
Abstract
BMI1, a constituent of polycomb repressive complex 1, is overexpressed in a variety of cancers, including neuroblastoma, highlighting its potential as a target for cancer therapeutics. Given the pivotal role of BMI1, a number of inhibitors have been synthesized and assessed for therapeutic efficacy across a spectrum of cancers. In our present study, the BMI1 inhibitors PTC-028 and PTC-209 exhibited selective antitumor activity against MYCN-amplified neuroblastoma. Notably, PTC-028, which exhibited toxicity at lower concentrations, triggered apoptosis in neuroblastoma cells and induced G1-phase accumulation, along with reductions in S-phase and G2/M-phase populations, thereby promoting cell cycle arrest. Thorough RNA sequencing analyses revealed that PTC-028 treatment activated the p53 signaling pathway, suggesting it plays a critical role in the mechanism of apoptosis induction. Moreover, PTC-028 treatment led to decreases in levels of anti-apoptotic proteins, including BCL2 and MCL1. Significantly, PTC-028 also exhibited antitumor efficacy in a mouse xenograft model of human neuroblastoma. These results suggest that BMI1 inhibitors, particularly PTC-028, are promising therapeutic agents for the management of aggressive MYCN-amplified neuroblastomas.
Similar content being viewed by others


Introduction
Neuroblastoma is one of the most common malignant tumors occurring in childhood, accounting for approximately 8% of all pediatric cancers1. Neuroblastoma tumors are thought to originate in the embryonal neural crest, with tumors primarily identified in the adrenal glands and sympathetic ganglia2,3. In addition, neuroblastoma exhibits a diverse range of clinical behaviors, from spontaneous regression to progression despite intensive treatments such as surgery, chemotherapy, and radiotherapy2,4. To guide prognosis and treatment strategies, patients with neuroblastoma are classified according to the International Neuroblastoma Risk Group staging system. This system incorporates tumor extent (including metastatic status), imaging-defined risk factors, patient age, and metastatic distribution to assign tumors to one of four pre-treatment stages: L1, L2, M, or MS. The staging classification, along with genomic features such as MYCN amplification, 11q deletion, and ploidy, are integrated to categorize patients into very-low-risk, low-risk, intermediate-risk, or high-risk groups2. The current 5-year overall survival rate for children diagnosed with advanced-stage neuroblastoma is < 50%5, underscoring the urgent need for new therapeutic strategies.
MYCN amplification is strongly associated with tumor progression and poor clinical outcomes in neuroblastoma. Importantly, it serves as an independent prognostic factor, regardless of disease stage or age at diagnosis, and it is used as a key biomarker in neuroblastoma risk stratification2. Amplification of the MYCN gene is observed in approximately 20% of all neuroblastomas and 50% of high-risk cases, and it is a key biomarker of poor prognosis1,2,6. MYCN, a member of the MYC family that also includes C-MYC and L-MYC, is predominantly expressed during embryonic development7. Aberrant MYCN expression during the perinatal period can initiate tumorigenesis, as demonstrated in TH-MYCN transgenic mice, in which MYCN overexpression in neural crest cells drives the formation of embryonal tumors8. In adults, MYCN expression is largely restricted to hematopoietic and neural tissues9,10. MYCN functions as a transcription factor by forming heterodimers with MYC-associated factor X and binding to E-box sequences, thereby regulating the expression of > 15% of all genes in the genome6,11,12. In cancer, MYCN overexpression—driven by gene amplification, enhanced transcription and translation, and increased protein stability—is associated with malignant progression in multiple tumor types13. However, due to its intrinsically disordered structure and complex regulatory mechanisms, MYCN remains a challenging target for direct therapeutic intervention14.
Polycomb repressive complex 1 (PRC1), which consists of various polycomb group proteins, controls gene expression epigenetically at the transcriptional level, primarily through histone H2A Lys119 monoubiquitination15,16. BMI1, which is a key component of PRC1, binds to the E3 ligase RING1B to regulate histone ubiquitination16. BMI1 is overexpressed in a wide range of cancers, including neuroblastoma and cancers of the stomach, ovaries, breast, head and neck, pancreas, and lung, and this overexpression is closely associated with tumor malignancy and patient prognosis17. BMI1 also plays a vital role in alleviating DNA replication stress by suppressing transcription-replication conflicts and common fragile site instability, as well as by promoting homologous recombination repair of damaged DNA18,19.
In neuroblastoma, BMI1 expression is regulated by MYCN, which binds directly to the promoter region of the BMI1 gene to positively control its expression20. Knockdown of BMI1 reportedly inhibits tumorigenicity in neuroblastoma21,22. Therefore, BMI1 represents a promising therapeutic target in the treatment of neuroblastoma. Currently, several BMI1 inhibitors are under development, such as PTC-028, PTC-209, and PTC596, with PTC-028 exerting antitumor effects in endometrial cancer and alveolar rhabdomyosarcoma23,24. However, the effects of these compounds on neuroblastomas have not been examined. This study investigated the effects of BMI1 inhibitors on both MYCN-amplified and non-amplified neuroblastomas and the underlying molecular mechanisms, with a view toward clinical application.
Results
PTC-028 exhibits specific antitumor effects against MYCN-amplified neuroblastoma
To investigate the effect of the BMI1 inhibitors PTC-209 and PTC-028 on neuroblastoma cell survival, WST-8 assays were conducted using four cell lines, including MYCN-amplified (IMR-32, LA–N-5) and MYCN–non-amplified (SHSY5Y, SKNSH) cells. PTC-209 and PTC-028 treatment resulted in a concentration-dependent decrease in the survival of MYCN-amplified cells, whereas the survival of MYCN–non-amplified cells was less affected by the drugs, even at higher concentrations (Fig. 1A,B). Due to the limited cytotoxicity observed in the MYCN–non-amplified group, the 50% inhibitory concentration (IC50) values could not be reliably determined for these cell lines. In contrast, the MYCN-amplified group exhibited low IC50 values, indicative of higher drug sensitivity. Furthermore, PTC-028 demonstrated greater potency than PTC-209, exerting cytotoxic effects at lower concentrations (Fig. 1C). Subsequent analyses thus focused on the anticancer activity of PTC-028, which was cytotoxic at lower concentrations than PTC-209.

Antitumor effects of PTC-209 and PTC-028 against MYCN-amplified or MYCN–non-amplified neuroblastoma. (A) Evaluation of the cytotoxicity of PTC-209 after 48 h of treatment against various neuroblastoma cell lines, as determined using the WST-8 assay. Surviving fraction is shown as a relative value normalized to the drug-free (0 µM) control group. Data are presented as mean ± SE (n = 6). (B) Evaluation of the cytotoxicity of PTC-028 after 48 h of treatment against various neuroblastoma cell lines, as determined using the WST-8 assay. Surviving fraction is shown as a relative value normalized to the drug-free (0 µM) control group. Data are presented as mean ± SE (n = 6). (C) IC50 values of PTC-209 and PTC-028 against MYCN-amplified neuroblastoma cell line.
PTC-028 induces apoptosis and cell cycle arrest in MYCN-amplified neuroblastoma
The expression of BMI1 in four neuroblastoma cell lines was confirmed by Western blotting, which showed expression in all four lines, with or without MYCN amplification (Fig. S1A). Treatment with PTC-028 resulted in a significant decrease in BMI1 protein levels in SK-N-SH, and SH-SY5Y cells after 24 h and in IMR-32 and LA–N-5 cells after 48 h (Fig. 2A–D and S1B, C). IMR-32 and LA–N-5 cells did not exhibit a reduction in BMI1 protein levels after 24 h of treatment with PTC-028 (data not shown).

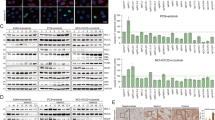
Apoptosis, cell cycle progression, and tubulin polymerization in MYCN-amplified neuroblastoma under PTC-028 treatment. (A) Western blotting analysis of BMI1, PARP, Cl-PARP, Caspase-3, and cleaved (Cl)-Caspase-3 in IMR-32 cells 48 h after initiation of PTC-028 treatment. Uncropped blots are shown in Figure S11. (B) Quantification of BMI1, Cl-PARP, and Cl-Caspase-3 in IMR-32 cells as determined by Western blotting analysis. Protein levels were normalized to that of actin. Data are presented as mean ± SE (n = 3; *p < 0.05, **p < 0.01; Student’s t-test). (C) Western blotting analysis of BMI1, PARP, Cl-PARP, Caspase-3, and Cl-Caspase-3 in LA–N-5 cells 48 h after initiation of PTC-028 treatment. Uncropped blots are shown in Figure S11. (D) Quantification of BMI1, Cl-PARP, and Cl-Caspase-3 in LA–N-5 cells as determined by Western blotting analysis. Protein levels were normalized to that of actin. Data are presented as mean ± SE (n = 3; *p < 0.05, **p < 0.01; Student’s t-test). (E) Percentage of IMR-32 cells in each cell cycle phase 24 h and 48 h after initiation of PTC-028 treatment, as determined by PI staining. Data are presented as mean ± SE (n = 3; *p < 0.05, vs. control; Tukey’s multiple comparison test). (F) Western blotting analysis of α-tubulin in polymerized (p) or soluble (s) fractions of IMR-32 cells 24 h after initiation of treatment with PTC-028 or vincristine (VCR, 100 nM). Uncropped blots are shown in Figure S11. (G) Ratio of the amount of tubulin in the soluble fraction to that in the polymerized fraction (s/p) as determined by Western blotting analysis. Data are presented as mean ± SE (n = 3; *p < 0.05, **p < 0.01 vs. control; Tukey’s multiple comparison test).
Experiments from this point forward were designed to explore the mechanism of cell death in PTC-028–sensitive MYCN-amplified neuroblastoma cells (IMR-32 and LA–N-5). Importantly, PTC-028 also induced an increase in levels of the apoptosis markers cleaved PARP and cleaved caspase-3 in IMR-32 and LA–N-5 cells (Fig. 2A–D). In addition to causing cell death via apoptosis, the possibility that PTC-028 inhibits cell proliferation by inducing cell cycle arrest was investigated using flow cytometry. This analysis revealed dynamic changes in cell cycle distribution over time. At 12 h post-treatment, a reduction in the G1-phase population and a corresponding increase in the M-phase population were observed (Fig. S2A-D). By 24 h, the G1-phase population increased, whereas the S-phase fraction decreased (Fig. 2E and Fig. S3). At 48 h, a similar pattern was observed, with an increase in G1-phase cells, a reduction in S-phase cells, and, notably, a decrease in the G2/M-phase population (Fig. 2E and Fig. S3). A previous study suggested that the antitumor effect of PTC-028 against myelodysplastic syndrome involves inhibition of tubulin polymerization25. The effect of PTC-028 on tubulin polymerization was thus examined in IMR-32 cells using Western blotting. IMR-32 cells were treated with PTC-028 or the tubulin polymerization inhibitor vincristine, after which soluble and insoluble fractions were prepared and analyzed for α-tubulin. Vincristine increased the ratio of non-polymerized (soluble fraction) to polymerized α-tubulin (insoluble fraction), but PTC-028 had no effect on this ratio (Fig. 2F,G).
PTC-028 activates the p53 pathway in MYCN-amplified neuroblastoma
The mechanism of the antitumor effect against MYCN-amplified neuroblastoma was investigated by comprehensively analyzing the expression of 410 genes that showed significant variation in expression in RNA-Seq analysis of IMR-32 cells treated with PTC-028 (false-discovery rate [FDR] < 0.01) (Fig. 3A). Of the 410 genes, 173 were upregulated, including CDKN1A (p21), whereas 237 genes were downregulated (Fig. 3A,B; Tables S1, S2). Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis revealed that PTC-028 treatment upregulated the “p53 signaling” and “Apoptosis” pathways and downregulated the “Cell cycle” pathway (Fig. 3C). Gene Set Enrichment Analysis (GSEA) confirmed these pathway changes (Fig. 3D). A cnet plot of the upregulated genes showed overlap between multiple genes related to p53 signaling and apoptosis (Fig. 3E). Regarding the p53 signaling pathway, genes involved in cell cycle control (e.g., CDK family and upstream of p53 like CHEK1 [CHK1], CHEK2 [CHK2]) were downregulated, whereas genes related to apoptosis and cell cycle arrest (e.g., p21, GADD45A, and apoptosis-related BAX, PAG608, DR5), DNA repair (e.g., p48, p53R2, sestrin family [SESN1-3]), and negative feedback regulators of the p53 pathway (e.g., MDM2, cyclin G, Wip1) were upregulated (Fig. 3F). Further analyses of clinical samples revealed significant downregulation of CDKN1A (p21 gene) expression in MYCN-amplified tumors and high-risk groups (Fig. S4A-D). To validate the RNA-Seq findings, reverse transcription-quantitative polymerase chain reaction (RT-qPCR) analysis was performed for p21 and BAX, both of which showed expression changes consistent with the transcriptomic analysis results (Fig. S5).

Comprehensive RNA-Seq analysis of gene expression changes in MYCN-amplified neuroblastoma treated with PTC-028. (A) Flowchart of RNA-Seq analysis (n = 3). (B) Volcano plot of gene expression changes. Top 30 genes with low q-values are indicated by labels. (C) Dot plot showing the results of KEGG enrichment analysis for genes exhibiting variable expression. The top 10 terms for each up- or downregulated gene are shown. (D) Gene Set Enrichment Analysis of the KEGG terms “p53 signaling pathway”, “Apoptosis”, and “Cell cycle”. (E) Cnet plot of the group of genes associated with the top 10 upregulated terms. (F) Heatmap showing the expression levels of genes associated with the KEGG term “p53 signaling pathway” in each sample. (F) Heatmap showing the expression levels of genes associated with the KEGG term “p53 signaling pathway” in each sample. Expression levels were standardized according to Z score.
PTC-028 induces p53-dependent cell death in MYCN-amplified neuroblastoma
To confirm that the p53 signaling pathway and apoptosis were altered at the protein level, expression changes in p53, p21, and apoptosis-related factors were evaluated using Western blotting (Figs. 4A,B and S6A, B). Protein levels of p53 and p21 increased after PTC-028 treatment, but no significant changes in the apoptosis-promoting factors NOXA and BAX were observed. Protein levels of the apoptosis-inhibiting factors BCL2 and MCL1 decreased after 24-h and 48-h PTC-028 treatment. PTC-028 treatment also reduced the ubiquitination of histone H2A (Lys119), a target of the PRC1 complex to which BMI1 belongs. These results suggested that PTC-028 induces cell death by activating the p53 pathway. Combination treatment of IMR-32 cells with the p53 inhibitor pifithrin-α and PTC-028 significantly attenuated the PTC-028–mediated reduction in cell survival (Fig. 4C). These experiments confirmed that pifithrin-α does not affect BMI1 expression in IMR-32 cells (Fig. S7A). WST-8 assays revealed that TGW MYCN-amplified neuroblastoma cells, which harbor a p53 functional mutation, were significantly less sensitive to PTC-028 compared with IMR-32 cells (Fig. S7B).

Expression of proteins related to the p53 and apoptosis pathways following treatment with PTC-028. (A) Western blotting analysis of p53, p21, NOXA, BAX, BCL2, MCL1, and H2Aub.(K119) in IMR-32 cells 24 or 48 h after initiation of PTC-028 treatment. Uncropped blots are shown in Figure S11. (B) Relative quantitation of each protein as determined by Western blotting analysis. Protein levels were normalized to that of actin. Data are presented as mean ± SE (n = 3; *p < 0.05, **p < 0.01; Student’s t-test). (C) Surviving fraction of IMR-32 cells under treatment with PTC-028 alone or combined PTC-028 and pifithrin-α (PFT-α) for 48 h, as determined using the WST-8 assay. Data are presented as mean ± SE (n = 6; *p < 0.05, **p < 0.01; two-way ANOVA).
PTC-028 induces mild DNA damage in MYCN-amplified neuroblastoma
DNA damage stabilizes p53 protein levels through post-translational modifications26,27. To determine if the PTC-028–mediated stabilization of p53 depends on DNA damage, changes in DNA damage markers were examined in PTC-028–treated LA–N-5 MYCN-amplified neuroblastoma cells using Western blotting and immunofluorescence analysis. Western blotting analysis indicated a trend toward increased phospho-histone H2A.X (γH2AX) protein levels following PTC-028 treatment, although the difference did not reach statistical significance (Fig. 5A–D). In contrast, the number of γH2AX-positive cells observed by immunofluorescence analysis was increased in cells treated with the CHK1 inhibitor LY2606368, a positive control, but not following PTC-028 stimulation (Figs. 5E,F and S8A, B). Similarly, the number of cells exhibiting multiple 53BP1 foci (another marker of DNA damage) increased following treatment with CHK1 inhibitor but not PTC-028 (Figs. 5G,H and S8C, D).

DNA damage in MYCN-amplified neuroblastoma under PTC-028 treatment. (A) Western blotting analysis of γH2AX in IMR-32 cells 24 h after initiation of PTC-028 treatment. Uncropped blots are shown in Figure S11. (B) Western blotting analysis of γH2AX in LA–N-5 cells 48 h after initiation of PTC-028 treatment. Uncropped blots are shown in Figure S11. (C) Relative quantitation of γ-H2AX in IMR-32 cells as determined by Western blotting analysis. Protein levels were normalized to that of actin. Data are presented as mean ± SE (n = 3; Student’s t-test). (D) Relative quantitation of γ-H2AX in LA–N-5 cells as determined by Western blotting analysis. Protein levels were normalized to that of actin. Data are presented as mean ± SE (n = 3; Student’s t-test). (E) Immunofluorescence staining of γH2AX (green) in LA–N-5 cells treated with PTC-028 for 24 h. DAPI = blue. Scale bar = 10 μm. (F) Ratio (%) of γH2AX-positive LA–N-5 cells. Data are presented as mean ± SE (n = 3; 3 fields/well; *p < 0.05, vs. control; Tukey’s multiple comparison test; N.D., not detected). (G) Immunofluorescence staining of 53BP1 (green) in LA–N-5 cells treated with PTC-028 for 24 h. DAPI = blue. Scale bar = 10 μm. (H) Ratio (%) of LA–N-5 cells with ≥ 4. 53BP1 foci. Data are presented as mean ± SE (n = 3; 3 fields/well; *p < 0.05, vs. control; Tukey’s multiple comparison test; N.D., not detected).
PTC-028 exhibits an antitumor effect in vivo
The antitumor effect of PTC-028 in vivo was examined using a mouse xenograft model of human neuroblastoma. LA–N-5 cells were subcutaneously implanted into NOD.Cg-PrkdcscidIl2rgtm1Sug/ShiJic (NOG) mice, and PTC-028 was administered intraperitoneally at a dose of 15 mg/kg after the tumors reached approximately 150 mm3 in size (Fig. 6A). Significant reductions in tumor volume and weight were observed in the PTC-028–treated group compared with the control (Fig. 6B–D). Excluding the tumor, no change in body weight was observed, with or without PTC-028 treatment (Fig. 6E). Finally, tumors from PTC-028–treated mice exhibited a decrease in the proportion of cells positive for Ki67, a marker of cell proliferation (Fig. S9).

Antitumor effects of PTC-028 on human tumor xenograft models. (A) Protocol for analysis of the effect of PTC-028 treatment on human tumor xenograft models. (B) Representative photographs of mice and tumors 14 days after the commencement of treatment. Photographs of all of the mice before tumor dissection and photographs of all of the dissected tumors are shown in Figure S12. The raw measurements of the tumors for every mouse are shown in Table S6. (C) Tumor volume of the control group and PTC-028 group monitored for 14 days. Data are presented as mean ± SE (n = 6; *p < 0.05, **p < 0.01; two-way ANOVA). (D) Weight of excised tumor in each group after 14 days of treatment. Data are presented as mean ± SE (n = 6; *p < 0.05, **p < 0.01; Student’s t-test). (E) Body weight of mice after tumor excision at 14 days after the commencement of treatment (n = 6; *p < 0.05, **p < 0.01; Student’s t-test).
Discussion
In this study, we found that the BMI1 inhibitors PTC-028 and PTC-209 exert selective antitumor activity against MYCN-amplified neuroblastoma. The induction of apoptosis is a key step in the antitumor mechanism of PTC-028. This process appears to involve reduced expression of the anti-apoptotic factors BCL2 and MCL1, which suppress the function of BAX. BAX is crucial for mediating the release of cytochrome c from mitochondria, relative to its expression level. Another component of the antitumor mechanism involves the accumulation of cells in the G1 phase, accompanied by decreases in both S-phase and G2/M-phase populations, consistent with cell cycle inhibition. p21 halts progression of the cell cycle in S phase by directly binding to PCNA and inhibiting DNA replication, in addition to suppressing the CDK2–cyclin E complex28,29. In the context of the present study, PTC-028 significantly enhanced p21 mRNA and protein expression in neuroblastoma cells, indicating the induction of cell cycle arrest via p21 activation. These findings suggest the p53 pathway plays a significant role in the antitumor activity. PTC-028 also exhibited antitumor efficacy in a mouse xenograft model. Thus, PTC-028 is potentially effective not only against cultured neuroblastoma cells but also tumors arising in vivo.
Interestingly, our findings indicate that the BMI1 inhibitors PTC-028 and PTC-209 specifically target MYCN-amplified neuroblastoma cells. In addition, PTC-028 triggered apoptosis and cell cycle arrest in MYCN-amplified neuroblastoma cells, as evidenced by elevated levels of the cell death (including apoptosis) markers cleaved PARP and cleaved caspase-3. The observation that BMI1 knockdown leads to apoptosis in neuroblastoma cells22 suggests that PTC-028 induces apoptosis via a reduction in BMI1 protein expression. Notably, PTC-028 reportedly induces apoptosis in ovarian cancer cells by causing a transient decrease in ATP and imbalance in mitochondrial redox following BMI1 downregulation23. In other contexts, PTC-028 suppresses the growth of myelodysplastic syndrome cells and induces their apoptosis by blocking microtubule polymerization25. As both microtubule destabilization and BMI1 inhibition have been proposed as potential mechanisms underlying the cytotoxicity of PTC-028, determining which pathway predominates in neuroblastoma is of particular importance. Therefore, we directly evaluated the effect of PTC-028 on tubulin polymerization in neuroblastoma cells. Vincristine served as a positive control and successfully disrupted microtubule polymerization, whereas PTC-028 had no observable effect, even at concentrations sufficient to induce apoptosis (25 nM and 2.5 μM). These results suggest that the pro-apoptotic activity of PTC-028 in neuroblastoma does not involve microtubule destabilization but is instead mediated via alternative pathways, such as BMI1 downregulation, p53 activation, or cell cycle arrest.
RNA-Seq analysis was conducted to assess the antitumor efficacy of PTC-028 and revealed that PTC-028 treatment activates the p53 pathway and related apoptotic pathways. Western blotting analysis confirmed an increase in p53 and p21 protein levels. However, p53 mRNA expression remained unchanged, indicating a post-translational regulation mechanism. Similarly, increased p21 mRNA levels were observed, suggesting enhanced transcription facilitated by activation of the p53 pathway. We hypothesized that activation of the p53 pathway plays a role in the antitumor mechanism of PTC-028. To test this hypothesis, we examined the combined use of PTC-028 with the p53 inhibitor pifithrin-α. These experiments revealed that pifithrin-α partially restored the decrease in the viability of IMR-32 cells induced by PTC-028. Additionally, pifithrin-α treatment did not alter BMI1 expression, and no changes in the viability of cells treated with pifithrin-α alone were noted visually. Furthermore, compared with IMR-32 cells, PTC-028 exhibited a weaker anti-tumor effect against TGW cells, which harbor p53 functional mutations. These findings suggest that the anti-tumor effect of PTC-028 is partially suppressed by the inhibition of p53 function. Taken together, these results indicate that the anti-tumor effect of PTC-028 is mediated in part by p53 activation, but other mechanisms are also likely involved.
It is important to consider alternative mechanisms that contribute to cellular regulation and response, beyond activation of the p53 pathway. Significantly, treatment with PTC-028 decreased protein expression levels of the anti-apoptotic factors MCL1 and BCL2. Importantly, BMI1 was shown to enhance MCL1 protein expression via the BMI1-HUWE1-MCL1 axis in non-small cell lung cancer30. MCL1 is overexpressed in various malignant neoplasms31,32, suggesting that BMI1-mediated modulation of MCL1 expression could also play a critical role in the pathophysiology of neuroblastoma cells. Furthermore, the PTC-028–mediated reduction in BCL2 expression in neuroblastoma cells is consistent with the antitumor effects of the BCL2 inhibitors ABT263 and ABT199 against neuroblastoma cells33,34. Other research showed that downregulation of MCL1 enhances the sensitivity of neuroblastoma cells to BCL2-family antagonists35. Another recent study reported that cisplatin-resistant neuroblastoma is characterized by a high reliance on MCL1 and diminished sensitivity to BCL2/BCL-XL inhibitors36. Furthermore, pharmacologic co-inhibition of BCL2/MCL1 has an antitumor effect in neuroblastoma cells37. Thus, concurrent downregulation of BCL2 and MCL1 using BMI1 inhibitors could induce more potent cell death in high-grade neuroblastoma. Given that both activation of the p53 pathway and inhibition of BCL2 and MCL1 expression reportedly play a role in synthetic lethality38, PTC-028 might activate a mechanism similar to synthetic lethality by simultaneously activating the p53 pathway and reducing BCL2 and MCL1 protein levels. This observation suggests that the inability of p53 inhibitors to fully reverse PTC-028–induced apoptosis derives from their capacity to block p53 activation but not counteract reductions in BCL2 and MCL1 levels. Activation of the p53 pathway thus plays a crucial role in apoptosis triggered by PTC-028.
Despite the observed significant increase in p53 protein levels following PTC-028 treatment, the underlying mechanism of this activation remains unclear and warrants further investigation. BMI1 was previously implicated in DNA repair mechanisms19, and p53 is known to enhance the stability and transcriptional activity of BMI1 in response to DNA damage through post-translational modifications such as increased acetylation or reduced ubiquitination26,27. To determine whether p53 activation reflects a secondary response to DNA damage, we examined the DNA damage response markers γH2AX and 53BP1. Western blot analysis revealed a trend toward increased γH2AX levels in IMR-32 and LA–N-5 cells following PTC-028 treatment, suggesting that BMI1 inhibition may induce mild DNA damage. However, γH2AX and 53BP1 signals were not detectable via immunofluorescence analysis in LA–N-5 cells, suggesting that the level of DNA damage induced by PTC-028 is so minor that it falls below the detection threshold of the immunofluorescence assay, which is less sensitive than Western blotting. Moreover, transcriptomic analysis revealed increased expression of genes in the p53 and apoptosis pathways following PTC-028 treatment, which is consistent with protein-level evidence of p53 stabilization. Although this may suggest activation of the DNA damage response, it is also plausible that these changes are a downstream consequence of direct p53 stabilization due to BMI1 inhibition, rather than a response to actual genotoxic stress. Interestingly, previous studies have reported both the presence and absence of γH2AX accumulation following BMI1 knockdown19,22, underscoring the complex and potentially context-dependent relationship between BMI1 and DNA damage response mechanisms. Thus, our data indicate the effects of PTC-028 involve mild DNA damage and activation of p53-related pathways; however, the causal relationship remains unresolved—specifically, whether the upregulation of genes typically responsive to DNA damage reflects actual DNA damage resulting from impaired DNA repair due to BMI1 inhibition, leading to p53 stabilization, or whether it instead reflects direct p53 reactivation due to the relief of BMI1-mediated suppression. Further studies employing time-course analyses, DNA damage response inhibitors, or p53-deficient models will be essential to clarify this mechanistic ambiguity.
Another potential mechanism for the upregulation of p53 involves direct regulation by BMI1. This pathway is of particular interest in the context of neuroblastoma, in which the majority of cases retain wild-type p53, suggesting an alternative mechanism of p53 functional suppression. Notably, p53 transcription is reportedly increased by MYCN in MYCN-amplified neuroblastoma but repressed at the protein level via the MDM2 pathway and other factors39. The inhibition of p53 ubiquitination by MDM2, using MDM2-p53 antagonists such as Nutlin-3 and MI-63, renders MYCN-amplified neuroblastoma more susceptible to apoptosis than MYCN–non-amplified neuroblastoma. MYCN siRNA knockdown and mutation of p53 attenuated the effect of the drugs, suggesting that activation of the p53 pathway, which is suppressed in MYCN-amplified neuroblastoma, efficiently induces apoptosis39. In contrast, the significance of p53 pathway activation may not be as pronounced in MYCN–non-amplified neuroblastoma. In neuroblastoma cells, BMI1 reportedly ubiquitinates p53 and promotes its degradation by binding to RING1A and RING1B40. Thus, ubiquitination of p53 by BMI1 in MYCN-amplified neuroblastoma may play an important role in attenuating p53 protein function and stability. Therefore, the antitumor effect of PTC-028 in MYCN-amplified neuroblastoma may involve BMI1 degradation and de-repression of the p53 pathway, along with reductions in BCL2 and MCL1 expression.
This study has several limitations. First, although treatment with the BMI1 inhibitor demonstrated antitumor effects, the impact on normal tissues has not been thoroughly examined. Consistent with previous reports23, administration of PTC-028 in mice did not result in weight loss or other signs of toxicity, suggesting the absence of lethal effects. Similarly, in our present mouse xenograft model experiment, no change in body weight (excluding the tumor) was observed, with or without treatment. Indeed, PTC-028 has been shown to have limited effects on normal cells, such as ovarian epithelial cells or fallopian tube epithelial cells23. In that study, the non-toxicity of PTC-028 was attributed to the low BMI1 expression in normal cells. However, when BMI1 is overexpressed in immortalized fallopian tube epithelial cell lines, such as FTE188, PTC-028 becomes toxic. BMI1 is expressed in a wide range of tissues, including the brain, kidney, gastrointestinal tract, placenta, thymus, parathyroid, salivary glands, gonads, and bone marrow41. BMI1 plays a crucial role in the maintenance and proliferation of stem cells, extending beyond oncogenic contexts42. Therefore, assessing the potential cytotoxicity of PTC-028 against both cancerous and non-cancerous cells and tissues exhibiting high BMI1 expression is critical. Furthermore, both long-term and short-term toxicity must be evaluated. This assessment is critical for understanding the broader implications of targeting BMI1 using therapeutic agents such as PTC-028 and ensuring the specificity and safety of such interventions in clinical settings.
Second, the precise mechanism by which PTC-028 exerts antitumor effects in neuroblastoma xenograft models remains unclear. We attempted to assess BMI1 protein levels in xenograft tumors using immunohistochemistry analysis; however, reproducible and quantifiable results could not be obtained due to technical limitations. This in turn limited our ability to directly compare our findings with those of a previous report from a study of ovarian cancer, in which PTC-028 was shown to reduce BMI1 expression in vivo23. Based on our in vitro observations, a reduction in BMI1 protein expression in vivo would be expected to coincide with activation of the p53 pathway and downregulation of anti-apoptotic proteins such as MCL1 and BCL2, ultimately leading to apoptosis. As we were unable to confirm this using the present in vivo model, further investigation will be necessary to clarify the relevance of BMI1 modulation in neuroblastoma tumor biology.
Third, the present study was unable to clarify why PTC-028 exhibited limited efficacy against MYCN–non-amplified neuroblastoma. As BMI1 expression is upregulated by MYCN, it was hypothesized that the survival of MYCN–non-amplified neuroblastoma cells, which should exhibit low BMI1 expression, does not depend on BMI1 or p53 suppression via ubiquitination, thereby leading to reduced drug efficacy. We therefore examined whether BMI1 expression is regulated by MYCN by performing siRNA-mediated knockdown of MYCN. However, the results were inconsistent and not sufficiently robust to be included in this study. Given these limitations, we next assessed endogenous MYCN and BMI1 expression levels across the four neuroblastoma cell lines. BMI1 expression was detected in all four neuroblastoma cell lines, with particularly high levels observed in both the MYCN-amplified LA–N-5 and the MYCN–non-amplified SH-SY5Y lines, indicating no clear correlation between MYCN amplification and BMI1 expression level. These findings suggest that BMI1 expression does not significantly affect drug sensitivity, at least in the cell lines examined in this study. These findings also suggest that BMI1 contributes to suppression of the p53 pathway in both MYCN-amplified and non-amplified cell lines and that PTC-028 treatment may activate the p53 pathway. However, minimal death of MYCN–non-amplified neuroblastoma cell lines was observed at low concentrations of PTC-028. This observation suggests that MYCN–non-amplified neuroblastoma cells are resistant to p53 activation. According to previous reports, neuroblastoma cell lines that express low levels of MYCN exhibit increased p53 protein expression upon treatment with MDM2-p53 antagonists, but apoptosis is only mildly induced compared with MYCN-amplified neuroblastoma cells39. These findings may explain the weak anti-tumor effect of PTC-028 against MYCN–non-amplified neuroblastoma, which appears less susceptible to p53 activation. Another point worth noting is that, when comparing SK-N-SH and SH-SY5Y, both of which are MYCN–non-amplified cell lines, SK-N-SH cells exhibit somewhat greater sensitivity to PTC-028 and PTC-209. Although the underlying cause of this difference remains unclear, it may be attributed to a greater resistance of SH-SY5Y cells to p53 activation. Future studies should focus on elucidating the mechanism by which MYCN amplification affects sensitivity to BMI1 inhibitors, as this will provide valuable insights for the development of new neuroblastoma therapies.
Fourth, several experiments presented technical challenges, resulting in ambiguous conclusions. For instance, although we attempted immunostaining for γH2AX and p53BP1 in IMR-32 cells, similar to LA–N-5 cells, this was not feasible due to the tendency of IMR-32 cells to detach during the procedure. Additionally, efforts to establish a xenograft model by transplanting IMR-32 cells into NOG mice, as was successfully performed with LA–N-5 cells, were unsuccessful because the tumors failed to engraft. We also attempted to investigate the localization and protein levels of BMI1 and p53 in tumor tissues via immunohistochemistry. However, we were unable to obtain clear data due to poor staining results. This may be attributable to technical issues, reagent quality, or inherently low expression levels, but the exact cause remains undetermined. Therefore, it is important to acknowledge that the absence of results from these experiments contributes to the limitations and ambiguity in the interpretation of this study’s conclusions.
Fifth, although functional outcomes of PTC-028 treatment, including gene expression changes and apoptosis, were clearly observed within 24 h, a measurable reduction in BMI1 protein levels was only detected after 48 h. Given that PTC-028 induces degradation of BMI1 via hyperphosphorylation, it is possible that functional inhibition precedes detectable protein loss. However, this hypothesis remains speculative, as no direct supporting evidence was obtained in the present study. Further investigation will be necessary to clarify the temporal dynamics and mechanism of BMI1 inhibition by PTC-028.
In conclusion, our findings suggest that the BMI1 inhibitor PTC-028 provides therapeutic benefits in high-risk MYCN-amplified neuroblastoma by activating the p53 pathway. This suggests that BMI1 inhibitors such as PTC-028 merit consideration as a potentially effective new therapy for neuroblastoma.
Methods
Cell culture
Human neuroblastoma cell lines (IMR-32 [JCRB Cell Bank, catalog no. JCRB9050, TGW [JCRB Cell Bank, catalog no. JCRB0618], LA–N-5 [RIKEN Cell Bank, catalog no. RCB0485], SK-N-SH [RIKEN Cell Bank, catalog no. RCB0426], and SH-SY5Y [ATCC, catalog no. CRL-2266]) were cultured in RPMI-1640 medium (Wako, catalog no. 183–02,023) containing 10% fetal bovine serum (FBS), 100 U/ml penicillin, and 100 mg/ml streptomycin at 37 °C in a 5% CO2 environment. The MYCN amplification status of each cell line was consistent with that described in previous reports: IMR-32 and LA–N-5 are MYCN-amplified, whereas SK-N-SH and SH-SY5Y are MYCN–non-amplified43,44,45. All cell lines were obtained from reputable public repositories (JCRB, RIKEN, or ATCC), and their identities were confirmed based on catalog information and established profiles. All experiments were performed using cells at low to moderate passage numbers (≤ 15) to ensure consistency and minimize variations due to culture-induced changes.
Reagents
BMI1 inhibitors used included PTC-028 (C19H12F5N5, Selleckchem, catalog no. S8662) and PTC-209 (C17H13Br2N5OS, Cayman Chemical, catalog no. 16277); structural information for these compounds, along with information for PTC596, is provided in Figure S10. Vincristine sulfate (C46H56N4O10・H2SO4, Cayman Chemical, catalog no. 11764) was used as a tubulin polymerization inhibitor, and pifithrin-α (C16H18N2OS・HBr, Cayman Chemical, catalog no. 13326) was used to inhibit p53. For assays, these compounds were dissolved in DMSO.
Cell viability assay
Cell proliferation was assessed using a WST-8 cell proliferation assay (Cell Counting Kit-8; Dojindo Laboratories, catalog no. 341–08,001). Cells (IMR-32, LA–N-5, SK-N-SH, SH-SY-5Y, TGW) were detached using 0.25% (w/v) trypsin–1 mM EDTA・4Na solution (Fujifilm Wako Pure Chemical Corp., catalog no. 209–16,941) or TrypLE™ Express Enzyme (1 ×) without phenol red (Gibco, catalog no. 12604013) diluted in RPMI-1640 (+ FBS, + penicillin, + streptomycin). The cells were then seeded in a 96-well plate (Corning, catalog no. 3599) at specific densities. After 24 h, PTC-028 or PTC-209, both diluted in RPMI-1640 (+ FBS, + penicillin, + streptomycin), was added to each well and incubated for 48 h. In some experiments, cells were also treated with pifithrin-α (10 µM) for 48 h. Post-treatment, Cell Counting Kit-8 solution diluted in RPMI-1640 (+ penicillin, + streptomycin) was added, and the plate was incubated at 37 °C for 1 h. Absorbance at 450 nm and 690 nm was measured using a multi-function plate reader (Nivo 3S; Perkin Elmer).
Western blotting analysis
IMR-32 cells were seeded in a 6-well plate (Falcon) at 3.2 × 105 cells/well, incubated for 24 h, and then treated with DMSO or PTC-028 (25 nM). Cells were harvested using 1 × SDS sample buffer (62.5 mM Tris–HCl [pH 6.8], 2% SDS, and 10% glycerol) and sonicated to reduce viscosity, followed by a 3-min incubation at 95 °C. Protein concentration was measured using the Pierce BCA Protein Assay kit (Thermo Scientific, catalog no. 23225) and equalized using 1 × SDS sample buffer with bromophenol blue and 5% β-mercaptoethanol. SDS-PAGE was performed, followed by semi-dry transfer onto a membrane in transfer buffer for 15 min at 40 V. A single full-length membrane was physically cut horizontally before antibody incubation to enable separate probing of the upper and lower molecular weight regions using different primary antibodies. The membrane was blocked with 5% skim milk and incubated at room temperature for 1 h. After washing with Tris-buffered saline with Tween-20 (TBS-T), the membrane was incubated with primary antibodies overnight at 4 °C. The membrane was then washed with TBS-T and incubated for 1 h at room temperature with secondary antibodies (anti-mouse IgG, HRP-conjugated; anti-rabbit IgG, HRP-conjugated, both from Cell Signaling Technology, catalog no. 7076) diluted 1:10,000 in TBS-T. After washing, proteins were detected using ECL Prime (GE Healthcare, catalog no. RPN2236) or West Femto (Thermo Scientific, catalog no. 34094) and imaged using an Amersham Imager 600 (GE Healthcare Life Sciences). Table S3 lists the primary antibodies used in this study. The antibody diluent was prepared by adding 0.3 g of bovine serum albumin (BSA) and 100 µl of 10% NaN3 to 10 ml of TBS-T. Uncropped blots are shown in Figure S11.
Detection of polymerized and non-polymerized tubulin
The protocol followed the report by Zhong et al.25. IMR-32 cells were seeded in a 6-well plate (Corning, catalog no. 353046) at a density of 3.2 × 105 cells/well and cultured for 24 h, followed by treatment with DMSO, PTC-028 (25 nM, 2.5 µM), or vincristine (100 nM) for 24 h. Cells were then collected using permeabilization buffer (200 µl; 80 mM PIPES–NaOH [pH 6.8], 1 mM MgCl2, 1 mM EGTA, 0.2% Triton X-100, 10% glycerol, and 1 × protease inhibitor cocktail) and incubated at 30 °C for 5 min. A small aliquot of each sample was mixed with 2 × SDS sample buffer, and proteins were quantified using the Pierce BCA Protein Assay kit (Thermo Scientific, catalog no. 23225). After equalizing protein concentrations, the samples were centrifuged to separate the soluble fraction (supernatant) and insoluble fraction (pellet). The supernatant was mixed with 4 × SDS sample buffer, and the pellet was dissolved in 1 × SDS sample buffer. Both were then boiled at 95 °C for 3 min. α-Tubulin levels in these samples were determined by Western blotting analysis.
Flow cytometry
IMR-32 cells were seeded in a 6-well plate (Corning, catalog no. 353046) at 5.0 × 105 cells/well and cultured for 24 h, then treated with DMSO or PTC-028 (25 nM) for an additional 24 h at 37 °C with 5% CO2. The cells were detached using TrypLE™ Express Enzyme and centrifuged to obtain 1.0 × 106 cells. For cell cycle analysis, pelleted cells were fixed overnight at − 20 °C in ice-cold 70% ethanol, washed with PBS, and then treated with FxCycle™ PI/RNase Staining Solution (Invitrogen, catalog no. F10797). Cells were incubated in the dark for 30 min and then analyzed using a FACS Lyric™ (BD Biosciences). Cell cycle analysis was performed using the cell cycle platform in FlowJo™ software (BD Biosciences). Debris was excluded by gating based on forward scatter (FSC) and side scatter parameters, followed by singlet discrimination using FSC-W versus FSC-H to remove doublets with high FSC-W signals. The analysis was conducted on singlet populations only. Cell cycle distribution was calculated using the Dean-Jett-Fox model46, with “CC Gates in Workspace” enabled to quantify the proportion of cells in each phase as displayed in the workspace.
RNA-Seq
IMR-32 cells were seeded in a 6-well plate (Corning, catalog no. 353046) at 2.0 × 105 cells/well and cultured for 24 h at 37 °C with 5% CO2. The cells were then treated with DMSO or PTC-028 (25 nM) for another 24 h, and total RNA was extracted using an RNeasy Mini Kit (QIAGEN, catalog no. 74104). Paired-end Illumina sequencing was performed by Novogene. Quantification of FASTQ files, quality control metrics, and differential expression analysis were conducted using RaNA-seq47 via its graphical user interface (GUI). The reference genome Human, Homo sapiens (GRCh38) was selected, and differential expression analysis was performed using DESeq2, with the statistical test set to Wald and the fit type set to Parametric. Genes with an FDR < 0.01 were considered significantly differentially expressed. Volcano plots were generated using the ggvolcano R package with a GUI (https://ggvolcanor.erc.monash.edu/)48. KEGG pathway enrichment analysis was performed using the clusterProfiler R package (version 4.8.3)49, with gene annotation provided by org.Hs.eg.db. Gene symbols were first converted to ENTREZ IDs using the bitr() function. Enrichment analysis was conducted using the enrichKEGG() function, using organism = “hsa” and qvalueCutoff = 0.05. The results were converted to human-readable gene names using setReadable(). Visualization was carried out using dotplot() and cnetplot(), displaying the top 10 enriched pathways (showCategory = 10). GSEA for KEGG pathways was conducted using the gseKEGG() function in the clusterProfiler R package, based on a ranked gene list sorted by log2 fold-change values in descending order. Gene symbols were first converted to ENTREZ IDs using the bitr() function with org.Hs.eg.db. GSEA was then performed using the following parameters: organism = “hsa”, keyType = “kegg”, exponent = 1, minGSSize = 10, maxGSSize = 500, eps = 1e-10, pvalueCutoff = 0.05, pAdjustMethod = “BH”, use_internal_data = FALSE, and seed = FALSE. Enrichment plots for selected pathways were generated using the gseaplot() function. Heatmaps were generated using the ComplexHeatmap R package (version 2.16.0), based on TPM-normalized gene expression data. Genes in the KEGG p53 signaling pathway were extracted by merging pathway gene sets with RNA-Seq expression data using gene symbols as the common identifier. Expression values were transformed to Z-scores by applying row-wise scaling (mean-centered and variance-standardized per gene), and rows with missing values were omitted. Heatmaps were constructed using the Heatmap() function with parameters to display row-wise clustering, black grid borders (rect_gp), column and row titles, and customized font sizes and dendrogram orientations. The RNA-Seq data generated in this study were deposited in the NCBI Gene Expression Omnibus (GEO accession number: GSE261020).
Clinical data analysis
Variations in gene expression in neuroblastoma clinical samples with/without MYCN amplification and at each stage were analyzed using the R2 Genomics Analysis and Visualization Platform and the SEQC (Tumor Neuroblastoma-SEQC-498-custom-ag44kcwolf) microarray data set.
Immunofluorescence analysis
LAN-5 cells were seeded in a 24-well plate (Falcon, catalog no. 353047) at 7.0 × 104 cells/well, cultured for 24 h, then stimulated for 48 h with DMSO, PTC-028 (25 nM), and Chk1 inhibitor (LY2606368; 10 nM). The cells were washed with PBS, fixed with 4% paraformaldehyde for 30 min at room temperature, then permeabilized for 10 min in PBS containing 0.2% Triton X-100. Blocking solution (PBS containing 3% BSA, 0.1% glycine, and 0.1% NaN3) was then added, and the cells were left to stand at room temperature for 1 h. The primary antibody (Table S4) was added, and the cells were left to stand overnight in the dark at 4 °C. Secondary antibody (Alexa Fluor™ 488F fragment of goat anti-rabbit; Invitrogen) was then added, and the cells were left to stand for 1 h in the dark at room temperature. Finally, the cells were mounted using Prolong™ Diamond Antifade Mountant with DAPI (Invitrogen, catalog no. P36962) and left to stand overnight in the dark at room temperature. Stained cells were observed using a confocal laser scanning microscope (LSM800 with Airyscan; Zeiss). Antibodies were diluted in blocking solution.
Human tumor xenograft models
This study was conducted with approval from the Animal Care and Use Committee of Nagoya City University Graduate School of Pharmaceutical Sciences, approval number 23–009. All experiments were performed in accordance with the guidelines for the care and use of laboratory animals established by the facility and the US National Institutes of Health. In addition, this study was conducted and reported in accordance with the ARRIVE guidelines (Animal Research: Reporting of In Vivo Experiments) to ensure transparency and reproducibility in animal research.
Female NOG mice (Central Institute for Experimental Animals) were subcutaneously implanted with LA–N-5 tumor cells suspended in PBS (1:1) at a density of 5.0 × 106 cells/200 μl in the lower-right abdominal region, thus successfully establishing tumor engraftment at the implantation site. Tumor growth was monitored regularly following transplantation, and the treatment was initiated (day 0) when the tumor volume reached approximately 150 mm3. On average, this occurred 2–3 weeks after tumor implantation. Tumor size was measured five times, on days 0, 3, 7, 10, and 14. PTC-028 was dissolved in DMSO, and the solution was adjusted to a final concentration of 5% ethanol, 0.5% HPMC, and 1% Tween-80 in PBS. Intraperitoneal administration of PTC-028 was performed on days 0, 3, 7, and 10. Tumors were transplanted subcutaneously into 6-week-old mice, and treatment was administered for 2 weeks, starting at 8 or 9 weeks of age. Tumors were excised at 10 or 11 weeks of age. Prior to tumor excision, the mice were euthanized via cervical dislocation.
Mice were routinely monitored for signs of pain, distress, weight loss, ulceration, or abnormal behavior, and euthanized if humane endpoints were met. No animals exhibited signs of significant suffering throughout the study.
Immunohistochemistry analysis
Collected tumors were embedded in OCT compound, and frozen sections were prepared using a cryostat (CM1850UV; LEICA) and dried on glass slides. Frozen sections were fixed in 4% paraformaldehyde for 30 min, then washed with PBS. Blocking solution (3% BSA, 0.1% glycine, 0.1% NaN3, 0.2% Tween-20) was added, and the sections were left to stand for 1 h at room temperature. Primary antibody (Table S2) was added, and the sections were left to stand overnight in the dark at 4 °C. Secondary antibody (goat anti-mouse IgG [H + L] highly cross-adsorbed secondary antibody, Alexa Fluor™ 488; Invitrogen, catalog no. A-11001) was added, and the sections were left to stand for 1 h in the dark at room temperature. Finally, the sections were mounted using Prolong™ Diamond Antifade Mountant with DAPI and observed using a Keyence microscope (BZ-X800). Antibodies were diluted in blocking solution.
RT-qPCR
Total RNA was extracted using RNAiso Plus (Takara Bio, Inc., catalog no. 9109) in accordance with the manufacturer’s protocol. For reverse transcription, 250 ng of total RNA was converted to cDNA using PrimeScript RT Master Mix (Takara Bio, Inc., catalog no. RR036B). Quantitative real-time PCR was carried out using GoTaq qPCR Master Mix (Promega Corp., catalog no. A6002), employing a 1:10 dilution of the synthesized cDNA and gene-specific forward (F) and reverse (R) primers. Primer sequences are provided in Table S5. Glyceraldehyde-3-phosphate dehydrogenase served as the internal reference gene.
Statistical analysis
All statistical analyses were performed using EZR (Saitama Medical Center, Jichi Medical University, Saitama, Japan), a graphical user interface for R. Specifically, EZR is a modified version of R Commander designed to add functions frequently used in biostatistics50. Student’s t-test was used for comparisons between two groups, and Tukey’s multiple comparison test was used after one-way analysis of variance (ANOVA) for comparisons between multiple groups. Cell viability and xenograft model data were compared using two-way ANOVA. Data are presented as the mean ± SE. Differences were considered statistically significant at p < 0.05.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request. The RNA-Seq data shown in this study were deposited in the NCBI Gene Expression Omnibus (GEO accession number: GSE261020).
References
Nakagawara, A. et al. Neuroblastoma. Jpn. J. Clin. Oncol. 48, 214–241 (2018).
Matthay, K. K. et al. Neuroblastoma. Nat. Rev. Dis. Primers 2, 1–21 (2016).
Johnsen, J. I., Dyberg, C. & Wickström, M. Neuroblastoma-A neural crest derived embryonal malignancy. Front. Mol. Neurosci. 12, 9 (2019).
Cohn, S. L. et al. The international neuroblastoma risk group (INRG) classification system: An INRG task force report. J. Clin. Orthod. 27, 289–297 (2009).
Takita, J. Molecular basis and clinical features of neuroblastoma. JMA J 4, 321–331 (2021).
Otte, J., Dyberg, C., Pepich, A. & Johnsen, J. I. MYCN function in neuroblastoma development. Front. Oncol. 10, 624079 (2020).
Zhang, J. T. et al. MycN Is critical for the maintenance of human embryonic stem cell-derived neural crest stem cells. PLoS ONE 11, e0148062 (2016).
Hansford, L. M. et al. Mechanisms of embryonal tumor initiation: Distinct roles for MycN expression and MYCN amplification. Proc. Natl. Acad. Sci. U. S. A. 101, 12664–12669 (2004).
Chen, J. & Guan, Z. Function of oncogene mycn in adult neurogenesis and oligodendrogenesis. Mol. Neurobiol. 59, 77–92 (2022).
Pession, A. & Tonelli, R. The MYCN oncogene as a specific and selective drug target for peripheral and central nervous system tumors. Curr. Cancer Drug Targets 5, 273–283 (2005).
Huang, M. & Weiss, W. A. Neuroblastoma and MYCN. Cold Spring Harb. Perspect. Med. 3, a014415 (2013).
Meyer, N. & Penn, L. Z. Reflecting on 25 years with MYC. Nat. Rev. Cancer 8, 976–990 (2008).
Liu, R., Shi, P., Wang, Z., Yuan, C. & Cui, H. Molecular mechanisms of MYCN dysregulation in cancers. Front. Oncol. 10, 625332 (2020).
Dang, C. V., Reddy, E. P., Shokat, K. M. & Soucek, L. Drugging the “undruggable” cancer targets. Nat. Rev. Cancer 17, 502–508 (2017).
Parreno, V., Martinez, A.-M. & Cavalli, G. Mechanisms of Polycomb group protein function in cancer. Cell Res. 32, 231–253 (2022).
Geng, Z. & Gao, Z. Mammalian PRC1 complexes: Compositional complexity and diverse molecular mechanisms. Int. J. Mol. Sci. 21, 8594 (2020).
Wang, M.-C. et al. BMI-1, a promising therapeutic target for human cancer. Oncol. Lett. 10, 583–588 (2015).
Sanchez, A. et al. Transcription-replication conflicts as a source of common fragile site instability caused by BMI1-RNF2 deficiency. PLoS Genet. 16, e1008524 (2020).
Fitieh, A. et al. BMI-1 regulates DNA end resection and homologous recombination repair. Cell Rep. 38, 110536 (2022).
Ochiai, H. et al. Bmi1 is a MYCN target gene that regulates tumorigenesis through repression of KIF1Bbeta and TSLC1 in neuroblastoma. Oncogene 29, 2681–2690 (2010).
Cui, H. et al. Bmi-1 is essential for the tumorigenicity of neuroblastoma cells. Am. J. Pathol. 170, 1370–1378 (2007).
Akita, N. et al. Polycomb group protein BMI1 protects neuroblastoma cells against DNA damage-induced apoptotic cell death. Exp. Cell Res. 422, 113412 (2023).
Dey, A. et al. Evaluating the mechanism and therapeutic potential of PTC-028, a novel inhibitor of BMI-1 function in ovarian cancer. Mol. Cancer Ther. 17, 39–49 (2018).
Shields, C. E. et al. Epigenetic regulator BMI1 promotes alveolar rhabdomyosarcoma proliferation and constitutes a novel therapeutic target. Mol. Oncol. 15, 2156–2171 (2021).
Zhong, C. et al. Efficacy of the novel tubulin polymerization inhibitor PTC-028 for myelodysplastic syndrome. Cancer Sci. 111, 4336–4347 (2020).
Abuetabh, Y. et al. DNA damage response revisited: The p53 family and its regulators provide endless cancer therapy opportunities. Exp. Mol. Med. 54, 1658–1669 (2022).
Nagasaka, M. et al. Insights into regulators of p53 acetylation. Cells 11, 3825 (2022).
Moldovan, G.-L., Pfander, B. & Jentsch, S. PCNA, the maestro of the replication fork. Cell 129, 665–679 (2007).
Abbas, T. & Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 9, 400–414 (2009).
Lin, E.-H. et al. Targeting cancer stemness mediated by BMI1 and MCL1 for non-small cell lung cancer treatment. J. Cell. Mol. Med. 26, 4305–4321 (2022).
Fernández-Marrero, Y., Spinner, S., Kaufmann, T. & Jost, P. J. Survival control of malignant lymphocytes by anti-apoptotic MCL-1. Leukemia 30, 2152–2159 (2016).
Belmar, J. & Fesik, S. W. Small molecule Mcl-1 inhibitors for the treatment of cancer. Pharmacol. Ther. 145, 76–84 (2015).
Lamers, F. et al. Targeted BCL2 inhibition effectively inhibits neuroblastoma tumour growth. Eur. J. Cancer 48, 3093–3103 (2012).
Bate-Eya, L. T. et al. High efficacy of the BCL-2 inhibitor ABT199 (venetoclax) in BCL-2 high-expressing neuroblastoma cell lines and xenografts and rational for combination with MCL-1 inhibition. Oncotarget 7, 27946–27958 (2016).
Lestini, B. J. et al. Mcl1 downregulation sensitizes neuroblastoma to cytotoxic chemotherapy and small molecule Bcl2-family antagonists. Cancer Biol. Ther. 8, 1587–1595 (2009).
Jacob, M. et al. Increased MCL1 dependency leads to new applications of BH3-mimetics in drug-resistant neuroblastoma. Br. J. Cancer 129, 1667–1678 (2023).
Klenke, S. et al. The small molecule Bcl-2/Mcl-1 inhibitor TW-37 shows single-agent cytotoxicity in neuroblastoma cell lines. BMC Cancer 19, 243 (2019).
Pan, R. et al. Synthetic lethality of combined Bcl-2 inhibition and p53 activation in AML: Mechanisms and superior antileukemic efficacy. Cancer Cell 32, 748-760.e6 (2017).
Gamble, L. D., Kees, U. R., Tweddle, D. A. & Lunec, J. MYCN sensitizes neuroblastoma to the MDM2-p53 antagonists Nutlin-3 and MI-63. Oncogene 31, 752–763 (2011).
Calao, M. et al. Direct effects of Bmi1 on p53 protein stability inactivates oncoprotein stress responses in embryonal cancer precursor cells at tumor initiation. Oncogene 32, 3616–3626 (2012).
Sánchez-Beato, M. et al. Variability in the expression of polycomb proteins in different normal and tumoral tissues. A pilot study using tissue microarrays. Mod. Pathol. 19, 684–694 (2006).
Park, I.-K., Morrison, S. J. & Clarke, M. F. Bmi1, stem cells, and senescence regulation. J. Clin. Invest. 113, 175–179 (2004).
Panachan, J. et al. Extracellular vesicle-based method for detecting MYCN amplification status of pediatric neuroblastoma. Cancers (Basel) 14, 2627 (2022).
Pandita, A. et al. Integrated cytogenetic and high-resolution array CGH analysis of genomic alterations associated with MYCN amplification. Cytogenet. Genome Res. 134, 27–39 (2011).
Lodrini, M. et al. Using droplet digital PCR to analyze MYCN and ALK copy number in plasma from patients with neuroblastoma. Oncotarget 8, 85234–85251 (2017).
Fox, M. H. A model for the computer analysis of synchronous DNA distributions obtained by flow cytometry. Cytometry 1, 71–77 (1980).
Prieto, C. & Barrios, D. RaNA-Seq: Interactive RNA-Seq analysis from FASTQ files to functional analysis. Bioinformatics 36, 1955–1956 (2019).
Mullan, K. A. et al. ggVolcanoR: A Shiny app for customizable visualization of differential expression datasets. Comput. Struct. Biotechnol. J. 19, 5735–5740 (2021).
Yu, G., Wang, L. G., Han, Y. & He, Q. Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 16, 284–287 (2012).
Kanda, Y. Investigation of the freely available easy-to-use software “EZR” for medical statistics. Bone Marrow Transplant. 48, 452–458 (2013).
Acknowledgements
We acknowledge the assistance of the Research Equipment Sharing Center at Nagoya City University. All illustrations were created using Biorender.com.
Funding
This work was supported in part by Grants-in-Aid for Scientific Research from the Japan Society for the Promotion of Science, KAKEN grant numbers 20K08211, 21K16403, 23K14984, and 23K15455.
Author information
Authors and Affiliations
Contributions
Conceptualization, H.A. and M.A.; data curation, M.H., E.Y., H.A., A.A., K.I., K.T., K.O., M.N., and Y.I.; formal analysis, M.H., E.Y., H.A., K.I., M.N., Y.I., and S.T.; resources, Y.I., H.H., S.T., H.K., and Y.Y.; methodology, A.A., K.O., M.N., Y.I., H.H., S.T., H.K., and Y.Y.; project administration, H.A. and M.A.; supervision, M.A.; funding acquisition, H.A., K.T., H.K., and M.A.; writing–original draft, M.H., E.Y., H.A., K.I., K.T., M.N., Y.I., H.H., S.T., H.K., Y.Y., and M.A. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Hirayama, M., Yamada, E., Aoki, H. et al. Pharmacologic inhibition of BMI1 exerts antitumor effects against MYCN-amplified neuroblastoma, with activation of the p53 pathway. Sci Rep 15, 22917 (2025). https://doi.org/10.1038/s41598-025-06922-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-06922-w
