weeks were injected with AAV-Renin or null vector (control) (Fig. 1A). Renin dosing was calibrated to achieve a 2-3x increased in circulating protein levels (Fig. 1B). HFD feeding reliably resulted in obesity, though AAV-Renin dosing at week 10 caused a modest flattening in weight gain (Fig. 1C). The obese mice had impaired glucose tolerance and insulin resistance (Fig. 1D and Supplementary Fig. S2A). This corresponded with fasting hyperglycemia, hyperinsulinemia, and hypercholesterolemia (Supplementary Fig. S2B). There were no observed metabolic effects due to AAV-Renin (see chow + Renin cohort).

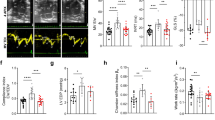
Novel “two-hit” HFpEF mouse model driven by renin overexpression and HFD. (A) Schema of HFD-Renin model, where C57BL/6NJ mice are preconditioned on HFD for 10 weeks prior to injection of AAV8-Renin with endpoints at week 16–18. Created with Biorender.com. (B) Serum renin levels one week post-injection. (C) Weight trend (left) and terminal weights (right). (D) Glucose tolerance test with associated area-under-curve (right) in four-hour fasted male littermate mice. (E) Echocardiographic measures at 16 weeks: LV ejection fraction (EF) and interventricular septal thickness in diastole (IVSd). (F) Tibia length-normalized gross biventricular weight (BiV/TL). (G) Representative m-mode echocardiograms, red bar notes LV posterior wall dimension in diastole. N = 10–15 per conditions. Data displayed as mean ± SEM. Statistical analyses used one-way analysis of variance (ANOVA) followed by multiple-comparisons test. P values < 0.05 displayed on graphs.
All cohorts had stable LV systolic function at six weeks post-injection (Fig. 1E and Supplementary Table S2). AAV-Renin resulted in 30–40% increase in LV intraventricular septal wall thickness (IVSd) and LV mass index (LVMI) as determined by echocardiography (Fig. 1E-G, and Supplementary Table S2). Normalized ventricular weights (biventricular weight to tibia length, BiV/TL) similarly increased, with an additive effect between HFD and renin expression (Fig. 1E). Despite the LVH, there was no significant change in ventricular volumes (Supplementary Table S2).
To assess diastolic function, multiple complimentary endpoints were utilized. HFD or AAV-Renin alone did not produce significant changes in echo-based parameters. An additive effect of HFD and AAV-Renin was observed in mitral valve inflow and tissue doppler imaging parameters (increased E/e’, a trend in Tei index, no change in E/A ratios, Fig. 2A and B) as well as global longitudinal and radial strain and strain rates (Fig. 2C and Supplementary Table S2). Radial strain appeared to be a more sensitive marker of dysfunction in this model. As observed in the human HFpEF condition, the mouse left atria (LA) was impacted, including significantly increased in normalized LA weight relative to the control (Fig. 2D) and trended toward increased LA volume measured by echo (Fig. 2E), consistent with LV diastolic dysfunction. LA strain, a standard human clinical parameter, was reduced. Similarly, pulmonary vein doppler flow was significantly decreased in the HFD Renin cohort, indicative of decreased LV compliance (Fig. 2F). Expression of elevated natriuretic peptides Nppb (e.g., BNP, brain natriuretic peptide) and Nppa, a diagnostic criterion in human disease, were significantly elevated in heart tissue from HFD Renin animals compared to other treatment groups (Fig. 2G).

HFD-Renin HFpEF model exhibits LV diastolic dysfunction, abnormal left atrial (LA) function, and exercise intolerance. (A) Representative echocardiographic mitral valve inflow and tissue doppler images with (B) measurements of diastolic function including E/A and E/e’ ratios, and Tei index (n = 5–15). (C) Echocardiographic left ventricular global longitudinal and radial strain (n = 4–8). (D) Tibia length-normalized LA gross weight. Representative (E) LA minimum volume and reservoir strain, and (F) Doppler echo-measured pulmonary vein flows during systole (S) and diastole (D). (G) QT-PCR from bulk ventricle RNA for indicated genes. (H) Treadmill exercise testing to exhaustion. Data displayed as mean ± SEM. One-way analysis of variance (ANOVA) followed by multiple-comparisons test. P values < 0.05 displayed on graphs.
Exercise intolerance is the most consistent symptom in patients with HFpEF, though the etiology is debated21,22. Mice underwent exercise testing to exhaustion to determine the combinatorial effects of DIO and hypertension. HFD Renin animals displayed greatly diminished exercise tolerance in both time and distance to exhaustion as well as total work, which adjusts for body mass differences (Fig. 2H). Chow AAV-Renin mice did not perform differently from control mice. Accordingly, the dominant effect was due to HFD, as has been demonstrated in prior studies7.
HFpEF is more prevalent in human females, yet as a group, they are relatively understudied in clinical trials23. We tested the cumulative effects of HFD and AAV-Renin in a cohort of female mice using the same protocol as that for males (Supplementary Fig. S3A and S3B). Female mice fed HFD became obese, but to a lesser extent than male mice, consistent with the pre-clinical rodent literature24,25,26. The HFD-fed groups developed insulin resistance (data not shown). Renin-treated groups had preserved systolic function (Supplementary Fig. S3C). Both HFD and AAV-Renin resulted in mild LVH with an additive degree (20–30%) in the HFD-Renin cohort (Supplementary Fig. S3D). Diastolic parameters were mixed: Echo parameters for diastolic dysfunction, including E/A, E/e’, and isovolumic relaxation time (IVRT), were unchanged in the HFD-Renin group compared to controls (Supplementary Fig. S3E). However, decreased pulmonary vein doppler flow and LA enlargement, consistent with decreased ventricular compliance, were observed in the HFD Renin group (Supplementary Fig. S3F and S3G). Lastly, female HFD-Renin mice had decreased exercise performance in an additive manner (Supplementary Fig. S3H). Taken together the HFpEF phenotype driven by HFD-Renin was milder in female mice.
Two-hit HFpEF model results in cardiac fibrosis
Cardiac fibrosis is a hallmark of human HFpEF, and variably demonstrated in current models6,9,14. Analysis of histologic sections of LV free wall myocardium stained with picrosirius red demonstrated a trend toward increased collagen deposition in the HFD-Renin group (Fig. 3A-B). Perivascular changes were more apparent than interstitial fibrosis, similar to the known effects of angiotensin II in rodents9. These observations correlated with increased expression of markers associated with fibrosis, fibroblast activation, and extracellular matrix production by QT-PCR (Fig. 3C).

HFD-Renin HFpEF results in increased cardiac fibrosis. Representative histology with picrosirius red and fast green counterstain of (A) whole heart (scale bar 3 mm) and (B) 20x magnification (scale bar 200 μm) demonstrating perivascular fibrosis, quantified from two separate regions in n = 4 independent samples. (C) QT-PCR from cardiac tissue for gene markers of fibrosis and myofibroblast activation, n = 6. Data displayed as mean ± SEM. One-way analysis of variance (ANOVA) followed by multiple-comparisons test. P values < 0.05 displayed on graphs.
Transcriptomic and metabolomic changes mirror observations in human HFpEF
Targeted gene expression changes suggested cardiac remodeling largely predicated on fibrotic and hypertrophic pathways (Figs. 2H and 3C). To further understand the gene expression signature of the HFD-Renin model, we performed bulk RNA sequencing (RNA-seq) of LV samples from control, DIO, hypertensive, and HFpEF samples. Principal component analysis and heat mapping suggested that the null-treated chow and HFD groups were notably similar, whereas the chow Renin group was quite distinct (Fig. 4A and B). The addition of HFD to renin caused a left shift, repositioning the HFpEF group to overlay with control and HFD only groups. We analyzed the effects of the individual drivers and combined effect on differentially expressed genes (DEGs). DIO primarily drove downregulation of genes associated with metabolic pathways and cell signaling (839 downregulated DEGs with adjusted p-value < 0.01) (Supplementary Fig. S4A). Conversely, AAV-Renin was responsible for a large upregulation of genes (927 DEGs) involved with proliferative growth and hypertrophic signaling pathways (Supplementary Fig. S4B). Interestingly, several pathways moved in opposing directions (e.g., cAMP signaling, calcium signaling, and circadian entrapment downregulated by HFD and upregulated by Renin). Consequently, some of these pathways were not present in the gene ontology and KEGG pathway analyses in the HFD-Renin condition. Compared to Chow null, the HFD-Renin cohort resulted in a unique set of 498 upregulated and 472 downregulated genes (Fig. 4C), with a separate set of differentially regulated pathways from either the DIO or hypertensive components. This suggests an additive effect, resulting in downregulated metabolic pathways and upregulation of ECM and cardiac hypertrophic growth (Fig. 4D). These signatures were consistent with published transcriptomic data from HFpEF patients19. Top DEGs are listed in Supplementary Figure S4C.

Cardiac transcriptomic and metabolomic profiling of HFpEF hearts shows strong signatures of reduced mitochondrial fuel (BCAA) and activation of ECM/fibrosis. (A) Principal component (PC) analysis and (B) heatmap of ventricular RNAseq comparing control and HFD-Renin groups (n = 5–6). (C) Volcano plot of differentially expressed genes in HFpEF (vs. control) hearts (n = 5–6). (D) KEGG pathway analysis of upregulated (yellow bars) and downregulated (blue bars) pathways. (E) Targeted metabolomics from heart tissue displaying short-chain acylcarnitine species (n = 5–7). Data displayed as mean ± SEM. Unpaired, 2-tailed Student’s t-test, with p values as displayed or * <0.05.
Given the observed changes in mitochondrial pathways, including downregulation of branch chain amino acid metabolism and oxidative phosphorylation in the HFD-Renin state, we performed targeted metabolomic analysis in control and HFpEF hearts. Short-chain acylcarnitines, including C03-DC and C04-OH isobutryl, intermediates in BCAA catabolism, were decreased, consistent with the observed reduction in BCAA degradation pathways in the RNA-seq dataset (Fig. 4E). There were few changes in organic acids or medium- and long-chain acylcarnitines (Supplementary Fig. S5).
Human HFpEF standard-of-care therapy improves a subset of pathophysiologic parameters in the HFD-Renin model
Since the HFD-Renin model demonstrated changes in metabolism, cardiac remodeling, and exercise tolerance endpoints that capture critical translatable features in HFpEF patients, we next assessed the effects of the newly approved HFpEF therapy sodium glucose cotransporter 2 inhibitor (SGLT2i) empagliflozin (empa) in this model27. Empa was administered concurrent with AAV injection as a preventive treatment intervention modality for a total of 14 weeks at the Pfizer site (Fig. 5A). Treatment with SGLT2i resulted in a slight but significant improvement in body weight, a trend toward reduced blood glucose, with significant reduction in hyperinsulinemia/HOMA-IR (Fig. 5B). No differences were observed in systolic function (Fig. 5C). There was a reduction in echocardiographic LVH and normalized ventricular gross weight following empa therapy (Fig. 5D). Radial strain and echocardiographic measures of diastolic dysfunction were largely unchanged (e.g., IVRT, Fig. 5E), but there was a trend toward diminished normalized LA mass, which correlated with reduced liver congestion. Exercise testing demonstrated significant improvement in maximal distance run prior to exhaustion (Fig. 5F). Confirmatory studies conducted in parallel at Penn with a shorter time course of treatment and lower dose demonstrated similar findings (Supplementary Fig. S6). These results suggest that SGLT2i may exert a positive benefit through effects on improved metabolic state, cardiac hypertrophic remodeling, and exercise tolerance and underscore the importance of a model that exhibits multiple pathophysiological responses to determine the impact of a given intervention. Importantly, this study design also demonstrated that the HFpEF characteristics do not degenerate into HF with reduced ejection fraction over > 12 weeks of HFD-Renin exposure (e.g., systolic function remains stable, albeit slightly lower than control animals).

SGLT2 inhibitor treatment improves HFpEF in mice. (A) Schema for empa treatment regimen with HFD-Renin HFpEF model conducted at two independent research sites. (B) Biophysical parameters of Pfizer cohort, n = 12–13. (C) Echocardiographic measures of systolic function and strain. (D) LV hypertrophy measures by echo and normalized biventricular weight. (E) Diastolic parameters including IVRT, atrial enlargement, and liver congestion. (F) Maximal exercise testing. Data displayed as mean ± SEM. One-way analysis of variance (ANOVA) followed by multiple-comparisons test. P values < 0.05 displayed on graphs. RWT regional wall thickness, IVRT isovolumic relaxation time, SR strain rate.
Discussion
Understanding the underlying pathophysiology and identifying new treatment strategies for HFpEF has been hindered, in part, by a lack of mouse models of HFpEF that faithfully recapitulate the human disease state. This challenge is further complicated by the existence of multiple human HFpEF phenogroups necessitating multiple pre-clinical models. The ease of genetic manipulation in mice allows rigorous testing of new targets using genetic loss-of-function approaches complemented by pharmacologic interventions. The two most common drivers in human HFpEF are obesity and hypertension—each to varying extent—ranging from obesity with dominant hypertension to modest chronic hypertension/renal and vascular dysfunction. We developed a “two-hit” HFpEF model that combines persistent, mild hypertension through renin overexpression with metabolic dysfunction from DIO to simulate the human phenotype. A unique feature of these studies is that two separate facilities assessed the models in parallel, increasing confidence in the results and reproducibility for the model. Following careful dose assessment, AAV-Renin could be administered to reliably effect a 20-40mmHg increase in systolic blood pressure and ~ 30% LV hypertrophic response without weight loss and preserved systolic function. Activation of the RAAS was the not the goal of this model, as many trials of RAAS antagonists have not shown benefit in HFpEF treatment;28,29,30 rather, we were co-opting a physiologic pathway to induce hypertension, a clear component of the disease state.
We comprehensively phenotyped this new HFpEF model for cardiac and extra-cardiac physiologic endpoints, confirming obesity, insulin resistance, hypertension, preserved systolic function, concentric cardiac hypertrophy, left atrial enlargement, diastolic dysfunction, elevated cardiac natriuretic peptide gene expression, and impaired exercise capacity. To assess diastolic dysfunction, we relied on a plurality of endpoints, including novel left atrial and classic mitral valve echocardiographic measures, gross and normalized tissue weights, exercise intolerance, and natriuretic peptide expression. Multiple metrics of abnormal diastology were present (ventricular strain, E/e’, increased trend in Tei index and IVRT), as well as novel echocardiographic parameters quantifying LA strain, LA volume, and pulmonary vein doppler patterns, which have been shown to robustly correlate with diastolic dysfunction9. Increased hepatic and pulmonary congestion and elevated BNP, a marker of cardiac congestion, mirror the exercise intolerance critical to the human disease. The need for a plurality of markers is critical since E/A or E/e’ individually can be an unreliable representation of diastolic dysfunction in mice31,32, requiring technical expertise to obtain heart rates slow enough to see E and A separation but not so slow as to affect ventricular function and loading conditions. In addition, multiple pathophysiological endpoints also allow for assessment of therapies that may only impact a subset as we observed with intervention of SGLT inhibitor therapy. We acknowledge the model is limited by modest sex differences, where male mice have greater evidence of HFpEF compared to female mice. This observation is disparate from the human condition but is similar to other mouse HFpEF models11,33, and highlights a need in the field for better female HFpEF modeling.
This model allows for assessing how individual pathophysiological interventions that are known drivers (LV pressure overload, caloric excess, abnormal neurohormonal axis) contribute to the development of HFpEF. The ability to isolate component drivers is a powerful technical aspect to the model, querying the specific and additive contributions of hypertension and metabolic dysfunction to the overall HFpEF phenotype. For some metrics, the hypertensive component was the dominant driver, such as measures of LA size and LV hypertrophy. A primary dietary effect was seen in assessing exercise capacity, where HFD largely determined response to exercise. However, for many endpoints, including hypertrophy, diastolic dysfunction, and fibrosis, a significant additive effect was observed beyond either constituent effect.
Ventricular transcriptomic interrogation (RNAseq) was used to assess component effects, identify signature transcriptional changes in the HFpEF state, and allow for comparison with recent published human data to further assess relevance19. Gene expression changes caused by the individual drivers had incomplete similarity to the patterns seen in the HFpEF state, further emphasizing the “syndrome” aspect to this disease—multiple, multiorgan, chronic inputs are required. Among the top upregulated genes in both datasets, many are implicated in extracellular matrix remodeling, cardiac fibrosis, and inflammation (e.g., Acta1, Edn3, Col8a1, Postn, Adamtsl2). For example, Acta1 has previously been demonstrated to be upregulated in mouse HFpEF LV34. Cartilage intermediate layer protein 1 (CILP1), reported as a novel sensitive biomarker for cardiac fibrosis35,36,37, and disintegrin-like and metalloproteinase domain with thrombospondin type 1 motifs-like (Adamtsl2), an extracellular matrix glycoprotein upregulated in human heart failure, act to modulate TGFβ signaling in cardiac fibrosis38. There was also strong evidence for altered mitochondrial function, including diminished branched chain amino acid catabolism. Targeted metabolomics from cardiac tissue bore out the decrease of BCAA metabolites in the HFpEF state, which is similar to observed changes in human HFpEF samples39.
To test the translational utility of this HFpEF model, mice were treated with empa, a potent sodium-glucose co-transporter 2 inhibitor (SGLT2i) which has demonstrated improvement in reducing heart failure hospitalization in HFpEF40,41. Preclinical data has shown variable benefit of SGLT2i in mouse models of HFpEF, including improvement of fibrosis42, improved hypertrophy with43 or without improved diastolic dysfunction7, and improved electrophysiologic parameters11. We demonstrate consistent, albeit subtle improvement in a subset of cardiac and extra-cardiac endpoints, including body weight, insulin sensitivity, LA and LV hypertrophy, and exercise performance, with trends toward improved strain. The benefit of a model that exhibits multiple phenotypic features of HFpEF is emphasized by these results given that therapeutic interventions will likely impact only a subset of the many drivers of this disease. The results are also reproducible in two separate experimental settings using testing protocols that are similar, not identical; for instance, exercise testing that measured sprint (Pfizer) versus high intensity endurance (Penn). These results highlight the value of this model for translational and mechanistic studies including the potential for genetic interventions that can provide insight into the relevant pathogenic origins of HFpEF.
In summary, we present a tractable, consistent model of mouse HFpEF that recapitulates multiple concerted endpoints observed in the human condition. Compared to the models currently available, this model utilizes AAV-Renin to create a stable, consistent hypertension not affected by food or water intake, or time-limited drug dosing via implantable minipumps6,8,9,14,44. Moreover, renin-driven hypertension is a well-known driver of human hypertension and cardiac remodeling. Additionally, there is no artificially imposed vascular dysfunction from an exogenous chemical, such as L-NAME. Hijacking the renin-angiotensin-aldosterone system as a tool to induce hypertension is physiologically relevant to human HFpEF patients, mimicking the significant increase of plasma renin levels observed in the TOPCAT HFpEF clinical trial phenogroup 3 patients17. Accepting the limitation of using rodents to model human disease, we believe this model, used in conjunction with others previously developed, will allow for critical investigation into a complex and poorly understood form of heart disease.
Methods
In vivo studies
Mouse studies were performed in parallel at two separate institutions: University of Pennsylvania (Penn) and Pfizer. Penn utilized male and female C57BL/6NJ mice (Jackson Laboratory, Stock 005304) age 8–10 weeks old, and randomized to chow or 60%kcal HFD (D12492i, Research Diets) arms for 10 weeks. Pfizer utilized 8-week-old C57BL/6 N male mice fed chow or 60% HFD-fed (Taconic, DIO-B6NTac) for 12 weeks. Mice were housed in a facility under a 12-hour light/12-hour dark cycle, at 72 degrees Fahrenheit and 40–50% humidity. For tissue collection, mice were fasted for four hours prior to anesthesia with intraperitoneal pentobarbital (Sagent, 100 mg/kg) and euthanasia by cardiectomy. Tissue weights were normalized to tibia length. Lungs and livers were excised and weighed “wet” and, for lungs only, following 48 h at 37°C “dry.” For empagliflozin (empa) studies, mice were randomized to treatment with drug (Advanced ChemBlocks Inc, Cat#: G-7261) admixed at 500 mg/kg (Pfizer) or 50 mg/kg (Penn) in 60%kcal HFD, validated to match the human pharmacologic effects45.
AAV8-Renin generation and treatment
Mouse Ren1d cDNA was purchased from Horizon Discovery (Clone ID 30313659, Sequence BC061053) and mutated at F61R and P65S as previously described (to enable prorenin cleavage to renin in non-renal locations, see Supplementary Fig. S1A) using the In-Fusion HD system (Clontech, #639650)20. Construct was subcloned in the pAAV.TBG.PI.Null.bGH (Addgene #105536) plasmid provided by the Vector Core at Penn. All cloning and mutagenesis primer sets are listed in Supplementary Table S1. pAAV.TBG.PI.mRen1(F61R)(P65S).bGH (AAV8-Renin) or null constructs were packaged at the Vector Core. For the Pfizer model, AAV was produced at the University of Massachusetts Medical Center Gene Therapy Center and Vector Core. Eighteen to 20-week-old lean or obese mice were randomized to receive AAV8-Renin or AAV8-null virus. Mice were anesthetized using 2.5ppm isoflurane prior to injection via the medial canthus into the retro-orbital venous plexus.
Based on extensive dose-response testing, doses were selected to generate a 2-3x increase over basal renin expression but without affecting weight gain (Supplementary Fig. S1). Given viral production occurred in two independent facilities, empiric doses were as follows: Penn lean mice received 3.0 × 10^10 and HFD-fed mice received 1.0 × 10^11 viral genomes (vg) diluted in sterile saline, injected in a volume of 100ul. Pfizer lean mice received 1.0 × 10^10 and HFD-fed 3 × 10^11 vg. Higher doses of AAV-Renin were required to raise plasma renin levels in obese mice, potentially due to lower transduction efficacy of AAVs in the fatty liver of the HFD model. Blood samples were obtained one week post-injection and assayed for circulating renin using the mRen1 DuoSet ELISA (Bio-techne, #DY4277) per manufacturer protocol. Samples from mice receiving AAV8-Renin were diluted 1:50. Aspartate aminotransferase (AST) and alanine aminotransferase (ALT) assays were performed from blood collected in EDTA tubes and placed on ice. Plasma was collected after centrifugation at 10,000 RPM for 10 min at 4°C, and stored at -80°C until use. AST/ALT assays were performed on the Siemens ADVIA Chemistry XPT system (ALT #07501976, and AST #07499718).
Telemetry study
Radiotelemetry recording of arterial pressure was performed in 8- to 10-week-old male C57BL/6 N mice, implanted with telemeters (PA-C10, Data Science) by Envigo (Invito, Indianapolis, IN). After one week, telemetry units were activated, and arterial pressure was recorded for baseline recording. Measurements were recorded for five seconds every minute for the length of the study. Mice were randomized and dosed with either 1 × 10^10 AAV null or 3 × 10^9, 1 × 10^10 or 3 × 10^10 AAV-Renin. Two weeks post-AAV dosing, mice were treated with enalapril (200 mg/L; Sigma) in the drinking water for three days, followed by switching back to normal drinking water.
Mouse echocardiography
Ultrasound examination of the left ventricle was performed independently at both study sites using a Fujifilm VisualSonics Ultrasound System. At the Rodent Cardiovascular Phenotyping Core of the University of Pennsylvania Cardiovascular Institute, adult mice were anesthetized with an intraperitoneal injection (0.05 mg/g) of 2% avertin (to maintain a heart rate close to 600 beats per minute or higher for the evaluation of LV systolic function). Hair was removed from the anterior chest using chemical hair remover, and the animals were placed on a warming pad in a left lateral decubitus position to maintain normothermia. Ultrasound gel was applied to the chest. Care was taken to maintain adequate contact while avoiding excessive pressure on the chest. 2D long-axis and short-axis M-mode images were obtained. Diastolic function–related parameters were evaluated using a modified four-chamber view. Transmitral inflow velocities, tissue Doppler, pulmonary vein Doppler, and left atrial strain measurements, as previously described9, were obtained following intraperitoneal injection of zatebradine (funny current inhibitor, 0.008 mg/gm body weight) to reduce heart rate to 425–475 bpm (15–25% heart rate reduction compared with baseline) when needed. After completion of the imaging studies, mice were allowed to recover from anesthesia and returned to their cages. At Pfizer, animals were anesthetized at 2% isoflurane (chow) or 3% isoflurane (HFD) with heart rate goals of 300-450 bpm for diastolic parameters and > 400 bpm for functional parameters. Images were obtained similarly. Imaging and image analysis was performed by blinded sonographers and analyzed using Vevo Lab software (VisualSonics).
Exercise testing
Mice were exercised to exhaustion utilizing similar protocols. At Penn, mice underwent three consecutive days of exercise on a five-lane treadmill with shock grid (Maze Engineering) starting with two days of an acclimation protocol of nine minutes at 10 m/min followed by one minute at 20 m/min at a grade of 6–8 degrees. Testing was performed on a 25-degree treadmill, starting at 10 m/min for three minutes followed by a ramp over three minutes to a final speed of 18 m/min, continuing to exhaustion. At Pfizer, animals were acclimated to the Exer3/6 Animal Treadmill (Columbus Instruments) for two days prior to testing (Day 1: five minutes off and five minutes at 2.5 m/min; Day 2: five minutes at 2.5 m/min followed by five minutes at 5 m/min). Testing was performed at a 5 degree incline starting at 5 m/min for two minutes, increased to 8 m/min for two minutes and then increased an additional 2 m/min for every two minutes until the animal was exhausted. Exhaustion was defined by more than five seconds in contact with shock pad, or more than 50% of the time off the treadmill.
Metabolic measurements
Glucose and insulin tolerance testing (GTT and ITT, respectively) was conducted after a five-hour fast. A bolus of glucose (1 g/kg) or insulin (0.75U/kg) was administered via intraperitoneal injection. A blood sample was obtained from the tail tip for the measurement of baseline glucose using a handheld glucometer (Accu-Chek Aviva Plus, Roche or AlphaTrak2 #71681-01) at 0, 15, 30, 45, 60, and 90 min following the injection. At Penn, plasma insulin was measured using Mouse Insulin ELISA kit (Mercodia). Cholesterol and triglycerides were measured using Infinity colorimetric assays (ThermoScientific). Plasma samples at Pfizer were analyzed using Insulin Rodent (Mouse/Rat) Chemiluminescence ELISA kit (ALPCO, #80-INSMR-CH01). HOMA-IR was calculated using the insulin and glucose measurements, using the formula: insulin (µU/ml)*glucose (mmol/L)/22.5. Targeted metabolomics was performed at the Metabolomics Core in the Penn Cardiovascular Institute as previously described, from flash frozen mouse hearts46.
Histology
The tissues were prepared as previously described46 including overnight fixation in 4% paraformaldehyde in PBS and dehydration through sequential ethanol washes. The tissues were then embedded in paraffin and sectioned as 6 mm. Picro Sirius Red staining was performed by the Histology and Gene Expression Co-Op at the University of Pennsylvania and imaged by the Children’s Hospital of Philadelphia Pathology Core. Collagen staining was quantified with Aperio ImageScope (Leica Biosystems).
RNA isolation and qRT-PCR
Total RNA was isolated using the RNeasy Mini Kit (QIAGEN) and the RNase-Free DNase Set (QIAGEN) according to the manufacturer’s instructions. cDNA was synthesized using the Affinity Script cDNA Synthesis Kit (Agilent Technologies) with 0.25 µg total RNA. PCR reactions were performed using Brilliant III Ultra-Fast SYBR Green QPCR Master Mix (Agilent Technologies) on a QuantStudio 6 Flex Real-Time PCR System (Applied Biosystems) with specific primers for each gene. The Penn primer sets are listed in Supplementary Table S1. The expression of target mRNAs was normalized by that of Rplp0 (36B4). Pfizer cardiac gene expression was measured by RT-PCR with TaqMan probes (Myh6: Mm00440359_m1, Myh7: Mm00600555_m1) normalized to Hprt (Mm03024075_m1). All experiments were repeated independently, with samples in either triplicate or quadruplicate.
RNA-seq library preparation, sequencing, and analysis
RNA library preparations and sequencing reactions were conducted at GENEWIZ, LLC. (South Plainfield, NJ), per their standard operating procedure and as previously described47,48. We used Salmon v1.10.2 to count reads in GRCm38 (mm10) reference transcripts49. The counts were summarized at gene level and differentially expressed genes were identified with DESeq2 1.40.250. Genes with an FDR below 0.05, when comparing conditions of interest, and at least a 1.2-fold expression change (up or down) were considered differentially expressed. We performed pathway enrichment analysis, using gProfiler web service with FDR < 0.05 cutoff and enrichment at least 251,52.
Statistics
For two-group comparisons, a Student’s t test was performed when the data were normally distributed based on a Shapiro-Wilk test (α = 0.05). For multiple comparisons, two-way ANOVA with Tukey’s post hoc test was performed. GraphPad Prism 7.04 or 8.03 (GraphPad Software) was used for graphing and statistical analysis.
Study approval
All animal experiments were performed in accordance with NIH and ARRIVE guidelines for the humane treatment of animals and approved by the Institutional Animal Care and Use Committees of the Perelman School of Medicine at the University of Pennsylvania and Pfizer.
Data availability
All data have been deposited in NCBI’s Gene Expression Omnibus Series accession number GSE269053.
References
Omote, K., Verbrugge, F. H. & Borlaug, B. A. Heart failure with preserved ejection fraction: mechanisms and treatment strategies. Annu. Rev. Med. 73, 321–337. https://doi.org/10.1146/annurev-med-042220-022745 (2022).
Roh, J., Hill, J. A., Singh, A., Valero-Munoz, M. & Sam, F. Heart failure with preserved ejection fraction: heterogeneous syndrome, diverse preclinical models. Circ. Res. 130, 1906–1925. https://doi.org/10.1161/CIRCRESAHA.122.320257 (2022).
Valero-Munoz, M., Backman, W. & Sam, F. Murine models of heart failure with preserved ejection fraction: a fishing expedition. JACC Basic Transl. Sci. 2, 770–789. https://doi.org/10.1016/j.jacbts.2017.07.013 (2017).
Smith, A. N. et al. Genomic, proteomic, and metabolic comparisons of small animal models of heart failure with preserved ejection fraction: a tale of mice, rats, and cats. J. Am. Heart Assoc. 11, e026071. https://doi.org/10.1161/JAHA.122.026071 (2022).
Withaar, C., Lam, C. S. P., Schiattarella, G. G., de Boer, R. A. & Meems, L. M. G. Heart failure with preserved ejection fraction in humans and mice: embracing clinical complexity in mouse models. Eur. Heart J. 42, 4420–4430. https://doi.org/10.1093/eurheartj/ehab389 (2021).
Deng, Y. et al. Targeting mitochondria-inflammation circuit by beta-hydroxybutyrate mitigates HFpEF. Circ. Res. 128, 232–245. https://doi.org/10.1161/CIRCRESAHA.120.317933 (2021).
Ranjbarvaziri, S. et al. Targeting HDAC6 to treat heart failure with preserved ejection fraction in mice. Nat. Commun. 15, 1352. https://doi.org/10.1038/s41467-024-45440-7 (2024).
Li, B. et al. Statins improve cardiac endothelial function to prevent heart failure with preserved ejection fraction through upregulating circRNA-RBCK1. Nat. Commun. 15, 2953. https://doi.org/10.1038/s41467-024-47327-z (2024).
Matsiukevich, D. et al. Characterization of a robust mouse model of heart failure with preserved ejection fraction. Am. J. Physiol. Heart Circ. Physiol. https://doi.org/10.1152/ajpheart.00038.2023 (2023).
Schauer, A. et al. ZSF1 rat as animal model for HFpEF: development of reduced diastolic function and skeletal muscle dysfunction. ESC Heart Fail. 7, 2123–2134. https://doi.org/10.1002/ehf2.12915 (2020).
Mira Hernandez, J. et al. Differential sex-dependent susceptibility to diastolic dysfunction and arrhythmia in cardiomyocytes from obese diabetic HFpEF model. Cardiovasc. Res. https://doi.org/10.1093/cvr/cvae070 (2024).
Hegyi, B. et al. Diabetes and excess aldosterone promote heart failure with preserved ejection fraction. J. Am. Heart Assoc. 11, e027164. https://doi.org/10.1161/JAHA.122.027164 (2022).
Gao, C. et al. Glucagon receptor antagonist for heart failure with preserved ejection fraction. Circ. Res. 135, 614–628. https://doi.org/10.1161/CIRCRESAHA.124.324706 (2024).
Schiattarella, G. G. et al. Nitrosative stress drives heart failure with preserved ejection fraction. Nature 568, 351–356. https://doi.org/10.1038/s41586-019-1100-z (2019).
LaPenna, K. B. et al. Combination sodium nitrite and hydralazine therapy attenuates heart failure with preserved ejection fraction severity in a 2-Hit murine model. J. Am. Heart Assoc. 12, e028480. https://doi.org/10.1161/JAHA.122.028480 (2023).
Li, Y. et al. Combining three independent pathological stressors induces a heart failure with preserved ejection fraction phenotype. Am. J. Physiol. Heart Circ. Physiol. 324, H443–H460. https://doi.org/10.1152/ajpheart.00594.2022 (2023).
Cohen, J. B. et al. Clinical phenogroups in heart failure with preserved ejection fraction: detailed phenotypes, prognosis, and response to spironolactone. JACC Heart Fail. 8, 172–184. https://doi.org/10.1016/j.jchf.2019.09.009 (2020).
Shah, S. J. et al. Phenomapping for novel classification of heart failure with preserved ejection fraction. Circulation 131, 269–279. https://doi.org/10.1161/CIRCULATIONAHA.114.010637 (2015).
Hahn, V. S. et al. Myocardial gene expression signatures in human heart failure with preserved ejection fraction. Circulation 143, 120–134. https://doi.org/10.1161/CIRCULATIONAHA.120.050498 (2021).
Harlan, S. M. et al. Viral transduction of renin rapidly establishes persistent hypertension in diverse murine strains. Am. J. Physiol. Regul. Integr. Comp. Physiol. 309, R467–474. https://doi.org/10.1152/ajpregu.00106.2015 (2015).
Sarma, S. et al. Challenging the hemodynamic hypothesis in heart failure with preserved ejection fraction: is exercise capacity limited by elevated pulmonary capillary wedge pressure? Circulation 147, 378–387. https://doi.org/10.1161/CIRCULATIONAHA.122.061828 (2023).
Reddy, Y. N. V., Olson, T. P., Obokata, M., Melenovsky, V. & Borlaug, B. A. Hemodynamic correlates and diagnostic role of cardiopulmonary exercise testing in heart failure with preserved ejection fraction. JACC Heart Fail. 6, 665–675. https://doi.org/10.1016/j.jchf.2018.03.003 (2018).
Beale, A. L. et al. Sex differences in heart failure with preserved ejection fraction pathophysiology: a detailed invasive hemodynamic and echocardiographic analysis. JACC Heart Fail. 7, 239–249. https://doi.org/10.1016/j.jchf.2019.01.004 (2019).
Casimiro, I., Stull, N. D., Tersey, S. A. & Mirmira, R. G. Phenotypic sexual dimorphism in response to dietary fat manipulation in C57BL/6J mice. J. Diabetes Complications 35, 107795. https://doi.org/10.1016/j.jdiacomp.2020.107795 (2021).
Oraha, J., Enriquez, R. F., Herzog, H. & Lee, N. J. Sex-specific changes in metabolism during the transition from chow to high-fat diet feeding are abolished in response to dieting in C57BL/6J mice. Int. J. Obes. (Lond.) 46, 1749–1758. https://doi.org/10.1038/s41366-022-01174-4 (2022).
Gelineau, R. R. et al. The behavioral and physiological effects of high-fat diet and alcohol consumption: sex differences in C57BL6/J mice. Brain Behav. 7, e00708. https://doi.org/10.1002/brb3.708 (2017).
Anker, S. D., Usman, M. S. & Butler, J. SGLT2 inhibitors: from antihyperglycemic agents to all-around heart failure therapy. Circulation 146, 299–302. https://doi.org/10.1161/CIRCULATIONAHA.122.060348 (2022).
Pitt, B. et al. Spironolactone for heart failure with preserved ejection fraction. N. Engl. J. Med. 370, 1383–1392. https://doi.org/10.1056/NEJMoa1313731 (2014).
Solomon, S. D. et al. Angiotensin-neprilysin inhibition in heart failure with preserved ejection fraction. N. Engl. J. Med. 381, 1609–1620. https://doi.org/10.1056/NEJMoa1908655 (2019).
Yusuf, S. et al. Effects of Candesartan in patients with chronic heart failure and preserved left-ventricular ejection fraction: the CHARM-Preserved trial. Lancet 362, 777–781. https://doi.org/10.1016/S0140-6736(03)14285-7 (2003).
Schnelle, M. et al. Echocardiographic evaluation of diastolic function in mouse models of heart disease. J. Mol. Cell. Cardiol. 114, 20–28. https://doi.org/10.1016/j.yjmcc.2017.10.006 (2018).
Kass, D. A. Assessing and interpreting diastolic function in animal models of heart disease. J. Mol. Cell. Cardiol. 197, 1–4. https://doi.org/10.1016/j.yjmcc.2024.10.001 (2024).
Tong, D. et al. Female sex is protective in a preclinical model of heart failure with preserved ejection fraction. Circulation 140, 1769–1771. https://doi.org/10.1161/CIRCULATIONAHA.119.042267 (2019).
Valero-Munoz, M. et al. Proteomic and phosphoproteomic profiling in heart failure with preserved ejection fraction (HFpEF). Front. Cardiovasc. Med. 9, 966968. https://doi.org/10.3389/fcvm.2022.966968 (2022).
van Nieuwenhoven, F. A. et al. Cartilage intermediate layer protein 1 (CILP1): a novel mediator of cardiac extracellular matrix remodelling. Sci. Rep. 7, 16042. https://doi.org/10.1038/s41598-017-16201-y (2017).
Park, S., Ranjbarvaziri, S., Zhao, P. & Ardehali, R. Cardiac Fibrosis is associated with decreased circulating levels of full-length CILP in heart failure. JACC Basic Transl. Sci. 5, 432–443. https://doi.org/10.1016/j.jacbts.2020.01.016 (2020).
Gross, S. & Thum, T. TGF-beta inhibitor CILP as a novel biomarker for cardiac fibrosis. JACC Basic Transl. Sci. 5, 444–446. https://doi.org/10.1016/j.jacbts.2020.03.013 (2020).
Rypdal, K. B. et al. The extracellular matrix glycoprotein ADAMTSL2 is increased in heart failure and inhibits TGFbeta signalling in cardiac fibroblasts. Sci. Rep. 11, 19757. https://doi.org/10.1038/s41598-021-99032-2 (2021).
Hahn, V. S. et al. Myocardial metabolomics of human heart failure with preserved ejection fraction. Circulation 147, 1147–1161. https://doi.org/10.1161/CIRCULATIONAHA.122.061846 (2023).
Anker, S. D. et al. Empagliflozin in heart failure with a preserved ejection fraction. N. Engl. J. Med. 385, 1451–1461. https://doi.org/10.1056/NEJMoa2107038 (2021).
Solomon, S. D. et al. Dapagliflozin in heart failure with mildly reduced or preserved ejection fraction. N. Engl. J. Med. 387, 1089–1098. https://doi.org/10.1056/NEJMoa2206286 (2022).
Withaar, C. et al. The effects of liraglutide and dapagliflozin on cardiac function and structure in a multi-hit mouse model of heart failure with preserved ejection fraction. Cardiovasc. Res. 117, 2108–2124. https://doi.org/10.1093/cvr/cvaa256 (2021).
Zhang, X. et al. Dapagliflozin attenuates heart failure with preserved ejection fraction remodeling and dysfunction by elevating beta-hydroxybutyrate-activated citrate synthase. J. Cardiovasc. Pharmacol. 82, 375–388. https://doi.org/10.1097/FJC.0000000000001474 (2023).
Zincarelli, C., Soltys, S., Rengo, G. & Rabinowitz, J. E. Analysis of AAV serotypes 1–9 mediated gene expression and tropism in mice after systemic injection. Mol. Ther. 16, 1073–1080. https://doi.org/10.1038/mt.2008.76 (2008).
Berger, J. H. et al. Sodium-glucose co-transporter 2 inhibitors Act independently of SGLT2 to confer benefit for heart failure with reduced ejection fraction in mice. bioRxiv. https://doi.org/10.1101/2024.04.29.591665 (2024).
Sakamoto, T. et al. A critical role for estrogen-related receptor signaling in cardiac maturation. Circ. Res. 126, 1685–1702. https://doi.org/10.1161/CIRCRESAHA.119.316100 (2020).
Sakamoto, T. et al. The nuclear receptor ERR cooperates with the cardiogenic factor GATA4 to orchestrate cardiomyocyte maturation. Nat. Commun. 13, 1991. https://doi.org/10.1038/s41467-022-29733-3 (2022).
Yamamoto, T. et al. RIP140 deficiency enhances cardiac fuel metabolism and protects mice from heart failure. J. Clin. Investig. 133. https://doi.org/10.1172/JCI162309 (2023).
Patro, R., Duggal, G., Love, M. I., Irizarry, R. A. & Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 14, 417–419. https://doi.org/10.1038/nmeth.4197 (2017).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of Fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. https://doi.org/10.1186/s13059-014-0550-8 (2014).
Kolberg, L. et al. G:profiler-interoperable web service for functional enrichment analysis and gene identifier mapping (2023 update). Nucleic Acids Res. 51, W207–W212. https://doi.org/10.1093/nar/gkad347 (2023).
Kanehisa, M., Furumichi, M., Sato, Y., Kawashima, M. & Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 51, D587–D592. https://doi.org/10.1093/nar/gkac963 (2023).
Acknowledgements
We thank Teresa C. Leone and Ling Lai at Penn, and Federico Damilano at Pfizer for technical assistance. Echocardiography was performed by the Rodent Cardiovascular Phenotyping Core (RRID: SCR_022419) at the University of Pennsylvania, supported by the Penn Cardiovascular Institute and NIH S10OD016393. AAV8 virus was generated by the Penn Vector Core (RRID: SCR_022432). Metabolomics studies were performed by the Metabolomics Core (RRID: SCR_022381) in the Penn Cardiovascular Institute and supported, in part, by NCI P30 CA016520 and NIH P30DK050306. Biorender.com was used for creation of figure schematics.
Funding
Funding provided by NIH/NICHD K12HD043245 and AHA 24CDA1269277 (https://doi.org/10.58275/AHA.24CDA1269277.pc.gr.193568) (JHB); R01 HL151345, R01 HL128349, R01 HL058493, and Pfizer Research Support (DPK).
Author information
Authors and Affiliations
Contributions
JHB, YS, DH-S, RM, BBZ, RRF, and DPK conceived and designed research; JHB, YS, KT, XC, MM, RK, TRM, JP, RT, RC, and JG performed experiments; JHB, YS, TRM, KB, and JG analyzed data; JHB, YS, RRF, and DPK interpreted results of experiments; JHB, KP, RT, and KB prepared figures; JHB, YS, and TRM drafted manuscript; and AK, RRF, and DPK edited and revised manuscript. All authors approved the final version of manuscript.
Corresponding author
Ethics declarations
Competing interests
DPK is a consultant for Pfizer Inc. YS, KT, XC, MM, RK, DH-S, RM, BBZ, RJRF are employees of Pfizer Inc. or were employees at the time the research was conducted. Multiple Pfizer authors own Pfizer stock. The other authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Berger, J.H., Shi, Y., Matsuura, T.R. et al. Two-hit mouse model of heart failure with preserved ejection fraction combining diet-induced obesity and renin-mediated hypertension. Sci Rep 15, 422 (2025). https://doi.org/10.1038/s41598-024-84515-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-84515-9
