
Abstract
The black soldier fly frass fertilizer (BSFFF) has gained global attention as a multipurpose input for soil fertilization and pest and disease management. However, there are limited studies that have examined its effects on insect pest resistance and the underlying mechanisms. We investigated the impact of amending soil with BSFFF on maize growth, defense gene expression and resistance to a polyphagous insect herbivore, Spodoptera frugiperda (Lepidoptera: Noctuidae) through larval feeding assay. Maize growth was evaluated by measuring plant height, chlorophyll concentration, and biomass accumulation in soils amended with BSFFF, synthetic fertilizers (Di-ammonium phosphate and Calcium ammonium nitrate) and unfertilized soils at various growth stages. Larval feeding assays were conducted using leaf discs from maize plants grown in different amended soils. The expression level of three maize defense genes: pathogenesis related protein 5 (pr-5), maize proteinase inhibitors (mpi), and lipoxygenase 3 (lox-3) were analyzed using quantitative polymerase chain reaction (qPCR) while yield was assessed through a field trial over two cropping seasons. Maize plants grown in BSFFF amended soils showed 30% more growth, higher chlorophyll, 0.93–2.86 t ha− 1 higher yield, and 48% better nitrogen use efficiency than from those in synthetic or unfertilized soils. Moreover, S. frugiperda larvae consumed significantly less leaf tissue from maize plants grown in BSFFF amended soils than synthetically fertilized and non-fertilized soils. Maize defense genes pr-5, mpi, and lox-3 were highly expressed both constitutively and inductively in maize planted in BSFFF amended soils compared to those grown in synthetically fertilized and non-fertilized soils. We observed a significant negative correlation between mpi gene expression and larval feeding, suggesting its role in maize resistance. Our results show that soil amendment with BSFFF strengthens plant defense systems and positively impacts plant growth and yield, contributing to increased agricultural productivity and sustainability.
Similar content being viewed by others
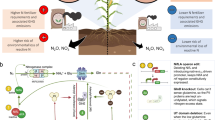


Introduction
The intensification of agriculture to meet the needs of a rapidly growing population is a continuous process that demands multi-dimensional approaches to improve crop yield, soil fertility, and resistance to insect pests. Soil amendments using organic and inorganic fertilizers are common strategies for replenishing depleted soil nutrients, thus improving soil fertility, plant growth and crop production1,2,3. However, the effectiveness of these amendments varies depending on the types and application strategies. For instance, revitalizing soil with inorganic fertilizers temporarily improves soil fertility, requiring multiple applications during growing seasons making it costly4. Additionally, continuous use of synthetic fertilizers has adverse impacts on human and environmental health, and can increase plant susceptibility to pathogens and herbivore pests5,6,7,8,9. For example, Culliney & Pimentel10 observed that plants grown in soils highly fertilized with inorganic nitrogen had increased populations of mites and aphids. This implies that soil amendment practices that improve soil fertility and plant growth do not necessarily increase plants’ capacity to resist herbivore pests.
Unlike inorganic fertilization, organic amendments maintain long-term enhancement of soil fertility1. In addition to its cost-effectiveness, organic fertilizers improve soil chemical properties by increasing availability and uptake of essential nutrients for crop growth and production4,11,12. Organic soil amendments also improve soil structure, reduce erosion while strengthening its water holding capacity13. They further enhance soil biological quality by promoting beneficial macro- and micro-biotic organisms such as nematodes, springtails, earthworms, ground beetles, bacteria, fungi, and protozoa14,15,16. This underscores the ability of organic soil amendments to restore overall soil health by improving its physical, chemical, and biological properties. Given that plant defenses are intrinsically linked to soil health, enhancing these properties through organic amendments may positively impact soil and plant health1,9,17,18,19. Indeed, soil amendment with organic fertilizer has been shown to improve plant resistance to herbivore pests8,20. For instance, European corn borer, Ostrinia nubilalis Hubner (Lepidoptera: Crambidae) female moths oviposited more on plants grown in chemically fertilized soils than those organically fertilized8. These studies suggest that not all soil amendments improve plant defense against herbivore pests21,22.
The use of black soldier fly (Hermetia illucens, L., Diptera: Stratiomyidae) frass fertilizer (BSFFF) as a novel approach for organic soil amendment is gaining global traction due to its pronounced agroecological benefits. It is generated by treating biodegradable waste with black soldier fly larvae which converts organic waste into safe and enriched organic fertilizer23,24. Like many other organic fertilizers, BSFFF positively impacts soil chemical properties by enhancing nutrient abundance and adsorption in the soil25. In addition, BSFFF strengthens soil biological quality by increasing populations of beneficial soil microbiota and improves soil physical properties by increasing organic matter and porosity25,26. Apart from being naturally derived and cost-effective, BSFFF has been shown to interfere with plant-pathogen interactions by reducing the spread of soil-borne fungal pathogens such as Fusarium oxysporum and Rhizoctonia solani27,28and aboveground and soil-dwelling pests29,30. However, its impact in plant-insect interactions, particularly invasive herbivore pests like fall armyworm (Spodoptera frugiperda (Lepidoptera: Noctuidae)), remains largely unexplored.
Spodoptera frugiperda is a highly destructive phytophagous pest, native to Americas but invasive in Africa31Asia32 and Australia33. It represents one of the major global threats to food security, attacking over 80 different agricultural crops including cereal grains (maize, rice, sorghum, groundnut, soybean, millet and cotton), vegetables (cabbage, tomato and potato) and legumes, thus threatening the livelihood of farmers34,35,36,37. The use of agrochemical strategies to control S. frugiperda has proven ineffective due to ecological hazards and the rapid development of pest resistance38,39,40. Ecologically sound pest management strategies that involve less synthetic chemical inputs and emphasize nature-based approaches, including the exploitation of plant resistance, offer a promising solution for mitigating the economic impacts of this pest20.
Generally, plants employ a diverse array of defense genes that are upregulated or downregulated depending on attacking insect species and nutrient availability41,42. Nitrogen, a key soil nutrient, plays a central role in plant growth and defense expression. Apparently, an increase in inorganic nitrogen negatively impacts plant physical and chemical defenses43,44,45. Lu et al.46 demonstrated how the quality and quantity of nitrogen fertilizers significantly affected gene expression under organic and inorganic fertilization. Similarly, Kavroulakis et al.47 and Tenea et al.48 found that plants grown in organically amended soils expressed more defense genes than those grown in conventionally amended soils. Since plant defense response and soil management are intrinsically dependent49soil amendments that influence plant physiological and biochemical components play a critical role in enhancing innate plant defenses against herbivore pests. Therefore, given that BSFFF stimulates soil quality and beneficial microorganisms26it has the potential to enhance both acquired and induced systemic plant resistance.
Numerous studies have investigated the impact of soil amendments with BSFFF on nutrient availability, plant growth, and resistance to pests26,29,30,46. However, little is known about the effects of soil amendment with BSFFF on maize resistance to the invasive S. frugiperda. This study, therefore aimed to investigate how BSFFF amended, synthetically fertilized and unfertilized soils affect the expression of plant defense-related genes. In addition, we evaluated direct plant resistance through larval feeding assays. To capture agronomic outcomes at the field level, we compared maize yields across the three soil treatments. We hypothesized that BSFFF amended soils enhance expression of plant defense genes, thereby increasing resistance to herbivore pests while promoting plant growth and yield.
Materials and methods
Plants and fertilizers
Maize seeds (SC Duma 43) were obtained from Simlaw seeds Ltd, Nairobi, Kenya. The experiments were carried out using two types of fertilizers: organic fertilizer (BSFFF) and synthetic fertilizers, specifically Calcium ammonium nitrate (CAN) and Di-ammonium phosphate (DAP). The BSFFF was obtained from a BSF colony at the Animal Rearing and Containment Unit (ARCU) at icipe, while the synthetic fertilizers were obtained from Kenya Farmers Association Ltd, Nairobi, Kenya. The BSFFF was produced upon feeding black soldier fly larvae on brewery spent grain sourced from Kenya Breweries Ltd, Nairobi, Kenya, and composted following the procedure outlined by Beesigamukama et al.26.
Insects
Spodoptera frugiperda moths were reared in 80 × 60 × 120 cm oviposition cages at the ARCU of icipe, Nairobi, Kenya. The cages were provided with three-week-old maize plants for adult moths to oviposit on. After two days, eggs were collected and transferred to rearing jars (1000 ml with steel-infused lids to allow airflow), where they were incubated until hatching. For all experiments that required neonates, one-day-old neonates were used. Rearing conditions were maintained at 25 ± 2 °C, 72 ± 5% relative humidity (RH), and a 12:12-hour light-dark photoperiod. Second-generation insects were used in all experiments and were mixed with field-collected colonies every two months to maintain their biological characteristics and prevent genetic degradation17.
Laboratory and greenhouse experiments
Laboratory and greenhouse experiments were conducted at Duduville Campus (1.2921° S, 36.8219° E; 1616 m above sea level), of the International Centre of Insect Physiology and Ecology (icipe) located at Nairobi, Kenya. Maize seeds were individually sown in 5 L plastic pots in a greenhouse under optimum conditions (25 ± 2 °C, 72 ± 5% RH; 12 L:12D photoperiod). For soil amendments, BSFFF was mixed with soil at a 1:3 ratio. Five days after planting, 10 g of DAP was applied to each pot with synthetic fertilizer treatment. As a top dressing, 10 g of CAN was added to each pot of synthetic fertilizer treatment 7 days after germination. Soil with no amendments was used as the control. Pots used for the experiment were thoroughly cleaned using 70% ethanol and household bleach (Jik) to prevent cross-contamination. Each 5 L pot was half-filled with fresh soil mixed with the different treatments and arranged in a randomized complete block design with 10 replicates per treatment. Maize seedlings were placed 70 cm apart and watered once a day with 0.2 L until they were four weeks old for use in the experiments.
Maize plant growth parameters were assessed by continuously measuring plant height and chlorophyll concentration weekly for four weeks after germination. Plant height was measured by positioning a tape measure from the soil surface to the arch of the uppermost leaf that had at least halfway emerged from the whorl region of the shoot. Maize chlorophyll concentration was recorded by measuring the average of the three newly and fully formed leaf using SPAD-502 chlorophyll meter (Konica Minolta corporation, Ltd, Osaka, Japan)50.
At the end of the experiment (four weeks), biomass accumulation was assessed by measuring dry shoot and root weight through destructive sampling. Maize plants were cut above the soil level, oven-dried at 80 °C for 48 h, and weighed afterwards for shoot weight. Similarly, below-ground parts of the maize plants were cut, cleaned with water to remove attached soil particles, dried in an oven at 80 °C for 48 h, and then weighed to determine root weight.
Gene expression
To determine the effects of soil amendments on gene expression, the pathogenesis related protein 5 (pr-5), maize proteinase inhibitors (mpi), and lipoxygenase 3 (lox-3) genes were quantified using qRT-PCR. For constitutive defense genes, 1 g of undamaged, newly developed leaf tissue was cut from maize plants grown in soil amended with BSFFF, synthetic fertilizer and non-fertilized soil. The leaf tissue was then placed into 2.0 mL Eppendorf tubes, freeze-dried using liquid nitrogen, and stored at -80 °C for later RNA extraction. To further understand the effect of soil amendments on induced defense genes, another set of maize plants were exposed to ten S. frugiperda neonates for 24 h. Afterwards, 1 g of the newly developed leaf tissue of herbivore damaged maize plant from each of the soil treatments was cut, placed into 2.0 mL Eppendorf tubes, freeze-dried in liquid nitrogen, and stored at -80 °C for later RNA extraction.
RNA extraction and cDNA synthesis
For sample processing, 100 mg of freeze-dried leaf tissue from each treatment was cut and placed into 2 mL Epperdorf tubes with glass beads (BioSpec Products Inc., Bartlesville, Oklahoma, USA) and mechanically homogenized using a Tissue Lyser II (Qiagen Retsch GmbH, Hannover, Germany). Total RNA was extracted using ISOLATE II RNA Mini Kit (Meridian Bioscience, UK) following the manufacturer’s instructions. DNAse treatment was done before elution to clear all DNA contaminants and the resultant RNA concentration and purity was determined using a Nanodrop 2000/2000c spectrophotometer (Thermo Fischer Scientific, Wilmington, USA), and samples were stored at -80 °C for downstream processes. Complementary DNA (cDNA) synthesis was performed using the High-Capacity cDNA Reverse Transcription kit (Applied Biosystems, Waltham, USA) following manufacturer’s instructions. Into a microcentrifuge tube, a 20 µL reaction mix consisting of 10X RT buffer, 8 mM dNTP mix, 0.5 pmol µL− 1 RT randoms primers, 45 ng µL− 1 of total RNA, 5 U/µL MultiScribe™ Reverse Transcriptase and nuclease free water were added. The reactions were set up in a Nexus Mastercycler gradient (Eppendorf, Hamburg, Germany), under the following thermal cycling conditions: initial activation for 10 min at 25 °C, cDNA synthesis for 120 min at 37 °C, the Reverse Transcriptase inactivation at 85 °C for 5 min, then a final holding step at 4°C47,51,.
qPCR analyses
The specific maize defense primers (pr-5, mpi, and lox-3) were selected based on those previously reported by Stratton et al.42. The specific primers were designed using Primer 3.0 software hosted by NCBI (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) (Table 1). Quantitative PCR (qPCR) was performed using a QuantStudio 5 Real-Time System (Thermo Fisher Scientific, Waltham, USA). The reactions were all in triplicates and a minimum of 4 biological replicates. These were set up in 10 µL final reaction volume consisting of 5 µL of SensiFAST SYBR® Hi-ROX Kit (Meridian Bioscience, London, UK), 0.5 pmol µL[- [1 of each primer, 2.5 µL of cDNA template, and 1.5 µL of nuclease free water. The qPCR cycling conditions consisted of initial activation of 95 °C for 2 min; followed by continuous 40 cycles at 95 °C for 15 s, annealing for 45 s, and extension and plate reading at 72 °C for 30 s. Beta tubulin was used as the reference gene to normalize the expression of target genes of interest. Relative gene expression was determined using the delta-delta Ct (2-ΔΔCt) method52.
Larval feeding
To evaluate the influence of soil amendments on insect feeding, larval feeding assays were conducted on undamaged and herbivore-damaged maize plants. One leaf disc of 2.0 cm diameter was cut from a newly formed leaf of maize plants grown in soil amended with BSFFF, synthetic fertilizer and non-fertilized soil. The newly formed leaves are known to contain high levels of defense compounds in maize53. The leaf discs were then placed in 30 mL clear plastic cups containing agar medium (Technical Agar #3) to maintain the physiological state. Ten naïve neonates of S. frugiperda were introduced into the 30 mL small cup containing a maize leaf disc from each treatment group. Afterwards, the small cups were gently sealed with parafilm paper to prevent neonates from escaping, and tiny holes were made using a thin needle at the cup lid to allow air circulation. The neonates were allowed to feed on the leaf discs for 24 h.
To determine the relationship between herbivore-damaged responses and soil amendments, maize plants were exposed to 10 S. frugiperda neonates for 24 h. Afterwards, 2.0 cm diameter leaf discs were cut from newly formed leaf of the exposed maize plants grown in all soil treatments as described in the above constitutive larval feeding assay. Six unique treatments were established, involving S. frugiperda neonates feeding on leaf discs from plants grown in soil amended with BSFFF, synthetic fertilizer and non-fertilized soils, for both constitutive and induced feeding responses. Each treatment was replicated nine times. Images of the leaf discs were taken, and the area fed on each leaf disc calculated using ImageJ software54]– [55.
Gene expression and larval feeding
To examine the relationship between the expression of anti-chewing gene (mpi) and larval feeding56Pearson’s correlation analysis was performed, and a scatter plot was constructed. The average feeding by S. frugiperda neonates on both damaged and undamaged plants was correlated with the expression levels of induced and constitutive mpi defense genes, respectively. Each treatment included four biological replicates.
Field experiments
Field experiments were conducted for two growing seasons; April to September 2023 and October 2023 to March 2024 at the Kenyatta University Teaching and Demonstration Farm (1°10′59″ S, 36°55′34″ E; 1580 m above sea level) in Kiambu County, Kenya. This region experiences bimodal rainfall, with an average annual precipitation of approximately 925 mm and mean monthly temperatures ranging from 21 to 28 °C (www.meteo.go.ke). The short rainy season often starts from March to June, while the long rainy season begins in October and extends to January. During the experimental periods, cumulative rainfall totals were 246 mm and 278 mm for the short and long rainy seasons, respectively. The treatments included: (i) BSFFF amended soil, (ii) synthetic fertilizer applied at a rate equivalent to 60 kg N ha[- [1, and (iii) an unfertilized control soil. To eliminate nutrient limitations from either organic or synthetic fertilizer, phosphorus (P) [supplied as triple super phosphate – TSP (46% P2O5)] and potassium (K) [supplied as muriate of potash (60% K2O)] were obtained from Kenya Farmers’ Association and applied at uniform rates of 60 kg P ha[- [1 and 50 kg K ha[- [1, respectively57. For organic fertilizers treatment, additional inorganic P and K were applied as top up to the nutrients content already present in the dry matter, used to supply the required N, ensuring equivalent nutrient supply across treatments57.
The maize variety SC Duma 43 was used as the test crop. The experiment followed a randomized complete block design (RCBD) with three replicates. Each plot measured 4 × 4 m (m) with border widths of 0.5 m and 1 m between the plots and blocks, respectively29. The TSP fertilizer was applied at planting, while urea and muriate of potash were applied in two equal splits: 50% at 4 weeks after planting and another 50% at 7 weeks after planting. Weeding was conducted three times using a hand hoe, and all plots were managed following standard agronomic practices up to crop maturity. Grain yield data was collected at the harvesting period from each plot area after all the ears had fully dried. Maize plants in the harvested area were cut at ground level and their ears threshed to determine grain and residue weights using a weighing scale. Grain samples were taken to the laboratory and air-dried to 12.5% moisture content for determination of grain yields per plot and on a hectare basis (t ha[- [1). Agronomic nitrogen use efficiency was determined following the method of Baligar et al.58.
Data analyses
Prior to statistical analyses, data were tested for normality and homogeneity of variance using Shapiro-Wilk test and Levene test, respectively. One way analysis of variance (ANOVA) was used to test whether soil amendments had effects on maize growth parameters (plant height, chlorophyll concentration, and shoot and root weight) and yield. Larval feeding and defense gene expression data were analyzed using generalized linear model (GLM) with a quasi-Poisson distribution. Means were compared and separated using Student-Newman-Keuls (SNK) test and Tukey post hoc test (P < 0.05). Two-sample student’s t-test (independent) was used to determine if there were differences between constitutive and induced larval feeding as well as differences between constitutive and induced expression of maize defense genes and agronomic nitrogen use efficiencies. The principal component analysis (PCA) was conducted to explore the relationship between soil amendments and growth parameters, as well as interactions between soil amendments, larval feeding, and expression of maize defense genes. Pearson’s correlation coefficient was performed to investigate the linear relationship between expression of anti-feeding gene (mpi) and larval feeding. All statistical analyses were performed using R software packages (v4.1.2)59with α set at 0.05.
Results
Growth parameters
We found differences regarding plant height, chlorophyll concentration, and biomass accumulation among maize plants grown in BSFFF amended soil, synthetically fertilized, and non-fertilized soils. Maize grew faster in BSFFF amended soil compared to those grown in synthetically fertilized and non-fertilized soils at 14, 21, and 28 days after germination (F2,57 = 22.38, P < 0.001, F2,57 = 33.78, P < 0.001, F2,57 = 75.95, P < 0.001, respectively, Fig. 1A). However, this difference was not observed in the first week after germination (F 2,57 = 5.24, P = 0.080, Fig. 1A). In addition, we observed higher chlorophyll concentrations in maize plants grown in BSFFF amended soil in comparison to those grown in synthetically fertilized and non-fertilized soils at 7, 14, 21, and 28 days after germination, (F 2,57 = 19.15, P < 0.001, F2,57 = 81.25, P < 0.001, F2,57 = 85.47, P < 0.001, F2,57 = 112.70, P = < 0.001, respectively, Fig. 1B). Chlorophyll concentration for maize plants grown in BSFFF amended soil and synthetically fertilized soil were comparable in first, second, and third week after germination (P = 0.140, P = 0.410 and P = 0.460, respectively, Fig. 1B). We observed higher shoot and root dry weight in the maize plants grown in BSFFF amended soil compared to those grown in synthetically fertilized and non-fertilized soils (F 2,27 = 81.18, P < 0.001, F 2,27 = 81.18, P < 0.001 respectively, Fig. 1C).

(A) maize plant height, (B) chlorophyll concentration at one, two, three and four weeks after germination, and (C) box plots representing maize root and shoot weight after four weeks of growth in soil amended with black soldier fly frass fertilizer (BSFFF) and synthetic fertilizers and non-fertilized soil. Different small letters above the error bars and upper whisker indicate a significant difference between the means of the treatments (P < 0.05).
The PCA indicated distinct variations in root and shoot weight, chlorophyll concentration, and plant height based on soil treatment (Fig. 2). For growth parameters, PC1 accounted for 84.3% of the total data variability, while PC2 explained 8.9%, for a combined total of 93.2% (Fig. 2). All measured growth parameters (root and shoot weight, chlorophyll concentration, and plant height) were highest in soil amended with BSFFF compared to the other soil treatments and showed a positive correlation with each other.

Principal component analysis (PCA) of growth parameters on soil amendments with black soldier fly frass fertilizer (BSFFF), synthetic fertilizers, and non-fertilized soils. PC1 = principal component 1; PC2 = principal component 2.
Gene expression
Undamaged maize plant grown in BSFFF amended soil constitutively expressed significantly higher mpi and lox-3 defense genes than those grown in soil amended with synthetic fertilizers and non-fertilized soils (F2,9 = 13.59, P < 0.001; F2,9 = 23.92, P < 0.001, Fig. 3A). However, these differences were not observed in constitutive expression of pr-5 defense gene among the different soil treatments (F2,9 = 0.39, P = 0.680, Fig. 3A). We noted significant high expression of induced defense genes (pr-5 and mpi) in maize plants grown in soil amended with BSFFF than those grown in synthetic fertilizers and non-fertilized soils (F2,9 = 3.82, P = 0.020, F2,9 = 6.65, P = 0.004, Fig. 3B). These differences were not apparent in induced expression of lox-3 gene in undamaged maize plant grown in BSFFF, synthetic fertilizer and non-fertilized soils (F2,9 = 0.78, P = 0.480, Fig. 3B).

Relative quantification of pathogenesis related protein 5 (pr-5), maize proteinase inhibitor (mpi) and lipoxygenase 3 (lox-3) maize defense genes on, (A) undamaged maize plants, (B) S. frugipreda-damaged maize plants, and (C) constitutive and induced expression of defense genes in maize plants grown in soil amended with black soldier fly frass fertilizer (BSFFF), synthetic fertilizers and non-fertilized soils. Different letters above the error bars and upper whisker indicate a significant difference between the means of the treatments (P < 0.05) for A and B. * indicate significant difference, while n.s indicates no significant difference (C).
Spodoptera frugiperda-damaged maize plants grown in soil amended with BSFFF inductively expressed significantly higher pr5, mpi, and lox-3 defense genes compared to undamaged maize plants grown in soil amended with BSFFF (P = 0.003, P = 0.010, P = 0.050, respectively, Fig. 3C). However, there were no differences observed between constitutive and induced expression of pr5, mpi, and lox-3 in maize plant grown in synthetically fertilized soil (P = 0.110, P = 0.150, and P = 0.630, respectively, Fig. 3C) as well as those planted in non-fertilized soils (P = 0.500, P = 0.200, and P = 0.150, respectively, Fig. 3C).
Larval feeding
Spodoptera frugiperda larvae fed significantly less leaf tissue from undamaged maize plants grown in soil amended with BSFFF in comparison to leaf tissue from maize plants grown in synthetically fertilized and non-fertilized soils (F2,42 = 62.73, P < 0.001, Fig. 4A). Similarly, S. frugiperda larvae consumed significantly less maize leaf tissue in initially damaged maize plants grown in BSFFF amended soil in comparison to those grown in soil amended with synthetically fertilized and non-fertilized soils (F2,42 = 103, P < 0.001, Fig. 4B). There was a significant difference in leaf area fed by S. frugiperda between undamaged and damaged maize plants in soil amended with BSFFF (P = 0.020, Fig. 4C). However, there were no significant differences in larval feeding between damaged and undamaged maize plants grown in synthetically fertilized (P = 0.140) and non-fertilized soils (P = 0.250, Fig. 4C).

Box plot representing mean leaf area consumed by Spodoptera frugiperda naïve neonates in a no-choice experiment on (A) undamaged maize plant, (B) damaged maize plant, and (C) comparison between undamaged and damaged maize plant grown in soil amended with black soldier fly frass fertilizer (BSFFF), synthetic fertilizers and non-fertilized soils.
Gene expression and larval feeding
A strong and significant negative correlation was observed between larval feeding and the relative expression of the mpi gene in maize plants grown in soil amended with BSFFF (R = -0.850, P < 0.010, Fig. 5). On the other hand, maize plants grown in soil amended with synthetically fertilized and non-fertilized soils showed no significant correlation and only a weak negative correlation between larval feeding and expression of mpi gene (R = -0.460, P = 0.250; and R = -0.160, P = 0.700, respectively, Fig. 5). Larval feeding and gene expression were explained by PC1 that accounted for 66.1% of the total data variability, and PC2 explained 17.5%, for a total of 83.6% (Fig. 6). Larval feeding was highest in non-fertilized soil while mpi and lox-3 gene expression were highest in soil amended with BSFFF, and pr-5 was highest in soil amended with synthetic fertilizer treatments. Expression of maize defense genes positively correlated with each other but negatively correlated with larval feeding (Fig. 6).

Scatter plot of Pearson correlation between larval feeding of Spodoptera frugiperda naïve neonates and expression of anti-chewing defense gene (maize proteinase inhibitor (mpi)).

Principal component analysis (PCA) of larval feeding and gene expression in soil amendments with black soldier fly frass fertilizer (BSFFF), synthetic fertilizers and unfertilized soils. PC1 = principal component 1; PC2 = principal component 2.
Impact of fertilizer treatments on maize yield and nitrogen use efficiency
The different fertilizer treatments had a significant effect on maize yield (F2,6 = 26.60, P = 0.001), but not on agronomic N use efficiency (T = 1.24, df = 3.99, P = 0.282). Soil amendment with BSFFF and synthetic fertilizer increased maize yield by 2.86 t ha− 1 (105%) and 1.93 t ha− 1 (71%), respectively, compared to the unfertilized control soil (Fig. 7A). Moreover, maize grown in BSFFF amended soil exhibited 0.93 t ha− 1 (20%) higher yield and 48% greater N use efficiency than those grown with synthetic fertilizer (Fig. 7B).

Grain yield (A) and agronomic nitrogen use efficiency (B) of maize grown in soil amended with black soldier fly frass fertilizer (BSFFF) and synthetic fertilizer. Different letters above the error bars indicate a significant difference at P < 0.05.
Discussion
The current study provides empirical evidence that soil amendment with BSFFF not only improves maize plant growth but also reduces larval feeding of an invasive insect pest, S. frugiperda by upregulating maize defense genes. Our findings suggest that BSFFF amended soil mediated alterations in plant defense genes that positively affect direct plant defense traits. To the best of our knowledge, this is the first study to show enhancement of plant defense genes through soil amendment using an insect frass fertilizer while simultaneously deterring insect foliar feeding.
Plant height, chlorophyll concentration, and biomass accumulation on soils amended with BSFFF increased considerably compared to those grown in synthetically fertilized and non-fertilized soils. Soil amendments that improve nitrogen, potassium, and phosphorous availability restore cell growth and promote plant growth and development60. Both organic and inorganic fertilizers enrich soil with essential nutrients that stimulate plant growth61. Possibly, the increased plant growth regardless of the amendment regime was associated with increased nutrient availability. Studies by Beesigamukama et al.25 and Tanga et al.62 have also reported higher plant growth of maize grown in soils amendment with BSFFF than unamended soils. Interestingly, plant height and chlorophyll concentration varied over time after maize germination. Notably, there were no differences in the early days after germination, likely due to slow release and uptake of nutrients in BSFFF amended soils63 resulting in initially low nutrient availability. Thus, the similar early growth rates across all treatments could be due to equal nutrient availability at the start of germination. These results concur with those obtained by Bashir et al.64who showed that maize plants grown in organic, inorganic, and non-fertilized soils attained similar growth rates within the first two weeks of germination. However, at later days of growth stages, plant height and chlorophyll concentration in maize plants grown in soil amended with BSFFF surpassed maize plants grown in synthetically fertilized and non-fertilized soils. This can be linked to the fact that nitrogen release from BSFFF or organically amended soils exhibit a slow pace during the initial stages of plant growth but sustained adequate levels for prolonged soil quality and plant growth25,63,65,66. On the other hand, synthetic fertilizers release nitrogen faster but only for a short period of time67thus, lower growth rate at later stages of plant growth. The increased plant growth at late stages of maize growth in soil amended with BSFFF is likely due to enhanced and sustained nutrient mineralization and adsorption68which not only supports plant growth but also promotes biomass accumulation.
Our study revealed significantly higher maize grain yields in fertilizer treated plots compared to the unfertilized control, highlighting the crucial role of fertilizers in enhancing crop productivity, particularly in the degraded soils of Kenya69. Increased maize yield and agronomic nitrogen use efficiency achieved using BSFFF treated plots, compared to plots treated with mineral fertilizers has been previously reported25,62. These improvements may be attributed to the more effective nutrient supply and availability from the newly introduced frass fertilizer65. Additionally, the high nutrient release associated with the rapid mineralisation rate of BSFFF has been reported to enhance plant growth and productivity26,68. Beyond nutrient supply, BSFFF may also contribute to improved crop performance by enhancing drought and salt stress tolerance, suppressing pests and diseases, and boosting plant defense mechanisms70. Generally, plant defense theory projects a trade-off between plant growth and defense, across species and genotypes51. It further predicts resource allocation in response to biotic and abiotic stresses with rapidly-growing plants typically exhibiting poor defense mechanisms51. Here, we demonstrated that maize grown in soil amended with BSFFF not only had superior growth but also exhibited upregulated maize defense genes, leading to reduced herbivore feeding. This increased direct resistance to S. frugiperda in BSFFF amended soil is both economically and ecologically important, since leaf tissue removal adversely affects photosynthetic activity and reduces yield17,71. Furthermore, the extent of herbivore damage can be associated with pathogen infections, further threatening plant health72,73,74.
What drives this increased resistance to herbivores in maize planted in BSFFF amended soil? In Poaceae species, including maize, plant defense genes such as proteinase inhibitors (PIs) are usually present in seeds and other plant parts where they are synthesized and stored75. These molecules have been identified as potent precursors that mediate resistance against pathogens and herbivore pests53,75. Plant defense genes are often activated by defense signaling pathways such as jasmonic acid, ethylene and salicylic acid to confer anti-chewing and anti-digestive properties against insect attack58,71,78. In this study, maize defense genes including pr-5, lox-3, and mpi were elevated in maize seedlings grown in soil amended with BSFFF. This elevated synthesis of these known defense genes especially the mpi in maize planted in BSFFF amended soil strongly correlated with reduced S. frugiperda larval feeding observed in our results. This reduction in herbivore leaf consumption in otherwise better growing plants in BSFFF amended soil constitutes a paradox. Generally, insect herbivores tend to feed poorly on nutrient-deficient plants and better on well fertilized plants like those grown in soil amended with BSFFF17,76. For example, S. exigua fed more on nitrogen-fertilized plants compared to those with low nutrient availability77. The observed differences are correlated with reduced production of defense molecules in plants fertilized with synthetic fertilizers, making them more susceptible to herbivore attack47,48. Here, we suggest that constitutive expression of higher defense especially the mpi in maize plants grown in BSFFF amended soil could also have higher defense response against the lepidopteran herbivores. Indeed, there is strong evidence that PIs play a key role in plant defense response against insect herbivores through inhibition of proteolytic enzymes such as elastase and chymotrypsin in insect herbivore midguts75. This enzymatic inhibition reduces food digestibility, leading to reduced insect feeding rates54. Our results indicate that BSFFF amended soil increases maize resistance to S. frugiperda by enhancing PIs synthesis, thus limiting insect feeding.
Moreover, exposure of maize plant to S. frugiperda larval feeding further increased expression of defense genes relative to undamaged maize plants in BSFFF amended soil. Consequently, herbivore feeding between insect damaged and undamaged maize plants was found to be significantly reduced in insect damaged maize plants grown in BSFFF amended soil as opposed to other soil treatments. Herbivore feeding not only causes physical damage to plants but also releases oral cues (saliva, regurgitant, and frass) which trigger defense mechanisms53], [78,79, . The observed increment in induction of defense on damaged plants than undamaged plants could therefore be associated with insect-derived elicitors produced by S. frugiperda larvae during feeding which further upregulated maize defense genes in maize grown in BSFFF amended soil. Indeed, plant damage by S. exigua and S. frugiperda has been shown to stimulate accumulation of mpi in plant tissue adjacent to the herbivore damaged parts53,75. What is interesting in the current results is that this accumulation of PIs following insect herbivore damage was only significantly higher in maize plants grown in soil amended with BSFFF as opposed to amendments with synthetically fertilized and non-fertilized soils.
What components in BSFFF-amended soil drive the upregulation of defense genes and subsequent reduction in herbivore feeding? Plants employ comprehensive defense mechanisms and their activation to initiate resistance to herbivore insects are often controlled by quantity and quality of soil nutrients among other factors44. Therefore, nutritional deficiency negatively impact plants’ ability to protect themselves against insect attack through expression of plant defense genes41,42. In addition, plant resistance to herbivore pests is intrinsically linked to not only soil physicochemical properties but mainly soil biological properties9,20. Mattoo & Abdul-Baki80 noted that plant genetic responses are likely influenced by soil microbial communities, shaping plant resistance traits. Given that soil amendment with BSFFF improves soil biological quality25,65,67 the heightened induction of defense genes in BSFFF amended soils could be explained by the effects of the amendment on soil health. Conversely, synthetic fertilizers adversely affect soil microbial diversity81 describing the decreased expression of defense genes. These results align with previous studies which demonstrated that plants grown in soil amended with organic fertilizers induced higher defense gene expression than those grown in inorganic fertilizers47,48.
Conclusions
This study aimed at assessing the impact of amending soil with BSFFF on plant growth, defense genes expression, herbivore pest resistance, agronomic nitrogen use efficiency and yield. The study demonstrates that soil amendment with BSFFF improves maize plant growth and upregulates defense genes, contributing to increased resistance against S. frugiperda, high nutrient use efficiency and grain yield. The elevated expression of maize defense genes plays an important role in plant-insect interactions, effectively reducing S. frugiperda larval feeding. The link between BSFFF soil amendment and the associated expression of plant defense genes demonstrates a new mechanism through which insect frass fertilizer can reduce plant damage by invasive S. frugiperda. This study provides empirical evidence that soil amendments can influence plant defense traits, offering a promising strategy for sustainable pest management and crop protection. Due to the rapid development of insect farming, further studies on the impact of bioactive compounds of BSFFF on soil microbiome need to be systematically explored with particular attention to above-and below-ground microbial shifts. Additionally, future studies should investigate the role of soil amendment with BSFFF on plant phytochemistry and subsequent attraction of insects’ natural enemies and repellence of herbivore pests. A deeper understanding of these mechanisms will provide valuable insights into how soil amendments shape plant growth, defense responses, and herbivore resistance, ultimately contributing to more sustainable agricultural practices and enhanced agroecosystem productivity.
Data availability
The data that support the findings of this study are currently available from the corresponding author upon reasonable request.
References
Alyokhin, A., Nault, B. & Brown, B. Soil conservation practices for insect pest management in highly disturbed agroecosystems – a review. Entomol. Exp. Appl. 168 (1), 7–27. https://doi.org/10.1111/eea.12863 (2020).
Doran, J. W. Soil health and global sustainability: translating science into practice. Agric. Ecosyst. Environ. 88, 119–127. https://doi.org/10.1007/11545163_1 (2002).
Stone, A., Scheuerell, S. J. & Darby, H. Suppression of soilborne diseases in field agricultural systems: organic matter management, cover cropping, and other cultural practices. Soil. Org. Matter Sustainable Agric. 131–177. https://doi.org/10.1201/9780203496374-9 (2004).
Akanmu, A. O. et al. Plant disease management: leveraging on the plant-microbe-soil interface in the biorational use of organic amendments. Front. Plant Sci. 12 https://doi.org/10.3389/fpls.2021.700507 (2021).
Maheshwari, D. K. et al. Integrated approach for disease management and growth enhancement of Sesamum indicum L. utilizing Azotobacter Chroococcum TRA2 and chemical fertilizer. World J. Microbiol. Biotechnol. 28 (10), 3015–3024. https://doi.org/10.1007/s11274-012-1112-4 (2012).
Mattson, W. J. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol. Syst. 11 (1), 119–161. https://doi.org/10.1146/annurev.es.11.110180.001003 (1980).
Slansky, F. Insect nutritional ecology as a basis for studying host plant resistance. Fla. Entomol. 73 (3), 359–378. https://doi.org/10.2307/3495455 (1990).
Phelan, P. L., Mason, J. F. & Stinner, B. R. Soil-fertility management and host preference by European corn borer, ostrinia nubilalis (Hübner), on Zea Mays L.: A comparison of organic and conventional chemical farming. Agric. Ecosyst. Environ. 56 (1), 1–8. https://doi.org/10.1016/0167-8809(95)00640-0 (1995).
Altieri, M. A. & Nicholls, C. I. Soil fertility management and insect pests: harmonizing soil and plant health in agroecosystems. Soil Tillage. Res. 72 (2), 203–211. https://doi.org/10.1016/S0167-1987(03)00089-8 (2003).
Culliney, T. W. & Pimentel, D. Ecological effects of organic agricultural practices on insect populations. Agric. Ecosyst. Environ. 15 (4), 253–266. https://doi.org/10.1016/0167-8809(86)90124-6 (1986).
Lawal, T. E. & Babalola, O. O. Relevance of biofertilizers to agriculture. J. Hum. Ecol. 47 (1), 35–43. https://doi.org/10.1080/09709274.2014.11906737 (2014).
Yadav, G. S. et al. Conservation tillage and nutrient management effects on productivity and soil carbon sequestration under double cropping of rice in North Eastern region of India. Ecol. Ind. 105, 303–315. https://doi.org/10.1016/j.ecolind.2017.08.071 (2019).
Chukwuka, K. S., Akanmu, A. O., Umukoro, B. O. J. & Asemoloye, M. D. Biochar: A vital source for sustainable agriculture. Biostimulants in Plant Science, eds (eds Mirmajlessi, S. M. & Radhakrishnan, R.) (London: IntechOpen). 51–72. https://doi.org/10.5772/intechopen.86568 (2020).
Porras-Alfaro, A., Herrera, J., Natvig, D. O., Lipinski, K. & Sinsabaugh, R. L. Diversity and distribution of soil fungal communities in a semiarid grassland. Mycologia 103 (1), 10–21. https://doi.org/10.3852/09-297 (2011).
Bonanomi, G., Lorito, M., Vinale, F. & Woo, S. L. Organic amendments, beneficial microbes, and soil microbiota: toward a unified framework for disease suppression. Annu. Rev. Phytopathol. 56, 1–20. https://doi.org/10.1146/annurev-phyto-080615-100046 (2018).
Bonanomi, G. et al. Mixtures of organic amendments and Biochar promote beneficial soil microbiota and affect Fusarium oxysporum. Spp. Lactucae, Rhizoctonia Solani and Sclerotinia minor disease suppression. Plant. Pathol. 71 (4), 818–829. https://doi.org/10.1111/ppa.13514 (2022).
Mutyambai, D. M. et al. Push-pull cropping system soil legacy alter maize metabolism and fall armyworm, Spodoptera Frugiperda (Lepidoptera: Noctuidae) resistance through tritrophic interactions. Plant. Soil. https://doi.org/10.1007/s11104-023-06467-9 (2024).
Maschinski, J. & Whitham, T. G. The continuum of plant responses to herbivory: the influence of plant association, nutrient availability, and timing. Am. Soc. Naturalists. 134 (1), 1–19. https://doi.org/10.1086/284962 (1989).
Meyer, G. A. Interactive effects of soil fertility and herbivory on Brassica Nigra. Oikos 88 (2), 433–441. https://doi.org/10.1034/j.1600-0706.2000.880221.x (2000).
Altieri, M., Nicholls, C. I. & Fritz, M. Manage insects on your farm: A guide to ecological strategies. Sustainable agriculture network handbook series, volume 7. Journal of Sustainable Agriculture and Education, 29, 2. College park, MD, north central US. (2005).
Kumar, V., Mills, D. J., Anderson, J. D. & Mattoo, A. K. An alternative agriculture system is defined by a distinct expression profile of select gene transcripts and proteins. Proc. Natl. Acad. Sci. U.S.A. 101 (29), 10535–10540. https://doi.org/10.1073/pnas.0403496101 (2004).
Rojas, C. M., Senthil-Kumar, M., Tzin, V. & Mysore, K. S. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 5, 1–12. https://doi.org/10.3389/fpls.2014.00017 (2014).
Erickson, M. C., Islam, M., Sheppard, C., Liao, J. & Doyle, M. P. Reduction of Escherichia coli O157:H7 and Salmonella enterica serovar enteritidis in chicken manure by larvae of the black soldier fly. J. Food. Prot. 67 (4), 685–690. https://doi.org/10.4315/0362-028X-67.4.685 (2004).
Lalander, C. H., Fidjeland, J., Diener, S., Eriksson, S. & Vinnerås, B. High waste-to-biomass conversion and efficient Salmonella spp. Reduction using black soldier fly for waste recycling. Agron. Sustain. Dev. 35 (1), 261–271. https://doi.org/10.1007/s13593-014-0235-4 (2015).
Beesigamukama, D. et al. Exploring black soldier fly Frass as novel fertilizer for improved growth, yield, and nitrogen use efficiency of maize under field conditions. Front. Plant Sci. 11, 574592. https://doi.org/10.3389/fpls.2020.574592 (2020a).
Beesigamukama, D. et al. In situ nitrogen mineralization and nutrient release by soil amended with black soldier fly Frass fertilizer. Sci. Rep. 11 (1), 1–14. https://doi.org/10.1038/s41598-021-94269-3 (2021).
Choi, S. & Hassanzadeh, N. BSFL drass: A novel biofertilizer for improving plant health while minimizing environmental impact. Candian Sci. Fair J. 2 (2), 41–46. https://doi.org/10.18192/csfj.v2i220194146 (2019).
Quilliam, R. S. et al. Integrating insect Frass biofertilisers into sustainable peri-urban agro-food systems. J. Insects as Food Feed. 6 (3), 315–322. https://doi.org/10.3920/JIFF2019.0049 (2020).
Anedo, E. et al. Unpacking the benefits of black soldier fly Frass fertilizer towards nematode suppression and potato production. Front. Plant. Sci. 16, 1509643. https://doi.org/10.3389/fpls.2025.1509643 (2025).
Kisaakye, J. et al. Chitin-enriched insect Frass fertilizer as a biorational alternative for root-knot nematode (Meloidogyne incognita) management. Front. Plant. Sci. 15, 1361739. https://doi.org/10.3389/fpls.2024.1361739 (2024).
Goergen, G., Kumar, P. L., Sankung, S. B., Togola, A. & Tamò, M. First report of outbreaks of the fall armyworm spodoptera Frugiperda (J. E. Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE. 11 (10), 1–9. https://doi.org/10.1371/journal.pone.0165632 (2016).
Nagoshi, R. N. et al. Southeastern Asia fall armyworms are closely related to populations in Africa and india, consistent with common origin and recent migration. Sci. Rep. 10 (1), 1–10. https://doi.org/10.1038/s41598-020-58249-3 (2020).
Qi, G. J. et al. Source regions of the first immigration of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) invading Australia. Insects 12 (12). https://doi.org/10.3390/insects12121104 (2021).
Day, R. et al. Fall armyworm: impacts and implications for africa. Outlooks on pest management. Outlooks Pest Manage. 28 (5), 196–201. https://doi.org/10.1564/v28 (2017).
De Groote, H. et al. Spread and impact of fall armyworm (Spodoptera Frugiperda J.E. Smith) in maize production areas of Kenya. Agric. Ecosyst. Environ. 292, 106804. https://doi.org/10.1016/j.agee.2019.106804 (2020).
Jalloh, A. A., Yusuf, A. A., Khamis, F., Subramanian, S. & Mutyambai, D. M. Soil legacies in maize-edible legume intercropping alter maize growth and reduce Spodoptera Frugiperda larval feeding. Front. Agron. 5, 1300545. https://doi.org/10.3389/fagro.2023.1300545 (2023).
Peter, E. et al. Crop mixtures influence fall armyworm infestation and natural enemy abundance in maize-based intercropping systems. J. Crop Health. 1–15. https://doi.org/10.1007/s10343-024-01061-0 (2024).
Baudron, F., Zaman-Allah, M. A., Chaipa, I., Chari, N. & Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera Frugiperda J. E. Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Prot. 120, 141–150. https://doi.org/10.1016/j.cropro.2019.01.028 (2019).
Kumela, T. et al. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. Int. J. Pest Manage. 65 (1), 1–9. https://doi.org/10.1080/09670874.2017.1423129 (2019).
Wan, J. et al. Biology, invasion and management of the agricultural invader: fall armyworm, Spodoptera Frugiperda (Lepidoptera: Noctuidae). J. Integr. Agric. 20 (3), 646–663. https://doi.org/10.1016/S2095-3119(20)63367-6 (2021).
Mason, C. J. et al. Plant nutrition influences resistant maize defense responses to the fall armyworm (Spodoptera frugiperda). BMC Plant Biol. 10 (1), 1–11. https://doi.org/10.3389/fevo.2022.844274 (2022).
Stratton, C. A. et al. Nutrition vs association: plant defenses are altered by arbuscular mycorrhizal fungi association not by nutritional provisioning alone. BMC Plant Biol. 22 (1), 1–11. https://doi.org/10.1186/s12870-022-03795-3 (2022).
Mur, L. A. J., Simpson, C., Kumari, A., Gupta, A. K. & Gupta, K. J. Moving nitrogen to the centre of plant defence against pathogens. Ann. Botany. 119 (5), 703–709. https://doi.org/10.1093/aob/mcw179 (2017).
Sun, Y., Wang, M., Mur, L. A. J., Shen, Q. & Guo, S. Unravelling the roles of nitrogen nutrition in plant disease defences. Int. J. Mol. Sci. 21 (2), 1–20. https://doi.org/10.3390/ijms21020572 (2020).
Traw, M. B. & Dawson, T. E. Differential induction of trichomes by three herbivores of black mustard. Oecologia 131 (4), 526–532. https://doi.org/10.1007/s00442-002-0924-6 (2002).
Lu, C. et al. Markedly different gene expression in wheat grown with organic or inorganic fertilizer. Proceedings of the Royal Society B: Biological Sciences, 272(1575), 1901–1908. (2005). https://doi.org/10.1098/rspb.2005.3161
Kavroulakis, N., Papadopoulou, K. K., Ntougias, S., Zervakis, G. I. & Ehaliotis, C. Cytological and other aspects of pathogenesis-related gene expression in tomato plants grown on a suppressive compost. Ann. Botany. 98 (3), 555–564. https://doi.org/10.1093/aob/mcl149 (2006).
Tenea, G. N., Raposo, C., Maquet, A. & F., & Comparative transcriptome profiling in winter wheat grown under different agricultural practices. J. Agric. Food Chem. 60 (44), 10970–10978. https://doi.org/10.1021/jf302705p (2012).
Mitchell, C., Brennan, R. M., Graham, J. & Karley, A. J. Plant defense against herbivorous pests: exploiting resistance and tolerance traits for sustainable crop protection. Front. Plant Sci. 2016, 1–8. https://doi.org/10.3389/fpls.2016.01132 (2016).
Yuan, Z. et al. Optimal leaf positions for SPAD meter measurement in rice. Front. Plant Sci. 7, 1–10. https://doi.org/10.3389/fpls.2016.00719 (2016).
Chuang, W. P. et al. Herbivore cues from the fall armyworm (Spodoptera frugiperda) larvae trigger direct defenses in maize. Mol. Plant Microbe Interact. 27 (5), 461–470. https://doi.org/10.1094/MPMI-07-13-0193-R (2014).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real- time quantitative PCR and the 2 (-Delta delta C(T)). Method 408, 402–408. https://doi.org/10.1006/meth.2001.1262 (2001).
Elvira, S. et al. Spodoptera frugiperda caterpillars suppress herbivore-induced volatile emissions in maize. Journal of Chemical Ecology, 46(3), 344–360. https://doi.org/10.1007/s10886-020-01153-x (2020).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH image to imageJ: 25 years of image analysis. Nat. Methods. 9 (7), 671–675. https://doi.org/10.1038/nmeth.2089 (2012).
Arena, E. T. et al. Quantitating the cell: turning images into numbers with ImageJ. WIREs Dev. Biol. https://doi.org/10.1002/wdev.260 (2017).
War, A. R. et al. Mechanisms of plant defense against insect herbivores. Plant. Signal. Behav. 7 (10), 1306–1320. https://doi.org/10.4161/psb.21663 (2012).
Tittonell, P., Corbeels, M., Van Wijk, M. T., Vanlauwe, B. & Giller, K. E. Combining organic and mineral fertilizers for integrated soil fertility management in smallholder farming systems of kenya: explorations using the crop-soil model FIELD. Agronomy J. 100, 1511–1526. https://doi.org/10.2134/agronj2007.0355 (2008).
Baligar, V. C., Fageria, N. K. & He, Z. L. Nutrient use efficiency in plants. Commun. Soil. Sci. Plant. Analysi. 32, 921–950. https://doi.org/10.1007/978-3-319-10635-9 (2001).
Team, R. D. C. A language and environment for statistical computing. (2020). https://doi.org/10.1603/EN11083
Zhang, Q. et al. Spatial variability of soil nutrients and GIS-based nutrient management in Yongji county, China. Int. J. Geogr. Inf. Sci. 24 (7), 965–981. https://doi.org/10.1080/13658810903257954 (2010).
Aina, O. E., Olowoyo, J. O., Mugivhisa, L. L. & Amoo, S. O. Effect of different soil amendments on growth performance and levels of copper and zinc in Lycopersicon esculentum. Nat. Environ. Pollution Technol. 17 (1), 255–259 (2018).
Tanga, C. M. et al. Performance of black soldier fly Frass fertiliser on maize (Zea Mays L.) growth, yield, nutritional quality, and economic returns. J. Insects as Food Feed. 8 (2), 185–196. https://doi.org/10.3920/JIFF2021.0012 (2022).
Loecke, T. D., Cambardella, C. A. & Liebman, M. Synchrony of net nitrogen mineralization and maize nitrogen uptake following applications of composted and fresh swine manure in the Midwest U.S. Nutr. Cycl. Agrosyst. 93 (1), 65–74. https://doi.org/10.1007/s10705-012-9500-6 (2012).
Bashir, K. A., Muhammad, A. Y., Muhammad, A. & Sada, S. M. Comparing the efficacy of organic and inorganic fertilizers on the growth of maize (Zea Mays L.) plant. J. Agricultural Res. Pesticides Biofertilizers. 3 (3), 1–5. http;//doi.org/01.2022/1.1058 (2022).
Beesigamukama, D. et al. Nitrogen fertilizer equivalence of black soldier fly Frass fertilizer and synchrony of nitrogen mineralization for maize production. Agronomy 10 (9), 5–8. https://doi.org/10.3390/agronomy10091395 (2020b).
Masunga, R. H. et al. Nitrogen mineralization dynamics of different valuable organic amendments commonly used in agriculture. Appl. Soil. Ecol. 101, 185–193. https://doi.org/10.1016/j.apsoil.2016.01.006 (2016).
Gebremikael, M. T., van Wickeren, N., Hosseini, P. S. & De Neve, S. The impacts of black soldier fly frass on nitrogen availability, microbial activities, C sequestration, and plant growth. Front. Sustainable Food Syst. 1–13. https://doi.org/10.3389/fsufs.2022.795950 (2022).
Adin Yéton, B. G. et al. K., M. Decomposition and nutrient release pattern of agro-processing by-products biodegraded by fly larvae in Acrisols. Archives of Agronomy and Soil Science, 65(11), 1610–1621. (2019). https://doi.org/10.1080/03650340.2019.1572118
Gachimbi, L. N. et al. Nutrient balances at farm level in Machakos (Kenya), using a participatory nutrient monitoring (NUTMON) approach. Land. Use Policy. 22, 13–22. https://doi.org/10.1016/j.landusepol.2003.07.002 (2005).
Barrag´ an-Fonseca, K. Y. et al. Insect Frass and exuviae to promote plant growth and health. Trends Plant. Sci. 1–9. https://doi.org/10.1016/j.tplants.2022.01.007 (2022).
Davidson-lowe, E., Ray, S., Murrell, E., Kaye, J. & Ali, J. G. Plant-insect interactions cover crop soil legacies alter phytochemistry and resistance to fall armyworm (Spodoptera frugiperda, lepidoptera: Noctuidae) in maize. Environ. Entomol. 50 (4), 958–967. https://doi.org/10.1093/ee/nvab047 (2021).
Hatcher, P. E. Three-way interactions between plant pathogenic fungi, herbivorous insects and their host plants. Biol. Rev. Camb. Philos. Soc. 70, 639–694 (1995).
Rostas, M., Simon, M. & Hilker, M. Ecological cross-effects of induced plant responses towards herbivores and phytopathogenic fungi. Basic Appl. Ecol. 4 (1), 43–62. https://doi.org/10.1078/1439-1791-00132 (2003).
Gossner, M. M., Beenken, L., Arend, K., Begerow, D. & Peršoh, D. Insect herbivory facilitates the establishment of an invasive plant pathogen. ISME Commun. 1 (1), 1–8. https://doi.org/10.1038/s43705-021-00004-4 (2021).
Tamayo, M. C., Rufat, M., Bravo, J. M. & Segundo, B. S. Accumulation of a maize proteinase inhibitor in response to wounding and insect feeding, and characterization of its activity toward digestive proteinases of Spodoptera littoralis larvae. Planta 211, 62–71. https://doi.org/10.1007/s004250000258 (2000).
Chen, Y., Olson, D. M. & Ruberson, J. R. Effects of nitrogen fertilization on tritrophic interactions. Arthropod-Plant Interact. 4 (2), 81–94. https://doi.org/10.1007/s11829-010-9092-5 (2010).
Ren, L. L., Hardy, G., Liu, Z. D., Wei, W. & Dai, H. G. Corn defense responses to nitrogen availability and subsequent performance and feeding preferences of beet armyworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 106 (3), 1240–1249. https://doi.org/10.1603/EC12091 (2013).
Fürstenberg-Hägg, J., Zagrobelny, M. & Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 14 (5), 10242–10297. https://doi.org/10.3390/ijms140510242 (2013).
Ray, S. et al. Turnabout is fair play: Herbivory-induced plant chitinases excreted in fall armyworm Frass suppress herbivore defenses in maize. Plant Physiol. 171 (1), 694–706. https://doi.org/10.1104/pp.15.01854 (2016).
Mattoo, A. & Abdul-Baki, A. Crop genetic responses to management: Evidence of root–shoot communication. Biological Approaches to Sustainable Soil Systems; Uphoff, N., Ball, A.S., Palm, C., Fernandes, E., Pretty, J., Herren, H., Sanchez, P., Husson, O., Sanginga, N., Laing, M., et al., Eds.; Taylor & Francis: Boca Raton, FL, USA; 221–230. (2006).
Nakhro, N. & Dkhar, M. S. Impact of organic and inorganic fertilizers on microbial populations and biomass carbon in paddy field soil. J. Agron. 9 (3), 102–110. https://doi.org/10.3923/ja.2010.102.110 (2010).
Acknowledgements
We are grateful to the Arthropod Pathology Unit (APU) and Molecular Biology and Bioinformatics Unit (MBBU) of the icipe. Many thanks go to Catherine Awuoche, and Faith Ndunge for insect rearing, Amos Mwangangi for his assistance in greenhouse operations. Our appreciation extends to Levi Ombura for his contributions in laboratory assistance. Gratitude to icipe’s Insects for Food Feed and Other Uses (INSEFF) Program team for providing the frass fertilizer used during the studies.
Funding
The authors gratefully acknowledge the financial support for this research by the following organizations and agencies: European Union (FAW-IPM, Grant No: DCI-FOOD/2017/), IKEA Foundation (Grant No: DN00151), Australian Centre for International Agricultural Research (ACIAR) (Protein Africa, Grant No: LS/2020/154), the Rockefeller Foundation (WAVE-IN, Grant No: 2021 FOD 030); Bill and Melinda Gates Foundation (INV-032416), European Commission (HORIZON 101060762 NESTLER and HORIZON 101136739 INNOECOFOOD), The French Ministry of Europe and Foreign Affairs (BIO Kenya project- FEF N°2024-53), Postkode Lottery, Sweden (Waste for Cash Eco Project (WACEP-PJ1651), the Curt Bergfors Foundation Food Planet Prize Award; the Swedish International Development Cooperation Agency (Sida); the Swiss Agency for Development and Cooperation (SDC); the Australian Centre for International Agricultural Research (ACIAR); the Government of Norway; the German Federal Ministry for Economic Cooperation and Development (BMZ); and the Government of the Republic of Kenya. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. The views expressed herein do not necessarily reflect the official opinion of the donors.
Author information
Authors and Affiliations
Contributions
DMM, JMM, and AAJ conceived the idea and designed the study; JMM collected data; DMM, AAJ, DB, and JMM analyzed data; and led the drafting of the manuscript; AK, DB, SS, TD, SE, and CMT read and reviewed the manuscript. DMM supervised the work. SS and CMT provided resources. All authors critically reviewed and approved the final.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Mutyambai, D.M., Mutua, J.M., Jalloh, A.A. et al. Insect frass fertilizer upregulates maize defence genes and resistance against an invasive herbivore pest. Sci Rep 15, 29978 (2025). https://doi.org/10.1038/s41598-025-14883-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-14883-3
Keywords
This article is cited by
-
Uncovering Ethical Blind Spots and Operational Pathways in Sustainable Maggot-Based Waste Management Systems
Journal of Agricultural and Environmental Ethics (2026)
