
Abstract
Inflammatory bowel disease (IBD) involves chronic gut inflammation and barrier damage. While natural flavonoids show promise for IBD treatment, cirsiliol’s effects were unknown. This study examined cirsiliol’s therapeutic potential in mice with DNBS-induced colitis and its mechanisms in vitro and in vivo. Mice received either ethanol (vehicle), DNBS, or cirsiliol (10 or 30 mg/kg). Disease severity was measured by weight loss, colon shortening, and Disease Activity Index (DAI). Lipopolysaccharide (LPS) and interferon-gamma (IFN-γ) stimulated rat intestinal cells (IEC-6) modeled inflammation in vitro. Oxidative stress, inflammatory factors, barrier proteins (Claudin-1, Occludin, E-cadherin), and NF-κB/MAPK pathways were analyzed. Cirsiliol significantly alleviated colitis symptoms, reducing weight loss and DAI while improving colon length. It suppressed pro-inflammatory cytokines (TNF-α, IL-6, IL-1β), boosted antioxidants (increased GSH, decreased MDA), and restored tight junction proteins. Cirsiliol also reduced apoptosis and enhanced cell migration. Mechanistically, it inhibited the activation of NF-κB and MAPK pathways seen in DNBS-induced mice. These findings demonstrate cirsiliol’s protective effects against colitis via anti-inflammatory, antioxidant, and barrier-enhancing actions, mediated by NF-κB and MAPK pathway inhibition, suggesting its potential as a novel IBD treatment.
Similar content being viewed by others
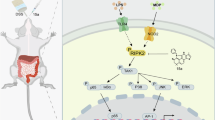


Introduction
Inflammatory bowel disease (IBD), including Crohn’s disease and ulcerative colitis (UC), is a group of gastrointestinal disorders characterized by chronic intestinal inflammation, which is usually accompanied by symptoms such as abdominal pain, diarrhea, weight loss, and fatigue1,2. The pathogenesis of IBD is complex with multiple factors involved, including abnormal activation of the immune system, impaired intestinal barrier function, and imbalance of the intestinal microbiota3,4. Notably, the intestinal barrier, composed of a single layer of epithelial cells joined by tight junctions, plays a crucial role in maintaining gut homeostasis by selectively regulating the passage of luminal contents while preventing the translocation of harmful substances and pathogens5. Disruption of this barrier integrity has been identified as a key initiating factor in IBD pathogenesis, with studies demonstrating increased intestinal permeability preceding clinical symptoms in both Crohn’s disease and ulcerative colitis patients6. Studies have shown that increased epithelial apoptosis and impaired tight junction assembly contribute to enhanced mucosal permeability, bacterial translocation, and subsequent immune activation7,8. Current clinical management of IBD primarily relies on anti-inflammatory and immunosuppressive therapies, including 5-aminosalicylic acid (5-ASA) preparations, corticosteroids, immunomodulators (azathioprine, methotrexate), and biological agents targeting Tumor Necrosis Factor-alpha (TNF-α), integrins, or interleukins9. However, these therapeutic approaches present several limitations, including variable efficacy, potential side effects, and high treatment costs10. Moreover, while these treatments primarily focus on controlling inflammation, they often fail to address the underlying epithelial barrier dysfunction, which may contribute to disease recurrence and treatment resistance11.
Flavonoid compounds are widely distributed in a variety of plants and have received much attention from the scientific community because of their multifaceted biological activities, including potent anti-inflammatory, antioxidant, anticancer, and antimutagenic properties, as well as the ability to modulate the function of key cellular enzymes12. Flavonoids are structurally diverse, with phenolic hydroxyl groups and conjugated double bonds, and are therefore effective in scavenging reactive oxygen species (ROS) and modulating various signaling pathways involved in inflammation and cellular homeostasis13,14. Cirsiliol is a natural flavonoid compound predominantly found in Salvia officinalis and Artemisia spp. with a typical flavonoid structure that has shown unique therapeutic potential for inflammation-related diseases and cancer15,16. For instance, Lim et al. reported that treatment with 20 µM cirsiliol for 12 h significantly alleviated IL-6-induced inflammation and dysfunction in Hep3B cells17. One of the earliest studies on cirsiliol revealed its ability to induce concentration-dependent relaxation of isolated rat ileum by inhibiting calcium (Ca²⁺) influx18. Another investigation showed that higher concentrations of cirsiliol (40 µM vs. 10 µM) effectively suppressed the viability of colorectal cancer cell lines HCT116 and SW48019. However, evidence regarding the application of cirsiliol in IBD remains extremely limited, and the molecular mechanisms through which it may exert its effects have yet to be elucidated.
However, the therapeutic effects of other flavonoids on intestinal diseases, especially IBD, have attracted the attention of researchers. For example, total flavonoids extracted from Abelmoschus manihot L. Medic (TFA) were shown to markedly alleviate inflammation in 2,4,6-Trinitrobenzenesulfonic acid (TNBS)-induced colitis in mice, while also inhibiting lipopolysaccharide (LPS)-induced activation of the NF-κB and MAPK signaling pathways in RAW264.7 macrophages20. In the dextrose sodium sulfate (DSS)-induced colitis model, both dietary antioxidants, rutin and the glycoside ligand quercetin, were shown to significantly ameliorate the symptoms of DSS-induced colitis by inhibiting oxidative stress and attenuating the expression of inflammatory factors in the colonic mucosa21. Moreover, in vitro studies using Caco-2 cell monolayers demonstrated that quercetin enhanced the expression of tight junction proteins such as Zonula Occludens-1 (ZO-1) and occludin, and improved transepithelial electrical resistance, suggesting its potential to restore intestinal barrier integrity22.
We aim to systematically investigate the protective effect of cirsiliol on inflammatory bowel disease (IBD), focusing on its regulatory role in excessive inflammation and intestinal barrier function in both cellular and animal models. Additionally, we seek to elucidate the underlying molecular mechanisms by examining its impact on key signaling pathways, such as nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and mitogen-activated protein kinase (MAPK) signaling pathways. This study is expected to provide valuable insights into the therapeutic potential of natural flavonoids in IBD and contribute to the development of novel flavonoid-based therapies for gastrointestinal diseases.
Methods
Reagents and instruments
DNBS, cirsiliol (purity > 98%), LPS, IFN-γ, purchased from Sigma Chemicals Company (Sigma, Milan, Italy). Antibodies: ZO-1, Occludin, Claudin-1, E-cadherin (Cell Signaling Technology, Danvers, MA, USA), Bax, Bcl-2, Caspase-3 (Abcam, Cambridge, UK), β-actin (Santa Cruz Biotechnology, Dallas, TX, USA). Microscope (Leica Microsystems, Wetzlar, Germany), microplate reader (BioTek Instruments, Winooski, VT, USA), Western blot electrophoresis system (Bio-Rad Laboratories, Hercules, CA, USA).
In vitro studies
Animal experiments
Male C57BL/6 mice aged 6–8 weeks (purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd.) were used. All mice were acclimatized for one week before the experiment. The DNBS-induced modeling approach was referenced from Peritore et al. and Rapa et al. related to IBD23,24,25,26. Mice were anesthetized using isoflurane (2–3%). DNBS solution (3 mg/mL in 50% ethanol) was administered into the colon via a catheter, approximately 4 cm from the anus. Control group mice received an equal volume of 50% ethanol. Mice were randomly divided into four groups: Ethanol 50% group, DNBS-induced IBD model group (DNBS group), IBD model with cirsiliol 10 mg/kg treatment group (Cir-Low group), and IBD model with cirsiliol 30 mg/kg treatment group (Cir-High group). Sulfasalazine-induced IBD model was used as positive control in evaluating Clinical and physiological assessment of normal and DNBS-induced colitis mouse models by HE staining. Due to the limited use of cirsiliol in mice, we refer to published studies in Wistar rats and set up two dose groups accordingly to cover the range of low and potentially high effects27,28. Cirsiliol was administered daily by gavage, while the Ethanol 50% group and DNBS group received an equal volume of solvent. After 10 days of continuous administration, subsequent evaluations were performed. From the first day of the study until its conclusion, changes in body weight, colon length, and disease activity index (DAI) were recorded. DAI is a composite score based on weight loss, stool consistency, and rectal bleeding29. The study was approved by the Institutional Animal Care and Use Committee of Anhui Medical University (LLSC20230734) and confirmed that all methods were performed in accordance with ARRIVE guidelines and other relevant guidelines and regulations.
Quantification of goblet cell percentage in intestinal tissue
Intestinal tissue samples were harvested and fixed in 4% paraformaldehyde (PFA) at 4 °C for 24 h. Following fixation, tissues were rinsed with phosphate-buffered saline (PBS), dehydrated through a graded ethanol series (70%, 80%, 95%, and 100%), cleared in xylene, and embedded in paraffin. Paraffin blocks were sectioned at a thickness of 5 μm using a rotary microtome. The sections were stained with hematoxylin and eosin (H&E) and observed under a light microscope at 200× magnification. Goblet cells were identified based on their typical morphology, including pale-staining cytoplasm and basally located nuclei. Quantification was performed by counting goblet cells in at least 10 randomly selected intestinal crypts per section. The percentage of goblet cells was calculated relative to the total number of epithelial cells within each crypt30. Meanwhile, we compared the cirsiliol-treated group to the conventional IBD drug Sulfasalazine treatment.
Detection of oxidative stress levels
Glutathione (GSH) levels were measured using the GSH Assay Kit (Abcam, #ab239727) according to the manufacturer’s protocol. 50 µL of tissue homogenization after centrifugation was mixed with 150 µL of reaction buffer (containing 5,5’-dithiobis-2-nitrobenzoic acid, DTNB). The mixture was incubated at 25 °C for 5 min, and absorbance was measured at 412 nm using the microplate reader (BioTek Synergy H1). Standard curve (0–20 µM GSH) was used for quantification. Lipid peroxidation (MDA) was assessed by MDA Assay Kit (Abcam, #ab118970). Samples were incubated with TBA reagent at 95 °C for 15 min, cooled, and centrifuged, and absorbance of the supernatant was read at 532 nm31.
Detection of pro-inflammatory factors in tissues
50 mg of colon tissue (avoiding necrotic areas) was taken and rinsed with pre-cooled PBS to remove intestinal contents. 500 µL of RIPA lysate (Thermo Fisher, #89900) containing protease inhibitors was added, homogenized on ice using a tissue homogenizer, and the supernatant was centrifuged. TNF-α (R&D Systems, #MTA00B), IL-6 (#M6000B), IL-1β (#MLB00C), MCP-1 (#MJE00) and iNOS (Abcam, #ab253217) were measured by ELISA according to the manufacturer’s protocol. Briefly, samples were incubated in antibody-coated plates, and then biotinylated detection antibodies and streptavidin-HRP were added. color development was performed with TMB substrate, the reaction was interrupted with H₂SO₄, and absorbance was measured at 450 nm (570 nm for iNOS)32.
In vitro studies
Cell culture and inflammatory injury model construction
Based on previous study, IEC-6 cells were treated with simultaneous exposure to two pro-inflammatory stimulants, LPS (10 µg/mL) and IFN-γ (10 U/mL), to establish a model of cellular inflammatory injury23,33. The experimental design included four groups: normal IEC-6 cells (Control group), inflammatory injury model cells (LPS/IFN-γ group), inflammatory injury cells treated with low-dose cirsiliol (10 µM), and with high-dose cirsiliol (50 µM). Therapeutic dose range selection for cirsiliol based on previous studies17,19.
Cell migration assay
IEC-6 cells were inoculated in a 6-well plate, and the cells were allowed to grow to 80% confluence and were delineated with a sterile lance tip. Different concentrations of cirsiliol (0, 10, 30, 50 µM) were added and photographed after 24 h of incubation. The width of the scratch was measured and the migration rate was calculated using ImageJ software34.
ELISA analysis
Cells were seeded in 96-well plates at a density of 4 × 103 cells/well in 100 µL culture medium and allowed to adhere for 24 h at 37 °C. After treatment with different concentrations of cirsiliol, the medium was removed and replaced with fresh medium containing the ELISA reagents for the corresponding antibodies. Following the treatment period, cell lysates were prepared and analyzed according to the manufacturer’s protocol. Absorbance measurements were taken at 450 nm using a microplate reader (Bio-Rad, CA). Protein activity and expression levels were quantified based on the measured absorbance values20.
Western blot analysis
Cells and tissues were rinsed with PBS and lysed using RIPA buffer supplemented with protease and phosphatase inhibitors. The lysates were centrifuged at 12,000 × g for 10 min at 4 °C, and the supernatants were collected and stored at −80 °C for further analysis. Protein concentrations were determined using a BCA Protein Assay Kit, following the manufacturer’s guidelines. Equal amounts of protein (20 µg) were separated on a 12% SDS-PAGE gel and subsequently transferred to a PVDF membrane. The membrane was blocked with 5% skimmed milk in TBST for 1 h at room temperature. The membranes were probed with primary antibodies targeting including Bax, Bcl-2, caspase 3, p-p65, E-cadherin etc. and GAPDH overnight at 4 °C. After washing with TBST, the membranes were incubated with species-specific horseradish peroxidase (HRP)-conjugated secondary antibodies for 1 h at room temperature. Following three washes with TBST, protein bands were visualized using an ECL Detection Kit. The chemiluminescent signals were captured, and GAPDH was used as an internal control for normalization20.
Statistical analysis
All values were expressed as mean ± SEM of three independent experiments. The statistical significance was performed using the two-tailed Student’s t test or one-way ANOVA. The P-value of less than 0.05 was considered significantly different.
Results
Protective effects of cirsiliol on DNBS-induced intestinal injury in mice
The weight of the Ethanol 50% group gradually increased throughout the experimental period. In contrast, DNBS-induced colitis mice showed significant weight loss (P < 0.001). Notably, the 10 mg/kg and 30 mg/kg doses of cirsiliol were effective in attenuating the weight loss compared with the DNBS group (P < 0.001), while the higher dose of cirsiliol exhibited a more pronounced protective effect (Fig. 1A). The effect of cirsiliol on intestinal morphology was assessed by measuring colon length. The colon was significantly shorter in the untreated DNBS group (P < 0.001), and both cirsiliol treatment groups (10 mg/kg and 30 mg/kg) significantly ameliorated this pathologic change (P < 0.001) (Fig. 1B). The DAI assessment showed an increase in the Disease Activity Index (DAI) over time in the DNBS group (P < 0.001). However, treatment with two concentrations (10 mg/kg and 30 mg/kg) of cirsiliol significantly reduced the DAI scores of mice with DNBS-induced colitis (P < 0.001), suggesting an improvement in disease status (Fig. 1C). HE staining showed a significant increase in infiltrating inflammatory cells and decrease in goblet cells in the intestinal tissues of mice with DNBS-induced colitis compared with the control group.The intestinal inflammation was alleviated and the proportion of goblet cells was restored significantly after treatment with cirsiliol, which was close to that in the group treated with the conventional IBD drug, Sulfasalazine (Fig. 1D-E).

Clinical and physiological assessment of normal and DNBS-induced colitis mouse models. (A) Body weight changes: Body weight changes in mice across different treatment groups in the colitis model. The blue line represents the normal group, the red line represents the DNBS-induced colitis mouse model group, the green line represents the DNBS model treated with 10 mg/kg cirsiliol, and the purple line represents the DNBS model treated with 30 mg/kg cirsiliol. (B) Colon length: Measurement of colon length in different groups. (C) Disease Activity Index (DAI) of the four groups. (D-E) HE staining of intestinal tissue and quantitative analysis of Goblet cell percentage. Cir-Low: 10 mg/kg of cirsiliol; Cir-High: 30 mg/kg of cirsiliol. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 indicated statistically significant differences compared to Ethanol 50% group.
Cirsiliol inhibits oxidative stress responses and improves intestinal mucosal barrier function
The relative protein expression levels of ICAM-1, P-selectin, and PAR were significantly elevated in the DNBS group (P < 0.001), compared to Ethanol 50% group. However, treatment with cirsiliol at both low (10 mg/kg) and high (30 mg/kg) doses significantly reduced the expression levels of these proteins (P < 0.001). Notably, the high-dose cirsiliol group demonstrated more pronounced inhibitory effects (Fig. 2A). Oxidative stress markers such as MDA levels were significantly increased and GSH levels were decreased in the DNBS model. Moreover, 30 mg/kg cirsiliol restored better than 10 mg/kg (Fig. 2B).

Therapeutic effects of cirsiliol on intestinal barrier function in vivo. (A) ELISA analysis of the relative protein expression levels of ICAM-1, P-selectin, and PAR across the four groups. (B) Analysis of levels of GSH, MDA expression. Cir-Low: 10 mg/kg of cirsiliol; Cir-High: 30 mg/kg of cirsiliol. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 indicate statistically significant differences compared to Ethanol 50% group.
Cirsiliol modulates multiple signaling pathways to attenuate inflammatory response and apoptosis
Comprehensive analysis of key signaling pathways revealed that significant upregulation of pro-inflammatory and pro-apoptotic proteins, including p-IκBα, p-p65, p-IKKα/β, p-p38, p-ERK, p-JNK, Bax, and Caspase-3 in the LPS/TNF-α induced cellular inflammatory injury model (P < 0.001). Notably, cirsiliol treatment at both concentrations (10 µM and 50 µM) demonstrated remarkable regulatory effects, significantly suppressing the expression of these proteins (P < 0.001). In contrast, the anti-apoptotic protein Bcl-2, which was markedly downregulated in the LPS/TNF-α group, showed significant restoration following cirsiliol administration (P < 0.001) (Fig. 3).

Effects of cirsiliol on NF-κB and MAPK pathways and inhibition of apoptosis. (A) Relative expression of critical protein in NF-κB pathways including p-IκBα, p-p65, and p-IKKα/β. And critical protein in MAPK pathways including p-p38, p-ERK, and p-JNK. (B) The expression levels of Bax, Bcl-2 and Caspase-3 in WB analysis. (C-E) Quantitative results of WB analysis. Cir-Low: 10µM of cirsiliol; Cir-High: 50µM of cirsiliol. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 indicate statistically significant differences compared to Control group.
Cirsiliol enhances intestinal barrier function and repairs damaged intestinal epithelial cells
The integrity of the intestinal epithelial barrier was significantly compromised in the inflammatory injury model group, as evidenced by marked downregulation of key tight junction proteins. Specifically, the expression levels of Claudin-1, Occludin, and E-cadherin were significantly reduced (P < 0.001) compared to normal controls. However, cirsiliol treatment at both concentrations (10 µM and 50 µM) effectively restored the expression of these critical barrier proteins (P < 0.001) (Fig. 4A-C).

Cirsiliol rescues intestinal barrier injury caused by IBD and promotes repair of the intestinal epithelium. (A-D) The expression level of ICAM-1, Claudin-1, Occludin, E-cadherin via WB analysis. (E) Quantitative analysis of cell migration assay. Cir-Low: 10µM of cirsiliol; Cir-High: 50µM of cirsiliol. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 indicate statistically significant differences compared to Control group.
The reparative capacity of intestinal epithelium was further evaluated through cell migration assays. The inflammatory injury model group exhibited significantly impaired cell migration (P < 0.001). Notably, cirsiliol treatment demonstrated a dose-dependent enhancement of epithelial migration, with increasing concentrations (0, 10, 30, and 50 µM) progressively improving migration rates (P < 0.001) (Fig. 4D). This dose-responsive effect suggests that cirsiliol not only strengthens the intestinal barrier through upregulation of tight junction proteins but also actively promotes epithelial restitution and wound healing.
Cirsiliol exhibits Anti-Inflammatory effects
The anti-inflammatory efficacy of cirsiliol was demonstrated through its significant modulation of key inflammatory mediators. In DNBS-induced colitis mice, pro-inflammatory cytokines TNF-α, IL-6, and IL-1β were markedly elevated (P < 0.001). However, cirsiliol treatment at both concentrations (10 mg/kg and 30 mg/kg) effectively suppressed their expression levels (P < 0.001) (Fig. 5A).

Anti-inflammatory efficacy of cirsiliol in vivo. (A) TNF-α, IL-6, IL-1β expression levels via ELISA analysis. (B) MCP-1 and iNOS expression levels via ELISA analysis. Cir-Low: 10 mg/kg of cirsiliol; Cir-High: 30 mg/kg of cirsiliol. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 indicate statistically significant differences compared to Ethanol 50% group.
Furthermore, cirsiliol demonstrated significant regulatory effects on inflammatory signaling molecules. The expression of MCP-1 and iNOS, which were substantially upregulated in DNBS group (P < 0.001), was significantly attenuated following cirsiliol administration at both 10 mg/kg and 30 mg/kg concentrations (P < 0.001) (Fig. 5B).
Discussion
The therapeutic potential of flavonoids in IBD has been extensively documented. For example, quercetin can alleviate DSS-induced colitis by inhibiting NF-κB pathway activation and enhancing tight junction protein expression35. Apigenin was found to target NRF2 to inhibit pro-inflammatory factor release and repair the intestinal epithelial barrier36. Baicalin, on the other hand, exerts anti-inflammatory and antioxidant effects by regulating intestinal microbiota and metabolism37. In addition, luteolin can significantly reduce the expression of inflammatory factors such as IL-6 and TNF-α in the colonic mucosa by inhibiting the NF-κB pathway and improve the pathological changes of DSS-induced experimental enteritis38. Based on the DSS-induced UC model in mice, Cao et al. found that naringin also down-regulated the phosphorylation level of MAPK, thereby reducing inflammation and oxidative damage in the intestinal mucosa39. These studies have demonstrated that flavonoids can play a wide range of roles in ameliorating intestinal inflammation and repairing the barrier function from different perspectives. Notably, cirsiliol seems to have a broader immunomodulatory function, especially in the regulation of apoptosis. In this study, we found that cirsiliol not only inhibited key inflammatory signaling pathways such as NF-κB and MAPK, but also regulated both endogenous and exogenous apoptotic pathways. Specifically, cirsiliol up-regulated the anti-apoptotic protein Bcl-2 and down-regulated the cleavage expression of pro-apoptotic proteins Bax and Caspase-3, suggesting that cirsiliol may further enhance the mucosal barrier function by maintaining the survival and stability of epithelial cells. Compared with other single-targeted flavonoids, the multi-targeted mechanism of action of cirsiliol gives it a greater potential for application in IBD treatment. Toxicological studies on cirsiliol are relatively limited, but preliminary results suggest that it is well tolerated and has low toxicity. For example, an acute toxicity study evaluating Echinops ritro L. extracts, of which cirsiliol is a major component, showed no hepatotoxicity40. Nonetheless, systematic chronic toxicity, pharmacokinetic, and population studies are lacking, and more comprehensive safety assessments are needed to further advance its drug development.
Cirsiliol is widely found in a wide range of aromatic plants, especially in high content in plants of the genus Salvia spp. and Baccharis spp. such as Salvia officinalis (medicinal sage), Salvia x jamensis and Baccharis trimera41. In phytochemical studies, cirsiliol is often used as an indicator component for specific plant species for differentiation and quality control due to its specificity and stability in specific plants. Several studies have used it in phytopharmaceutical standardization processes, indicating its potential as an important phytochemical and pharmacological marker42,43. Moreover, cirsiliol-rich plants have a long history of use in traditional ethnomedicines in several regions of the world. For example, in the Mediterranean region, Salvia officinalis has long been used to treat dyspepsia, diarrhea, stomach cramps, and inflammatory bowel symptoms44. And in Arabia and South America, Salvia officinalis and Baccharis trimera are widely used in folk remedies for the relief of gastrointestinal disorders, loss of appetite, and chronic gastritis45,46. The ethnobotanical uses of these plants provide historical support for the study of their active ingredient, cirsiliol, in the treatment of gastrointestinal disorders, and further strengthens the scientific link between the traditional uses of plant-derived medicines and the exploration of modern pharmacological mechanisms.
The intestinal barrier represents a sophisticated defense system, comprising epithelial cells, tight junctions, and the mucus layer, which collectively maintain mucosal homeostasis. In IBD pathogenesis, barrier dysfunction manifests as increased intestinal permeability, allowing luminal antigens and pathogens to access the lamina propria, thereby triggering and perpetuating inflammatory responses47. Our findings demonstrate that cirsiliol significantly enhances barrier integrity through upregulation of tight junction proteins (Claudin-1, Occludin, E-cadherin) and promotion of epithelial restitution. These results align with previous studies on other flavonoids, such as quercetin and Juglanin, which have shown similar barrier-protective effects through modulation of tight junction protein expression and distribution48,49. Notably, the observed enhancement of epithelial migration by cirsiliol parallels findings with baicalein, another flavonoid that accelerates wound healing through activation of NF-κB pathways50.
While our study provides compelling evidence for cirsiliol’s therapeutic potential in IBD, several limitations should be acknowledged. First, the exclusive use of a DNBS-induced colitis model, while well-established for studying acute inflammation, may not fully recapitulate the complex pathophysiology of human IBD, particularly the chronic and relapsing nature of the disease51. Future studies should incorporate additional experimental models, including DSS-induced chronic colitis and genetically engineered mouse models (e.g., IL-10 knockout mice), to better evaluate cirsiliol’s efficacy across different disease stages and phenotypes. Second, our investigation focused primarily on epithelial and immune responses, while the potential effects of cirsiliol on gut microbiota composition and function remain unexplored. Given the crucial role of microbiota in IBD pathogenesis52, future research should employ metagenomic sequencing and metabolomic analyses to assess cirsiliol’s impact on microbial communities and their metabolic activities. Third, the pharmacokinetic profile of cirsiliol, including its bioavailability, tissue distribution, and metabolic fate, requires systematic investigation to facilitate its clinical translation. Recent advances in flavonoid delivery systems, such as nanoparticle encapsulation and prodrug strategies, could be explored to optimize cirsiliol’s therapeutic efficacy. Finally, while our mechanistic studies focused on NF-κB and MAPK pathways, the potential involvement of other signaling pathways, such as JAK-STAT and Nrf2-Keap1, warrants further investigation. Addressing these limitations through comprehensive preclinical studies will be crucial for advancing cirsiliol toward clinical trials and potential therapeutic applications in IBD management.
Conclusion
Our study demonstrated that cirsiliol exerted significant protective effects against DNBS-induced colitis in mice as well as LPS/TNF-α-induced cellular inflammation model, including inhibition of inflammatory response, restoration of intestinal barrier function, reduction of oxidative stress and inhibition of epithelial cell apoptosis. More importantly, this study revealed that cirsiliol played a therapeutic role by inhibiting the activation of NF-κB and MAPK pathways in vitro and in vivo. These findings highlighted the potential of cirsiliol as a therapeutic candidate for IBD and exemplify the research value of cirsiliol in alleviating gastrointestinal diseases. However, further validation of the efficacy and safety of cirsiliol in chronic inflammation models and preclinical studies was needed to lay the foundation for its possible clinical translation.
Data availability
Our study was approved by the Institutional Animal Care and Use Committee of Anhui Medical University (LLSC20230734) and confirmed that all methods were performed in accordance with ARRIVE guidelines and other relevant guidelines and regulations. The data included in this study can be obtained upon request from the corresponding author.
References
Neurath, M. F., Artis, D. & Becker, C. The intestinal barrier: a pivotal role in health, inflammation, and cancer. Lancet Gastroenterol. Hepatol. 10 (6), 73–592 (2025).
Rath, T. et al. Intestinal barrier healing is superior to endoscopic and histologic remission for predicting major adverse outcomes in inflammatory bowel disease: the prospective ERIca trial. Gastroenterology 164 (2), 241–255 (2023).
Fu, E. et al. Biomimetic supramolecular assembly with IGF-1 C delivery ameliorates inflammatory bowel disease (IBD) by restoring intestinal barrier integrity. Adv. Sci. (Weinh). 11 (36), e2403075 (2024).
Wu, Y. et al. Reducing severity of inflammatory bowel disease through colonization of Lactiplantibacillus plantarum and its extracellular vesicles release. J. Nanobiotechnol. 23 (1), 227 (2025).
Cheng, Y. et al. Targeting uPA-uPAR interaction to improve intestinal epithelial barrier integrity in inflammatory bowel disease. EBioMedicine 75, 103758 (2022).
Zhang, H., Wu, J., Li, N., Wu, R. & Chen, W. Microbial influence on triggering and treatment of host cancer: an intestinal barrier perspective. Biochim. Biophys. Acta Rev. Cancer. 1878 (6), 188989 (2023).
Xu, D. et al. Amygdalin alleviates DSS-Induced colitis by restricting cell death and inflammatory response, maintaining the intestinal barrier, and modulating intestinal flora. Cells 13 (5), 444 (2024).
Huang, Y. et al. E3 ligase TRIM65 alleviates intestinal ischemia/reperfusion injury through Inhibition of TOX4-mediated apoptosis. Cell. Death Dis. 15 (1), 29 (2024).
Arebi, N., Dyall, L. & Kamperidis, N. A user’s guide to De-Escalating Immunomodulator and biologic therapy in inflammatory bowel disease. Clin. Gastroenterol. Hepatol. 19 (6), 1300–1301 (2021).
Ventura, I. et al. Therapeutic and Immunologic effects of Short-Chain fatty acids in inflammatory bowel disease: A systematic review. Int. J. Mol. Sci. 25 (20), 10879 (2024).
Covello, C. et al. Parenteral nutrition, inflammatory bowel disease, and gut barrier: an intricate plot. Nutrients 16 (14), 2288 (2024).
Lv, H. W. et al. Phytochemistry and Pharmacology of natural prenylated flavonoids. Arch. Pharm. Res. 46 (4), 207–272 (2023).
Liu, G. et al. GmSTOP1-3 regulates flavonoid synthesis to reduce ROS accumulation and enhance aluminum tolerance in soybean. J. Hazard. Mater. 480, 136074 (2024).
Slika, H. et al. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomed. Pharmacother. 146, 112442 (2022).
Luo, M. et al. Cirsiliol induces autophagy and mitochondrial apoptosis through the AKT/FOXO1 axis and influences methotrexate resistance in osteosarcoma. J. Transl Med. 21 (1), 907 (2023).
Jia, X. et al. Cirsiliol targets tyrosine kinase 2 to inhibit esophageal squamous cell carcinoma growth in vitro and in vivo. J. Exp. Clin. Cancer Res. 40 (1), 105 (2021).
Lim, H. J. et al. In vitro inhibitory effects of cirsiliol on IL-6-induced STAT3 activation through anti-inflammatory activity. Bioorg. Med. Chem. Lett. 29 (13), 1586–1592 (2019).
Mustafa, E. H., Abu Zarga, M. & Abdalla, S. Effects of cirsiliol, a flavone isolated from Achillea fragrantissima, on rat isolated ileum. Gen. Pharmacol. 23 (3), 555–560 (1992).
Jiang, T. et al. Cirsiliol regulates mitophagy in colon cancer cells via STAT3 signaling. Cancer Cell. Int. 22 (1), 304 (2022).
Zhang, D. et al. Total flavone of Abelmoschus manihot ameliorates crohn’s disease by regulating the NF–kappaB and MAPK signaling pathways. Int. J. Mol. Med. 44 (1), 324–334 (2019).
Kwon, K. H., Murakami, A., Tanaka, T. & Ohigashi, H. Dietary rutin, but not its aglycone quercetin, ameliorates dextran sulfate sodium-induced experimental colitis in mice: Attenuation of pro-inflammatory gene expression. Biochem. Pharmacol. 69 (3), 395–406 (2005).
Wang, X. et al. Quercetin ameliorates ulcerative colitis by activating Aryl hydrocarbon receptor to improve intestinal barrier integrity. Phytother Res. 38 (1), 253–264 (2024).
Rapa, S. F. et al. Plumericin protects against experimental inflammatory bowel disease by restoring intestinal barrier function and reducing apoptosis. Biomedicines 9 (1), 67 (2021).
Peritore, A. F. et al. PEA/Polydatin: Anti-Inflammatory and antioxidant approach to counteract DNBS-Induced colitis. Antioxid. (Basel) 10 (3), 464 (2021).
Cruz-Munoz, J. R. et al. Ethanolic extract from Lepidium virginicum L. ameliorates DNBS-induced colitis in rats. J. Ethnopharmacol. 289, 115056 (2022).
Ardizzone, A. et al. Ulva pertusa, a Marine Green Alga, Attenuates DNBS-Induced Colitis Damage via NF-kappaB/Nrf2/SIRT1 Signaling Pathways. J Clin Med. ;11(15). (2022).
Stefkov, G. et al. Effects of Teucrium polium spp. Capitatum flavonoids on the lipid and carbohydrate metabolism in rats. Pharm. Biol. 49 (9), 885–892 (2011).
Barkaoui, T. et al. Alleviated actions of Plantago albicans extract on lead Acetate-Produced hepatic damage in rats through antioxidant and free radical scavenging capacities. J. Med. Food. 23 (11), 1201–1215 (2020).
Sahu, B. D., Kumar, J. M. & Sistla, R. Fisetin, a dietary flavonoid, ameliorates experimental colitis in mice: relevance of NF-kappaB signaling. J. Nutr. Biochem. 28, 171–182 (2016).
Hegarty, L. M., Jones, G. R. & Bain, C. C. Macrophages in intestinal homeostasis and inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 20 (8), 538–553 (2023).
Esiringu, F. et al. Investigation of the effect of intracolonic melatonin gel formulation on acetic acid-induced colitis. Drug Deliv. 23 (7), 2318–2326 (2016).
Chami, B. & Witting, P. K. Immunomodulatory effects of intestinal mesenchymal cells in the context of inflammatory bowel disease. Acta Physiol. (Oxf). 233 (2), e13721 (2021).
Adesso, S. et al. The food contaminants Nivalenol and Deoxynivalenol induce inflammation in intestinal epithelial cells by regulating reactive oxygen species release. Nutrients 9 (12), 1343 (2017).
Basilicata, M. G. et al. Anti-Inflammatory and antioxidant properties of dehydrated Potato-Derived bioactive compounds in intestinal cells. Int. J. Mol. Sci. 20 (23), 6087 (2019).
Wei, Q. et al. Quercetin protected the gut barrier in ulcerative colitis by activating Aryl hydrocarbon receptor. Phytomedicine 140, 156633 (2025).
Xu, B. et al. Apigenin alleviates intestinal ischemia/reperfusion injury via upregulating Nrf2-Mediated tight junction integrity. Mol. Nutr. Food Res. 69 (9), e70043 (2025).
Hu, H. et al. Integrated Microbiome and metabolomics revealed the protective effect of Baicalin on alveolar bone inflammatory resorption in aging. Phytomedicine 124, 155233 (2024).
Xue, L. et al. Luteolin ameliorates DSS-induced colitis in mice via suppressing macrophage activation and chemotaxis. Int. Immunopharmacol. 124(Pt B), 110996 (2023).
Cao, H. et al. Protective effect of naringin on DSS-Induced ulcerative colitis in mice. J. Agric. Food Chem. 66 (50), 13133–13140 (2018).
Zheleva-Dimitrova, D. et al. Antioxidant and hepatoprotective potential of Echinops ritro L. Extracts on induced oxidative stress in vitro/in vivo. Int. J. Mol. Sci. 24 (12), 9999 (2023).
Yanagimichi, M. et al. Analyses of putative anti-cancer potential of three STAT3 signaling inhibitory compounds derived from Salvia officinalis. Biochem. Biophys. Rep. 25, 100882 (2021).
Frezza, C. et al. Phytochemical analysis on the aerial parts of Teucrium capitatum L. with aspects of chemosystematics and ethnobotany. Nat. Prod. Res. 37 (14), 2398–2407 (2023).
Lekmine, S. et al. Investigation of photoprotective, Anti-Inflammatory, antioxidant capacities and LC-ESI-MS phenolic profile of Astragalus gombiformis Pomel. Foods 10 (8), 1937 (2021).
Jaric, S. et al. An ethnobotanical survey of traditionally used plants on Suva planina mountain (south-eastern Serbia). J. Ethnopharmacol. 175, 93–108 (2015).
Qnais, E. Y., Abu-Dieyeh, M., Abdulla, F. A. & Abdalla, S. S. The antinociceptive and anti-inflammatory effects of Salvia officinalis leaf aqueous and butanol extracts. Pharm. Biol. 48 (10), 1149–1156 (2010).
Garcia, F. A. et al. A comparative study of two clerodane diterpenes from Baccharis trimera (Less.) DC. on the influx and mobilization of intracellular calcium in rat cardiomyocytes. Phytomedicine 21 (8–9), 1021–1025 (2014).
Llewellyn, S. R. et al. Interactions between diet and the intestinal microbiota alter intestinal permeability and colitis severity in mice. Gastroenterology 154 (4), 1037–46e2 (2018).
Deng, S. et al. Quercetin alleviates lipopolysaccharide–induced acute lung injury by inhibiting ferroptosis via the Sirt1/Nrf2/Gpx4 pathway. Int. J. Mol. Med. 52 (6), 118 (2023).
Liu, J., Chen, L., Zhang, X., Pan, L. & Jiang, L. The protective effects of Juglanin in cerebral ischemia reduce Blood-Brain barrier permeability via Inhibition of VEGF/VEGFR2 signaling. Drug Des. Devel Ther. 14, 3165–3175 (2020).
Guo, C. et al. Baicalein alleviates non-alcoholic fatty liver disease in mice by ameliorating intestinal barrier dysfunction. Food Funct. 14 (4), 2138–2148 (2023).
Cao, L. et al. Oral enzyme-responsive nanoprobes for targeted theranostics of inflammatory bowel disease. J. Nanobiotechnol. 22 (1), 484 (2024).
Bao, M. et al. ROS scavenging and inflammation-directed polydopamine nanoparticles regulate gut immunity and flora therapy in inflammatory bowel disease. Acta Biomater. 161, 250–264 (2023).
Funding
This research was funded by Natural Science Foundation of Hunan Province (S2023JJBMLH1190).
Author information
Authors and Affiliations
Contributions
Methodology and Formal analysis, JG; Sofware, Validation, Resources, ZQW; Investigations, Data curation, HYL; Writing—original draf, GFJ, LP; Writing—review and editing, XHL; Visualization and Supervision, ZQW, ZXJ; Project administration and Funding acquisition, JG; All authors agreed to submit this version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Gao, J., Li, H., Wang, Z. et al. Cirsiliol alleviates experimental inflammatory bowel disease via restoration of intestinal barrier integrity and inhibition of NF-κB and MAPK pathways. Sci Rep 15, 24408 (2025). https://doi.org/10.1038/s41598-025-09896-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-09896-x
